QTL Mapping of Candidate Genes Involved in Cd Accumulation in Rice Grain
Chenyang Pan1, Hanfei Ye1, Weiyong Zhou3, Sheng Wang1, Mengjia Li1, Mei Lu1, Sanfeng Li2, Xudong Zhu2, Yuexing Wang2, Yuchun Rao,1,*, Gaoxing Dai,3,*1College of Chemistry and Life Sciences, Zhejiang Normal University, Jinhua 321004, China 2State Key Laboratory of Rice Biology, China National Rice Research Institute, Hangzhou 310006, China 3Guangxi Academy of Agricultural Sciences, Nanning 530007, China
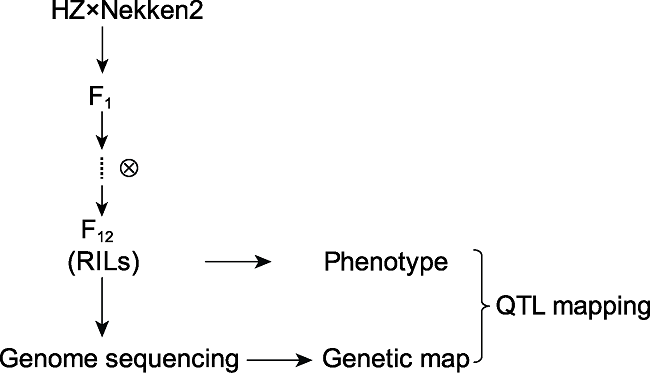
Abstract Rice is one of the world’s most important crops, but cadmium (Cd) pollution and Cd accumulation in rice fields pose a serious threat to its yield and quality as well as to human health. How to reduce Cd accumulation in rice has become a major concern. In this study, a japonica rice variety Nekken2 was used as the maternal parent, and an indica rice variety HZ was used as the paternal parent. After F1 generation was obtained by cross-breeding, successive selfings were performed for 120 recombinant inbred lines (RILs) population as the QTL mapping populations. We measured the Cd content in rice grains of each line at the maturity stage. Meanwhile, QTL mapping was performed using genetic map. As a result, a total of 7 QTLs were detected, which were located on chromosome 2, 3, 9, and 12 of rice, with the highest LOD score of 4.97. The qRT-PCR analysis on the candidate genes involved in metal ion stress resistance in these QTL regions showed that the expression level of LOC_Os02g50240, LOC_Os02g52780, LOC_Os09g31200, LOC_Os09g35030 and LOC_Os09g37949 was significantly different from the parents. Combined with the data of concentration of different metal ions in the parents, we speculated that the high expression of LOC_Os02g50240, LOC_Os09g31200 and LOC_Os09g35030 might greatly improve the absorption and tolerance of rice to Cd ion stress. And these genes may be related to the accumulation of Cd in rice grains, and may affect the ability of rice to tolerate Cd stress, which could help for further breeding and screening for rice varieties to tolerate Cd stress, and exploring the molecular regulation mechanism of Cd accumulation in rice. Keywords:rice;cadmium accumulation;QTL mapping;candidate genes;ion stress;genetic map
PDF (1630KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 潘晨阳, 叶涵斐, 周维永, 王盛, 李梦佳, 路梅, 李三峰, 朱旭东, 王跃星, 饶玉春, 戴高兴. 水稻籽粒镉积累QTL定位及候选基因分析. 植物学报, 2021, 56(1): 25-32 doi:10.11983/CBB20148 Pan Chenyang, Ye Hanfei, Zhou Weiyong, Wang Sheng, Li Mengjia, Lu Mei, Li Sanfeng, Zhu Xudong, Wang Yuexing, Rao Yuchun, Dai Gaoxing. QTL Mapping of Candidate Genes Involved in Cd Accumulation in Rice Grain. Chinese Bulletin of Botany, 2021, 56(1): 25-32 doi:10.11983/CBB20148
AbeT, NonoueY, OnoN, OmotenoM, KuramataM, FukuokaS, YamamotoT, YanoM, IshikawaS (2013). Detection of QTLs to reduce cadmium content in rice grains using LAC23/Koshihikari chromosome segment substitution lines Breed Sci 63, 284-291. URLPMID:24273423 [本文引用: 1]
ArisawaK, UemuraH, HiyoshiM, TakedaH, SaitoH, SodaM (2007). Cadmium-induced renal dysfunction and mortality in two cohorts: disappearance of the association in a generation born later Toxicol Lett 169, 214-221. URLPMID:17336471 [本文引用: 1]
ChengYW, QiYC, ZhuQ, ChenX, WangN, ZhaoX, ChenHY, CuiXJ, XuLL, ZhangW (2009). New changes in the plasma-membrane-associated proteome of rice roots under salt stress Proteomics 9, 3100-3114. [本文引用: 1]
DubouzetJG, SakumaY, ItoY, KasugaM, DubouzetEG, MiuraS, SekiM, ShinozakiK, Yamaguchi-ShinozakiK (2003). OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought-, high-salt- and cold-responsive gene expression Plant J 33, 751-763. [本文引用: 1]
FlorijnPJ, van BeusichemML (1993). Uptake and distribution of cadmium in maize inbred lines Plant Soil 150, 25-32. [本文引用: 1]
FujimakiS, SuzuiN, IshiokaNS, KawachiN, ItoS, ChinoM, NakamuraS (2010). Tracing cadmium from culture to spikelet: noninvasive imaging and quantitative characterization of absorption, transport, and accumulation of cadmium in an intact rice plant Plant Physiol 152, 1796-1806. [本文引用: 1]
HoriguchiH, TeranishiH, NiiyaK, AoshimaK, KatohK, SakuragawaN, KasuyaM (1994). Hypoproduction of erythropoietin contributes to anemia in chronic cadmium intoxication: clinical study on Itai-itai disease in Japan Arch Toxicol 68, 632-636. URLPMID:7857202 [本文引用: 1]
HuDW, ShengZH, LiQL, ChenW, WeiXJ, XieLH, JiaoGA, ShaoGN, WangJL, TangSQ, HuPS (2018). Identification of QTLs associated with cadmium concentration in rice grains J Integr Agric 17, 1563-1573. [本文引用: 2]
IshikawaS, AbeT, KuramataM, YamaguchiM, AndoT, YamamotoT, YanoM (2010). A major quantitative trait locus for increasing cadmium-specific concentration in rice grain is located on the short arm of chromosome 7 J Exp Bot 61, 923-934. URLPMID:20022924 [本文引用: 1]
LiuWQ, PanXW, LiYC, DuanYH, MinJ, LiuSX, LiuLC, ShengXN, LiXX (2019). Identification of QTLs and validation of qCd-2 associated with grain cadmium concentrations in rice Rice Sci 26, 42-49. [本文引用: 2]
LivakKJ, SchmittgenTD (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2?ΔΔCT method Methods 25, 402-408. URLPMID:11846609 [本文引用: 1]
MaoCP, SongYX, ChenLX, JiJF, LiJZ, YuanXY, YangZF, AyokoGA, FrostRL, TheissF (2019). Human health risks of heavy metals in paddy rice based on transfer characteristics of heavy metals from soil to rice Catena 175, 339-348. [本文引用: 1]
MiyadateH, AdachiS, HiraizumiA, TezukaK, NakazawaN, KawamotoT, KatouK, KodamaI, SakuraiK, TakahashiH, Satoh-NagasawaN, WatanabeA, FujimuraT, AkagiH (2011). OsHMA3, a P1B-type of ATPase affects root-to-shoot cadmium translocation in rice by mediating efflux into vacuoles New Phytol 189, 190-199. DOI:10.1111/j.1469-8137.2010.03459.xURLPMID:20840506 [本文引用: 1] * The cadmium (Cd) over-accumulating rice (Oryza sativa) cv Cho-Ko-Koku was previously shown to have an enhanced rate of root-to-shoot Cd translocation. This trait is controlled by a single recessive allele located at qCdT7. * In this study, using positional cloning and transgenic strategies, heavy metal ATPase 3 (OsHMA3) was identified as the gene that controls root-to-shoot Cd translocation rates. The subcellular localization and Cd-transporting activity of the gene products were also investigated. * The allele of OsHMA3 that confers high root-to-shoot Cd translocation rates (OsHMA3mc) encodes a defective P(1B) -ATPase transporter. OsHMA3 fused to green fluorescent protein was localized to vacuolar membranes in plants and yeast. An OsHMA3 transgene complemented Cd sensitivity in a yeast mutant that lacks the ability to transport Cd into vacuoles. By contrast, OsHMA3mc did not complement the Cd sensitivity of this yeast mutant, indicating that the OsHMA3mc transport function was lost. * We propose that the root cell cytoplasm of Cd-overaccumulating rice plants has more Cd available for loading into the xylem as a result of the lack of OsHMA3-mediated transportation of Cd to the vacuoles. This defect results in Cd translocation to the shoots in higher concentrations. These data demonstrate the importance of vacuolar sequestration for Cd accumulation in rice.
MukhopadhyayA, VijS, TyagiAK (2004). Overexpression of a zinc-finger protein gene from rice confers tolerance to cold, dehydration, and salt stress in transgenic tobacco Proc Natl Acad Sci USA 101, 6309-6314. URLPMID:15079051 [本文引用: 1]
NakanishiH, OgawaI, IshimaruY, MoriS, NishizawaNK (2006). Iron deficiency enhances cadmium uptake and translocation mediated by the Fe2+ transporters OsIRT1 and OsIRT2 in rice Soil Sci Plant Nut 52, 464-469. [本文引用: 1]
SatarugS, GarrettSH, SensMA, SensDA (2010). Cadmium, environmental exposure, and health outcomes Environ Health Perspect 118, 182-190. DOI:10.1289/ehp.0901234URLPMID:20123617 [本文引用: 1] OBJECTIVES: We provide an update of the issues surrounding health risk assessment of exposure to cadmium in food. DATA SOURCES: We reviewed epidemiologic studies published between 2004 and 2009 concerning the bioavailability of cadmium in food, assessment of exposure, and body burden estimate, along with exposure-related effects in nonoccupationally exposed populations. DATA EXTRACTION AND SYNTHESIS: Bioavailability of ingested cadmium has been confirmed in studies of persons with elevated dietary exposure, and the findings have been strengthened by the substantial amounts of cadmium accumulated in kidneys, eyes, and other tissues and organs of environmentally exposed individuals. We hypothesized that such accumulation results from the efficient absorption and systemic transport of cadmium, employing multiple transporters that are used for the body's acquisition of calcium, iron, zinc, and manganese. Adverse effects of cadmium on kidney and bone have been observed in environmentally exposed populations at frequencies higher than those predicted from models of exposure. Increasing evidence implicates cadmium in the risk of diseases that involve other tissues and organ systems at cadmium concentrations that do not produce effects on bone or renal function. CONCLUSIONS: Population data raise concerns about the validity of the current safe intake level that uses the kidney as the sole target in assessing the health risk from ingested cadmium. The data also question the validity of incorporating the default 5% absorption rate in the threshold-type risk assessment model, known as the provisional tolerable weekly intake (PTWI), to derive a safe intake level for cadmium.
Satoh-NagasawaN, MoriM, NakazawaN, KawamotoT, NagatoY, SakuraiK, TakahashiH, WatanabeA, AkagiH (2012). Mutations in rice (Oryza sativa) heavy metal ATPase 2 (OsHMA2) restrict the translocation of zinc and cadmium Plant Cell Physiol 53, 213-224. [本文引用: 1]
ShiJC, YuXL, ZhangMK, LuSG, WuWH, WuJJ, XuJM (2011). Potential risks of copper, zinc, and cadmium pollution due to pig manure application in a soil-rice system under intensive farming: a case study of Nanhu, China J Environ Qual 40, 1695-1704. DOI:10.2134/jeq2010.0316URLPMID:22031551 [本文引用: 1] Heavy metal (copper [Cu], zinc [Zn], and cadmium [Cd]) pollution of soils from pig manures in soil-rice ( L.) systems under intensive farming was investigated, taking Nanhu, China, as the case study area. Two hundred pig manures and 154 rice straws, brown rice samples, and corresponding surface soil (0-15 cm) samples were collected in paddy fields from 150 farms in 16 major villages within the study area. The mean Cu and Zn concentrations in pig manures consistently exceeded the related standard. About 44 and 60% of soil samples exceed the Chinese Soil Cu and Cd Environmental Quality Standards, respectively. The concentration of Cu, Zn, and Cd in brown rice did not exceed the Chinese Food Hygiene Standard. There was a significant positive correlation between total Cu and Zn contents in soil and application rate of pig manures. Strong correlation was observed between the extractable Cu, Zn, and Cd in soil and the Cu, Zn, and Cd contents in the brown rice. The spatial distribution maps of Cu and Zn concentrations in brown rice, straw, and extractable soil Cu and Zn concentration also showed similar geographical trends. Further analyses on heavy metals loading flux and accumulation rates from pig manure applied suggested that Cu and Cd contents in soil currently have already exceeded the maximum permissible limit, and Zn, if still at current manure application rates, will reach the ceiling concentration limits in 9 yr. This study assists in understanding the risk of heavy metals accumulating from pig manure applications to agricultural soils.
SinghKK, GhoshS (2013). Regulation of glutamine synthetase isoforms in two differentially drought-tolerant rice (Oryza sativa L.) cultivars under water deficit conditions Plant Cell Rep 32, 183-193. [本文引用: 1]
UenoD, KonoI, YokoshoK, AndoT, YanoM, MaJF (2009a). A major quantitative trait locus controlling cadmium translocation in rice (Oryza sativa) New Phytol 182, 644-653. URLPMID:19309445 [本文引用: 1]
UenoD, KoyamaE, KonoI, AndoT, YanoM, MaJF (2009b). Identification of a novel major quantitative trait locus controlling distribution of Cd between roots and shoots in rice Plant Cell Physiol 50, 2223-2233. [本文引用: 2]
UraguchiS, KamiyaT, SakamotoT, KasaiK, SatoY, NagamuraY, YoshidaA, KyozukaJ, IshikawaS, FujiwaraT (2011). Low-affinity cation transporter (OsLCT1) regulates cadmium transport into rice grains Proc Natl Acad Sci USA 108, 20959-20964. DOI:10.1073/pnas.1116531109URLPMID:22160725 [本文引用: 1] Accumulation of cadmium (Cd) in rice (Oryza sativa L.) grains poses a potential health problem, especially in Asia. Most Cd in rice grains accumulates through phloem transport, but the molecular mechanism of this transport has not been revealed. In this study, we identified a rice Cd transporter, OsLCT1, involved in Cd transport to the grains. OsLCT1-GFP was localized at the plasma membrane in plant cells, and OsLCT1 showed Cd efflux activity in yeast. In rice plants, strong OsLCT1 expression was observed in leaf blades and nodes during the reproductive stage. In the uppermost node, OsLCT1 transcripts were detected around large vascular bundles and in diffuse vascular bundles. RNAi-mediated knockdown of OsLCT1 did not affect xylem-mediated Cd transport but reduced phloem-mediated Cd transport. The knockdown plants of OsLCT1 accumulated approximately half as much Cd in the grains as did the control plants. The content of other metals in rice grains and plant growth were not negatively affected by OsLCT1 suppression. These results suggest that OsLCT1 functions at the nodes in Cd transport into grains and that in a standard japonica cultivar, the regulation of OsLCT1 enables the generation of
WangFJ, WangM, LiuZB, ShiY, HanTQ, YeYY, GongN, SunJW, ZhuC (2015). Different responses of low grain-Cd-accumulating and high grain-Cd-accumulating rice cultivars to Cd stress Plant Physiol Biochem 96, 261-269. [本文引用: 1]
YuH, WangJL, FangW, YuanJG, YangZY (2006). Cadmium accumulation in different rice cultivars and screening for pollution-safe cultivars of rice Sci Total Environ 370, 302-309. DOI:10.1016/j.scitotenv.2006.06.013URLPMID:16870236 [本文引用: 1] Large areas of contaminated land are being used for agricultural production in some countries due to the high demand for food. To minimize the influx of pollutants to the human food chain through consumption of agricultural products, we propose the concept of pollution-safe cultivars (PSCs), i.e. cultivars whose edible parts accumulate a specific pollutant at a level low enough for safe consumption, even when grown in contaminated soil. We tested the feasibility of the PSC concept by growing 43 cultivars of paddy rice (Oryza sativa L., including 20 normal and 23 hybrid cultivars) under a high (75.69-77.55 mg kg(-1)) and a low (1.75-1.85 mg kg(-1)) cadmium (Cd) exposure. These pot experiments took place in the spring and summer of 2004. At the low level of Cd exposure, 30 out of the 43 tested cultivars were found to be Cd-PSCs. Grain Cd concentrations were highly correlated (p<0.01) between the two experiments, suggesting that Cd accumulation in rice grain is genotype-dependent and that the selection of PSCs is possible, at least at a certain level of soil contamination. No Cd-PSCs were found under the high level of Cd exposure. Yield was enhanced in some cultivars and depressed in others in response to elevated soil Cd, indicating that farmers cannot rely on yield depression as an indicator of toxicity of the grains. It is therefore important and feasible to screen for PSCs and to establish PSC breeding programs to effectively and efficiently reduce the risk of human exposure to soil pollutants, such as Cd, through crop consumption.
ZhangXQ, ZhangGP, GuoLB, WangHZ, ZengDL, DongGJ, QianQ, XueDW (2011). Identification of quantitative trait loci for Cd and Zn concentrations of brown rice grown in Cd-polluted soils Euphytica 180, 173-179. [本文引用: 1]
ZongW, TangN, YangJ, PengL, MaSQ, XuY, LiGL, XiongLZ (2016). Feedback regulation of ABA signaling and biosynthesis by a bZIP transcription factor targets drought-resistance-related genes Plant Physiol 171, 2810-2825. [本文引用: 1]
植物防御素调控水稻镉积累的新机制 1 2018
... 镉(Cd)在土壤中移动性高, 毒性极强, 是最具危害的重金属元素之一, 极易被植物吸收并运输储存至不同部位, 通过食物链进入生物体并在体内累积, 引起人畜慢性中毒, 给动物及人类的生存带来巨大威胁(Florijn and van Beusichem, 1993).当人摄入过量Cd时会引起肾小管功能障碍、骨质疏松、骨痛(itai- itai)等疾病(Horiguchi et al., 1994; Arisawa et al., 2007), 甚至可能增加癌症的发病率(Satarug et al., 2010).20世纪以来, 随着工业的快速发展, 过度施肥和污染物排放等问题不断恶化, 稻田Cd污染程度日益加剧(Yu et al., 2006; Shi et al., 2011).随着土壤pH值的下降, 水稻(Oryza sativa) Cd污染问题愈加严重(Mao et al., 2019), 世界许多地区均出现水稻Cd含量严重超标的情况.稻田Cd污染的治理包括人为Cd排放量的控制和水稻中Cd积累的抑制两方面.鉴于我国人多地少, 在中、轻度污染土壤上进行低Cd积累作物的种植, 符合经济与生态双重效益, 边生产边修复的新理念成为共识(黄新元和赵方杰, 2018).要实现这一目标, 需挖掘作物阻控Cd相关基因, 深入剖析水稻Cd元素累积的遗传机制, 以阻滞Cd通过食物链富集. ...
Cadmium-induced renal dysfunction and mortality in two cohorts: disappearance of the association in a generation born later 1 2007
... 镉(Cd)在土壤中移动性高, 毒性极强, 是最具危害的重金属元素之一, 极易被植物吸收并运输储存至不同部位, 通过食物链进入生物体并在体内累积, 引起人畜慢性中毒, 给动物及人类的生存带来巨大威胁(Florijn and van Beusichem, 1993).当人摄入过量Cd时会引起肾小管功能障碍、骨质疏松、骨痛(itai- itai)等疾病(Horiguchi et al., 1994; Arisawa et al., 2007), 甚至可能增加癌症的发病率(Satarug et al., 2010).20世纪以来, 随着工业的快速发展, 过度施肥和污染物排放等问题不断恶化, 稻田Cd污染程度日益加剧(Yu et al., 2006; Shi et al., 2011).随着土壤pH值的下降, 水稻(Oryza sativa) Cd污染问题愈加严重(Mao et al., 2019), 世界许多地区均出现水稻Cd含量严重超标的情况.稻田Cd污染的治理包括人为Cd排放量的控制和水稻中Cd积累的抑制两方面.鉴于我国人多地少, 在中、轻度污染土壤上进行低Cd积累作物的种植, 符合经济与生态双重效益, 边生产边修复的新理念成为共识(黄新元和赵方杰, 2018).要实现这一目标, 需挖掘作物阻控Cd相关基因, 深入剖析水稻Cd元素累积的遗传机制, 以阻滞Cd通过食物链富集. ...
New changes in the plasma-membrane-associated proteome of rice roots under salt stress 1 2009
... 已有研究表明, 基因LOC_Os02g52780和LOC_ Os09g35030分别属于bZIP转录因子和AP2/EREBP转录因子, 前者正向调控OsPP2C49和9-顺式-环氧类胡萝卜素双加氧酶基因OsNCED4, 是ABA信号合成及抗旱性的中心调控因子(Zong et al., 2016), 通过ABA依赖途径调节胁迫相关基因的表达, 增强抗旱耐盐性; 后者与干旱响应元件特异结合, 控制许多胁迫诱导基因的表达, 具有培育抗旱耐盐和耐冷转基因水稻的潜力(Dubouzet et al., 2003).基因LOC_ Os02g50240编码胞质谷氨酰胺合成酶, 参与多种生物过程的调控, 进而调节胁迫应答.干旱条件下, LOC_Os02g50240的表达可能对Khitish品种的耐旱性有贡献(Singh and Ghosh, 2013).基因LOC_ Os09g31200编码一种锌指蛋白, 过表达该基因的转基因烟草(Nicotiana tabacum)对冷、干旱和盐胁迫具有耐受性(Mukhopadhyay et al., 2004).基因LOC_Os09g37949编码一种受体蛋白激酶, 其表达受低温、干旱与脱落酸诱导, 可能在响应盐等逆境胁迫中起重要作用(Cheng et al., 2009). ...
OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought-, high-salt- and cold-responsive gene expression 1 2003
... 已有研究表明, 基因LOC_Os02g52780和LOC_ Os09g35030分别属于bZIP转录因子和AP2/EREBP转录因子, 前者正向调控OsPP2C49和9-顺式-环氧类胡萝卜素双加氧酶基因OsNCED4, 是ABA信号合成及抗旱性的中心调控因子(Zong et al., 2016), 通过ABA依赖途径调节胁迫相关基因的表达, 增强抗旱耐盐性; 后者与干旱响应元件特异结合, 控制许多胁迫诱导基因的表达, 具有培育抗旱耐盐和耐冷转基因水稻的潜力(Dubouzet et al., 2003).基因LOC_ Os02g50240编码胞质谷氨酰胺合成酶, 参与多种生物过程的调控, 进而调节胁迫应答.干旱条件下, LOC_Os02g50240的表达可能对Khitish品种的耐旱性有贡献(Singh and Ghosh, 2013).基因LOC_ Os09g31200编码一种锌指蛋白, 过表达该基因的转基因烟草(Nicotiana tabacum)对冷、干旱和盐胁迫具有耐受性(Mukhopadhyay et al., 2004).基因LOC_Os09g37949编码一种受体蛋白激酶, 其表达受低温、干旱与脱落酸诱导, 可能在响应盐等逆境胁迫中起重要作用(Cheng et al., 2009). ...
Uptake and distribution of cadmium in maize inbred lines 1 1993
... 镉(Cd)在土壤中移动性高, 毒性极强, 是最具危害的重金属元素之一, 极易被植物吸收并运输储存至不同部位, 通过食物链进入生物体并在体内累积, 引起人畜慢性中毒, 给动物及人类的生存带来巨大威胁(Florijn and van Beusichem, 1993).当人摄入过量Cd时会引起肾小管功能障碍、骨质疏松、骨痛(itai- itai)等疾病(Horiguchi et al., 1994; Arisawa et al., 2007), 甚至可能增加癌症的发病率(Satarug et al., 2010).20世纪以来, 随着工业的快速发展, 过度施肥和污染物排放等问题不断恶化, 稻田Cd污染程度日益加剧(Yu et al., 2006; Shi et al., 2011).随着土壤pH值的下降, 水稻(Oryza sativa) Cd污染问题愈加严重(Mao et al., 2019), 世界许多地区均出现水稻Cd含量严重超标的情况.稻田Cd污染的治理包括人为Cd排放量的控制和水稻中Cd积累的抑制两方面.鉴于我国人多地少, 在中、轻度污染土壤上进行低Cd积累作物的种植, 符合经济与生态双重效益, 边生产边修复的新理念成为共识(黄新元和赵方杰, 2018).要实现这一目标, 需挖掘作物阻控Cd相关基因, 深入剖析水稻Cd元素累积的遗传机制, 以阻滞Cd通过食物链富集. ...
Tracing cadmium from culture to spikelet: noninvasive imaging and quantitative characterization of absorption, transport, and accumulation of cadmium in an intact rice plant 1 2010
Potential risks of copper, zinc, and cadmium pollution due to pig manure application in a soil-rice system under intensive farming: a case study of Nanhu, China 1 2011
... 镉(Cd)在土壤中移动性高, 毒性极强, 是最具危害的重金属元素之一, 极易被植物吸收并运输储存至不同部位, 通过食物链进入生物体并在体内累积, 引起人畜慢性中毒, 给动物及人类的生存带来巨大威胁(Florijn and van Beusichem, 1993).当人摄入过量Cd时会引起肾小管功能障碍、骨质疏松、骨痛(itai- itai)等疾病(Horiguchi et al., 1994; Arisawa et al., 2007), 甚至可能增加癌症的发病率(Satarug et al., 2010).20世纪以来, 随着工业的快速发展, 过度施肥和污染物排放等问题不断恶化, 稻田Cd污染程度日益加剧(Yu et al., 2006; Shi et al., 2011).随着土壤pH值的下降, 水稻(Oryza sativa) Cd污染问题愈加严重(Mao et al., 2019), 世界许多地区均出现水稻Cd含量严重超标的情况.稻田Cd污染的治理包括人为Cd排放量的控制和水稻中Cd积累的抑制两方面.鉴于我国人多地少, 在中、轻度污染土壤上进行低Cd积累作物的种植, 符合经济与生态双重效益, 边生产边修复的新理念成为共识(黄新元和赵方杰, 2018).要实现这一目标, 需挖掘作物阻控Cd相关基因, 深入剖析水稻Cd元素累积的遗传机制, 以阻滞Cd通过食物链富集. ...
Regulation of glutamine synthetase isoforms in two differentially drought-tolerant rice (Oryza sativa L.) cultivars under water deficit conditions 1 2013
... 已有研究表明, 基因LOC_Os02g52780和LOC_ Os09g35030分别属于bZIP转录因子和AP2/EREBP转录因子, 前者正向调控OsPP2C49和9-顺式-环氧类胡萝卜素双加氧酶基因OsNCED4, 是ABA信号合成及抗旱性的中心调控因子(Zong et al., 2016), 通过ABA依赖途径调节胁迫相关基因的表达, 增强抗旱耐盐性; 后者与干旱响应元件特异结合, 控制许多胁迫诱导基因的表达, 具有培育抗旱耐盐和耐冷转基因水稻的潜力(Dubouzet et al., 2003).基因LOC_ Os02g50240编码胞质谷氨酰胺合成酶, 参与多种生物过程的调控, 进而调节胁迫应答.干旱条件下, LOC_Os02g50240的表达可能对Khitish品种的耐旱性有贡献(Singh and Ghosh, 2013).基因LOC_ Os09g31200编码一种锌指蛋白, 过表达该基因的转基因烟草(Nicotiana tabacum)对冷、干旱和盐胁迫具有耐受性(Mukhopadhyay et al., 2004).基因LOC_Os09g37949编码一种受体蛋白激酶, 其表达受低温、干旱与脱落酸诱导, 可能在响应盐等逆境胁迫中起重要作用(Cheng et al., 2009). ...
 ,1,*, 戴高兴
,1,*, 戴高兴

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT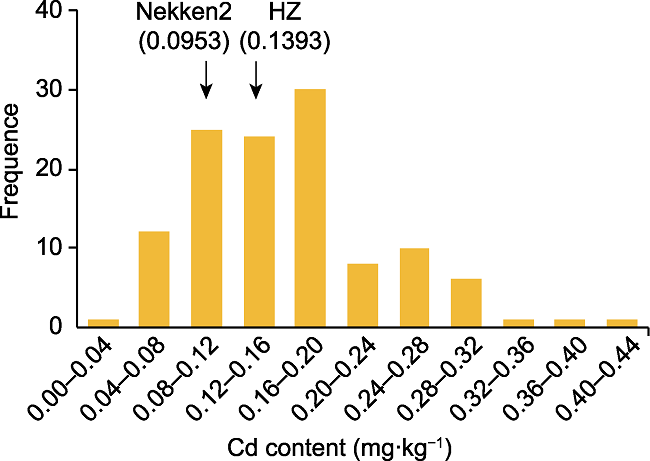 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT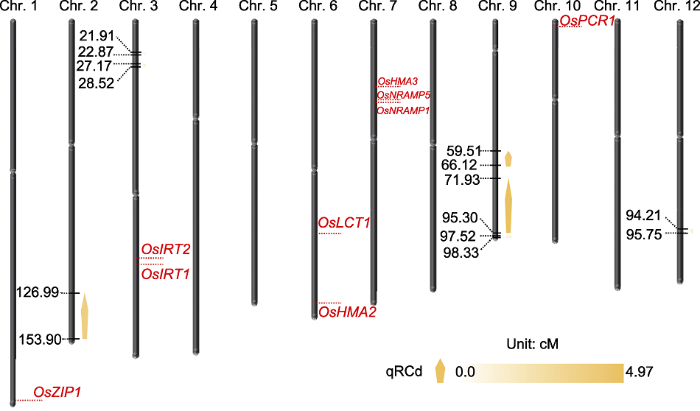 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT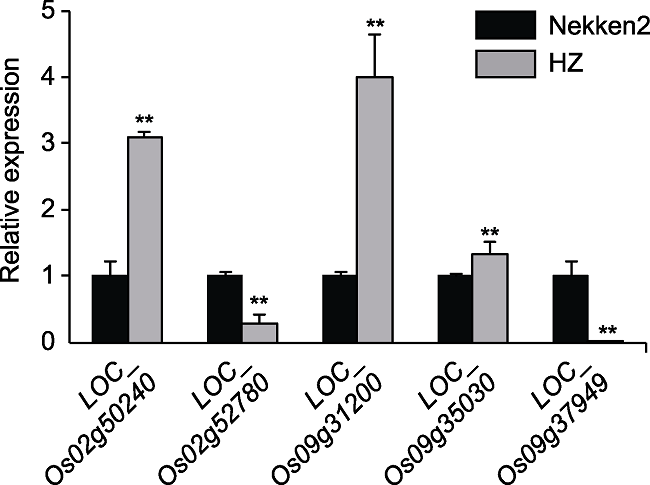 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}