 ,2,*, 白史且,3,4,*
,2,*, 白史且,3,4,*Establishment of Biolistic Mediated Transformation System for Elymus sibiricus
Pengfei Du1,2,3, Yu Wang2, Yingping Cao2, Song Yang2, Zhichao Sun2, Decai Mao3,4, Jiajun Yan3,4, Daxu Li3,4, Meizhen Sun5, Chunxiang Fu,2,*, Shiqie Bai,3,4,*通讯作者: E-mail:fucx@qibebt.ac.cn;610852681@qq.com
责任编辑: 朱亚娜
收稿日期:2020-11-2接受日期:2021-01-5网络出版日期:2021-01-01
| 基金资助: |
Corresponding authors: E-mail:fucx@qibebt.ac.cn;610852681@qq.com
Received:2020-11-2Accepted:2021-01-5Online:2021-01-01

摘要
川草2号老芒麦(Elymus sibiricus)是青藏高原地区治理荒漠化和建设高产人工草地的主要栽培草种。用川草2号老芒麦5种外植体诱导愈伤组织, 经分化测试, 仅幼穗愈伤组织能分化再生。以当代培养25天和35天的结构致密坚硬的幼穗愈伤组织为受体, 分别进行农杆菌侵染和基因枪转化, 结果只有基因枪能转化成功。在基因枪转化过程中, 采用高渗培养和滤纸干燥2种方式预处理愈伤组织, 结果表明滤纸干燥处理比高渗处理转化效率高。当代诱导25天的幼穗愈伤组织, 滤纸干燥处理2小时转化效率最高, 达40%。该研究成功获得了基因枪转化的以川草2号老芒麦幼穗愈伤为受体的阳性愈伤组织。
关键词:
Abstract
Elymus sibiricus cv. ‘Chuancao No.2’ is the main cultivated grass species for desertification control and construction of high-yield and high-quality pasture in northwest Sichuan Plateau. In this study, we tested five explants of E. sibiricus cv. ‘Chuancao No.2’ for callus induction, and found that only inflorescence calli were able to differentiate and regenerate. The calli of inflorescence with dense and hard structure cultured for 25 d and 35 d were used for Agrobacterium and biolistic mediated transformation respectively. The results showed that only biolistic-mediated transformation could produce positive transgenic calli of ‘Chuancao No.2’. In the process of biolistic-mediated transformation, the calli was pretreated in two ways: hyperosmotic culture and filter paper drying. The results revealed that the transformation efficiency of filter paper drying was higher than that of hyperosmotic treatment. For the inflorescence callus after 25 d induction, the transformation efficiency under the condition of 2 h drying of filter paper was highest which reached about 40%. In short, we applied the biolistic technology in ‘Chuancao No.2’ for the first time and successfully obtained the positive transgenic inflorescence calli. This work will lead to establishment of the robust transformation system for E. sibiricus in future.
Keywords:
PDF (3053KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
杜鹏飞, 王玉, 曹英萍, 杨松, 孙志超, 毛德才, 鄢家俊, 李达旭, 孙美贞, 付春祥, 白史且. 基因枪介导的老芒麦遗传转化体系的建立. 植物学报, 2021, 56(1): 62-70 doi:10.11983/CBB20174
Du Pengfei, Wang Yu, Cao Yingping, Yang Song, Sun Zhichao, Mao Decai, Yan Jiajun, Li Daxu, Sun Meizhen, Fu Chunxiang, Bai Shiqie.

老芒麦(Elymus sibiricus)又名西伯利亚披碱草(Siberian wildrye), 属于禾本科(Poaceae)小麦族(Triticeae)披碱草属的一种多年生自花授粉、异源四倍体疏丛型旱生牧草。老芒麦在我国主要分布于东北、华北、西北和青藏高原等地区, 是青藏高原地区高寒草甸草原和草甸群落中的优势种和建群种(陈默君和贾慎修, 2002; 白史且和鄢家俊, 2020)。老芒麦植株分蘖好、再生力强、返青早、叶量丰富、适口性好, 其粗蛋白含量高达11.13%, 消化率达80%以上, 青草和调制的干草均为马、牛、羊所喜食(周国栋, 2011)。Dv?ráorak (1981)研究表明, 老芒麦具有耐旱、耐寒、耐湿、耐土壤贫瘠和耐盐等特性。此外, 老芒麦在退化草地修复改良和栽培草地建植中也具有重要作用(吴召林等, 2020)。川草2号老芒麦是四川省草原科学研究院以阿坝本地老芒麦为原始材料, 经过10年选育成功的一种多年生禾草新品种, 因其抗逆性强、产草量高、利用年限长、饲草品质优, 现已成为青藏高原牧区建设高产优质牧草基地的主要栽培草种(王元富等, 1995; 李才旺等, 1999; 李达旭, 2006)。因此, 对其深入研究将对我国北方草原和青藏高原高寒草甸的退化草地改良, 及发展草地畜牧业均具有重要意义(鄢家俊等, 2007)。
近年来, 随着分子生物技术的发展, 植物的功能基因研究成为热点, 为分子育种奠定了基础, 尤其是CRISPR/Cas9技术的产生, 大大加快了作物分子育种的进程, 如玉米(Zea mays) (Liang et al., 2014)、水稻(Oryza sativa) (Shan et al., 2013)、小麦(Triticum aestivum) (Shan et al., 2013)和烟草(Nicotiana tabacum) (Li et al., 2013)。但由于遗传转化体系的限制, 目前植物功能基因研究和分子育种主要集中在一些大作物和模式植物(Mao et al., 2013; Feng et al., 2013; Gao et al., 2015), 草业作物及其它经济作物遗传转化体系进展缓慢, 因此急需构建配套的遗传转化体系, 服务重要作物的基础研究及育种工作。
目前, 关于老芒麦组培快繁与遗传转化体系建立的报道相对较少。Lee等(2012)利用内蒙古野生老芒麦成熟种子诱导愈伤组织并分化形成再生植株; 王宇(2014)利用甘肃野生老芒麦幼穗诱导愈伤组织建立组培再生体系。李达旭等(2006)、Li等(2006)以及李达旭(2006)以成熟种子诱导的愈伤组织为转化受体, 建立了农杆菌介导的川草2号老芒麦遗传转化体系, 转化效率为11%。然而, 由于种子遗传背景复杂且质量参差不齐, 目前老芒麦的遗传转化仍存在体系不稳定、重复性差、难以实现批量转化等问题。与农杆菌介导的遗传转化体系相比, 基因枪介导的遗传转化体系不受物种和外植体类型的限制, 操作简单, 可控性强, 而且不会将目标序列之外的微生物基因引入植物细胞中, 因此该体系的效率与安全性较高。随着该技术的不断发展和完善, 目前已经能够通过控制质粒浓度有效克服基因枪转化导致的基因多拷贝串联问题。因此, 基因枪介导的遗传转化体系在植物尤其是难以遗传转化的禾本科植物中占据重要位置。例如, 在主要采用基因枪法和农杆菌介导法进行遗传转化的小麦中, 基因枪法的使用占68.8% (喻修道等, 2010), 然而该技术至今尚未在老芒麦中进行尝试。
本研究以优质老芒麦品种川草2号为研究对象, 旨在建立其稳定的遗传转化体系。分别以川草2号老芒麦成熟种子、根、茎、下胚轴和幼穗为外植体诱导愈伤组织, 仅幼穗愈伤组织能够分化再生成完整植株。以挑选的幼穗胚性愈伤系为受体, 分别进行农杆菌和基因枪转化, 获得了基因枪转化的阳性愈伤组织。研究结果为建立稳定的川草2号老芒麦遗传转化体系奠定了基础。
1 材料与方法
1.1 实验材料
川草2号老芒麦(Elymus sibiricus L. cv. ‘Chuancao No.2’)种子由四川省草原科学研究院提供, 生长地位于四川省阿坝藏族自治州红原县四川省草原科学研究院青藏高原草原科技创新基地。种子收获年限为2018年。1.2 载体
农杆菌和基因枪转化过程中所用载体为PANIC系列(Mann et al., 2012), 载体T-DNA区段如图1所示。图1
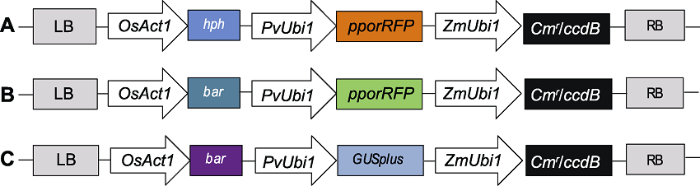 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1PANIC6A (A)、PANIC6D (B)和PANIC6E (C) T-DNA区段
LB: T-DNA区段左边界; OsAct1: 水稻启动子; hph/bar: 筛选标记基因; PvUbi1: 柳枝稷启动子; pporRFP/GUSplus: 红色荧光/葡萄糖苷酸酶报告基因; ZmUbi1: 玉米启动子; Cmr: 氯霉素抗性基因; ccdB: 目的基因; RB: T-DNA区段右边界
Figure 1T-DNA segment of PANIC6A (A), PANIC6D (B) and PANIC6E (C)
LB: Left boundary of T-DNA segment; OsAct1: Rice promoter; hph/bar: Screening marker genes; PvUbi1: Switchgrass promoter; pporRFP/GUSplus: Red fluorescence/glucuronidase reporter gene; ZmUbi1: Maize promoter; Cmr: Chloramphenicol resistant gene; ccdB: Target gene; RB: Right boundary of T-DNA segment
1.3 方法
1.3.1 外植体准备选取籽粒饱满、结构完整且状态一致的川草2号老芒麦种子, 剥去颖壳。用100 mg?L-1赤霉素(GA3)浸泡24小时, 无菌水冲洗3次, 加2滴Tween-20, 无菌水冲洗至无泡沫。然后用75%乙醇浸泡50秒, 无菌水冲洗3次, 加入5% NaClO浸泡8-10分钟。最后用无菌水冲洗3-5次后置于超净台的无菌滤纸上吹干。将洗净晾干的老芒麦种子(约5万粒)按每皿30粒接种于愈伤组织诱导培养基, 26°C暗培养。同时, 将部分种子接种于生根培养基, 7天后剪取其细根(将其切成3-4 mm的小段; 约300皿, 每皿10段)和下胚轴(约200皿, 每皿10个)接种于诱导培养基上, 26°C暗培养。
部分川草2号老芒麦种子于中国科学院青岛生物能源与过程研究所能源作物分子育种团队基地进行土播(3月下旬), 到孕穗期且未抽穗(6月中旬)时收集老芒麦植株上半部。去掉外层的叶片后在无菌条件下用75%乙醇消毒8分钟后晾干, 用镊子小心剥取幼穗, 将其切成4-5 mm的小段后接种于愈伤组织诱导培养基上(约500皿, 每皿10段)进行26°C暗培养。另选取幼嫩带茎节的部位, 从茎节中间切开, 然后切成3- 4 mm的小段, 带切口一端接触愈伤组织诱导培养基(约100皿, 每皿10段)进行26°C暗培养。
统计上述愈伤组织的诱导时间以及诱导率。诱导出愈伤组织30天后转入继代培养基, 每20天继代1次, 待长出具有边界、结构致密且呈颗粒状的淡黄色胚性愈伤组织后接种于分化培养基, 30天后开始统计分化率(每皿30个愈伤组织, 重复5次)。待分化出小苗后接种于生根培养基, 生根完成, 炼苗3-5天后移栽。
诱导率=(出愈外植体数/接种外植体数)×100%
分化率=(出芽愈伤组织数/接种愈伤组织数)× 100%
1.3.2 培养基成分
基本培养基: MS培养基, 蔗糖30 g?L-1, 琼脂7.5- 7.8 g?L-1, pH值均为5.9-6.0。诱导培养基: 基本培养基+5 mg?L-1 2,4-D+0.15 mg?L-1 6-BA。继代培养基: 基本培养基+3 mg?L-1 2,4-D+0.15 mg?L-1 6-BA。分化培养基: 基本培养基+0.5 mg?L-1 6-BA+2 mg?L-1 KT。生根培养基: 1/2MS。高渗培养基: MS基本培养基+72 g?L-1甘露醇。培养基均经过液体高压灭菌(121°C, 15分钟)。
1.3.3 农杆菌转化
用热激法(Mann et al., 2012)将PANIC6A/6D/6E (图1A-C)分别转化根癌农杆菌EHA105 (本研究所用农杆菌和大肠杆菌菌株由中国科学院青岛生物能源与过程研究所能源与作物分子育种团队提供), 涂板约48小时后, 挑取单克隆接种到添加50 mg?L-1卡那霉素和50 mg?L-1利福平的600 μL LB培养液中, 于28°C、每分钟200转的摇床中摇菌约14小时。菌液检测呈阳性后, 取200 μL菌液加入含有50 mg?L-1卡那霉素和50 mg?L-1利福平的50 mL LB培养液中, 于28°C、每分钟200转的摇床中摇至OD600=0.4, 然后加入乙酰丁香酮(AS)至终浓度为100 μmol?L-1。继续摇约2小时至OD600=0.6时, 851 ×g 离心15分钟收集菌体。用侵染液(未加琼脂的继代培养基)重悬菌体, 至终浓度OD600=0.3, 乙酰丁香酮浓度为100 μmol?L-1时开始侵染。侵染受体为结构致密且状态较一致的川草2号老芒麦幼穗愈伤组织(约培养30天)和幼穗。农杆菌侵染方法参照Wang和Ge (2005)的方法并进行改动, 4种侵染方案具体见表1。
Table 1
表1
表14种农杆菌侵染方案
Table 1
| Infect schemes | Infection methods | |||
|---|---|---|---|---|
| Predrying treatment (10 min) | Vacuum treatment (10 min) | Ultrasonic treatment (5 min) | Vacuum treatment (10 min) | |
| 1 | - | + | + | + |
| 2 | - | + | + | - |
| 3 | - | - | + | + |
| 4 | + | + | + | + |
新窗口打开|下载CSV
用4种侵染方案侵染后, 分别将愈伤组织碎片和农杆菌轻轻摇动孵育10分钟, 孵育后用无菌滤纸吸干愈伤组织表面菌液并晾干。将愈伤置于用侵染液微湿的无菌滤纸上(每个滤纸上的愈伤组织块数大约等于20个原始完整愈伤组织), 23°C暗培养2-3天。在体式显微镜(SZX16, 日本奥林巴斯公司)下进行红色荧光跟踪观测与拍照(RFP荧光激发块型号为U-RFL-T, 日本奥林巴斯公司), 统计荧光愈伤比率(每皿接种20粒愈伤, 重复3次)。GUS染色操作参照Wang和Ge等(2005)的方法(每皿接种20粒染色愈伤, 重复3次), 统计GUS染色比率。
荧光愈伤比率=(带有红色荧光的愈伤数/接种愈伤数)×100%
GUS染色比率=(带有蓝绿色的愈伤数/接种染色愈伤数)×100%
1.3.4 基因枪转化
1.3.4.1 愈伤受体准备和预处理
以川草2号老芒麦幼穗诱导约25天、35天的愈伤组织为受体。挑选结构致密且状态较一致的胚性愈伤组织置于无菌培养皿中, 用镊子捏碎, 混匀备用。分2种预处理方式: (1) 高渗处理, 将愈伤在高渗培养基上培养6小时; (2) 滤纸干燥, 将愈伤在2层无菌滤纸上干燥处理2小时, 滤纸使用之前置于愈伤诱导培养基表面微浸湿, 用Pirifilm膜(PM-96, 美国Bemis公司) 封口。
1.3.4.2 菌株和质粒
用热激法将PANIC6A (图1A)和PANIC6D (图1B)两种质粒(Mann et al., 2012)转化DH5α大肠杆菌。用试剂盒(Ml13101M, 莫纳生物公司)提取质粒, 经琼脂糖凝胶电泳检测后, 用紫外吸收法检测质粒DNA的浓度和纯度, 并置于-20°C冰箱中保存备用。
1.3.4.3 基因枪转化过程
用Bio-Rad公司PDS-1000He型基因枪轰击, 参照王关林和方宏筠(2002)的方法。具体过程如下: 洗涤微载体(金粉); 实验当天用DNA分子包埋微载体(DNA溶液浓度为1 μg?μL-1, 用2.5 mol?L-1 CaCl2 (氯化钙)和0.1 mol?L-1 C7H22CL3N3 (亚精胺)作沉淀剂)。每次轰击6 μL (500 μg微载体以及0.83 μg DNA), 轰击压力为7.58×106 Pa (1 100 psi), 轰击距离为9 cm, 每皿愈伤轰击1次。将轰击12小时后的愈伤组织转移到继代培养基上恢复培养, 并于体式显微镜下进行荧光跟踪观测与拍照, 统计荧光愈伤比率(每皿接种20粒愈伤, 重复3次)。
荧光愈伤比率=(带有红色荧光的愈伤数/接种愈伤数)×100%
2 结果与讨论
2.1 不同外植体愈伤诱导率以及出愈时长
用老芒麦种子(图2A)、根(图2B)、下胚轴(图2C)和茎(图2D)诱导的愈伤组织均呈水渍化, 海绵状, 结构较疏松; 除根诱导的愈伤呈白色外, 其余外植体诱导的愈伤均为深黄色。幼穗愈伤(图2E)结构相对致密, 具有明显的边界, 用镊子夹取发现其质地坚硬, 呈淡黄色。不同外植体诱导出愈伤所用时长和诱导率均有差异(图3)。根部出愈时间最长(约52天), 下胚轴、茎和种子次之; 幼穗出愈时间最短(约23天)。幼穗愈伤诱导率最高(达90%), 种子、根和茎次之, 下胚轴愈伤诱导率最低(约25%)。综合出愈时长、愈伤诱导率和愈伤状态, 表明幼穗作外植体最佳, 出愈时间短且诱导率高。图2
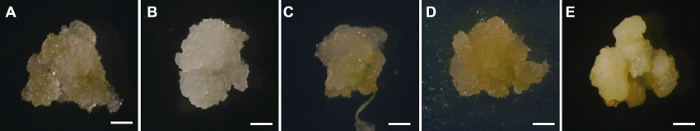 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同外植体诱导出愈伤后约30天的显微图
(A)-(E) 分别表示用种子(成熟胚)、根、下胚轴、茎和幼穗诱导出的愈伤。Bars=50 μm
Figure 2Micrographs of calli from different explants (30 d)
(A)-(E) Callus induced by seeds (mature embryo), roots, hypocotyl, stem and inflorescence, respectively. Bars=50 μm
图3
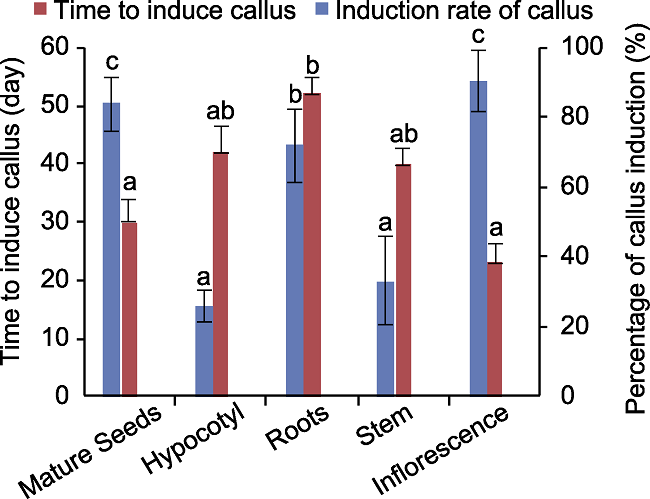 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3川草2号老芒麦不同外植体愈伤诱导所用时间及诱导率
误差线为标准误(SE), 样本容量为60。不同小写字母表示差异显著(P<0.05)。
Figure 3Callus induction time and induction rate of different explants in Elymus sibiricus cv. ‘Chuancao No.2’
The error bars indicate standard error (SE), the sample size was 60. Different lowercase letters indicate significant differences (P<0.05).
2.2 川草2号老芒麦幼穗组培再生体系建立
愈伤再生结果表明, 仅幼穗愈伤能分化出绿点和小苗(图4A-F)。幼穗愈伤在分化培养基继代30天后开始分化, 分化苗生根需25天, 从愈伤诱导到分化共需约115天。然而种子外植体诱导的愈伤无法获得再生苗(图4G-K), 根、茎、下胚轴均未成功建立再生体系(数据未列出)。在一定时间范围内, 幼穗愈伤分化率随再生时间推移而增加, 将幼穗愈伤置于分化培养基后约30天出现绿点, 分化率为30%; 50天后幼穗分化率不再增加, 达到峰值(约63%) (图4L)。图4
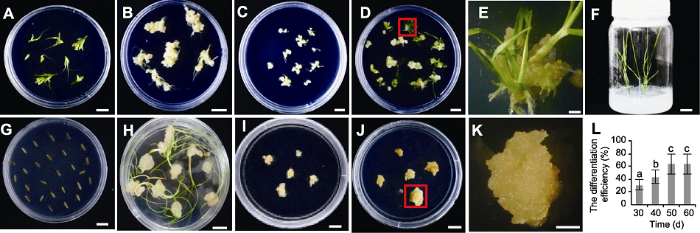 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4川草2号老芒麦组培再生体系建立及幼穗愈伤分化效率随继代时间的变化
(A) 幼穗接种于愈伤诱导培养基; (B) 幼穗诱导35天后的愈伤; (C) 挑选图(B)中结构致密坚硬的愈伤置于继代培养基15天; (D) 图(C)愈伤置于分化培养基约35天; (E) 图(D)中红色框内愈伤分化显微图; (F) 图(D)中分化苗置于生根培养基约30天; (G) 种子接种于愈伤诱导培养基; (H) 种子诱导35天后的愈伤; (I) 挑选图(H)中结构致密坚硬的愈伤置于继代培养基15天; (J) 图(I)愈伤置于分化培养基约35天; (K) 图(J)中红色框内愈伤分化显微图; (L) 不同再生时间下的幼穗愈伤分化效率(误差线为标准误, 样本容量为30, 不同小写字母表示差异显著(P<0.05))。(D), (F), (G), (J) Bars=2 cm; (A)-(C), (E), (H), (I), (K) Bars=1 cm
Figure 4The process diagram of tissue culture and regeneration system and the plot of callus differentiation efficiency of Elymus sibiricus cv. ‘Chuancao No.2’ with subculture time
(A) Inflorescence of ‘Chuancao No.2’ placed in callus induction medium; (B) Callus induced by inflorescence of ‘Chuancao No.2’ for 35 d; (C) The callus with dense and hard structure in figure (B) was selected and placed in medium for 15 d; (D) Callus in figure (C) cultured in differentiation medium for 35 d; (E) The micrograph of callus in red frame in figure (D); (F) Differentiated seedlings in figure (D) placed in rooting medium for 30 d; (G) Seeds placed in callus induction medium; (H) Callus induced by seeds for 35 d; (I) The callus with dense and hard structure in figure (H) was selected and placed in subculture medium for 15 d; (J) Callus in figure (I) cultured in differentiation medium for 35 d; (K) The micrograph of callus in red frame in figure (J); (L) The differentiation efficiency of callus induced by inflorescence at different time periods (The error line is the standard error, and the sample size is 30; Different lowercase letters indicate significant differences at P<0.05). Bars in (D), (F), (G), (J)=2 cm; Bars in (A)-(C), (E), (H), (I), (K)=1 cm
2.3 农杆菌侵染
挑选川草2号老芒麦幼穗诱导出的结构致密愈伤, 采用4种不同侵染方案(表1)进行农杆菌侵染。结果表明, GUS染色均呈阴性(图5A, B), RFP荧光观测也均未观测到红色荧光亮点(图5C, D)。用农杆菌直接侵染幼穗, GUS染色结果均呈阴性(图5E, F), RFP荧光观测均未观测到红色荧光亮点(图5G, H)。以上结果表明, 用农杆菌直接侵染幼穗和用农杆菌侵染幼穗愈伤均未能获得阳性愈伤。图5
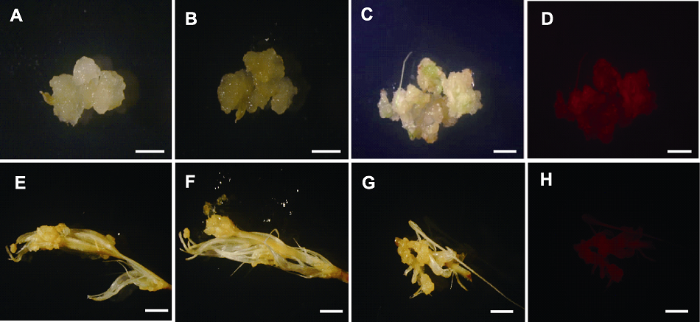 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5根癌农杆菌侵染川草2号老芒麦幼穗和幼穗愈伤
(A), (B) 农杆菌侵染幼穗愈伤GUS染色前后图片; (C), (D) 农杆菌侵染幼穗愈伤RFP荧光下明场和暗场图片; (E), (F) 农杆菌直接侵染幼穗后GUS染色前后图片; (G), (H) 农杆菌直接侵染幼穗后RFP下明场和暗场图片。Bars=50 μm
Figure 5Callus of inflorescence and inflorescence infected by Agrobacterium tumefaciens in Elymus sibiricus cv. ‘Chuancao No.2’
(A), (B) Callus induced by inflorescence before and after GUS staining after Agrobacterium infection; (C), (D) The bright and dark field images of RFP fluorescence after Agrobacterium infection of callus induced by inflorescence; (E), (F) The images before and after GUS staining directly after Agrobacterium infection with inflorescence; (G), (H) The bright and dark field images of RFP fluorescence after Agrobacterium infection of inflorescence, respectively. Bars=50 μm
2.4 基因枪转化
基因枪转化结果(图6A-H)表明, 对于诱导25天的幼穗愈伤, 经高渗处理后, 愈伤被轰击后第4天可观察到红色荧光(图6C, D), 轰击后4-7天荧光愈伤百分比呈上升趋势, 轰击7天后开始下降, 第9天趋于平稳, 荧光愈伤百分比约为20%。对于诱导25天的幼穗愈伤, 经滤纸干燥处理, 愈伤被轰击后第3天可观察到红色荧光(图6E, F), 轰击后3-6天荧光愈伤百分比持续增加, 轰击后6-8天荧光愈伤百分比不再增加, 轰击8天后开始下降, 到第9天趋于稳定, 荧光愈伤百分比约为40%。对于诱导35天的幼穗愈伤, 经滤纸干燥处理, 愈伤被轰击后第2天可观察到红色荧光(图6G, H), 轰击后2-6天荧光愈伤百分比呈上升趋势, 轰击7天后开始下降, 到第9天趋于稳定, 荧光愈伤百分比约为20% (图7)。综上表明, 在其它条件一致的情况下, 滤纸干燥处理相比常规高渗处理荧光愈伤比率高。对于2种预处理方式, 轰击后在一定时间段内(1-10天)其荧光愈伤比率的总趋势均为先增加后减少并最终趋于稳定, 轰击后2-4天可观察到红色荧光, 轰击后6-7天荧光愈伤比率达到高峰, 7-8天后呈减少趋势, 第9天荧光愈伤比率基本稳定。25天的幼穗愈伤对应各时间点荧光愈伤比率均高于35天的幼穗愈伤, 表明靶愈伤的状态是影响基因枪转化效率的关键因素之一, 且幼嫩幼穗愈伤转化效率更高。图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6不同诱导时间和预处理方式下的幼穗愈伤基因枪轰击后的荧光显微图
(A), (B) 25天幼穗愈伤(未进行轰击); (C), (D) 25天幼穗愈伤常规高渗处理; (E), (F) 25天幼穗愈伤滤纸干燥处理2小时; (G), (H) 35天的幼穗愈伤滤纸干燥处理2小时。箭头所示为阳性转化愈伤。Bars=20 μm
Figure 6Fluorescence micrographs of inflorescence callus after bombardment with different induction time and pretreatment methods
(A), (B) Callus of inflorescence on 25 d (no bombardment); (C), (D) Conventional hyperosmotic treatment of inflorescence callus at 25 d; (E), (F) Inflorescence callus filter paper dried for 2 h on 25 d; (G), (H) Inflorescence callus filter paper dried for 2 h on 35 d. Arrows indicate the positive transgenic calli. Bars =20 μm
图7
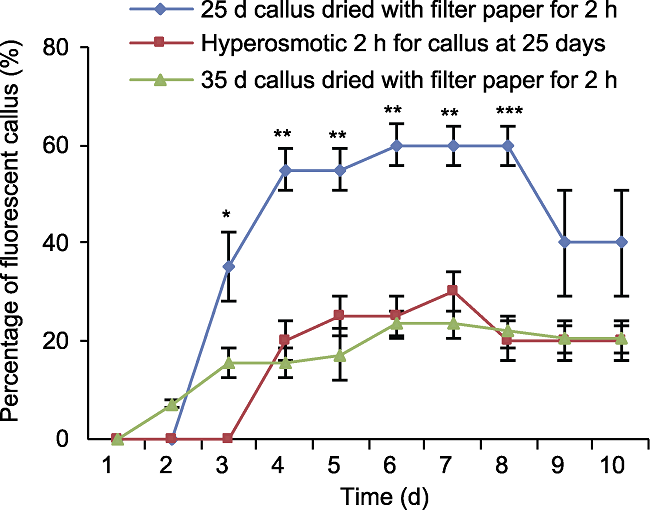 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7基因枪轰击后荧光愈伤百分比随持续观测时间的变化趋势
误差线为标准误差, 样本容量为30。25天幼穗愈伤滤纸干燥2小时荧光愈伤比率均显著高于其它2种处理方式(* P<0.05, ** P<0.01, *** P<0.001)。
Figure 7The trend of percentage of fluorescent callus after gene gun bombardment with continuous observation time
The error line is the standard error, and the sample size is 30. The fluorescence callus ratio of the 25-day inflorescence callus after 2 h drying on filter paper was significantly higher than that of the other two treatments (* P<0.05, ** P<0.01, *** P<0.001).
2.5 讨论
相比双子叶植物, 单子叶植物的遗传转化研究少且起步晚(杨静, 2019)。禾本科牧草愈伤再生困难(Vasil, 1994)是限制其遗传转化体系建立的主要因素。李达旭等(2006)用川草2号老芒麦种子(成熟胚)诱导出愈伤并建立遗传转化体系。本研究团队利用川草2号老芒麦约5万粒种子(成熟胚)诱导愈伤, 出现的愈伤均为海绵状且水渍化严重, 以该类型的愈伤进行分化实验, 均不能再生成完整植株或出现绿点(图4J)。推测实验结果不同是由于老芒麦种子的基因型不同所致。已有研究表明, 基因型是影响愈伤生长速率及分化再生的关键因素之一(Cheng et al., 2004)。禾本科牧草具自交不亲和及异源多倍体等遗传特性, 每粒种子均代表一个基因型(Wang and Ge, 2006; Li et al., 2006)。故来自每一粒种子的愈伤均可能属于不同基因型, 因此极有可能出现不同批次种子愈伤表现不同的情况, 这也是老芒麦遗传转化体系不稳定的主要原因之一。张童(2009)研究表明, 不同组织器官来源的愈伤其分化能力和接受农杆菌侵染的能力也不同。我们通过对川草2号老芒麦大量的成熟胚、下胚轴、幼茎和幼穗等组织的愈伤进行比较, 发现大部分幼穗来源的愈伤组织均可分化, 分化效率最高可达63%, 这与王宇(2014)的研究结果一致, 也与前人认为幼穗是禾本科牧草建立组培再生体系较为理想的外植体候选材料一致(霍秀文等, 2004; 郭夏宇等, 2011; 周妍彤等, 2018)。因此, 与成熟胚愈伤相比, 以幼穗来源的愈伤组织作为受体材料, 较容易建立老芒麦稳定的遗传转化体系。本实验还发现, 老芒麦幼穗愈伤的分化能力随着愈伤继代时间的推移而下降(结果未显示)。因此继续筛选幼穗愈伤, 得到可以继代增殖且分化能力不受影响的幼穗愈伤系是老芒麦遗传转化的关键环节。
单子叶植物不是农杆菌的天然宿主(de Cleene and de Ley, 1976), 因此限制了农杆菌在常规条件下对单子叶植物的遗传转化。目前, 单子叶植物诱导愈伤组织, 虽然可以实现农杆菌对某些禾本科植物的遗传转化, 但整个遗传体系的建立需要花费较长时间, 且往往只针对某一特定品种。本研究对川草2号幼穗愈伤进行农杆菌侵染和基因枪转化, 结果表明, 尽管川草2号幼穗的愈伤可以分化, 但并不能被农杆菌侵染(图5A-D)。基因枪转化实验中, 本研究采取常规高渗和滤纸干燥2种预处理方式。结果表明滤纸干燥处理转化效率高于高渗处理(图7)。用滤纸干燥预处理的转化效率与愈伤状态有较大关系, 25天幼穗愈伤的转化效率高于35天的幼穗(图7)。
本研究通过对川草2号老芒麦成熟种子、根、茎、下胚轴和幼穗5种外植体进行筛选, 确认幼穗愈伤具有较稳定的分化效率, 可以作为转基因的受体材料。农杆菌不能成功侵染本实验室当前获得的老芒麦愈伤系, 利用基因枪转化初步获得了荧光标记的阳性愈伤, 可为进一步建立川草2号老芒麦稳定遗传转化体系奠定基础。
(责任编辑: 朱亚娜)
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3321/j.issn:1000-7091.2004.03.004URL [本文引用: 1]

利用113对SSR多态性引物研究了来源比较广泛的6个爆裂玉米自交系的遗传多样性,并初步进行了杂种优势群划分.结果表明:供试自交系间存在较丰富的SSR多态性;6个爆裂玉米自交系可划分为6个杂种优势群,其中Ⅰ,Ⅴ,Ⅵ类群以国内种质选系为主,Ⅱ,Ⅲ,Ⅳ类群主要为外引杂交种选系,第Ⅰ类群又可分为3个亚群.该分群结果与杂交组合的组配效果相吻合,高优势组合的亲本均属于不同的优势类群(亚群),而在类群(亚群)内未组配出优良组合.SSR标记可以用于研究爆裂玉米自交系的种质基础.
[本文引用: 1]
[本文引用: 4]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
近年来植物转基因技术研究迅速发展,大豆、玉米、油菜和棉花等转基因品种已大面积推广应用,取得巨大的经济和社会效益。小麦是世界上主要的粮食作物,自1992年第一株转基因小麦诞生以来,小麦转基因技术发展较快,为开展小麦分子育种奠定了基础。目前,小麦遗传转化主要采用基因枪法和农杆菌介导法,分别占68.8%和15.9%。研究涉及抗病、抗虫、抗逆、品质改良、提高产量等方面,其中研究较多的为抗病(39.7%)和品质改良(25.6%),部分转基因小麦品系已进行环境释放及生产性试验。本文综述了小麦转基因技术的发展和应用现状,讨论了小麦转基因研究存在的主要问题和今后的发展方向。
URL [本文引用: 1]
阐述了几种禾本科植物离体再生体系研究现状.着重介绍了外植体选择、褐化现象及其防治措施以及离体再生体系建立的主要步骤,最后对离体再生培养技术做了展望.
URL [本文引用: 1]
我国有着极其丰富的老芒麦(Elymus sibiricus)种质资源,这对于今后研究老芒麦种质资源具有非常重要的作用。目前国内外对老芒麦种质资源的研究主要集中在其系统分类和遗传多样性方面,而老芒麦种质资源的育种和遗传完整性方面的研究还不够深入,尤其是遗传完整性方面还未见报道。本研究主要综述了老芒麦种质资源的植物学和生物学特征、遗传多样性、育种等国内外的研究进展,并提出了老芒麦种质资源研究中存在的主要问题,为老芒麦种质资源的进一步研究提供参考依据。
[本文引用: 1]
New York: Springer. pp.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
URLPMID:24576457 [本文引用: 1]
DOI:10.1111/j.1467-7652.2011.00658.xURLPMID:21955653 [本文引用: 3]
Switchgrass (Panicum virgatum L.) is a C4 perennial grass and has been identified as a potential bioenergy crop for cellulosic ethanol because of its rapid growth rate, nutrient use efficiency and widespread distribution throughout North America. The improvement of bioenergy feedstocks is needed to make cellulosic ethanol economically feasible, and genetic engineering of switchgrass is a promising approach towards this goal. A crucial component of creating transgenic switchgrass is having the capability of transforming the explants with DNA sequences of interest using vector constructs. However, there are limited options with the monocot plant vectors currently available. With this in mind, a versatile set of Gateway-compatible destination vectors (termed pANIC) was constructed to be used in monocot plants for transgenic crop improvement. The pANIC vectors can be used for transgene overexpression or RNAi-mediated gene suppression. The pANIC vector set includes vectors that can be utilized for particle bombardment or Agrobacterium-mediated transformation. All the vectors contain (i) a Gateway cassette for overexpression or silencing of the target sequence, (ii) a plant selection cassette and (iii) a visual reporter cassette. The pANIC vector set was functionally validated in switchgrass and rice and allows for high-throughput screening of sequences of interest in other monocot species as well.
[本文引用: 1]
[本文引用: 1]
URLPMID:23929338 [本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
1
2020
... 老芒麦(Elymus sibiricus)又名西伯利亚披碱草(Siberian wildrye), 属于禾本科(Poaceae)小麦族(Triticeae)披碱草属的一种多年生自花授粉、异源四倍体疏丛型旱生牧草.老芒麦在我国主要分布于东北、华北、西北和青藏高原等地区, 是青藏高原地区高寒草甸草原和草甸群落中的优势种和建群种(
1
2002
... 老芒麦(Elymus sibiricus)又名西伯利亚披碱草(Siberian wildrye), 属于禾本科(Poaceae)小麦族(Triticeae)披碱草属的一种多年生自花授粉、异源四倍体疏丛型旱生牧草.老芒麦在我国主要分布于东北、华北、西北和青藏高原等地区, 是青藏高原地区高寒草甸草原和草甸群落中的优势种和建群种(
南荻的组织培养与快速繁殖技术
1
2011
...
冰草属植物组织培养再生体系的建立
1
2004
...
提高多年生人工草地建植当年产草量的研究
1
1999
... 老芒麦(Elymus sibiricus)又名西伯利亚披碱草(Siberian wildrye), 属于禾本科(Poaceae)小麦族(Triticeae)披碱草属的一种多年生自花授粉、异源四倍体疏丛型旱生牧草.老芒麦在我国主要分布于东北、华北、西北和青藏高原等地区, 是青藏高原地区高寒草甸草原和草甸群落中的优势种和建群种(
几种禾本科牧草遗传转化体系的建立和抗虫转基因研究
4
2006
... 老芒麦(Elymus sibiricus)又名西伯利亚披碱草(Siberian wildrye), 属于禾本科(Poaceae)小麦族(Triticeae)披碱草属的一种多年生自花授粉、异源四倍体疏丛型旱生牧草.老芒麦在我国主要分布于东北、华北、西北和青藏高原等地区, 是青藏高原地区高寒草甸草原和草甸群落中的优势种和建群种(
... 目前, 关于老芒麦组培快繁与遗传转化体系建立的报道相对较少.
... 以及
... 相比双子叶植物, 单子叶植物的遗传转化研究少且起步晚(
根癌农杆菌介导转化川草二号老芒麦胚性愈伤组织
0
2006
1
2002
... 用Bio-Rad公司PDS-1000He型基因枪轰击, 参照
几种牧草再生体系和遗传转化体系的优化
2
2014
... 目前, 关于老芒麦组培快繁与遗传转化体系建立的报道相对较少.
...
川草2号老芒麦选育报告
1
1995
... 老芒麦(Elymus sibiricus)又名西伯利亚披碱草(Siberian wildrye), 属于禾本科(Poaceae)小麦族(Triticeae)披碱草属的一种多年生自花授粉、异源四倍体疏丛型旱生牧草.老芒麦在我国主要分布于东北、华北、西北和青藏高原等地区, 是青藏高原地区高寒草甸草原和草甸群落中的优势种和建群种(
氮素形态及其配比对老芒麦生长及生理特性的影响
1
2020
... 老芒麦(Elymus sibiricus)又名西伯利亚披碱草(Siberian wildrye), 属于禾本科(Poaceae)小麦族(Triticeae)披碱草属的一种多年生自花授粉、异源四倍体疏丛型旱生牧草.老芒麦在我国主要分布于东北、华北、西北和青藏高原等地区, 是青藏高原地区高寒草甸草原和草甸群落中的优势种和建群种(
老芒麦遗传多样性及育种研究进展
1
2007
... 老芒麦(Elymus sibiricus)又名西伯利亚披碱草(Siberian wildrye), 属于禾本科(Poaceae)小麦族(Triticeae)披碱草属的一种多年生自花授粉、异源四倍体疏丛型旱生牧草.老芒麦在我国主要分布于东北、华北、西北和青藏高原等地区, 是青藏高原地区高寒草甸草原和草甸群落中的优势种和建群种(
根癌农杆菌介导单子叶植物遗传转化研究进展
1
2019
... 相比双子叶植物, 单子叶植物的遗传转化研究少且起步晚(
小麦转基因技术研究及其应用
1
2010
... 目前, 关于老芒麦组培快繁与遗传转化体系建立的报道相对较少.
禾本科植物离体再生体系研究进展
1
2009
...
老芒麦种质资源的研究进展
1
2011
... 老芒麦(Elymus sibiricus)又名西伯利亚披碱草(Siberian wildrye), 属于禾本科(Poaceae)小麦族(Triticeae)披碱草属的一种多年生自花授粉、异源四倍体疏丛型旱生牧草.老芒麦在我国主要分布于东北、华北、西北和青藏高原等地区, 是青藏高原地区高寒草甸草原和草甸群落中的优势种和建群种(
长穗偃麦草幼穗离体培养高频再生体系的建立
1
2018
...
Advances in New Technology for Targeted Modification of Plant Genomes
1
2015
... 近年来, 随着分子生物技术的发展, 植物的功能基因研究成为热点, 为分子育种奠定了基础, 尤其是CRISPR/Cas9技术的产生, 大大加快了作物分子育种的进程, 如玉米(Zea mays) (
Factors influencing Agrobacterium-mediated transformation of monocotyledonous species
1
2004
... 相比双子叶植物, 单子叶植物的遗传转化研究少且起步晚(
The host range of crown gall
1
1976
... 单子叶植物不是农杆菌的天然宿主(
Efficient genome editing in plants using a CRISPR/Cas system
1
2013
... 近年来, 随着分子生物技术的发展, 植物的功能基因研究成为热点, 为分子育种奠定了基础, 尤其是CRISPR/Cas9技术的产生, 大大加快了作物分子育种的进程, 如玉米(Zea mays) (
Callus induction and plant regeneration from mature seeds of Siberian wildrye grass (Elymus sibiricus L.)
1
2012
... 目前, 关于老芒麦组培快繁与遗传转化体系建立的报道相对较少.
Plant regeneration via somatic embryogenesis of Elymus sibiricus cv. ‘Chuancao No.2’
2
2006
... 目前, 关于老芒麦组培快繁与遗传转化体系建立的报道相对较少.
... 相比双子叶植物, 单子叶植物的遗传转化研究少且起步晚(
Multiplex and homologous recombination-mediated genome editing in Arabidopsis and Nicotiana benthamiana using guide RNA and Cas9
1
2013
... 近年来, 随着分子生物技术的发展, 植物的功能基因研究成为热点, 为分子育种奠定了基础, 尤其是CRISPR/Cas9技术的产生, 大大加快了作物分子育种的进程, 如玉米(Zea mays) (
Targeted mutagenesis in Zea mays using TALENs and the CRISPR/ Cas system
1
2014
... 近年来, 随着分子生物技术的发展, 植物的功能基因研究成为热点, 为分子育种奠定了基础, 尤其是CRISPR/Cas9技术的产生, 大大加快了作物分子育种的进程, 如玉米(Zea mays) (
Gateway-compatible vectors for high-throughput gene functional analysis in switchgrass (Panicum virgatum L.) and other monocot species
3
2012
... 农杆菌和基因枪转化过程中所用载体为PANIC系列(
... 用热激法(
... 用热激法将PANIC6A (
Application of the CRISPR-Cas system for efficient genome engineering in plants
1
2013
... 近年来, 随着分子生物技术的发展, 植物的功能基因研究成为热点, 为分子育种奠定了基础, 尤其是CRISPR/Cas9技术的产生, 大大加快了作物分子育种的进程, 如玉米(Zea mays) (
High salt tolerance potential in wheatgrasses
1
1981
... 老芒麦(Elymus sibiricus)又名西伯利亚披碱草(Siberian wildrye), 属于禾本科(Poaceae)小麦族(Triticeae)披碱草属的一种多年生自花授粉、异源四倍体疏丛型旱生牧草.老芒麦在我国主要分布于东北、华北、西北和青藏高原等地区, 是青藏高原地区高寒草甸草原和草甸群落中的优势种和建群种(
Targeted genome modification of crop plants using a CRISPR- Cas system
2
2013
... 近年来, 随着分子生物技术的发展, 植物的功能基因研究成为热点, 为分子育种奠定了基础, 尤其是CRISPR/Cas9技术的产生, 大大加快了作物分子育种的进程, 如玉米(Zea mays) (
... ) (
Molecular improvement of cereals
1
1994
... 相比双子叶植物, 单子叶植物的遗传转化研究少且起步晚(
Agrobacterium-mediated high efficiency transformation of tall fescue (Festuca arundinacea)
2
2005
... 用热激法(
... 用4种侵染方案侵染后, 分别将愈伤组织碎片和农杆菌轻轻摇动孵育10分钟, 孵育后用无菌滤纸吸干愈伤组织表面菌液并晾干.将愈伤置于用侵染液微湿的无菌滤纸上(每个滤纸上的愈伤组织块数大约等于20个原始完整愈伤组织), 23°C暗培养2-3天.在体式显微镜(SZX16, 日本奥林巴斯公司)下进行红色荧光跟踪观测与拍照(RFP荧光激发块型号为U-RFL-T, 日本奥林巴斯公司), 统计荧光愈伤比率(每皿接种20粒愈伤, 重复3次).GUS染色操作参照
Recent advances in genetic transformation of forage and turf grasses
1
2006
... 相比双子叶植物, 单子叶植物的遗传转化研究少且起步晚(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}