 ,*南京农业大学生命科学学院, 南京 210095
,*南京农业大学生命科学学院, 南京 210095Establishment and Optimization of a Shoot Tip-based Genetic Transformation System for Foxtail Millet
Lan Yang, Ya Liu, Yang Xiang, Xiujuan Sun, Jingwei Yan, Aying Zhang,*College of Life Science, Nanjing Agricultural University, Nanjing 210095, China通讯作者: E-mail:ayzhang@njau.edu.cn
责任编辑: 朱亚娜
收稿日期:2020-07-2接受日期:2020-10-14网络出版日期:2021-01-01
| 基金资助: |
Corresponding authors: *E-mail:ayzhang@njau.edu.cn
Received:2020-07-2Accepted:2020-10-14Online:2021-01-01

摘要
以谷子(Seteria italica)豫谷一号为实验材料, 建立了一套简便、稳定的体外茎尖遗传转化体系。通过根癌农杆菌(Agrobacterium tumefaciens)介导的茎尖转化法, 对转化受体采取不同的处理方式, 待拟转化株长到三叶期后进行PCR鉴定。探明了草丁膦(Basta)喷施处理用于谷子转基因幼苗筛选的最适浓度, 以及2种不同检测方式(直接PCR和喷施Basta+PCR)鉴定转基因植株的效果。在上述基础上, 对影响谷子遗传转化体系的多种因素进行优化。结果表明, 菌液浓度(OD600)=1.4、侵染液中乙酰丁香酮浓度为800 μmol?L -1、侵染压强为0.05 MPa、侵染40分钟有利于谷子茎尖的遗传转化。同时, 采用上述优化系统获得谷子转SiCBL4基因植株, 通过喷施草丁膦和实时荧光定量PCR对T2代转基因植株进行遗传稳定性分析, 可节约检测时间。综上, 该研究初步建立了稳定的谷子体外茎尖遗传转化体系, 并开发了一种便捷的检测后代转基因植株的组合方法。
关键词:
Abstract
In this study, a simple and stable genetic transformation system of foxtail millet (Seteria italica) was established and optimized, in which shoot tips were used as the explant. We transformed Yugu 1, an elite millet cultivar, by Agrobacterium-mediated transformation, and tested different treatments to boost transformation efficiency. We used a PCR-based assay to screen transformants in third-leaf stage seedlings. We determined an optimal lethal concentration of glufosinate (Basta) when sprayed to millet seedlings, and tested the different PCR-based genotyping methods with or without Basta spary. Using the newly established pipeline, we further optimized various crucial factors that affect genetic transformation efficiency. We found that an optimal concentration of bacterial culture was OD600=1.4, an optimal concentration of acetolsyringone was 800 μmol?L -1. We also obtained high transformation efficiency with an infecting pressure at 0.05 MPa, and an infecting time of 40 min. We used the above-mentioned transformation method to transform a Seteria italica calcineurin B-like protein 4 (SiCBL4) overexpression construct. Genetic stability analysis on T2 generation transformed plants was performed by the combination assay of Basta resistance and real-time quantitative fluorescence RT-PCR, which can save the time of genotyping. Altogether, this study establishes a shoot tip-based stable genetic transformation system for foxtail millets, and also develops a robust pipeline to detect transgenic offsprings.
Keywords:
PDF (7785KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
杨澜, 刘雅, 项阳, 孙秀娟, 颜景畏, 张阿英. 谷子茎尖体外遗传转化体系的建立与优化. 植物学报, 2021, 56(1): 71-79 doi:10.11983/CBB20119
Yang Lan, Liu Ya, Xiang Yang, Sun Xiujuan, Yan Jingwei, Zhang Aying.

谷子(Seteria italica)是起源于中国的古老农作物, 在中国北方的驯化栽培历史悠久(刁现民和程汝宏, 2017), 具有抗旱性强、适应性广、营养丰富均衡以及饲草蛋白含量高等特点(李顺国等, 2012; 宋利军, 2019)。近年来, 随着干旱愈发严重、畜牧业大力发展以及人们饮食习惯逐渐偏爱杂粮, 谷子的市场需求量不断增加, 其必将成为农业供给侧结构性改革的重要方向之一(李顺国等, 2014, 2018)。
随着转基因技术的发展, 通过遗传转化等技术提高作物产量和品质已成为一种广泛应用的途径。然而, 与水稻(Oryza sativa)和玉米(Zea mays)等主要农作物相比, 谷子功能基因研究相对滞后, 其中一个主要原因是谷子遗传转化体系发展较为缓慢。因此, 建立并进一步优化谷子遗传转化体系十分重要。
谷子遗传转化的方法有很多, 主要包括基因枪法、农杆菌介导法、PEG介导法、子房注射法和花粉管通道法等(李臻等, 2015)。目前研究最多且使用较广泛的是农杆菌介导法和基因枪法。基因枪法的优点是对宿主没有明显的特定要求, 可将外源基因直接导入受体, 转化各种组织和器官, 缺点是转化率不高, 易导致基因沉默且成本较高。农杆菌介导法不仅能够完整稳定地表达外源基因, 而且操作简单、成本低。通常情况下, 用于农杆菌介导的谷子遗传转化受体材料包括愈伤组织、幼穗、幼胚及成熟胚等(王永芳等, 2003; 陈倩楠等, 2018; 李颜方等, 2019; Sood et al., 2020), 但这些受体材料均需进行组织培养, 获得受体时间较长且有较强的基因型依赖性。因此, 近年来以茎尖生长点为受体进行遗传转化的方法备受关注, 已在玉米(Li et al., 2002; Ma et al., 2018)、水稻(Arockiasamy and Ignacimuthu, 2007; 张笑寒等, 2016)、高粱(Sorghum bicolor) (张明洲等, 2006; Yellisetty et al., 2015)和珍珠粟(Pennisetum glaucum) (Jha et al., 2011)等多种禾本科植物中成功应用。其主要优点是不受季节限制, 可大大缩短育种时间。因此, 本研究以豫谷一号黄化苗裸露出的茎尖为受体材料, 利用农杆菌介导法侵染茎尖, 获得阳性植株, 随后进一步优化侵染条件和检测手段, 旨在建立一套高效稳定的遗传转化体系, 为加快谷子转基因育种进程奠定基础。
1 材料与方法
1.1 材料
供试材料谷子(Seteria italica (L.) Beauv.)品种豫谷一号由本实验室保存。遗传转化所用的大肠杆菌(Escherichia coli)菌株为DH5α、根癌农杆菌(Agrobacterium tumefaciens)选用LBA4404, 质粒为pCUN- NHF, 分别携带Bar基因, 均由本实验室保存。pCUN- NHF质粒T-DNA区如图1所示。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1pCUN-NHF质粒T-DNA区段示意图
LB: LB T-DNA重复; Ubi promoter: 玉米Ubiquitin1启动子; HA: HA标签; MCS: 多克隆位点; 35S promoter: CaMV35S启动子; Bar: 抗草铵膦基因; RB: RB T-DNA重复
Figure 1The T-DNA region of plasmid pCUN-NHF
LB: LB T-DNA repeat; Ubi promoter: Zea mays Ubiquitin1 promoter; HA: HA tag; MCS: Multiple cloning site; 35S promoter: CaMV35S promoter; Bar: Herbicide bialaphos-resistance gene; RB: RB T-DNA repeat
1.2 方法
1.2.1 构建pCUN-NHF-SiCBL4过表达载体以及pCUN-NHF-GUS载体利用一步克隆法(Jeong et al., 2012)构建所需载体。分别以谷子cDNA或pGWB3载体为模板, 利用引物(SiCBL4-F: 5′-taccgtcgacgagctaagcttgATGGGGTGCATACCAACAA-3′, SiCBL4-R: 5′-gggaaattcgagctcggtaccTTACATTTCAGGTTTGGCAGCC-3′; GUS- F: 5′-taccgtcgacgagctaagcttgAACCGACGACTCGTCCGTC-3′, GUS-R: 5′-gggaaattcgagctcggtaccTGTTTGCCTCCCTGCTGC-3′)进行PCR扩增反应, 同时将pCUN-NHF载体线性化, 然后将得到的片段与线性化的载体回收, 利用一步克隆专用酶进行连接并转化, 通过菌落PCR和测序验证载体构建成功。
1.2.2 制备侵染液
利用刘颖慧等(2005)的方法将上述载体分别转入农杆菌LBA4404, 经菌落PCR验证后, 用接种环蘸取部分菌液在含有100 mg?mL-1卡那霉素和50 mg?mL-1利福平的固体LB培养基上划线。培养36-48小时后挑取单菌落接种于5 mL含有相同抗生素的YEP液体培养基中, 30°C摇床过夜培养。然后转接到100 mL YEP液体培养基中, 振荡培养约36小时至OD600值为1.0, 6 400 ×g离心10分钟后收集菌体, 利用重悬液进行重悬, 备用。
1.2.3 初步建立农杆菌介导的谷子茎尖遗传转化体系
参考贺美林(2018)的方法并进行修改。选取健康饱满的谷子种子于水中浸泡, 置于4°C冰箱春化2-3天后, 将种子均匀撒在铺有湿润滤纸的培养皿中, 置于30°C培养箱暗培养约3天, 期间不定期喷水以保证滤纸湿润。待谷子黄化苗茎长到约1 cm, 采取3种不同的方式对转化受体进行处理: (1) 利用刀片在谷子茎尖生长点处呈45°斜切一刀, 去掉胚芽鞘尖端(称之为一刀法); (2) 在谷子茎尖生长点处横竖各切一刀(称之为两刀法); (3) 不需要做切除处理, 直接将整株幼苗进行侵染(称之为未切法)。随后将不同处理方式的谷子置于含400 μmol?L-1乙酰丁香酮的侵染液中, 在0.04 MPa真空压力下侵染20分钟。侵染完成后直接将谷子铺在装有营养基质(营养土:蛭石=1:1, v/v)的盆中, 并在谷子表面铺撒少量的营养基质(营养土:蛭石=1:2, v/v), 以避免根部缺水死亡。置于28°C暗培养1天后取出, 在28°C温室中继续正常培养。
1.2.4 GUS组织化学染色及观察
参考Yan等(2020)的方法进行GUS活性的组织化学检测。将侵染3天的幼苗置于GUS染色液中并抽真空30分钟, 37°C恒温避光染色过夜, 用75%乙醇完成脱色后置于体式显微镜下观察。
1.2.5 转基因植株的PCR鉴定
待谷子长到三叶期, 取新鲜幼嫩的叶片, 利用CTAB法(Dellaporta et al., 1983)提取谷子叶片基因组DNA, 并通过PCR方法检测Bar基因。扩增引物: F: 5′- ATCGAGACAAGCACGGTCAA-3′, R: 5′-AAACCCACGTCATGCCAGTT-3′。扩增程序为: 95°C预变性3分钟; 94°C变性25秒, 52°C退火25秒, 72°C延伸25秒, 33个循环; 72°C延伸5分钟, 4°C保存。取10 μL PCR产物进行琼脂糖凝胶电泳检测。
1.2.6 草丁膦对谷子幼苗最适致死浓度的确定
待野生型谷子长到三叶期, 分别用0、5、12.5、25和50 mg?L-1五种不同浓度的草丁膦(Basta)均匀喷施植株表面, 每个处理约100株苗。7天后统计存活率。存活率=(处理后存活株数/处理前正常生长株数)× 100%。实验重复3次。
1.2.7 单因素试验
分别选取菌液浓度、乙酰丁香酮浓度、侵染压强和侵染时间4个因素设计单因素试验, 考察各因素对谷子遗传转化率的影响。将谷子在0.04 MPa压力下抽真空20分钟。其中, 选取OD600梯度为0.8、1.0、1.2、1.4和1.6; 在相同压强和时间处理下, 乙酰丁香酮浓度梯度设置为0、200、400、600、800和1 000 μmol?L-1; 将谷子置于OD600值为1.0且已添加400 μmol?L-1乙酰丁香酮的菌液中进行侵染, 侵染压强分别设置为0、0.03、0.04、0.05、0.06和0.07 MPa; 在相同菌液及乙酰丁香酮浓度下, 将侵染时间梯度设置为10、20、30、40和50分钟, 分别统计不同侵染条件的转化率。转化率=(阳性植株数/侵染植株数)×100%。实验重复3次。
1.2.8 谷子转基因植株后代遗传稳定性检测
以构建SiCBL4过表达材料为例, 将得到的SiCBL4过表达材料T2代的种子进行播种, 喷施适宜浓度的草丁膦并统计纯合株系。之后随机选取3个株系, 利用Trizol法(冷秋思, 2019)提取三叶期幼苗总RNA, 并反转录成cDNA, 设计引物(F: 5′-GGGGTGCATACCAACAAAGC-3′, R: 5′-AGCCCTTGCTGCTGAAACAT-3′)并利用谷子内参基因β-actin特异引物(F: 5′-GTGCTTTCCCTCTACGCCAGTG-3′, R: 5′-ACCGCTGAGCACAATGTTACCA-3′)进行qRT-PCR (刘宝玲等, 2016), 以检测目的基因的相对表达量。每个反应设置3次重复。
2 结果与讨论
2.1 谷子茎尖遗传转化体系的初步建立
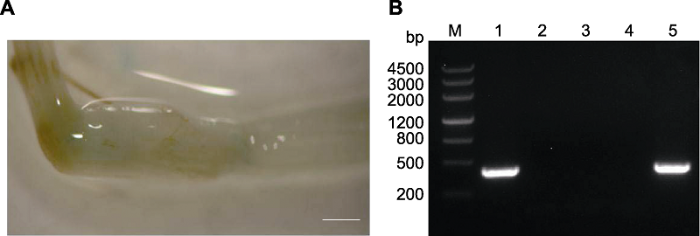
为初步建立农杆菌介导的谷子茎尖遗传转化体系, 分别采取一刀法、两刀法和未切法3种方式处理转化受体, 每种方式各获得至少400株幼苗。侵染3天后, 对其中200株幼苗进行GUS染色。结果(图2A)表明, 采取一刀法和两刀法获得的谷子没有染出蓝色, 而直接利用未切法可以染出蓝色(2.5%, 5株/200株)。随后提取另外200株谷子幼苗(三叶期)的新叶DNA并进行Bar基因检测, 结果(图2B)发现, 采取一刀法和两刀法获得的谷子没有鉴定出Bar基因, 而直接利用未切法虽然转化率较低, 却可以检测到Bar基因(3%, 6株/ 200株), 这与GUS染色结果基本一致。因此我们推测, 造成2种检测方法转化率略有差异的原因可能是: (1) GUS染色不如Bar基因检测灵敏; (2) 可能两组(各200株)本身的转化率就不完全一致。因此可以确定, 当以谷子茎尖为转化受体建立遗传转化体系时, 采取未切法较为合适。其体系为: 制备转化菌液置于30°C培养箱待用, 同时将谷子均匀铺在含有湿润滤纸的培养皿中(图3A), 然后置于30°C培养箱避光处理3天。待谷子茎长到约1 cm (图3B)时, 直接将谷子置于菌液浓度(OD600)为1.0的侵染液中, 并加入400 μmol?L-1乙酰丁香酮, 在0.04 MPa真空压强下处理20分钟。侵染完成后直接将谷子铺在装有营养基质的盆中, 置于28°C暗培养1天后取出, 在28°C温室中继续培养(图3C)。待长到三叶期进行PCR鉴定, 筛选阳性转基因植株。然后将转基因植株移栽到较大的花盆中进行正常生长管理, 经历拔节期(图3D)和抽穗期(图3E)等过程, 3-4个月后得到成熟种子(图3F)。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2谷子转基因植株鉴定
(A) 谷子幼苗GUS染色图(bar=0.1 cm); (B) PCR鉴定(1: 阳性对照; 2: 阴性对照; 3: 切一刀的植株叶片; 4: 切两刀的植株叶片; 5: 不做切除处理的植株叶片)
Figure 2Identification of transgenic foxtail millet
(A) Image from GUS staining of foxtail millet seedlings (bar=0.1 cm); (B) PCR identification (1: Positive control; 2: Negative control; 3: A leaf with one wound cut; 4: A leaf with two wounds cut; 5: A leaf without wound)
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3谷子茎尖遗传转化体系的建立
(A) 待萌发的种子; (B) 萌发3天后的种子; (C) 转化5天后的幼苗; (D) 拔节期植株; (E) 抽穗期植株; (F) 成熟种子。(A), (C), (E) Bars=1.0 cm; (B) Bar=0.5 cm; (D) Bar=10.0 cm; (F) Bar=5.0 cm
Figure 3Establishment of the genetic transformation system for foxtail millet using shoot tip
(A) Seeds to be germinated; (B) Seeds after germination for 3 d; (C) Seedlings after transformation for 5 d; (D) Plants at jointing stage; (E) Plants at heading stage; (F) Mature seeds. (A), (C), (E) Bars=1.0 cm; (B) Bar=0.5 cm; (D) Bar=10.0 cm; (F) Bar=5.0 cm
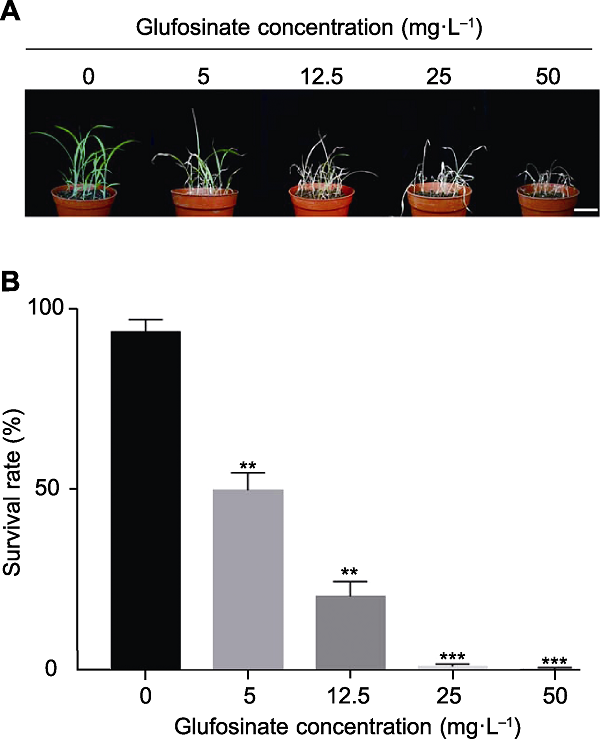
2.2 草丁膦喷施处理谷子最适致死浓度的确定
用不同浓度的草丁膦喷施处理谷子种子, 结果(图4A, B)表明, 随着草丁膦浓度的增加, 野生型谷子存活率急剧下降。当处理浓度为5 mg?L-1时, 存活率约为50%, 致死效果不明显; 当采用12.5 mg?L-1草丁膦喷施野生型谷子后, 其存活率约为20%, 仍较高; 当草丁膦浓度为25和50 mg?L-1时, 处理后的野生型幼苗几乎全部死亡, 且3次重复实验结果无显著差异。考虑到喷施过量草丁膦可能会对阳性植株后续生长造成伤害, 因此将草丁膦喷施处理谷子的最适浓度确定为25 mg?L-1。图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同浓度的草丁膦对谷子幼苗存活率的影响
(A) 不同浓度的草丁膦处理谷子的表型图(bar=5.0 cm); (B) 不同浓度草丁膦处理下谷子的存活率。数据为平均值±标准差(n=3)。Student’s t-test, ** P<0.01, *** P<0.001
Figure 4Effects of glufosinate concentrations on survival rate of foxtail millet seedlings
(A) Phenotypes of foxtail millet seedlings treated with glufosinate at different concentrations (bar=5.0 cm); (B) The survival rate of foxtail millet seedlings treated with different concentrations of glufosinate. Data are means±SD (n=3). Student’s t-test, ** P<0.01, *** P<0.001
2.3 转基因植株的检测
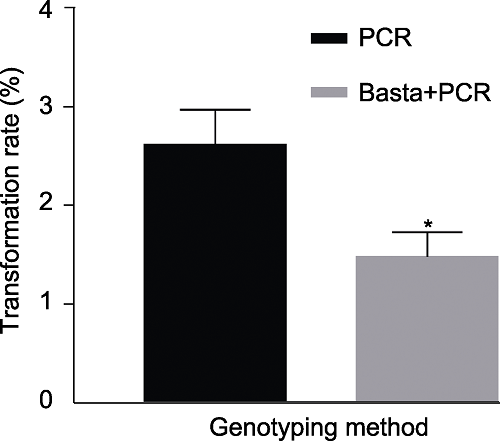
为了优化检测手段, 我们将三叶期的待检测幼苗平均分成两部分, 并采取2种方式进行检测: 取一半幼苗不做处理直接进行PCR检测, 另外一半均匀喷施浓度为25 mg?L-1草丁膦后, 将存活植株进行PCR检测, 统计2种检测方法的转化率(图5), 结果表明, 直接进行PCR检测的转基因植株幼苗的转化率约为2.5%; 采取喷施Basta+PCR组合的方法转化率约为1.5%, 且3次重复结果无显著差异。尽管两种检测方法存在一定差异, 但采取组合方法进行检测, 不仅经济实惠, 而且可以节约时间。因此, 本研究后续实验检测转基因植株均采用喷施Basta+PCR组合的方法。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5两种不同方式检测转基因植株的转化率
数据为平均值±标准差(n=3)。Student’s t-test, * P<0.05
Figure 5Transformation rate obtained using two different methods
Data are means±SD (n=3). Student’s t-test, * P<0.05
2.4 谷子茎尖遗传转化体系的优化
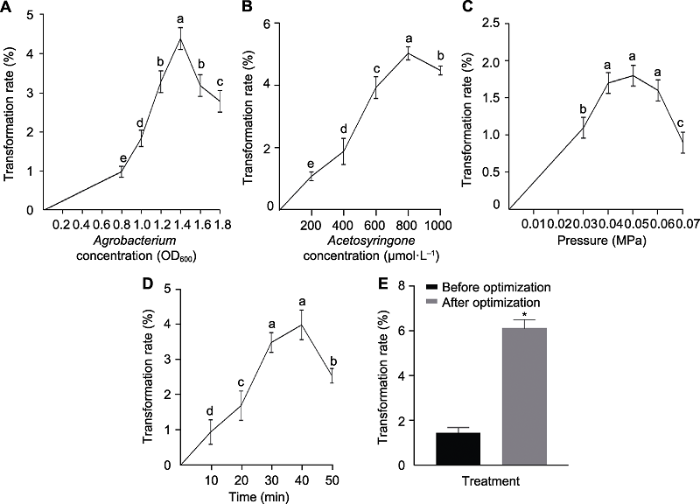
为了研究菌液浓度对谷子遗传转化率的影响, 我们控制其它变量, 对菌液浓度设置了梯度。结果(图6A)表明, 随着农杆菌浓度的升高, 遗传转化率呈现先升后降的趋势, 当菌液OD600=1.4时, 转化率达到最大。农杆菌浓度较低时, 由于菌落数量较少, 导致转化效率较低, 而农杆菌浓度过高, 会因营养供应不足导致菌落活性有所下降, 且3次重复结果无显著差异。因此, 我们确定谷子遗传转化的最适农杆菌浓度为OD600= 1.4。图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6农杆菌浓度(A)、乙酰丁香酮浓度(B)、侵染压强(C)、侵染时间(D)及同时改变4个变量(E)对谷子茎尖转化率的影响
数据为平均值±标准差(n=3)。Student’s t-test, * P<0.05。不同小写字母表示差异显著(P<0.05)。
Figure 6Effects of Agrobacterium concentration (A), acetosyringone concentration (B), infecting pressure (C), infecting time (D) and changing four variables (E) on transformation rates for foxtail millet using shoot tip
Data are means±SD (n=3). Student’s t-test, * P<0.05. Different lowercase letters indicat significant differences (P<0.05).
为了研究乙酰丁香酮浓度对谷子遗传转化率的影响, 我们控制其它变量, 对乙酰丁香酮浓度设置了梯度。结果(图6B)表明, 乙酰丁香酮的添加对谷子遗传转化效果的影响非常显著。在不添加乙酰丁香酮的情况下, 转化率为0; 在乙酰丁香酮浓度较低时, 随着浓度的增加, 转化率不断提高; 当乙酰丁香酮浓度为800 μmol?L-1时, 转化率达到最高(5%), 之后乙酰丁香酮的浓度继续增加, 转化率反而出现下降趋势, 且3次重复结果无显著差异。因此, 我们确定添加乙酰丁香酮的最佳浓度为800 μmol?L-1。
为了研究真空压强对谷子遗传转化率的影响, 我们控制其它变量, 对真空压强设置了不同梯度。结果(图6C)表明, 当压强处于0?0.04 MPa时, 随着压强的升高, 转化率逐渐提高, 当压强达到0.04 MPa时, 转化率暂时达到稳定。在压强为0.04、0.05和0.06 MPa 时转化率较高, 分别为1.7%、1.8%和1.6%, 三者相差较小; 之后压强继续升高, 转化率下降幅度更大,且3次重复结果无显著差异。因此, 综合考虑, 最佳压强确定为0.05 MPa。
为了研究侵染时间对谷子遗传转化率的影响, 我们控制其它变量, 对侵染时间设置了不同梯度。结果(图6D)表明, 转化率随着侵染时间的延长呈现先上升后下降趋势, 当侵染时间为40分钟时, 转化率最高(4%), 且3次重复结果无显著差异。因此, 侵染时间确定为40分钟。
由上述单因素试验确定了影响谷子遗传转化效率4个因素的最佳条件, 为得到最优的遗传转化体系, 与之前初步建立的体系条件进行了对比(表1)。利用优化前的条件进行侵染转化率约为1.5%, 而采用单因素试验优化后的条件转化率约6.2% (图6E)。因此, 优化菌液浓度、乙酰丁香酮浓度、真空压强及处理时间4个因素后, 遗传转化率得到明显提升。
Table 1
表1
表1初步建立的体系和优化后的体系条件对比
Table 1
| Agrobacterium concentration (OD600) | Acetosyringone concentration (μmol?L-1) | Pressure (MPa) | Time (min) | |
|---|---|---|---|---|
| Before optimization | 1.0 | 400 | 0.04 | 20 |
| After optimization | 1.4 | 800 | 0.05 | 40 |
新窗口打开|下载CSV
2.5 谷子转化体系的遗传稳定性分析
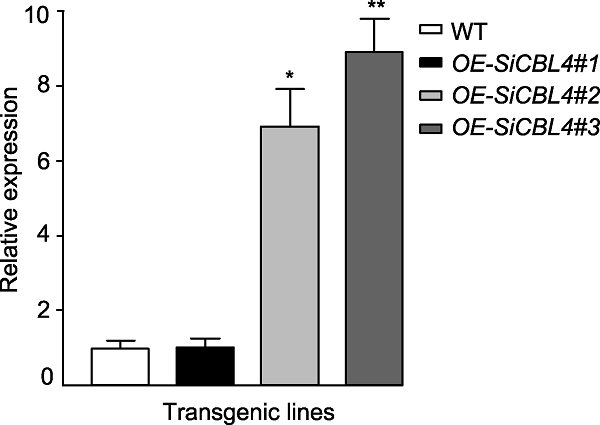
为了检测转化体系的遗传稳定性, 利用该体系构建了SiCBL4基因过表达材料, 共获得18株T0代谷子。进一步鉴定发现, 只有12株T1代谷子叶片中可以检测到Bar基因, 我们进一步通过鉴定获得了T2代转基因植株。首先, 对T2代转基因植株进行抗草丁膦分析, 将25 mg?L-1草丁膦均匀喷施于植株表面, 可以获得表现出抗性的纯合植株。随后利用qRT-PCR分析T2代转基因植株中SiCBL4的相对表达量(图7), 结果表明, SiCBL4过表达株系#2和#3中SiCBL4表达量均明显高于野生型, 说明SiCBL4基因已经在转基因植株中成功过表达。研究结果进一步证实SiCBL4基因可以在谷子转基因后代中稳定表达。图7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7qRT-PCR分析谷子转基因植株中SiCBL4基因表达量
数据为平均值±标准差(n=3)。Student’s t-test, * P<0.05, ** P<0.01
Figure 7qRT-PCR analysis of the expression of SiCBL4 in transgenic foxtail millet
Data are means±SD (n=3). Student’s t-test, * P<0.05, ** P<0.01
2.6 讨论
在谷子遗传转化中, 受体材料的选择一直是重要障碍。目前谷子遗传转化的受体主要有幼穗、幼胚、成熟胚及其愈伤组织等(王永芳等, 2003; 陈倩楠等, 2018; 李颜方等, 2019; Sood et al., 2020; Zhao et al., 2020)。幼穗的取材受季节及谷子生长阶段影响很大; 而谷子幼胚体积非常小, 操作受限; 成熟胚诱导愈伤组织再生能力较弱且差异大。现阶段谷子的遗传转化研究侧重于愈伤组织的诱导及再生组培体系的优化, 虽然可以提高转化率, 但再生率低、操作复杂及耗时较长的问题仍未得到解决。本研究以谷子黄化苗裸露的茎尖为转化受体, 无需切除胚芽鞘尖端, 直接利用农杆菌进行侵染。与以其它组织为受体相比, 该方法不仅不受季节和基因型的影响, 而且还可以缩短时间、节约组织培养的成本。由于茎尖转化法必须依赖生殖途径才有可能获得稳定遗传的材料, 因此, 为确定该方法的遗传稳定性, 利用此体系构建了SiCBL4基因过表达材料, 并通过草丁膦筛选和qRT-PCR验证了SiCBL4基因已经成功在转基因植株中稳定表达。
目前有关遗传转化后代植株的鉴定有多种方式。李明浩等(2010)利用了PCR扩增; 张园等(2020)通过PCR检测与GUS组织化学染色进行鉴定; 刘颖慧等(2005)采用分子检测(PCR扩增和Southern杂交)和GUS组织化学染色结合的方法。由于实验误差等原因, 无论是GUS染色、分子鉴定还是草丁膦喷施, 单独使用都可能会存在一定的假阳性, 采取几种技术相结合的方法可使检测结果更可靠, 并节约后代检测及筛选转基因植株的时间; 而采用分子与GUS相结合的鉴定方法, 大量筛选转基因植株时过程太烦琐。本研究利用喷施Basta+PCR组合检测的方法进行转基因植株鉴定, 先通过喷施草丁膦进行初步筛选, 存活下来的植株再进行PCR检测, 这样不仅进一步降低了假阳性出现的概率, 还可以避免提取大量DNA的复杂过程, 大大节约时间。
我们的研究发现, 一刀法和两刀法都未能成功获得转基因植株。原因可能是谷子幼苗较小, 难以准确找到茎尖生长点进行切除; 其次, 采取两刀法会对植株造成较大伤害, 影响其正常生长。因此, 本研究只采用未切法构建遗传转化体系。随后, 我们通过控制单一变量, 从菌液浓度、乙酰丁香酮浓度、压强和侵染时间4个方面优化侵染体系, 得到最佳侵染条件: 在谷子黄化苗茎长约1 cm时, 直接置于菌液浓度(OD600)为1.4的侵染液中, 并添加800 μmol?L-1乙酰丁香酮, 在0.05 MPa压强下侵染40分钟。后续研究中, 我们会进一步摸索两种方法(一刀法和两刀法)的具体条件, 以比较与本文所用方法效率的差别。
(责任编辑: 朱亚娜)
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s00299-007-0377-9URLPMID:17593368 [本文引用: 1]

We have established a reproducible procedure for transformation of shoot apices and regeneration of transgenic plants for two indica rice cultivars, white ponni (WP) and Pusa Basmathi 1 (PB 1). Four-day-old shoot apex explants were transformed by cocultivation with Agrobacterium tumefaciens strain EHA 101 harbouring a binary plasmid pRIT1. The vector contained an improved hygromycin phosphotransferase (hpt) gene for hygromycin resistance driven by actin 1 promoter and the reporter gene beta-glucuronidase intron (INT-GUS) controlled by CaMV 35S promoter. Rice shoots were induced on media containing 0.1 mg/l napthalene acetic acid (NAA), 1.0 mg/l kinetin (kn), 1.0 mg/l N(6)-benzyleaminopurin (BAP), 300 mg/l casaminoacid, 500 mg/l proline, 50 mg/l hygromycin and 500 mg/l cefotaxime. Transgenic plants were raised in pots and seeds were collected. Histochemical and polymerase chain reaction (PCR) analyses of field established transgenic rice plants and their offsprings confirmed the presence of GUS gene. Integration of T-DNA into the genome of putative transgenics was further confirmed by southern analysis. The transformation efficiency of WP was found to be ranging from 5.6 to 6.2% whereas in the case of PB1, it was from 7 to 8%. Progeny analysis of these plants showed a pattern of classical Mendelian inheritance for both hpt and GUS gene.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
以抗除草剂Bar基因稳定转化谷子技术研究
2
2018
... 谷子遗传转化的方法有很多, 主要包括基因枪法、农杆菌介导法、PEG介导法、子房注射法和花粉管通道法等(
... 在谷子遗传转化中, 受体材料的选择一直是重要障碍.目前谷子遗传转化的受体主要有幼穗、幼胚、成熟胚及其愈伤组织等(
十五年区试数据分析展示谷子糜子育种现状
1
2017
... 谷子(Seteria italica)是起源于中国的古老农作物, 在中国北方的驯化栽培历史悠久(
谷子成熟胚再生体系建立及EPSPS基因遗传转化研究
1
2018
... 参考
绿绒蒿属植物不同RNA提取方式的比较分析
1
2019
... 以构建SiCBL4过表达材料为例, 将得到的SiCBL4过表达材料T2代的种子进行播种, 喷施适宜浓度的草丁膦并统计纯合株系.之后随机选取3个株系, 利用Trizol法(
普通小麦品种Alondra's遗传转化体系的建立
1
2010
... 目前有关遗传转化后代植株的鉴定有多种方式.
新时期中国谷子产业发展技术需求与展望
1
2018
... 谷子(Seteria italica)是起源于中国的古老农作物, 在中国北方的驯化栽培历史悠久(
我国谷子产业现状、发展趋势及对策建议
1
2014
... 谷子(Seteria italica)是起源于中国的古老农作物, 在中国北方的驯化栽培历史悠久(
河北省谷子产业现状和技术需求及发展对策
1
2012
... 谷子(Seteria italica)是起源于中国的古老农作物, 在中国北方的驯化栽培历史悠久(
农杆菌介导谷子成熟胚遗传转化体系的建立与优化
2
2019
... 谷子遗传转化的方法有很多, 主要包括基因枪法、农杆菌介导法、PEG介导法、子房注射法和花粉管通道法等(
... 在谷子遗传转化中, 受体材料的选择一直是重要障碍.目前谷子遗传转化的受体主要有幼穗、幼胚、成熟胚及其愈伤组织等(
谷子遗传转化体系研究进展
1
2015
... 谷子遗传转化的方法有很多, 主要包括基因枪法、农杆菌介导法、PEG介导法、子房注射法和花粉管通道法等(
谷子bZIP转录因子的全基因组鉴定及其在干旱和盐胁迫下的表达分析
1
2016
... 以构建SiCBL4过表达材料为例, 将得到的SiCBL4过表达材料T2代的种子进行播种, 喷施适宜浓度的草丁膦并统计纯合株系.之后随机选取3个株系, 利用Trizol法(
根癌农杆菌介导谷子的遗传转化
2
2005
... 利用
... 目前有关遗传转化后代植株的鉴定有多种方式.
谷子高产栽培技术分析
1
2019
... 谷子(Seteria italica)是起源于中国的古老农作物, 在中国北方的驯化栽培历史悠久(
根癌农杆菌共培养转化谷子技术体系的建立
2
2003
... 谷子遗传转化的方法有很多, 主要包括基因枪法、农杆菌介导法、PEG介导法、子房注射法和花粉管通道法等(
... 在谷子遗传转化中, 受体材料的选择一直是重要障碍.目前谷子遗传转化的受体主要有幼穗、幼胚、成熟胚及其愈伤组织等(
高粱茎尖再生体系及其遗传转化影响因子的研究
1
2006
... 谷子遗传转化的方法有很多, 主要包括基因枪法、农杆菌介导法、PEG介导法、子房注射法和花粉管通道法等(
农杆菌介导McCHIT1基因遗传转化水稻茎尖研究
1
2016
... 谷子遗传转化的方法有很多, 主要包括基因枪法、农杆菌介导法、PEG介导法、子房注射法和花粉管通道法等(
芦笋茎尖遗传转化体系的建立与优化
1
2020
... 目前有关遗传转化后代植株的鉴定有多种方式.
Regeneration of transgenic plants from two indica rice (Oryza sativa L.) cultivars using shoot apex explants
1
2007
... 谷子遗传转化的方法有很多, 主要包括基因枪法、农杆菌介导法、PEG介导法、子房注射法和花粉管通道法等(
A plant DNA minipreparation: version II
1
1983
... 待谷子长到三叶期, 取新鲜幼嫩的叶片, 利用CTAB法(
One-step sequence- and ligation-in- dependent cloning as a rapid and versatile cloning method for functional genomics studies
1
2012
... 利用一步克隆法(
Efficient Agrobacterium-mediated transformation of Pennisetum glaucum (L.) R. Br. using shoot apices as explant source
1
2011
... 谷子遗传转化的方法有很多, 主要包括基因枪法、农杆菌介导法、PEG介导法、子房注射法和花粉管通道法等(
Developmental, tissue culture, and genotypic factors affecting plant regeneration from shoot apical meristems of germinated Zea mays L. seedlings
1
2002
... 谷子遗传转化的方法有很多, 主要包括基因枪法、农杆菌介导法、PEG介导法、子房注射法和花粉管通道法等(
ZmbZIP4 contributes to stress resistance in maize by regulating ABA synthesis and root development
1
2018
... 谷子遗传转化的方法有很多, 主要包括基因枪法、农杆菌介导法、PEG介导法、子房注射法和花粉管通道法等(
An efficient Agrobacterium-mediated genetic transformation method for foxtail millet (Setaria italica L.)
2
2020
... 谷子遗传转化的方法有很多, 主要包括基因枪法、农杆菌介导法、PEG介导法、子房注射法和花粉管通道法等(
... 在谷子遗传转化中, 受体材料的选择一直是重要障碍.目前谷子遗传转化的受体主要有幼穗、幼胚、成熟胚及其愈伤组织等(
Abscisic acid positively regulates L-arabinose metabolism to inhibit seed germination through ABSCISIC ACID INSENSITIVE4-mediated transcriptional promotions of MUR4 in Arabidopsis thaliana
1
2020
... 参考
In planta transformation of sorghum (Sorghum bicolor (L.) Moench) using TPS1 gene for enhancing tolerance to abiotic stresses
1
2015
... 谷子遗传转化的方法有很多, 主要包括基因枪法、农杆菌介导法、PEG介导法、子房注射法和花粉管通道法等(
DROOPY LEAF 1 controls leaf architecture by orchestrating early brassinosteroid signaling
1
2020
... 在谷子遗传转化中, 受体材料的选择一直是重要障碍.目前谷子遗传转化的受体主要有幼穗、幼胚、成熟胚及其愈伤组织等(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}