 ,1,*
,1,*Optimization of the Regeneration System from Somatic Embryogenesis in Larix olgensis
Jianfei Liu1, Yan Liu1, Kejian Liu2, Yang Chi3, Zhifa Huo3, Yonghong Huo3, Xiangling You,1,*通讯作者: E-mail:185064633@qq.com
责任编辑: 朱亚娜
收稿日期:2020-02-26接受日期:2020-06-5网络出版日期:2020-09-01
| 基金资助: |
Corresponding authors: E-mail:185064633@qq.com
Received:2020-02-26Accepted:2020-06-5Online:2020-09-01

摘要
以长白落叶松(Larix olgensis)未成熟合子胚为外植体诱导胚性愈伤组织, 通过调节影响体胚发生的营养物质和植物生长调节剂配比, 进行愈伤组织的胚性恢复与保持以及体胚发生再生体系的优化。结果表明: 不同无性系之间胚性愈伤组织诱导率差异显著, 胚性愈伤组织在S+0.2 mg·L -1NAA+0.5 mg·L -1BA+0.5 mg·L -1KT+0.5 g·L -1谷氨酰胺+0.5 g·L -1水解酪蛋白+30 g·L -1蔗糖及3.0 g·L -1植物凝胶培养条件下, 可以恢复胚性并长久保持。在S+20 mg·L -1ABA+60 g·L -1PEG4000+60 g·L -1蔗糖及3.0 g·L -1植物凝胶条件下分化培养6周, 体胚发生率可达100%。将正常发育的体胚先在WPM+ 6 mg·L -1间苯三酚+1.0 g·L -1活性炭+3.0 mg·L -1VB1+20 g·L -1蔗糖及3.0 g·L -1植物凝胶条件下培养2周, 再转接至B5+ 0.4 mg·L -1NAA+1.0 mg·L -1IBA+0.5 mg·L -1GA3+2.0 mg·L -1VB1+1.0 g·L -1活性炭+20 g·L -1蔗糖及3.0 g·L -1植物凝胶条件下培养2周, 可见子叶舒展、下胚轴伸长且根系正常的体胚苗。该研究建立了长白落叶松胚性愈伤组织胚性恢复与保持方法, 并进一步优化了体胚发生的植株再生体系, 为林木资源快速繁育和遗传改良奠定了基础。
关键词:
Abstract
In this study, immature zygotic embryos of Larix olgensis were used as explants to induce embryogenic callus and optimize the regeneration system from somatic embryogenesis. The rejuvenation and preservation of the embryogenic potential of embryogenic callus, the somatic embryogenesis and plant regeneration were investigated through adjusting the nutrition and plant growth regulator. The results showed that the generation rates of embryogenic callus were significantly different among different lines. Under the conditions of S+0.2 mg·L -1NAA+0.5 mg·L -1BA+0.5 mg·L -1KT+0.5 g·L -1glutamine+0.5 g·L -1hydrolyzed casein+30 g·L -1sucrose and 3.0 g·L -1vegetable gel, the embryogenic potential of embryonic callus could be recovered and maintained for a long time. Somatic embryogenesis were induced from embryonic callus cultured in S+20 mg·L -1ABA+60 g·L -1PEG4000+60 g·L -1sucrose and 3.0 g·L -1vegetable gel for 6 weeks, and the generation rate of somatic embryo reached 100%. The normal somatic embryos were first cultured for 2 weeks under the conditions of WPM+6 mg·L -1phloglucinol+1.0 g·L -1active carbon+3.0 mg·L -1VB1+20 g·L -1sucrose and 3.0 g·L -1vegetable gel, and then transferred to B5+0.4 mg·L -1NAA+1.0 mg·L -1IBA+0.5 mg·L -1GA3+2.0 mg·L -1VB1+1.0 g·L -1active carbon+20 g·L -1sucrose and 3.0 g·L -1vegetable gel. After 2 weeks, somatic embryo plantlets with cotyledon stretch, hypocotyl elongation and normal root system were observed. This study established a method for the recovery and maintenance of embryogenic callus from larch, and further optimized the somatic embryogenic pathway, which will lay a foundation for the rapid breeding and genetic improvement of L. olgensis.
Keywords:
PDF (1755KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
刘建飞, 刘炎, 刘克俭, 池阳, 霍志发, 霍永洪, 由香玲. 长白落叶松体胚发生再生体系优化. 植物学报, 2020, 55(5): 605-612 doi:10.11983/CBB20030
Liu Jianfei, Liu Yan, Liu Kejian, Chi Yang, Huo Zhifa, Huo Yonghong, You Xiangling.

体细胞胚胎发生(简称体胚发生)是一项重要的植物生物技术, 是指在离体培养条件下, 体细胞经过类似胚胎发育的过程, 完成体细胞向胚胎发生途径转变的重编程, 最终形成完整植株(Yang and Zhang, 2010)。该过程是植物发育中的独特现象, 是植物细胞表达全能性的经典方式。由于裸子植物为无性繁殖, 相对于有性繁殖较困难, 因此该技术广泛应用于针叶树的无性繁殖。Chalupa等(1976)最早在班克松(Pinus banksiana)离体培养中发现类胚结构, 开启了针叶树体胚发生途径的研究。之后, Hakman等(1985)利用挪威云杉(Picea abies)的未成熟合子胚诱导出胚性愈伤组织, 经分化培养获得体胚苗, 取得了云杉属体胚发生的突破。Durzan和Gupta (1987)以花旗松(Pseudotsuga menziesii)的未成熟合子胚为外植体诱导出体胚并再生出完整植株, 取得了松属体胚发生的突破。据不完全统计, 目前已有6科14属针叶树在无性系体胚发生繁育技术上取得重大进展(Klimaszewska et al., 2016)。
落叶松(Larix gmelinii)是中国东北地区荒山造林和森林更新的重要速生树种。20世纪80年代, 欧洲率先建立落叶松属体胚发生再生体系。von Aderkas等(1987)以欧洲落叶松(L. decidua)的雌配子体为外植体, 首次诱导出胚性愈伤组织并在分化培养基中发生体胚, 但未见再生植株。Klimaszewska (1989)以日本×欧洲落叶松(L. leptolepis × L. decidua)的未成熟合子胚为材料, 诱导出胚性愈伤组织, 经分化培养后体胚发生, 并获得再生植株。美西落叶松(L. occidentalis) (Thompson and von Aderkas, 1992)、日本×高代落叶松(L. leptolepis × L. eurolepis) (Lelu et al., 1994)、北美落叶松(L. laricina) (Klimaszewska et al., 1997)、日本落叶松(L. kaempferi) (Kim et al., 1998)、华北落叶松(L. principis-rupprechtii) (齐力旺, 2000)、日本×长白; 日本×兴安杂种落叶松(L. kaempferi × L. olgensis; L. kaempferi × L. gemlinii) (王伟达等, 2009)、西伯利亚落叶松(L. sibirica) (Tret’yakova and Barsukova, 2012)以及长白落叶松(L. olgensis) (宋跃等, 2016)等落叶松属树种, 相继利用其未成熟合子胚诱导出胚性愈伤组织, 并建立了体胚发生途径。但随着增殖继代次数的增加遇到了胚性愈伤组织的胚性下降、消失及体胚苗生根难等问题, 严重制约了落叶松的植株再生和遗传改良。
在长白落叶松体胚发生过程中, 我们也发现了愈伤组织胚性下降问题。通过调节植物生长调节剂配比来恢复和维持愈伤组织的胚性; 同时阐明影响体胚发生的营养和植物生长调节剂等因素, 旨在进一步优化体胚发生再生体系, 为长白落叶松组培快繁和遗传改良奠定基础。
1 植物材料
2017年7月9日从吉林省四平市林木种子园中选取4棵长白落叶松(Larix olgensis Henry) (L061、L091、L058和L024)优树, 从优树上获得未成熟合子胚。首先用75%乙醇消毒3分钟, 灭菌水清洗3次; 然后用5% NaClO消毒10分钟, 灭菌水清洗3次。2 培养基成分与培养条件
2.1 胚性愈伤组织诱导
诱导培养条件为S (齐力旺, 2000)+1.0 mg·L-1 2,4-D+ 0.5 mg·L-1 BA+0.5 mg·L-1 KT+0.5 g·L-1谷氨酰胺+ 0.5 g·L-1水解酪蛋白+30 g·L-1蔗糖及3.0 g·L-1植物凝胶(pH6.0±0.2, 下同)。用灭菌滤纸吸干种子表面的水分, 将未成熟合子胚接种在诱导培养基上, 每50 mL的锥形瓶中(含20 mL培养基)接种7个外植体, 切口向下贴近培养基并做好标记, 于温室暗培养, 温度为(25±2)°C。期间观察外植体的结构、颜色及形态等特征变化, 培养10天时统计各无性系愈伤组织的诱导率。2.2 胚性愈伤组织的胚性恢复与保持
将上述诱导出的胚性愈伤组织增殖培养(S+0.1 mg·L-1 2,4-D+0.04 mg·L-1 BA+0.02 mg·L-1 KT+0.5 g·L-1谷氨酰胺+0.5 g·L-1水解酪蛋白+30 g·L-1蔗糖及3.0 g·L-1植物凝胶) 6个月, 体胚分化率急剧下降。我们采取去除2,4-D、逐渐降低NAA浓度以及提高细胞分裂素浓度的措施来恢复与保持愈伤组织的胚性。在BA和KT浓度升高到0.5 mg·L-1, NAA浓度从0.5、0.4、0.3、0.2、0.1 mg·L-1依次降低的增殖培养基中进行胚性恢复培养。待每个浓度梯度的胚性愈伤组织可以继代时, 一部分转入下一个浓度梯度, 另一部分转入体胚发生培养基, 根据体胚发生率来确定恢复愈伤组织胚性的最佳NAA浓度。2.3 体胚诱导
将上述恢复胚性的愈伤组织接入分化培养基(S+ 15 mg·L-1 ABA+80 g·L-1 PEG4000+80 g·L-1蔗糖+ 5 g·L-1 AgNO3+0.4 g·L-1谷氨酰胺+0.2 g·L-1水解酪蛋白+3.0 g·L-1植物凝胶)中进行暗培养。本研究主要从影响体胚发生的关键因素包括ABA浓度(10- 30 mg·L-1)、PEG4000浓度(20-80 g·L-1)以及蔗糖和麦芽糖浓度(20-80 g·L-1)进行体胚发生的优化。2.4 体胚萌发及壮苗
体胚萌发的基本培养基: WPM+6 mg·L-1间苯三酚+ 1.0 g·L-1活性炭+3.0 mg·L-1 VB1+20 g·L-1蔗糖及3.0 g·L-1植物凝胶。促进体胚苗生根和下胚轴生长的培养基: B5+2.0 mg·L-1 VB1+1.0 g·L-1活性炭+20 g·L-1蔗糖及3.0 g·L-1植物凝胶。本研究主要利用植物生长调节剂IBA、NAA和GA3对体胚萌发进行优化。将萌发状态良好的体胚苗先后接入含有0.4-0.8 mg·L-1 NAA、0.5-1.5 mg·L-1 IBA、0-1.5 mg·L-1 GA3的上述B5培养基中, 4周后统计成苗率、茎高及主根长度。体胚萌发时所用培养瓶为100 mL三角瓶, 内装50 mL培养基, 每次24瓶, 每瓶接种5个子叶胚; 生根优化所用培养瓶为广口瓶, 内装100 mL培养基, 每个处理10瓶, 每瓶接种6个早期体胚苗。培养环境均为组培室光照培养, 光周期为16小时光照/8小时黑暗, 光照强度为50 μmol·m-2·s-1, 温度为(25±2)°C。2.5 数据处理
愈伤组织诱导率=(诱导出愈伤组织的接种数/接种总数)×100%; 体胚发生率=(发生体胚的接种数/接种总数)×100%; 体胚萌发率=(萌发的体胚数/接种总数)× 100%; 生根率=(生根的体胚数/接种总数)×100%; 成苗率=(生根长茎的体胚数/接种总数)×100%。采用软件IBM SPSS Statistics19.0对数据进行方差分析, 用Duncan法进行多重比较(0.05检测水平)。利用GraphPad Prism6.0软件作图。3 结果与讨论
3.1 胚性愈伤组织诱导
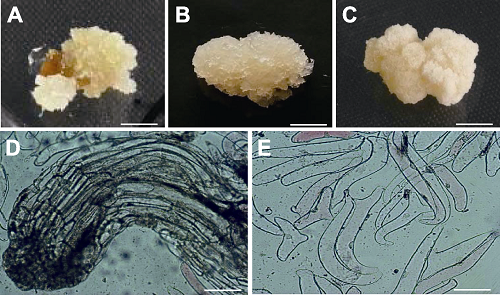
将未成熟合子胚接种到诱导培养基上, 1周后可见浅黄色愈伤组织(图1A)。该愈伤组织在继代过程中逐渐分化为2类愈伤组织, 一类是白色透明、质感较软、稍带黏性、表面呈细丝状的愈伤组织(图1B), 经光学显微镜观测, 愈伤组织内有呈穗条状排列紧密的细胞团(图1D), 经成熟培养后体胚发生, 为胚性愈伤组织; 另一类是质硬、黏稠、表面有棉花絮状的雪白色愈伤组织(图1C), 愈伤组织内有呈短棒状排列疏松的细胞(图1E), 经分化培养后体胚未发生, 为非胚性愈伤组织。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1长白落叶松愈伤组织及镜检结构
(A) 愈伤组织; (B) 胚性愈伤组织; (C) 非胚性愈伤组织; (D) 胚性愈伤组织显微图像; (E) 非胚性愈伤组织显微图像。 (A)- (C) Bars=3 mm; (D) Bar=200 μm; (E) Bar=500 μm
Figure 1Callus and its microscopic images of Larix olgensis
(A) Callus; (B) Embryogenic callus; (C) Non-embryogenic callus; (D) The image of embryogenic callus; (E) The image of non-embryogenic callus. (A)-(C) Bars=3 mm; (D) Bar= 200 μm; (E) Bar=500 μm
同一时间采集的未成熟合子胚, 不同无性系之间愈伤组织诱导率无显著差异(均在80%以上) (表1)。胚性愈伤组织诱导率差异显著, 4个无性系中只有058- 842号优树诱导出胚性愈伤组织(诱导率为4.5%), 其余3个无性系均未诱导出胚性愈伤组织。
Table 1
表1
表1不同长白落叶松株系愈伤组织与胚性愈伤组织诱导(平均值±标准差)
Table 1
| Excellent trees | Callus induction rate (%) | Embryogenic callus formation rate (%) |
|---|---|---|
| 061-1009 | 89.43±48.59 a | 0 |
| 091-1008 | 81.56±47.91 c | 0 |
| 058-842 | 91.25±53.17 a | 4.53±2.73 a |
| 024-922 | 85.91±37.29 b | 0 |
新窗口打开|下载CSV
3.2 胚性愈伤组织继代过程中胚性恢复和保持
将上述诱导的胚性愈伤组织进行增殖培养(S+0.1 mg·L-1 2,4-D+0.04 mg·L-1 BA+0.02 mg·L-1 KT+ 0.5 g·L-1谷氨酰胺+0.5 g·L-1水解酪蛋白+30 g·L-1蔗糖及3.0 g·L-1植物凝胶)。继代6个月后, 发现胚性愈伤组织的体胚分化率从75.4%急剧下降至45.3%。提高BA (0.5 mg·L-1)和KT (0.5 mg·L-1)的浓度, 用NAA替代2,4-D, 并逐渐降低NAA浓度, 然后统计体胚发生率(表2)。结果表明, 与对照组相比, 用NAA代替2,4-D, 同时提高BA和KT的浓度, 可以明显提高胚性愈伤组织的体胚发生率和体胚数量。NAA浓度在0.5-0.1 mg·L?1范围内, 随着NAA浓度的逐渐递减, 体胚发生率呈现先缓慢提高再降低的趋势, 体胚数量也先增多后减少。当NAA浓度为0.2 mg·L-1时胚性愈伤组织胚性恢复效果最好, 体胚发生率高达72.4%, 体胚数量最多达48.6个·g-1。目前, 胚性愈伤组织在S+0.2 mg·L-1 NAA+0.5 mg·L-1 BA+0.5 mg·L-1 KT+ 0.5 g·L-1谷氨酰胺+0.5 g·L-1水解酪蛋白+30 g·L-1蔗糖及3.0 g·L-1植物凝胶培养条件下继代胚性依然很好。Table 2
表2
表2不同浓度配比的植物生长调节剂对长白落叶松体胚发生率和体胚数量的影响(平均值±标准差)
Table 2
| The combination of plant growth regulators in the medium | Somatic embryogenesis (%) | Somatic embryo amount (numbers·g-1) |
|---|---|---|
| 0.1 mg?L-1 2,4-D+0.04 mg·L-1 BA+0.02 mg·L-1 KT | 45.27±23.31 | 16.47±1.67 |
| 0.5 mg?L-1 NAA+0.5 mg·L-1 BA+0.5 mg·L-1 KT | 52.65±31.83 b | 25.31±9.85 c |
| 0.4 mg?L-1 NAA+0.5 mg·L-1 BA+0.5 mg·L-1 KT | 57.38±15.41 ab | 31.53±5.37 b |
| 0.3 mg?L-1 NAA+0.5 mg·L-1 BA+0.5 mg·L-1 KT | 64.41±11.79 a | 39.74±4.29 ab |
| 0.2 mg?L-1 NAA+0.5 mg·L-1 BA+0.5 mg·L-1 KT | 72.36±10.57 a | 48.56±3.77 a |
| 0.1 mg?L-1 NAA+0.5 mg·L-1 BA+0.5 mg·L-1 KT | 63.70±43.86 a | 41.92±7.22 a |
新窗口打开|下载CSV
3.3 体胚诱导
将胚性愈伤组织接种到分化培养基培养2周后, 可见胚性愈伤组织略有生长, 表面布满白色细丝棒状凸起(图2A), 胚性细胞团逐渐黏稠; 培养4周, 愈伤组织表面大部分的白色细丝棒状逐渐消失, 出现浅黄色圆柱棒状凸起的早期子叶胚(图2B); 培养6周, 以上浅黄色棒状凸起逐渐伸长(图2C), 此时已经发育至子叶胚期。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2长白落叶松体胚成熟过程
(A) 培养2周; (B) 培养4周; (C) 培养6周。Bars=2.5 mm
Figure 2Maturation of somatic embryos of Larix olgensis
(A) Cultured for 2 weeks; (B) Cultured for 4 weeks; (C) Cultured for 6 weeks. Bars=2.5 mm
3.3.1 ABA处理对愈伤组织体胚发生的影响
将胚性愈伤组织分别接种到添加10、15、20、25和30 mg·L-1 ABA的分化培养基中, 诱导体胚发生。结果(图3A)表明, 不同ABA浓度梯度下体胚发生率差异显著。ABA浓度在10-20 mg·L-1范围内, 体胚发生率均高于60%; 当ABA浓度为20 mg·L-1时, 体胚发生率最高达84.9%; ABA浓度在25-30 mg·L-1范围内, 体胚发生率均低于50%。因此, 20 mg·L-1 ABA为胚性愈伤组织体胚发生的最适浓度。
图3
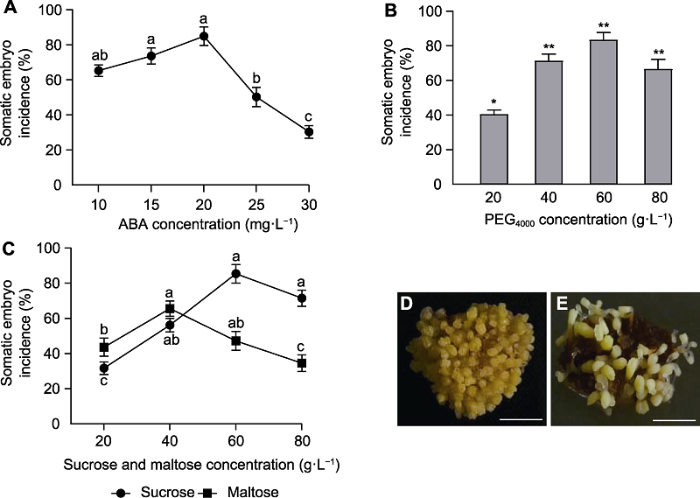 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同因素对长白落叶松体胚发生率的影响(平均值±标准误)
(A) ABA; (B) PEG4000; (C) 蔗糖和麦芽糖; (D) 添加蔗糖后培养6周; (E) 添加麦芽糖后培养6周。 (D), (E) Bars=2.5 mm。不同小写字母表示差异显著(P<0.05)。* P<0.05; ** P<0.01
Figure 3Effects of different factors on the rate of somatic embryogenesis in Larix olgensis (means±SE)
(A) ABA; (B) PEG4000; (C) Sucrose and maltose; (D) Cultured with sucrose for 6 weeks; (E) Cultured with maltose for 6 weeks. (D), (E) Bars=2.5 mm. Different lowercase letters indicate significant differences (P<0.05). * P<0.05; ** P<0.01
3.3.2 PEG4000处理对愈伤组织体胚发生的影响
在20 mg·L-1 ABA条件下, 将胚性愈伤组织分别接种到添加20、40、60和80 g·L-1 PEG4000的分化培养基中, 诱导体胚发生。结果(图3B)表明, 在分化培养基中添加不同浓度的PEG4000, 愈伤组织体胚发生率差异显著。当PEG4000浓度为40-80 g·L-1时, 体胚发生率均高于60%; 当PEG4000浓度为60 g·L-1时, 体胚发生率最高, 达83.4%。因此, 60 g·L-1 PEG4000为胚性愈伤组织体胚发生的最适浓度。
3.3.3 碳源处理对愈伤组织体胚发生的影响
在含有20 mg·L-1 ABA和60 g·L-1 PEG4000的分化培养基中分别添加20、40、60、80 g·L-1蔗糖和麦芽糖后培养6周, 诱导体胚发生。结果(图3C)表明, 碳源浓度在20-80 g·L-1范围内, 体胚发生率均随着蔗糖和麦芽糖浓度的升高先增加后降低, 但转折点不同。当蔗糖含量为60 g·L-1时, 体胚发生率最高(85.4%); 而麦芽糖浓度为40 g·L-1时, 体胚发生率最高为65.6%, 比添加蔗糖体胚发生率低近20%, 表明60 g·L-1蔗糖为体胚发生最适浓度。通过对比发现, 尽管添加蔗糖的分化培养基中体胚发生率高于添加麦芽糖, 且体胚发生数量多(图3D), 但不如添加麦芽糖的体胚发生质量高(图3E), 而高质量的体胚有利于后期健壮体胚苗的产生。
3.4 体胚萌发及壮苗
从分化培养基上选取发育状态良好、长度(约1 cm)一致的子叶胚(图4A)转接到萌发培养基, 2周后可见子叶膨大伸展, 下胚轴略有伸长, 胚柄类似物完全消失, 原始主根系透明发亮, 为早期体胚苗(图4B)。将早期体胚苗转接入B5培养基2周后, 对照组的早期体胚苗表现为根部褐化、下胚轴伸长及子叶生长稍舒展(图4C); 优化后的早期体胚苗表现为根部正常、下胚轴伸长及子叶生长舒展(图4D)。从B5培养基中选取粗壮的体胚苗(图4E), 剔除表面的培养基, 移栽至含有黑土:蛭石:草木灰=3:1:1 (v/v/v)等营养物质混匀的组培土里(图4F), 用少量B5液体培养基浇灌, 使土壤营养更充分。图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4长白落叶松不同发育状态的体胚苗及其移栽
(A) 子叶胚; (B) 子叶胚萌发; (C) 对照体胚苗; (D) 优化体胚苗; (E) 生根体胚苗; (F) 移栽体胚苗。 (A) Bar=1.5 mm; (B) Bar=6.5 mm; (C), (D) Bars=1.0 cm; (E), (F) Bars=1.5 cm
Figure 4Somatic embryo seedlings at different developmental states of Larix olgensis and their transplanting
(A) Cotyledonary embryo; (B) Cotyledons germinate; (C) Control of somatic embryo seedling; (D) Optimization somatic embryo seedling; (E) Rooting somatic embryo seedlings; (F) Transplanting somatic embryo seedling. (A) Bar=1.5 mm; (B) Bar=6.5 mm; (C), (D) Bars=1.0 cm; (E), (F) Bars=1.5 cm
3.4.1 IBA和NAA联合处理对体胚苗生长的影响
当NAA浓度为0.4 mg·L-1时, 一定范围内随着IBA浓度的增加, 成苗率、茎高和主根长呈现先增加后减少趋势; 当IBA浓度为1.0 mg·L-1时, 成苗率最高(46.2%), 茎高最高(1.8 mm), 主根最长(19.2 mm) (表3)。当NAA浓度为0.8 mg·L-1时, 一定范围内随着IBA浓度的增加, 主根长呈现先增加后降低趋势, 成苗率和茎高基本保持不变; 当IBA浓度为1.0 mg·L-1时, 主根最长(15.2 mm), 表明NAA浓度为0.8 mg·L-1时, IBA浓度变化对成苗率和茎高2个指标影响不明显。综上, 0.4 mg·L-1 NAA搭配1.0 mg·L-1 IBA在茎高这一指标上与0.8 mg·L-1 NAA搭配1.0 mg·L-1 IBA相差甚微, 但成苗率和主根长这2个指标明显优于0.8 mg·L-1 NAA组合1.0 mg·L-1 IBA。由此确定0.4 mg·L-1 NAA和1.0 mg·L-1 IBA联合作用时体胚苗生长效果最好。
Table 3
表3
表3生长调节剂组合对长白落叶松体胚苗生长的影响(平均值±标准差)
Table 3
| Growth regulator combination | Somatic embryo seedlings | |||
|---|---|---|---|---|
| NAA (mg·L-1) | IBA (mg·L-1) | Seedling rate (%) | Stem length (mm) | Main root length (mm) |
| 0.4 | 0.5 | 38.83±29.38 ab | 1.59±1.44 c | 16.38±12.74 b |
| 0.4 | 1.0 | 46.21±41.29 a | 1.75±0.77 ab | 19.17±14.52 a |
| 0.4 | 1.5 | 35.37±33.34 b | 1.67±1.75 b | 12.60±10.34 c |
| 0.8 | 0.5 | 22.57±25.43 c | 1.76±0.91 ab | 12.31±9.54 c |
| 0.8 | 1.0 | 22.65±15.83 c | 1.81±0.88 a | 15.18±13.37 bc |
| 0.8 | 1.5 | 21.43±19.68 c | 1.78±0.67 ab | 11.40±8.53 c |
新窗口打开|下载CSV
3.4.2 GA3处理对体胚苗生长的影响
我们选择体胚苗生长最优培养基(添加0.4 mg·L-1 NAA和1.0 mg·L-1 IBA), 发现在一定范围内随着GA3浓度的上调, 成苗率和茎高均呈现先上升后下降趋势(表4)。当GA3浓度为0.5 mg·L-1时, 成苗率最高(55.4%), 茎高最高(2.7 mm), 但随着GA3浓度进一步提高, 主根长度受到严重影响, 主根平均长度从18.0 mm降至8.7 mm。综合表明, 植株再生时选择0.5 mg·L-1 GA3、0.4 mg·L-1 NAA以及1.0 mg·L-1 IBA联合作用对体胚苗生长更有效。
Table 4
表4
表4不同浓度GA3对长白落叶松体胚苗生长的影响(平均值±标准差)
Table 4
| Plant regeneration | GA3 concentration (mg·L-1) | ||
|---|---|---|---|
| 0 | 0.5 | 1.0 | |
| Seedling rate (%) | 45.61±52.27 b | 55.44±37.91 a | 45.91±35.81 b |
| Stem length (mm) | 1.63±1.33 b | 2.65±2.61 a | 0.91±1.11 c |
| Main root length (mm) | 17.98±14.22 a | 12.41±16.55 b | 8.65±5.49 c |
新窗口打开|下载CSV
3.5 讨论
针叶树胚性愈伤组织的诱导培养基中多含有高浓度的生长素和细胞分裂素, 而增殖继代多采用低浓度的生长素和细胞分裂素(Stasolla and Yeung, 2003)。但胚性愈伤组织长期在含有2,4-D的增殖培养基中继代会导致胚性下降, 体胚发生率降低(Tautorus et al., 1991)。本研究将用2,4-D诱导后胚性下降的长白落叶松的胚性愈伤组织, 在用NAA代替2,4-D并持续降低NAA浓度至0.2 mg·L-1, 同时提高细胞分裂素(BA和KT)浓度至0.5 mg·L-1条件下继代, 可长久维持胚性愈伤组织的分化能力, 极大地提高了胚性愈伤组织的增殖效率及体胚发生率。齐力旺(2000)用0.5 mg·L-1 NAA代替2,4-D提高了华北落叶松体胚发生数量, 但未能抑制体胚发生率的下降。这可能与增殖继代过程中胚性愈伤组织内激素积累有关(Pla?ková et al., 2015)。有的落叶松树种利用较低浓度2,4-D进行诱导, 也可保持愈伤组织的胚性, 如杂种落叶松(2,4-D浓度为0.5 mg·L-1, BA和KT浓度为0.5 mg·L-1)和长白落叶松(2,4-D浓度为0.3 mg·L-1, BA和KT浓度为0.1 mg·L-1) (王伟达等, 2009; 宋跃等, 2016), 而黑云杉(P. mariana)仅用3.6 μmol·L-1 BA便可维持其胚性(Li et al., 2019)。
针叶树体胚成熟过程中ABA和渗透调节剂是必备条件。研究表明, 目前影响针叶树体胚发生的主要因素是ABA、蔗糖和PEG4000浓度。ABA能够有效调控处于增殖状态的胚性愈伤组织向成熟状态发育, 抑制裂生多胚现象发生, 阻止胚胎过早发育, 促进后期原胚的成熟(Nunes et al., 2018)。同时, 外源ABA能协调内源ABA共同参与体胚发生蛋白合成通路, 为体胚发生作铺垫(Klimaszewska and Smith, 1997)。目前, 针叶树中松属体胚成熟需要的ABA浓度范围为10-150 μmol·L-1 (Montalbán et al., 2010), 落叶松属多集中在15-135 μmol·L-1。本研究设定的ABA浓度范围为35-115 μmol·L-1, 在这一范围内确定的ABA最佳浓度与华北落叶松相近(齐力旺, 2000)。在体胚成熟过程中添加一定量的渗透调节剂也十分必要。渗透调节剂可以辅助ABA, 阻止体胚提前萌发变绿, 使体胚缓慢充分发育, 有利于提高体胚的品质(Lulsdorf et al., 1993)。PEG4000和蔗糖的作用是利用高渗胁迫使材料脱水, 从而使胚性愈伤组织处于一种向分化方向过渡的生理状态(Attree et al., 1992)。此外, 蔗糖还为胚性愈伤组织增殖提供碳源。本研究表明, 在一定范围内, 体胚发生率会随着ABA、蔗糖和PEG4000浓度的增加呈现先升高后降低趋势。最佳分化培养条件为: S+20 mg·L-1 ABA+60 g·L-1 PEG4000+60 mg·L-1蔗糖。用相同浓度的麦芽糖代替蔗糖, 体细胞胚胎发生质量明显提高, 但体胚数量有所下降。这一结果与对华北落叶松的研究结果一致(齐力旺, 2000)。此外, 也有研究表明, 在分化培养基中添加适量的IBA和活性炭, 以及在培养环境中添加适量CO2, 均有利于增加成熟胚的数量, 促进产生高质量的子叶胚(Roberts et al., 1990; Kvaalen and von Arnold, 1991; 黄健秋等, 1995)。
本研究建立了一种胚性愈伤组织胚性恢复与长久保持的方法, 进一步优化了长白落叶松体胚发生途径, 可为林木资源快速繁育和遗传改良奠定基础。
(责任编辑: 朱亚娜)
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 5]
[本文引用: 2]
[本文引用: 2]
DOI:10.1007/BF00195664URLPMID:24178081 [本文引用: 1]

In order to enhance post-germinative vigour, somatic embryos of Picea glauca (Moench) Voss. were matured under in-vitro conditions that stimulated triacylglycerol (TAG) biosynthesis. In P. glauca seeds over 90% of the TAG was stored within the megagametophyte, and isolated zygotic embryos contained twice the amount of TAG of somatic embryos cultured for four weeks on basal medium containing 16 muM abscisic acid (ABA). Polyethylene glycol-4000 (PEG) as a non-permeating osmoticum with ABA promoted TAG biosynthesis by somatic embryos and sustained maturation throughout an eight-week culture period. Treatments that promoted TAG biosynthesis also prevented precocious germination and promoted desiccation tolerance. Thus, the optimal culture conditions for maturation, desiccation survival, and plantlet regeneration were 16-24 muM ABA and 7.5% PEG for eight weeks, followed by desiccation. Under these conditions the levels of TAG per somatic embryo were raised ninefold to about five times the zygotic-embryo level, and the TAG fatty-acid composition became similar to that of zygotic embryos. A study of sectioned material, using light and transmission electron microscopy, showed that the structure and distribution of lipid bodies within these somatic embryos and the degree of embryo development were similar to mature zygotic embryos. Up to 81% of the desiccated somatic embryos regenerated to plantlets during which time the TAG was utilised in a manner similar to zygotic seedlings.
DOI:10.1139/b76-043URL [本文引用: 1]
DOI:10.1016/0168-9452(87)90056-2URL [本文引用: 1]
DOI:10.1016/0168-9452(85)90079-2URL [本文引用: 1]
DOI:10.1023/A:1006120302512URL [本文引用: 1]
DOI:10.1016/0168-9452(89)90105-2URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/ppl.1997.100.issue-4URL [本文引用: 1]
DOI:10.1007/BF00048206URL [本文引用: 1]
DOI:10.1139/x94-015URL [本文引用: 1]
DOI:10.1007/s00468-018-1793-xURL [本文引用: 1]
DOI:10.1007/BF00234697URLPMID:24197337 [本文引用: 1]
Interior spruce (Picea glauca engelmannii complex) and black spruce (Picea mariana Mill.) cotyledonary somatic embryos were encapsulated in sodium alginate. Somatic embryo viability was retained, but germination occurred at a reduced frequency compared with the equivalent zygotic embryos. The addition of 0.5% (w/v) activated charcoal to the alginate capsule significantly enhanced root development and germination for somatic embryos but not for zygotic embryos. The possibility of developing an artiflcal endosperm was also investigated, by addition of Litvay (Litvay et al. 1981) nutrients with or without 90 mM sucrose to the alginate-charcoal capsule. This treatment significantly enhanced root development for all embryo categories with the exception of black spruce somatic embryos. Encapsulated and non-encapsulated somatic embryos survived one month cold storage at 4 degrees C without reduction in germination frequency.
DOI:10.1007/s00468-010-0477-yURL [本文引用: 1]
DOI:10.1007/s11240-017-1311-7URL [本文引用: 1]
DOI:10.1007/s10725-015-0044-zURL [本文引用: 1]
DOI:10.1139/b90-239URL [本文引用: 1]
DOI:10.1023/A:1023345803336URL [本文引用: 1]
DOI:10.1139/b91-237URL [本文引用: 1]
DOI:10.1007/BF00234365URLPMID:24201537 [本文引用: 1]
Somatic embryogenesis was initiated from immature embryos of western larch (Larix occidentalis Nutt.) on media containing 2,4-dichlorophenoxyacetic acid and N6- benzyladenine. The effects of explant type and ammonium nitrate and glutamine concentrations on initiation were tested. Although 21-93% of explants rendered cultures in various experiments, only 3% yielded sustainable embryogenic lines. Excised embryos at the early cotyledonary stage were optimal for initiation. Maturation of somatic embryos was promoted by abscisic acid. Response to abscisic acid concentrations and duration of exposure to abscisic acid varied with genotype. Maximal results were obtained with 0.025 mu M abscisic acid for 1 to 2 weeks followed by individual culture on medium without growth regulators. Mature somatic embryos developed into shoots with roots. Plantlets have been established in peat.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
马尾松成熟合子胚的体细胞胚胎发生和植株再生
1
1995
... 针叶树体胚成熟过程中ABA和渗透调节剂是必备条件.研究表明, 目前影响针叶树体胚发生的主要因素是ABA、蔗糖和PEG4000浓度.ABA能够有效调控处于增殖状态的胚性愈伤组织向成熟状态发育, 抑制裂生多胚现象发生, 阻止胚胎过早发育, 促进后期原胚的成熟(
华北落叶松体细胞胚胎发生与遗传转化系统建立的研究
5
2000
... 落叶松(Larix gmelinii)是中国东北地区荒山造林和森林更新的重要速生树种.20世纪80年代, 欧洲率先建立落叶松属体胚发生再生体系.
... 诱导培养条件为S (
... 本研究将用2,4-D诱导后胚性下降的长白落叶松的胚性愈伤组织, 在用NAA代替2,4-D并持续降低NAA浓度至0.2 mg·L-1, 同时提高细胞分裂素(BA和KT)浓度至0.5 mg·L-1条件下继代, 可长久维持胚性愈伤组织的分化能力, 极大地提高了胚性愈伤组织的增殖效率及体胚发生率.
... 针叶树体胚成熟过程中ABA和渗透调节剂是必备条件.研究表明, 目前影响针叶树体胚发生的主要因素是ABA、蔗糖和PEG4000浓度.ABA能够有效调控处于增殖状态的胚性愈伤组织向成熟状态发育, 抑制裂生多胚现象发生, 阻止胚胎过早发育, 促进后期原胚的成熟(
... 蔗糖.用相同浓度的麦芽糖代替蔗糖, 体细胞胚胎发生质量明显提高, 但体胚数量有所下降.这一结果与对华北落叶松的研究结果一致(
长白落叶松胚性愈伤组织诱导及体细胞胚胎发生
2
2016
... 落叶松(Larix gmelinii)是中国东北地区荒山造林和森林更新的重要速生树种.20世纪80年代, 欧洲率先建立落叶松属体胚发生再生体系.
... 本研究将用2,4-D诱导后胚性下降的长白落叶松的胚性愈伤组织, 在用NAA代替2,4-D并持续降低NAA浓度至0.2 mg·L-1, 同时提高细胞分裂素(BA和KT)浓度至0.5 mg·L-1条件下继代, 可长久维持胚性愈伤组织的分化能力, 极大地提高了胚性愈伤组织的增殖效率及体胚发生率.
杂种落叶松未成熟胚的体细胞胚发生和植株再生
2
2009
... 落叶松(Larix gmelinii)是中国东北地区荒山造林和森林更新的重要速生树种.20世纪80年代, 欧洲率先建立落叶松属体胚发生再生体系.
... 本研究将用2,4-D诱导后胚性下降的长白落叶松的胚性愈伤组织, 在用NAA代替2,4-D并持续降低NAA浓度至0.2 mg·L-1, 同时提高细胞分裂素(BA和KT)浓度至0.5 mg·L-1条件下继代, 可长久维持胚性愈伤组织的分化能力, 极大地提高了胚性愈伤组织的增殖效率及体胚发生率.
Manipulation of conditions for the culture of somatic embryos of white spruce for improved triacylglycerol biosynthesis and desiccation tolerance
1
1992
... 针叶树体胚成熟过程中ABA和渗透调节剂是必备条件.研究表明, 目前影响针叶树体胚发生的主要因素是ABA、蔗糖和PEG4000浓度.ABA能够有效调控处于增殖状态的胚性愈伤组织向成熟状态发育, 抑制裂生多胚现象发生, 阻止胚胎过早发育, 促进后期原胚的成熟(
Growth and metabolism of cells and tissue of Jack pine ( Pinus banksiana). 2. The quantitative analysis of the growth of callus from hypocotyls and radicles
1
1976
... 体细胞胚胎发生(简称体胚发生)是一项重要的植物生物技术, 是指在离体培养条件下, 体细胞经过类似胚胎发育的过程, 完成体细胞向胚胎发生途径转变的重编程, 最终形成完整植株(
Somatic embryogenesis and polyembryogenesis in Douglas-fir cell suspension cultures
1
1987
... 体细胞胚胎发生(简称体胚发生)是一项重要的植物生物技术, 是指在离体培养条件下, 体细胞经过类似胚胎发育的过程, 完成体细胞向胚胎发生途径转变的重编程, 最终形成完整植株(
The development of somatic embryos in tissue cultures initiated from immature embryos of Picea abies (Norway Spruce)
1
1985
... 体细胞胚胎发生(简称体胚发生)是一项重要的植物生物技术, 是指在离体培养条件下, 体细胞经过类似胚胎发育的过程, 完成体细胞向胚胎发生途径转变的重编程, 最终形成完整植株(
Somatic embryogenesis and plant regeneration from immature zygotic embryos of Japanese larch (Larix leptolepis)
1
1998
... 落叶松(Larix gmelinii)是中国东北地区荒山造林和森林更新的重要速生树种.20世纪80年代, 欧洲率先建立落叶松属体胚发生再生体系.
Plantlet development from immature zygotic embryos of hybrid larch through somatic embryogenesis
1
1989
... 落叶松(Larix gmelinii)是中国东北地区荒山造林和森林更新的重要速生树种.20世纪80年代, 欧洲率先建立落叶松属体胚发生再生体系.
Larix laricinar (tamarack): somatic embryogenesis and genetic transformation
1
1997
... 落叶松(Larix gmelinii)是中国东北地区荒山造林和森林更新的重要速生树种.20世纪80年代, 欧洲率先建立落叶松属体胚发生再生体系.
1
2016
... 体细胞胚胎发生(简称体胚发生)是一项重要的植物生物技术, 是指在离体培养条件下, 体细胞经过类似胚胎发育的过程, 完成体细胞向胚胎发生途径转变的重编程, 最终形成完整植株(
Maturation of somatic embryos of Pinus strobus is promoted by a high concentration of gellan gum
1
1997
... 针叶树体胚成熟过程中ABA和渗透调节剂是必备条件.研究表明, 目前影响针叶树体胚发生的主要因素是ABA、蔗糖和PEG4000浓度.ABA能够有效调控处于增殖状态的胚性愈伤组织向成熟状态发育, 抑制裂生多胚现象发生, 阻止胚胎过早发育, 促进后期原胚的成熟(
Effects of various partial pressures of oxygen and carbon dioxide on different stages of somatic embryogenesis in Picea abies
1
1991
... 针叶树体胚成熟过程中ABA和渗透调节剂是必备条件.研究表明, 目前影响针叶树体胚发生的主要因素是ABA、蔗糖和PEG4000浓度.ABA能够有效调控处于增殖状态的胚性愈伤组织向成熟状态发育, 抑制裂生多胚现象发生, 阻止胚胎过早发育, 促进后期原胚的成熟(
Somatic embryogenesis from immature and mature zygotic embryos and from cotyledons and needles of somatic plantlets of Larix
1
1994
... 落叶松(Larix gmelinii)是中国东北地区荒山造林和森林更新的重要速生树种.20世纪80年代, 欧洲率先建立落叶松属体胚发生再生体系.
Dynamics of physiological and miRNA changes after long-term proliferation in somatic embryogenesis of Picea balfouriana
1
2019
... 本研究将用2,4-D诱导后胚性下降的长白落叶松的胚性愈伤组织, 在用NAA代替2,4-D并持续降低NAA浓度至0.2 mg·L-1, 同时提高细胞分裂素(BA和KT)浓度至0.5 mg·L-1条件下继代, 可长久维持胚性愈伤组织的分化能力, 极大地提高了胚性愈伤组织的增殖效率及体胚发生率.
Germination of encapsulated embryos of interior spruce (Picea glauca engelmannii complex) and black spruce (Picea mariana Mill.)
1
1993
... 针叶树体胚成熟过程中ABA和渗透调节剂是必备条件.研究表明, 目前影响针叶树体胚发生的主要因素是ABA、蔗糖和PEG4000浓度.ABA能够有效调控处于增殖状态的胚性愈伤组织向成熟状态发育, 抑制裂生多胚现象发生, 阻止胚胎过早发育, 促进后期原胚的成熟(
Bottlenecks in Pinus radiata somatic embryogenesis: improving maturation and germination
1
2010
... 针叶树体胚成熟过程中ABA和渗透调节剂是必备条件.研究表明, 目前影响针叶树体胚发生的主要因素是ABA、蔗糖和PEG4000浓度.ABA能够有效调控处于增殖状态的胚性愈伤组织向成熟状态发育, 抑制裂生多胚现象发生, 阻止胚胎过早发育, 促进后期原胚的成熟(
Somatic embryogenesis of hybrid Pinus elliottii var. elliottii × P. caribaea var. hondurensis and ploidy assessment of somatic plants
1
2018
... 针叶树体胚成熟过程中ABA和渗透调节剂是必备条件.研究表明, 目前影响针叶树体胚发生的主要因素是ABA、蔗糖和PEG4000浓度.ABA能够有效调控处于增殖状态的胚性愈伤组织向成熟状态发育, 抑制裂生多胚现象发生, 阻止胚胎过早发育, 促进后期原胚的成熟(
Cytokinin profiling of long-term in vitro pea (Pisum sativum L.) shoot cultures
1
2015
... 本研究将用2,4-D诱导后胚性下降的长白落叶松的胚性愈伤组织, 在用NAA代替2,4-D并持续降低NAA浓度至0.2 mg·L-1, 同时提高细胞分裂素(BA和KT)浓度至0.5 mg·L-1条件下继代, 可长久维持胚性愈伤组织的分化能力, 极大地提高了胚性愈伤组织的增殖效率及体胚发生率.
Erratum: synchronous and high frequency germination of interior spruce somatic embryos following partial drying at high relative humidity
1
1990
... 针叶树体胚成熟过程中ABA和渗透调节剂是必备条件.研究表明, 目前影响针叶树体胚发生的主要因素是ABA、蔗糖和PEG4000浓度.ABA能够有效调控处于增殖状态的胚性愈伤组织向成熟状态发育, 抑制裂生多胚现象发生, 阻止胚胎过早发育, 促进后期原胚的成熟(
Recent advances in conifer somatic embryogenesis: improving somatic embryo quality
1
2003
... 针叶树胚性愈伤组织的诱导培养基中多含有高浓度的生长素和细胞分裂素, 而增殖继代多采用低浓度的生长素和细胞分裂素(
Somatic embryogenesis in conifers
1
1991
... 针叶树胚性愈伤组织的诱导培养基中多含有高浓度的生长素和细胞分裂素, 而增殖继代多采用低浓度的生长素和细胞分裂素(
Somatic embryogenesis and plant regeneration from immature embryos of western larch
1
1992
... 落叶松(Larix gmelinii)是中国东北地区荒山造林和森林更新的重要速生树种.20世纪80年代, 欧洲率先建立落叶松属体胚发生再生体系.
Somatic embryogenesis in in vitro culture of three larch species
1
2012
... 落叶松(Larix gmelinii)是中国东北地区荒山造林和森林更新的重要速生树种.20世纪80年代, 欧洲率先建立落叶松属体胚发生再生体系.
Promotion of embryogenesis in cultured megagametophytes of Larix decidua
1
1987
... 落叶松(Larix gmelinii)是中国东北地区荒山造林和森林更新的重要速生树种.20世纪80年代, 欧洲率先建立落叶松属体胚发生再生体系.
Regulation of somatic embryogenesis in higher plants
1
2010
... 体细胞胚胎发生(简称体胚发生)是一项重要的植物生物技术, 是指在离体培养条件下, 体细胞经过类似胚胎发育的过程, 完成体细胞向胚胎发生途径转变的重编程, 最终形成完整植株(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}