许淑娟1,2, 种康1,2,*, 1中国科学院植物研究所, 植物分子生理学重点实验室, 北京 1000932中国科学院大学, 北京 100049 XuShujuan1,2, ChongKang1,2,*, 1Key Laboratory of Plant Molecular Physiology, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China2University of the Chinese Academy of Sciences, Beijing 100049, China 引用本文 许淑娟, 种康. 【-逻*辑*与-】#x0201c;先驱【-逻*辑*与-】#x0201d;转录因子LEC1在早期胚胎重置春化状态的机制. 植物学报, 2018, 53(1): 1-4
Abstract: Flowering is an important process for plants to switch from vegetative to reproductive phase. Vernalization is a process whereby plants acquire the ability to flower after exposure to a prolonged cold temperature. In Arabidopsis, inhibitor-type transcription factor FLOWERING LOCUS C (FLC) is a critical point in vernalization-mediated flowering pathway. Previous studies in Arabidopsis thaliana revealed that two homologous epigenome readers, VAL1 and VAL2, re- cognize a cis DNA element in the nucleation region for Polycomb group (PcG) silencing at the key floral repressor FLC, engaging Polycomb group proteins to induce epigenetic silencing of FLC by histone 3 lysine trimethylation (H3K27me3) during vernalization. This silencing is maintained in subsequent growth and development under normal temperature, namely vernalization memory. How to delete vernalization memory in the next generation to de novo activate FLC expression, preventing the offspring from flowering before or during winter, is not clear. Recently, Chinese scientist have found that a seed-specific transcription factor LEAFY COTYLEDON1 (LEC1) functions in deleting vernalization memory and reactivating the expression of FLC in the pro-embryo by resetting the chromatin states from the silenced state (marked by H3K27me3) to an active state (H3K36me3). This study provides important understanding of molecular and genetic mechanisms for flowering control by vernalization, and a novel strategy to genetically manipulate crop flowering times for the benefit of agricultural production, which is a great breakthrough of this field.
中国科学院上海生命科学研究院植物逆境生物学研究中心何跃辉研究组近期揭示了有关植物早期胚胎染色质状态重编程的新机制, 同时阐述了胚胎中的基因激活传递到后胚胎发育时期的表观遗传机理(图1), 该研究是开花调控分子与遗传机制的重要突破(Tao et al., 2017)。开花是植物由营养生长向生殖生长转变的重要过程。此过程受到外部环境因子和植物内在发育状态的双重复杂精准调控。这种内外协调的作用模式使得植物能在合适的时刻开花, 从而最大程度地确保生殖发育的成功及繁衍。拟南芥(Ara- bidopsis thaliana)中, 开花诱导主要由4种途径调控, 即赤霉素途径、自主途径、光周期途径和春化途径(Fornara et al., 2010)。其中, 春化作用在禾本科植物和拟南芥中都得到了广泛研究。
冬性和二年生植物需要经历一段时间持续环境低温的诱导, 才能从营养生长转入生殖生长, 该过程即为春化作用。一年生植物接受春化后在当代会产生记忆, 减数分裂形成的配子受精后, 此记忆会被擦除, 进而产生新的一代, 即形成FLC (FLOWERING LOCUS C)沉默的解除状态。春化作用的分子与表观遗传控制机理在双子叶(拟南芥)和单子叶(小麦(Triticum aestivum))植物中是完全不同的模式。小麦春化模式代表了一类春化调控分子遗传机制。在小麦中, 春化促进开花途径包括VRN1 (VERNALIZATION1)、VRN2、VRN3和VRN-D4 (VERNALIZATION D4)等春化基因的调控(Yan et al., 2003, 2006; Dubcovsky et al., 2006; Kippes et al., 2015)。其中, VRN1编码1个类似FRUITFULL的MADS-box转录因子, 该转录因子在春化过程中起至关重要的促进作用。小麦春化调控基因VER2 (VERNALIZATION- RELATED 2)具有部分取代春化的功能(Yong et al., 2003), 其编码的凝集素蛋白VER2在春化处理后发生磷酸化修饰, 并进入细胞核识别糖基化修饰的RNA结合蛋白TaGRP2 (Glycine-Rich RNA-binding Protein), TaGRP2进一步识别春化基因TaVRN1 Pre- mRNA第1内含子的critical region中RIP3基序, 进而控制其转录与开花启动(Xing et al., 2009; Xiao et al., 2014)。TaGRP2结合的RIP3基序决定着小麦的春冬性, 在小麦育种中具有非常重要的作用(Kippes et al., 2015)。关键蛋白的磷酸化(如VER2)和糖基化(如GRP2)可能参与复杂的春化记忆过程。
拟南芥开花受到FLC基因的抑制, 而FLC对开花的抑制作用随着低温处理时间的延长会被解除。在低温条件下, 组蛋白修饰和非编码RNA共同调控FLC的基因沉默(Hepworth and Dean, 2015)。非编码FLC转录本COLDAIR可能通过募集PRC2 (Polycomb Re- pressive Complex 2)复合体, 使FLC基因染色质组蛋白H3第27位的赖氨酸三甲基化(H3K27me3)修饰增加而发生基因沉默(Heo and Sung, 2011; Csorba et al., 2014)。2016年, 何跃辉团队和John Innes Centre的Caroline Dean团队分别发现了1个记忆顺式元件和反式作用因子协同Polycomb复合体, 它们对FLC周围的组蛋白进行H3K27me3表观修饰, 导致FLC基因表达关闭, 使植物在温暖的季节能够从营养生长阶段转入生殖生长阶段。在拟南芥营养生长时期, 一类B3转录因子VAL1或VAL2可以识别负调控开花的关键基因FLC成核区的顺式DNA元件, 并招募PRC2复合体催化H3K27me3, 从而沉默FLC的表达, 且在恢复常温条件后FLC基因的沉默能够被维持, 即春化记忆作用。正是春化记忆作用的存在使得植物具备了在经历漫长冬天低温处理后, 能够在来年春季开花的能力(Qüesta et al., 2016; Yuan et al., 2016)。然而在开花后及下一代种子中春化记忆如何被擦除, FLC基因如何被重新激活并不清楚。
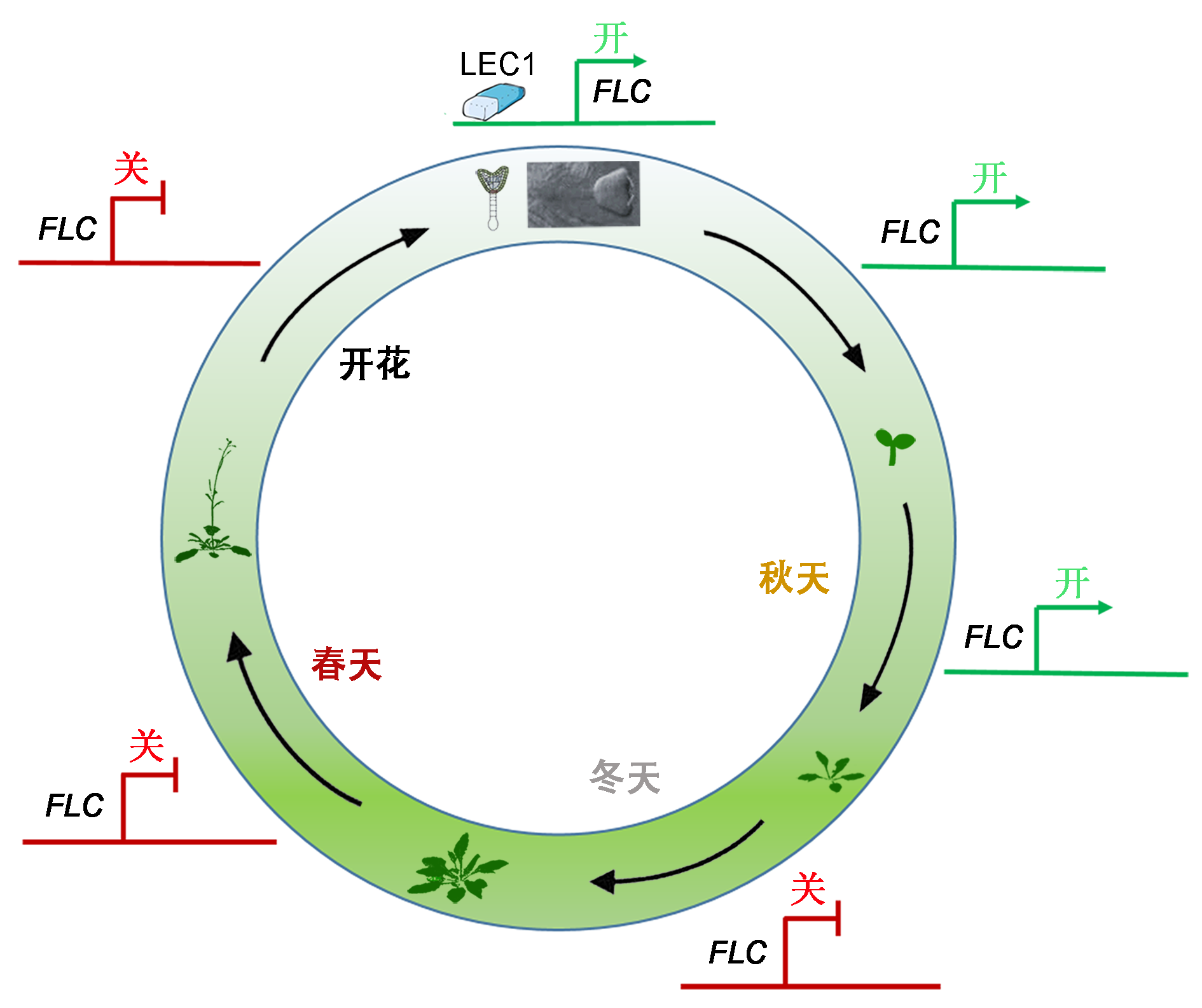
图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-1/img_1.png<b>图1</b> “先驱”转录因子LEC1 NF-Y在拟南芥生命周期中控制<i>FLC</i>表达示意图<br/>在胚胎发育早期, 1个种子特有的“先驱”转录因子LEC1重新激活<i>FLC</i>的表达, 并通过H3K36me3修饰促进染色体活性状态的建立, 擦除H3K27me3修饰的沉默状态。<i>FLC</i>的激活状态通过细胞分裂传递, 持续整个胚胎发育时期以及发育后期(幼苗期), 形成了苗期的“胚胎<i>FLC</i>表达记忆”。冬天低温使得<i>FLC</i>染色体状态因H3K27me3修饰转变成沉默状态, 且在春季回暖后<i>FLC</i>基因沉默能够被维持, 即形成春化记忆作用, 该春化记忆作用直到胚胎发育早期才被LEC1擦除。<br/><b>Figure 1</b> Schematic model for the seed-specific LEC1 NF-Y-mediated the expression of <i>FLC</i> throughout the life cy- cle in Arabidopsis<br/>The “pioneer” transcription factor LEC1 NF-Y expressed in pro-embryo de novo activates <i>FLC </i>expression through the establishment of the active chromatin state marked with H3K36me3 and consequently resetting the silenced state marked with H3K27me3, which is maintained in seed deve- lopment and passed on to seedling stages, leading to an embryonic memory of <i>FLC</i> activation in post-embryonic life. After a prolonged cold expose (winter), <i>FLC</i> was silenced by the increase of the repressive mark H3K27me3 at <i>FLC</i>. And this silenced state is maintained upon returning to warmth, named “vernalization memory”, which was deleted by LEC1 in proembryo. Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-1/img_1.png<b>图1</b> “先驱”转录因子LEC1 NF-Y在拟南芥生命周期中控制<i>FLC</i>表达示意图<br/>在胚胎发育早期, 1个种子特有的“先驱”转录因子LEC1重新激活<i>FLC</i>的表达, 并通过H3K36me3修饰促进染色体活性状态的建立, 擦除H3K27me3修饰的沉默状态。<i>FLC</i>的激活状态通过细胞分裂传递, 持续整个胚胎发育时期以及发育后期(幼苗期), 形成了苗期的“胚胎<i>FLC</i>表达记忆”。冬天低温使得<i>FLC</i>染色体状态因H3K27me3修饰转变成沉默状态, 且在春季回暖后<i>FLC</i>基因沉默能够被维持, 即形成春化记忆作用, 该春化记忆作用直到胚胎发育早期才被LEC1擦除。<br/><b>Figure 1</b> Schematic model for the seed-specific LEC1 NF-Y-mediated the expression of <i>FLC</i> throughout the life cy- cle in Arabidopsis<br/>The “pioneer” transcription factor LEC1 NF-Y expressed in pro-embryo de novo activates <i>FLC </i>expression through the establishment of the active chromatin state marked with H3K36me3 and consequently resetting the silenced state marked with H3K27me3, which is maintained in seed deve- lopment and passed on to seedling stages, leading to an embryonic memory of <i>FLC</i> activation in post-embryonic life. After a prolonged cold expose (winter), <i>FLC</i> was silenced by the increase of the repressive mark H3K27me3 at <i>FLC</i>. And this silenced state is maintained upon returning to warmth, named “vernalization memory”, which was deleted by LEC1 in proembryo.
图1 “先驱”转录因子LEC1 NF-Y在拟南芥生命周期中控制FLC表达示意图 在胚胎发育早期, 1个种子特有的“先驱”转录因子LEC1重新激活FLC的表达, 并通过H3K36me3修饰促进染色体活性状态的建立, 擦除H3K27me3修饰的沉默状态。FLC的激活状态通过细胞分裂传递, 持续整个胚胎发育时期以及发育后期(幼苗期), 形成了苗期的“胚胎FLC表达记忆”。冬天低温使得FLC染色体状态因H3K27me3修饰转变成沉默状态, 且在春季回暖后FLC基因沉默能够被维持, 即形成春化记忆作用, 该春化记忆作用直到胚胎发育早期才被LEC1擦除。 Figure 1 Schematic model for the seed-specific LEC1 NF-Y-mediated the expression of FLC throughout the life cy- cle in Arabidopsis The “pioneer” transcription factor LEC1 NF-Y expressed in pro-embryo de novo activates FLC expression through the establishment of the active chromatin state marked with H3K36me3 and consequently resetting the silenced state marked with H3K27me3, which is maintained in seed deve- lopment and passed on to seedling stages, leading to an embryonic memory of FLC activation in post-embryonic life. After a prolonged cold expose (winter), FLC was silenced by the increase of the repressive mark H3K27me3 at FLC. And this silenced state is maintained upon returning to warmth, named “vernalization memory”, which was deleted by LEC1 in proembryo.
近期, 何跃辉研究组发现了1个种子独有的“先驱”转录因子(pioneer transcription factor) LEC1 (L- EAFY COTYLEDON1)能够在受精后使得沉默状态的FLC基因被重新激活(Tao et al., 2017)。一年生越冬生态型拟南芥的FRI (FRIGIDA), 在低温来临之前通过促进FLC基因组蛋白发生活化状态修饰的酶类(如H3K36甲基转移酶EFS (SDG8))富集, 使FLC基因上调到一个很高的水平(Kim et al., 2009; He, 2012; Crevillén et al., 2014 )。低温状态下, PRC2复合体在FLC成核区富集, 并使其周围组蛋白发生H3K27- me3修饰, FLC基因处于沉默状态, 且在恢复常温后通过有丝分裂依然维持这种沉默状态。之前有报道, H3K27去甲基化酶ELF6 (EARLY FLOWERING 6)参与受精后在下一代重置FLC沉默状态(Crevillén et al., 2014), 但通过对弱突变体elf6-3和敲除突变体elf6-4中FLC基因在春化前后及受精后的表达情况进行分析, 发现春化记忆仍被擦除(Tao et al., 2017), 可见有其它因子主导FLC的重新激活。
CrevillénP, YangHC, CuiX, GreeffC, TrickM, QiuQ, CaoXF, DeanC (2014). Epigenetic reprogramming that prevents transgenerational inheritance of the vernalized state.Nature 515, 587-590. [本文引用: 1]
[2]
CsorbaT, QuestaJI, SunQ, DeanC (2014). Antisense COOLAIR mediates the coordinated switching of chromatin states at FLC during vernalization. Proc Natl Acad Sci USA 111, 16160-16165. [本文引用: 1]
[3]
DubcovskyJ, LoukoianovA, FuDL, ValarikM, SanchezA, YanLL (2006). Effect of photoperiod on the regulation of wheat vernalization genes VRN1 and VRN2. Plant Mol Biol 60, 469-480. [本文引用: 1]
[4]
FornaraF, de MontaiguA, CouplandG (2010). SnapShot: control of flowering in Arabidopsis. Cell 141, 550-550. e2 [本文引用: 1]
HeoJB, SungS (2011). Vernalization-mediated epigenetic silencing by a long intronic noncoding RNA.Science 331, 76-79. [本文引用: 1]
[7]
HepworthJ, DeanC (2015). Flowering Locus C's lessons: conserved chromatin switches underpinning developmental timing and adaptation.Plant Physiol 168, 1237-1245. [本文引用: 1]
[8]
KimDH, DoyleMR, SungS, AmasinoRM (2009). Vernalization: winter and the timing of flowering in plants.An- nu Rev Cell Dev Biol 25, 277-299. [本文引用: 1]
[9]
KippesN, DebernardiJM, Vasquez-GrossHA, AkpinarBA, BudakH, KatoK, ChaoS, AkhunovE, DubcovskyJ (2015). Identification of the VERNALIZATION 4 gene reveals the origin of spring growth habit in ancient wheats from South Asia. Proc Natl Acad Sci USA 112, E5401-E5410. [本文引用: 2]
[10]
KwongRW, BuiAQ, LeeH, KwongLW, FischerRL, GoldbergRB, HaradaJJ (2003). LEAFY COTYLEDON1-LIKE defines a class of regulators essential for em- bryo development.Plant Cell 15, 5-18. [本文引用: 1]
TaoZ, ShenLS, GuXF, WangYZ, YuH, HeYH (2017). Embryonic epigenetic reprogramming by a pioneer trans- cription factor in plants.Nature 551, 124-128. [本文引用: 3]
[13]
XiaoJ, XuSJ, LiCH, XuYY, XingLJ, NiuYD, HuanQ, TangYM, ZhaoCP, WagnerD, GaoCX, ChongK (2014). O-GlcNAc-mediated interaction between VER2 and TaGRP2 elicits TaVRN1 mRNA accumulation during ver- nalization in winter wheat. Nat Commun 5, 4572. [本文引用: 1]
[14]
XingLJ, LiJ, XuYY, XuZH, ChongK (2009). Phosphorylation modification of wheat lectin VER2 is associated with vernalization-induced O-GlcNAc signaling and intracellular motility. PLoS One 4, e4854. [本文引用: 1]
[15]
YanL, FuD, LiC, BlechlA, TranquilliG, BonafedeM, SanchezA, ValarikM, YasudaS, DubcovskyJ (2006). The wheat and barley vernalization gene VRN3 is an orthologue of FT. Proc Natl Acad Sci USA 103, 19581-19586. [本文引用: 1]
[16]
YanL, LoukoianovA, TranquilliG, HelgueraM, FahimaT, DubcovskyJ (2003). Positional cloning of the wheat vernalization gene VRN1. Proc Natl Acad Sci USA 100, 6263-6268. [本文引用: 1]
[17]
YongWD, XuYY, XuWZ, WangX, LiN, WuJS, LiangTB, ChongK, XuZH, TanKH, ZhuZQ (2003). Vernalization-induced flowering in wheat is mediated by a lectin-like gene VER2. Planta 217, 261-270. [本文引用: 1]
[18]
YuanWY, LuoX, LiZC, YangWN, WangYZ, LiuR, DuiJM, HeYH (2016). A cis cold memory element and a trans epigenome reader mediate Polycomb silencing of FLC by vernalization in Arabidopsis. Nat Genet 48, 1527-1534. [本文引用: 1]
1 2014
... 近期, 何跃辉研究组发现了1个种子独有的“先驱”转录因子(pioneer transcription factor) LEC1 (L- EAFY COTYLEDON1)能够在受精后使得沉默状态的FLC基因被重新激活(Tao et al., 2017).一年生越冬生态型拟南芥的FRI (FRIGIDA), 在低温来临之前通过促进FLC基因组蛋白发生活化状态修饰的酶类(如H3K36甲基转移酶EFS (SDG8))富集, 使FLC基因上调到一个很高的水平(Kim et al., 2009; He, 2012; Crevillén et al., 2014 ).低温状态下, PRC2复合体在FLC成核区富集, 并使其周围组蛋白发生H3K27- me3修饰, FLC基因处于沉默状态, 且在恢复常温后通过有丝分裂依然维持这种沉默状态.之前有报道, H3K27去甲基化酶ELF6 (EARLY FLOWERING 6)参与受精后在下一代重置FLC沉默状态(Crevillén et al., 2014), 但通过对弱突变体elf6-3和敲除突变体elf6-4中FLC基因在春化前后及受精后的表达情况进行分析, 发现春化记忆仍被擦除(Tao et al., 2017), 可见有其它因子主导FLC的重新激活. ...
1 2014
... 拟南芥开花受到FLC基因的抑制, 而FLC对开花的抑制作用随着低温处理时间的延长会被解除.在低温条件下, 组蛋白修饰和非编码RNA共同调控FLC的基因沉默(Hepworth and Dean, 2015).非编码FLC转录本COLDAIR可能通过募集PRC2 (Polycomb Re- pressive Complex 2)复合体, 使FLC基因染色质组蛋白H3第27位的赖氨酸三甲基化(H3K27me3)修饰增加而发生基因沉默(Heo and Sung, 2011; Csorba et al., 2014).2016年, 何跃辉团队和John Innes Centre的Caroline Dean团队分别发现了1个记忆顺式元件和反式作用因子协同Polycomb复合体, 它们对FLC周围的组蛋白进行H3K27me3表观修饰, 导致FLC基因表达关闭, 使植物在温暖的季节能够从营养生长阶段转入生殖生长阶段.在拟南芥营养生长时期, 一类B3转录因子VAL1或VAL2可以识别负调控开花的关键基因FLC成核区的顺式DNA元件, 并招募PRC2复合体催化H3K27me3, 从而沉默FLC的表达, 且在恢复常温条件后FLC基因的沉默能够被维持, 即春化记忆作用.正是春化记忆作用的存在使得植物具备了在经历漫长冬天低温处理后, 能够在来年春季开花的能力(Qüesta et al., 2016; Yuan et al., 2016).然而在开花后及下一代种子中春化记忆如何被擦除, FLC基因如何被重新激活并不清楚. ...
1 2006
... 冬性和二年生植物需要经历一段时间持续环境低温的诱导, 才能从营养生长转入生殖生长, 该过程即为春化作用.一年生植物接受春化后在当代会产生记忆, 减数分裂形成的配子受精后, 此记忆会被擦除, 进而产生新的一代, 即形成FLC (FLOWERING LOCUS C)沉默的解除状态.春化作用的分子与表观遗传控制机理在双子叶(拟南芥)和单子叶(小麦(Triticum aestivum))植物中是完全不同的模式.小麦春化模式代表了一类春化调控分子遗传机制.在小麦中, 春化促进开花途径包括VRN1 (VERNALIZATION1)、VRN2、VRN3和VRN-D4 (VERNALIZATION D4)等春化基因的调控(Yan et al., 2003, 2006; Dubcovsky et al., 2006; Kippes et al., 2015).其中, VRN1编码1个类似FRUITFULL的MADS-box转录因子, 该转录因子在春化过程中起至关重要的促进作用.小麦春化调控基因VER2 (VERNALIZATION- RELATED 2)具有部分取代春化的功能(Yong et al., 2003), 其编码的凝集素蛋白VER2在春化处理后发生磷酸化修饰, 并进入细胞核识别糖基化修饰的RNA结合蛋白TaGRP2 (Glycine-Rich RNA-binding Protein), TaGRP2进一步识别春化基因TaVRN1 Pre- mRNA第1内含子的critical region中RIP3基序, 进而控制其转录与开花启动(Xing et al., 2009; Xiao et al., 2014).TaGRP2结合的RIP3基序决定着小麦的春冬性, 在小麦育种中具有非常重要的作用(Kippes et al., 2015).关键蛋白的磷酸化(如VER2)和糖基化(如GRP2)可能参与复杂的春化记忆过程. ...
1 2010
... 中国科学院上海生命科学研究院植物逆境生物学研究中心何跃辉研究组近期揭示了有关植物早期胚胎染色质状态重编程的新机制, 同时阐述了胚胎中的基因激活传递到后胚胎发育时期的表观遗传机理(图1), 该研究是开花调控分子与遗传机制的重要突破(Tao et al., 2017).开花是植物由营养生长向生殖生长转变的重要过程.此过程受到外部环境因子和植物内在发育状态的双重复杂精准调控.这种内外协调的作用模式使得植物能在合适的时刻开花, 从而最大程度地确保生殖发育的成功及繁衍.拟南芥(Ara- bidopsis thaliana)中, 开花诱导主要由4种途径调控, 即赤霉素途径、自主途径、光周期途径和春化途径(Fornara et al., 2010).其中, 春化作用在禾本科植物和拟南芥中都得到了广泛研究. ...
1 2012
... 近期, 何跃辉研究组发现了1个种子独有的“先驱”转录因子(pioneer transcription factor) LEC1 (L- EAFY COTYLEDON1)能够在受精后使得沉默状态的FLC基因被重新激活(Tao et al., 2017).一年生越冬生态型拟南芥的FRI (FRIGIDA), 在低温来临之前通过促进FLC基因组蛋白发生活化状态修饰的酶类(如H3K36甲基转移酶EFS (SDG8))富集, 使FLC基因上调到一个很高的水平(Kim et al., 2009; He, 2012; Crevillén et al., 2014 ).低温状态下, PRC2复合体在FLC成核区富集, 并使其周围组蛋白发生H3K27- me3修饰, FLC基因处于沉默状态, 且在恢复常温后通过有丝分裂依然维持这种沉默状态.之前有报道, H3K27去甲基化酶ELF6 (EARLY FLOWERING 6)参与受精后在下一代重置FLC沉默状态(Crevillén et al., 2014), 但通过对弱突变体elf6-3和敲除突变体elf6-4中FLC基因在春化前后及受精后的表达情况进行分析, 发现春化记忆仍被擦除(Tao et al., 2017), 可见有其它因子主导FLC的重新激活. ...
1 2011
... 拟南芥开花受到FLC基因的抑制, 而FLC对开花的抑制作用随着低温处理时间的延长会被解除.在低温条件下, 组蛋白修饰和非编码RNA共同调控FLC的基因沉默(Hepworth and Dean, 2015).非编码FLC转录本COLDAIR可能通过募集PRC2 (Polycomb Re- pressive Complex 2)复合体, 使FLC基因染色质组蛋白H3第27位的赖氨酸三甲基化(H3K27me3)修饰增加而发生基因沉默(Heo and Sung, 2011; Csorba et al., 2014).2016年, 何跃辉团队和John Innes Centre的Caroline Dean团队分别发现了1个记忆顺式元件和反式作用因子协同Polycomb复合体, 它们对FLC周围的组蛋白进行H3K27me3表观修饰, 导致FLC基因表达关闭, 使植物在温暖的季节能够从营养生长阶段转入生殖生长阶段.在拟南芥营养生长时期, 一类B3转录因子VAL1或VAL2可以识别负调控开花的关键基因FLC成核区的顺式DNA元件, 并招募PRC2复合体催化H3K27me3, 从而沉默FLC的表达, 且在恢复常温条件后FLC基因的沉默能够被维持, 即春化记忆作用.正是春化记忆作用的存在使得植物具备了在经历漫长冬天低温处理后, 能够在来年春季开花的能力(Qüesta et al., 2016; Yuan et al., 2016).然而在开花后及下一代种子中春化记忆如何被擦除, FLC基因如何被重新激活并不清楚. ...
1 2015
... 拟南芥开花受到FLC基因的抑制, 而FLC对开花的抑制作用随着低温处理时间的延长会被解除.在低温条件下, 组蛋白修饰和非编码RNA共同调控FLC的基因沉默(Hepworth and Dean, 2015).非编码FLC转录本COLDAIR可能通过募集PRC2 (Polycomb Re- pressive Complex 2)复合体, 使FLC基因染色质组蛋白H3第27位的赖氨酸三甲基化(H3K27me3)修饰增加而发生基因沉默(Heo and Sung, 2011; Csorba et al., 2014).2016年, 何跃辉团队和John Innes Centre的Caroline Dean团队分别发现了1个记忆顺式元件和反式作用因子协同Polycomb复合体, 它们对FLC周围的组蛋白进行H3K27me3表观修饰, 导致FLC基因表达关闭, 使植物在温暖的季节能够从营养生长阶段转入生殖生长阶段.在拟南芥营养生长时期, 一类B3转录因子VAL1或VAL2可以识别负调控开花的关键基因FLC成核区的顺式DNA元件, 并招募PRC2复合体催化H3K27me3, 从而沉默FLC的表达, 且在恢复常温条件后FLC基因的沉默能够被维持, 即春化记忆作用.正是春化记忆作用的存在使得植物具备了在经历漫长冬天低温处理后, 能够在来年春季开花的能力(Qüesta et al., 2016; Yuan et al., 2016).然而在开花后及下一代种子中春化记忆如何被擦除, FLC基因如何被重新激活并不清楚. ...
1 2009
... 近期, 何跃辉研究组发现了1个种子独有的“先驱”转录因子(pioneer transcription factor) LEC1 (L- EAFY COTYLEDON1)能够在受精后使得沉默状态的FLC基因被重新激活(Tao et al., 2017).一年生越冬生态型拟南芥的FRI (FRIGIDA), 在低温来临之前通过促进FLC基因组蛋白发生活化状态修饰的酶类(如H3K36甲基转移酶EFS (SDG8))富集, 使FLC基因上调到一个很高的水平(Kim et al., 2009; He, 2012; Crevillén et al., 2014 ).低温状态下, PRC2复合体在FLC成核区富集, 并使其周围组蛋白发生H3K27- me3修饰, FLC基因处于沉默状态, 且在恢复常温后通过有丝分裂依然维持这种沉默状态.之前有报道, H3K27去甲基化酶ELF6 (EARLY FLOWERING 6)参与受精后在下一代重置FLC沉默状态(Crevillén et al., 2014), 但通过对弱突变体elf6-3和敲除突变体elf6-4中FLC基因在春化前后及受精后的表达情况进行分析, 发现春化记忆仍被擦除(Tao et al., 2017), 可见有其它因子主导FLC的重新激活. ...
2 2015
... 冬性和二年生植物需要经历一段时间持续环境低温的诱导, 才能从营养生长转入生殖生长, 该过程即为春化作用.一年生植物接受春化后在当代会产生记忆, 减数分裂形成的配子受精后, 此记忆会被擦除, 进而产生新的一代, 即形成FLC (FLOWERING LOCUS C)沉默的解除状态.春化作用的分子与表观遗传控制机理在双子叶(拟南芥)和单子叶(小麦(Triticum aestivum))植物中是完全不同的模式.小麦春化模式代表了一类春化调控分子遗传机制.在小麦中, 春化促进开花途径包括VRN1 (VERNALIZATION1)、VRN2、VRN3和VRN-D4 (VERNALIZATION D4)等春化基因的调控(Yan et al., 2003, 2006; Dubcovsky et al., 2006; Kippes et al., 2015).其中, VRN1编码1个类似FRUITFULL的MADS-box转录因子, 该转录因子在春化过程中起至关重要的促进作用.小麦春化调控基因VER2 (VERNALIZATION- RELATED 2)具有部分取代春化的功能(Yong et al., 2003), 其编码的凝集素蛋白VER2在春化处理后发生磷酸化修饰, 并进入细胞核识别糖基化修饰的RNA结合蛋白TaGRP2 (Glycine-Rich RNA-binding Protein), TaGRP2进一步识别春化基因TaVRN1 Pre- mRNA第1内含子的critical region中RIP3基序, 进而控制其转录与开花启动(Xing et al., 2009; Xiao et al., 2014).TaGRP2结合的RIP3基序决定着小麦的春冬性, 在小麦育种中具有非常重要的作用(Kippes et al., 2015).关键蛋白的磷酸化(如VER2)和糖基化(如GRP2)可能参与复杂的春化记忆过程. ... ... ).TaGRP2结合的RIP3基序决定着小麦的春冬性, 在小麦育种中具有非常重要的作用(Kippes et al., 2015).关键蛋白的磷酸化(如VER2)和糖基化(如GRP2)可能参与复杂的春化记忆过程. ...


{kind=link}
{kind=link}