 ,1,2,*
,1,2,*Advances in Genetic Studies of the Awn in Cereal Crops
Fei Qi1, Piyi Xing2, Yinguang Bao1,2, Honggang Wang1,2, Xingfeng Li,1,2,*通讯作者: E-mail:lixf@sdau.edu.cn
责任编辑: 白羽红
收稿日期:2019-12-11接受日期:2020-03-23网络出版日期:2020-09-01
| 基金资助: |
Corresponding authors: E-mail:lixf@sdau.edu.cn
Received:2019-12-11Accepted:2020-03-23Online:2020-09-01

摘要
芒是许多禾本科作物穗部的重要结构, 不仅可以作为区分不同品种以及基因定位的重要形态标记, 而且在禾谷类作物的种子传播、籽粒灌浆、蒸腾作用及产量形成等方面起重要作用。该文综述了小麦(Triticum aestivum)、大麦(Hordeum vulgare)和水稻(Oryza sativa)芒的结构、功能与遗传调控机制研究进展, 以期为芒性状遗传机理的进一步研究及其在育种中的应用提供参考。
关键词:
Abstract
The awn is an important structure on the panicles of many cereal crops, which can not only be used as an important morphological character to distinguish different varieties and as a morphological marker for genetic mapping, but also play an important role in seed transmission, grain filling, transpiration and yield. Here we summarize the recent advancement of studies on the structure, function and genetic mechanism of the awn in wheat, barley and rice, which may provide a reference for the further studies on the genetic mechanism of the awn development, and for the application in breeding.
Keywords:
PDF (3939KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
亓斐, 邢丕一, 鲍印广, 王洪刚, 李兴锋. 禾本科作物芒遗传研究进展. 植物学报, 2020, 55(5): 613-622 doi:10.11983/CBB19236
Qi Fei, Xing Piyi, Bao Yinguang, Wang Honggang, Li Xingfeng.

芒是禾本科作物穗部的重要结构, 属于叶的变态, 是植物长期适应环境和进化的产物。不同材料存在芒的有无及芒长度的差异, 可作为区分禾谷类作物不同品种以及基因定位的重要形态标记(Watkins and Ellerton, 1940; Cuthbert et al., 2008; Qureshi et al., 2018)。此外, 芒在禾谷类作物种子传播、籽粒灌浆、蒸腾作用及产量形成方面也起到至关重要的作用(Grundbacher, 1963; 王忠等, 1993; Elbaum et al., 2007; Rebetzke et al., 2016)。因此, 充分了解芒的形态特征、功能及其遗传机制对挖掘禾本科作物光合潜力、提高蒸腾效率及促进产量形成有重要意义, 对作物品质改良和遗传育种也有重要参考价值。
1 芒的形态结构特征
1.1 芒的解剖学特征
芒是禾本科植物小花外稃或颖片顶端的一种针状延伸, 从基部到顶端逐渐变细。其表皮含有不同形态的细胞, 如较狭长的细胞、卵状或方形的短细胞和厚壁而尖细的单细胞。其中较狭长的细胞具有波状加厚的壁; 卵状或方形的短细胞表现为芒表皮上的乳头状突起; 厚壁而尖细的单细胞向前斜生, 使得芒表面较为粗糙(Sorensen, 1986; 杜斌等, 2010)。芒的远轴端两个侧面上分布着带状排列的气孔, 基部表面排列的气孔最多, 其次为中部, 上部的气孔最少。气孔作为植物气体交换和蒸腾作用的重要通道, 在水分代谢和光合作用中起重要作用。气孔带下方是绿色组织, 这些绿色细胞对光合作用至关重要(王忠等, 1993)。芒的近轴端无气孔和绿色细胞, 是芒的非绿色区域, 其表皮细胞细胞壁明显增厚, 这对于减少水分散失和维持芒的形态起到重要作用(王忠等, 1993; 李晓娟, 2006; 杜斌等, 2010)。芒的横切面一般呈三角形。例如, 小麦(Triticum aestivum)芒的横切面为锐角三角形, 大麦(Hordeum vulgare)为钝角三角形。其横切面内部含有3条维管束, 中间较粗, 两边较细。中间的维管束具有完整的结构, 其外有2层维管束鞘, 内有木质部和韧皮部。外鞘细胞中与绿色组织相连的部分含有少量叶绿体, 不与绿色组织相连的外鞘细胞及内鞘细胞不含叶绿体。木质部中有少量薄壁细胞和厚壁导管。韧皮部中有筛管与伴胞, 且一一对应(图1) (王忠等, 1993; 李晓娟, 2006; 杜斌等, 2010)。
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1芒的横切面结构
(A) 大麦芒横切面; (B) 普通小麦芒横切面。S: 气孔; V: 维管束; Pa: 薄壁组织; Sc: 厚壁组织
Figure 1Cross-section of the awn
(A) Cross-section of the awn of Hordeum vulgare; (B) Cross-section of the awn of Triticum aestivum. S: Stoma; V: Vascular bundle; Pa: Parenchyma; Sc: Sclerenchyma
1.2 芒中叶绿体的超显微结构
在小麦和大麦芒绿色组织中, 绿色细胞常呈不规则形状, 含有丰富的叶绿体。不同时期其叶绿体形态结构存在明显变化(王忠等, 1993)。在抽穗期, 叶绿体尚未完全分化, 部分可称之为前质体, 该时期已出现类囊体(图2A)。到了开花期, 叶绿体体积开始增大, 形状更为细长, 出现基粒(图2B)。在灌浆期, 叶绿体体积显著增大, 出现大量的基粒及少量的淀粉粒和质体小球(图2C)。在干物质形成期, 叶绿体体积达到峰值, 淀粉粒和质体小球的数量显著增多(图2D)。在成熟期, 叶绿体的体积开始逐渐变小, 类囊体出现膨胀、松散和扭曲现象, 淀粉粒的含量也明显减少(图2E)。在收获期, 叶绿体逐渐裂解, 类囊体开始解体直至消失, 只有少数保持垛叠状态(李晓娟, 2006)。芒中叶绿体的动态变化及其与光合作用的关系值得深入研究。图2
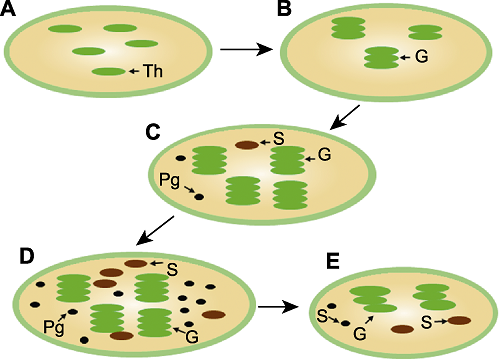 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同发育时期芒中叶绿体的结构
(A) 抽穗期; (B) 开花期; (C) 灌浆期; (D) 干物质形成期; (E) 成熟期。Th: 类囊体; G: 叶绿体基粒; Pg: 质体小球; S: 淀粉粒
Figure 2Chloroplast structure of the awn at different developmental stages
(A) Heading stage; (B) Anthesis stage; (C) Filling stage; (D) Dry matter formation stage; (E) Ripening stage. Th: Thylakoid; G: Granum; Pg: Plastoglobuli; S: Starch
2 芒的功能
2.1 芒在生物进化中的意义
芒是禾谷类作物一种重要的驯化性状, 其表面的倒刺可以附着在动物皮毛上, 借助动物的运动进行传播(Sorensen, 1986)。禾谷类作物的种子从母穗上脱落时, 每个种子播散单元中2个明显的芒可以起到保持平衡的作用, 使得成熟种子的胚端先着地, 促进种子萌发。研究表明, 种子上2个芒的张合角度可以随日夜交替中空气湿度的变化而变化, 这一吸湿特性可以促进种子下扎进入土壤, 而芒表面斜生的倒刺可固定在土壤中, 阻止种子向上移动(图3), 显示芒在促进种子繁殖上起到“自我掩埋”作用(Elbaum et al., 2007)。同时, 芒表面坚硬的硅质毛可阻碍害虫和鸟类飞落, 减少害虫在穗上产卵的机会, 有利于预防鸟害和虫害。尖锐的芒还可阻止动物摄食, 起到保护种子的作用(Grundbacher, 1963)。此外, 芒的有无及形状可作为区分不同品种及基因定位的重要形态学标记(Watkins and Ellerton, 1940; Cuthbert et al., 2008; Qureshi et al., 2017)。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3具芒种子下扎土壤情况(改自Elbaum et al., 2007)
d: 深度
Figure 3The awned seeds were planted in the soil (modified from Elbaum et al., 2007)
d: Depth
2.2 芒与光合作用
芒中的叶绿体与叶片中的叶绿体具有相似的结构和功能。芒作为穗部重要的光合器官, 与叶片相比, 在光合作用上具有许多优势。首先, 芒的光合面积大, 且着生于穗部顶端, 利于截获光能和气体(巴青松等, 2010)。其次, 芒的维管束与籽粒直接相连, 有利于光合产物运往最近的籽粒, 具有运输距离短和速度快的特性, 该特性既有利于促进光合作用, 又能促进同化物的转运(Evans and Rawson, 1970; Evans et al., 1972; Teare et al., 1972; Olugbemi et al., 1976; Olugbemi, 1978)。此外, 小麦发育后期旗叶衰老且光合速率开始下降, 芒不易老化的特性使其仍具有较高的光合速率(Teare et al., 1972; 王忠等, 1993; 李寒冰等, 2002)。在干旱条件下, 叶片的光合速率显著降低, 而芒的光合速率仍能保持相对稳定的状态, 因此干旱条件下有芒小麦相较于无芒小麦增产约1/4 (Evans et al., 1972)。此外, 芒不存在光合午休现象(杜斌等, 2010)。研究表明, 芒的光合产物约占整个穗部光合产物的1/3, 光合量占整个穗部的1/2 (李晓娟, 2006)。在大麦中, 芒作为穗部主要的光合器官, 优先表达与光合作用相关的基因(Abebe et al., 2009)。2.3 芒的蒸腾作用
气孔的主要作用是与外界进行气体交换, 与旗叶相比, 芒的气孔总数少且体积小(张永平等, 2006)。芒的气孔可以根据外界环境的变化来调节其张合度, 这一特性使植株在水分散失最少的条件下获取最多的CO2 (Brown and Escombe, 1901)。芒的蒸腾比率、相对含水量、电子运输速率和水分利用效率远高于旗叶(Maydup et al., 2014)。芒表面的气孔占穗部气孔数的1/2。在相同条件下, 去芒穗的蒸腾作用比未去芒穗低约75%。在炎热干旱条件下, 有芒植株通过增强蒸腾作用, 迅速降低温度, 增强吸水动力, 促进根系下扎土壤以吸收更多的水分和矿物质, 提高水分利用率, 促进水分向穗部转运, 增强植株抗旱能力(张永平等, 2006; 巴青松等, 2010; Maydup et al., 2014)。研究表明, 干旱条件下, 大麦芒中抗旱相关基因的表达量高于其内稃和外稃。因此, 在干旱和半干旱环境下, 芒对禾谷类作物籽粒产量的维持具有重要意义(Abebe et al., 2010)。
2.4 芒对禾谷类作物产量的贡献
芒对产量的贡献虽然没有旗叶显著, 但两者结合比单独作用对产量的贡献更大(Teare et al., 1972)。芒通过光合作用将光合产物运往籽粒, 促进产量形成; 通过蒸腾作用促进植株吸收水分, 提高耐旱性。研究表明, 有芒作物的产量一般显著高于同类无芒作物, 增产约1/10 (Biscoe et al., 1973; Motzo and Giunta, 2002; Guo and Schnurbusch, 2016)。早期的研究多采用剪芒处理的方法。陈培元和李英(1981)对不同小麦品种进行剪芒处理, 发现千粒重均有下降。Rebetzke等(2016)通过构建小麦有芒、无芒近等基因系, 发现有芒小麦产量显著高于无芒小麦。大麦中的芒也通过其光合作用促进产量的增加, 芒缺失则同样伴随其产量显著下降(Milner et al., 2019)。在水稻(Oryza sativa)中, 大量与芒相关的基因不仅影响芒的发生发育, 而且对水稻穗粒数、粒长及单株分蘖等性状也有一定影响, 而这些性状都对水稻产量形成起关键作用(Luo et al., 2013)。
3 控制芒性状的基因及其遗传机制
3.1 小麦芒的遗传学研究
在普通小麦中, B1 (5AL)、B2 (6BL)和Hd (4AL)是目前已知的芒显性抑制基因, 它们的不同组合导致芒表型发生变化(Sourdille et al., 2004; Mackay et al., 2014)。3个隐性等位基因b1、b2和hd的纯合子为有芒表型, 1个显性等位基因的存在会抑制芒的伸长和发育, 含有2个显性抑制等位基因的植株是无芒表型(Antonyuk et al., 2012) (表1)。Table 1
表1
表1小麦中已定位的控制芒功能的基因
Table 1
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| B1 | 5AL | 分生组织的 维持 | DeWitt et al., 2020; Huang et al., 2020 |
| B2 | 6BL | 抑制芒伸长 | 金迪等, 2019 |
| Hd (Wknox1a) | 4AL | 抑制芒伸长 | Milner et al., 2019 |
| Wknox1b Wknox1d | 4BS 4DS | 茎尖分生组织的形成和维持 | Morimoto et al., 2005 |
新窗口打开|下载CSV
研究表明, 小麦B1基因对芒长度的影响最强, 其突变体的表型为穗基部和中部产生短芒或无芒, 芒最长仅为1 cm (Yoshioka et al., 2017; Huang et al., 2020; Wang et al., 2020; DeWitt et al., 2020)。对B1近等基因系不同材料间的小穗数、粒长、粒重和容重进行统计, 发现无芒个体的小穗数高于有芒个体, 但有芒个体的粒重和容重较无芒个体分别增加了6.3%和11.6%, 表明B1基因与小麦小穗数、粒重和容重显著相关(DeWitt et al., 2020)。
美国北卡罗来纳州立大学的Gina Brown-Guedira团队通过对14 567个GBS标记进行关联分析, 将B1位点定位于5A染色体681 455 268-706 705 101 bp之间25 Mb的区间内, 其中SNP (5A28417)位点与小麦品系中芒的表型共分离且与小麦容重显著关联, 该位点位于预测基因TraesCS5A02G542800上游219 bp处。小麦有芒突变体在5AL远端区域发生缺失, 缺失片段包含候选基因TraesCS5A02G542700和TraesCS5A02G542800, 利用特异标记分析最终确定TraesCS5A02G542800为B1的候选基因(DeWitt et al., 2020)。加拿大J Allan Feurtado团队利用硬粒小麦构建F2群体并进行RNA-seq分析, 他们开发标记并进行精细定位, 结合突变体验证, 最终确定编码C2H2锌指的TraesCS5A02G542800即为B1芒抑制基因。定量分析表明, 该基因在无芒小麦中的表达量为有芒小麦的8倍。此外, 过表达TraesCS5A02G542800基因, 芒长受到显著抑制, 进一步证明TraesCS5A02G542800作为B1基因的候选基因具有显著的芒抑制效应(Huang et al., 2020)。
中国科学院遗传与发育生物学研究所张爱民研究团队初步将B1定位于5A染色体长臂125 kb的物理区间内, 其中包含2个基因(TraesCS5A02G542800和TraesCS5A02G542900)。表达谱和序列分析表明, 短芒材料中TraesCS5A02G542800的表达量远高于长芒材料, 而TraesCS5A02G542900在短芒和长芒材料间表达量无显著差异。测序结果表明, TraesCS5A02G542900序列在短芒和长芒材料中一致, TraesCS5A02G542800编码区序列一致, 但在启动子区有5个相同差异的SNP, B1/B1基因型材料表现为TAGA单体型, b1/b1基因型材料表现为CGAG单体型。转基因功能验证进一步表明, TraesCS5A02G542800启动子为B1芒长抑制位点的控制基因, 将其命名为ALI-1 (Wang et al., 2020)。
ALI-1编码1个C2H2锌指转录因子, 具有转录抑制活性。对B1近等基因系的芒进行纵切观察, 结果显示两者细胞长度无显著差异, 但短芒材料细胞数目仅为长芒材料的1/10, 表明短芒材料芒原基的细胞分裂被抑制。顺式调控元件预测结果表明, ALI-1区间内SNPs导致该基因缺失顺式元件BOXCPSAS1、LTRE1HVBLT49、SORLIP2AT和SITEIIATCYTC, 这些顺式元件参与调控分生组织和增殖细胞的基因表达(Hudson and Quail, 2003; Welchen and Gonzalez, 2006; DeWitt et al., 2020; Huang et al., 2020; Wang et al., 2020)。启动子中的SNPs上调了B1的表达, 导致CDC25微量表达。CDC25是细胞周期负调控因子, 可抑制细胞分裂。在植物体内, 细胞周期调节因子与植物激素相互作用。CDC25同源物被认为是细胞分裂素作用的早期靶点, 因此CDC25降低了细胞分裂素的含量(Lipavská et al., 2011; Wang et al., 2020)。在降低细胞分裂素含量的同时, ALI-1抑制细胞分裂素的信号转导, 导致细胞增殖停滞, 细胞数量减少, 进而使芒的伸长受到抑制, 表现为极显著的芒抑制作用(Wang et al., 2020) (图4)。此外, 单体型CGAG还表现出粒长和千粒重显著增加。
图4
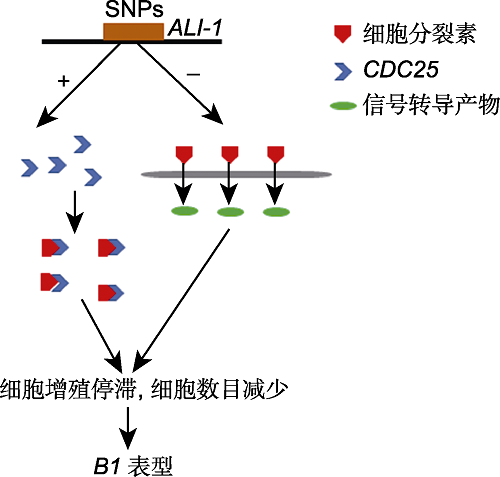 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4小麦B1基因的调控机制
Figure 4Regulation mechanism of wheat B1
截至目前, 虽然B1基因已被克隆, 但其表达和抑芒机制仍然未知。目前, 对其突变位点及功能研究仍然存在较大争议, 还需要进一步的实验验证(DeWitt et al., 2020; Huang et al., 2020; Wang et al., 2020)。
人们对于芒作用的认识现今主要集中在芒可以通过促进光合作用影响千粒重和产量。此外, 芒还可以直接调控种子大小, 扩大库容量(Wang et al., 2019)。
小麦B2基因表型为整个穗部的短芒。一般情况下, 穗部各部分的芒长一致, 少数情况下会出现穗中部芒比较长(Yoshioka et al., 2017)。研究表明, B2基因定位于小麦6B染色体上0.29 cM的遗传区间内, 其对应的物理距离为4.84 Mb, 该区间有61个注释基因, 这为B2基因的精细定位和克隆奠定了基础(金迪等, 2019)。
小麦Hd基因与大麦HvKNOX3基因表型相似, 其突变体芒缩短、基部弯曲且膨大成膜状(Milner et al., 2019)。已发现小麦HvKNOX3的3个同源基因Wknox1a、Wknox1b和Wknox1d。Wknox1基因定位于第4同源群, 属于KNOX I类基因家族, 其内含子调控基因表达水平和组织特异性, 内含子突变导致表达水平升高或基因异位表达。定位于小麦4A染色体上的Wknox1a基因被确定为Hd的候选基因, 该基因第4内含子中有大量的转座子插入、复制、缺失和SNPs, 这些突变都可能导致Hd表型(Takumi et al., 2000; Morimoto et al., 2005)。
3.2 水稻芒的遗传学研究
在水稻驯化过程中, 由于无芒品种便于种子的收集和储存, 因此, 人类倾向于选育无芒的水稻品种。目前, 许多参与调控芒形成和伸长的基因在水稻中已被克隆, 如An-1、An-2、LABA1、GAD1、GLA、TOB1、DL、SHL2、SHO1、SHO2、WAF1和OsETT2 (表2) (Itoh et al., 2000; Satoh et al., 2003; Yamaguchi et al., 2004; Abe et al., 2010; Tanaka et al., 2012; Luo et al., 2013; Toriba and Hirano, 2014; Gu et al., 2015; Hua et al., 2015; Jin et al., 2016; Zhang et al., 2019)。Table 2
表2
表2水稻中已定位的控制芒功能的基因
Table 2
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| AN1 | 4 | 促进细胞分裂和芒原基形成、谷粒伸长及穗粒数减少 | Luo et al., 2013 |
| AN2 | 4 | 促进细胞分裂素的合成 | Gu et al., 2015 |
| LABA1 | 4 | 增强芒原基细胞分裂活性及促进芒伸长和芒刺形成 | Hua et al., 2015 |
| GAD1 | 8 | 促进芒原基细胞分裂和芒形成 | Jin et al., 2016 |
| GLA | 8 | 促进谷粒伸长 | Zhang et al., 2019 |
| TOB1 | 4 | 促进外稃和内稃形成和生长、分生组织的维持及花器官数量减少 | Tanaka et al., 2012 |
| DL | 3 | 分生组织细胞的激活、促进芒形成和伸长及调节花器官的发育 | Yamaguchi et al., 2004 |
| SHO2 | - | 参与TAS3途径中ta-siRNA的合成 | Toriba and Hirano, 2014 |
| SHL2 | 1 | 参与TAS3途径中ta-siRNA的合成 | Satoh et al., 2003 |
| SHO1 | 4 | 参与TAS3途径中ta-siRNA的合成 | Itoh et al., 2000 |
| WAF1 | 7 | 参与TAS3途径中ta-siRNA的合成 | Abe et al., 2010 |
| OsETT2 | 1 | 外稃生长活化 | Itoh et al., 2008 |
新窗口打开|下载CSV
水稻An-1基因位于4号染色体上, 编码1个bHLH转录因子, 可促进细胞分裂和芒原基形成。An-1基因上调表达导致芒和穗粒伸长以及穗粒数减少, 在水稻人工选择中, An-1是关键靶基因(Takumi et al., 2000)。An-2基因编码的OsLOGL6蛋白通过促进野生稻细胞分裂素合成而促进芒伸长(Gu et al., 2015)。研究表明, 在水稻芒的驯化过程中, An-1基因优先被驯化, An-2基因紧随其后(Zhang et al., 2019)。
水稻LABA1基因编码1个细胞分裂素激活酶(cytokinin riboside 5°-monophosphate phosphoribohydrolase), 该酶通过提高芒原基的活性细胞分裂素含量增强芒原基的细胞分裂素活性, 促进芒的伸长和芒刺形成(Hua et al., 2015)。GAD1基因编码1个分泌信号肽, 属于EPFL基因家族成员, 通过促进芒原基细胞分裂从而促进芒的形成(Jin et al., 2016)。GLA基因是GAD1/RAE2的新等位基因, 也是EPFL基因家族成员, 其编码区缺乏InDel3导致短芒或无芒表型。GLA是造成水稻品质差异的重要基因, 控制水稻的粒长和芒长, 在水稻遗传改良中发挥重要作用(Zhang et al., 2019)。
YABBY基因家族的TOB1和DL也参与调控水稻芒的形成。YABBY基因编码的转录因子具有螺旋-环-螺旋和锌指两种保守性。这些基因在植物花器官的发育和分生组织的维持中起重要调控作用。其中YABBY蛋白对分生组织起间接作用, YABBY蛋白产生的次级信号进入分生组织细胞并激活分裂信号 (Yamaguchi et al., 2004; Tanaka et al., 2012)。
TOB1基因位于水稻4号染色体, 编码OsYABBY5转录因子, 该基因突变有多种表型, 其中I类突变型直接表现为促进外稃上芒的形成(Tanaka et al., 2012)。OsYABBY5通过间接激活细胞分裂, 增强分生组织的活性, 启动外稃和内稃的形成和生长, 促进芒的形成。
水稻DL基因定位于3号染色体, 在叶原基中心区域的细胞和芒原基中表达, 通过诱导芒原基细胞分裂促进芒的伸长(Yamaguchi et al., 2004)。然而单独的DL基因不足以形成芒, 还需要OsETT2基因的协助。OsETT2指定芒原基细胞, DL激活芒原基中的细胞增殖, 使芒具有固有的形态(Toriba and Hirano, 2014)。
水稻中芒发育相关基因SHO2、SHL2、SHO1及WAF1编码的蛋白参与TAS3 ta-siRNA调控途径。该途径调控幼叶向成熟叶过渡及侧根生长发育等过程, OsETT2基因受该途径调控(Itoh et al., 2000, 2008; Satoh et al., 2003; Abe et al., 2010; Toriba and Hirano, 2014)。
3.3 大麦芒的遗传学研究
大麦也是研究芒性状遗传及作用的重要模式作物。野生型大麦多为具芒表型, 突变体主要表现为芒表型的变化或发育缺陷, 目前研究较多的基因为HvKNOX3、Lks2、SuK和ROUGH AWN1等(表3) (Müller et al., 1995, 2000; Santi et al., 2003; Roig et al., 2004; Yuo et al., 2012; Milner et al., 2019)。Table 3
表3
表3大麦中已定位的控制芒功能的基因
Table 3
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| HvKNOX3 | 4H | 分生组织的维持 | Müller et al., 1995, 2000; Santi et al., 2003 |
| Lks2 | 7H | 抑制芒伸长和控制雌蕊形态 | Yuo et al., 2012 |
| SuKD SuKB SuKC SuKE SuKF | 5H 7H 7H 7H 7H | 抑制HvKNOX3的表达 | Roig et al., 2004 |
| ROUGH AWN1 | 5H | 控制芒倒钩 | Milner et al., 2019 |
新窗口打开|下载CSV
大麦HvKNOX3基因位于4H染色体, HvKNOX3突变植株表型为反向的花取代芒, 这种表型被称为“帽子”。由于其启动子和第4个内含子序列中的(GA)8重复的存在, 进而结合BBR位点激活基因的表达, 因此内含子中BBR结合位点的(GA)8重复可能导致HvKNOX3表达过度激活。也有报道推测, (GA)8重复破坏了第4个内含子中沉默子的另一个调控序列, 进而使该沉默子失活, 导致HvKNOX3过表达(Müller et al., 1995, 2000; Santi et al., 2003)。通过研究HvKNOX3基因的上位基因, Lks2基因和SuK作为突变位点, Lks2和SuK基因产物通过与HvKNOX3第4内含子序列结合调控HvKNOX3基因的表达(Müller et al., 2000)。
大麦Lks2基因定位于7H染色体, 编码SHI基因家族的转录因子, 控制芒的长度和雌蕊形态, 显性纯合状态下, 芒的长度会缩短1/2。该基因编码的蛋白包含锌指结构和IGGH结构域(Yuo et al., 2012)。在HvKNOX3突变和Lks2隐性突变的植株中, 不存在“帽子”结构, 仅形成短芒表型。
SuK位点位于5H (SuKD)和7H (SuKB、SuKC、SuKE和SuKF)染色体上。与野生型大麦相比, 具有隐性突变的SuK植株和显性突变的HvKNOX3植株芒更短且不存在“帽子”表型。一般认为SuK无表型, 仅有抑制HvKNOX3的作用(Roig et al., 2004)。大麦ROUGH AWN1基因位于5H染色体, 控制芒表面倒钩性状, 其候选基因(HORVU5Hr1G086520)与水稻LABA1基因同源(Milner et al., 2019)。
4 研究展望
芒作为禾本科作物典型的穗部形态特征, 是不可忽视的关键性状。基于现有的研究, 探讨不同作物中芒发育相关基因之间如何进行互作, 以及不同物种中芒调控基因的作用方式及分子机制, 对于我们理解禾本科作物芒的产生和发育具有重要意义。在水稻中, An-1、An-2、LABA1、GAD1、GLA、TOB1和DL都通过促进细胞分裂或提高细胞分裂素的含量对芒发育或相关功能进行调控。在小麦中, 有证据表明B1可能通过抑制生长素相关途径而抑制芒的发育和细胞增殖。在大麦中, Lks2短芒突变通过影响生长素进而影响芒细胞的分裂。由此推测, 细胞分裂素及生长素的含量通过影响芒原基细胞分裂活性, 进而促进芒的伸长和芒刺形成, 是影响芒生长发育的主要因素。但禾本科作物特有器官——芒的产生和生长发育的分子机制还有待深入研究阐明。目前, 在水稻和大麦中已克隆多个控制芒发育的主效基因并进行了功能验证。但在小麦中, 受其庞大的基因组限制, 芒功能基因的定位与克隆进展缓慢。虽然已定位到部分主效基因, B1基因也已被克隆, 但小麦中控制芒性状的基因位点还有很多, B1基因的作用机制也尚存争议。随着普通小麦参考基因组数据的补充、基因组组装和注释的完善, 结合转录组测序、RNA-seq和重测序等多种高通量测序手段, 可实现对小麦功能基因快速定位, 这将促进小麦中芒重要功能基因的定位和功能研究。此外, 利用在全基因组水平上开发的大量资源以及在水稻、大麦中挖掘出的与芒相关的SNPs, 可寻找芒发育调控基因并探索其发生、发育和功能的分子机制。
目前, 对芒性状及其功能的研究已取得突破性进展, 如芒器官在光合、蒸腾中的作用及其对产量的贡献, 然而多方面限制条件使得芒相关研究的深度和广度还远远不够。未来值得关注的问题包括: 芒原基如何起始进而伸长, 芒中叶绿体的动态变化及其在光合作用中的功能, 光合基因表达与产物合成有何特异性, 如何开发和利用芒的光合和蒸腾特性, 芒对产量的影响及其影响产量的内在机制。这些问题都有待于进一步解答。因此, 加强芒的发生、发育和功能研究, 有利于充分了解其价值, 发挥其光合潜能, 对禾谷类作物的遗传改良具有十分重要的理论意义和应用价值。
(责任编辑: 白羽红)
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 2]
[本文引用: 1]
[本文引用: 4]
[本文引用: 2]
[本文引用: 1]
[本文引用: 4]
[本文引用: 6]
[本文引用: 2]
[本文引用: 3]
DOI:10.1007/s10142-009-0149-4URL [本文引用: 1]
DOI:10.3835/plantgenome.2009.07.0019URL [本文引用: 1]
[本文引用: 1]
DOI:10.1111/aab.1973.75.issue-2URL [本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 6]
DOI:10.1126/science.1140097URLPMID:17495170 [本文引用: 4]

The dispersal unit of wild wheat bears two pronounced awns that balance the unit as it falls. We discovered that the awns are also able to propel the seeds on and into the ground. The arrangement of cellulose fibrils causes bending of the awns with changes in humidity. Silicified hairs that cover the awns allow propulsion of the unit only in the direction of the seeds. This suggests that the dead tissue is analogous to a motor. Fueled by the daily humidity cycle, the awns induce the motility required for seed dispersal.
DOI:10.1111/aab.1972.70.issue-1URL [本文引用: 2]
[本文引用: 1]
DOI:10.1007/BF02860824URL [本文引用: 2]
URLPMID:26283047 [本文引用: 3]
DOI:10.1093/jxb/erw140URLPMID:27162273 [本文引用: 1]
DOI:10.1105/tpc.15.00260URLPMID:26082172 [本文引用: 3]
Common wild rice (Oryza rufipogon), the wild relative of Asian cultivated rice (Oryza sativa), flaunts long, barbed awns, which are necessary for efficient propagation and dissemination of seeds. By contrast, O. sativa cultivars have been selected to be awnless or to harbor short, barbless awns, which facilitate seed processing and storage. The transition from long, barbed awns to short, barbless awns was a crucial event in rice domestication. Here, we show that the presence of long, barbed awns in wild rice is controlled by a major gene on chromosome 4, LONG AND BARBED AWN1 (LABA1), which encodes a cytokinin-activating enzyme. A frame-shift deletion in LABA1 of cultivated rice reduces the cytokinin concentration in awn primordia, disrupting barb formation and awn elongation. Sequencing analysis demonstrated low nucleotide diversity and a selective sweep encompassing an approximately 800-kb region around the derived laba1 allele in cultivated rice. Haplotype analysis revealed that the laba1 allele originated in the japonica subspecies and moved into the indica gene pool via introgression, suggesting that humans selected for this locus in early rice domestication. Identification of LABA1 provides new insights into rice domestication and also sheds light on the molecular mechanism underlying awn development.
[本文引用: 5]
DOI:10.1104/pp.103.030437URLPMID:14681527 [本文引用: 1]
Several hundred Arabidopsis genes, transcriptionally regulated by phytochrome A (phyA), were previously identified using an oligonucleotide microarray. We have now identified, in silico, conserved sequence motifs in the promoters of these genes by comparing the promoter sequences to those of all the genes present on the microarray from which they were sampled. This was done using a Perl script (called Sift) that identifies over-represented motifs using an enumerative approach. The utility of Sift was verified by analysis of circadian-regulated promoters known to contain a biologically significant motif. Several elements were then identified in phyA-responsive promoters by their over-representation. Five previously undescribed motifs were detected in the promoters of phyA-induced genes. Four novel motifs were found in phyA-repressed promoters, plus a motif that strongly resembles the DE1 element. The G-box, CACGTG, was a prominent hit in both induced and repressed phyA-responsive promoters. Intriguingly, two distinct flanking consensus sequences were observed adjacent to the G-box core sequence: one predominating in phyA-induced promoters, the other in phyA-repressed promoters. Such different conserved flanking nucleotides around the core motif in these two sets of promoters may indicate that different members of the same family of DNA-binding proteins mediate phyA induction and repression. An increased abundance of G-box sequences was observed in the most rapidly phyA-responsive genes and in the promoters of phyA-regulated transcription factors, indicating that G-box-binding transcription factors are upstream components in a transcriptional cascade that mediates phyA-regulated development.
DOI:10.1105/tpc.12.11.2161URLPMID:11090216 [本文引用: 4]
The mechanism regulating the pattern of leaf initiation was analyzed by using shoot organization (sho) mutants derived from three loci (SHO1, SHO2, and SHO3). In the early vegetative phase, sho mutants show an increased rate of leaf production with random phyllotaxy. The resulting leaves are malformed, threadlike, or short and narrow. Their shoot apical meristems are relatively low and wide, that is, flat shaped, although their shape and size are highly variable among plants of the same genotype. Statistical analysis reveals that the shape of the shoot meristem rather than its size is closely correlated with the variations of plastochron and phyllotaxy. Rapid and random leaf production in sho mutants is correlated with the frequent and disorganized cell divisions in the shoot meristem and with a reduction of expression domain of a rice homeobox gene, OSH1. These changes in the organization and behavior of the shoot apical meristems suggest that sho mutants have fewer indeterminate cells and more determinate cells than wild type, with many cells acting as leaf founder cells. Thus, the SHO genes have an important role in maintaining the proper organization of the shoot apical meristem, which is essential for the normal initiation pattern of leaf primordia.
[本文引用: 1]
DOI:10.1105/tpc.16.00379URLPMID:27634315 [本文引用: 3]
Cultivated rice (Oryza sativa) was domesticated from wild rice (Oryza rufipogon), which typically displays fewer grains per panicle and longer grains than cultivated rice. In addition, wild rice has long awns, whereas cultivated rice has short awns or lacks them altogether. These changes represent critical events in rice domestication. Here, we identified a major gene, GRAIN NUMBER, GRAIN LENGTH AND AWN DEVELOPMENT1 (GAD1), that regulates those critical changes during rice domestication. GAD1 is located on chromosome 8 and is predicted to encode a small secretary signal peptide belonging to the EPIDERMAL PATTERNING FACTOR-LIKE family. A frame-shift insertion in gad1 destroyed the conserved cysteine residues of the peptide, resulting in a loss of function, and causing the increased number of grains per panicle, shorter grains, and awnless phenotype characteristic of cultivated rice. Our findings provide a useful paradigm for revealing functions of peptide signal molecules in plant development and helps elucidate the molecular basis of rice domestication.
[本文引用: 1]
DOI:10.1105/tpc.113.113589URLPMID:24076974 [本文引用: 3]
Long awns are important for seed dispersal in wild rice (Oryza rufipogon), but are absent in cultivated rice (Oryza sativa). The genetic mechanism involved in loss-of-awn in cultivated rice remains unknown. We report here the molecular cloning of a major quantitative trait locus, An-1, which regulates long awn formation in O. rufipogon. An-1 encodes a basic helix-loop-helix protein, which regulates cell division. The nearly-isogenic line (NIL-An-1) carrying a wild allele An-1 in the genetic background of the awnless indica Guangluai4 produces long awns and longer grains, but significantly fewer grains per panicle compared with Guangluai4. Transgenic studies confirmed that An-1 positively regulates awn elongation, but negatively regulates grain number per panicle. Genetic variations in the An-1 locus were found to be associated with awn loss in cultivated rice. Population genetic analysis of wild and cultivated rice showed a significant reduction in nucleotide diversity of the An-1 locus in rice cultivars, suggesting that the An-1 locus was a major target for artificial selection. Thus, we propose that awn loss was favored and strongly selected by humans, as genetic variations at the An-1 locus that cause awn loss would increase grain numbers and subsequently improve grain yield in cultivated rice.
DOI:10.1534/g3.114.012963URLPMID:25237112 [本文引用: 1]
MAGIC populations represent one of a new generation of crop genetic mapping resources combining high genetic recombination and diversity. We describe the creation and validation of an eight-parent MAGIC population consisting of 1091 F7 lines of winter-sown wheat (Triticum aestivum L.). Analyses based on genotypes from a 90,000-single nucleotide polymorphism (SNP) array find the population to be well-suited as a platform for fine-mapping quantitative trait loci (QTL) and gene isolation. Patterns of linkage disequilibrium (LD) show the population to be highly recombined; genetic marker diversity among the founders was 74% of that captured in a larger set of 64 wheat varieties, and 54% of SNPs segregating among the 64 lines also segregated among the eight founder lines. In contrast, a commonly used reference bi-parental population had only 54% of the diversity of the 64 varieties with 27% of SNPs segregating. We demonstrate the potential of this MAGIC resource by identifying a highly diagnostic marker for the morphological character
[本文引用: 2]
DOI:10.1038/s41588-018-0266-xURLPMID:30420647 [本文引用: 6]
Genebanks hold comprehensive collections of cultivars, landraces and crop wild relatives of all major food crops, but their detailed characterization has so far been limited to sparse core sets. The analysis of genome-wide genotyping-by-sequencing data for almost all barley accessions of the German ex situ genebank provides insights into the global population structure of domesticated barley and points out redundancies and coverage gaps in one of the world's major genebanks. Our large sample size and dense marker data afford great power for genome-wide association scans. We detect known and novel loci underlying morphological traits differentiating barley genepools, find evidence for convergent selection for barbless awns in barley and rice and show that a major-effect resistance locus conferring resistance to bymovirus infection has been favored by traditional farmers. This study outlines future directions for genomics-assisted genebank management and the utilization of germplasm collections for linking natural variation to human selection during crop evolution.
[本文引用: 2]
DOI:10.1071/AR02008URL [本文引用: 1]
DOI:10.1007/s004240000073URL [本文引用: 3]
[本文引用: 3]
DOI:10.1111/aab.1978.90.issue-1URL [本文引用: 1]
[本文引用: 1]
[本文引用: 2]
DOI:10.1093/jxb/erw081URLPMID:26976817 [本文引用: 2]
Genotypic variation in ear morphology is linked to differences in photosynthetic potential to influence grain yield in winter cereals. Awns contribute to photosynthesis, particularly under water-limited conditions when canopy assimilation is restricted. We assessed performance of up to 45 backcross-derived, awned-awnletted NILs representing four diverse genetic backgrounds in 25 irrigated or rainfed, and droughted environments in Australia and Mexico. Mean environment grain yields were wide-ranging (1.38-7.93 t ha(-1)) with vegetative and maturity biomass, plant height, anthesis date, spike number, and harvest index all similar (P >0.05) for awned and awnletted NILs. Overall, grain yields of awned-awnletted sister-NILs were equivalent, irrespective of yield potential and genetic background. Awnletted wheats produced significantly more grains per unit area (+4%) and per spike (+5%) reflecting more fertile spikelets and grains in tertiary florets. Increases in grain number were compensated for by significant reductions in grain size (-5%) and increased frequency (+0.8%) of small, shrivelled grains ('screenings') to reduce seed-lot quality of awnletted NILs. Post-anthesis canopies of awnletted NILs were marginally warmer over all environments (+0.27 degrees C) but were not different and were sometimes cooler than awned NILs at cooler air temperatures. Awns develop early and represented up to 40% of total spikelet biomass prior to ear emergence. We hypothesize that the allocation of assimilate to large and rapidly developing awns decreases spikelet number and floret fertility to reduce grain number, particularly in distal florets. Individual grain size is increased to reduce screenings and to increase test weight and milling quality, particularly in droughted environments. Despite the average reduction in grain size, awnless lines could be identified that combined higher grain yield with larger grain size, increased grain protein concentration, and reduced screenings.
[本文引用: 3]
[本文引用: 3]
[本文引用: 3]
[本文引用: 2]
[本文引用: 1]
DOI:10.1016/s0378-1119(00)00164-5URLPMID:10831851 [本文引用: 2]
The plant knotted1 (kn1)-like homeobox genes are known to play important roles in the maintenance of shoot apical meristem (SAM), determination of cell fate and differentiation of vegetative tissues. To study structural diversity of the three homologous loci encoding a KN1-like homeobox protein in the hexaploid wheat genome, we isolated clones from a cDNA library of young spikes of Japanese common wheat cultivar 'Norin 26'. Three different but highly homologous cDNAs were isolated and their sequences were determined. The mean homology of the deduced amino acid sequences was 96% as compared to the barley ortholog KNOX3. The wheat kn1-like homeobox proteins named WKNOX1 are encoded by a single set of homologous genes on the homologous group 4 chromosomes in the three component genomes of common wheat, i.e. 4A, 4B and 4D. The nucleotide sequence data and the Southern blot pattern suggested that the three homologous loci of wknox1 genes are highly conserved through polyploid evolution of wheat. They were expressed in SAM-containing shoots and young spikes but not in developed leaves, glumes and lemmas and callus tissues. The ectopic expression of the wknox1 was observed in lemma of wheat Hooded (Hd) mutants. The result suggested that the Hd gene is a dominant allele of the wknox1 locus on chromosome 4A.
[本文引用: 4]
DOI:10.4141/cjps72-165URL [本文引用: 3]
[本文引用: 4]
[本文引用: 7]
[本文引用: 2]
DOI:10.1104/pp.105.075366URLPMID:16760496 [本文引用: 1]
[本文引用: 4]
DOI:10.1371/journal.pone.0176148URLPMID:28437453 [本文引用: 2]
The awn is a long needle-like structure formed at the tip of the lemma in the florets of some grass species. It plays a role in seed dispersal and protection against animals, and can contribute to the photosynthetic activity of spikes. Three main dominant inhibitors of awn development (Hd, B1 and B2) are known in hexaploid wheat, but the causal genes have not been cloned yet and a genetic association with awn length diversity has been found only for the B1 allele. To analyze the prevalence of these three awning inhibitors, we attempted to predict the genotypes of 189 hexaploid wheat varieties collected worldwide using markers tightly linked to these loci. Using recombinant inbred lines derived from two common wheat cultivars, Chinese Spring and Mironovskaya 808, both with short awns, and a high-density linkage map, we performed quantitative trait locus analysis to identify tightly linked markers. Because this linkage map was constructed with abundant array-based markers, we converted the linked markers to PCR-based markers and determined the genotypes of 189 hexaploids. A significant genotype-phenotype correlation was observed at the Hd and B1 regions. We also found that interaction among these three awning inhibitors is involved in development of a membranous outgrowth at the base of awn resembling the Hooded mutants of barley. For the hooded awn phenotype, presence of the Hd dominant allele was essential but not sufficient, so B2 and other factors appear to act epistatically to produce the ectopic tissue. On the other hand, the dominant B1 allele acted as a suppressor of the hooded phenotype. These three awning inhibitors largely contribute to the genetic variation in awn length and shape of common wheat.
[本文引用: 3]
[本文引用: 4]
小麦芒的研究
2
2010
... 芒中的叶绿体与叶片中的叶绿体具有相似的结构和功能.芒作为穗部重要的光合器官, 与叶片相比, 在光合作用上具有许多优势.首先, 芒的光合面积大, 且着生于穗部顶端, 利于截获光能和气体(
... 芒表面的气孔占穗部气孔数的1/2.在相同条件下, 去芒穗的蒸腾作用比未去芒穗低约75%.在炎热干旱条件下, 有芒植株通过增强蒸腾作用, 迅速降低温度, 增强吸水动力, 促进根系下扎土壤以吸收更多的水分和矿物质, 提高水分利用率, 促进水分向穗部转运, 增强植株抗旱能力(
小麦芒的功能及去芒对籽粒重的影响
1
1981
... 研究表明, 有芒作物的产量一般显著高于同类无芒作物, 增产约1/10 (
小麦芒长抑制基因B1近等基因系的鉴定及遗传分析
4
2010
... 芒是禾本科植物小花外稃或颖片顶端的一种针状延伸, 从基部到顶端逐渐变细.其表皮含有不同形态的细胞, 如较狭长的细胞、卵状或方形的短细胞和厚壁而尖细的单细胞.其中较狭长的细胞具有波状加厚的壁; 卵状或方形的短细胞表现为芒表皮上的乳头状突起; 厚壁而尖细的单细胞向前斜生, 使得芒表面较为粗糙(
... ;
... 芒的横切面一般呈三角形.例如, 小麦(Triticum aestivum)芒的横切面为锐角三角形, 大麦(Hordeum vulgare)为钝角三角形.其横切面内部含有3条维管束, 中间较粗, 两边较细.中间的维管束具有完整的结构, 其外有2层维管束鞘, 内有木质部和韧皮部.外鞘细胞中与绿色组织相连的部分含有少量叶绿体, 不与绿色组织相连的外鞘细胞及内鞘细胞不含叶绿体.木质部中有少量薄壁细胞和厚壁导管.韧皮部中有筛管与伴胞, 且一一对应(
... 芒中的叶绿体与叶片中的叶绿体具有相似的结构和功能.芒作为穗部重要的光合器官, 与叶片相比, 在光合作用上具有许多优势.首先, 芒的光合面积大, 且着生于穗部顶端, 利于截获光能和气体(
小麦芒长抑制基因B2的精细定位与候选基因分析
2
2019
... The genes involved in the function of the awn in Triticum aestivum identified by genetic mapping
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| B1 | 5AL | 分生组织的 维持 | |
| B2 | 6BL | 抑制芒伸长 | |
| Hd (Wknox1a) | 4AL | 抑制芒伸长 | |
| Wknox1b Wknox1d | 4BS 4DS | 茎尖分生组织的形成和维持 |
... 小麦B2基因表型为整个穗部的短芒.一般情况下, 穗部各部分的芒长一致, 少数情况下会出现穗中部芒比较长(
小麦芒和旗叶叶绿体结构及低温荧光发射光谱的比较研究
1
2002
... 芒中的叶绿体与叶片中的叶绿体具有相似的结构和功能.芒作为穗部重要的光合器官, 与叶片相比, 在光合作用上具有许多优势.首先, 芒的光合面积大, 且着生于穗部顶端, 利于截获光能和气体(
高产小麦旗叶和芒细胞结构及其光合性能的研究
4
2006
... 芒是禾本科植物小花外稃或颖片顶端的一种针状延伸, 从基部到顶端逐渐变细.其表皮含有不同形态的细胞, 如较狭长的细胞、卵状或方形的短细胞和厚壁而尖细的单细胞.其中较狭长的细胞具有波状加厚的壁; 卵状或方形的短细胞表现为芒表皮上的乳头状突起; 厚壁而尖细的单细胞向前斜生, 使得芒表面较为粗糙(
... 芒的横切面一般呈三角形.例如, 小麦(Triticum aestivum)芒的横切面为锐角三角形, 大麦(Hordeum vulgare)为钝角三角形.其横切面内部含有3条维管束, 中间较粗, 两边较细.中间的维管束具有完整的结构, 其外有2层维管束鞘, 内有木质部和韧皮部.外鞘细胞中与绿色组织相连的部分含有少量叶绿体, 不与绿色组织相连的外鞘细胞及内鞘细胞不含叶绿体.木质部中有少量薄壁细胞和厚壁导管.韧皮部中有筛管与伴胞, 且一一对应(
... 在小麦和大麦芒绿色组织中, 绿色细胞常呈不规则形状, 含有丰富的叶绿体.不同时期其叶绿体形态结构存在明显变化(
... 芒中的叶绿体与叶片中的叶绿体具有相似的结构和功能.芒作为穗部重要的光合器官, 与叶片相比, 在光合作用上具有许多优势.首先, 芒的光合面积大, 且着生于穗部顶端, 利于截获光能和气体(
麦芒的结构及其光合特性
6
1993
... 芒是禾本科作物穗部的重要结构, 属于叶的变态, 是植物长期适应环境和进化的产物.不同材料存在芒的有无及芒长度的差异, 可作为区分禾谷类作物不同品种以及基因定位的重要形态标记(
... 芒是禾本科植物小花外稃或颖片顶端的一种针状延伸, 从基部到顶端逐渐变细.其表皮含有不同形态的细胞, 如较狭长的细胞、卵状或方形的短细胞和厚壁而尖细的单细胞.其中较狭长的细胞具有波状加厚的壁; 卵状或方形的短细胞表现为芒表皮上的乳头状突起; 厚壁而尖细的单细胞向前斜生, 使得芒表面较为粗糙(
... ).芒的近轴端无气孔和绿色细胞, 是芒的非绿色区域, 其表皮细胞细胞壁明显增厚, 这对于减少水分散失和维持芒的形态起到重要作用(
... 芒的横切面一般呈三角形.例如, 小麦(Triticum aestivum)芒的横切面为锐角三角形, 大麦(Hordeum vulgare)为钝角三角形.其横切面内部含有3条维管束, 中间较粗, 两边较细.中间的维管束具有完整的结构, 其外有2层维管束鞘, 内有木质部和韧皮部.外鞘细胞中与绿色组织相连的部分含有少量叶绿体, 不与绿色组织相连的外鞘细胞及内鞘细胞不含叶绿体.木质部中有少量薄壁细胞和厚壁导管.韧皮部中有筛管与伴胞, 且一一对应(
... 在小麦和大麦芒绿色组织中, 绿色细胞常呈不规则形状, 含有丰富的叶绿体.不同时期其叶绿体形态结构存在明显变化(
... 芒中的叶绿体与叶片中的叶绿体具有相似的结构和功能.芒作为穗部重要的光合器官, 与叶片相比, 在光合作用上具有许多优势.首先, 芒的光合面积大, 且着生于穗部顶端, 利于截获光能和气体(
不同供水条件下小麦不同绿色器官的气孔特性研究
2
2006
... 气孔的主要作用是与外界进行气体交换, 与旗叶相比, 芒的气孔总数少且体积小(
... 芒表面的气孔占穗部气孔数的1/2.在相同条件下, 去芒穗的蒸腾作用比未去芒穗低约75%.在炎热干旱条件下, 有芒植株通过增强蒸腾作用, 迅速降低温度, 增强吸水动力, 促进根系下扎土壤以吸收更多的水分和矿物质, 提高水分利用率, 促进水分向穗部转运, 增强植株抗旱能力(
WAVY LEAF1, an ortholog of Arabidopsis HEN1, regulates shoot development by maintaining microRNA and trans-acting small interfering RNA accumulation in rice.
3
2010
... 在水稻驯化过程中, 由于无芒品种便于种子的收集和储存, 因此, 人类倾向于选育无芒的水稻品种.目前, 许多参与调控芒形成和伸长的基因在水稻中已被克隆, 如An-1、An-2、LABA1、GAD1、GLA、TOB1、DL、SHL2、SHO1、SHO2、WAF1和OsETT2 (
... The genes involved in the function of the awn in Oryza sativa identified by genetic mapping
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| AN1 | 4 | 促进细胞分裂和芒原基形成、谷粒伸长及穗粒数减少 | |
| AN2 | 4 | 促进细胞分裂素的合成 | |
| LABA1 | 4 | 增强芒原基细胞分裂活性及促进芒伸长和芒刺形成 | |
| GAD1 | 8 | 促进芒原基细胞分裂和芒形成 | |
| GLA | 8 | 促进谷粒伸长 | |
| TOB1 | 4 | 促进外稃和内稃形成和生长、分生组织的维持及花器官数量减少 | |
| DL | 3 | 分生组织细胞的激活、促进芒形成和伸长及调节花器官的发育 | |
| SHO2 | - | 参与TAS3途径中ta-siRNA的合成 | |
| SHL2 | 1 | 参与TAS3途径中ta-siRNA的合成 | |
| SHO1 | 4 | 参与TAS3途径中ta-siRNA的合成 | |
| WAF1 | 7 | 参与TAS3途径中ta-siRNA的合成 | |
| OsETT2 | 1 | 外稃生长活化 |
... 水稻中芒发育相关基因SHO2、SHL2、SHO1及WAF1编码的蛋白参与TAS3 ta-siRNA调控途径.该途径调控幼叶向成熟叶过渡及侧根生长发育等过程, OsETT2基因受该途径调控(
Drought response in the spikes of barley: gene expression in the lemma, palea, awn, and seed
1
2010
... 芒表面的气孔占穗部气孔数的1/2.在相同条件下, 去芒穗的蒸腾作用比未去芒穗低约75%.在炎热干旱条件下, 有芒植株通过增强蒸腾作用, 迅速降低温度, 增强吸水动力, 促进根系下扎土壤以吸收更多的水分和矿物质, 提高水分利用率, 促进水分向穗部转运, 增强植株抗旱能力(
Comparative transcriptional profiling established the awn as the major photosynthetic organ of the barley spike while the lemma and the palea primarily protect the seed
1
2009
... 芒中的叶绿体与叶片中的叶绿体具有相似的结构和功能.芒作为穗部重要的光合器官, 与叶片相比, 在光合作用上具有许多优势.首先, 芒的光合面积大, 且着生于穗部顶端, 利于截获光能和气体(
Identification of the genes promoting awnedness in the Triticum aestivum/Aegilops umbellulata introgressive line.
1
2012
... 在普通小麦中, B1 (5AL)、B2 (6BL)和Hd (4AL)是目前已知的芒显性抑制基因, 它们的不同组合导致芒表型发生变化(
Stomatal control of gas exchange in barley awns
1
1973
... 研究表明, 有芒作物的产量一般显著高于同类无芒作物, 增产约1/10 (
Static diffusion of gases and liquids in relation to the assimilation of carbon and translocation in plants
1
1901
... 气孔的主要作用是与外界进行气体交换, 与旗叶相比, 芒的气孔总数少且体积小(
Molecular mapping of quantitative trait loci for yield and yield components in spring wheat (Triticum aestivum L.).
2
2008
... 芒是禾本科作物穗部的重要结构, 属于叶的变态, 是植物长期适应环境和进化的产物.不同材料存在芒的有无及芒长度的差异, 可作为区分禾谷类作物不同品种以及基因定位的重要形态标记(
... 芒是禾谷类作物一种重要的驯化性状, 其表面的倒刺可以附着在动物皮毛上, 借助动物的运动进行传播(
Sequence-based mapping identifies a candidate transcription repressor underlying awn suppression at the B1 locus in wheat.
6
2020
... The genes involved in the function of the awn in Triticum aestivum identified by genetic mapping
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| B1 | 5AL | 分生组织的 维持 | |
| B2 | 6BL | 抑制芒伸长 | |
| Hd (Wknox1a) | 4AL | 抑制芒伸长 | |
| Wknox1b Wknox1d | 4BS 4DS | 茎尖分生组织的形成和维持 |
... 研究表明, 小麦B1基因对芒长度的影响最强, 其突变体的表型为穗基部和中部产生短芒或无芒, 芒最长仅为1 cm (
... 基因与小麦小穗数、粒重和容重显著相关(
... 美国北卡罗来纳州立大学的Gina Brown-Guedira团队通过对14 567个GBS标记进行关联分析, 将B1位点定位于5A染色体681 455 268-706 705 101 bp之间25 Mb的区间内, 其中SNP (5A28417)位点与小麦品系中芒的表型共分离且与小麦容重显著关联, 该位点位于预测基因TraesCS5A02G542800上游219 bp处.小麦有芒突变体在5AL远端区域发生缺失, 缺失片段包含候选基因TraesCS5A02G542700和TraesCS5A02G542800, 利用特异标记分析最终确定TraesCS5A02G542800为B1的候选基因(
... ALI-1编码1个C2H2锌指转录因子, 具有转录抑制活性.对B1近等基因系的芒进行纵切观察, 结果显示两者细胞长度无显著差异, 但短芒材料细胞数目仅为长芒材料的1/10, 表明短芒材料芒原基的细胞分裂被抑制.顺式调控元件预测结果表明, ALI-1区间内SNPs导致该基因缺失顺式元件BOXCPSAS1、LTRE1HVBLT49、SORLIP2AT和SITEIIATCYTC, 这些顺式元件参与调控分生组织和增殖细胞的基因表达(
... 截至目前, 虽然B1基因已被克隆, 但其表达和抑芒机制仍然未知.目前, 对其突变位点及功能研究仍然存在较大争议, 还需要进一步的实验验证(
The role of wheat awns in the seed dispersal unit
4
2007
... 芒是禾本科作物穗部的重要结构, 属于叶的变态, 是植物长期适应环境和进化的产物.不同材料存在芒的有无及芒长度的差异, 可作为区分禾谷类作物不同品种以及基因定位的重要形态标记(
... 芒是禾谷类作物一种重要的驯化性状, 其表面的倒刺可以附着在动物皮毛上, 借助动物的运动进行传播(
... 具芒种子下扎土壤情况(改自
... d: 深度
Effect of awns and drought on the supply of photosynthate and its distribution within wheat ears
2
1972
... 芒中的叶绿体与叶片中的叶绿体具有相似的结构和功能.芒作为穗部重要的光合器官, 与叶片相比, 在光合作用上具有许多优势.首先, 芒的光合面积大, 且着生于穗部顶端, 利于截获光能和气体(
... ).在干旱条件下, 叶片的光合速率显著降低, 而芒的光合速率仍能保持相对稳定的状态, 因此干旱条件下有芒小麦相较于无芒小麦增产约1/4 (
Photosynthesis and respiration by the flag leaf and components of the ear during grain development in wheat
1
1970
... 芒中的叶绿体与叶片中的叶绿体具有相似的结构和功能.芒作为穗部重要的光合器官, 与叶片相比, 在光合作用上具有许多优势.首先, 芒的光合面积大, 且着生于穗部顶端, 利于截获光能和气体(
The physiological function of the cereal awn
2
1963
... 芒是禾本科作物穗部的重要结构, 属于叶的变态, 是植物长期适应环境和进化的产物.不同材料存在芒的有无及芒长度的差异, 可作为区分禾谷类作物不同品种以及基因定位的重要形态标记(
... 芒是禾谷类作物一种重要的驯化性状, 其表面的倒刺可以附着在动物皮毛上, 借助动物的运动进行传播(
An-2 encodes a cytokinin synthesis enzyme that regulates awn length and grain production in rice
3
2015
... 在水稻驯化过程中, 由于无芒品种便于种子的收集和储存, 因此, 人类倾向于选育无芒的水稻品种.目前, 许多参与调控芒形成和伸长的基因在水稻中已被克隆, 如An-1、An-2、LABA1、GAD1、GLA、TOB1、DL、SHL2、SHO1、SHO2、WAF1和OsETT2 (
... The genes involved in the function of the awn in Oryza sativa identified by genetic mapping
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| AN1 | 4 | 促进细胞分裂和芒原基形成、谷粒伸长及穗粒数减少 | |
| AN2 | 4 | 促进细胞分裂素的合成 | |
| LABA1 | 4 | 增强芒原基细胞分裂活性及促进芒伸长和芒刺形成 | |
| GAD1 | 8 | 促进芒原基细胞分裂和芒形成 | |
| GLA | 8 | 促进谷粒伸长 | |
| TOB1 | 4 | 促进外稃和内稃形成和生长、分生组织的维持及花器官数量减少 | |
| DL | 3 | 分生组织细胞的激活、促进芒形成和伸长及调节花器官的发育 | |
| SHO2 | - | 参与TAS3途径中ta-siRNA的合成 | |
| SHL2 | 1 | 参与TAS3途径中ta-siRNA的合成 | |
| SHO1 | 4 | 参与TAS3途径中ta-siRNA的合成 | |
| WAF1 | 7 | 参与TAS3途径中ta-siRNA的合成 | |
| OsETT2 | 1 | 外稃生长活化 |
... 水稻An-1基因位于4号染色体上, 编码1个bHLH转录因子, 可促进细胞分裂和芒原基形成.An-1基因上调表达导致芒和穗粒伸长以及穗粒数减少, 在水稻人工选择中, An-1是关键靶基因(
Costs and benefits of awns
1
2016
... 研究表明, 有芒作物的产量一般显著高于同类无芒作物, 增产约1/10 (
LABA1, a domestication gene associated with long, barbed awns in wild rice
3
2015
... 在水稻驯化过程中, 由于无芒品种便于种子的收集和储存, 因此, 人类倾向于选育无芒的水稻品种.目前, 许多参与调控芒形成和伸长的基因在水稻中已被克隆, 如An-1、An-2、LABA1、GAD1、GLA、TOB1、DL、SHL2、SHO1、SHO2、WAF1和OsETT2 (
... The genes involved in the function of the awn in Oryza sativa identified by genetic mapping
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| AN1 | 4 | 促进细胞分裂和芒原基形成、谷粒伸长及穗粒数减少 | |
| AN2 | 4 | 促进细胞分裂素的合成 | |
| LABA1 | 4 | 增强芒原基细胞分裂活性及促进芒伸长和芒刺形成 | |
| GAD1 | 8 | 促进芒原基细胞分裂和芒形成 | |
| GLA | 8 | 促进谷粒伸长 | |
| TOB1 | 4 | 促进外稃和内稃形成和生长、分生组织的维持及花器官数量减少 | |
| DL | 3 | 分生组织细胞的激活、促进芒形成和伸长及调节花器官的发育 | |
| SHO2 | - | 参与TAS3途径中ta-siRNA的合成 | |
| SHL2 | 1 | 参与TAS3途径中ta-siRNA的合成 | |
| SHO1 | 4 | 参与TAS3途径中ta-siRNA的合成 | |
| WAF1 | 7 | 参与TAS3途径中ta-siRNA的合成 | |
| OsETT2 | 1 | 外稃生长活化 |
... 水稻LABA1基因编码1个细胞分裂素激活酶(cytokinin riboside 5°-monophosphate phosphoribohydrolase), 该酶通过提高芒原基的活性细胞分裂素含量增强芒原基的细胞分裂素活性, 促进芒的伸长和芒刺形成(
Dominant inhibition of awn development by a putative zinc-finger transcriptional repressor expressed at the B1 locus in wheat.
5
2020
... The genes involved in the function of the awn in Triticum aestivum identified by genetic mapping
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| B1 | 5AL | 分生组织的 维持 | |
| B2 | 6BL | 抑制芒伸长 | |
| Hd (Wknox1a) | 4AL | 抑制芒伸长 | |
| Wknox1b Wknox1d | 4BS 4DS | 茎尖分生组织的形成和维持 |
... 研究表明, 小麦B1基因对芒长度的影响最强, 其突变体的表型为穗基部和中部产生短芒或无芒, 芒最长仅为1 cm (
... 美国北卡罗来纳州立大学的Gina Brown-Guedira团队通过对14 567个GBS标记进行关联分析, 将B1位点定位于5A染色体681 455 268-706 705 101 bp之间25 Mb的区间内, 其中SNP (5A28417)位点与小麦品系中芒的表型共分离且与小麦容重显著关联, 该位点位于预测基因TraesCS5A02G542800上游219 bp处.小麦有芒突变体在5AL远端区域发生缺失, 缺失片段包含候选基因TraesCS5A02G542700和TraesCS5A02G542800, 利用特异标记分析最终确定TraesCS5A02G542800为B1的候选基因(
... ALI-1编码1个C2H2锌指转录因子, 具有转录抑制活性.对B1近等基因系的芒进行纵切观察, 结果显示两者细胞长度无显著差异, 但短芒材料细胞数目仅为长芒材料的1/10, 表明短芒材料芒原基的细胞分裂被抑制.顺式调控元件预测结果表明, ALI-1区间内SNPs导致该基因缺失顺式元件BOXCPSAS1、LTRE1HVBLT49、SORLIP2AT和SITEIIATCYTC, 这些顺式元件参与调控分生组织和增殖细胞的基因表达(
... 截至目前, 虽然B1基因已被克隆, 但其表达和抑芒机制仍然未知.目前, 对其突变位点及功能研究仍然存在较大争议, 还需要进一步的实验验证(
Identification of promoter motifs involved in the network of phytochrome a-regulated gene expression by combined analysis of genomic sequence and microarray data
1
2003
... ALI-1编码1个C2H2锌指转录因子, 具有转录抑制活性.对B1近等基因系的芒进行纵切观察, 结果显示两者细胞长度无显著差异, 但短芒材料细胞数目仅为长芒材料的1/10, 表明短芒材料芒原基的细胞分裂被抑制.顺式调控元件预测结果表明, ALI-1区间内SNPs导致该基因缺失顺式元件BOXCPSAS1、LTRE1HVBLT49、SORLIP2AT和SITEIIATCYTC, 这些顺式元件参与调控分生组织和增殖细胞的基因表达(
SHOOT ORGANIZATION genes regulate shoot apical meristem organization and the pattern of leaf primordium initiation in rice
4
2000
... 在水稻驯化过程中, 由于无芒品种便于种子的收集和储存, 因此, 人类倾向于选育无芒的水稻品种.目前, 许多参与调控芒形成和伸长的基因在水稻中已被克隆, 如An-1、An-2、LABA1、GAD1、GLA、TOB1、DL、SHL2、SHO1、SHO2、WAF1和OsETT2 (
... The genes involved in the function of the awn in Oryza sativa identified by genetic mapping
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| AN1 | 4 | 促进细胞分裂和芒原基形成、谷粒伸长及穗粒数减少 | |
| AN2 | 4 | 促进细胞分裂素的合成 | |
| LABA1 | 4 | 增强芒原基细胞分裂活性及促进芒伸长和芒刺形成 | |
| GAD1 | 8 | 促进芒原基细胞分裂和芒形成 | |
| GLA | 8 | 促进谷粒伸长 | |
| TOB1 | 4 | 促进外稃和内稃形成和生长、分生组织的维持及花器官数量减少 | |
| DL | 3 | 分生组织细胞的激活、促进芒形成和伸长及调节花器官的发育 | |
| SHO2 | - | 参与TAS3途径中ta-siRNA的合成 | |
| SHL2 | 1 | 参与TAS3途径中ta-siRNA的合成 | |
| SHO1 | 4 | 参与TAS3途径中ta-siRNA的合成 | |
| WAF1 | 7 | 参与TAS3途径中ta-siRNA的合成 | |
| OsETT2 | 1 | 外稃生长活化 |
...
... 水稻中芒发育相关基因SHO2、SHL2、SHO1及WAF1编码的蛋白参与TAS3 ta-siRNA调控途径.该途径调控幼叶向成熟叶过渡及侧根生长发育等过程, OsETT2基因受该途径调控(
The SHOOT ORGANIZATION 2 gene coordinates leaf domain development along the central-marginal axis in rice.
1
2008
... 水稻中芒发育相关基因SHO2、SHL2、SHO1及WAF1编码的蛋白参与TAS3 ta-siRNA调控途径.该途径调控幼叶向成熟叶过渡及侧根生长发育等过程, OsETT2基因受该途径调控(
GAD1 encodes a secreted peptide that regulates grain number, grain length, and awn development in rice domestication
3
2016
... 在水稻驯化过程中, 由于无芒品种便于种子的收集和储存, 因此, 人类倾向于选育无芒的水稻品种.目前, 许多参与调控芒形成和伸长的基因在水稻中已被克隆, 如An-1、An-2、LABA1、GAD1、GLA、TOB1、DL、SHL2、SHO1、SHO2、WAF1和OsETT2 (
... The genes involved in the function of the awn in Oryza sativa identified by genetic mapping
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| AN1 | 4 | 促进细胞分裂和芒原基形成、谷粒伸长及穗粒数减少 | |
| AN2 | 4 | 促进细胞分裂素的合成 | |
| LABA1 | 4 | 增强芒原基细胞分裂活性及促进芒伸长和芒刺形成 | |
| GAD1 | 8 | 促进芒原基细胞分裂和芒形成 | |
| GLA | 8 | 促进谷粒伸长 | |
| TOB1 | 4 | 促进外稃和内稃形成和生长、分生组织的维持及花器官数量减少 | |
| DL | 3 | 分生组织细胞的激活、促进芒形成和伸长及调节花器官的发育 | |
| SHO2 | - | 参与TAS3途径中ta-siRNA的合成 | |
| SHL2 | 1 | 参与TAS3途径中ta-siRNA的合成 | |
| SHO1 | 4 | 参与TAS3途径中ta-siRNA的合成 | |
| WAF1 | 7 | 参与TAS3途径中ta-siRNA的合成 | |
| OsETT2 | 1 | 外稃生长活化 |
... 水稻LABA1基因编码1个细胞分裂素激活酶(cytokinin riboside 5°-monophosphate phosphoribohydrolase), 该酶通过提高芒原基的活性细胞分裂素含量增强芒原基的细胞分裂素活性, 促进芒的伸长和芒刺形成(
Regulatory dephosphorylation of CDK at G2/M in plants: yeast mitotic phosphatase cdc25 induces cytokinin-like effects in transgenic tobacco morphogenesis.
1
2011
... ALI-1编码1个C2H2锌指转录因子, 具有转录抑制活性.对B1近等基因系的芒进行纵切观察, 结果显示两者细胞长度无显著差异, 但短芒材料细胞数目仅为长芒材料的1/10, 表明短芒材料芒原基的细胞分裂被抑制.顺式调控元件预测结果表明, ALI-1区间内SNPs导致该基因缺失顺式元件BOXCPSAS1、LTRE1HVBLT49、SORLIP2AT和SITEIIATCYTC, 这些顺式元件参与调控分生组织和增殖细胞的基因表达(
An-1 encodes a basic helix-loop-helix protein that regulates awn development, grain size, and grain number in rice
3
2013
... 研究表明, 有芒作物的产量一般显著高于同类无芒作物, 增产约1/10 (
... 在水稻驯化过程中, 由于无芒品种便于种子的收集和储存, 因此, 人类倾向于选育无芒的水稻品种.目前, 许多参与调控芒形成和伸长的基因在水稻中已被克隆, 如An-1、An-2、LABA1、GAD1、GLA、TOB1、DL、SHL2、SHO1、SHO2、WAF1和OsETT2 (
... The genes involved in the function of the awn in Oryza sativa identified by genetic mapping
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| AN1 | 4 | 促进细胞分裂和芒原基形成、谷粒伸长及穗粒数减少 | |
| AN2 | 4 | 促进细胞分裂素的合成 | |
| LABA1 | 4 | 增强芒原基细胞分裂活性及促进芒伸长和芒刺形成 | |
| GAD1 | 8 | 促进芒原基细胞分裂和芒形成 | |
| GLA | 8 | 促进谷粒伸长 | |
| TOB1 | 4 | 促进外稃和内稃形成和生长、分生组织的维持及花器官数量减少 | |
| DL | 3 | 分生组织细胞的激活、促进芒形成和伸长及调节花器官的发育 | |
| SHO2 | - | 参与TAS3途径中ta-siRNA的合成 | |
| SHL2 | 1 | 参与TAS3途径中ta-siRNA的合成 | |
| SHO1 | 4 | 参与TAS3途径中ta-siRNA的合成 | |
| WAF1 | 7 | 参与TAS3途径中ta-siRNA的合成 | |
| OsETT2 | 1 | 外稃生长活化 |
An eight-parent multiparent advanced generation inter- cross population for winter-sown wheat: creation, properties, and validation
1
2014
... 在普通小麦中, B1 (5AL)、B2 (6BL)和Hd (4AL)是目前已知的芒显性抑制基因, 它们的不同组合导致芒表型发生变化(
The contribution of the awns of bread wheat (Triticum aestivum L.) to grain filling: responses to water deficit and the effects of awns on ear temperature and hydraulic conductance.
2
2014
... 气孔的主要作用是与外界进行气体交换, 与旗叶相比, 芒的气孔总数少且体积小(
... 芒表面的气孔占穗部气孔数的1/2.在相同条件下, 去芒穗的蒸腾作用比未去芒穗低约75%.在炎热干旱条件下, 有芒植株通过增强蒸腾作用, 迅速降低温度, 增强吸水动力, 促进根系下扎土壤以吸收更多的水分和矿物质, 提高水分利用率, 促进水分向穗部转运, 增强植株抗旱能力(
Genebank genomics highlights the diversity of a global barley collection
6
2019
... 研究表明, 有芒作物的产量一般显著高于同类无芒作物, 增产约1/10 (
... The genes involved in the function of the awn in Triticum aestivum identified by genetic mapping
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| B1 | 5AL | 分生组织的 维持 | |
| B2 | 6BL | 抑制芒伸长 | |
| Hd (Wknox1a) | 4AL | 抑制芒伸长 | |
| Wknox1b Wknox1d | 4BS 4DS | 茎尖分生组织的形成和维持 |
... 小麦Hd基因与大麦HvKNOX3基因表型相似, 其突变体芒缩短、基部弯曲且膨大成膜状(
... 大麦也是研究芒性状遗传及作用的重要模式作物.野生型大麦多为具芒表型, 突变体主要表现为芒表型的变化或发育缺陷, 目前研究较多的基因为HvKNOX3、Lks2、SuK和ROUGH AWN1等(
... The genes involved in the function of the awn in Hordeum vulgare identified by genetic mapping
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| HvKNOX3 | 4H | 分生组织的维持 | |
| Lks2 | 7H | 抑制芒伸长和控制雌蕊形态 | |
| SuKD SuKB SuKC SuKE SuKF | 5H 7H 7H 7H 7H | 抑制HvKNOX3的表达 | |
| ROUGH AWN1 | 5H | 控制芒倒钩 |
... SuK位点位于5H (SuKD)和7H (SuKB、SuKC、SuKE和SuKF)染色体上.与野生型大麦相比, 具有隐性突变的SuK植株和显性突变的HvKNOX3植株芒更短且不存在“帽子”表型.一般认为SuK无表型, 仅有抑制HvKNOX3的作用(
Intragenic diversity and functional conservation of the three homoeologous loci of the KN1-type homeobox gene Wknox1 in common wheat.
2
2005
... The genes involved in the function of the awn in Triticum aestivum identified by genetic mapping
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| B1 | 5AL | 分生组织的 维持 | |
| B2 | 6BL | 抑制芒伸长 | |
| Hd (Wknox1a) | 4AL | 抑制芒伸长 | |
| Wknox1b Wknox1d | 4BS 4DS | 茎尖分生组织的形成和维持 |
... 小麦Hd基因与大麦HvKNOX3基因表型相似, 其突变体芒缩短、基部弯曲且膨大成膜状(
Awnedness affects grain yield and kernel weight in near-isogenic lines of durum wheat
1
2002
... 研究表明, 有芒作物的产量一般显著高于同类无芒作物, 增产约1/10 (
Molecular analysis of homeotic genes involved in barley development
3
2000
... 大麦也是研究芒性状遗传及作用的重要模式作物.野生型大麦多为具芒表型, 突变体主要表现为芒表型的变化或发育缺陷, 目前研究较多的基因为HvKNOX3、Lks2、SuK和ROUGH AWN1等(
... 大麦HvKNOX3基因位于4H染色体, HvKNOX3突变植株表型为反向的花取代芒, 这种表型被称为“帽子”.由于其启动子和第4个内含子序列中的(GA)8重复的存在, 进而结合BBR位点激活基因的表达, 因此内含子中BBR结合位点的(GA)8重复可能导致HvKNOX3表达过度激活.也有报道推测, (GA)8重复破坏了第4个内含子中沉默子的另一个调控序列, 进而使该沉默子失活, 导致HvKNOX3过表达(
... 基因的表达(
The barley Hooded mutation caused by a duplication in a homeobox gene intron.
3
1995
... 大麦也是研究芒性状遗传及作用的重要模式作物.野生型大麦多为具芒表型, 突变体主要表现为芒表型的变化或发育缺陷, 目前研究较多的基因为HvKNOX3、Lks2、SuK和ROUGH AWN1等(
... The genes involved in the function of the awn in Hordeum vulgare identified by genetic mapping
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| HvKNOX3 | 4H | 分生组织的维持 | |
| Lks2 | 7H | 抑制芒伸长和控制雌蕊形态 | |
| SuKD SuKB SuKC SuKE SuKF | 5H 7H 7H 7H 7H | 抑制HvKNOX3的表达 | |
| ROUGH AWN1 | 5H | 控制芒倒钩 |
... 大麦HvKNOX3基因位于4H染色体, HvKNOX3突变植株表型为反向的花取代芒, 这种表型被称为“帽子”.由于其启动子和第4个内含子序列中的(GA)8重复的存在, 进而结合BBR位点激活基因的表达, 因此内含子中BBR结合位点的(GA)8重复可能导致HvKNOX3表达过度激活.也有报道推测, (GA)8重复破坏了第4个内含子中沉默子的另一个调控序列, 进而使该沉默子失活, 导致HvKNOX3过表达(
Distribution of carbon-14 assimilated by wheat awns
1
1978
... 芒中的叶绿体与叶片中的叶绿体具有相似的结构和功能.芒作为穗部重要的光合器官, 与叶片相比, 在光合作用上具有许多优势.首先, 芒的光合面积大, 且着生于穗部顶端, 利于截获光能和气体(
Ear and flag leaf photosynthesis of awned and awnless Triticum species.
1
1976
... 芒中的叶绿体与叶片中的叶绿体具有相似的结构和功能.芒作为穗部重要的光合器官, 与叶片相比, 在光合作用上具有许多优势.首先, 芒的光合面积大, 且着生于穗部顶端, 利于截获光能和气体(
Genetic relationship of stripe rust resistance genes Yr34 and Yr48 in wheat and identification of linked KASP markers.
2
2018
... 芒是禾本科作物穗部的重要结构, 属于叶的变态, 是植物长期适应环境和进化的产物.不同材料存在芒的有无及芒长度的差异, 可作为区分禾谷类作物不同品种以及基因定位的重要形态标记(
... 芒是禾谷类作物一种重要的驯化性状, 其表面的倒刺可以附着在动物皮毛上, 借助动物的运动进行传播(
Awns reduce grain number to increase grain size and harvestable yield in irrigated and rainfed spring wheat
2
2016
... 芒是禾本科作物穗部的重要结构, 属于叶的变态, 是植物长期适应环境和进化的产物.不同材料存在芒的有无及芒长度的差异, 可作为区分禾谷类作物不同品种以及基因定位的重要形态标记(
... 研究表明, 有芒作物的产量一般显著高于同类无芒作物, 增产约1/10 (
Genetics of barley hooded suppression.
3
2004
... 大麦也是研究芒性状遗传及作用的重要模式作物.野生型大麦多为具芒表型, 突变体主要表现为芒表型的变化或发育缺陷, 目前研究较多的基因为HvKNOX3、Lks2、SuK和ROUGH AWN1等(
... The genes involved in the function of the awn in Hordeum vulgare identified by genetic mapping
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| HvKNOX3 | 4H | 分生组织的维持 | |
| Lks2 | 7H | 抑制芒伸长和控制雌蕊形态 | |
| SuKD SuKB SuKC SuKE SuKF | 5H 7H 7H 7H 7H | 抑制HvKNOX3的表达 | |
| ROUGH AWN1 | 5H | 控制芒倒钩 |
... SuK位点位于5H (SuKD)和7H (SuKB、SuKC、SuKE和SuKF)染色体上.与野生型大麦相比, 具有隐性突变的SuK植株和显性突变的HvKNOX3植株芒更短且不存在“帽子”表型.一般认为SuK无表型, 仅有抑制HvKNOX3的作用(
The GA octodinucleotide repeat binding factor BBR participates in the transcriptional regulation of the homeobox gene BKn3.
3
2003
... 大麦也是研究芒性状遗传及作用的重要模式作物.野生型大麦多为具芒表型, 突变体主要表现为芒表型的变化或发育缺陷, 目前研究较多的基因为HvKNOX3、Lks2、SuK和ROUGH AWN1等(
... The genes involved in the function of the awn in Hordeum vulgare identified by genetic mapping
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| HvKNOX3 | 4H | 分生组织的维持 | |
| Lks2 | 7H | 抑制芒伸长和控制雌蕊形态 | |
| SuKD SuKB SuKC SuKE SuKF | 5H 7H 7H 7H 7H | 抑制HvKNOX3的表达 | |
| ROUGH AWN1 | 5H | 控制芒倒钩 |
... 大麦HvKNOX3基因位于4H染色体, HvKNOX3突变植株表型为反向的花取代芒, 这种表型被称为“帽子”.由于其启动子和第4个内含子序列中的(GA)8重复的存在, 进而结合BBR位点激活基因的表达, 因此内含子中BBR结合位点的(GA)8重复可能导致HvKNOX3表达过度激活.也有报道推测, (GA)8重复破坏了第4个内含子中沉默子的另一个调控序列, 进而使该沉默子失活, 导致HvKNOX3过表达(
The SHOOTLESS2 and SHOOTLESS1 genes are involved in both initiation and maintenance of the shoot apical meristem through regulating the number of indeterminate cells.
3
2003
... 在水稻驯化过程中, 由于无芒品种便于种子的收集和储存, 因此, 人类倾向于选育无芒的水稻品种.目前, 许多参与调控芒形成和伸长的基因在水稻中已被克隆, 如An-1、An-2、LABA1、GAD1、GLA、TOB1、DL、SHL2、SHO1、SHO2、WAF1和OsETT2 (
... The genes involved in the function of the awn in Oryza sativa identified by genetic mapping
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| AN1 | 4 | 促进细胞分裂和芒原基形成、谷粒伸长及穗粒数减少 | |
| AN2 | 4 | 促进细胞分裂素的合成 | |
| LABA1 | 4 | 增强芒原基细胞分裂活性及促进芒伸长和芒刺形成 | |
| GAD1 | 8 | 促进芒原基细胞分裂和芒形成 | |
| GLA | 8 | 促进谷粒伸长 | |
| TOB1 | 4 | 促进外稃和内稃形成和生长、分生组织的维持及花器官数量减少 | |
| DL | 3 | 分生组织细胞的激活、促进芒形成和伸长及调节花器官的发育 | |
| SHO2 | - | 参与TAS3途径中ta-siRNA的合成 | |
| SHL2 | 1 | 参与TAS3途径中ta-siRNA的合成 | |
| SHO1 | 4 | 参与TAS3途径中ta-siRNA的合成 | |
| WAF1 | 7 | 参与TAS3途径中ta-siRNA的合成 | |
| OsETT2 | 1 | 外稃生长活化 |
... 水稻中芒发育相关基因SHO2、SHL2、SHO1及WAF1编码的蛋白参与TAS3 ta-siRNA调控途径.该途径调控幼叶向成熟叶过渡及侧根生长发育等过程, OsETT2基因受该途径调控(
Seed dispersal by adhesion
2
1986
... 芒是禾本科植物小花外稃或颖片顶端的一种针状延伸, 从基部到顶端逐渐变细.其表皮含有不同形态的细胞, 如较狭长的细胞、卵状或方形的短细胞和厚壁而尖细的单细胞.其中较狭长的细胞具有波状加厚的壁; 卵状或方形的短细胞表现为芒表皮上的乳头状突起; 厚壁而尖细的单细胞向前斜生, 使得芒表面较为粗糙(
... 芒是禾谷类作物一种重要的驯化性状, 其表面的倒刺可以附着在动物皮毛上, 借助动物的运动进行传播(
Microsatellite-based deletion bin system for the establishment of genetic-physical map relationships in wheat (Triticum aestivum L.).
1
2004
... 在普通小麦中, B1 (5AL)、B2 (6BL)和Hd (4AL)是目前已知的芒显性抑制基因, 它们的不同组合导致芒表型发生变化(
Molecular cloning of three homoeologous cDNAs encoding orthologs of the maize KNOTTED1 homeobox protein from young spikes of hexaploid wheat
2
2000
... 小麦Hd基因与大麦HvKNOX3基因表型相似, 其突变体芒缩短、基部弯曲且膨大成膜状(
... 水稻An-1基因位于4号染色体上, 编码1个bHLH转录因子, 可促进细胞分裂和芒原基形成.An-1基因上调表达导致芒和穗粒伸长以及穗粒数减少, 在水稻人工选择中, An-1是关键靶基因(
The YABBY gene TONGARI-BOUSHI1 is involved in lateral organ development and maintenance of meristem organization in the rice spikelet.
4
2012
... 在水稻驯化过程中, 由于无芒品种便于种子的收集和储存, 因此, 人类倾向于选育无芒的水稻品种.目前, 许多参与调控芒形成和伸长的基因在水稻中已被克隆, 如An-1、An-2、LABA1、GAD1、GLA、TOB1、DL、SHL2、SHO1、SHO2、WAF1和OsETT2 (
... The genes involved in the function of the awn in Oryza sativa identified by genetic mapping
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| AN1 | 4 | 促进细胞分裂和芒原基形成、谷粒伸长及穗粒数减少 | |
| AN2 | 4 | 促进细胞分裂素的合成 | |
| LABA1 | 4 | 增强芒原基细胞分裂活性及促进芒伸长和芒刺形成 | |
| GAD1 | 8 | 促进芒原基细胞分裂和芒形成 | |
| GLA | 8 | 促进谷粒伸长 | |
| TOB1 | 4 | 促进外稃和内稃形成和生长、分生组织的维持及花器官数量减少 | |
| DL | 3 | 分生组织细胞的激活、促进芒形成和伸长及调节花器官的发育 | |
| SHO2 | - | 参与TAS3途径中ta-siRNA的合成 | |
| SHL2 | 1 | 参与TAS3途径中ta-siRNA的合成 | |
| SHO1 | 4 | 参与TAS3途径中ta-siRNA的合成 | |
| WAF1 | 7 | 参与TAS3途径中ta-siRNA的合成 | |
| OsETT2 | 1 | 外稃生长活化 |
... YABBY基因家族的TOB1和DL也参与调控水稻芒的形成.YABBY基因编码的转录因子具有螺旋-环-螺旋和锌指两种保守性.这些基因在植物花器官的发育和分生组织的维持中起重要调控作用.其中YABBY蛋白对分生组织起间接作用, YABBY蛋白产生的次级信号进入分生组织细胞并激活分裂信号 (
... TOB1基因位于水稻4号染色体, 编码OsYABBY5转录因子, 该基因突变有多种表型, 其中I类突变型直接表现为促进外稃上芒的形成(
Comparative data on the rate of photosynthesis, respiration, and transpiration of different organs in awned and awnless isogenic lines of wheat
3
1972
... 芒中的叶绿体与叶片中的叶绿体具有相似的结构和功能.芒作为穗部重要的光合器官, 与叶片相比, 在光合作用上具有许多优势.首先, 芒的光合面积大, 且着生于穗部顶端, 利于截获光能和气体(
... ).此外, 小麦发育后期旗叶衰老且光合速率开始下降, 芒不易老化的特性使其仍具有较高的光合速率(
... 芒对产量的贡献虽然没有旗叶显著, 但两者结合比单独作用对产量的贡献更大(
The DROOPING LEAF and OsETTIN2 genes promote awn development in rice.
4
2014
... 在水稻驯化过程中, 由于无芒品种便于种子的收集和储存, 因此, 人类倾向于选育无芒的水稻品种.目前, 许多参与调控芒形成和伸长的基因在水稻中已被克隆, 如An-1、An-2、LABA1、GAD1、GLA、TOB1、DL、SHL2、SHO1、SHO2、WAF1和OsETT2 (
... The genes involved in the function of the awn in Oryza sativa identified by genetic mapping
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| AN1 | 4 | 促进细胞分裂和芒原基形成、谷粒伸长及穗粒数减少 | |
| AN2 | 4 | 促进细胞分裂素的合成 | |
| LABA1 | 4 | 增强芒原基细胞分裂活性及促进芒伸长和芒刺形成 | |
| GAD1 | 8 | 促进芒原基细胞分裂和芒形成 | |
| GLA | 8 | 促进谷粒伸长 | |
| TOB1 | 4 | 促进外稃和内稃形成和生长、分生组织的维持及花器官数量减少 | |
| DL | 3 | 分生组织细胞的激活、促进芒形成和伸长及调节花器官的发育 | |
| SHO2 | - | 参与TAS3途径中ta-siRNA的合成 | |
| SHL2 | 1 | 参与TAS3途径中ta-siRNA的合成 | |
| SHO1 | 4 | 参与TAS3途径中ta-siRNA的合成 | |
| WAF1 | 7 | 参与TAS3途径中ta-siRNA的合成 | |
| OsETT2 | 1 | 外稃生长活化 |
... 水稻DL基因定位于3号染色体, 在叶原基中心区域的细胞和芒原基中表达, 通过诱导芒原基细胞分裂促进芒的伸长(
... 水稻中芒发育相关基因SHO2、SHL2、SHO1及WAF1编码的蛋白参与TAS3 ta-siRNA调控途径.该途径调控幼叶向成熟叶过渡及侧根生长发育等过程, OsETT2基因受该途径调控(
Natural variations in the promoter of Awn Length Inhibitor 1(ALI-1) are associated with awn elongation and grain length in common wheat.
7
2020
... 研究表明, 小麦B1基因对芒长度的影响最强, 其突变体的表型为穗基部和中部产生短芒或无芒, 芒最长仅为1 cm (
... 中国科学院遗传与发育生物学研究所张爱民研究团队初步将B1定位于5A染色体长臂125 kb的物理区间内, 其中包含2个基因(TraesCS5A02G542800和TraesCS5A02G542900).表达谱和序列分析表明, 短芒材料中TraesCS5A02G542800的表达量远高于长芒材料, 而TraesCS5A02G542900在短芒和长芒材料间表达量无显著差异.测序结果表明, TraesCS5A02G542900序列在短芒和长芒材料中一致, TraesCS5A02G542800编码区序列一致, 但在启动子区有5个相同差异的SNP, B1/B1基因型材料表现为TAGA单体型, b1/b1基因型材料表现为CGAG单体型.转基因功能验证进一步表明, TraesCS5A02G542800启动子为B1芒长抑制位点的控制基因, 将其命名为ALI-1 (
... ALI-1编码1个C2H2锌指转录因子, 具有转录抑制活性.对B1近等基因系的芒进行纵切观察, 结果显示两者细胞长度无显著差异, 但短芒材料细胞数目仅为长芒材料的1/10, 表明短芒材料芒原基的细胞分裂被抑制.顺式调控元件预测结果表明, ALI-1区间内SNPs导致该基因缺失顺式元件BOXCPSAS1、LTRE1HVBLT49、SORLIP2AT和SITEIIATCYTC, 这些顺式元件参与调控分生组织和增殖细胞的基因表达(
... ;
... 抑制细胞分裂素的信号转导, 导致细胞增殖停滞, 细胞数量减少, 进而使芒的伸长受到抑制, 表现为极显著的芒抑制作用(
... 截至目前, 虽然B1基因已被克隆, 但其表达和抑芒机制仍然未知.目前, 对其突变位点及功能研究仍然存在较大争议, 还需要进一步的实验验证(
... 人们对于芒作用的认识现今主要集中在芒可以通过促进光合作用影响千粒重和产量.此外, 芒还可以直接调控种子大小, 扩大库容量(
Variation and genetics of the awn in Triticum.
2
1940
... 芒是禾本科作物穗部的重要结构, 属于叶的变态, 是植物长期适应环境和进化的产物.不同材料存在芒的有无及芒长度的差异, 可作为区分禾谷类作物不同品种以及基因定位的重要形态标记(
... 芒是禾谷类作物一种重要的驯化性状, 其表面的倒刺可以附着在动物皮毛上, 借助动物的运动进行传播(
Overrepresentation of elements recognized by TCP-domain transcription factors in the upstream regions of nuclear genes encoding components of the mitochondrial oxidative phosphorylation machinery
1
2006
... ALI-1编码1个C2H2锌指转录因子, 具有转录抑制活性.对B1近等基因系的芒进行纵切观察, 结果显示两者细胞长度无显著差异, 但短芒材料细胞数目仅为长芒材料的1/10, 表明短芒材料芒原基的细胞分裂被抑制.顺式调控元件预测结果表明, ALI-1区间内SNPs导致该基因缺失顺式元件BOXCPSAS1、LTRE1HVBLT49、SORLIP2AT和SITEIIATCYTC, 这些顺式元件参与调控分生组织和增殖细胞的基因表达(
The YABBY gene DROOPING LEAF regulates carpel specification and midrib development in Oryza sativa.
4
2004
... 在水稻驯化过程中, 由于无芒品种便于种子的收集和储存, 因此, 人类倾向于选育无芒的水稻品种.目前, 许多参与调控芒形成和伸长的基因在水稻中已被克隆, 如An-1、An-2、LABA1、GAD1、GLA、TOB1、DL、SHL2、SHO1、SHO2、WAF1和OsETT2 (
... The genes involved in the function of the awn in Oryza sativa identified by genetic mapping
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| AN1 | 4 | 促进细胞分裂和芒原基形成、谷粒伸长及穗粒数减少 | |
| AN2 | 4 | 促进细胞分裂素的合成 | |
| LABA1 | 4 | 增强芒原基细胞分裂活性及促进芒伸长和芒刺形成 | |
| GAD1 | 8 | 促进芒原基细胞分裂和芒形成 | |
| GLA | 8 | 促进谷粒伸长 | |
| TOB1 | 4 | 促进外稃和内稃形成和生长、分生组织的维持及花器官数量减少 | |
| DL | 3 | 分生组织细胞的激活、促进芒形成和伸长及调节花器官的发育 | |
| SHO2 | - | 参与TAS3途径中ta-siRNA的合成 | |
| SHL2 | 1 | 参与TAS3途径中ta-siRNA的合成 | |
| SHO1 | 4 | 参与TAS3途径中ta-siRNA的合成 | |
| WAF1 | 7 | 参与TAS3途径中ta-siRNA的合成 | |
| OsETT2 | 1 | 外稃生长活化 |
... YABBY基因家族的TOB1和DL也参与调控水稻芒的形成.YABBY基因编码的转录因子具有螺旋-环-螺旋和锌指两种保守性.这些基因在植物花器官的发育和分生组织的维持中起重要调控作用.其中YABBY蛋白对分生组织起间接作用, YABBY蛋白产生的次级信号进入分生组织细胞并激活分裂信号 (
... 水稻DL基因定位于3号染色体, 在叶原基中心区域的细胞和芒原基中表达, 通过诱导芒原基细胞分裂促进芒的伸长(
Three dominant awnless genes in common wheat: fine mapping, interaction and contribution to diversity in awn shape and length
2
2017
... 研究表明, 小麦B1基因对芒长度的影响最强, 其突变体的表型为穗基部和中部产生短芒或无芒, 芒最长仅为1 cm (
... 小麦B2基因表型为整个穗部的短芒.一般情况下, 穗部各部分的芒长一致, 少数情况下会出现穗中部芒比较长(
A SHORT INTERNODES (SHI) family transcription factor gene regulates awn elongation and pistil morphology in barley.
3
2012
... 大麦也是研究芒性状遗传及作用的重要模式作物.野生型大麦多为具芒表型, 突变体主要表现为芒表型的变化或发育缺陷, 目前研究较多的基因为HvKNOX3、Lks2、SuK和ROUGH AWN1等(
... The genes involved in the function of the awn in Hordeum vulgare identified by genetic mapping
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| HvKNOX3 | 4H | 分生组织的维持 | |
| Lks2 | 7H | 抑制芒伸长和控制雌蕊形态 | |
| SuKD SuKB SuKC SuKE SuKF | 5H 7H 7H 7H 7H | 抑制HvKNOX3的表达 | |
| ROUGH AWN1 | 5H | 控制芒倒钩 |
... 大麦Lks2基因定位于7H染色体, 编码SHI基因家族的转录因子, 控制芒的长度和雌蕊形态, 显性纯合状态下, 芒的长度会缩短1/2.该基因编码的蛋白包含锌指结构和IGGH结构域(
Natural alleles of GLA for grain length and awn development were differently domesticated in rice subspecies japonica and indica.
4
2019
... 在水稻驯化过程中, 由于无芒品种便于种子的收集和储存, 因此, 人类倾向于选育无芒的水稻品种.目前, 许多参与调控芒形成和伸长的基因在水稻中已被克隆, 如An-1、An-2、LABA1、GAD1、GLA、TOB1、DL、SHL2、SHO1、SHO2、WAF1和OsETT2 (
... The genes involved in the function of the awn in Oryza sativa identified by genetic mapping
| 基因 | 染色体 | 功能 | 参考文献 |
|---|---|---|---|
| AN1 | 4 | 促进细胞分裂和芒原基形成、谷粒伸长及穗粒数减少 | |
| AN2 | 4 | 促进细胞分裂素的合成 | |
| LABA1 | 4 | 增强芒原基细胞分裂活性及促进芒伸长和芒刺形成 | |
| GAD1 | 8 | 促进芒原基细胞分裂和芒形成 | |
| GLA | 8 | 促进谷粒伸长 | |
| TOB1 | 4 | 促进外稃和内稃形成和生长、分生组织的维持及花器官数量减少 | |
| DL | 3 | 分生组织细胞的激活、促进芒形成和伸长及调节花器官的发育 | |
| SHO2 | - | 参与TAS3途径中ta-siRNA的合成 | |
| SHL2 | 1 | 参与TAS3途径中ta-siRNA的合成 | |
| SHO1 | 4 | 参与TAS3途径中ta-siRNA的合成 | |
| WAF1 | 7 | 参与TAS3途径中ta-siRNA的合成 | |
| OsETT2 | 1 | 外稃生长活化 |
... 水稻An-1基因位于4号染色体上, 编码1个bHLH转录因子, 可促进细胞分裂和芒原基形成.An-1基因上调表达导致芒和穗粒伸长以及穗粒数减少, 在水稻人工选择中, An-1是关键靶基因(
... 水稻LABA1基因编码1个细胞分裂素激活酶(cytokinin riboside 5°-monophosphate phosphoribohydrolase), 该酶通过提高芒原基的活性细胞分裂素含量增强芒原基的细胞分裂素活性, 促进芒的伸长和芒刺形成(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}