Identification of a New OsBRI1 Weak Allele and Analysis of its Function in Grain Size Control
Liurong Guan1,2, Zupei Liu2,3, Ran Xu2, Penggen Duan2, Guozheng Zhang2,3, Haiyue Yu2,3, Jing Li2,3, Yuehua Luo,1,*, Yunhai Li,2,*1Hainan Key Laboratory for Sustainable Utilization of Tropical Bioresources, College of Tropical Crops, Hainan University, Haikou 570228, China 2State Key Laboratory of Plant Cell and Chromosome Engineering, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing 100101, China 3University of Chinese Academy of Sciences, Beijing 100039, China
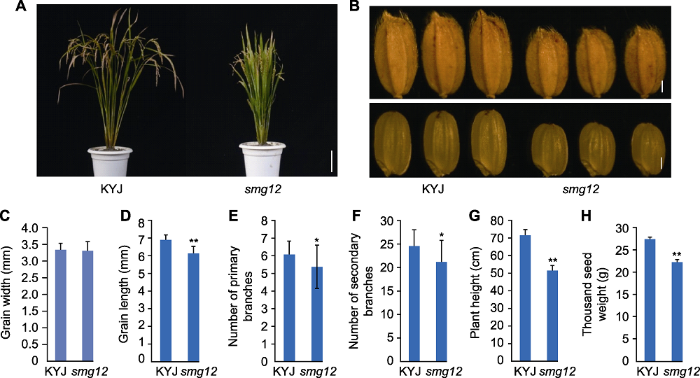
Abstract Rice (Oryza sativa) grain size and grain weight are key agronomic traits that affect rice yield. Cloning and study of grain size genes are helpful to increase rice production. In order to further understand the mechanism of rice grain size control, a set of mutants with altered grain size from an EMS-treated elite japonica cultivar KYJ (Kuanyejing) were isolated. smg12 exhibits small grains, short plants, and reduced number of primary branches and secondary branches. Genetic analyses show that the smg12 mutant phenotypes are controlled by a single recessive gene. Our celluar analyses show that the small grain size phenotype of smg12 is caused by the decrease in cell size of glumes, indicating that SMG12 affects cell expansion. By using the Mutmap method, we reveal that the candidate gene for SMG12 is OsBRI1, which encodes a brassinolide receptor kinase. The smg12 mutant causes a substitution of the 2 074th base (C to T) in OsBRI1, which results in an amino acid change (proline to serine). Therefore, this study identified a new mutant allele of OsBRI1 and provides a cellular and molecular basis for BR-mediated grain size control in rice. Keywords:rice;grain size;cell expansion;Mutmap;OsBRI1
PDF (1346KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 管柳蓉, 刘祖培, 徐冉, 段朋根, 张国政, 于海跃, 李静, 罗越华, 李云海. 一个新的OsBRI1弱等位突变体的鉴定及其调控种子大小的功能研究. 植物学报, 2020, 55(3): 279-286 doi:10.11983/CBB19239 Guan Liurong, Liu Zupei, Xu Ran, Duan Penggen, Zhang Guozheng, Yu Haiyue, Li Jing, Luo Yuehua, Li Yunhai. Identification of a New OsBRI1 Weak Allele and Analysis of its Function in Grain Size Control. Chinese Bulletin of Botany, 2020, 55(3): 279-286 doi:10.11983/CBB19239
水稻(Oryza sativa)作为最重要的谷物粮食之一, 是全世界超过一半人口的主要食物来源。然而, 现阶段人口和粮食需求不断增加, 使农业面临巨大挑战 , 且随着可利用土地面积和自然资源逐渐减少, 需要提高作物的生产效率, 其中增加单位面积产量尤为重要(Li et al., 2018)。高等植物的种子由胚、胚乳和种皮三部分组成, 它们分别由受精卵、受精卵中心细胞和母体细胞发育而来, 共同协调控制种子大小。水稻胚乳占据成熟种子大部分空间, 主要成分为淀粉, 是人类的主要食物来源, 也是决定粮食重量的直接因素(James et al., 2003; Zhou et al., 2013)。影响水稻产量的主要因素是单位面积有效穗数、每穗粒数、千粒重和灌浆速率(Sakamoto and Matsuoka, 2008; Xing and Zhang, 2010)。粒重受水稻籽粒宽度、长度和粒厚协调控制, 说明水稻籽粒大小与产量息息相关。在单子叶植物水稻中, 籽粒长在颖壳内且籽粒大小受颖壳限制(Song et al., 2007; Shomura et al., 2008)。籽粒大小受颖壳细胞数目和细胞大小调控, 细胞数目主要通过细胞增殖改变, 细胞大小受细胞扩展影响。近年来, 与水稻籽粒大小相关的基因逐渐被克隆, 其中一些基因对于提高水稻产量具有重要意义(宫李辉等, 2011)。
目前的研究已经揭示了多条调控水稻籽粒大小的信号途径, 包括泛素蛋白酶体途径、G蛋白信号途径、丝裂原活化蛋白激酶(mitogen-activated protein kinase, MAPK)途径、转录调控因子及植物激素等(Zuo and Li, 2014; Li et al., 2019)。控制粒宽和粒重的基因GW2编码1个E3泛素连接酶, 负向调控细胞分裂。GW2功能的丧失导致颖壳细胞数目增加, 产生更大更宽的颖壳并加快灌浆速率, 从而使籽粒更宽更重, 产量更高(Song et al., 2007)。GS3编码异源三聚体G蛋白的Gγ亚基, 是植物籽粒大小和器官大小的负调控因子。野生型等位基因对应中等大小籽粒, N端OSR (organ size regulation)结构域功能缺失突变体籽粒变长, 而C端的TNFR (tumor necrosis factor receptor)/NGFR (nerve growth factor receptor)和VWFC (von Willebrand factor type C)结构域对OSR结构域的功能有抑制作用, C端结构域功能缺失则会产生非常短的籽粒(Fan et al., 2006; Mao et al., 2010)。丝裂原活化蛋白激酶信号通路在真核生物中的作用非常保守, 且与植物生长发育以及防御反应的多个过程相关(Meng and Zhang, 2013)。MAPK途径组成包括MAPK激酶激酶(MKKK)、MAPK激酶(MKK)和MAPK, 这3种激酶依次激活控制下游基因的表达, 调控植物生长发育。水稻OsMKKK10、OsMKK4和OsMAPK6作为1个级联信号通路促进细胞增殖, 从而正向调控水稻籽粒大小和粒重(Xu et al., 2018b)。其功能缺失突变体osmkkk10、osmkk4和osmapk6由于颖壳细胞增殖过程受到限制, 导致籽粒变小(Liu et al., 2015b; Xu et al., 2018a, 2018b)。OsSPL16编码1种细胞增殖的正调控蛋白, 过表达该基因会促进细胞分裂和灌浆, 使水稻籽粒变宽, 产量增加(Wang et al., 2012)。水稻中重要的数量位点GLW7编码植物特异性转录因子OsSPL13, 通过正向调控颖壳细胞大小提高籽粒长度和产量(Si et al., 2016)。植物激素对种子发育有重要作用并直接影响籽粒大小。BG1是正向调控籽粒大小的膜蛋白, 有研究显示BG1能够响应生长素变化, 且与生长素转运过程相关, 表明其可能通过生长素信号来调控籽粒生长。BG1的激活促进细胞扩展, 从而增加籽粒大小和粒重, 而BG1的表达被抑制时会形成小的籽粒(Liu et al., 2015a)。近年来, 油菜素内酯对种子发育的调控机制已经在水稻、拟南芥(Arabidopsis thaliana)及其它植物中进行了深入研究。油菜素内酯对株高、叶夹角、分蘖数、籽粒大小和产量都有不同程度的影响(Morinaka et al., 2006)。水稻中油菜素内酯受体基因D61的突变和油菜素内酯信号的负调控基因GSK2的过表达植株都会产生更小的籽粒(Tanabe et al., 2005; Tong et al., 2012)。BU1编码1个具有螺旋-环-螺旋结构的转录因子, 作为响应油菜素内酯信号的正调控因子作用于D61基因的下游, BU1基因过表达导致水稻籽粒变大(Tanaka et al., 2009)。上述结果表明, 籽粒大小调控是非常复杂的过程, 涉及多种信号转导以及生化代谢途径。
(A) Outer epidermal cell number in the grain-length direction of the glume; (B) Outer epidermal cell number in the grain-width direction of the glume; (C) The longitudinal length of a single cell in the glume; (D) Transverse width of a single cell in the glume; (E) Cytological analysis diagram (Bars= 0.1 mm). ** indicates extremely significant difference between the mutant and the wild type (P<0.01).
(A) The dCAPS marker was developed to detect the smg12 mutation; (B) Population linkage analysis of the candidate gene LOC_Os01g52050; (C) The LOC_Os01g52050 gene structure, open boxes show the 5' and 3' untranslated regions, the closed box shows the coding sequence, and the start codon (ATG), the stop codon (TAG) and the LOC_Os01g52050 mutation site (C/T) are indicated; (D) Schematic of the LOC_Os01g52050 protein. P/S indicated the LOC_Os01g52050 mutation site.
AbeA, KosugiS, YoshidaK, NatsumeS, TakagiH, KanzakiH, MatsumuraH, YoshidaK, MitsuokaC, TamiruM, InnanH, CannoL, KamounS, TerauchiR (2012). Genome sequencing reveals agronomically important loci in rice using Mutmap Nat Biotechnol 30, 174-178. URLPMID:22267009 [本文引用: 3]
FanCC, XingYZ, MaoHL, LuTT, HanB, XuCG, LiXH, ZhangQF (2006). GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein Theor Appl Genet 112, 1164-1171. URLPMID:16453132 [本文引用: 1]
JamesMG, DenyerK, MyersAM (2003). Starch synthesis in the cereal endosperm Curr Opin Plant Biol 6, 215-222. [本文引用: 1]
LiN, XuR, DuanPG, LiYH (2018). Control of grain size in rice Plant Reprod 31, 237-251. [本文引用: 1]
LiN, XuR, LiYH (2019). Molecular networks of seed size control in plants Annu Rev Plant Biol 70, 435-463. URLPMID:30795704 [本文引用: 1]
LiuLC, TongHN, XiaoYH, CheRH, XuF, HuB, LiangCZ, ChuJF, LiJY, ChuCC (2015a). Activation of Big Grain1 significantly improves grain size by regulating auxin transport in rice Proc Natl Acad Sci USA 112, 11102-11107. URLPMID:26283354 [本文引用: 1]
LiuSY, HuaL, DongSJ, ChenHQ, ZhuXD, JiangJE, ZhangF, LiYH, FangXH, ChenF (2015b). OsMAPK6, a mitogen-activated protein kinase, influences rice grain size and biomass production Plant J 84, 672-681. DOI:10.1111/tpj.13025URLPMID:26366992 [本文引用: 1] Grain size is an important agronomic trait in determining grain yield. However, the molecular mechanisms that determine the final grain size are not well understood. Here, we report the functional analysis of a rice (Oryza sativa L.) mutant, dwarf and small grain1 (dsg1), which displays pleiotropic phenotypes, including small grains, dwarfism and erect leaves. Cytological observations revealed that the small grain and dwarfism of dsg1 were mainly caused by the inhibition of cell proliferation. Map-based cloning revealed that DSG1 encoded a mitogen-activated protein kinase (MAPK), OsMAPK6. OsMAPK6 was mainly located in the nucleus and cytoplasm, and was ubiquitously distributed in various organs, predominately in spikelets and spikelet hulls, consistent with its role in grain size and biomass production. As a functional kinase, OsMAPK6 interacts strongly with OsMKK4, indicating that OsMKK4 is likely to be the upstream MAPK kinase of OsMAPK6 in rice. In addition, hormone sensitivity tests indicated that the dsg1 mutant was less sensitive to brassinosteroids (BRs). The endogenous BR levels were reduced in dsg1, and the expression of several BR signaling pathway genes and feedback-inhibited genes was altered in the dsg1 mutant, with or without exogenous BRs, indicating that OsMAPK6 may contribute to influence BR homeostasis and signaling. Thus, OsMAPK6, a MAPK, plays a pivotal role in grain size in rice, via cell proliferation, and BR signaling and homeostasis.
MaoHL, SunSY, YaoJL, WangCR, YuSB, XuCG, LiXH, ZhangQF (2010). Linking differential domain functions of the GS3 protein to natural variation of grain size in rice Proc Natl Acad Sci USA 107, 19579-19584. [本文引用: 1]
MengXZ, ZhangSQ (2013). MAPK cascades in plant disease resistance signaling Annu Rev Phytopathol 51, 245-266. URLPMID:23663002 [本文引用: 1]
MiuraK, AshikariM, MatsuokaM (2011). The role of QTLs in the breeding of high-yielding rice Trends Plant Sci 16, 319-326. URLPMID:21429786 [本文引用: 1]
MorinakaY, SakamotoT, InukaiY, AgetsumaM, KitanoH, AshikariM, MatsuokaM (2006). Morphological alteration caused by brassinosteroid insensitivity increases the biomass and grain production of rice Plant Physiol 141, 924-931. [本文引用: 1]
NakamuraA, FujiokaS, SunoharaH, KamiyaN, HongZ, InukaiY, MiuraK, TakatsutoS, YoshidaS, Ueguchi- TanakaM, HasegawaY, KitanoH, MatsuokaM (2006). The role of OsBRI1 and its homologous genes, OsBRL1 and OsBRL3, in rice Plant Physiol 140, 580-590. DOI:10.1104/pp.105.072330URLPMID:16407447 [本文引用: 2] Since first identifying two alleles of a rice (Oryza sativa) brassinosteroid (BR)-insensitive mutant, d61, that were also defective in an orthologous gene in Arabidopsis (Arabidopsis thaliana) BRASSINOSTEROID INSENSITIVE1 (BRI1), we have isolated eight additional alleles, including null mutations, of the rice BRI1 gene OsBRI1. The most severe mutant, d61-4, exhibited severe dwarfism and twisted leaves, although pattern formation and differentiation were normal. This severe shoot phenotype was caused mainly by a defect in cell elongation and the disturbance of cell division after the determination of cell fate. In contrast to its severe shoot phenotype, the d61-4 mutant had a mild root phenotype. Concomitantly, the accumulation of castasterone, the active BR in rice, was up to 30-fold greater in the shoots, while only 1.5-fold greater in the roots. The homologous genes for OsBRI1, OsBRL1 and OsBRL3, were highly expressed in roots but weakly expressed in shoots, and their expression was higher in d61-4 than in the wild type. Based on these observations, we conclude that OsBRI1 is not essential for pattern formation or organ initiation, but is involved in organ development through controlling cell division and elongation. In addition, OsBRL1 and OsBRL3 are at least partly involved in BR perception in the roots.
SakamotoT, MatsuokaM (2008). Identifying and exploiting grain yield genes in rice Curr Opin Plant Biol 11, 209-214. DOI:10.1016/j.pbi.2008.01.009URLPMID:18343712 [本文引用: 1] Improved grain yield has been a major focus of crop breeding programs around the world. With the accomplishments of the Rice Genome Project, genes regulating several agronomically important traits related to grain yield, such as tiller number, grain number, grain size, and plant height, have recently been identified. Although these findings have not been enough to fully characterize the mechanisms that regulate each trait, these genes and knowledge of the molecular mechanisms involved provide a set of tools that can be combined to achieve tailor-made breeding suitable for various programs aimed at higher grain yield.
ShomuraA, IzawaT, EbanaK, EbitaniT, KanegaeH, KonishiS, YanoM (2008). Deletion in a gene associated with grain size increased yields during rice domestication Nat Genet 40, 1023-1028. URLPMID:18604208 [本文引用: 1]
SongXJ, HuangW, ShiM, ZhuMZ, LinHX (2007). A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase Nat Genet 39, 623-630. DOI:10.1038/ng2014URLPMID:17417637 [本文引用: 2] Grain weight is one of the most important components of grain yield and is controlled by quantitative trait loci (QTLs) derived from natural variations in crops. However, the molecular roles of QTLs in the regulation of grain weight have not been fully elucidated. Here, we report the cloning and characterization of GW2, a new QTL that controls rice grain width and weight. Our data show that GW2 encodes a previously unknown RING-type protein with E3 ubiquitin ligase activity, which is known to function in the degradation by the ubiquitin-proteasome pathway. Loss of GW2 function increased cell numbers, resulting in a larger (wider) spikelet hull, and it accelerated the grain milk filling rate, resulting in enhanced grain width, weight and yield. Our results suggest that GW2 negatively regulates cell division by targeting its substrate(s) to proteasomes for regulated proteolysis. The functional characterization of GW2 provides insight into the mechanism of seed development and is a potential tool for improving grain yield in crops.
TanabeS, AshikariM, FujiokaS, TakatsutoS, YoshidaS, YanoM, YoshimuraA, KotanoH, MatsuokaM, FujisawaY, KatoH, LwasakiY (2005). A novel cytochrome P450 is implicated in brassinosteroid biosynthesis via the characterization of a rice dwarf mutant, dwarf11, with reduced seed length Plant Cell 17, 776-790. DOI:10.1105/tpc.104.024950URLPMID:15705958 [本文引用: 2] We have characterized a rice (Oryza sativa) dwarf mutant, dwarf11 (d11), that bears seeds of reduced length. To understand the mechanism by which seed length is regulated, the D11 gene was isolated by a map-based cloning method. The gene was found to encode a novel cytochrome P450 (CYP724B1), which showed homology to enzymes involved in brassinosteroid (BR) biosynthesis. The dwarf phenotype of d11 mutants was restored by the application of the brassinolide (BL). Compared with wild-type plants, the aberrant D11 mRNA accumulated at higher levels in d11 mutants and was dramatically reduced by treatment with BL, implying that the gene is feedback-regulated by BL. Precise determination of the defective step(s) in BR synthesis in d11 mutants proved intractable because of tissue specificity and the complex control of BR accumulation in plants. However, 6-deoxotyphasterol (6-DeoxoTY) and typhasterol (TY), but not any upstream intermediates before these compounds, effectively restored BR response in d11 mutants in a lamina joint bending assay. Multiple lines of evidence together suggest that the D11/CYP724B1 gene plays a role in BR synthesis and may be involved in the supply of 6-DeoxoTY and TY in the BR biosynthesis network in rice.
TanakaA, NakagawaH, TomitaC, ShimataniZ, OhtakeM, NomuraT, JiangCJ, DubozetJG, KikuchiS, SekimotoH, YokotaT, AsamiT, KamakuraT, MoriM (2009). BRASSINOSTEROID UPREGULATED1, encoding a helix-loop-helix protein, is a novel gene involved in brassinosteroid signaling and controls bending of the lamina joint in rice Plant Physiol 151, 669-680. DOI:10.1104/pp.109.140806URLPMID:19648232 [本文引用: 1] Brassinosteroids (BRs) are involved in many developmental processes and regulate many subsets of downstream genes throughout the plant kingdom. However, little is known about the BR signal transduction and response network in monocots. To identify novel BR-related genes in rice (Oryza sativa), we monitored the transcriptomic response of the brassinosteroid deficient1 (brd1) mutant, with a defective BR biosynthetic gene, to brassinolide treatment. Here, we describe a novel BR-induced rice gene BRASSINOSTEROID UPREGULATED1 (BU1), encoding a helix-loop-helix protein. Rice plants overexpressing BU1 (BU1:OX) showed enhanced bending of the lamina joint, increased grain size, and resistance to brassinazole, an inhibitor of BR biosynthesis. In contrast to BU1:OX, RNAi plants designed to repress both BU1 and its homologs displayed erect leaves. In addition, compared to the wild type, the induction of BU1 by exogenous brassinolide did not require de novo protein synthesis and it was weaker in a BR receptor mutant OsbriI (Oryza sativa brassinosteroid insensitive1, d61) and a rice G protein alpha subunit (RGA1) mutant d1. These results indicate that BU1 protein is a positive regulator of BR response: it controls bending of the lamina joint in rice and it is a novel primary response gene that participates in two BR signaling pathways through OsBRI1 and RGA1. Furthermore, expression analyses showed that BU1 is expressed in several organs including lamina joint, phloem, and epithelial cells in embryos. These results indicate that BU1 may participate in some other unknown processes modulated by BR in rice.
TongHN, LiuLC, JinY, DuL, YinYH, QianQ, ZhuLH, ChuCC (2012). DWARF AND LOW-TILLERING acts as a direct downstream target of a GSK3/SHAGGY-like kinase to mediate brassinosteroid responses in rice Plant Cell 24, 2562-2577. URLPMID:22685166 [本文引用: 1]
WangSK, WuK, YuanQB, LiuXY, LiuZB, LinXY, ZengRZ, ZhuHT, DongGJ, QianQ, ZhangGQ, FuXD (2012). Control of grain size, shape and quality by OsSPL16 in rice Nat Genet 44, 950-954. DOI:10.1038/ng.2327URLPMID:22729225 [本文引用: 1] Grain size and shape are important components of grain yield and quality and have been under selection since cereals were first domesticated. Here, we show that a quantitative trait locus GW8 is synonymous with OsSPL16, which encodes a protein that is a positive regulator of cell proliferation. Higher expression of this gene promotes cell division and grain filling, with positive consequences for grain width and yield in rice. Conversely, a loss-of-function mutation in Basmati rice is associated with the formation of a more slender grain and better quality of appearance. The correlation between grain size and allelic variation at the GW8 locus suggests that mutations within the promoter region were likely selected in rice breeding programs. We also show that a marker-assisted strategy targeted at elite alleles of GS3 and OsSPL16 underlying grain size and shape can be effectively used to simultaneously improve grain quality and yield.
XingYZ, ZhangQF (2010). Genetic and molecular bases of rice yield Annu Rev Plant Biol 61, 421-442. [本文引用: 1]
XuR, DuanPG, YuHY, ZhouZK, ZhangBL, WangRC, LiJ, ZhangGZ, ZhuangSS, LyuJ, LiN, ChaiTY, TianZX, YaoSG, LiYH (2018a). Control of grain size and weight by the OsMKKK10-OsMKK4-OsMAPK6 signaling pathway in rice Mol Plant 11, 860-873. [本文引用: 1]
XuR, YuHY, WangJM, DuanPG, ZhangBL, LiJ, LiY, XuJS, LyuJ, LiN, ChaiTY, LiYH (2018b). A mitogen-activated protein kinase phosphatase influences grain size and weight in rice Plant J 95, 937-946. DOI:10.1111/tpj.13971URLPMID:29775492 [本文引用: 2] Grain size and weight are directly associated with grain yield in crops. However, the molecular mechanisms that set final grain size and weight remain largely unknown. Here, we characterize two large grain mutants, large grain8-1 (large8-1) and large grain8-2 (large8-2). LARGE8 encodes the mitogen-activated protein kinase phosphatase1 (OsMKP1). Loss of function mutations in OsMKP1 results in large grains, while overexpression of OsMKP1 leads to small grains. OsMKP1 determines grain size by restricting cell proliferation in grain hulls. OsMKP1 directly interacts with and deactivates the mitogen-activated protein kinase 6 (OsMAPK6). Taken together, we identify OsMKP1 as a crucial factor that influences grain size by deactivating OsMAPK6, indicating that the reversible phosphorylation of OsMAPK6 plays important roles in determining grain size in rice.
ZhangC, BaiMY, ChongK (2014). Brassinosteroid-mediated regulation of agronomic traits in rice Plant Cell Rep 33, 683-696. [本文引用: 1]
ZhaoJF, WuCX, YuanSJ, YinL, SunW, ZhaoQL, ZhaoBH, LiXY (2013). Kinase activity of OsBRI1 is essential for brassinosteroids to regulate rice growth and development Plant Sci 199-200, 113-120. DOI:10.1016/j.plantsci.2012.10.011URLPMID:23265324 [本文引用: 1] Brassinosteroids (BRs) are steroid hormones that participate in multiple biological processes. In this paper, we characterized a classic rice mutant Fn189 (dwarf54, d54) showing semi-dwarf stature and erect leaves. The coleoptile elongation and root growth was less affected in Fn189 than wild-type plant by the exogenous application of eBL, the most active form of BRs. Lamina joint inclination assay and morphological analysis in darkness further showed that Fn189 mutant plant was insensitive to exogenous eBL. Through map-based cloning, Fn189 was found to be a novel allelic mutant of the DWARF 61 (D61) gene, which encodes the putative BRs receptor OsBRI1. A single base mutation caused the I834F substitution in the OsBRI1 kinase domain. Consequently, kinase activity of OsBRI1 was found to decrease dramatically. Taken together, the kinase activity of OsBRI1 is essential for brassinosteroids to regulate normal plant growth and development in rice.
ZhouSR, YinLL, XueHW (2013). Functional genomics based understanding of rice endosperm development Curr Opin Plant Biol 16, 236-246. URLPMID:23582455 [本文引用: 1]
ZuoJR, LiJY (2014). Molecular genetic dissection of quantitative trait loci regulating rice grain size Annu Rev Genet 48, 99-118. [本文引用: 1]
水稻粒形遗传的研究进展 1 2011
... 水稻(Oryza sativa)作为最重要的谷物粮食之一, 是全世界超过一半人口的主要食物来源.然而, 现阶段人口和粮食需求不断增加, 使农业面临巨大挑战 , 且随着可利用土地面积和自然资源逐渐减少, 需要提高作物的生产效率, 其中增加单位面积产量尤为重要(Li et al., 2018).高等植物的种子由胚、胚乳和种皮三部分组成, 它们分别由受精卵、受精卵中心细胞和母体细胞发育而来, 共同协调控制种子大小.水稻胚乳占据成熟种子大部分空间, 主要成分为淀粉, 是人类的主要食物来源, 也是决定粮食重量的直接因素(James et al., 2003; Zhou et al., 2013).影响水稻产量的主要因素是单位面积有效穗数、每穗粒数、千粒重和灌浆速率(Sakamoto and Matsuoka, 2008; Xing and Zhang, 2010).粒重受水稻籽粒宽度、长度和粒厚协调控制, 说明水稻籽粒大小与产量息息相关.在单子叶植物水稻中, 籽粒长在颖壳内且籽粒大小受颖壳限制(Song et al., 2007; Shomura et al., 2008).籽粒大小受颖壳细胞数目和细胞大小调控, 细胞数目主要通过细胞增殖改变, 细胞大小受细胞扩展影响.近年来, 与水稻籽粒大小相关的基因逐渐被克隆, 其中一些基因对于提高水稻产量具有重要意义(宫李辉等, 2011). ...
油菜素内酯(BR)促进植物生长机理研究进展 1 2001
... 油菜素内酯影响植株生长(侯雷平和李梅兰, 2001).之前报道过OsBRI1的多个等位突变体, 其中仅编码的氨基酸发生改变的弱等位突变体表型变化比较微弱, 使得水稻株高变矮和籽粒变小, 其突变体表型与smg12类似(Nakamura et al., 2006).但另外一些由于重要氨基酸改变或者编码提前终止将会导致表型严重受损甚至不育.突变体d61-4在OsBRI1的第847个氨基酸处由谷氨酸变为终止密码子, 造成编码提前终止, 使其缺失了几乎所有蛋白激酶结构域, 表现为植株明显矮化、叶片严重扭曲(Nakamura et al., 2006).因此smg12可能是OsBRI1的一个新的弱等位突变.此外, OsBRI1调控种子大小的细胞学基础并不清楚, 本研究表明其通过调控细胞大小从而决定种子大小. ...
Genome sequencing reveals agronomically important loci in rice using Mutmap 3 2012
... 为深入研究水稻籽粒大小的调控机制, 我们在品种宽叶粳(KYJ)背景下筛选到1个小粒突变体smg12.利用Mutmap方法对smg12的候选基因进行克隆(Abe et al., 2012; Takagi et al., 2015), 鉴定到候选基因为OsBRI1. ...
... 本研究采用Mutmap方法对候选基因进行克隆(Abe et al., 2012; Takagi et al., 2015).将小粒突变体smg12与野生型KYJ杂交获得F1, F1自交获得F2分离群体.在抽穗期以后取50株小粒矮秆表型植株的叶片, 等重量叶片混合后提取DNA并进行全基因组重测序, 并以F2代中与野生型表型相似的50株植株作为对照进行基因组重测序, 然后进行SNP分析. ...
A novel cytochrome P450 is implicated in brassinosteroid biosynthesis via the characterization of a rice dwarf mutant, dwarf11, with reduced seed length 2 2005
... 目前的研究已经揭示了多条调控水稻籽粒大小的信号途径, 包括泛素蛋白酶体途径、G蛋白信号途径、丝裂原活化蛋白激酶(mitogen-activated protein kinase, MAPK)途径、转录调控因子及植物激素等(Zuo and Li, 2014; Li et al., 2019).控制粒宽和粒重的基因GW2编码1个E3泛素连接酶, 负向调控细胞分裂.GW2功能的丧失导致颖壳细胞数目增加, 产生更大更宽的颖壳并加快灌浆速率, 从而使籽粒更宽更重, 产量更高(Song et al., 2007).GS3编码异源三聚体G蛋白的Gγ亚基, 是植物籽粒大小和器官大小的负调控因子.野生型等位基因对应中等大小籽粒, N端OSR (organ size regulation)结构域功能缺失突变体籽粒变长, 而C端的TNFR (tumor necrosis factor receptor)/NGFR (nerve growth factor receptor)和VWFC (von Willebrand factor type C)结构域对OSR结构域的功能有抑制作用, C端结构域功能缺失则会产生非常短的籽粒(Fan et al., 2006; Mao et al., 2010).丝裂原活化蛋白激酶信号通路在真核生物中的作用非常保守, 且与植物生长发育以及防御反应的多个过程相关(Meng and Zhang, 2013).MAPK途径组成包括MAPK激酶激酶(MKKK)、MAPK激酶(MKK)和MAPK, 这3种激酶依次激活控制下游基因的表达, 调控植物生长发育.水稻OsMKKK10、OsMKK4和OsMAPK6作为1个级联信号通路促进细胞增殖, 从而正向调控水稻籽粒大小和粒重(Xu et al., 2018b).其功能缺失突变体osmkkk10、osmkk4和osmapk6由于颖壳细胞增殖过程受到限制, 导致籽粒变小(Liu et al., 2015b; Xu et al., 2018a, 2018b).OsSPL16编码1种细胞增殖的正调控蛋白, 过表达该基因会促进细胞分裂和灌浆, 使水稻籽粒变宽, 产量增加(Wang et al., 2012).水稻中重要的数量位点GLW7编码植物特异性转录因子OsSPL13, 通过正向调控颖壳细胞大小提高籽粒长度和产量(Si et al., 2016).植物激素对种子发育有重要作用并直接影响籽粒大小.BG1是正向调控籽粒大小的膜蛋白, 有研究显示BG1能够响应生长素变化, 且与生长素转运过程相关, 表明其可能通过生长素信号来调控籽粒生长.BG1的激活促进细胞扩展, 从而增加籽粒大小和粒重, 而BG1的表达被抑制时会形成小的籽粒(Liu et al., 2015a).近年来, 油菜素内酯对种子发育的调控机制已经在水稻、拟南芥(Arabidopsis thaliana)及其它植物中进行了深入研究.油菜素内酯对株高、叶夹角、分蘖数、籽粒大小和产量都有不同程度的影响(Morinaka et al., 2006).水稻中油菜素内酯受体基因D61的突变和油菜素内酯信号的负调控基因GSK2的过表达植株都会产生更小的籽粒(Tanabe et al., 2005; Tong et al., 2012).BU1编码1个具有螺旋-环-螺旋结构的转录因子, 作为响应油菜素内酯信号的正调控因子作用于D61基因的下游, BU1基因过表达导致水稻籽粒变大(Tanaka et al., 2009).上述结果表明, 籽粒大小调控是非常复杂的过程, 涉及多种信号转导以及生化代谢途径. ...
BRASSINOSTEROID UPREGULATED1, encoding a helix-loop-helix protein, is a novel gene involved in brassinosteroid signaling and controls bending of the lamina joint in rice 1 2009
... 目前的研究已经揭示了多条调控水稻籽粒大小的信号途径, 包括泛素蛋白酶体途径、G蛋白信号途径、丝裂原活化蛋白激酶(mitogen-activated protein kinase, MAPK)途径、转录调控因子及植物激素等(Zuo and Li, 2014; Li et al., 2019).控制粒宽和粒重的基因GW2编码1个E3泛素连接酶, 负向调控细胞分裂.GW2功能的丧失导致颖壳细胞数目增加, 产生更大更宽的颖壳并加快灌浆速率, 从而使籽粒更宽更重, 产量更高(Song et al., 2007).GS3编码异源三聚体G蛋白的Gγ亚基, 是植物籽粒大小和器官大小的负调控因子.野生型等位基因对应中等大小籽粒, N端OSR (organ size regulation)结构域功能缺失突变体籽粒变长, 而C端的TNFR (tumor necrosis factor receptor)/NGFR (nerve growth factor receptor)和VWFC (von Willebrand factor type C)结构域对OSR结构域的功能有抑制作用, C端结构域功能缺失则会产生非常短的籽粒(Fan et al., 2006; Mao et al., 2010).丝裂原活化蛋白激酶信号通路在真核生物中的作用非常保守, 且与植物生长发育以及防御反应的多个过程相关(Meng and Zhang, 2013).MAPK途径组成包括MAPK激酶激酶(MKKK)、MAPK激酶(MKK)和MAPK, 这3种激酶依次激活控制下游基因的表达, 调控植物生长发育.水稻OsMKKK10、OsMKK4和OsMAPK6作为1个级联信号通路促进细胞增殖, 从而正向调控水稻籽粒大小和粒重(Xu et al., 2018b).其功能缺失突变体osmkkk10、osmkk4和osmapk6由于颖壳细胞增殖过程受到限制, 导致籽粒变小(Liu et al., 2015b; Xu et al., 2018a, 2018b).OsSPL16编码1种细胞增殖的正调控蛋白, 过表达该基因会促进细胞分裂和灌浆, 使水稻籽粒变宽, 产量增加(Wang et al., 2012).水稻中重要的数量位点GLW7编码植物特异性转录因子OsSPL13, 通过正向调控颖壳细胞大小提高籽粒长度和产量(Si et al., 2016).植物激素对种子发育有重要作用并直接影响籽粒大小.BG1是正向调控籽粒大小的膜蛋白, 有研究显示BG1能够响应生长素变化, 且与生长素转运过程相关, 表明其可能通过生长素信号来调控籽粒生长.BG1的激活促进细胞扩展, 从而增加籽粒大小和粒重, 而BG1的表达被抑制时会形成小的籽粒(Liu et al., 2015a).近年来, 油菜素内酯对种子发育的调控机制已经在水稻、拟南芥(Arabidopsis thaliana)及其它植物中进行了深入研究.油菜素内酯对株高、叶夹角、分蘖数、籽粒大小和产量都有不同程度的影响(Morinaka et al., 2006).水稻中油菜素内酯受体基因D61的突变和油菜素内酯信号的负调控基因GSK2的过表达植株都会产生更小的籽粒(Tanabe et al., 2005; Tong et al., 2012).BU1编码1个具有螺旋-环-螺旋结构的转录因子, 作为响应油菜素内酯信号的正调控因子作用于D61基因的下游, BU1基因过表达导致水稻籽粒变大(Tanaka et al., 2009).上述结果表明, 籽粒大小调控是非常复杂的过程, 涉及多种信号转导以及生化代谢途径. ...
DWARF AND LOW-TILLERING acts as a direct downstream target of a GSK3/SHAGGY-like kinase to mediate brassinosteroid responses in rice 1 2012
... 目前的研究已经揭示了多条调控水稻籽粒大小的信号途径, 包括泛素蛋白酶体途径、G蛋白信号途径、丝裂原活化蛋白激酶(mitogen-activated protein kinase, MAPK)途径、转录调控因子及植物激素等(Zuo and Li, 2014; Li et al., 2019).控制粒宽和粒重的基因GW2编码1个E3泛素连接酶, 负向调控细胞分裂.GW2功能的丧失导致颖壳细胞数目增加, 产生更大更宽的颖壳并加快灌浆速率, 从而使籽粒更宽更重, 产量更高(Song et al., 2007).GS3编码异源三聚体G蛋白的Gγ亚基, 是植物籽粒大小和器官大小的负调控因子.野生型等位基因对应中等大小籽粒, N端OSR (organ size regulation)结构域功能缺失突变体籽粒变长, 而C端的TNFR (tumor necrosis factor receptor)/NGFR (nerve growth factor receptor)和VWFC (von Willebrand factor type C)结构域对OSR结构域的功能有抑制作用, C端结构域功能缺失则会产生非常短的籽粒(Fan et al., 2006; Mao et al., 2010).丝裂原活化蛋白激酶信号通路在真核生物中的作用非常保守, 且与植物生长发育以及防御反应的多个过程相关(Meng and Zhang, 2013).MAPK途径组成包括MAPK激酶激酶(MKKK)、MAPK激酶(MKK)和MAPK, 这3种激酶依次激活控制下游基因的表达, 调控植物生长发育.水稻OsMKKK10、OsMKK4和OsMAPK6作为1个级联信号通路促进细胞增殖, 从而正向调控水稻籽粒大小和粒重(Xu et al., 2018b).其功能缺失突变体osmkkk10、osmkk4和osmapk6由于颖壳细胞增殖过程受到限制, 导致籽粒变小(Liu et al., 2015b; Xu et al., 2018a, 2018b).OsSPL16编码1种细胞增殖的正调控蛋白, 过表达该基因会促进细胞分裂和灌浆, 使水稻籽粒变宽, 产量增加(Wang et al., 2012).水稻中重要的数量位点GLW7编码植物特异性转录因子OsSPL13, 通过正向调控颖壳细胞大小提高籽粒长度和产量(Si et al., 2016).植物激素对种子发育有重要作用并直接影响籽粒大小.BG1是正向调控籽粒大小的膜蛋白, 有研究显示BG1能够响应生长素变化, 且与生长素转运过程相关, 表明其可能通过生长素信号来调控籽粒生长.BG1的激活促进细胞扩展, 从而增加籽粒大小和粒重, 而BG1的表达被抑制时会形成小的籽粒(Liu et al., 2015a).近年来, 油菜素内酯对种子发育的调控机制已经在水稻、拟南芥(Arabidopsis thaliana)及其它植物中进行了深入研究.油菜素内酯对株高、叶夹角、分蘖数、籽粒大小和产量都有不同程度的影响(Morinaka et al., 2006).水稻中油菜素内酯受体基因D61的突变和油菜素内酯信号的负调控基因GSK2的过表达植株都会产生更小的籽粒(Tanabe et al., 2005; Tong et al., 2012).BU1编码1个具有螺旋-环-螺旋结构的转录因子, 作为响应油菜素内酯信号的正调控因子作用于D61基因的下游, BU1基因过表达导致水稻籽粒变大(Tanaka et al., 2009).上述结果表明, 籽粒大小调控是非常复杂的过程, 涉及多种信号转导以及生化代谢途径. ...
Control of grain size, shape and quality by OsSPL16 in rice 1 2012
... 目前的研究已经揭示了多条调控水稻籽粒大小的信号途径, 包括泛素蛋白酶体途径、G蛋白信号途径、丝裂原活化蛋白激酶(mitogen-activated protein kinase, MAPK)途径、转录调控因子及植物激素等(Zuo and Li, 2014; Li et al., 2019).控制粒宽和粒重的基因GW2编码1个E3泛素连接酶, 负向调控细胞分裂.GW2功能的丧失导致颖壳细胞数目增加, 产生更大更宽的颖壳并加快灌浆速率, 从而使籽粒更宽更重, 产量更高(Song et al., 2007).GS3编码异源三聚体G蛋白的Gγ亚基, 是植物籽粒大小和器官大小的负调控因子.野生型等位基因对应中等大小籽粒, N端OSR (organ size regulation)结构域功能缺失突变体籽粒变长, 而C端的TNFR (tumor necrosis factor receptor)/NGFR (nerve growth factor receptor)和VWFC (von Willebrand factor type C)结构域对OSR结构域的功能有抑制作用, C端结构域功能缺失则会产生非常短的籽粒(Fan et al., 2006; Mao et al., 2010).丝裂原活化蛋白激酶信号通路在真核生物中的作用非常保守, 且与植物生长发育以及防御反应的多个过程相关(Meng and Zhang, 2013).MAPK途径组成包括MAPK激酶激酶(MKKK)、MAPK激酶(MKK)和MAPK, 这3种激酶依次激活控制下游基因的表达, 调控植物生长发育.水稻OsMKKK10、OsMKK4和OsMAPK6作为1个级联信号通路促进细胞增殖, 从而正向调控水稻籽粒大小和粒重(Xu et al., 2018b).其功能缺失突变体osmkkk10、osmkk4和osmapk6由于颖壳细胞增殖过程受到限制, 导致籽粒变小(Liu et al., 2015b; Xu et al., 2018a, 2018b).OsSPL16编码1种细胞增殖的正调控蛋白, 过表达该基因会促进细胞分裂和灌浆, 使水稻籽粒变宽, 产量增加(Wang et al., 2012).水稻中重要的数量位点GLW7编码植物特异性转录因子OsSPL13, 通过正向调控颖壳细胞大小提高籽粒长度和产量(Si et al., 2016).植物激素对种子发育有重要作用并直接影响籽粒大小.BG1是正向调控籽粒大小的膜蛋白, 有研究显示BG1能够响应生长素变化, 且与生长素转运过程相关, 表明其可能通过生长素信号来调控籽粒生长.BG1的激活促进细胞扩展, 从而增加籽粒大小和粒重, 而BG1的表达被抑制时会形成小的籽粒(Liu et al., 2015a).近年来, 油菜素内酯对种子发育的调控机制已经在水稻、拟南芥(Arabidopsis thaliana)及其它植物中进行了深入研究.油菜素内酯对株高、叶夹角、分蘖数、籽粒大小和产量都有不同程度的影响(Morinaka et al., 2006).水稻中油菜素内酯受体基因D61的突变和油菜素内酯信号的负调控基因GSK2的过表达植株都会产生更小的籽粒(Tanabe et al., 2005; Tong et al., 2012).BU1编码1个具有螺旋-环-螺旋结构的转录因子, 作为响应油菜素内酯信号的正调控因子作用于D61基因的下游, BU1基因过表达导致水稻籽粒变大(Tanaka et al., 2009).上述结果表明, 籽粒大小调控是非常复杂的过程, 涉及多种信号转导以及生化代谢途径. ...
Genetic and molecular bases of rice yield 1 2010
... 水稻(Oryza sativa)作为最重要的谷物粮食之一, 是全世界超过一半人口的主要食物来源.然而, 现阶段人口和粮食需求不断增加, 使农业面临巨大挑战 , 且随着可利用土地面积和自然资源逐渐减少, 需要提高作物的生产效率, 其中增加单位面积产量尤为重要(Li et al., 2018).高等植物的种子由胚、胚乳和种皮三部分组成, 它们分别由受精卵、受精卵中心细胞和母体细胞发育而来, 共同协调控制种子大小.水稻胚乳占据成熟种子大部分空间, 主要成分为淀粉, 是人类的主要食物来源, 也是决定粮食重量的直接因素(James et al., 2003; Zhou et al., 2013).影响水稻产量的主要因素是单位面积有效穗数、每穗粒数、千粒重和灌浆速率(Sakamoto and Matsuoka, 2008; Xing and Zhang, 2010).粒重受水稻籽粒宽度、长度和粒厚协调控制, 说明水稻籽粒大小与产量息息相关.在单子叶植物水稻中, 籽粒长在颖壳内且籽粒大小受颖壳限制(Song et al., 2007; Shomura et al., 2008).籽粒大小受颖壳细胞数目和细胞大小调控, 细胞数目主要通过细胞增殖改变, 细胞大小受细胞扩展影响.近年来, 与水稻籽粒大小相关的基因逐渐被克隆, 其中一些基因对于提高水稻产量具有重要意义(宫李辉等, 2011). ...
Control of grain size and weight by the OsMKKK10-OsMKK4-OsMAPK6 signaling pathway in rice 1 2018
... 目前的研究已经揭示了多条调控水稻籽粒大小的信号途径, 包括泛素蛋白酶体途径、G蛋白信号途径、丝裂原活化蛋白激酶(mitogen-activated protein kinase, MAPK)途径、转录调控因子及植物激素等(Zuo and Li, 2014; Li et al., 2019).控制粒宽和粒重的基因GW2编码1个E3泛素连接酶, 负向调控细胞分裂.GW2功能的丧失导致颖壳细胞数目增加, 产生更大更宽的颖壳并加快灌浆速率, 从而使籽粒更宽更重, 产量更高(Song et al., 2007).GS3编码异源三聚体G蛋白的Gγ亚基, 是植物籽粒大小和器官大小的负调控因子.野生型等位基因对应中等大小籽粒, N端OSR (organ size regulation)结构域功能缺失突变体籽粒变长, 而C端的TNFR (tumor necrosis factor receptor)/NGFR (nerve growth factor receptor)和VWFC (von Willebrand factor type C)结构域对OSR结构域的功能有抑制作用, C端结构域功能缺失则会产生非常短的籽粒(Fan et al., 2006; Mao et al., 2010).丝裂原活化蛋白激酶信号通路在真核生物中的作用非常保守, 且与植物生长发育以及防御反应的多个过程相关(Meng and Zhang, 2013).MAPK途径组成包括MAPK激酶激酶(MKKK)、MAPK激酶(MKK)和MAPK, 这3种激酶依次激活控制下游基因的表达, 调控植物生长发育.水稻OsMKKK10、OsMKK4和OsMAPK6作为1个级联信号通路促进细胞增殖, 从而正向调控水稻籽粒大小和粒重(Xu et al., 2018b).其功能缺失突变体osmkkk10、osmkk4和osmapk6由于颖壳细胞增殖过程受到限制, 导致籽粒变小(Liu et al., 2015b; Xu et al., 2018a, 2018b).OsSPL16编码1种细胞增殖的正调控蛋白, 过表达该基因会促进细胞分裂和灌浆, 使水稻籽粒变宽, 产量增加(Wang et al., 2012).水稻中重要的数量位点GLW7编码植物特异性转录因子OsSPL13, 通过正向调控颖壳细胞大小提高籽粒长度和产量(Si et al., 2016).植物激素对种子发育有重要作用并直接影响籽粒大小.BG1是正向调控籽粒大小的膜蛋白, 有研究显示BG1能够响应生长素变化, 且与生长素转运过程相关, 表明其可能通过生长素信号来调控籽粒生长.BG1的激活促进细胞扩展, 从而增加籽粒大小和粒重, 而BG1的表达被抑制时会形成小的籽粒(Liu et al., 2015a).近年来, 油菜素内酯对种子发育的调控机制已经在水稻、拟南芥(Arabidopsis thaliana)及其它植物中进行了深入研究.油菜素内酯对株高、叶夹角、分蘖数、籽粒大小和产量都有不同程度的影响(Morinaka et al., 2006).水稻中油菜素内酯受体基因D61的突变和油菜素内酯信号的负调控基因GSK2的过表达植株都会产生更小的籽粒(Tanabe et al., 2005; Tong et al., 2012).BU1编码1个具有螺旋-环-螺旋结构的转录因子, 作为响应油菜素内酯信号的正调控因子作用于D61基因的下游, BU1基因过表达导致水稻籽粒变大(Tanaka et al., 2009).上述结果表明, 籽粒大小调控是非常复杂的过程, 涉及多种信号转导以及生化代谢途径. ...
A mitogen-activated protein kinase phosphatase influences grain size and weight in rice 2 2018
... 目前的研究已经揭示了多条调控水稻籽粒大小的信号途径, 包括泛素蛋白酶体途径、G蛋白信号途径、丝裂原活化蛋白激酶(mitogen-activated protein kinase, MAPK)途径、转录调控因子及植物激素等(Zuo and Li, 2014; Li et al., 2019).控制粒宽和粒重的基因GW2编码1个E3泛素连接酶, 负向调控细胞分裂.GW2功能的丧失导致颖壳细胞数目增加, 产生更大更宽的颖壳并加快灌浆速率, 从而使籽粒更宽更重, 产量更高(Song et al., 2007).GS3编码异源三聚体G蛋白的Gγ亚基, 是植物籽粒大小和器官大小的负调控因子.野生型等位基因对应中等大小籽粒, N端OSR (organ size regulation)结构域功能缺失突变体籽粒变长, 而C端的TNFR (tumor necrosis factor receptor)/NGFR (nerve growth factor receptor)和VWFC (von Willebrand factor type C)结构域对OSR结构域的功能有抑制作用, C端结构域功能缺失则会产生非常短的籽粒(Fan et al., 2006; Mao et al., 2010).丝裂原活化蛋白激酶信号通路在真核生物中的作用非常保守, 且与植物生长发育以及防御反应的多个过程相关(Meng and Zhang, 2013).MAPK途径组成包括MAPK激酶激酶(MKKK)、MAPK激酶(MKK)和MAPK, 这3种激酶依次激活控制下游基因的表达, 调控植物生长发育.水稻OsMKKK10、OsMKK4和OsMAPK6作为1个级联信号通路促进细胞增殖, 从而正向调控水稻籽粒大小和粒重(Xu et al., 2018b).其功能缺失突变体osmkkk10、osmkk4和osmapk6由于颖壳细胞增殖过程受到限制, 导致籽粒变小(Liu et al., 2015b; Xu et al., 2018a, 2018b).OsSPL16编码1种细胞增殖的正调控蛋白, 过表达该基因会促进细胞分裂和灌浆, 使水稻籽粒变宽, 产量增加(Wang et al., 2012).水稻中重要的数量位点GLW7编码植物特异性转录因子OsSPL13, 通过正向调控颖壳细胞大小提高籽粒长度和产量(Si et al., 2016).植物激素对种子发育有重要作用并直接影响籽粒大小.BG1是正向调控籽粒大小的膜蛋白, 有研究显示BG1能够响应生长素变化, 且与生长素转运过程相关, 表明其可能通过生长素信号来调控籽粒生长.BG1的激活促进细胞扩展, 从而增加籽粒大小和粒重, 而BG1的表达被抑制时会形成小的籽粒(Liu et al., 2015a).近年来, 油菜素内酯对种子发育的调控机制已经在水稻、拟南芥(Arabidopsis thaliana)及其它植物中进行了深入研究.油菜素内酯对株高、叶夹角、分蘖数、籽粒大小和产量都有不同程度的影响(Morinaka et al., 2006).水稻中油菜素内酯受体基因D61的突变和油菜素内酯信号的负调控基因GSK2的过表达植株都会产生更小的籽粒(Tanabe et al., 2005; Tong et al., 2012).BU1编码1个具有螺旋-环-螺旋结构的转录因子, 作为响应油菜素内酯信号的正调控因子作用于D61基因的下游, BU1基因过表达导致水稻籽粒变大(Tanaka et al., 2009).上述结果表明, 籽粒大小调控是非常复杂的过程, 涉及多种信号转导以及生化代谢途径. ...
... , 2018b).OsSPL16编码1种细胞增殖的正调控蛋白, 过表达该基因会促进细胞分裂和灌浆, 使水稻籽粒变宽, 产量增加(Wang et al., 2012).水稻中重要的数量位点GLW7编码植物特异性转录因子OsSPL13, 通过正向调控颖壳细胞大小提高籽粒长度和产量(Si et al., 2016).植物激素对种子发育有重要作用并直接影响籽粒大小.BG1是正向调控籽粒大小的膜蛋白, 有研究显示BG1能够响应生长素变化, 且与生长素转运过程相关, 表明其可能通过生长素信号来调控籽粒生长.BG1的激活促进细胞扩展, 从而增加籽粒大小和粒重, 而BG1的表达被抑制时会形成小的籽粒(Liu et al., 2015a).近年来, 油菜素内酯对种子发育的调控机制已经在水稻、拟南芥(Arabidopsis thaliana)及其它植物中进行了深入研究.油菜素内酯对株高、叶夹角、分蘖数、籽粒大小和产量都有不同程度的影响(Morinaka et al., 2006).水稻中油菜素内酯受体基因D61的突变和油菜素内酯信号的负调控基因GSK2的过表达植株都会产生更小的籽粒(Tanabe et al., 2005; Tong et al., 2012).BU1编码1个具有螺旋-环-螺旋结构的转录因子, 作为响应油菜素内酯信号的正调控因子作用于D61基因的下游, BU1基因过表达导致水稻籽粒变大(Tanaka et al., 2009).上述结果表明, 籽粒大小调控是非常复杂的过程, 涉及多种信号转导以及生化代谢途径. ...
Brassinosteroid-mediated regulation of agronomic traits in rice 1 2014
... OsBRI1编码1个由1 211个氨基酸残基组成的油菜素内酯受体激酶, 包含跨膜结构域、亮氨酸富集结构域和蛋白激酶结构域, 是拟南芥BRI1基因的直系同源基因.油菜素内酯在植物种子萌发、株高增加和生长发育等过程中都有重要作用(Zhang et al., 2014). ...
Kinase activity of OsBRI1 is essential for brassinosteroids to regulate rice growth and development 1 2013
 ,1,*, 李云海
,1,*, 李云海
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT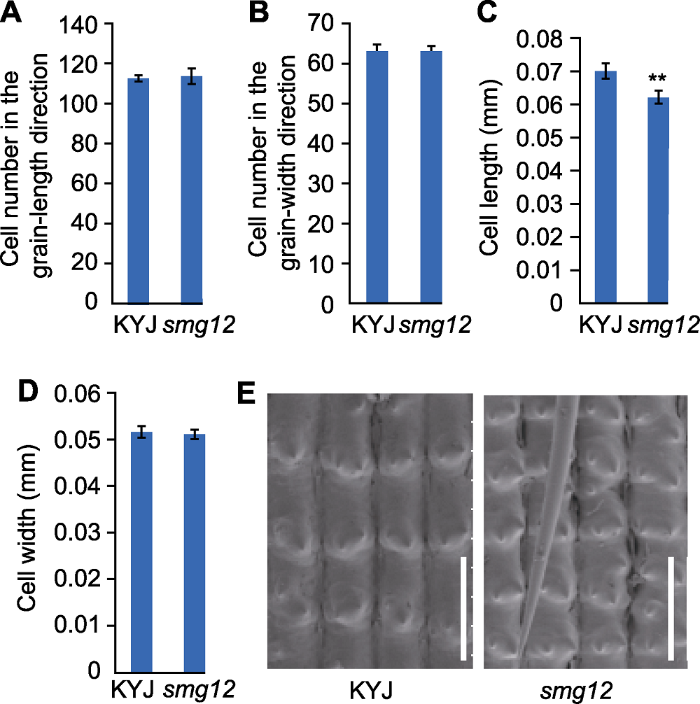 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT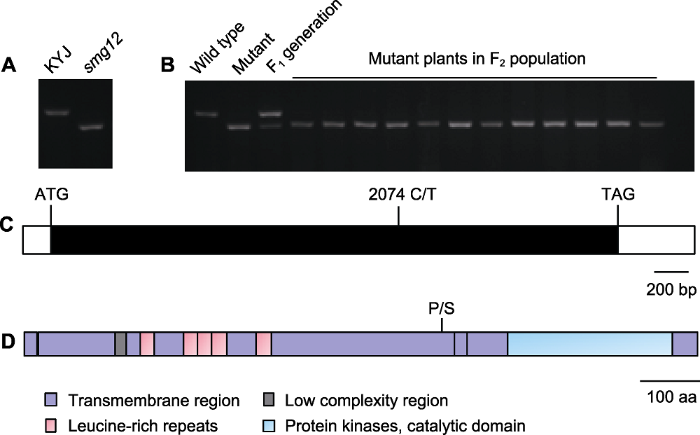 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}