 ,1,*
,1,*Response of AtR8 lncRNA to Salt Stress and Its Regulation on Seed Germination in Arabidopsis
Nan Zhang1, Ziguang Liu2, Shichen Sun3, Shengyi Liu4, Jianhui Lin1, Yifang Peng5, Xiaoxu Zhang1, He Yang1, Xi Cen1, Juan Wu,1,*通讯作者:
责任编辑: 孙冬花
收稿日期:2019-12-18接受日期:2020-04-15网络出版日期:2020-07-01
| 基金资助: |
Corresponding authors:
Received:2019-12-18Accepted:2020-04-15Online:2020-07-01

摘要
关键词:
Abstract
Keywords:
PDF (1148KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
张楠, 刘自广, 孙世臣, 刘圣怡, 林建辉, 彭疑芳, 张晓旭, 杨贺, 岑曦, 吴娟. 拟南芥AtR8 lncRNA对盐胁迫响应及其对种子萌发的调节作用. 植物学报, 2020, 55(4): 421-429 doi:10.11983/CBB19244
Zhang Nan, Liu Ziguang, Sun Shichen, Liu Shengyi, Lin Jianhui, Peng Yifang, Zhang Xiaoxu, Yang He, Cen Xi, Wu Juan.
种子是植物特有的繁殖器官。种子萌发是指有活力的种子吸胀后, 重新开始物质合成与代谢活动, 促使胚根露出种皮的过程。种子萌发极易受到光照、温度、水分和盐分等外部环境因素的影响, 同时受多基因调控。例如, 脱落酸(abscisic acid, ABA)胁迫下, WRKY41通过直接调节ABI3 (ABA-insensitive 3)的表达影响拟南芥(Arabidopsis thaliana)种子萌发(Ding et al., 2014); WRKY2通过影响ABA信号通路中ABI3、ABI5 (ABA-insensitive 5)、EM1和EM6 (early methionine-labeled 1 and 6)的表达, 抑制拟南芥种子萌发(Jiang and Yu, 2009); WRKY6通过影响RAV1 (related to ABA-insensitive3/vivparous1)及其下游ABI3、ABI4和ABI5的表达调节拟南芥的种子萌发(Huang et al., 2016)。此外, 还有研究表明, RSM1 (radialis-like sant/MYB 1)与HY5 (elongated hypocotyl 5)/HYH (HY5 homolog)结合后聚集在ABI5启动子上, 调节ABI5及其下游ABA应答基因的表达, 从而改变拟南芥萌发种子对ABA、NaCl以及甘露醇的敏感性(Yang et al., 2018)。CAMTA6 (calmodulin-bining transcription activator 6)通过调节Na+稳态和盐胁迫耐受性相关基因的表达, 影响拟南芥萌发种子对ABA的敏感性和盐胁迫的耐受性(Shkolnik et al., 2019)。尽管如此, 种子萌发的详细分子机制仍不十分清楚。
非编码RNA (non-coding RNA, ncRNAs)不编码蛋白质, 但却大量存在于生物体中发挥重要作用。长度大于200个核苷酸的非编码RNA为长链非编码RNA (long non-coding RNA, lncRNA), 它们通过充当诱饵、支架和增强子的方式调节许多基因的表达, 在剂量补偿(dosage compensation)、基因组印记(genomic imprinting)、X染色体失活(X chromosome inactivation)、发育和环境胁迫等生物过程中发挥重要作用(黄小庆等, 2015)。现已发现植物中Pol II转录的一些功能性lncRNAs, 并对其作用机制进行了解析。AtIPS1 (induced by phosphate starvation 1)通过模仿miR399的靶基因PHO2抑制miR399活性, 影响磷酸盐饥饿条件下拟南芥根的生长(Franco-Zorrilla et al., 2007; 黄小庆等, 2015); COOLAIR (cold induced antisense intragenic RNA)通过启动子转录干扰方式抑制开花调控基因FLC的表达, 进而调节植物开花时间(Swiezewski et al., 2009; Heo and Sung, 2011; 黄小庆等, 2015); COLDAIR (cold assisted intronic noncoding RNA)能引起FLC基因位点的组蛋白修饰, 导致FLC表观遗传沉默, 从而诱导植物快速开花(Kim and Sung, 2012; 黄小庆等, 2015); 水稻(Oryza sativa) LDMAR (long-day-specific male- fertility-associated RNA)序列中C碱基突变成G导致其启动子区域甲基化, 抑制其表达, 引起NK58S雄性不育(Ding et al., 2012; 黄小庆等, 2015)。热胁迫下, RNA测序分析发现了白菜(Brassica pekinensis)中4 594个lncRNA, 其中lncRNA (TCONS_00048391)通过模拟miR164a的靶基因NAC1, 影响白菜的耐热性(Wang et al., 2019)。Wu等(2019b)通过RNA-seq分析了梅花(Armeniaca mume)雌蕊发育中lncRNA的表达谱, 发现一些lncRNAs及其靶基因在雌蕊分化和花发育中发挥作用, 其中XR_514690.2和TCONS_ 00032517两个lncRNA可能在梅花多个雌蕊形成过程中发挥作用。Zhang等(2018)通过高通量测序, 发现LNC1和LNC2两个lncRNA可以作为miR156a和miR828a的靶基因, 分别抑制SPL9和诱导MYB114的表达, 调节沙棘(Hippophae rhamnoides)果实中花青素的合成。Qin等(2017)通过转录组分析发现, DRIR (drought induced lncRNA)能够增强拟南芥对干旱和盐胁迫的耐受性及对ABA的敏感性。Zhao等(2018)研究表明, MAS (MAF4 (mads affecting flowe-
ring 4)的反义转录lncRNA)通过与WDR5a相互作用激活MAF4的转录, 从而调节拟南芥的开花时间。Liu等(2019)研究发现, T5120 lncRNA过表达促进拟南芥硝酸盐同化并提高氮的利用率, 进而促进根生长。Zhang等(2019)研究表明, lncRNA973通过调节一系列盐胁迫相关基因的表达进而调控棉花(Gossypium spp.)对盐胁迫的响应。
近年来, 萌发相关ncRNA逐渐被发现并被解析, 如miR156、miR159、miR167和miR9678。拟南芥miR156基因下调SPL13 (SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE13)的表达, 延迟萌发后幼苗的发育(Martin et al., 2010a, 2010b)。miR159通过影响MYB101和MYB33的表达介导拟南芥的种子萌发(Reyes and Chua, 2007)。miR167通过调节ARF基因的表达影响生长素的信号转导, 在种子萌发以及胚发育过程中起重要作用(Rhoades et al., 2002)。miR9678通过调节GA/ABA信号转导影响小麦(Triticum aestivum)的种子萌发(Guo et al., 2018)。近几年, 参与种子萌发的长链非编码RNA也被发现。Xu等(2018)研究表明, 萌发的蓖麻(Ricinus communis)种子中多个DNA甲基化相关的lncRNA在胚乳和胚胎发育中起重要作用。Zhu等(2017)在玉米(Zea mays)种子中鉴定出753个lncRNA, 其中7个新lncRNA可能参与了玉米种子的发育和代谢过程。Yin等(2018)从发育的牡丹(Paeonia suffruticosa)种子中鉴定出22 430个lncRNA, 并预测39个lncRNA可能参与种子的脂肪酸合成和脂质代谢过程。BoNR8 lncRNA是甘蓝(B. oleracea)中RNA聚合酶III转录的长链非编码RNA, 拟南芥中BoNR8过表达影响ABA信号通路中重要基因的表达, 并抑制根生长和角果发育, 降低萌发种子对ABA的敏感性和盐胁迫的耐受性(Wu et al., 2019a)。
前期研究中, 我们根据RNA聚合酶III的转录活性及其转录的非编码RNA基因结构特征, 发现了拟南芥中RNA聚合酶III转录的AtR8 lncRNA (259 nt), 其在幼苗根端细胞质中大量表达, 并响应低氧胁迫(Wu et al., 2012); 且水杨酸(salicylic acid, SA)处理诱导萌发种子中AtR8 lncRNA的表达, AtR8 lncRNA缺失降低SA胁迫下的种子萌发(Li et al., 2016)。我们进一步研究发现, AtR8 lncRNA转录区域内包含保守的盐胁迫响应元件, 盐胁迫处理影响萌发种子中AtR8 lncRNA的表达, 且AtR8 lncRNA缺失降低了萌发种子对盐胁迫的耐受性, 表明AtR8 lncRNA在拟南芥种子萌发期盐胁迫中起重要作用。
1 材料与方法
1.1 植物材料
实验材料为野生型拟南芥(Arabidopsis thaliana L.)和AtR8 lncRNA部分缺失型拟南芥突变体(atr8, FLAG410H04), 后者购自凡尔赛拟南芥储备中心(versailles Arabidopsis stock center (http://publiclines.versailles.inra.fr/)) (Li et al., 2016)。1.2 各种应激应答处理及萌发率、鲜重和干重统计
将相同批次拟南芥野生型和atr8各50粒种子经70% (v/v)乙醇溶液和5% (v/v)次氯酸钠溶液表面消毒后, 播种在1/2MS固体培养基和分别含有50、100、150和200 mmol·L-1 NaCl的培养基上。4°C吸胀处理72小时后, 于22°C (16小时光照/8小时黑暗)培养箱中培养; 统计1、2、3、4、5、6和7天的种子萌发率(以胚根突破种皮为萌发标准)。称量萌发7天种子的鲜重, 70°C烘箱处理2天后, 称量萌发种子的干重。每个实验重复3次, 然后进行统计学分析。1.3 RNA提取方法
RNA提取参照Martin等(2005)和刘春晓等(2019)文献所述方法。称取0.1 g拟南芥干种子或萌发种子于液氮中彻底研磨后, 加入1 mL RNA提取液(45.5% (v/v)苯酚, 9% (v/v)氯仿, 0.45% (w/v) SDS, 41 mmol·L-1 LiCl, 2 mmol·L-1 EDTA, 5.9 mmol·L-1 β-巯基乙醇, 82 mmol·L-1 Tris-HCl), 混匀后离心。吸取上清, 加入等体积的PCI溶液(苯酚:氯仿:异戊醇=25:24:1, v/v/v); 离心后取上层溶液, 加入等体积的氯仿, 室温孵育; 离心取上层溶液加入1/3体积的8 mol·L-1 LiCl, 于-20°C静置过夜。次日, 离心取上清并加入1/4体积的异丙醇, 于-20°C静置30分钟; 离心取上清并加入3/5体积的异丙醇, -20°C静置30分钟; 离心后得RNA沉淀, 使用75%乙醇漂洗后加入适量的焦碳酸二乙酯(diethyl pyrocarbonate, DEPC)水, 于-80°C冰箱保存。1.4 Northern分析
将经6%聚丙烯酰胺凝胶电泳分离的RNA转移至尼龙膜上, UV交联固定后, 于42°C杂交箱预杂交1小时; 加入变性地高辛标记AtR8 lncRNA特异性RNA探针, 于42°C过夜杂交; 杂交后的膜于2× SSC (含0.1% SDS)及0.2× SSC (含0.1% SDS)洗液中清洗2次; 室温封闭1小时后, 与稀释2 000倍的Anti-Digoxigenin- AP抗体反应2小时; 马来酸洗液洗膜3次; CDP-Star暗处反应15分钟后, 使用LAS-4 000化学发光系统检测信号。1.5 整体原位杂交(in situ)
将萌发2天的拟南芥种子经固定液(4% (w/v)多聚甲醛, 15% (v/v) DMSO, 0.1% (v/v) tween 20, 0.08 mol·L-1 EGTA (pH8.0)) 4°C固定3天后, 进行脱水、水饱和及蛋白酶K处理。与地高辛标记AtR8 lncRNA特异性探针于42°C过夜杂交; 杂交后的样品经水洗加入blocking封闭2小时, 于4°C与稀释1 500倍的Anti-Digoxigenin-AP抗体反应过夜, 经BM Purple检测信号。2 结果与讨论
2.1 种子萌发过程中AtR8 lncRNA的表达特性分析
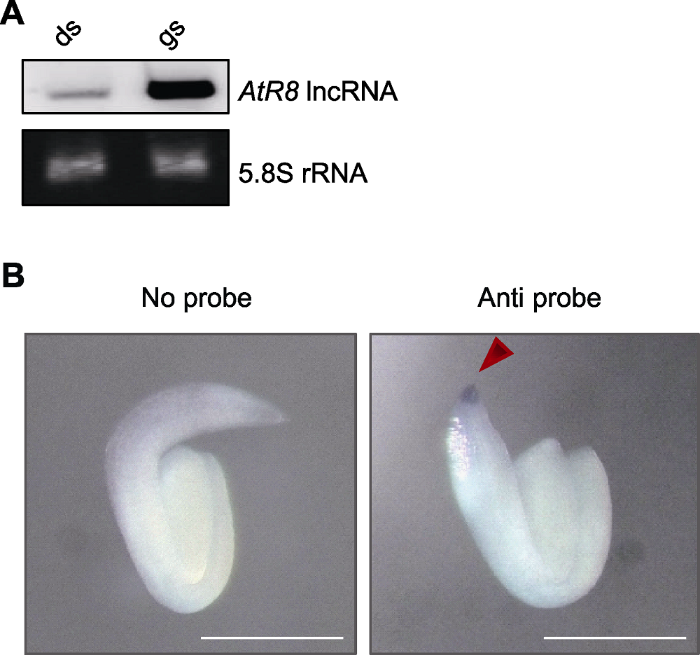
为确定拟南芥种子萌发过程中AtR8 lncRNA的表达特性, 提取其干种子和萌发48小时种子的RNA进行Northern分析, 发现AtR8 lncRNA在萌发种子中大量表达(图1A)。整体原位杂交实验进一步确认AtR8 lncRNA于萌发种子的根尖端大量表达(图1B), 表明拟南芥萌发种子中AtR8 lncRNA的表达具有高度的组织特异性。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1拟南芥种子萌发过程中AtR8 lncRNA的表达特性分析
(A) 拟南芥种子萌发过程中AtR8 lncRNA表达特性的Northern分析(以5.8S rRNA作为上样对照); (B) 拟南芥种子萌发过程中AtR8 lncRNA组织表达特性的整体原位杂交(箭头指示AtR8 lncRNA信号) (Bars=200 μm)。
Figure 1Analysis of AtR8 lncRNA expression during seed germination in Arabidopsis thaliana
(A) Northern blotting analysis of AtR8 lncRNA expression during seed germination (expression of 5.8S rRNA serves as loading controls); (B) In situ hybridization of AtR8 lncRNA expression during seed germination (arrow indicates AtR8 lncRNA signal) (Bars=200 μm).
2.2 种子萌发过程中AtR8 lncRNA表达响应盐胁迫
UCC盐胁迫响应元件(TCTTCTTCTTTA)是盐应答基因中高度保守的序列。Di等(2014)证明长链非编码RNA中存在该保守UCC元件。因此, 我们比较了AtR8 lncRNA序列与UCC元件, 发现AtR8 lncRNA转录区域内存在UCC盐胁迫响应元件, 序列相似性达75% (9 bp/12 bp) (图2A), 这表明AtR8 lncRNA可能响应盐胁迫。RNAlogo (http://rnalogo.mbc.nctu.edu.tw/index.php)预测到UCC元件存在于AtR8 lncRNA二级结构的茎环相连位置(图2B)。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2拟南芥种子萌发过程中AtR8 lncRNA表达响应NaCl逆境胁迫
(A) AtR8 lncRNA与UCC盐胁迫响应元件的序列比较分析(USE、TATA启动子序列为大写、加粗并加框, AtR8 lncRNA转录区域为大写并加粗, 保守的盐胁迫响应元件用星号标注)。(B) AtR8 lncRNA二级结构中盐胁迫响应元件存在位置的RNAlogo预测(盐胁迫响应元件为大写、加粗并用箭头指出)。(C) 拟南芥种子萌发过程中不同浓度NaCl处理下AtR8 lncRNA表达特性的Northern分析, 以5.8S rRNA作为上样对照。下方为Northern半定量分析, 2个独立的实验给出了相似的结果, 并显示了1个代表性的例子。值为平均值±标准误(t-检验, *P<0.05, **P<0.01)。(D) 拟南芥种子萌发过程中150 mmol·L-1 NaCl处理不同时间AtR8 lncRNA表达特性的Northern分析, 以5.8S rRNA作为上样对照。下方为Northern半定量分析, 2个独立的实验给出了相似的结果, 并显示了1个代表性的例子。值为平均值±标准误(t-检验, **P<0.01)。
Figure 2AtR8 lncRNA expression during seed germination of Arabidopsis thaliana after NaCl treatment
(A) Sequence comparison between AtR8 lncRNA and UCC salt stress-responsive element (the USE and TATA promoter sequences are capitalized, bloded and framed; the AtR8 lncRNA transcriptional region is capitalized and bolded; and the conserved salt stress-responsive element is marked with asterisk). (B) The location of the salt stress-responsive element in the secondary structure of AtR8 lncRNA predicted by RNAlogo (the salt stress-responsive motif is capitalized, bloded and indicated with an arrow). (C) Northern blotting analysis of AtR8 lncRNA expression in germinating seeds under different NaCl treatments, 5.8S rRNA was used as a loading control. The lower panel shows semi-quantitative analysis of the Northern blotting signals. Two independent experiments gave similar results, and a representative example is shown. Values are means ± SE (t-test, *P<0.05, **P<0.01). (D) Northern blotting analysis of AtR8 lncRNA in germinating seeds under different periods of 150 mmol·L-1 NaCl treatment, 5.8S rRNA was used as a loading control. The lower panel shows semi-quantitative analysis of the Northern blotting signals. Two independent experiments gave similar results, and a representative example is shown. Values are means ± SE (t-test, **P<0.01).
此外, 实验结果表明, 50、100、150和200 mmol·L-1 NaCl处理均抑制AtR8 lncRNA的表达(图2C), 且150 mmol·L-1 NaCl的抑制作用最明显。150 mmol·L-1 NaCl分别处理0、8、16、24、32和40小时的Northern分析结果进一步表明, 处理时间越长, AtR8 lncRNA表达受抑制越明显(图2D)。
2.3 盐胁迫下AtR8 lncRNA缺失抑制种子萌发
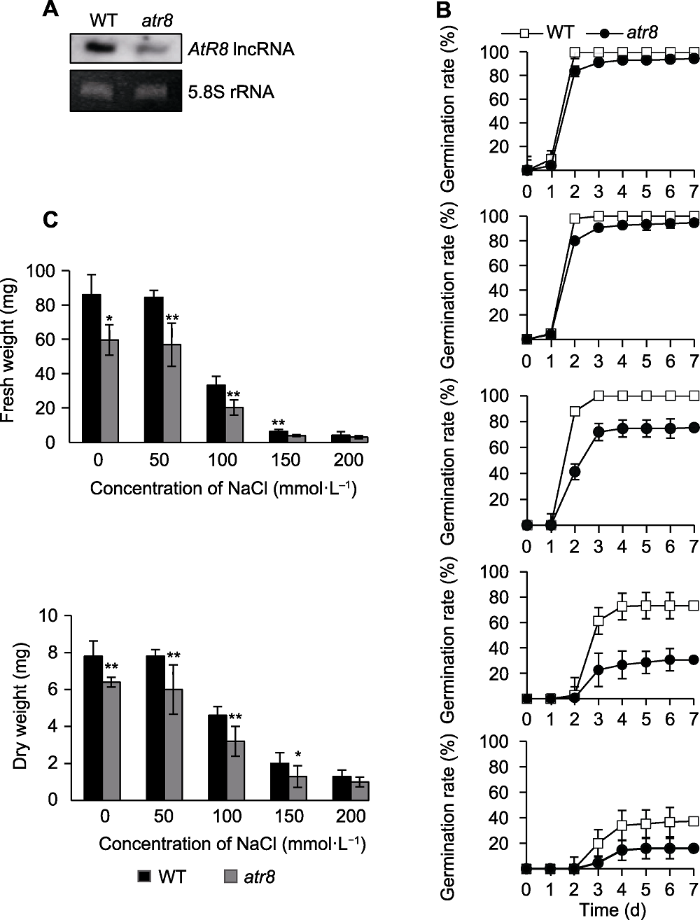
乔慧萍等(2007)研究表明, 盐胁迫对种子萌发具有抑制作用。我们使用AtR8 lncRNA部分缺失型突变体(atr8), 调查了盐胁迫下AtR8 lncRNA缺失是否影响种子萌发(图3A) (Li et al., 2016)。结果表明, 正常培养条件下, atr8种子萌发率低于野生型。50 mmol·L-1 NaCl处理时, 野生型和atr8种子萌发率与正常培养条件类似。100、150和200 mmol·L-1 NaCl处理时, 野生型和atr8种子萌发均受到抑制, 但对atr8种子萌发的抑制作用更明显(图3B)。对萌发7天的野生型与atr8种子鲜重和干重的分析结果表明, 正常培养条件下野生型的鲜重和干重均大于atr8。随着NaCl浓度的增加, 二者的鲜重和干重均明显降低, 但atr8的鲜重和干重仍明显低于野生型(图3C), 表明NaCl胁迫下, AtR8 lncRNA缺失进一步抑制了拟南芥的种子萌发。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3盐胁迫下AtR8 lncRNA缺失抑制拟南芥种子萌发
(A) Northern分析鉴定AtR8 lncRNA缺失突变体atr8 (以5.8S rRNA作为上样对照); (B) 不同浓度NaCl处理下, 野生型和atr8种子生长状况及萌发率统计分析(数据为3次独立试验的平均值, 误差为标准误); (C) 不同浓度NaCl处理下, 野生型和atr8萌发7天种子的鲜重及干重(数值为3次独立试验的平均值, 误差为标准误, 星号表示atr8与野生型的显著性差异(t-检验, *P<0.05, **P<0.01))。WT: 野生型
Figure 3Loss of AtR8 lncRNA inhibits Arabidopsis thaliana seed germination under salt stress
(A) Northern blotting analysis of the AtR8 lncRNA loss-of-function mutant atr8 (5.8S rRNA was used as the loading control); (B) Statistical analysis of growth and germination rate of the wild-type and atr8 seeds under different concentrations of NaCl (data are average of three independent experiments, and bars indicate standard error); (C) Fresh and dry weight of the wild-type and atr8 seeds after 7 d of germination under NaCl treatment (the values are average of three independent experiments, and bars indicate standard error (the asterisk indicate significant differences between atr8 and wild type (t-test, * P < 0.05, ** P < 0.01)). WT: Wild type
2.4 讨论
高盐环境会对植物造成离子毒害、渗透胁迫和矿质营养缺失, 使其生理代谢紊乱, 导致种子萌发和植株生长受到阻碍, 严重降低植物的产量和品质(韩志平等, 2015)。植物耐盐性是多基因控制的数量性状, 是多种耐盐生理性状的综合体现(孙兰菊等, 2001; 陈洁和林栖凤, 2003; 陆玉建等, 2012)。苏永全和吕迎春(2007)研究表明, 盐胁迫通过增效、负效和完全阻抑效应影响种子的萌发。低盐可增强种子的呼吸作用, 提高蛋白酶和脂肪酶活性, 促进贮藏物质的转化, 进而促进种子萌发和生长; 高盐条件下, 由于盐形成的渗透势阻碍种子吸水(盐浓度越高, 阻碍作用越强), 严重影响了种子内蛋白质等大分子物质的分解和合成进程, 降低种子的发芽率、发芽指数和活力指数(苏永全和吕迎春, 2007; 郝雪峰等, 2013)。盐胁迫下, γ-氨基丁酸(GABA)能够增强淀粉酶的活性, 使种子获得更多营养和能量, 改善种子的萌发质量(王泳超, 2016)。NaHS显著缓解了盐处理对种子萌发过程中水解酶活性的抑制作用(窦伟, 2010)。拟南芥U-Box泛素连接酶AtPUB18与AtPUB19双突变后降低了种子对高盐的敏感性(Bergler and Hoth, 2011; 张新宇等, 2014)。NAC57过表达拟南芥的种子发芽率、超氧化物歧化酶和过氧化物酶活性较高, 种子的耐盐性增强(Yao et al., 2018)。目前, 盐胁迫调节种子萌发的详细分子机制尚不十分清楚。前期研究表明, RNA聚合酶III转录的AtR8 Inc- RNA同源物BoNR8 lncRNA大量存在于甘蓝萌发种子的根部(Wu et al., 2019a)。本研究发现, AtR8 lncRNA在拟南芥萌发种子的根尖端特异表达, 表明AtR8 lncRNA与BoNR8 lncRNA均在种子萌发阶段特异性表达, 这表明种子萌发过程中RNA聚合酶III具有较高的转录活性, 可以转录与萌发相关的特殊非编码RNA。BoNR8 lncRNA和AtR8 lncRNA转录区域内都存在盐胁迫响应元件。盐胁迫下, BoNR8 lncRNA被诱导表达, AtR8 lncRNA的表达则被抑制, 表明AtR8 lncRNA与BoNR8 lncRNA均响应盐胁迫但表达趋势不同。拟南芥中BoNR8 lncRNA过表达影响ABA信号中RAV1、ABI3、ABI5、EM1和EM6等重要基因的表达, 抑制正常培养条件下的种子萌发、角果发育和幼苗根生长, 高盐胁迫进一步抑制种子萌发。AtR8 lncRNA缺失抑制正常培养条件下的种子萌发, 高盐胁迫也进一步抑制种子萌发, 但不影响根生长, 表明BoNR8 lncRNA与AtR8 lncRNA均参与拟南芥的生长发育和盐胁迫响应过程, 但它们的生物学功能存在差异。后续, 我们将从分子水平及遗传学角度详细分析种子萌发过程中AtR8 lncRNA参与盐胁迫的作用机制, 为培育优质丰产的耐盐作物新品种提供理论依据。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
硕士论文.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 5]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
博士论文.
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/j.1438-8677.2010.00431.xURLPMID:21815976 [本文引用: 1]

Plant U-box armadillo repeat (PUB-ARM) proteins represent a type of E3 ubiquitin ligase. PUB-ARM proteins have various functions in plant development, plant defence and plant stress responses. The so far uncharacterised PUB-ARM proteins AtPUB18 and AtPUB19 are highly homologous to each other and regulated on the transcript level by ABA and NaCl. To investigate their physiological function, we isolated and characterised two Arabidopsis mutants and named them pub18-1 and pub19-1. However, these single mutants did not show any ABA- or salt-dependent phenotype. In contrast, pub18-1/pub19-1 double mutant seeds were less sensitive to ABA and NaCl inhibition of seed germination compared to wild-type seeds, indicating that both PUB-ARM proteins may coordinately function as regulatory components in ABA or salt inhibition of germination.
DOI:10.1111/tpj.12679URLPMID:25256571 [本文引用: 1]
Recently, in addition to poly(A)+ long non-coding RNAs (lncRNAs), many lncRNAs without poly(A) tails, have been characterized in mammals. However, the non-polyA lncRNAs and their conserved motifs, especially those associated with environmental stresses, have not been fully investigated in plant genomes. We performed poly(A)- RNA-seq for seedlings of Arabidopsis thaliana under four stress conditions, and predicted lncRNA transcripts. We classified the lncRNAs into three confidence levels according to their expression patterns, epigenetic signatures and RNA secondary structures. Then, we further classified the lncRNAs to poly(A)+ and poly(A)- transcripts. Compared with poly(A)+ lncRNAs and coding genes, we found that poly(A)- lncRNAs tend to have shorter transcripts and lower expression levels, and they show significant expression specificity in response to stresses. In addition, their differential expression is significantly enriched in drought condition and depleted in heat condition. Overall, we identified 245 poly(A)+ and 58 poly(A)- lncRNAs that are differentially expressed under various stress stimuli. The differential expression was validated by qRT-PCR, and the signaling pathways involved were supported by specific binding of transcription factors (TFs), phytochrome-interacting factor 4 (PIF4) and PIF5. Moreover, we found many conserved sequence and structural motifs of lncRNAs from different functional groups (e.g. a UUC motif responding to salt and a AU-rich stem-loop responding to cold), indicated that the conserved elements might be responsible for the stress-responsive functions of lncRNAs.
DOI:10.1073/pnas.1121374109URLPMID:22308482 [本文引用: 1]
Hybrid rice has greatly contributed to the global increase of rice productivity. A major component that facilitated the development of hybrids was a mutant showing photoperiod-sensitive male sterility (PSMS) with its fertility regulated by day length. Transcriptome studies have shown that large portions of the eukaryotic genomic sequences are transcribed to long noncoding RNAs (lncRNAs). However, the potential roles for only a few lncRNAs have been brought to light at present. Thus, great efforts have to be invested to understand the biological functions of lncRNAs. Here we show that a lncRNA of 1,236 bases in length, referred to as long-day-specific male-fertility-associated RNA (LDMAR), regulates PSMS in rice. We found that sufficient amount of the LDMAR transcript is required for normal pollen development of plants grown under long-day conditions. A spontaneous mutation causing a single nucleotide polymorphism (SNP) between the wild-type and mutant altered the secondary structure of LDMAR. This change brought about increased methylation in the putative promoter region of LDMAR, which reduced the transcription of LDMAR specifically under long-day conditions, resulting in premature programmed cell death (PCD) in developing anthers, thus causing PSMS. Thus, a lncRNA could directly exert a major effect on a trait like a structure gene, and a SNP could alter the function of a lncRNA similar to amino acid substitution in structural genes. Molecular elucidating of PSMS has important implications for understanding molecular mechanisms of photoperiod regulation of many biological processes and also for developing male sterile germplasms for hybrid crop breeding.
URLPMID:24946881 [本文引用: 1]
DOI:10.1038/ng2079URLPMID:17643101 [本文引用: 1]
MicroRNAs (miRNA) regulate key aspects of development and physiology in animals and plants. These regulatory RNAs act as guides of effector complexes to recognize specific mRNA sequences based on sequence complementarity, resulting in translational repression or site-specific cleavage. In plants, most miRNA targets are cleaved and show almost perfect complementarity with the miRNAs around the cleavage site. Here, we examined the non-protein coding gene IPS1 (INDUCED BY PHOSPHATE STARVATION 1) from Arabidopsis thaliana. IPS1 contains a motif with sequence complementarity to the phosphate (Pi) starvation-induced miRNA miR-399, but the pairing is interrupted by a mismatched loop at the expected miRNA cleavage site. We show that IPS1 RNA is not cleaved but instead sequesters miR-399. Thus, IPS1 overexpression results in increased accumulation of the miR-399 target PHO2 mRNA and, concomitantly, in reduced shoot Pi content. Engineering of IPS1 to be cleavable abolishes its inhibitory activity on miR-399. We coin the term 'target mimicry' to define this mechanism of inhibition of miRNA activity. Target mimicry can be generalized beyond the control of Pi homeostasis, as demonstrated using artificial target mimics.
DOI:10.1105/tpc.17.00842URLPMID:29567662 [本文引用: 1]
Seed germination is important for grain yield and quality and rapid, near-simultaneous germination helps in cultivation; however, cultivars that germinate too readily can undergo preharvest sprouting (PHS), which causes substantial losses in areas that tend to get rain around harvest time. Moreover, our knowledge of mechanisms regulating seed germination in wheat (Triticum aestivum) remains limited. In this study, we analyzed function of a wheat-specific microRNA 9678 (miR9678), which is specifically expressed in the scutellum of developing and germinating seeds. Overexpression of miR9678 delayed germination and improved resistance to PHS in wheat through reducing bioactive gibberellin (GA) levels; miR9678 silencing enhanced germination rates. We provide evidence that miR9678 targets a long noncoding RNA (WSGAR) and triggers the generation of phased small interfering RNAs that play a role in the delay of seed germination. Finally, we found that abscisic acid (ABA) signaling proteins bind the promoter of miR9678 precursor and activate its expression, indicating that miR9678 affects germination by modulating the GA/ABA signaling.
URLPMID:21127216 [本文引用: 1]
DOI:10.1371/journal.pgen.1005833URLPMID:26829043 [本文引用: 1]
The phytohormone abscisic acid (ABA) plays important roles during seed germination and early seedling development. Here, we characterized the function of the Arabidopsis WRKY6 transcription factor in ABA signaling. The transcript of WRKY6 was repressed during seed germination and early seedling development, and induced by exogenous ABA. The wrky6-1 and wrky6-2 mutants were ABA insensitive, whereas WRKY6-overexpressing lines showed ABA-hypersensitive phenotypes during seed germination and early seedling development. The expression of RAV1 was suppressed in the WRKY6-overexpressing lines and elevated in the wrky6 mutants, and the expression of ABI3, ABI4, and ABI5, which was directly down-regulated by RAV1, was enhanced in the WRKY6-overexpressing lines and repressed in the wrky6 mutants. Electrophoretic mobility shift and chromatin immunoprecipitation assays showed that WRKY6 could bind to the RAV1 promoter in vitro and in vivo. Overexpression of RAV1 in WRKY6-overexpressing lines abolished their ABA-hypersensitive phenotypes, and the rav1 wrky6-2 double mutant showed an ABA-hypersensitive phenotype, similar to rav1 mutant. Together, the results demonstrated that the Arabidopsis WRKY6 transcription factor played important roles in ABA signaling by directly down-regulating RAV1 expression.
URLPMID:19622176 [本文引用: 1]
DOI:10.1016/j.pbi.2011.10.004URLPMID:22078062 [本文引用: 1]
In Arabidopsis, the role of the vernalization pathway is to repress expression of a potent floral repressor, FLOWERING LOCUS C (FLC), after a sufficient period of winter cold has been perceived. Following winter, the lack of FLC expression allows unimpeded operation of the photoperiod pathway and hence rapid flowering of vernalized plants in spring via the activation of floral integrator genes. Molecular studies revealed that regulation of the key floral repressor, FLC, is under the control of the interplay between Trithorax group (TrxG)-mediated activation and Polycomb group (PcG)-mediated repression. On-off switch of genes by TrxG and PcG is an evolutionarily conserved mechanism to coordinate cellular identity in eukaryotes. Regulation of FLC by external cues provides an excellent model system to study mechanisms in which cell identity is influenced by environment. In this review, we discuss coordinated contributions by protein and long noncoding RNA components to this environmentally induced epigenetic switch of a developmental program in plants.
[本文引用: 3]
DOI:10.1111/nph.16038URLPMID:31264223 [本文引用: 1]
Long noncoding RNAs (lncRNAs) are crucial regulators in many plant biological processes. However, it remains unknown whether lncRNAs can respond to nitrate or function in nitrate regulation. We detected 695 lncRNAs, 480 known and 215 novel, in Arabidopsis seedling roots; six showed altered expression in response to nitrate treatment, among which T5120 showed the highest induction. Overexpression of T5120 in Arabidopsis promoted the response to nitrate, enhanced nitrate assimilation and improved biomass and root development. Biochemical and molecular analyses revealed that NLP7, a master nitrate regulatory transcription factor, directly bound to the nitrate-responsive cis-element (NRE)-like motif of the T5120 promoter and activated T5120 transcription. In addition, T5120 partially restored the nitrate signalling and assimilation phenotypes of nlp7 mutant, suggesting that T5120 is involved in NLP7-mediated nitrate regulation. Interestingly, the expression of T5120 was regulated by the nitrate sensor NRT1.1. Therefore, T5120 is modulated by NLP7 and NRT1.1 to regulate nitrate signalling. Our work reveals a new regulatory mechanism in which lncRNA T5120 functions in nitrate regulation, providing new insights into the nitrate signalling network. Importantly, lncRNA T5120 can promote nitrate assimilation and plant growth to improve nitrogen use efficiency.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1104/pp.17.00574URLPMID:28887353 [本文引用: 1]
Long noncoding RNAs (lncRNAs) affect gene expression through a wide range of mechanisms and are considered as important regulators in many essential biological processes. A large number of lncRNA transcripts have been predicted or identified in plants in recent years. However, the biological functions for most of them are still unknown. In this study, we identified an Arabidopsis (Arabidopsis thaliana) lncRNA, DROUGHT INDUCED lncRNA (DRIR), as a novel positive regulator of the plant response to drought and salt stress. DRIR was expressed at a low level under nonstress conditions but can be significantly activated by drought and salt stress as well as by abscisic acid (ABA) treatment. We identified a T-DNA insertion mutant, drir(D) , which had higher expression of the DRIR gene than the wild-type plants. The drir(D) mutant exhibits increased tolerance to drought and salt stress. Overexpressing DRIR in Arabidopsis also increased tolerance to drought and salt stress of the transgenic plants. The drir(D) mutant and the overexpressing seedlings are more sensitive to ABA than the wild type in stomata closure and seedling growth. Genome-wide transcriptome analysis demonstrated that the expression of a large number of genes was altered in drir(D) and the overexpressing plants. These include genes involved in ABA signaling, water transport, and other stress-relief processes. Our study reveals a mechanism whereby DRIR regulates the plant response to abiotic stress by modulating the expression of a series of genes involved in the stress response.
DOI:10.1111/j.1365-313X.2006.02980.xURLPMID:17217461 [本文引用: 1]
Upon seed imbibition, abscisic acid (ABA) levels decrease to allow embryos to germinate and develop into seedlings. However, under abiotic stress conditions, ABA levels remain high, and growth and development are arrested. Several transcription factors, including abscisic acid-insensitive (ABI)3 and ABI5, are known to control this developmental checkpoint. Here, we show that, in germinating Arabidopsis thaliana seeds, ABA induces the accumulation of microRNA 159 (miR159) in an ABI3-dependent fashion, and miRNA159 mediates cleavage of MYB101 and MYB33 transcripts in vitro and in vivo. The two MYB transcription factors function as positive regulators of ABA responses, as null mutants of myb33 and myb101 show hyposensitivity to the hormone. Consistent with this, miR159 over-expression suppresses MYB33 and MYB101 transcript levels and renders plants hyposensitive to ABA, whereas transgenic plants over-expressing cleavage-resistant forms of MYB33 and MYB101 are hypersensitive, as are plants over-expressing the Turnip mosaic virus (TuMV) P1/HC-Pro viral protein that is known to inhibit miRNA function. Our results suggest that ABA-induced accumulation of miR159 is a homeostatic mechanism to direct MYB33 and MYB101 transcript degradation to desensitize hormone signaling during seedling stress responses.
DOI:10.1016/s0092-8674(02)00863-2URLPMID:12202040 [本文引用: 1]
We predict regulatory targets for 14 Arabidopsis microRNAs (miRNAs) by identifying mRNAs with near complementarity. Complementary sites within predicted targets are conserved in rice. Of the 49 predicted targets, 34 are members of transcription factor gene families involved in developmental patterning or cell differentiation. The near-perfect complementarity between plant miRNAs and their targets suggests that many plant miRNAs act similarly to small interfering RNAs and direct mRNA cleavage. The targeting of developmental transcription factors suggests that many plant miRNAs function during cellular differentiation to clear key regulatory transcripts from daughter cell lineages.
DOI:10.1104/pp.19.00119URLPMID:30894419 [本文引用: 1]
Salinity impairs seed germination and seedling establishment. We investigated the role of Arabidopsis (Arabidopsis thaliana) CALMODULIN-BINDING TRANSCRIPTION ACTIVATOR 6 (CAMTA6) in salinity stress responses during early germination. Compared with the wild type, the camta6-4 and camta6-5 mutants were more tolerant to NaCl and abscisic acid (ABA) and accumulated less Na(+) In contrast, 4- to 11-d-old camta6 seedlings were more sensitive to NaCl. In camta6, expression of HIGH-AFFINITY K(+) TRANSPORTER1 (AtHKT1;1), encoding an Na(+)/K(+) transporter, was restricted to the radicles and was not enhanced by NaCl or ABA. During germination, the camta6 hkt1 double mutant was as sensitive as the wild type and hkt1 to NaCl, suggesting that HKT1;1 is crucial for the salt tolerance of camta6 An ABA response element in the HKT1;1 promoter was found to be indispensable for the enhanced expression of the gene in response to NaCl and to ABA. Transcriptome analysis of the wild type and camta6-5 with and without salt treatment revealed 1,020 up-regulated and 1,467 down-regulated salt-responsive genes in the wild type. Among these, 638 up-regulated and 1,242 down-regulated genes were classified as CAMTA6-dependent. Expression of several known salt stress-associated genes, including SALT OVERLY SENSITIVE1 and Na(+)/H(+) ANTIPORTER, was impaired in camta6 mutants. Bioinformatics analysis of the 5' upstream sequences of the salt-responsive CAMTA6-dependent up-regulated genes revealed the CACGTGTC motif as the most prominent element, representing an ABA response element and a potential CAMTA-binding site. We suggest that CAMTA6 regulates, directly or indirectly, the expression of most of the salt-responsive genes in germinating seeds, including genes that are crucial for Na(+) homeostasis and salt stress tolerance.
DOI:10.1038/nature08618URLPMID:20010688 [本文引用: 1]
Transcription in eukaryotic genomes generates an extensive array of non-protein-coding RNA, the functional significance of which is mostly unknown. We are investigating the link between non-coding RNA and chromatin regulation through analysis of FLC - a regulator of flowering time in Arabidopsis and a target of several chromatin pathways. Here we use an unbiased strategy to characterize non-coding transcripts of FLC and show that sense/antisense transcript levels correlate in a range of mutants and treatments, but change independently in cold-treated plants. Prolonged cold epigenetically silences FLC in a Polycomb-mediated process called vernalization. Our data indicate that upregulation of long non-coding antisense transcripts covering the entire FLC locus may be part of the cold-sensing mechanism. Induction of these antisense transcripts occurs earlier than, and is independent of, other vernalization markers and coincides with a reduction in sense transcription. We show that addition of the FLC antisense promoter sequences to a reporter gene is sufficient to confer cold-induced silencing of the reporter. Our data indicate that cold-induced FLC antisense transcripts have an early role in the epigenetic silencing of FLC, acting to silence FLC transcription transiently. Recruitment of the Polycomb machinery then confers the epigenetic memory. Antisense transcription events originating from 3' ends of genes might be a general mechanism to regulate the corresponding sense transcription in a condition/stage-dependent manner.
DOI:10.1038/s41598-019-41428-2URLPMID:30899041 [本文引用: 1]
Long non-coding RNAs (lncRNAs) mediate important epigenetic regulation in various biological processes related to the stress response in plants. However, the systematic analysis of the lncRNAs expressed in Brassica rapa under heat stress has been elusive. In this study, we performed a genome-wide analysis of the lncRNA expression profiles in non-heading Chinese cabbage leaves using strand-specific RNA-sequencing. A total of 4594 putative lncRNAs were identified with a comprehensive landscape of dynamic lncRNA expression networks under heat stress. Co-expression networks of the interactions among the differentially expressed lncRNAs, mRNAs and microRNAs revealed that several phytohormones were associated with heat tolerance, including salicylic acid (SA) and brassinosteroid (BR) pathways. Of particular importance is the discovery of 25 lncRNAs that were highly co-expressed with 10 heat responsive genes. Thirty-nine lncRNAs were predicted as endogenous target mimics (eTMs) for 35 miRNAs, and five of them were validated to be involved in the heat tolerance of Chinese cabbage. Heat responsive lncRNA (TCONS_00048391) is an eTM for bra-miR164a, that could be a sponge for miRNA binding and may be a competing endogenous RNA (ceRNA) for the target gene NAC1 (Bra030820), affecting the expression of bra-miR164a in Chinese cabbage. Thus, these findings provide new insights into the functions of lncRNAs in heat tolerance and highlight a set of candidate lncRNAs for further studies in non-heading Chinese cabbage.
DOI:10.1093/pcp/pcy220URLPMID:30462304 [本文引用: 2]
Long non-coding RNAs (lncRNAs) are non-protein-coding transcripts longer than 200 nt that are distributed widely in organisms and play many physiological roles. The BoNR8 lncRNA is a 272 nt long transcript yielded by RNA polymerase III in cabbage that was identified as the closest homolog of the AtR8 lncRNA in Arabidopsis. The BoNR8 lncRNA was expressed extensively in the epidermal tissue in the root elongation zone of germinated seeds, and its accumulation was induced by abiotic stresses, auxins and ABA. To investigate the correlation between the BoNR8 lncRNA and germination, BoNR8-overexpressing Arabidopsis plants (BoNR8-AtOX) were prepared. Three independent BoNR8-AtOX lines showed less primary root elongation, incomplete silique development and decreased germination rates. The germination efficiencies were affected strongly by ABA and slightly by salt stress, and ABA-related gene expression was changed in the BoNR8-AtOX lines.
DOI:10.4161/rna.19101URL [本文引用: 1]
Recently, a large number of non-coding RNAs (ncRNAs) have been found in a wide variety of organisms, but their biological functions are poorly understood, except for several tiny RNAs. To identify novel ncRNAs with essential functions in flowering plants, we focused attention on RNA polymerase III (Pol III) and its transcriptional activity, because most Pol III-transcribed RNAs contribute to key processes relating to cell activities, and have highly conserved promoter elements: upstream sequence elements, a TATA-like sequence, and a poly(T) stretch as a transcription terminator. After in silico prediction from the Arabidopsis genome, 20 novel ncRNAs candidates were obtained. AtR8 RNA (approx. 260 nt) and AtR18 RNA (approx. 160 nt) were identified by efficient in vitro transcription by Pol III in tobacco nuclear extracts. AtR8 RNA was conserved among six additional taxa of Brassicaceae, and the secondary structure of the RNA was also conserved among the orthologs. Abundant accumulation of AtR8 RNA was observed in the plant roots and cytosol of cultured cells. The RNA was not processed into a smaller fragment and no short open reading frame was included. Remarkably, expression of the AtR8 RNA responded negatively to hypoxic stress, and this regulation evidently differed from that of U6 snRNA.
DOI:10.1186/s12870-019-1672-7URLPMID:30744565 [本文引用: 1]
BACKGROUND: Long non-coding RNAs (lncRNAs) are transcripts more than 200 bp in length do not encode proteins. Up to the present, it has been reported that lncRNAs play an essential role in developmental processes through their regulatory functions. However, their characteristics, expression inheritance patterns, and functions in Prunus mume are quite unidentified. RESULTS: In this present study, we exposed the specific characters of pistil development process between single pistil cv 'Qingjia No.2' (QJN2) and multiple pistils cv 'Da Yu' (DY). We found that early October is the key stage for pistil differentiation. The similarity epidermis was observed between two types of pistil. We also further investigated a complete pistil development lncRNA profiles through RNA-seq in Prunus mume. 2572 unique lncRNAs and 24,648 genes mapped to Prunus mume genome, furthermore, 591 novel lncRNAs were predicted. Both unique lncRNAs and novel lncRNAs are shorter in length than the mRNAs, and the overall expression level of lncRNAs was lower than mRNAs in Prunus mume. 186 known lncRNAs, 1638 genes and 89 novel lncRNAs were identified as significant differential expressed in QJN2 compared with DY. We predicted 421 target genes of differentially expressed known lncRNAs (DEKLs) and 254 target genes of differentially expressed novel lncRNAs (DENLs). 153 miRNAs were predicted interacted with 100 DEKLs while 112 miRNAs were predicted interacted with 55 DENLs. Further analysis of the DEKLs showed that the lncRNA of XR_514690.2 down-regulated its target ppe-miR172d, and up-regulated AP2, respectively. Meanwhile, the other lncRNA of TCONS_00032517 induced cytokinin negative regulator gene A-ARR expression via repressing its target miRNA ppe-miR160a/b in DY. At the same time we found that the AP2 expression was significantly up-regulated by zeatin (ZT) treatment in flower buds. Our experiments suggest that the two lncRNAs of XR_514690.2 and TCONS_00032517 might contribute the formation of multiple pistils in Prunus mume. CONCLUSION: This study shows the first characterization of lncRNAs involved in pistil development and provides new indications to elucidate how lncRNAs and their targets play role in pistil differentiation and flower development in Prunus mume.
DOI:10.1111/tpj.13953URLPMID:29738104 [本文引用: 1]
Long non-coding RNAs (lncRNAs) serve as versatile regulators of plant growth and development. The potential functions and inheritance patterns of lncRNAs, as well as the epigenetic regulation of lncRNA itself, remain largely uncharacterized in plant seeds, especially in the persistent endosperm of the dicotyledons. In this study, we investigated diverse RNA-seq data and catalogued 5356 lncRNAs in castor bean seeds. A small fraction of lncRNAs were transcribed from the same direction as the promoters of protein-coding genes (PCgenes) and exhibited strongly coordinated expression with the nearby PCgene. Co-expression analysis with weighted gene co-expression network analysis (WGCNA) showed these lncRNAs to be involved in differential transcription networks between the embryo and endosperm in the early developing seed. Genomic DNA methylation analyses revealed that the expression level of lncRNAs was tightly linked to DNA methylation and that endosperm hypomethylation could promote the expression of linked lncRNAs. Intriguingly, upon hybridization, most lncRNAs with divergent genome sequences between two parents could be reconciled and were expressed according to their parental genome contribution; however, some deviation in the expression of allelic lncRNAs was observed and found to be partially dependent on parental effects. In triploid endosperm, the expression of most lncRNAs was not dosage sensitive, as only 20 lncRNAs had balanced dosage. Our findings not only demonstrate that lncRNAs play potential roles in regulating the development of castor bean endosperm and embryo, but also provide novel insights into the parental effects, allelic expression and epigenetic regulation of lncRNAs in dicotyledonous seeds.
DOI:10.1371/journal.pgen.1007839URL [本文引用: 1]
DOI:10.3389/fpls.2018.01121URLPMID:30233602 [本文引用: 1]
NAC domain genes belong to a large plant-specific transcription factor family, which is well-known to be associated with multiple stress responses and plant developmental processes. In this study, we screened differentially expressed genes (DEGs) and detected mRNA abundance of NAC family by RNA-Seq in the poplar leaves under salt stress condition. A total of 276 up-regulated DEGs and 159 down-regulated DEGs were identified to be shared in Populus alba x Populus glandulosa and Populus simonii x Populus nigra. Among 170 NAC members, NAC57 gene was significantly up-regulated in response to salt stress in the two species. Tissue-specific and salt-responsive analyses indicated the expression pattern of NAC57 gene was spatial and temporal in poplar under salt stress. Particle bombardment results showed subcellular localization of NAC57 was not solely nucleus-targeted. Full-length cDNA sequence of the NAC57 gene was cloned from P. alba x P. glandulosa and transformed into Arabidopsis thaliana. Under salt stress, transgenic Arabidopsis overexpressing NAC57 showed higher seed germination rate, root length, and fresh weight than wild type plants. In addition, the transgenic plants displayed higher superoxide dismutase activity and peroxidase activity, and lower malondialdehyde content and relative electrical conductivity than the wild type under salt stress condition. Furthermore, histochemical staining indicated reactive oxygen species accumulation was lower in the transgenic plants than that in the wild type under salt stress. All the results indicated that the NAC57 gene plays an important role in salt stress responses.
DOI:10.1016/j.gene.2018.05.011URLPMID:29738839 [本文引用: 1]
MicroRNAs (miRNAs) and long noncoding RNAs (lncRNAs) act as important molecular regulators in a wide range of biological processes during plant development and seed formation, including oil production. Tree peony seeds contain >90% unsaturated fatty acids (UFAs) and high proportions of alpha-linolenic acid (ALA, > 40%). To dissect the non-coding RNAs (ncRNAs) pathway involved in fatty acids synthesis in tree peony seeds, we construct six small RNA libraries and six transcriptome libraries from developing seeds of two cultivars (J and S) containing different content of fatty acid compositions. After deep sequencing the RNA libraries, the ncRNA expression profiles of tree peony seeds in two cultivars were systematically and comparatively analyzed. A total of 318 known and 153 new miRNAs and 22,430 lncRNAs were identified, among which 106 conserved and 9 novel miRNAs and 2785 lncRNAs were differentially expressed between the two cultivars. In addition, potential target genes of the microRNA and lncRNAs were also predicted and annotated. Among them, 9 miRNAs and 39 lncRNAs were predicted to target lipid related genes. Results showed that all of miR414, miR156b, miR2673b, miR7826, novel-m0027-5p, TR24651|c0_g1, TR24544|c0_g15, and TR27305|c0_g1 were up-regulated and expressed at a higher level in high-ALA cultivar J when compared to low-ALA cultivar S, suggesting that these ncRNAs and target genes are possibly involved in different fatty acid synthesis and lipid metabolism through post-transcriptional regulation. These results provide a better understanding of the roles of ncRNAs during fatty acid biosynthesis and metabolism in tree peony seeds.
URLPMID:29873696 [本文引用: 1]
DOI:10.1186/s12870-019-2088-0URLPMID:31666019 [本文引用: 1]
BACKGROUND: Long non-coding (lnc) RNAs are a class of functional RNA molecules greater than 200 nucleotides in length, and lncRNAs play important roles in various biological regulatory processes and response to the biotic and abiotic stresses. LncRNAs associated with salt stress in cotton have been identified through RNA sequencing, but the function of lncRNAs has not been reported. We previously identified salt stress-related lncRNAs in cotton (Gossypium spp.), and discovered the salt-related lncRNA-lncRNA973. RESULTS: In this study, we identified the expression level, localization, function, and preliminary mechanism of action of lncRNA973. LncRNA973, which was localized in the nucleus, was expressed at a low level under nonstress conditions but can be significantly increased by salt treatments. Here lncRNA973 was transformed into Arabidopsis and overexpressed. Along with the increased expression compared with wild type under salt stress conditions in transgenic plants, the seed germination rate, fresh weights and root lengths of the transgenic plants increased. We also knocked down the expression of lncRNA973 using virus-induced gene silencing technology. The lncRNA973 knockdown plants wilted, and the leaves became yellowed and dropped under salt-stress conditions, indicating that the tolerance to salt stress had decreased compared with wild type. LncRNA973 may be involved in the regulation of reactive oxygen species-scavenging genes, transcription factors and genes involved in salt stress-related processes in response to cotton salt stress. CONCLUSIONS: LncRNA973 was localized in the nucleus and its expression was increased by salt treatment. The lncRNA973-overexpression lines had increased salt tolerance compared with the wild type, while the lncRNA973 knockdown plants had reduced salt tolerance. LncRNA973 regulated cotton responses to salt stress by modulating the expression of a series of salt stress-related genes. The data provides a basis for further studies on the mechanisms of lncRNA973-associated responses to salt stress in cotton.
DOI:10.1038/s41467-018-07500-7URLPMID:30498193 [本文引用: 1]
Long non-coding RNAs (lncRNAs) have emerged as important regulators of gene expression and plant development. Here, we identified 6,510 lncRNAs in Arabidopsis under normal or stress conditions. We found that the expression of natural antisense transcripts (NATs) that are transcribed in the opposite direction of protein-coding genes often positively correlates with and is required for the expression of their cognate sense genes. We further characterized MAS, a NAT-lncRNA produced from the MADS AFFECTING FLOWERING4 (MAF4) locus. MAS is induced by cold and indispensable for the activation of MAF4 transcription and suppression of precocious flowering. MAS activates MAF4 by interacting with WDR5a, one core component of the COMPASS-like complexes, and recruiting WDR5a to MAF4 to enhance histone 3 lysine 4 trimethylation (H3K4me3). Our study greatly extends the repertoire of lncRNAs in Arabidopsis and reveals a role for NAT-lncRNAs in regulating gene expression in vernalization response and likely in other biological processes.
[本文引用: 1]
植物耐盐生理及耐盐机理研究进展
1
2003
... 高盐环境会对植物造成离子毒害、渗透胁迫和矿质营养缺失, 使其生理代谢紊乱, 导致种子萌发和植株生长受到阻碍, 严重降低植物的产量和品质(
硫化氢对盐和铝胁迫下小麦种子萌发及氧化损伤的缓解效应
1
2010
... 高盐环境会对植物造成离子毒害、渗透胁迫和矿质营养缺失, 使其生理代谢紊乱, 导致种子萌发和植株生长受到阻碍, 严重降低植物的产量和品质(
盐胁迫对植物的影响及植物对盐胁迫的适应性
1
2015
... 高盐环境会对植物造成离子毒害、渗透胁迫和矿质营养缺失, 使其生理代谢紊乱, 导致种子萌发和植株生长受到阻碍, 严重降低植物的产量和品质(
盐胁迫对大豆种子萌发及生理的影响
1
2013
... 高盐环境会对植物造成离子毒害、渗透胁迫和矿质营养缺失, 使其生理代谢紊乱, 导致种子萌发和植株生长受到阻碍, 严重降低植物的产量和品质(
植物长链非编码RNA研究进展
5
2015
... 非编码RNA (non-coding RNA, ncRNAs)不编码蛋白质, 但却大量存在于生物体中发挥重要作用.长度大于200个核苷酸的非编码RNA为长链非编码RNA (long non-coding RNA, lncRNA), 它们通过充当诱饵、支架和增强子的方式调节许多基因的表达, 在剂量补偿(dosage compensation)、基因组印记(genomic imprinting)、X染色体失活(X chromosome inactivation)、发育和环境胁迫等生物过程中发挥重要作用(
ring 4)的反义转录lncRNA)通过与WDR5a相互作用激活MAF4的转录, 从而调节拟南芥的开花时间.
... ;
ring 4)的反义转录lncRNA)通过与WDR5a相互作用激活MAF4的转录, 从而调节拟南芥的开花时间.
... ;
ring 4)的反义转录lncRNA)通过与WDR5a相互作用激活MAF4的转录, 从而调节拟南芥的开花时间.
... ;
ring 4)的反义转录lncRNA)通过与WDR5a相互作用激活MAF4的转录, 从而调节拟南芥的开花时间.
... ;
ring 4)的反义转录lncRNA)通过与WDR5a相互作用激活MAF4的转录, 从而调节拟南芥的开花时间.
十字花科植物种子低分子RNA提取方法比较
1
2019
... RNA提取参照
盐胁迫对拟南芥种子萌发的影响
1
2012
... 高盐环境会对植物造成离子毒害、渗透胁迫和矿质营养缺失, 使其生理代谢紊乱, 导致种子萌发和植株生长受到阻碍, 严重降低植物的产量和品质(
植物盐胁迫生理及其适应性调控机制的研究进展
1
2007
...
盐分胁迫对植物的影响研究简述
2
2007
... 高盐环境会对植物造成离子毒害、渗透胁迫和矿质营养缺失, 使其生理代谢紊乱, 导致种子萌发和植株生长受到阻碍, 严重降低植物的产量和品质(
... 研究表明, 盐胁迫通过增效、负效和完全阻抑效应影响种子的萌发.低盐可增强种子的呼吸作用, 提高蛋白酶和脂肪酶活性, 促进贮藏物质的转化, 进而促进种子萌发和生长; 高盐条件下, 由于盐形成的渗透势阻碍种子吸水(盐浓度越高, 阻碍作用越强), 严重影响了种子内蛋白质等大分子物质的分解和合成进程, 降低种子的发芽率、发芽指数和活力指数(
植物耐盐机制的研究进展
1
2001
... 高盐环境会对植物造成离子毒害、渗透胁迫和矿质营养缺失, 使其生理代谢紊乱, 导致种子萌发和植株生长受到阻碍, 严重降低植物的产量和品质(
γ-氨基丁酸(GABA)调控盐胁迫下玉米种子萌发和幼苗生长的机制
1
2016
... 高盐环境会对植物造成离子毒害、渗透胁迫和矿质营养缺失, 使其生理代谢紊乱, 导致种子萌发和植株生长受到阻碍, 严重降低植物的产量和品质(
盐胁迫对拟南芥AtPUB18基因的诱导表达及其启动子分析
1
2014
... 高盐环境会对植物造成离子毒害、渗透胁迫和矿质营养缺失, 使其生理代谢紊乱, 导致种子萌发和植株生长受到阻碍, 严重降低植物的产量和品质(
Plant U-box armadillo repeat proteins AtPUB18 and AtPUB19 are involved in salt inhibition of germination in Arabidopsis
1
2011
... 高盐环境会对植物造成离子毒害、渗透胁迫和矿质营养缺失, 使其生理代谢紊乱, 导致种子萌发和植株生长受到阻碍, 严重降低植物的产量和品质(
Characterization of stress-responsive lncRNAs in Arabidopsis thaliana by integrating expression, epigenetic and structural features
1
2014
... UCC盐胁迫响应元件(TCTTCTTCTTTA)是盐应答基因中高度保守的序列.
A long noncoding RNA regulates photoperiod-sensitive male sterility, an essential component of hybrid rice
1
2012
... 非编码RNA (non-coding RNA, ncRNAs)不编码蛋白质, 但却大量存在于生物体中发挥重要作用.长度大于200个核苷酸的非编码RNA为长链非编码RNA (long non-coding RNA, lncRNA), 它们通过充当诱饵、支架和增强子的方式调节许多基因的表达, 在剂量补偿(dosage compensation)、基因组印记(genomic imprinting)、X染色体失活(X chromosome inactivation)、发育和环境胁迫等生物过程中发挥重要作用(
ring 4)的反义转录lncRNA)通过与WDR5a相互作用激活MAF4的转录, 从而调节拟南芥的开花时间.
WRKY41 controls Arabidopsis seed dormancy via direct regulation of ABI3 transcript levels not downstream of ABA
1
2014
... 种子是植物特有的繁殖器官.种子萌发是指有活力的种子吸胀后, 重新开始物质合成与代谢活动, 促使胚根露出种皮的过程.种子萌发极易受到光照、温度、水分和盐分等外部环境因素的影响, 同时受多基因调控.例如, 脱落酸(abscisic acid, ABA)胁迫下, WRKY41通过直接调节ABI3 (ABA-insensitive 3)的表达影响拟南芥(Arabidopsis thaliana)种子萌发(
Target mimicry provides a new me- chanism for regulation of microRNA activity
1
2007
... 非编码RNA (non-coding RNA, ncRNAs)不编码蛋白质, 但却大量存在于生物体中发挥重要作用.长度大于200个核苷酸的非编码RNA为长链非编码RNA (long non-coding RNA, lncRNA), 它们通过充当诱饵、支架和增强子的方式调节许多基因的表达, 在剂量补偿(dosage compensation)、基因组印记(genomic imprinting)、X染色体失活(X chromosome inactivation)、发育和环境胁迫等生物过程中发挥重要作用(
ring 4)的反义转录lncRNA)通过与WDR5a相互作用激活MAF4的转录, 从而调节拟南芥的开花时间.
Wheat miR9678 affects seed germination by generating phased siRNAs and modulating abscisic acid/gibberellin signaling
1
2018
... 近年来, 萌发相关ncRNA逐渐被发现并被解析, 如miR156、miR159、miR167和miR9678.拟南芥miR156基因下调SPL13 (SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE13)的表达, 延迟萌发后幼苗的发育(
Vernalization-mediated epigenetic silencing by a long intronic noncoding RNA
1
2011
... 非编码RNA (non-coding RNA, ncRNAs)不编码蛋白质, 但却大量存在于生物体中发挥重要作用.长度大于200个核苷酸的非编码RNA为长链非编码RNA (long non-coding RNA, lncRNA), 它们通过充当诱饵、支架和增强子的方式调节许多基因的表达, 在剂量补偿(dosage compensation)、基因组印记(genomic imprinting)、X染色体失活(X chromosome inactivation)、发育和环境胁迫等生物过程中发挥重要作用(
ring 4)的反义转录lncRNA)通过与WDR5a相互作用激活MAF4的转录, 从而调节拟南芥的开花时间.
Arabidopsis WRKY6 transcription factor acts as a positive regulator of abscisic acid signaling during seed germination and early seedling development
1
2016
... 种子是植物特有的繁殖器官.种子萌发是指有活力的种子吸胀后, 重新开始物质合成与代谢活动, 促使胚根露出种皮的过程.种子萌发极易受到光照、温度、水分和盐分等外部环境因素的影响, 同时受多基因调控.例如, 脱落酸(abscisic acid, ABA)胁迫下, WRKY41通过直接调节ABI3 (ABA-insensitive 3)的表达影响拟南芥(Arabidopsis thaliana)种子萌发(
Arabidopsis WRKY2 transcription factor mediates seed germination and postgermination arrest of development by abscisic acid
1
2009
... 种子是植物特有的繁殖器官.种子萌发是指有活力的种子吸胀后, 重新开始物质合成与代谢活动, 促使胚根露出种皮的过程.种子萌发极易受到光照、温度、水分和盐分等外部环境因素的影响, 同时受多基因调控.例如, 脱落酸(abscisic acid, ABA)胁迫下, WRKY41通过直接调节ABI3 (ABA-insensitive 3)的表达影响拟南芥(Arabidopsis thaliana)种子萌发(
Environmentally coordinated epigenetic silencing of FLC by protein and long noncoding RNA components
1
2012
... 非编码RNA (non-coding RNA, ncRNAs)不编码蛋白质, 但却大量存在于生物体中发挥重要作用.长度大于200个核苷酸的非编码RNA为长链非编码RNA (long non-coding RNA, lncRNA), 它们通过充当诱饵、支架和增强子的方式调节许多基因的表达, 在剂量补偿(dosage compensation)、基因组印记(genomic imprinting)、X染色体失活(X chromosome inactivation)、发育和环境胁迫等生物过程中发挥重要作用(
ring 4)的反义转录lncRNA)通过与WDR5a相互作用激活MAF4的转录, 从而调节拟南芥的开花时间.
Effect of AtR8 lncRNA partial deletion on Arabidopsis seed germination
3
2016
... 前期研究中, 我们根据RNA聚合酶III的转录活性及其转录的非编码RNA基因结构特征, 发现了拟南芥中RNA聚合酶III转录的AtR8 lncRNA (259 nt), 其在幼苗根端细胞质中大量表达, 并响应低氧胁迫(
... 实验材料为野生型拟南芥(Arabidopsis thaliana L.)和AtR8 lncRNA部分缺失型拟南芥突变体(atr8, FLAG410H04), 后者购自凡尔赛拟南芥储备中心(versailles Arabidopsis stock center (http://publiclines.versailles.inra.fr/)) (
...
The long noncoding RNA T5120 regulates nitrate response and assimilation in Arabidopsis
1
2019
... 非编码RNA (non-coding RNA, ncRNAs)不编码蛋白质, 但却大量存在于生物体中发挥重要作用.长度大于200个核苷酸的非编码RNA为长链非编码RNA (long non-coding RNA, lncRNA), 它们通过充当诱饵、支架和增强子的方式调节许多基因的表达, 在剂量补偿(dosage compensation)、基因组印记(genomic imprinting)、X染色体失活(X chromosome inactivation)、发育和环境胁迫等生物过程中发挥重要作用(
ring 4)的反义转录lncRNA)通过与WDR5a相互作用激活MAF4的转录, 从而调节拟南芥的开花时间.
Simple purification of small RNAs from seeds and efficient detection of multiple microRNAs expressed in Arabidopsis thaliana and tomato(Lycopersicon esculentum) seeds
1
2005
... RNA提取参照
The regulation of post-germinative transition from the cotyledon- to vegetative-leaf stages by microRNA-targeted SQUAMOSA PROMOTER-BINDING PROTEIN LIKE13 in Arabidopsis
1
2010
... 近年来, 萌发相关ncRNA逐渐被发现并被解析, 如miR156、miR159、miR167和miR9678.拟南芥miR156基因下调SPL13 (SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE13)的表达, 延迟萌发后幼苗的发育(
The microRNA156 and microRNA172 gene regulation cascades at post-germinative stages in Arabidopsis
1
2010
... 近年来, 萌发相关ncRNA逐渐被发现并被解析, 如miR156、miR159、miR167和miR9678.拟南芥miR156基因下调SPL13 (SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE13)的表达, 延迟萌发后幼苗的发育(
A nucleus-localized long non-coding RNA enhances drought and salt stress tolerance
1
2017
... 非编码RNA (non-coding RNA, ncRNAs)不编码蛋白质, 但却大量存在于生物体中发挥重要作用.长度大于200个核苷酸的非编码RNA为长链非编码RNA (long non-coding RNA, lncRNA), 它们通过充当诱饵、支架和增强子的方式调节许多基因的表达, 在剂量补偿(dosage compensation)、基因组印记(genomic imprinting)、X染色体失活(X chromosome inactivation)、发育和环境胁迫等生物过程中发挥重要作用(
ring 4)的反义转录lncRNA)通过与WDR5a相互作用激活MAF4的转录, 从而调节拟南芥的开花时间.
ABA induction of miR159 controls transcript levels of two MYB factors during Arabidopsis seed germination
1
2007
... 近年来, 萌发相关ncRNA逐渐被发现并被解析, 如miR156、miR159、miR167和miR9678.拟南芥miR156基因下调SPL13 (SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE13)的表达, 延迟萌发后幼苗的发育(
Prediction of plant microRNA targets
1
2002
... 近年来, 萌发相关ncRNA逐渐被发现并被解析, 如miR156、miR159、miR167和miR9678.拟南芥miR156基因下调SPL13 (SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE13)的表达, 延迟萌发后幼苗的发育(
CALMODULIN-BINDING TRANSCRIPTION ACTIVATOR 6: a key regulator of Na+ homeostasis during germination
1
2019
... 种子是植物特有的繁殖器官.种子萌发是指有活力的种子吸胀后, 重新开始物质合成与代谢活动, 促使胚根露出种皮的过程.种子萌发极易受到光照、温度、水分和盐分等外部环境因素的影响, 同时受多基因调控.例如, 脱落酸(abscisic acid, ABA)胁迫下, WRKY41通过直接调节ABI3 (ABA-insensitive 3)的表达影响拟南芥(Arabidopsis thaliana)种子萌发(
Cold-induced silencing by long antisense transcripts of an Arabidopsis polycomb target
1
2009
... 非编码RNA (non-coding RNA, ncRNAs)不编码蛋白质, 但却大量存在于生物体中发挥重要作用.长度大于200个核苷酸的非编码RNA为长链非编码RNA (long non-coding RNA, lncRNA), 它们通过充当诱饵、支架和增强子的方式调节许多基因的表达, 在剂量补偿(dosage compensation)、基因组印记(genomic imprinting)、X染色体失活(X chromosome inactivation)、发育和环境胁迫等生物过程中发挥重要作用(
ring 4)的反义转录lncRNA)通过与WDR5a相互作用激活MAF4的转录, 从而调节拟南芥的开花时间.
Genome-wide analysis of long non-coding RNAs unveils the regulatory roles in the heat tolerance of Chinese cabbage (Brassica rapa ssp. chinensis)
1
2019
... 非编码RNA (non-coding RNA, ncRNAs)不编码蛋白质, 但却大量存在于生物体中发挥重要作用.长度大于200个核苷酸的非编码RNA为长链非编码RNA (long non-coding RNA, lncRNA), 它们通过充当诱饵、支架和增强子的方式调节许多基因的表达, 在剂量补偿(dosage compensation)、基因组印记(genomic imprinting)、X染色体失活(X chromosome inactivation)、发育和环境胁迫等生物过程中发挥重要作用(
ring 4)的反义转录lncRNA)通过与WDR5a相互作用激活MAF4的转录, 从而调节拟南芥的开花时间.
Pol III-dependent cabbage BoNR8 long ncRNA affects seed germination and growth in Arabidopsis
2
2019
... 近年来, 萌发相关ncRNA逐渐被发现并被解析, 如miR156、miR159、miR167和miR9678.拟南芥miR156基因下调SPL13 (SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE13)的表达, 延迟萌发后幼苗的发育(
... 前期研究表明, RNA聚合酶III转录的AtR8 Inc- RNA同源物BoNR8 lncRNA大量存在于甘蓝萌发种子的根部(
A novel hypoxic stress-responsive long non-coding RNA transcribed by RNA polymerase III in Arabidopsis
1
2012
... 前期研究中, 我们根据RNA聚合酶III的转录活性及其转录的非编码RNA基因结构特征, 发现了拟南芥中RNA聚合酶III转录的AtR8 lncRNA (259 nt), 其在幼苗根端细胞质中大量表达, 并响应低氧胁迫(
Genome-wide discovery and characterization of flower development related long non-coding RNAs in Prunus mume
1
2019
... 非编码RNA (non-coding RNA, ncRNAs)不编码蛋白质, 但却大量存在于生物体中发挥重要作用.长度大于200个核苷酸的非编码RNA为长链非编码RNA (long non-coding RNA, lncRNA), 它们通过充当诱饵、支架和增强子的方式调节许多基因的表达, 在剂量补偿(dosage compensation)、基因组印记(genomic imprinting)、X染色体失活(X chromosome inactivation)、发育和环境胁迫等生物过程中发挥重要作用(
ring 4)的反义转录lncRNA)通过与WDR5a相互作用激活MAF4的转录, 从而调节拟南芥的开花时间.
Differential expression networks and inheritance patterns of long non-coding RNAs in castor bean seeds
1
2018
... 近年来, 萌发相关ncRNA逐渐被发现并被解析, 如miR156、miR159、miR167和miR9678.拟南芥miR156基因下调SPL13 (SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE13)的表达, 延迟萌发后幼苗的发育(
RSM1, an Arabidopsis MYB protein, interacts with HY5/ HYH to modulate seed germination and seedling development in response to abscisic acid and salinity
1
2018
... 种子是植物特有的繁殖器官.种子萌发是指有活力的种子吸胀后, 重新开始物质合成与代谢活动, 促使胚根露出种皮的过程.种子萌发极易受到光照、温度、水分和盐分等外部环境因素的影响, 同时受多基因调控.例如, 脱落酸(abscisic acid, ABA)胁迫下, WRKY41通过直接调节ABI3 (ABA-insensitive 3)的表达影响拟南芥(Arabidopsis thaliana)种子萌发(
Transcriptome analysis of poplar under salt stress and over-expression of transcription factor NAC57 gene confers salt tolerance in transgenic Arabidopsis
1
2018
... 高盐环境会对植物造成离子毒害、渗透胁迫和矿质营养缺失, 使其生理代谢紊乱, 导致种子萌发和植株生长受到阻碍, 严重降低植物的产量和品质(
Identification of microRNAs and long non-coding RNAs involved in fatty acid biosynthesis in tree peony seeds
1
2018
... 近年来, 萌发相关ncRNA逐渐被发现并被解析, 如miR156、miR159、miR167和miR9678.拟南芥miR156基因下调SPL13 (SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE13)的表达, 延迟萌发后幼苗的发育(
Transcriptomic and functional analyses unveil the role of long non-coding RNAs in anthocyanin biosynthesis during sea buckthorn fruit ripening
1
2018
... 非编码RNA (non-coding RNA, ncRNAs)不编码蛋白质, 但却大量存在于生物体中发挥重要作用.长度大于200个核苷酸的非编码RNA为长链非编码RNA (long non-coding RNA, lncRNA), 它们通过充当诱饵、支架和增强子的方式调节许多基因的表达, 在剂量补偿(dosage compensation)、基因组印记(genomic imprinting)、X染色体失活(X chromosome inactivation)、发育和环境胁迫等生物过程中发挥重要作用(
ring 4)的反义转录lncRNA)通过与WDR5a相互作用激活MAF4的转录, 从而调节拟南芥的开花时间.
The long non-coding RNA lncRNA 973 is involved in cotton response to salt stress
1
2019
... 非编码RNA (non-coding RNA, ncRNAs)不编码蛋白质, 但却大量存在于生物体中发挥重要作用.长度大于200个核苷酸的非编码RNA为长链非编码RNA (long non-coding RNA, lncRNA), 它们通过充当诱饵、支架和增强子的方式调节许多基因的表达, 在剂量补偿(dosage compensation)、基因组印记(genomic imprinting)、X染色体失活(X chromosome inactivation)、发育和环境胁迫等生物过程中发挥重要作用(
ring 4)的反义转录lncRNA)通过与WDR5a相互作用激活MAF4的转录, 从而调节拟南芥的开花时间.
Global identification of Arabidopsis lncRNAs reveals the regulation of MAF4 by a natural antisense RNA
1
2018
... 非编码RNA (non-coding RNA, ncRNAs)不编码蛋白质, 但却大量存在于生物体中发挥重要作用.长度大于200个核苷酸的非编码RNA为长链非编码RNA (long non-coding RNA, lncRNA), 它们通过充当诱饵、支架和增强子的方式调节许多基因的表达, 在剂量补偿(dosage compensation)、基因组印记(genomic imprinting)、X染色体失活(X chromosome inactivation)、发育和环境胁迫等生物过程中发挥重要作用(
ring 4)的反义转录lncRNA)通过与WDR5a相互作用激活MAF4的转录, 从而调节拟南芥的开花时间.
Transcriptomic analysis of long non-coding RNAs and coding genes uncovers a complex regulatory network that is involved in maize seed development
1
2017
... 近年来, 萌发相关ncRNA逐渐被发现并被解析, 如miR156、miR159、miR167和miR9678.拟南芥miR156基因下调SPL13 (SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE13)的表达, 延迟萌发后幼苗的发育(
备案号: 京ICP备16067583号-21
版权所有 © 2021 《植物学报》编辑部
地址:北京香山南辛村20号 邮编:100093
电话:010-62836135 010-62836131 E-mail:cbb@ibcas.ac.cn
本系统由北京玛格泰克科技发展有限公司设计开发

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}