 ,1,*, 尚琛晶,1,3,*
,1,*, 尚琛晶,1,3,*Expression of Amaranthin-like Lectins Gene and Responses to Abiotic Stresses in Cucumber
Fei Zhao1, Liuyi Dang2, Minhui Wei1, Chunying Liu1, Wei Leng,1,*, Chenjing Shang,1,3,*通讯作者: *E-mail:LWselt@126.com;cjshang@szu.edu.cn
收稿日期:2020-11-1接受日期:2021-03-1网络出版日期:2021-03-01
| 基金资助: |
Corresponding authors: *E-mail:LWselt@126.com;cjshang@szu.edu.cn
Received:2020-11-1Accepted:2021-03-1Online:2021-03-01

摘要
凝集素是一类具有特异性糖结合活性的蛋白质, 通常具有1个或多个非催化的糖结合结构域。凝集素在植物对病原菌的防御反应中发挥重要作用。由于其抗细菌、真菌、病毒和昆虫等的活性, 凝集素在农业和生物医药领域都具有很大的应用潜力。作为最小的凝集素家族之一, 苋科凝集素的研究较少。该文通过对重要经济作物黄瓜(Cucumis sativus)的基因组进行分析, 对16种苋科凝集素基因在黄瓜基因组中的分布和位置进行研究, 并分析相关基因的外显子/内含子组成。进一步通过启动子分析, 阐明了苋科凝集素基因对非生物胁迫的响应情况。最后, 通过实时荧光定量PCR, 检测了黄瓜中4种苋科凝集素基因对低温、高盐、干旱和ABA处理的响应情况。研究结果可为揭示苋科凝集素的生理功能及其在植物胁迫响应中的作用提供参考。
关键词:
Abstract
Lectins are globular proteins with carbohydrate-binding sites which enable them to specifically recognize and bind single or more particular carbohydrate structures. Previous studies have shown that lectins play critical roles in plant defense against their herbivores and pathogens. Due to their toxicity towards organisms like bacteria, fungi, viruses and insects, lectins are believed to have great potential in both agricultural and medical applications. As one of the smallest lectin families, there has been very limited research on amaranthin-like lectins. Here, we analyzed the distributions and intron/exon structures from 16 different amaranthin-like genes in the genome of cucumber, an important economic crop. To evaluate their responses against different stresses/plant hormones, promoter analysis was performed for all amaranthin-like genes. Finally, real-time quantitative PCR were performed on stress-treated plants to analyze the responses of amaranthin-like genes towards cold, salt, drought stresses and ABA treatment. This work provides valuable information for the study of physiological roles of amaranthin-like proteins and their involvement in plant defense.
Keywords:
PDF (910KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
赵菲, 党刘毅, 魏敏惠, 刘春莹, 冷伟, 尚琛晶. 黄瓜苋科凝集素基因的表达分析与逆境调控. 植物学报, 2021, 56(2): 183-190 doi:10.11983/CBB20177
Zhao Fei, Dang Liuyi, Wei Minhui, Liu Chunying, Leng Wei, Shang Chenjing.
凝集素是具有特异性糖结合活性的一类蛋白质, 具有1个或多个糖结合活性结构域, 能够特异性并且可逆地与糖链发生相互作用(Tsaneva and Van Damme, 2020)。虽然大部分已知的凝集素均为植物凝集素, 但凝集素也存在于动物、病毒、真菌和细菌中(Van Holle and Van Damme, 2019)。通过分析已经完成测序物种的基因组和转录组数据, 发现凝集素存在于已知的所有植物中。截至目前, 已有数百种植物凝集素被发现、纯化和表征。已有报道的凝集素具有多种糖结合活性。有的凝集素分子可以识别和结合单糖, 如甘露糖、葡萄糖、半乳糖和岩藻糖, 大部分凝集素与复杂糖链发生作用, 如N-连接糖链或O-连接糖链(Ghazarian et al.2011)。研究表明, 凝集素在农业(如植物抗逆与抗菌)和生物医学(如癌症治疗)等领域都具有重要的潜在应用价值(曾日中和黎瑜, 1998; 王志斌等, 2000)。
凝集素家族包含所有与糖链特异性相互作用而不改变糖链结构的蛋白质。近几十年来, 生物学家和化学家一直在努力建立适宜的植物凝集素分类系统。目前, 基于凝集素结构域的蛋白质序列相似性和进化关系, 可将凝集素分为12个家族, 包括双孢蘑菇类凝集素家族(ABA)、苋科类凝集素家族(Amaranthin)、几丁质酶相关凝集素家族(CRA)、蓝藻凝集素家族(Cyanovirin)、卫矛凝集素家族(Euonymus euro- paeus)、雪花莲凝集素家族(GNA)、橡胶蛋白家族(Hevein)、木菠萝凝集素家族(Jacalins)、豆科凝集素家族(Legume)、具有赖氨酸基序的凝集素家族(Lysin- related lectin)、烟草凝集素家族(Nictaba)和蓖麻毒素B凝集素家族(Ricin-B)。
黄瓜(Cucumis sativus)基因组测序已于2009年完成, 但其中的凝集素家族却一直报道较少(Huang et al.2009)。直到2016年, 我们对黄瓜基因组中不同凝集素家族的分布进行分析, 表明黄瓜中主要包含10种不同的凝集素家族, 并对其进化关系进行了研究(Dang and Van Damme, 2016), 探讨了其在应对非生物胁迫中的作用。研究表明, 虽然苋科凝集素在多种植物的基因组中都有发现, 但目前其生物学功能研究仍很缺乏。对于黄瓜而言, 低温、高盐和干旱均为其在自然生存环境中常见的非生物胁迫, 与其生长发育密切相关。因此, 我们对黄瓜中苋科凝集素基因的表达情况及其在逆境胁迫中的作用进行研究, 旨在为揭示苋科凝集素的生理功能及其在植物响应胁迫中的作用提供参考。
1 材料与方法
1.1 实验材料
本研究所用材料为黄瓜(Cucumis sativus L.)品种Vert Petit de Paris。首先, 将种子播种在湿润的滤纸上, 28°C黑暗中萌发。然后, 将萌发的种子转移到1/2 Hoagland’s营养液中, 在16小时光照(28°C)/8小时黑暗(22°C)条件下生长, 待幼苗生长至双叶期进行逆境处理。均设置实验组与对照组。低温处理: 将黄瓜幼苗置于光照培养箱中, 实验组培养温度设定为4°C, 对照组按原设定温度(28°C); 高盐、干旱和ABA处理: 将黄瓜幼苗分别浸泡在含有200 mmol?L-1 NaCl、100 mmol?L-1甘露醇或100 μmol?L -1 ABA的1/2Hoagland’s营养液溶液中, 对照组为1/2Hoag- land’s营养液。经1、3、6、12和24小时处理后, 根据处理方式的不同, 黄瓜植株逐渐呈现出受害相关表型(Wanget al.2015; Yan et al.2016)。于各时间点, 对不同处理组和对照组, 分别取4株植物的相同位置叶片合并为一个样品, 放入液氮中速冻, 然后保存于-80°C冰箱, 备用。所有实验均包含2次生物学重复。1.2 苋科凝集素的生物信息学分析
黄瓜中苋科凝集素基因的分布与表达分析所用数据库为CuGenDB (Cucumber (Chinese Long) genome v3,启动子分析: 提取所有苋科凝集素基因上游2 000 bp的启动子序列, 并使用植物DNA顺式作用调控元件数据库(PLACE)对典型的启动子元件进行分析(Higo et al.1999), 其中包括CAAT、TATA框和转录起始位点(TSS)。
1.3 RNA提取与cDNA合成
使用TRIzol试剂(Sigma-Aldrich)提取RNA, 并使用DNase I处理以去除DNA污染, 详细步骤参考试剂使用说明书。使用M-MLV反转录酶(Thermo Fisher)合成cDNA。使用nanodrop测量RNA浓度。1.4 实时荧光定量PCR (qRT-PCR)
采用qRT-PCR方法检测非生物胁迫处理前后黄瓜中4种苋科凝集素基因的表达水平, 以评估其对非生物胁迫的响应情况。qRT-PCR反应在CFX Connect实时定量PCR仪(Bio-Rad)上进行。反应体系总体积为20 μL, 其中包括10 μL SYBR Green supermix (Bio- Rad), 10 μmol?L -1正向与反向引物各1 μL, 1 μL 20 ng?μL -1 cDNA模板及7 μL超纯水。所用引物见 表1, 包含2对内参基因引物(CACS和PP2A)及4对目标基因引物。qRT-PCR反应程序: 95°C预变性10分钟; 96°C变性25秒, 58°C退火25秒, 72°C延伸20秒, 45次循环。qRT-PCR结果使用REST-384软件进行分析, 以评估其统计学显著性(Pfafflet al.2002)。所有样品均包含2次生物学重复, 每个生物学重复包括3次技术重复。Table 1
表1
表1实时荧光定量PCR引物列表
Table 1
| Primer name | Forward primer (5'-3') | Reverse primer (5'-3') |
|---|---|---|
| CACS | TGGGAAGATTCTTA- TGAAGTGC | CTCGTCAAATTT- ACACATTGGT |
| PP2A | CAACAGGTGATATT- GGATTATGAT | GCCAGCTCATCC- TCATATAAG |
| AAT4 | TTCGTACACGCAAC- GAGA | TGAAGAGGGTAA- GGCTTG |
| NAAT1 | GAGGAGCTGTGAAA- GGAGCA | CCCTCCACGACA- GTTCCAAT |
| AAT9 | CAGAAACAGCGAAC- CAGAGC | AACTTCATCCCCA- CCGAGTT |
| AAT14 | GGGAATAGAGACGA- TCCGAACT | GCGCAGAAGGCA- GTGTTT |
All primers for reference genes were chosen based on lite- rature (Migocka and Papierniak,
新窗口打开|下载CSV
2 结果与讨论
2.1 黄瓜中苋科凝集素基因的分布
通过对黄瓜基因组进行检索, 发现黄瓜中包含16种苋科凝集素基因, 均位于黄瓜第6号染色体上, 并成簇排列(表2) (Dang et al.2017)。其中, 8个基因的序列在染色体上反向排列, 分别为CucsaAAT1、CucsaAAT2、CucsaAAT3、CucsaAAT4、CucsaAAT- 11、CucsaAAT12、CucsaNAAT1和CucsaNAAT2。Table 2
表2
表2黄瓜中苋科凝集素基因及其在6号染色体上的位置
Table 2
| Gene | Start position | End position | Orientation | Gene | Start position | End position | Orientation |
|---|---|---|---|---|---|---|---|
| CucsaAAT1 | 5496646 | 5495195 | Reverse | CucsaAAT9 | 5654159 | 5656355 | Forward |
| CucsaAAT2 | 5583941 | 5582553 | Reverse | CucsaAAT10 | 5681664 | 5683484 | Forward |
| CucsaAAT3 | 5590531 | 5589140 | Reverse | CucsaAAT11 | 5691672 | 5690257 | Reverse |
| CucsaAAT4 | 5596447 | 5595167 | Reverse | CucsaAAT12 | 7077986 | 7075008 | Reverse |
| CucsaAAT5 | 5605793 | 5607184 | Forward | CucsaAAT13 | 7083801 | 7085207 | Forward |
| CucsaAAT6 | 5619299 | 5621364 | Forward | CucsaAAT14 | 7090825 | 7094963 | Forward |
| CucsaAAT7 | 5630016 | 5631894 | Forward | CucsaNAAT1 | 5648115 | 5646235 | Reverse |
| CucsaAAT8 | 5636899 | 5638943 | Forward | CucsaNAAT2 | 5670107 | 5668263 | Reverse |
新窗口打开|下载CSV
通过分析基因序列中外显子与内含子的分布, 我们发现有4个基因序列(CucsaAAT6、CucsaAAT7、CucsaAAT12和CucsaAAT14)包含1个内含子, 3个基因序列(CucsaAAT8、CucsaAAT9和CucsaAAT10)包含2个内含子(图1)。
图1
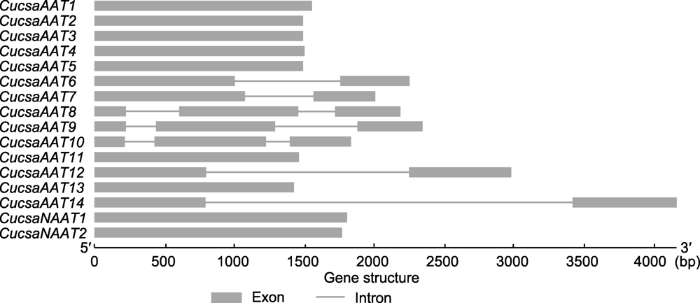 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1黄瓜中苋科凝集素基因的外显子/内含子组成及分布
Figure 1Exon/intron composition and distribution of amaranthin-like genes in cucumber
已有研究表明, 凝集素结构域常与具有不同生物学活性的其它结构域相结合, 从而使其功能更加多样化(王梦龙等, 2020)。对黄瓜中苋科凝集素的氨基酸序列进行分析, 发现所有苋科凝集素均包含2个苋科凝集素结构域(amaranthin domain)和1个气单胞菌溶素结构域(aerolysin domain), 因此将其命名为Cucsa AAT1-14; 另有2个基因在AAT结构域以外还包含1个N端功能未知的结构域, 被命名为Cucsa- NAAT1-2。所有苋科凝集素蛋白都不含有信号肽, 其理化性质见表3。苋科凝集素结构域和气单胞菌溶素结构域组成的嵌合蛋白在亚麻(Linum usitatissimum)中也被发现。研究表明, 具有该结构域的蛋白质编码基因在经过植物激素(如茉莉酸甲酯和水杨酸)诱导时发生高表达, 因此可能在植物的防御响应中起重要作用(Faruque et al.2015)。
Table 3
表3
表3黄瓜中苋科凝集素蛋白的理化性质
Table 3
| Protein | Domain architecture | No. of amino acids | Protein size (kDa) | Theoretical pI |
|---|---|---|---|---|
| CucsaAAT1 | AAT | 484 | 54.9 | 8.3 |
| CucsaAAT2 | AAT | 463 | 53.6 | 7.9 |
| CucsaAAT3 | AAT | 463 | 53.5 | 8.0 |
| CucsaAAT4 | AAT | 466 | 53.7 | 8.2 |
| CucsaAAT5 | AAT | 463 | 53.3 | 7.5 |
| CucsaAAT6 | AAT | 464 | 53.4 | 7.5 |
| CucsaAAT7 | AAT | 471 | 53.5 | 5.1 |
| CucsaAAT8 | AAT | 475 | 54.7 | 8.3 |
| CucsaAAT9 | AAT | 474 | 55.0 | 6.7 |
| CucsaAAT10 | AAT | 476 | 54.9 | 5.7 |
| CucsaAAT11 | AAT | 482 | 55.4 | 7.0 |
| CucsaAAT12 | AAT | 502 | 57.6 | 9.0 |
| CucsaAAT13 | AAT | 468 | 53.9 | 7.7 |
| CucsaAAT14 | AAT | 505 | 58.2 | 6.8 |
| CucsaNAAT1 | NAAT | 626 | 70.4 | 5.1 |
| CucsaNAAT2 | NAAT | 614 | 69.4 | 4.9 |
新窗口打开|下载CSV
根据这些序列的结构域及其外显子/内含子组成, 结合基因的进化关系(Dang and Van Damme, 2016), 我们选择4个基因进行后续分析(图2)。4个基因对应的蛋白结构域组成均包含AAT结构, 但其中CucsaAAT4无内含子; CucsaAAT14包含1个内含子而CucsaAAT9包含2个内含子; CucsaNAAT1同样无内含子, 但其对应蛋白中除AAT外另包含1个N端未知功能的结构域。
图2
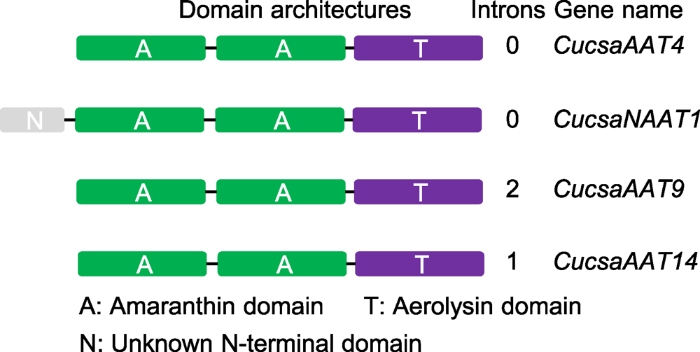 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2黄瓜中4种苋科凝集素蛋白的结构域构成
Figure 2Domain architectures of four amaranthin-like proteins in cucumber
2.2 黄瓜中苋科凝集素基因的启动子分析
作为基因的重要组成部分, 启动子的主要功能是控制基因表达(转录)的起始时间和表达水平。但启动子本身并不控制基因活动, 而是通过与转录因子结合进而控制基因活动(Al Atalah et al.2013)。对启动子进行分析, 可以获得影响基因表达的相关因子。我们选择苋科凝集素基因中起始密码子上游2 000 bp的序列用于启动子分析(promoter analysis) (图3)。结果表明, 在4种苋科凝集素基因的启动子序列中都鉴定到多种已知的非生物胁迫相关顺式作用元件。其中, 与光响应有关的元件(light-responsive elements)在所有苋科凝集素基因中都最为丰富。此外, 6种胁迫相关顺式作用元件的发生频率均比较接近, 包括与盐(salt)、干旱(drought)、脱落酸(ABA)、赤霉素(gibberellin)、病原体感染(pathogen)和低温(cold)胁迫相关的启动子元件。由此表明, 这4种苋科凝集素基因的表达很可能受到黄瓜生长发育中常见的胁迫, 如低温、高盐和干旱以及植物激素脱落酸的调控, 在植物对胁迫的响应中发挥重要作用。图3
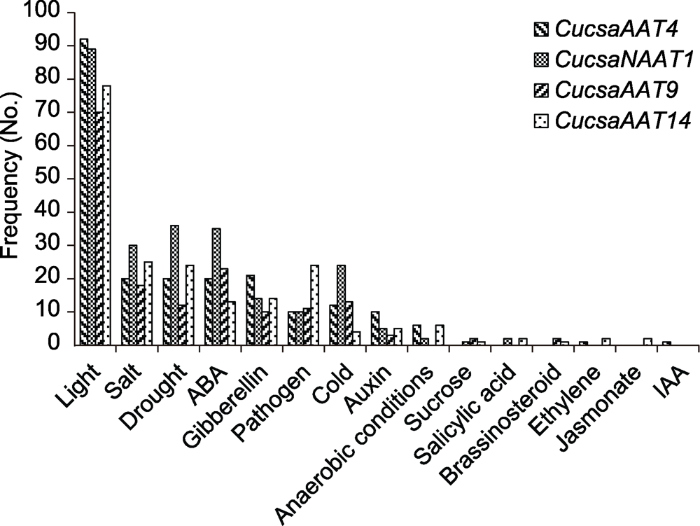 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3苋科凝集素基因中不同启动子应激调节因子的频率
Figure 3Frequencies of stress regulatory elements in the amaranthin-like genes promoter regions
2.3 苋科凝集素基因在非生物胁迫下的表达变化
为进一步验证启动子分析的结果, 我们利用实时荧光定量PCR对苋科凝集素基因在植物经不同胁迫处理后的表达水平变化进行了研究。对2周龄黄瓜幼苗进行低温、高盐、干旱胁迫和脱落酸处理, 在处理后的不同时间点分析4种黄瓜基因(CucsaAAT4、Cucsa- NAAT1、CucsaAAT9和CucsaAAT14)的转录水平, 以未处理的幼苗作为对照(图4)。所有数据均为处理组/对照组的相对表达水平。图4
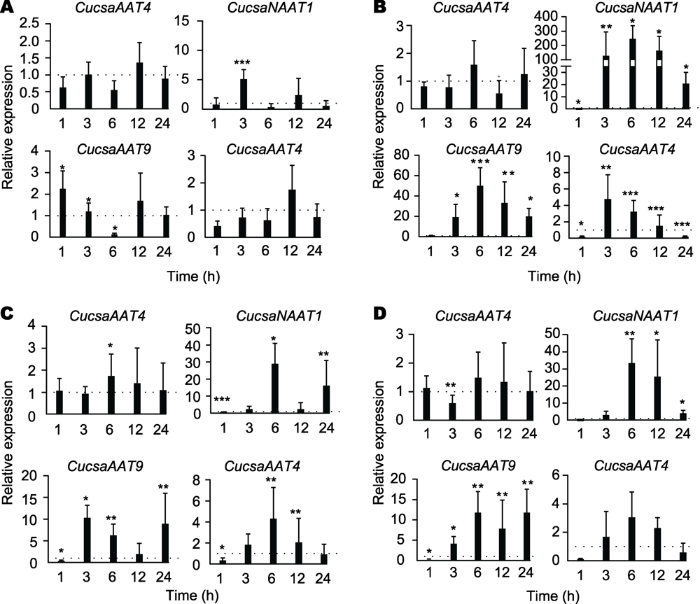 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同胁迫处理后黄瓜苋科凝集素基因的表达水平
(A)低温处理; (B)高盐处理;(C)干旱处理; (D)脱落酸处理。柱状图代表了2次生物学重复的平均值和标准偏差, 每个重复中包含实验组与对照组各4株植物; 星号表示处理组与对照组之间的统计学显著性(* P<0.05, ** P<0.01, ***P<0.001)。
Figure 4Relative expression levels for amaranthin-like genes in cucumber plants subjected to different abiotic stress conditions
(A)Cold treatment; (B) Salt treatment; (C) Drought treatment;(D) ABA treatment. Bars represent means and standard errors from two independent biological replicates, each replicate containing a pool of 4 plants for stress-treated groups as well as control groups. Asterisks indicate statistically signi?cant differences compared to the plants with mock treatments (* P<0.05,** P<0.01, ***P<0.001).
经低温处理后, 黄瓜幼苗在3小时后即呈现出萎蔫脱水的症状。叶片首先出现轻微受害症状, 叶缘发黄, 并随处理时间的增加叶片出现逐渐增大的脱水斑, 24小时后叶片出现大面积脱水斑, 冻害严重。转录水平上, 在低温处理3小时后可以检测到Cucsa- NAAT1和CucsaAAT9的表达水平上调, 而Cucsa- AAT4和CucsaAAT14的转录水平在24小时内未发生显著变化(图4A)。其中, CucsaAAT9的转录水平在处理1小时后即发生显著上调(2倍), 而CucsaNAAT1的转录水平在3小时后发生显著上调。
高盐处理会扰乱植物的渗透压平衡和离子平衡, 同时, 过量盐分会抑制和诱导多种酶系统的表达和活力, 进而影响植物的正常生长。经200 mmol?L-1 NaCl处理12小时后, 黄瓜幼苗根系侧根数目和长度相比对照组明显受到抑制, 并且随着时间的推移差别逐渐增大, 同时叶缘出现发黄现象。转录水平上, 除CucsaAAT4外, 另3种苋科凝集素基因的转录水平均发生明显上调(图4B)。尤其是CucsaNAAT1在高盐处理3小时后转录水平上调超过100倍, 然后在6小时达到最高水平, 表达上调超过200倍。CucsaAAT9的表达也发生了类似变化, 其表达水平在处理6小时后上调约50倍, 并且在处理后24小时仍保持约20倍上调表达。此外, 在高盐处理3小时后, CucsaAAT14的表达水平上调了4.7倍, 但之后逐渐恢复到正常水平。
我们通过在植物培养液中加入100 mmol?L-1甘露醇模拟干旱逆境。干旱处理后, 植物叶片表型改变最为显著, 6小时后黄瓜叶片出现轻度脱水, 并在12小时后叶片面积出现萎缩, 24小时后黄瓜幼苗叶片脱水严重并且发黄萎缩, 表现出显著的生长受抑制。黄瓜叶片经干旱处理6小时后, 所有黄瓜苋科凝集素基因均发生不同程度的上调表达(图4C)。其中, CucsaAAT4表达上调1.7倍, 而CucsaNAAT1表达上调近30倍。
当黄瓜幼苗经脱落酸处理后, 未观察到明显的表型变化, 而苋科凝集素基因表达水平的变化与干旱处理后非常类似(图4D), 这可能是由于ABA在植物对干旱胁迫的响应过程中发挥重要作用, 其在植物干旱胁迫时也会发生累积。
2.4 讨论
自从100多年前发现第一种植物凝集素以来, 科学家已经对大量植物凝集素进行了详细研究。目前, 苋科凝集素家族是最小的植物凝集素家族之一, 而几乎所有相关研究都集中在Amaranthus caudatus和一些相关物种中。随着各种植物基因组测序的完成, 苋科凝集素在不同物种中的研究成为可能。通过对已经完成测序的基因组数据进行分析, 发现该凝集素家族在不同植物家族中的分布显著不同(Dang et al.2017)。我们之前对84个已测序物种的基因组进行分析, 结果表明绝大部分苋科凝集素除含有1个或多个苋科凝集素结构域外都包含其它功能的结构域。其中, 气单胞菌溶素结构域是最常见的与苋科凝集素结构域共同存在的结构域。气单胞菌溶素是一类由微生物、植物或动物分泌的细胞毒素, 能够与真核细胞结合、聚集并在细胞膜上穿孔进而导致细胞裂解(Szczesny et al.2011)。因此, 气单胞菌溶素结构域的存在可能会导致细胞膜通透性提高。根据该类蛋白的结构域组成, 科学家推测其可能在植物对于特定病原菌的抗性中发挥重要作用(Shang et al.2017)。
本研究表明, 在植物应对非生物胁迫(低温、干旱和高盐)和脱落酸处理时, 苋科凝集素基因表达水平也会发生改变。作为一种广泛栽培的作物, 黄瓜对低温(Chen et al.2013)、干旱(Li et al.2020)和盐胁迫(Zhu et al.2019)均非常敏感。我们首先对苋科凝集素基因启动子进行分析, 结果表明其表达可能受到高盐、干旱和低温等的调控。进一步通过荧光定量PCR分析, 证实不同的苋科凝集素基因表达水平确实会受到高盐、干旱和低温等胁迫及脱落酸的调控, 这表明该家族蛋白在植物非生物胁迫响应中可能发挥重要作用。考虑到同一基因家族对非生物胁迫的不同反应, 我们推测不同的蛋白在植物中发挥的作用可能具有一定的特异性, 以应对不同的胁迫类型。此外, 作为一种重要的植物激素, 脱落酸主要作为调节植物水分平衡和渗透胁迫耐受性的内源信使, 采用脱落酸对植物进行处理, 也在一定程度上模拟了植物可能遭受的胁迫。由于多种非生物胁迫最终导致细胞干燥和渗透不平衡, 因此在低温、干旱和高盐处理后, 应激相关基因的表达与脱落酸处理后也存在一定程度的类似(Tuteja, 2007)。
目前, 对于苋科凝集素的研究报道较少, 本研究可为后续深入揭示苋科凝集素的生理功能及其在植物胁迫响应中的作用提供参考。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
In: Gopalakrishnakone P, Carlini RC, Ligabue-Braun R, eds.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
植物凝集素类受体蛋白激酶研究进展
1
2020
... 已有研究表明, 凝集素结构域常与具有不同生物学活性的其它结构域相结合, 从而使其功能更加多样化(王梦龙等,
在转基因植物中利用植物凝集素防治害虫的研究
1
2000
... 凝集素是具有特异性糖结合活性的一类蛋白质, 具有1个或多个糖结合活性结构域, 能够特异性并且可逆地与糖链发生相互作用(Tsaneva and Van Damme,
橡胶蛋白—一种与胶乳凝固有关的具有抗真菌活性的植物凝集素
1
1998
... 凝集素是具有特异性糖结合活性的一类蛋白质, 具有1个或多个糖结合活性结构域, 能够特异性并且可逆地与糖链发生相互作用(Tsaneva and Van Damme,
Promoter analysis for three types of EUL-related rice lectins in transgenic Arabidopsis
1
2013
... 作为基因的重要组成部分, 启动子的主要功能是控制基因表达(转录)的起始时间和表达水平.但启动子本身并不控制基因活动, 而是通过与转录因子结合进而控制基因活动(Al Atalah et al.
Arbuscular mycorrhizal fungi (AMF) increase growth and secondary metabolism in cucumber subjected to low temperature stress
1
2013
... 本研究表明, 在植物应对非生物胁迫(低温、干旱和高盐)和脱落酸处理时, 苋科凝集素基因表达水平也会发生改变.作为一种广泛栽培的作物, 黄瓜对低温(Chen et al.
Amaranthin-like proteins with aerolysin domains in plants
2
2017
... 通过对黄瓜基因组进行检索, 发现黄瓜中包含16种苋科凝集素基因, 均位于黄瓜第6号染色体上, 并成簇排列(
... 自从100多年前发现第一种植物凝集素以来, 科学家已经对大量植物凝集素进行了详细研究.目前, 苋科凝集素家族是最小的植物凝集素家族之一, 而几乎所有相关研究都集中在Amaranthus caudatus和一些相关物种中.随着各种植物基因组测序的完成, 苋科凝集素在不同物种中的研究成为可能.通过对已经完成测序的基因组数据进行分析, 发现该凝集素家族在不同植物家族中的分布显著不同(Dang et al.
Genome-wide identification and domain organization of lectin domains in cucumber
2
2016
... 黄瓜(Cucumis sativus)基因组测序已于2009年完成, 但其中的凝集素家族却一直报道较少(Huang et al.
... 根据这些序列的结构域及其外显子/内含子组成, 结合基因的进化关系(Dang and Van Damme,
The amaranthin-like lectin (LuALL) genes of flax: a unique gene family with members inducible by defence hormones
1
2015
... 已有研究表明, 凝集素结构域常与具有不同生物学活性的其它结构域相结合, 从而使其功能更加多样化(王梦龙等,
A glycobiology review: carbohydrates, lectins and implications in cancer therapeutics
1
2011
... 凝集素是具有特异性糖结合活性的一类蛋白质, 具有1个或多个糖结合活性结构域, 能够特异性并且可逆地与糖链发生相互作用(Tsaneva and Van Damme,
Plant cis-acting regulatory DNA elements (PLACE) database: 1999
1
1999
... 启动子分析: 提取所有苋科凝集素基因上游2 000 bp的启动子序列, 并使用植物DNA顺式作用调控元件数据库(PLACE)对典型的启动子元件进行分析(Higo et al.
The genome of the cucumber, Cucumis sativus L
2
2009
... 黄瓜(Cucumis sativus)基因组测序已于2009年完成, 但其中的凝集素家族却一直报道较少(Huang et al.
... 黄瓜中苋科凝集素基因的分布与表达分析所用数据库为CuGenDB (Cucumber (Chinese Long) genome v3,
CO2 enrichment enhanced drought resistance by regulating growth, hydraulic conductivity and phytohormone contents in the root of cucumber seedlings
1
2020
... 本研究表明, 在植物应对非生物胁迫(低温、干旱和高盐)和脱落酸处理时, 苋科凝集素基因表达水平也会发生改变.作为一种广泛栽培的作物, 黄瓜对低温(Chen et al.
Identification of suitable reference genes for studying gene expression in cucumber plants subjected to abiotic stress and growth regulators
2
2011
... 内参基因引物序列来自文献(Migocka and Papierniak,
... All primers for reference genes were chosen based on lite- rature (Migocka and Papierniak,
Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR
1
2002
... 采用qRT-PCR方法检测非生物胁迫处理前后黄瓜中4种苋科凝集素基因的表达水平, 以评估其对非生物胁迫的响应情况.qRT-PCR反应在CFX Connect实时定量PCR仪(Bio-Rad)上进行.反应体系总体积为20 μL, 其中包括10 μL SYBR Green supermix (Bio- Rad), 10 μmol?L -1正向与反向引物各1 μL, 1 μL 20 ng?μL -1 cDNA模板及7 μL超纯水.所用引物见
Plant AB toxins with lectin domains
1
2017
... 我们之前对84个已测序物种的基因组进行分析, 结果表明绝大部分苋科凝集素除含有1个或多个苋科凝集素结构域外都包含其它功能的结构域.其中, 气单胞菌溶素结构域是最常见的与苋科凝集素结构域共同存在的结构域.气单胞菌溶素是一类由微生物、植物或动物分泌的细胞毒素, 能够与真核细胞结合、聚集并在细胞膜上穿孔进而导致细胞裂解(Szczesny et al.
Extending the aerolysin family: from bacteria to vertebrates
1
2011
... 我们之前对84个已测序物种的基因组进行分析, 结果表明绝大部分苋科凝集素除含有1个或多个苋科凝集素结构域外都包含其它功能的结构域.其中, 气单胞菌溶素结构域是最常见的与苋科凝集素结构域共同存在的结构域.气单胞菌溶素是一类由微生物、植物或动物分泌的细胞毒素, 能够与真核细胞结合、聚集并在细胞膜上穿孔进而导致细胞裂解(Szczesny et al.
130 years of plant lectin research
1
2020
... 凝集素是具有特异性糖结合活性的一类蛋白质, 具有1个或多个糖结合活性结构域, 能够特异性并且可逆地与糖链发生相互作用(Tsaneva and Van Damme,
Abscisic acid and abiotic stress signaling
1
2007
... 本研究表明, 在植物应对非生物胁迫(低温、干旱和高盐)和脱落酸处理时, 苋科凝集素基因表达水平也会发生改变.作为一种广泛栽培的作物, 黄瓜对低温(Chen et al.
Messages from the past: new insights in plant lectin evolution
1
2019
... 凝集素是具有特异性糖结合活性的一类蛋白质, 具有1个或多个糖结合活性结构域, 能够特异性并且可逆地与糖链发生相互作用(Tsaneva and Van Damme,
Genome-wide identification of MAPK, MAPKK, and MAPKKK gene families and transcriptional profiling analysis during development and stress response in cucumber
1
2015
... 本研究所用材料为黄瓜(Cucumis sativus L.)品种Vert Petit de Paris.首先, 将种子播种在湿润的滤纸上, 28°C黑暗中萌发.然后, 将萌发的种子转移到1/2 Hoagland’s营养液中, 在16小时光照(28°C)/8小时黑暗(22°C)条件下生长, 待幼苗生长至双叶期进行逆境处理.均设置实验组与对照组.低温处理: 将黄瓜幼苗置于光照培养箱中, 实验组培养温度设定为4°C, 对照组按原设定温度(28°C); 高盐、干旱和ABA处理: 将黄瓜幼苗分别浸泡在含有200 mmol?L-1 NaCl、100 mmol?L-1甘露醇或100 μmol?L -1 ABA的1/2Hoagland’s营养液溶液中, 对照组为1/2Hoag- land’s营养液.经1、3、6、12和24小时处理后, 根据处理方式的不同, 黄瓜植株逐渐呈现出受害相关表型(Wanget al.
Different cucumber CsYUC genes regulate response to abiotic stresses and flower development
1
2016
... 本研究所用材料为黄瓜(Cucumis sativus L.)品种Vert Petit de Paris.首先, 将种子播种在湿润的滤纸上, 28°C黑暗中萌发.然后, 将萌发的种子转移到1/2 Hoagland’s营养液中, 在16小时光照(28°C)/8小时黑暗(22°C)条件下生长, 待幼苗生长至双叶期进行逆境处理.均设置实验组与对照组.低温处理: 将黄瓜幼苗置于光照培养箱中, 实验组培养温度设定为4°C, 对照组按原设定温度(28°C); 高盐、干旱和ABA处理: 将黄瓜幼苗分别浸泡在含有200 mmol?L-1 NaCl、100 mmol?L-1甘露醇或100 μmol?L -1 ABA的1/2Hoagland’s营养液溶液中, 对照组为1/2Hoag- land’s营养液.经1、3、6、12和24小时处理后, 根据处理方式的不同, 黄瓜植株逐渐呈现出受害相关表型(Wanget al.
Identification of cucumber circular RNAs responsive to salt stress
1
2019
... 本研究表明, 在植物应对非生物胁迫(低温、干旱和高盐)和脱落酸处理时, 苋科凝集素基因表达水平也会发生改变.作为一种广泛栽培的作物, 黄瓜对低温(Chen et al.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}