Peipei Liu, Geng Zhang, Xiaojuan Li,*Beijing Advanced Innovation Center for Tree Breeding by Molecular Design, College of Biological Science and Technology, Beijing Forestry University, Beijing 100083, China
Abstract As one of the plant cell wall polysaccharides, pectin has a very complex structure and function. Pectin is mainly composed by homogalacturonan (HG), rhamngalacturonan I (RGI), rhamngalacturonan II (RGII). Pectin plays an important role in maintaining the integrity of cell wall structure, intercellular adhesion and signal transduction. Therefore, studying the structure, distribution and roles of pectin components is of great significance for understanding the construction and function of cell wall. However, it is not clear how these three components of pectin cross-link to form high structure and perform biological function in the cell wall. This review will focus on the biosynthesis, functions of HG, RGI, RGII as well as the microscopic imaging techniques of pectin, aiming to provide a theoretical basis for the study of the structure and function of plant pectin. Keywords:biosynthesis;function of pectin;microscopic imaging;pectin components
PDF (1031KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 刘佩佩, 张耿, 李晓娟. 植物果胶的生物合成与功能. 植物学报, 2021, 56(2): 191-200 doi:10.11983/CBB20179 Liu Peipei, Zhang Geng, Li Xiaojuan. Biosynthesis and Function of Plant Pectin. Bulletin of Botany, 2021, 56(2): 191-200 doi:10.11983/CBB20179
细胞壁作为植物细胞的屏障, 在维持细胞膨压、完整性以及细胞间信号交流方面具有重要作用(Cosgrove, 2005)。细胞壁包括初生细胞壁和次生细胞壁, 均由多糖和少量蛋白组成(Logan et al.2014), 但组成成分和功能有所不同。其中初生细胞壁主要由纤维素、半纤维素和果胶组成(解敏敏等, 2015), 具有可塑性, 可以适应植物细胞生长过程中的动态变化, 对植物细胞的形态建成具有重要作用(Sakamoto et al.2018)。次生细胞壁主要由纤维素、半纤维素和木质素组成(Zhong and Ye, 2015), 刚性较强, 可赋予植物强度和硬度(Cosgrove and Jarvis, 2012; Li et al.2016; 张雨等, 2020)。初生细胞壁纤维素含量较少, 果胶多糖含量较多, 在双子叶和非禾本科植物中, 果胶约占初生细胞壁的35% (Cosgrove and Jarvis, 2012)。次生细胞壁的果胶含量尚不明确, 但有研究表明, 杨树(Populus trichocarpa, P. nigra × P. maximowiczii, P. trichocarpa×P. koreana)成熟木质部中有类果胶(pectin-like)组分在次生细胞壁中沉积(Arend et al.2008)。
作为植物细胞壁的重要组分, 果胶广泛存在于高等植物的根、茎和叶等器官的细胞壁和细胞间隙中, 在植物生长发育、防御和细胞壁完整性等方面具有重要作用。果胶多糖是自然界中结构最复杂的多糖家族, 组成成分主要包括同型半乳糖醛酸聚糖(homogalacturonan, HG)、鼠李半乳糖醛酸聚糖I (rhamngalacturonan I, RGI)和鼠李半乳糖醛酸聚糖II (rhamngalacturonan II, RGII), 不同的果胶结构域在细胞壁中彼此共价连接(Atmodjo et al.2013)。然而, 这3种组分的交联方式与其功能之间的关系尚不明确。因此, 研究果胶的结构与功能有利于更加深刻理解细胞壁高级结构的构建与功能。本文主要从果胶HG、RGI和RGII的生物合成、功能以及果胶的显微成像3个方面进行综述。
1 果胶多糖的生物合成
果胶组分在高尔基体中合成和修饰, 然后通过共价键相互连接形成果胶网络。果胶的合成过程需要多种酶类参与。已有研究表明, 果胶的合成至少需要67种不同的转移酶, 包括糖基转移酶、甲基转移酶和乙酰转移酶(Mohnen, 2008)。这些酶在顺式高尔基体、中间高尔基体和反式高尔基体中参与构建果胶多糖的复杂高级结构(Nebenfuhr and Staehelin, 2001)。
1.1 HG的生物合成
HG是α-1,4连接的半乳糖醛酸(α-1,4-linked D-galacturonic acid, GalA)线性同聚物, 在果胶中含量最高(Mohnen,2008)。HG可以在C-6羧基处进行甲基化, 参与甲基化修饰的酶包括HG半乳糖醛酸转移酶(galacturonosyltransferases, GAUT)和果胶甲基转移酶(pectin methyl-transferase, PMT) (Sebastian et al.2009)。在数据库CAZy中检索发现, GAUT蛋白属于糖基转移酶(glycosyltransferases, GT)家族8, 该家族在拟南芥(Arabidopsis thaliana)中有15个成员。其中参与果胶生物合成的酶有GAUT1 (Sterling et al.2006)、GAUT7 (Atmodjo et al.2011)、GAUT4 (Biswal et al.2018)和GAUT8 (QUA1) (Bouton et al.2002)。GAUT1是首个被鉴定的HG半乳糖醛酸转移酶, 它能在体外合成聚半乳糖醛酸(Sterling et al.2006); GAUT7是GAUT1的同源蛋白, 二者形成GAUT1:GAUT7复合物, 共同参与细胞壁基质多糖的生物合成(Atmodjo et al.2011); GAUT4可以合成HG, 其表达下调使细胞壁中的HG和RG-II减少(Biswal et al.2018); qua1突变体中HG含量极低, 表明QUA1可能参与果胶的生物合成(Bouton et al.2002)。PMT通过催化S-腺苷-L-甲硫氨酸的甲基转移到果胶HG中α-1,4连接的半乳糖醛酸残基的羧基上, 使HG发生甲基化修饰(Liuet al.2015)。PMT与GAUT可能作为异源复合体参与HG的聚合, 使HG完全甲基化(即甲基化程度达到80%)。此外, HG的一些GalA残基在O-2或O-3处可发生乙酰化修饰(易建勇等, 2020)。甲基化和乙酰化的酯化程度是动态可变的, 其修饰的程度影响果胶的理化性质。例如, 在Ca2+的作用下, 两条HG链之间相互作用形成二聚体。值得注意的是, 至少需要9个未甲基化修饰的GalA残基的连续延伸才能实现此同型二聚体(Scheller et al.2007)。
1.2 RGI的生物合成
RGI占果胶成分的20%-35%, 与HG和RGII不同, 它的主链由双糖重复链[4)-α-D-GalA-(1,2)-α-L-Rha- (1, ]n组成。RGI主链中的大部分或全部GalA在O-2或O-3处被乙酰化修饰, 大多数鼠李糖(rhamnose, Rha)残基可在O-4处被线性或支链寡糖或多糖取代(Atmodjo et al.2013)。RGI侧链包括β-1,4-半乳聚糖、阿拉伯聚糖或阿拉伯半乳聚糖等。RGI生物合成过程中, 其主链和侧链合成所需酶不尽相同, 主链合成需要半乳糖醛酸转移酶(galacturonosyltransferase, GalAT)和鼠李糖基转移酶(rhamnosyltransferase, RhaT)参与, 而侧链合成需要半乳糖基转移酶(galactosyltransferases, GalTs)和阿拉伯糖基转移酶(arabinosyltransferases, AraTs)。目前在拟南芥中已有报道的GalTs和AraTs编码基因包括GT92家族的GALS1 (GALACTAN SYNTHASE 1) (Liwanag et al.2012)和GT47家族B亚群中的ARAD1 (ARABINAN DEFICIENT 1) (Harholt et al.2006)及其同源基因ARAD2(Harholt et al.2012)。其中GALS1基因功能缺失和过表达分别导致细胞壁半乳聚糖的减少和增加(Liwanag et al.2012)。对arad1突变体的生化分析显示, 突变体细胞壁中的阿拉伯糖数量明显低于野生型(Harholt et al.2006); 而对arad2突变体细胞壁的生化分析显示, 其单糖组成与野生型无明显差异, 且过表达ARAD2并不能完全恢复arad1的表型, 说明虽然ARAD1与ARAD2同源, 但是二者功能并不冗余, 推测这2个糖基转移酶可能在二硫键连接的复合物形成中发挥作用(Harholt et al.2012)。
1.3 RGII的生物合成
RGII的结构非常复杂, 由至少12种不同的单糖构成, 其主干是HG, 主干上带有4个不同的侧链(A-D), 链A和B在拟南芥中分别被描述为高度分化的八糖和七糖, 链C和D是双糖(Glushka et al.2003; Pabst et al.2013)。值得注意的是, 侧链A和B的单糖组成中都含有芹菜糖残基(apiosyl, Api), 而侧链A的芹菜糖残基是形成硼酸二酯键的关键位点(Funakawa and Miwa, 2015)。目前, 对RGII生物合成的研究较少, 已确定参与RGII生物合成的酶有鼠李半乳糖醛酸木糖基转移酶1-4 (rhamno galacturonan xylosyltransferase, RGXT1-4), 它们属于GT77的B亚群, 具有α-1,3-木糖基转移酶活性, 能将UDP-Xyl转移到岩藻糖(fucose, Fuc)上。RGXT1和RGXT2在拟南芥幼苗和成熟植株的营养组织中均有表达, 二者突变体的生长表型与野生型相比均无明显变化(Egelund et al.2006); RGXT3仅在成熟植株的叶片和角质层中表达(Egelund et al.2008); RGXT4在许多器官和组织中都有表达, 其功能缺失会严重影响花粉管和根的生长, 导致植株不育和幼苗死亡(Liu et al.2011)。
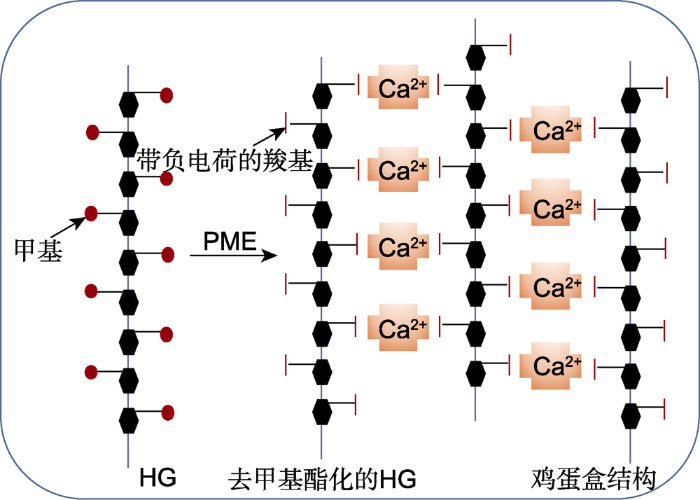
高甲基酯化的HG被果胶甲基酯酶(PME)去甲基酯化, 产生带负电荷的羧基。在Ca2+存在时, 2条HG链的羧基与Ca2+相互作用, 形成“鸡蛋盒”结构。 Figure 1An “egg-box” model of homogalacturonan (HG) polysaccharide and Ca 2+ cross-linking (modified from Micheli, 2001; Braccini and Peréz,2001)
Hypermethylated HG is de-methylated by pectin methylesterase (PME) to produce negatively charged carboxyl groups. In the presence of Ca2+, the carboxyl groups of the two HG chains interact with Ca2+ to form an “egg box” structure.
2.1.2 RGI侧链参与细胞重塑以及增强细胞壁硬度
RGI侧链包括β-1,4-半乳聚糖、阿拉伯聚糖或阿拉伯半乳聚糖, 其组成和结构影响细胞壁的强度、弹性和柔韧性等机械性能。例如, 侧链阿拉伯聚糖和阿拉伯半乳聚糖的长度和数量部分决定初生细胞壁的相对灵活性, 而半乳聚糖含量增加有助于增强细胞壁的硬度和刚度(Ulvskovet al.2005; Moore et al.2008)。Cankar等(2014)通过表型观察和组分分析, 对编码果胶修饰、主链降解和侧链类型等酶的多个转基因马铃薯(Solanum tuberosum)株系进行研究, 发现果胶主链被鼠李半乳糖醛酸裂解酶(rhamno galacturonanlyase, RGL)裂解为片段的转基因株系, 以及阿拉伯聚糖含量降低70%的转基因株系中, 成熟花粉粒的内壁在亲和孔的位置外翻突起, 最终导致花粉粒破裂和细胞质泄漏, 这些结果证实在花粉粒成熟和脱水过程中, 果胶阿拉伯聚糖侧链在细胞壁重塑过程中发挥了重要作用。McCartney等(2000)通过对(1→4)- β-D-半乳聚糖出现前后的豌豆(Pisum sativum cv. ‘Avola’)子叶进行机械压应力测试, 发现细胞壁富含半乳聚糖的子叶硬度明显增强。此外, RGI侧链的组成和结构与细胞和组织的发育阶段密切相关。例如, 在胡萝卜( Daucus carotacv. ‘Early Nantes’)由细胞分裂向细胞伸长过渡时, 伴随着细胞壁果胶半乳聚糖的出现和阿拉伯聚糖减少; 豌豆子叶在发育中随着果胶半乳聚糖的出现表现出组织硬度增强(Willats et al.1999; McCartney et al.2000)。
2.1.3 RGII二聚体有助于增强细胞壁强度
RGII结构虽然复杂, 但其结构在植物中高度保守(Matsunaga et al.2004)。在植物细胞壁中, 90%以上的RGII以二聚体形式存在。如图2所示, RGII二聚体由侧链A上的芹菜糖残基之间的硼酸二酯键共价交联形成(O’Neillet al.2004; Atmodjo et al.2013), 侧链A的完整结构对RGII二聚体的形成至关重要。当RGII完整性缺失时会影响植物的生长, 导致植物矮化(O’Neillet al.2001)和花粉管伸长缺陷(Delmas et al.2008), 严重时可导致植物细胞死亡(Ahn et al.2006)。在拟南芥中, MUR1 (MURUS1)编码GDP-甘露糖-4,6-脱水酶, 催化由GDP-D-mannose到GDP- L-focuse的第1步合成。GDP-L-focuse是L-岩藻糖(L- focuse, 6-deoxy-L-galactose)的活化核苷酸糖形式, L-岩藻糖是RGII侧链A的单糖组成之一, 也是其它多种结构多糖和糖蛋白的组成部分(Zhang et al.2019)。MUR1功能缺失突变体mur1植株矮化, RGII交联率降低(O’Neillet al.2001), 可能是由于侧链A截断所致(Pabst et al.2013)。进一步研究发现, mur1突变体侧链A中的L-focuse被L-Gal取代后, 全长侧链的比例降低, 导致RGII对硼的亲和力降低, 二聚体形成减少, 造成mur1植株地上部矮化(Reuhs et al.2004)。在植物细胞壁中, 硼将2条RGII链连接在一起形成二聚体, 增强了细胞之间的黏附和机械强度。研究发现, 通过补充硼可增加RGII的交联比例, 同时减弱mur1突变体的矮化表型, 表明硼酸盐引起的RGII交联对植物正常生长至关重要(O’Neillet al.2001)。
图2
新窗口打开|下载原图ZIP|生成PPT 图2拟南芥野生型和mur1突变体中的鼠李半乳糖醛酸聚糖II (RGII)二聚体(改自O’Neill et al.2004; Atmodjo et al.2013)
野生型植株中的鼠李半乳糖醛酸聚糖II二聚体由侧链A的芹菜糖残基通过硼酸二酯键共价交联而成。MUR1催化GDP-L-岩藻糖从头合成的第一步, MUR1功能缺失导致GDP-L-岩藻糖合成受阻。因此, mur1突变体中的鼠李半乳糖醛酸聚糖II侧链A被截断, 这一缺陷导致鼠李半乳糖醛酸聚糖II二聚体的形成减少。HG: 同型半乳糖醛酸聚糖; Api: 芹菜糖; Rha: L-鼠李糖; Fuc: L-岩藻糖; GlcA: D-葡萄糖醛酸; Gal: L-半乳糖; GalA: D-半乳糖醛酸; Me xyl: 2-O-甲基-D-木糖 Figure 2Rhamngalacturonan II (RGII) dimer in Arabidopsis wild-type and mur1 mutants (modified from O’Neill et al.2004; Atmodjo et al.2013)
In wild-type plants, RGII dimers are formed between Apiosyl residue of side chain A which are cross-linked covalently by the diester borate bonds. MUR1 catalyzes the first step in the de novo synthesis of GDP-L-fucose. The loss of MUR1 function leads to the block of GDP-L-fucose synthesis. Therefore, the RGII side chain A was truncated in mur1 mutants which resulted in reduced formation of RG-II dimers. HG: Homogalacturonan; Api: D-apiosyl; Rha: L-rhamnose; Fuc: L-fucose; GlcA: D-glucuronic acid; Gal: L-galactose; GalA: D-galacturonic acid; Me xyl: 2-O-methyl-D-xylose
2.2 果胶类多糖参与介导防御反应
细胞壁作为抵御病原体的屏障, 其完整性为植物防御病原体所必需(Hamann, 2015)。植物细胞壁果胶被细菌和真菌分泌的果胶降解酶降解, 其中HG降解后产生去甲基酯化的果胶片段或寡聚半乳糖醛酸(oligogalacturonides, OGs) (Ferrari et al.2013; 郁有健等, 2014)。OGs作为激发子可被植物体内的受体识别, 从而激活一系列防御反应, 起到保护植物的作用。研究表明, OGs的信号转导通过细胞壁关联激酶(wall-associated kinases, WAKs)发挥作用, WAKs能感知OGs, 进而激活植物的应激反应(Brutus et al.2010; Kohorn et al.2014)。此外, 病原体入侵会激活PME, PME催化HG去甲基酯化过程中产生具有挥发性的甲醇, 甲醇可作为信号分子参与启动邻近植物的免疫反应, 增强邻近植物对细菌病原体的抵抗力(Komarova et al.2014)。
果胶多糖原位成像的方法之一是使用糖抗体, 如单克隆抗体(JIM系列和LM系列等)可以高分辨率和高灵敏度地结合特定的多糖结构(Moller et al.2008; Pattathil et al.2010; Ralet et al.2010)。使用荧光素标记单克隆抗体, 当抗体与特定的果胶多糖结构结合后形成带有荧光素的复合体, 在荧光显微镜下使用合适的激发光, 即可观察多糖结构的定位与分布(Pattathil et al.2010)。
在果胶多糖的3种组分中, 对于HG的标记成像研究较多, 用于标记HG的单克隆抗体有JIM5、JIM7和2F4等。其中, JIM5的最佳识别表位可能是4个以上相邻的未酯化GalA残基, 这些残基与甲基酯化的GalA残基相邻或在其两侧(Clausen et al.2003); JIM7识别表位的重要特征是每隔一段残基上都有甲基酯化的GalA残基(Clausen et al.2003); 2F4可结合钙离子交联的未酯化的GalA残基(Sujkowska-Rybkowska and Borucki, 2015)。我们用JIM5和JIM7对青扦(Picea wilsonii)花粉管细胞壁中的果胶进行标记, 在激光共聚焦显微镜下用488 nm激发并在500-550 nm处采集发射信号, 可观察到在花粉管细胞壁中, JIM7标记的高甲基酯化的果胶聚集在生长的花粉管顶端, JIM5标记的低甲基酯化的果胶分布在花粉管伸长处(Cui et al.2015)。此外, 我们利用JIM5和JIM7免疫标记, 还检测了杉木(Cunninghamia lanceolata)维管形成层细胞壁多糖在季节周期中的变化, 结果显示, 休眠期形成层细胞壁中富含低甲基酯化的HG, 而活跃期形成层细胞壁中高甲基酯化的HG较多(Wu et al.2016)。Sujkowska-Rybkowska和Borucki (2015)使用JIM5、JIM7和2F4分别对低甲基酯化果胶、高甲基酯化果胶和钙交联果胶进行免疫标记, 检测铝处理的豌豆根瘤中果胶酯化和分布模式。Mravec等(2014)利用低聚壳聚糖(chitosan oligosaccharides, COS)与去酯化的HG果胶可以特异性结合的特点, 开发了非免疫标记分子探针。应用Alexa Fluor 488偶联COS (COS488)标记去甲基酯化HG, 可以对其分布进行高分辨率荧光成像观察; COS与纳米金粒子耦合(COSAuNP)后, 可以用于在透射电子显微镜下对HG定位进行超微结构水平的观察。与抗体免疫标记相比较, COS488具有穿透速度快和对酯化程度的变化敏感等特点, 可用于活细胞实时成像。
对RGI的标记研究表明, 其主链和侧链的抗体标记有所不同, 其中中性侧链可被LM5半乳糖和LM6/ LM13阿拉伯糖抗体探针标记(Wu et al.2016), 主链可被INRA-RU1和INRA-RU2标记(Ralet et al.2010)。标记主链的INRA-RU2需要至少2个双糖重复序列才能被结合, INRA-RU1需要至少6个双糖重复才能被结合, 两个抗体的最佳表位均为7个双糖重复序列(Ralet et al.2010)。例如, Lee等(2013)使用LM5、LM6和INRA-RU1对烟草(Nicotiana tabacum)种子胚乳细胞壁进行原位分析和色谱检测分析, 研究RGI结构在单个组织中的相对发生情况, 证明RGI聚合物在组织水平和单细胞壁水平上存在空间异质性。
BoschM, CheungAY, HeplerPK (2005). Pectin methylesterase, a regulator of pollen tube growth Plant Physiol138,1334-1346. DOI:10.1104/pp.105.059865URL [本文引用: 1]
BoutonS, LeboeufE, MouilleG, LeydeckerMT, TalbotecJ, GranierF, LahayeM, H?fteH, TruongHN (2002). QUASIMODO1 encodes a putative membrane-bound glycosyltransferase required for normal pectin synthesis and cell adhesion in Arabidopsis Plant Cell14, 2577-2590. DOI:10.1105/tpc.004259URL [本文引用: 2]
BracciniI, PerézS (2001). Molecular basis of Ca 2+-induced gelation in alginates and pectins: the egg-box model revisited Biomacromolecules2,1089-1096. DOI:10.1021/bm010008gURL [本文引用: 3]
BrutusA, SiciliaF, MaconeA, CervoneF, DeLorenzo G (2010). A domain swap approach reveals a role of the plant wall-associated kinase 1 (WAK1) as a receptor of oligogalacturonides Proc Natl Acad Sci USA107,9452- 9457. DOI:10.1073/pnas.1000675107URL [本文引用: 1]
CaffallKH, MohnenD (2009). The structure, function, and biosynthesis of plant cell wall pectic polysaccharides Carbohydr Res344,1879-1900. DOI:10.1016/j.carres.2009.05.021URL [本文引用: 1]
CankarK, KortsteeA, ToonenMAJ, Wolters-ArtsM, HoubeinR, MarianiC, UlvskovP, JorgensenB, ScholsHA, VisserRGF, TrindadeLM (2014). Pectic arabinan side chains are essential for pollen cell wall integrity during pollen development Plant Biotechnol J12,492-502. DOI:10.1111/pbi.12156URL [本文引用: 1] Pectin is a complex polysaccharide and an integral part of the primary plant cell wall and middle lamella, contributing to cell wall mechanical strength and cell adhesion. To understand the structure-function relationships of pectin in the cell wall, a set of transgenic potato lines with altered pectin composition was analysed. The expression of genes encoding enzymes involved in pectin acetylation, degradation of the rhamnogalacturonan backbone and type and length of neutral side chains, arabinan and galactan in particular, has been altered. Upon crossing of different transgenic lines, some transgenes were not transmitted to the next generation when these lines were used as a pollen donor, suggesting male sterility. Viability of mature pollen was severely decreased in potato lines with reduced pectic arabinan, but not in lines with altered galactan side chains. Anthers and pollen of different developmental stages were microscopically examined to study the phenotype in more detail. Scanning electron microscopy of flowers showed collapsed pollen grains in mature anthers and in earlier stages cytoplasmic protrusions at the site of the of kin pore, eventually leading to bursting of the pollen grain and leaking of the cytoplasm. This phenomenon is only observed after the microspores are released and the tapetum starts to degenerate. Timing of the phenotype indicates a role for pectic arabinan side chains during remodelling of the cell wall when the pollen grain is maturing and dehydrating.
CosgroveDJ (2005). Growth of the plant cell wall Nat Rev Mol Cell Biol6,850-861. [本文引用: 1]
CosgroveDJ, JarvisMC (2012). Comparative structure and biomechanics of plant primary and secondary cell walls Front Plant Sci3,204. [本文引用: 2]
CuiYN, LingY, ZhouJH, LiXJ (2015). Interference of the histone deacetylase inhibits pollen germination and pollen tube growth in Picea wilsonii mast PLoS One10,e0145661. [本文引用: 1]
DelmasF, SévenoM, NortheyJGB, HernouldM, LerougeP, McCourtP, ChevalierC (2008). The synthesis of the rhamnogalacturonan II component 3-deoxy-D- manno-2- octulosonic acid (Kdo) is required for pollen tube growth and elongation J Exp Bot59,2639-2647. [本文引用: 1]
DumontM, LehnerA, VauzeillesB, MalassisJ, MarchantA, SmythK, LinclauB, BaronA, PonsJM, AndersonCT, SchapmanD, GalasL, MolletJC, LerougeP (2016). Plant cell wall imaging by metabolic click-mediated labelling of rhamnogalacturonan II using azido 3-deoxy-D- manno-oct-2-ulosonic acid Plant J85,437- 447. [本文引用: 2]
EgelundJ, DamagerI, FaberK, OlsenCE, UlvskovP, PetersenBL (2008). Functional characterisation of a putative rhamnogalacturonan II specific xylosyltransferase FEBS Lett582,3217-3222. [本文引用: 1]
EgelundJ, PetersenBL, MotawiaMS, DamagerI, FaikA, OlsenCE, IshiiT, ClausenH, UlvskovP, GeshiN (2006). Arabidopsis thaliana RGXT1 and RGXT2 encode golgi-localized (1,3)-α-D-xylosyltransferases involved in the synthesis of pectic rhamnogalacturonan-II Plant Cell18,2593-2607. [本文引用: 1]
FerrariS, SavatinDV, SiciliaF, GramegnaG, CervoneF, DeLorenzo G (2013). Oligogalacturonides: plant damage-associated molecular patterns and regulators of growth and development Front Plant Sci4,49. [本文引用: 1]
FunakawaH, MiwaK (2015). Synthesis of borate cross- linked rhamnogalacturonan II Front Plant Sci6,223. [本文引用: 1]
GlushkaJN, TerrellM, YorkWS, O'NeillMA, GucwaA, DarvillAG, AlbersheimP, PrestegardJH (2003). Primary structure of the 2- O-methyl-α-L-fucose-containing side chain of the pectic polysaccharide, rhamnogalacturonan II Carbohydr Res338,341-352. [本文引用: 1]
HamannT (2015). The plant cell wall integrity maintenance mechanism—a case study of a cell wall plasma membrane signaling network Phytochemistry112,100-109. [本文引用: 1]
HarholtJ, JensenJK, S?rensenSO, OrfilaC, PaulyM, SchellerHV (2006). ARABINAN DEFICIENT 1 is a putative arabinosyltransferase involved in biosynthesis of pectic arabinan in Arabidopsis Plant Physiol140,49-58. [本文引用: 2]
HarholtJ, JensenJK, VerhertbruggenY, S?gaardC, BernardS, NafisiM, PoulsenCP, GeshiN, SakuragiY, DriouichA, KnoxJP, SchellerHV (2012). ARAD proteins associated with pectic arabinan biosynthesis form complexes when transiently overexpressed in planta Planta236,115-128. [本文引用: 2]
Jiménez-MaldonadoM, Tiznado-HernándezM, Rascón-Chu A, Carvajal-MillánE, Lizardi-MendozaJ, Troncoso-RojasR (2018). Analysis of rhamnogalacturonan I fragments as elicitors of the defense mechanism in tomato fruit Chil J Agric Res78,339-349. [本文引用: 1]
KohornBD, KohornSL, SabaNJ, MartinezVM (2014). Requirement for pectin methyl esterase and preference for fragmented over native pectins for wall-associated kinase-activated, eds1/pad4-dependent stress response in Arabidopsis J Biol Chem289,18978-18986. [本文引用: 1]
KomarovaTV, SheshukovaEV, DorokhovYL (2014). Cell wall methanol as a signal in plant immunity Front Plant Sci5,101. [本文引用: 1]
LeeKJD, CornuaultV, ManfieldIW, RaletMC, KnoxJP (2013). Multi-scale spatial heterogeneity of pectic rhamnogalacturonan I (RG-I) structural features in tobacco seed endosperm cell walls Plant J75,1018-1027. [本文引用: 1]
LiZ, FernieAR, PerssonS (2016). Transition of primary to secondary cell wall synthesis Sci Bull61,838-846. [本文引用: 1]
LiuXL, LiuLF, NiuQK, XiaCA, YangKZ, LiR, ChenLQ, ZhangXQ, ZhouYH, YeD (2011). MALE GAMETOPHYTE DEFECTIVE 4 encodes a rhamnogalacturonan II xylosyltransferase and is important for growth of pollen tubes and roots in Arabidopsis Plant J65,647-660. [本文引用: 1]
LiwanagAJM, EbertB, VerhertbruggenY, RennieEA, RautengartenC, OikawaA, AndersenMCF, ClausenMH, SchellerHV (2012). Pectin biosynthesis: GALS1 in Arabidopsis thaliana is a β-1,4-Galactan β-1,4-Galactosyltransferase Plant Cell24,5024-5036. [本文引用: 2]
LoganB, LeiL, LiSD, GuY (2014). Cell wall, cytoskeleton, and cell expansion in higher plants Mol Plant7,586-600. [本文引用: 1]
MatsunagaT, IshiiT, MatsumotoS, HiguchiM, DarvillA, AlbersheimP, O'NeillMA (2004). Occurrence of the primary cell wall polysaccharide rhamnogalacturonan II in pteridophytes, lycophytes, and bryophytes. Implications for the evolution of vascular plants Plant Physiol134,339- 351. [本文引用: 1]
McCartneyL, OrmerodAP, GidleyMJ, KnoxJP (2000). Temporal and spatial regulation of pectic (1→4)-β-D-galactan in cell walls of developing pea cotyledons: implications for mechanical properties Plant J22,105-113. [本文引用: 2]
MicheliF (2001). Pectin methylesterases: cell wall enzymes with important roles in plant physiology Trends Plant Sci6,414-419. [本文引用: 3]
MollerI, MarcusSE, HaegerA, VerhertbruggenY, VerhoefR, ScholsH, UlvskovP, MikkelsenJD, KnoxJP, WillatsW (2008). High-throughput screening of monoclonal antibodies against plant cell wall glycans by hierarchical clustering of their carbohydrate microarray binding profiles Glycoconj J25,37-48. [本文引用: 1]
MooreJP, FarrantJM, DriouichA (2008). A role for pectin-associated arabinans in maintaining the flexibility of the plant cell wall during water deficit stress Plant Signal Behav3,102-104. [本文引用: 1]
MravecJ, Kra?unSK, RydahlMG, WesterengB, MiartF, ClausenMH, FangelJU, DaugaardM, VanCutsem P, De Fine LichtHH, HofteH, MalinovskyFG, DomozychDS, WillatsWGT (2014). Tracking developmentally regulated post-synthetic processing of homogalacturonan and chitin using reciprocal oligosaccharide probes Development141,4841-4850. [本文引用: 1]
NebenfuhrA, StaehelinLA (2001). Mobile factories: golgi dynamics in plant cells Trends Plant Sci6,160-167. [本文引用: 1]
O’NeillMA, EberhardS, AlbersheimP, DarvillAG (2001). Requirement of borate cross-linking of cell wall rhamnogalacturonan II for Arabidopsis growth Science294,846- 849. [本文引用: 3]
O’NeillMA, IshiiT, AlbersheimP, DarvillAG (2004). RHAMNOGALACTURONAN II: structure and function of a borate cross-linked cell wall pectic polysaccharide Annu Rev Plant Biol55,109-139. [本文引用: 3]
PabstM, FischlRM, BreckerL, MorelleW, FaulandA, K?felerH, AltmannF, LéonardR (2013). Rhamnogalacturonan II structure shows variation in the side chains monosaccharide composition and methylation status within and across different plant species Plant J76,61-72. [本文引用: 2]
PanterPE, KentO, DaleM, SmithSJ, SkipseyM, ThorlbyG, CumminsI, RamsayN, BegumRA, SanhuezaD, FrySC, KnightMR, KnightH (2019). MUR1-mediated cell- wall fucosylation is required for freezing tolerance in Arabidopsis thaliana New Phytol224, 1518-1531. [本文引用: 1]
Sujkowska-RybkowskaM, BoruckiW (2015). Pectins esterification in the apoplast of aluminum-treated pea root nodules J Plant Physiol184,1-7. [本文引用: 2]
Torn?eCW, ChristensenC, MeldalM (2002). Peptidotriazoles on solid phase: [1,2,3]-triazoles by regiospecific copper(I)-catalyzed 1,3-dipolar cycloadditions of terminal alkynes to azides J Org Chem67,3057-3064. [本文引用: 1]
UlvskovP, WiumH, BruceD, J?rgensenB, QvistKB, Skj?tM, HepworthD, BorkhardtB, S?rensenSO (2005). Biophysical consequences of remodeling the neutral side chains of rhamnogalacturonan I in tubers of transgenic potatoes Planta220,609-620. [本文引用: 1]
WillatsWGT, Steele-KingCG, MarcusSE, KnoxJP (1999). Side chains of pectic polysaccharides are regulated in relation to cell proliferation and cell differentiation Plant J20,619-628. [本文引用: 1]
WormitA, UsadelB (2018). The multifaceted role of pectin methylesterase inhibitors (PMEIs) Int J Mol Sci19,2878. [本文引用: 1]
WuHY, XuHM, LiHY, WeiDM, LinJX, LiXJ (2016). Seasonal development of cambial activity in relation to xylem formation in Chinese fir J Plant Physiol195,23-30. [本文引用: 2]
ZhangL, PaaschBC, ChenJ, DayB, HeSY (2019). An important role of L-fucose biosynthesis and protein fucosylation genes in Arabidopsis immunity New Phytol222,981-994. [本文引用: 1]
ZhongRQ, YeZH (2015). Secondary cell walls: biosynthesis, patterned deposition and transcriptional regulation Plant Cell Physiol56,195-214. [本文引用: 1]
参与木葡聚糖合成的糖基转移酶基因研究进展 1 2015
... 细胞壁作为植物细胞的屏障, 在维持细胞膨压、完整性以及细胞间信号交流方面具有重要作用(Cosgrove, 2005).细胞壁包括初生细胞壁和次生细胞壁, 均由多糖和少量蛋白组成(Logan et al.2014), 但组成成分和功能有所不同.其中初生细胞壁主要由纤维素、半纤维素和果胶组成(解敏敏等, 2015), 具有可塑性, 可以适应植物细胞生长过程中的动态变化, 对植物细胞的形态建成具有重要作用(Sakamoto et al.2018).次生细胞壁主要由纤维素、半纤维素和木质素组成(Zhong and Ye, 2015), 刚性较强, 可赋予植物强度和硬度(Cosgrove and Jarvis, 2012; Li et al.2016; 张雨等, 2020).初生细胞壁纤维素含量较少, 果胶多糖含量较多, 在双子叶和非禾本科植物中, 果胶约占初生细胞壁的35% (Cosgrove and Jarvis, 2012).次生细胞壁的果胶含量尚不明确, 但有研究表明, 杨树(Populus trichocarpa, P. nigra × P. maximowiczii, P. trichocarpa×P. koreana)成熟木质部中有类果胶(pectin-like)组分在次生细胞壁中沉积(Arend et al.2008). ...
果胶结构域精细结构研究进展 1 2020
... HG是α-1,4连接的半乳糖醛酸(α-1,4-linked D-galacturonic acid, GalA)线性同聚物, 在果胶中含量最高(Mohnen,2008).HG可以在C-6羧基处进行甲基化, 参与甲基化修饰的酶包括HG半乳糖醛酸转移酶(galacturonosyltransferases, GAUT)和果胶甲基转移酶(pectin methyl-transferase, PMT) (Sebastian et al.2009).在数据库CAZy中检索发现, GAUT蛋白属于糖基转移酶(glycosyltransferases, GT)家族8, 该家族在拟南芥(Arabidopsis thaliana)中有15个成员.其中参与果胶生物合成的酶有GAUT1 (Sterling et al.2006)、GAUT7 (Atmodjo et al.2011)、GAUT4 (Biswal et al.2018)和GAUT8 (QUA1) (Bouton et al.2002).GAUT1是首个被鉴定的HG半乳糖醛酸转移酶, 它能在体外合成聚半乳糖醛酸(Sterling et al.2006); GAUT7是GAUT1的同源蛋白, 二者形成GAUT1:GAUT7复合物, 共同参与细胞壁基质多糖的生物合成(Atmodjo et al.2011); GAUT4可以合成HG, 其表达下调使细胞壁中的HG和RG-II减少(Biswal et al.2018); qua1突变体中HG含量极低, 表明QUA1可能参与果胶的生物合成(Bouton et al.2002).PMT通过催化S-腺苷-L-甲硫氨酸的甲基转移到果胶HG中α-1,4连接的半乳糖醛酸残基的羧基上, 使HG发生甲基化修饰(Liuet al.2015).PMT与GAUT可能作为异源复合体参与HG的聚合, 使HG完全甲基化(即甲基化程度达到80%).此外, HG的一些GalA残基在O-2或O-3处可发生乙酰化修饰(易建勇等, 2020).甲基化和乙酰化的酯化程度是动态可变的, 其修饰的程度影响果胶的理化性质.例如, 在Ca2+的作用下, 两条HG链之间相互作用形成二聚体.值得注意的是, 至少需要9个未甲基化修饰的GalA残基的连续延伸才能实现此同型二聚体(Scheller et al.2007). ...
植物细胞壁同聚半乳糖醛酸的代谢与功能 1 2014
... 细胞壁作为抵御病原体的屏障, 其完整性为植物防御病原体所必需(Hamann, 2015).植物细胞壁果胶被细菌和真菌分泌的果胶降解酶降解, 其中HG降解后产生去甲基酯化的果胶片段或寡聚半乳糖醛酸(oligogalacturonides, OGs) (Ferrari et al.2013; 郁有健等, 2014).OGs作为激发子可被植物体内的受体识别, 从而激活一系列防御反应, 起到保护植物的作用.研究表明, OGs的信号转导通过细胞壁关联激酶(wall-associated kinases, WAKs)发挥作用, WAKs能感知OGs, 进而激活植物的应激反应(Brutus et al.2010; Kohorn et al.2014).此外, 病原体入侵会激活PME, PME催化HG去甲基酯化过程中产生具有挥发性的甲醇, 甲醇可作为信号分子参与启动邻近植物的免疫反应, 增强邻近植物对细菌病原体的抵抗力(Komarova et al.2014). ...
果胶酯酶的研究进展 1 2010
... HG在高尔基体中以高甲基酯化的形式合成, 随后由果胶甲基酯酶(pectin methylesterase, PME)选择性地去甲基酯化(Caffall and Mohnen, 2009), 同时释放甲醇和质子(查笑君等, 2010), 生成带负电荷的羧基(Wormit and Usadel, 2018).去甲基酯化的HG可以增强细胞壁的刚性.例如, PMEs催化HG去甲基酯化后可形成连续的GalAs, 在Ca2+存在时, 带负电荷的羧基与Ca2+相互作用, 形成“鸡蛋盒” (Egg Box Model)结构(图1).该结构可以通过增强细胞壁刚性来维持细胞壁的完整性(Micheli, 2001; Braccini and Peréz,2001); 花粉管的顶端区域几乎只存在甲基酯化的果胶, 外源施加PME时可诱导其尖端细胞壁增厚, 从而抑制花粉管生长(Bosch et al.2005).此外, PME去甲基酯化HG的过程中会导致局部pH降低, 激活多种细胞壁降解酶, 从而促进细胞壁的扩张, 使其弹性增强.相反, 当植物体缺少PME时, 细胞壁的弹性降低.研究发现, 果胶甲基酯酶6 (PME6)在保卫细胞中表达量较高, 野生型保卫细胞富含未酯化的HG, 而突变体pme6-1的保卫细胞壁富含甲基酯化的果胶, 其气孔开启/关闭动态范围缩小, 表明异常的果胶甲基酯化改变了保卫细胞的机械特性, 导致气孔无法根据环境信号进行适度的开放与关闭(Amsbury et al.2016). ...
植物次生细胞壁生物合成的转录调控网络 1 2020
... 细胞壁作为植物细胞的屏障, 在维持细胞膨压、完整性以及细胞间信号交流方面具有重要作用(Cosgrove, 2005).细胞壁包括初生细胞壁和次生细胞壁, 均由多糖和少量蛋白组成(Logan et al.2014), 但组成成分和功能有所不同.其中初生细胞壁主要由纤维素、半纤维素和果胶组成(解敏敏等, 2015), 具有可塑性, 可以适应植物细胞生长过程中的动态变化, 对植物细胞的形态建成具有重要作用(Sakamoto et al.2018).次生细胞壁主要由纤维素、半纤维素和木质素组成(Zhong and Ye, 2015), 刚性较强, 可赋予植物强度和硬度(Cosgrove and Jarvis, 2012; Li et al.2016; 张雨等, 2020).初生细胞壁纤维素含量较少, 果胶多糖含量较多, 在双子叶和非禾本科植物中, 果胶约占初生细胞壁的35% (Cosgrove and Jarvis, 2012).次生细胞壁的果胶含量尚不明确, 但有研究表明, 杨树(Populus trichocarpa, P. nigra × P. maximowiczii, P. trichocarpa×P. koreana)成熟木质部中有类果胶(pectin-like)组分在次生细胞壁中沉积(Arend et al.2008). ...
Depletion of UDP-D-apiose/ UDP-D-xylose synthases results in rhamnogalacturonan- II deficiency, cell wall thickening, and cell death in higher plants 1 2006
... RGII结构虽然复杂, 但其结构在植物中高度保守(Matsunaga et al.2004).在植物细胞壁中, 90%以上的RGII以二聚体形式存在.如图2所示, RGII二聚体由侧链A上的芹菜糖残基之间的硼酸二酯键共价交联形成(O’Neillet al.2004; Atmodjo et al.2013), 侧链A的完整结构对RGII二聚体的形成至关重要.当RGII完整性缺失时会影响植物的生长, 导致植物矮化(O’Neillet al.2001)和花粉管伸长缺陷(Delmas et al.2008), 严重时可导致植物细胞死亡(Ahn et al.2006).在拟南芥中, MUR1 (MURUS1)编码GDP-甘露糖-4,6-脱水酶, 催化由GDP-D-mannose到GDP- L-focuse的第1步合成.GDP-L-focuse是L-岩藻糖(L- focuse, 6-deoxy-L-galactose)的活化核苷酸糖形式, L-岩藻糖是RGII侧链A的单糖组成之一, 也是其它多种结构多糖和糖蛋白的组成部分(Zhang et al.2019).MUR1功能缺失突变体mur1植株矮化, RGII交联率降低(O’Neillet al.2001), 可能是由于侧链A截断所致(Pabst et al.2013).进一步研究发现, mur1突变体侧链A中的L-focuse被L-Gal取代后, 全长侧链的比例降低, 导致RGII对硼的亲和力降低, 二聚体形成减少, 造成mur1植株地上部矮化(Reuhs et al.2004).在植物细胞壁中, 硼将2条RGII链连接在一起形成二聚体, 增强了细胞之间的黏附和机械强度.研究发现, 通过补充硼可增加RGII的交联比例, 同时减弱mur1突变体的矮化表型, 表明硼酸盐引起的RGII交联对植物正常生长至关重要(O’Neillet al.2001). ...
Stomatal function requires pectin de- methyl-esterification of the guard cell wall 1 2016
... HG在高尔基体中以高甲基酯化的形式合成, 随后由果胶甲基酯酶(pectin methylesterase, PME)选择性地去甲基酯化(Caffall and Mohnen, 2009), 同时释放甲醇和质子(查笑君等, 2010), 生成带负电荷的羧基(Wormit and Usadel, 2018).去甲基酯化的HG可以增强细胞壁的刚性.例如, PMEs催化HG去甲基酯化后可形成连续的GalAs, 在Ca2+存在时, 带负电荷的羧基与Ca2+相互作用, 形成“鸡蛋盒” (Egg Box Model)结构(图1).该结构可以通过增强细胞壁刚性来维持细胞壁的完整性(Micheli, 2001; Braccini and Peréz,2001); 花粉管的顶端区域几乎只存在甲基酯化的果胶, 外源施加PME时可诱导其尖端细胞壁增厚, 从而抑制花粉管生长(Bosch et al.2005).此外, PME去甲基酯化HG的过程中会导致局部pH降低, 激活多种细胞壁降解酶, 从而促进细胞壁的扩张, 使其弹性增强.相反, 当植物体缺少PME时, 细胞壁的弹性降低.研究发现, 果胶甲基酯酶6 (PME6)在保卫细胞中表达量较高, 野生型保卫细胞富含未酯化的HG, 而突变体pme6-1的保卫细胞壁富含甲基酯化的果胶, 其气孔开启/关闭动态范围缩小, 表明异常的果胶甲基酯化改变了保卫细胞的机械特性, 导致气孔无法根据环境信号进行适度的开放与关闭(Amsbury et al.2016). ...
Metabolic click-labeling with a fucose analog reveals pectin delivery, architecture, and dynamics in Arabidopsis cell walls 1 2012
Unique occurrence of pectin-like fibrillar cell wall deposits in xylem fibres of poplar 1 2008
... 细胞壁作为植物细胞的屏障, 在维持细胞膨压、完整性以及细胞间信号交流方面具有重要作用(Cosgrove, 2005).细胞壁包括初生细胞壁和次生细胞壁, 均由多糖和少量蛋白组成(Logan et al.2014), 但组成成分和功能有所不同.其中初生细胞壁主要由纤维素、半纤维素和果胶组成(解敏敏等, 2015), 具有可塑性, 可以适应植物细胞生长过程中的动态变化, 对植物细胞的形态建成具有重要作用(Sakamoto et al.2018).次生细胞壁主要由纤维素、半纤维素和木质素组成(Zhong and Ye, 2015), 刚性较强, 可赋予植物强度和硬度(Cosgrove and Jarvis, 2012; Li et al.2016; 张雨等, 2020).初生细胞壁纤维素含量较少, 果胶多糖含量较多, 在双子叶和非禾本科植物中, 果胶约占初生细胞壁的35% (Cosgrove and Jarvis, 2012).次生细胞壁的果胶含量尚不明确, 但有研究表明, 杨树(Populus trichocarpa, P. nigra × P. maximowiczii, P. trichocarpa×P. koreana)成熟木质部中有类果胶(pectin-like)组分在次生细胞壁中沉积(Arend et al.2008). ...
Evolving views of pectin biosynthesis 5 2013
... 作为植物细胞壁的重要组分, 果胶广泛存在于高等植物的根、茎和叶等器官的细胞壁和细胞间隙中, 在植物生长发育、防御和细胞壁完整性等方面具有重要作用.果胶多糖是自然界中结构最复杂的多糖家族, 组成成分主要包括同型半乳糖醛酸聚糖(homogalacturonan, HG)、鼠李半乳糖醛酸聚糖I (rhamngalacturonan I, RGI)和鼠李半乳糖醛酸聚糖II (rhamngalacturonan II, RGII), 不同的果胶结构域在细胞壁中彼此共价连接(Atmodjo et al.2013).然而, 这3种组分的交联方式与其功能之间的关系尚不明确.因此, 研究果胶的结构与功能有利于更加深刻理解细胞壁高级结构的构建与功能.本文主要从果胶HG、RGI和RGII的生物合成、功能以及果胶的显微成像3个方面进行综述. ...
... RGI占果胶成分的20%-35%, 与HG和RGII不同, 它的主链由双糖重复链[4)-α-D-GalA-(1,2)-α-L-Rha- (1, ]n组成.RGI主链中的大部分或全部GalA在O-2或O-3处被乙酰化修饰, 大多数鼠李糖(rhamnose, Rha)残基可在O-4处被线性或支链寡糖或多糖取代(Atmodjo et al.2013).RGI侧链包括β-1,4-半乳聚糖、阿拉伯聚糖或阿拉伯半乳聚糖等.RGI生物合成过程中, 其主链和侧链合成所需酶不尽相同, 主链合成需要半乳糖醛酸转移酶(galacturonosyltransferase, GalAT)和鼠李糖基转移酶(rhamnosyltransferase, RhaT)参与, 而侧链合成需要半乳糖基转移酶(galactosyltransferases, GalTs)和阿拉伯糖基转移酶(arabinosyltransferases, AraTs).目前在拟南芥中已有报道的GalTs和AraTs编码基因包括GT92家族的GALS1 (GALACTAN SYNTHASE 1) (Liwanag et al.2012)和GT47家族B亚群中的ARAD1 (ARABINAN DEFICIENT 1) (Harholt et al.2006)及其同源基因ARAD2(Harholt et al.2012).其中GALS1基因功能缺失和过表达分别导致细胞壁半乳聚糖的减少和增加(Liwanag et al.2012).对arad1突变体的生化分析显示, 突变体细胞壁中的阿拉伯糖数量明显低于野生型(Harholt et al.2006); 而对arad2突变体细胞壁的生化分析显示, 其单糖组成与野生型无明显差异, 且过表达ARAD2并不能完全恢复arad1的表型, 说明虽然ARAD1与ARAD2同源, 但是二者功能并不冗余, 推测这2个糖基转移酶可能在二硫键连接的复合物形成中发挥作用(Harholt et al.2012). ...
... RGII结构虽然复杂, 但其结构在植物中高度保守(Matsunaga et al.2004).在植物细胞壁中, 90%以上的RGII以二聚体形式存在.如图2所示, RGII二聚体由侧链A上的芹菜糖残基之间的硼酸二酯键共价交联形成(O’Neillet al.2004; Atmodjo et al.2013), 侧链A的完整结构对RGII二聚体的形成至关重要.当RGII完整性缺失时会影响植物的生长, 导致植物矮化(O’Neillet al.2001)和花粉管伸长缺陷(Delmas et al.2008), 严重时可导致植物细胞死亡(Ahn et al.2006).在拟南芥中, MUR1 (MURUS1)编码GDP-甘露糖-4,6-脱水酶, 催化由GDP-D-mannose到GDP- L-focuse的第1步合成.GDP-L-focuse是L-岩藻糖(L- focuse, 6-deoxy-L-galactose)的活化核苷酸糖形式, L-岩藻糖是RGII侧链A的单糖组成之一, 也是其它多种结构多糖和糖蛋白的组成部分(Zhang et al.2019).MUR1功能缺失突变体mur1植株矮化, RGII交联率降低(O’Neillet al.2001), 可能是由于侧链A截断所致(Pabst et al.2013).进一步研究发现, mur1突变体侧链A中的L-focuse被L-Gal取代后, 全长侧链的比例降低, 导致RGII对硼的亲和力降低, 二聚体形成减少, 造成mur1植株地上部矮化(Reuhs et al.2004).在植物细胞壁中, 硼将2条RGII链连接在一起形成二聚体, 增强了细胞之间的黏附和机械强度.研究发现, 通过补充硼可增加RGII的交联比例, 同时减弱mur1突变体的矮化表型, 表明硼酸盐引起的RGII交联对植物正常生长至关重要(O’Neillet al.2001). ...
... 野生型植株中的鼠李半乳糖醛酸聚糖II二聚体由侧链A的芹菜糖残基通过硼酸二酯键共价交联而成.MUR1催化GDP-L-岩藻糖从头合成的第一步, MUR1功能缺失导致GDP-L-岩藻糖合成受阻.因此, mur1突变体中的鼠李半乳糖醛酸聚糖II侧链A被截断, 这一缺陷导致鼠李半乳糖醛酸聚糖II二聚体的形成减少.HG: 同型半乳糖醛酸聚糖; Api: 芹菜糖; Rha: L-鼠李糖; Fuc: L-岩藻糖; GlcA: D-葡萄糖醛酸; Gal: L-半乳糖; GalA: D-半乳糖醛酸; Me xyl: 2-O-甲基-D-木糖 Rhamngalacturonan II (RGII) dimer in Arabidopsis wild-type and mur1 mutants (modified from O’Neill <i>et al.</i> <xref ref-type="bibr" rid="b52">2004</xref>; Atmodjo <i>et al.</i> <xref ref-type="bibr" rid="b10">2013</xref>) In wild-type plants, RGII dimers are formed between Apiosyl residue of side chain A which are cross-linked covalently by the diester borate bonds. MUR1 catalyzes the first step in the de novo synthesis of GDP-L-fucose. The loss of MUR1 function leads to the block of GDP-L-fucose synthesis. Therefore, the RGII side chain A was truncated in mur1 mutants which resulted in reduced formation of RG-II dimers. HG: Homogalacturonan; Api: D-apiosyl; Rha: L-rhamnose; Fuc: L-fucose; GlcA: D-glucuronic acid; Gal: L-galactose; GalA: D-galacturonic acid; Me xyl: 2-O-methyl-D-xylose ...
Galacturonosyltransferase (GAUT)1 and GAUT7 are the core of a plant cell wall pectin biosynthetic homogalacturonan: galacturonosyltransferase complex 2 2011
... HG是α-1,4连接的半乳糖醛酸(α-1,4-linked D-galacturonic acid, GalA)线性同聚物, 在果胶中含量最高(Mohnen,2008).HG可以在C-6羧基处进行甲基化, 参与甲基化修饰的酶包括HG半乳糖醛酸转移酶(galacturonosyltransferases, GAUT)和果胶甲基转移酶(pectin methyl-transferase, PMT) (Sebastian et al.2009).在数据库CAZy中检索发现, GAUT蛋白属于糖基转移酶(glycosyltransferases, GT)家族8, 该家族在拟南芥(Arabidopsis thaliana)中有15个成员.其中参与果胶生物合成的酶有GAUT1 (Sterling et al.2006)、GAUT7 (Atmodjo et al.2011)、GAUT4 (Biswal et al.2018)和GAUT8 (QUA1) (Bouton et al.2002).GAUT1是首个被鉴定的HG半乳糖醛酸转移酶, 它能在体外合成聚半乳糖醛酸(Sterling et al.2006); GAUT7是GAUT1的同源蛋白, 二者形成GAUT1:GAUT7复合物, 共同参与细胞壁基质多糖的生物合成(Atmodjo et al.2011); GAUT4可以合成HG, 其表达下调使细胞壁中的HG和RG-II减少(Biswal et al.2018); qua1突变体中HG含量极低, 表明QUA1可能参与果胶的生物合成(Bouton et al.2002).PMT通过催化S-腺苷-L-甲硫氨酸的甲基转移到果胶HG中α-1,4连接的半乳糖醛酸残基的羧基上, 使HG发生甲基化修饰(Liuet al.2015).PMT与GAUT可能作为异源复合体参与HG的聚合, 使HG完全甲基化(即甲基化程度达到80%).此外, HG的一些GalA残基在O-2或O-3处可发生乙酰化修饰(易建勇等, 2020).甲基化和乙酰化的酯化程度是动态可变的, 其修饰的程度影响果胶的理化性质.例如, 在Ca2+的作用下, 两条HG链之间相互作用形成二聚体.值得注意的是, 至少需要9个未甲基化修饰的GalA残基的连续延伸才能实现此同型二聚体(Scheller et al.2007). ...
... 2011); GAUT4可以合成HG, 其表达下调使细胞壁中的HG和RG-II减少(Biswal et al.2018); qua1突变体中HG含量极低, 表明QUA1可能参与果胶的生物合成(Bouton et al.2002).PMT通过催化S-腺苷-L-甲硫氨酸的甲基转移到果胶HG中α-1,4连接的半乳糖醛酸残基的羧基上, 使HG发生甲基化修饰(Liuet al.2015).PMT与GAUT可能作为异源复合体参与HG的聚合, 使HG完全甲基化(即甲基化程度达到80%).此外, HG的一些GalA残基在O-2或O-3处可发生乙酰化修饰(易建勇等, 2020).甲基化和乙酰化的酯化程度是动态可变的, 其修饰的程度影响果胶的理化性质.例如, 在Ca2+的作用下, 两条HG链之间相互作用形成二聚体.值得注意的是, 至少需要9个未甲基化修饰的GalA残基的连续延伸才能实现此同型二聚体(Scheller et al.2007). ...
Fluorescence visualization of cellulose and pectin in the primary plant cell wall 1 2020
Molecular basis of Ca 2+-induced gelation in alginates and pectins: the egg-box model revisited 3 2001
... HG在高尔基体中以高甲基酯化的形式合成, 随后由果胶甲基酯酶(pectin methylesterase, PME)选择性地去甲基酯化(Caffall and Mohnen, 2009), 同时释放甲醇和质子(查笑君等, 2010), 生成带负电荷的羧基(Wormit and Usadel, 2018).去甲基酯化的HG可以增强细胞壁的刚性.例如, PMEs催化HG去甲基酯化后可形成连续的GalAs, 在Ca2+存在时, 带负电荷的羧基与Ca2+相互作用, 形成“鸡蛋盒” (Egg Box Model)结构(图1).该结构可以通过增强细胞壁刚性来维持细胞壁的完整性(Micheli, 2001; Braccini and Peréz,2001); 花粉管的顶端区域几乎只存在甲基酯化的果胶, 外源施加PME时可诱导其尖端细胞壁增厚, 从而抑制花粉管生长(Bosch et al.2005).此外, PME去甲基酯化HG的过程中会导致局部pH降低, 激活多种细胞壁降解酶, 从而促进细胞壁的扩张, 使其弹性增强.相反, 当植物体缺少PME时, 细胞壁的弹性降低.研究发现, 果胶甲基酯酶6 (PME6)在保卫细胞中表达量较高, 野生型保卫细胞富含未酯化的HG, 而突变体pme6-1的保卫细胞壁富含甲基酯化的果胶, 其气孔开启/关闭动态范围缩小, 表明异常的果胶甲基酯化改变了保卫细胞的机械特性, 导致气孔无法根据环境信号进行适度的开放与关闭(Amsbury et al.2016). ...
... ; Braccini and Peréz,2001) 高甲基酯化的HG被果胶甲基酯酶(PME)去甲基酯化, 产生带负电荷的羧基.在Ca2+存在时, 2条HG链的羧基与Ca2+相互作用, 形成“鸡蛋盒”结构. ...
... 高甲基酯化的HG被果胶甲基酯酶(PME)去甲基酯化, 产生带负电荷的羧基.在Ca2+存在时, 2条HG链的羧基与Ca2+相互作用, 形成“鸡蛋盒”结构. An “egg-box” model of homogalacturonan (HG) polysaccharide and Ca <sup>2+</sup> cross-linking (modified from Micheli, <xref ref-type="bibr" rid="b45">2001</xref>; Braccini and Peréz,<xref ref-type="bibr" rid="b16">2001</xref>) Hypermethylated HG is de-methylated by pectin methylesterase (PME) to produce negatively charged carboxyl groups. In the presence of Ca2+, the carboxyl groups of the two HG chains interact with Ca2+ to form an “egg box” structure. ...
A domain swap approach reveals a role of the plant wall-associated kinase 1 (WAK1) as a receptor of oligogalacturonides 1 2010
... 细胞壁作为抵御病原体的屏障, 其完整性为植物防御病原体所必需(Hamann, 2015).植物细胞壁果胶被细菌和真菌分泌的果胶降解酶降解, 其中HG降解后产生去甲基酯化的果胶片段或寡聚半乳糖醛酸(oligogalacturonides, OGs) (Ferrari et al.2013; 郁有健等, 2014).OGs作为激发子可被植物体内的受体识别, 从而激活一系列防御反应, 起到保护植物的作用.研究表明, OGs的信号转导通过细胞壁关联激酶(wall-associated kinases, WAKs)发挥作用, WAKs能感知OGs, 进而激活植物的应激反应(Brutus et al.2010; Kohorn et al.2014).此外, 病原体入侵会激活PME, PME催化HG去甲基酯化过程中产生具有挥发性的甲醇, 甲醇可作为信号分子参与启动邻近植物的免疫反应, 增强邻近植物对细菌病原体的抵抗力(Komarova et al.2014). ...
The structure, function, and biosynthesis of plant cell wall pectic polysaccharides 1 2009
... HG在高尔基体中以高甲基酯化的形式合成, 随后由果胶甲基酯酶(pectin methylesterase, PME)选择性地去甲基酯化(Caffall and Mohnen, 2009), 同时释放甲醇和质子(查笑君等, 2010), 生成带负电荷的羧基(Wormit and Usadel, 2018).去甲基酯化的HG可以增强细胞壁的刚性.例如, PMEs催化HG去甲基酯化后可形成连续的GalAs, 在Ca2+存在时, 带负电荷的羧基与Ca2+相互作用, 形成“鸡蛋盒” (Egg Box Model)结构(图1).该结构可以通过增强细胞壁刚性来维持细胞壁的完整性(Micheli, 2001; Braccini and Peréz,2001); 花粉管的顶端区域几乎只存在甲基酯化的果胶, 外源施加PME时可诱导其尖端细胞壁增厚, 从而抑制花粉管生长(Bosch et al.2005).此外, PME去甲基酯化HG的过程中会导致局部pH降低, 激活多种细胞壁降解酶, 从而促进细胞壁的扩张, 使其弹性增强.相反, 当植物体缺少PME时, 细胞壁的弹性降低.研究发现, 果胶甲基酯酶6 (PME6)在保卫细胞中表达量较高, 野生型保卫细胞富含未酯化的HG, 而突变体pme6-1的保卫细胞壁富含甲基酯化的果胶, 其气孔开启/关闭动态范围缩小, 表明异常的果胶甲基酯化改变了保卫细胞的机械特性, 导致气孔无法根据环境信号进行适度的开放与关闭(Amsbury et al.2016). ...
Pectic arabinan side chains are essential for pollen cell wall integrity during pollen development 1 2014
... RGI侧链包括β-1,4-半乳聚糖、阿拉伯聚糖或阿拉伯半乳聚糖, 其组成和结构影响细胞壁的强度、弹性和柔韧性等机械性能.例如, 侧链阿拉伯聚糖和阿拉伯半乳聚糖的长度和数量部分决定初生细胞壁的相对灵活性, 而半乳聚糖含量增加有助于增强细胞壁的硬度和刚度(Ulvskovet al.2005; Moore et al.2008).Cankar等(2014)通过表型观察和组分分析, 对编码果胶修饰、主链降解和侧链类型等酶的多个转基因马铃薯(Solanum tuberosum)株系进行研究, 发现果胶主链被鼠李半乳糖醛酸裂解酶(rhamno galacturonanlyase, RGL)裂解为片段的转基因株系, 以及阿拉伯聚糖含量降低70%的转基因株系中, 成熟花粉粒的内壁在亲和孔的位置外翻突起, 最终导致花粉粒破裂和细胞质泄漏, 这些结果证实在花粉粒成熟和脱水过程中, 果胶阿拉伯聚糖侧链在细胞壁重塑过程中发挥了重要作用.McCartney等(2000)通过对(1→4)- β-D-半乳聚糖出现前后的豌豆(Pisum sativum cv. ‘Avola’)子叶进行机械压应力测试, 发现细胞壁富含半乳聚糖的子叶硬度明显增强.此外, RGI侧链的组成和结构与细胞和组织的发育阶段密切相关.例如, 在胡萝卜( Daucus carotacv. ‘Early Nantes’)由细胞分裂向细胞伸长过渡时, 伴随着细胞壁果胶半乳聚糖的出现和阿拉伯聚糖减少; 豌豆子叶在发育中随着果胶半乳聚糖的出现表现出组织硬度增强(Willats et al.1999; McCartney et al.2000). ...
Synthetic methyl hexagalacturonate hapten inhibitors of anti-homogalacturonan monoclonal antibodies LM7, JIM5 and JIM7 2 2003
... 在果胶多糖的3种组分中, 对于HG的标记成像研究较多, 用于标记HG的单克隆抗体有JIM5、JIM7和2F4等.其中, JIM5的最佳识别表位可能是4个以上相邻的未酯化GalA残基, 这些残基与甲基酯化的GalA残基相邻或在其两侧(Clausen et al.2003); JIM7识别表位的重要特征是每隔一段残基上都有甲基酯化的GalA残基(Clausen et al.2003); 2F4可结合钙离子交联的未酯化的GalA残基(Sujkowska-Rybkowska and Borucki, 2015).我们用JIM5和JIM7对青扦(Picea wilsonii)花粉管细胞壁中的果胶进行标记, 在激光共聚焦显微镜下用488 nm激发并在500-550 nm处采集发射信号, 可观察到在花粉管细胞壁中, JIM7标记的高甲基酯化的果胶聚集在生长的花粉管顶端, JIM5标记的低甲基酯化的果胶分布在花粉管伸长处(Cui et al.2015).此外, 我们利用JIM5和JIM7免疫标记, 还检测了杉木(Cunninghamia lanceolata)维管形成层细胞壁多糖在季节周期中的变化, 结果显示, 休眠期形成层细胞壁中富含低甲基酯化的HG, 而活跃期形成层细胞壁中高甲基酯化的HG较多(Wu et al.2016).Sujkowska-Rybkowska和Borucki (2015)使用JIM5、JIM7和2F4分别对低甲基酯化果胶、高甲基酯化果胶和钙交联果胶进行免疫标记, 检测铝处理的豌豆根瘤中果胶酯化和分布模式.Mravec等(2014)利用低聚壳聚糖(chitosan oligosaccharides, COS)与去酯化的HG果胶可以特异性结合的特点, 开发了非免疫标记分子探针.应用Alexa Fluor 488偶联COS (COS488)标记去甲基酯化HG, 可以对其分布进行高分辨率荧光成像观察; COS与纳米金粒子耦合(COSAuNP)后, 可以用于在透射电子显微镜下对HG定位进行超微结构水平的观察.与抗体免疫标记相比较, COS488具有穿透速度快和对酯化程度的变化敏感等特点, 可用于活细胞实时成像. ...
... 细胞壁作为植物细胞的屏障, 在维持细胞膨压、完整性以及细胞间信号交流方面具有重要作用(Cosgrove, 2005).细胞壁包括初生细胞壁和次生细胞壁, 均由多糖和少量蛋白组成(Logan et al.2014), 但组成成分和功能有所不同.其中初生细胞壁主要由纤维素、半纤维素和果胶组成(解敏敏等, 2015), 具有可塑性, 可以适应植物细胞生长过程中的动态变化, 对植物细胞的形态建成具有重要作用(Sakamoto et al.2018).次生细胞壁主要由纤维素、半纤维素和木质素组成(Zhong and Ye, 2015), 刚性较强, 可赋予植物强度和硬度(Cosgrove and Jarvis, 2012; Li et al.2016; 张雨等, 2020).初生细胞壁纤维素含量较少, 果胶多糖含量较多, 在双子叶和非禾本科植物中, 果胶约占初生细胞壁的35% (Cosgrove and Jarvis, 2012).次生细胞壁的果胶含量尚不明确, 但有研究表明, 杨树(Populus trichocarpa, P. nigra × P. maximowiczii, P. trichocarpa×P. koreana)成熟木质部中有类果胶(pectin-like)组分在次生细胞壁中沉积(Arend et al.2008). ...
Comparative structure and biomechanics of plant primary and secondary cell walls 2 2012
... 细胞壁作为植物细胞的屏障, 在维持细胞膨压、完整性以及细胞间信号交流方面具有重要作用(Cosgrove, 2005).细胞壁包括初生细胞壁和次生细胞壁, 均由多糖和少量蛋白组成(Logan et al.2014), 但组成成分和功能有所不同.其中初生细胞壁主要由纤维素、半纤维素和果胶组成(解敏敏等, 2015), 具有可塑性, 可以适应植物细胞生长过程中的动态变化, 对植物细胞的形态建成具有重要作用(Sakamoto et al.2018).次生细胞壁主要由纤维素、半纤维素和木质素组成(Zhong and Ye, 2015), 刚性较强, 可赋予植物强度和硬度(Cosgrove and Jarvis, 2012; Li et al.2016; 张雨等, 2020).初生细胞壁纤维素含量较少, 果胶多糖含量较多, 在双子叶和非禾本科植物中, 果胶约占初生细胞壁的35% (Cosgrove and Jarvis, 2012).次生细胞壁的果胶含量尚不明确, 但有研究表明, 杨树(Populus trichocarpa, P. nigra × P. maximowiczii, P. trichocarpa×P. koreana)成熟木质部中有类果胶(pectin-like)组分在次生细胞壁中沉积(Arend et al.2008). ...
... ).初生细胞壁纤维素含量较少, 果胶多糖含量较多, 在双子叶和非禾本科植物中, 果胶约占初生细胞壁的35% (Cosgrove and Jarvis, 2012).次生细胞壁的果胶含量尚不明确, 但有研究表明, 杨树(Populus trichocarpa, P. nigra × P. maximowiczii, P. trichocarpa×P. koreana)成熟木质部中有类果胶(pectin-like)组分在次生细胞壁中沉积(Arend et al.2008). ...
Interference of the histone deacetylase inhibits pollen germination and pollen tube growth in Picea wilsonii mast 1 2015
... 在果胶多糖的3种组分中, 对于HG的标记成像研究较多, 用于标记HG的单克隆抗体有JIM5、JIM7和2F4等.其中, JIM5的最佳识别表位可能是4个以上相邻的未酯化GalA残基, 这些残基与甲基酯化的GalA残基相邻或在其两侧(Clausen et al.2003); JIM7识别表位的重要特征是每隔一段残基上都有甲基酯化的GalA残基(Clausen et al.2003); 2F4可结合钙离子交联的未酯化的GalA残基(Sujkowska-Rybkowska and Borucki, 2015).我们用JIM5和JIM7对青扦(Picea wilsonii)花粉管细胞壁中的果胶进行标记, 在激光共聚焦显微镜下用488 nm激发并在500-550 nm处采集发射信号, 可观察到在花粉管细胞壁中, JIM7标记的高甲基酯化的果胶聚集在生长的花粉管顶端, JIM5标记的低甲基酯化的果胶分布在花粉管伸长处(Cui et al.2015).此外, 我们利用JIM5和JIM7免疫标记, 还检测了杉木(Cunninghamia lanceolata)维管形成层细胞壁多糖在季节周期中的变化, 结果显示, 休眠期形成层细胞壁中富含低甲基酯化的HG, 而活跃期形成层细胞壁中高甲基酯化的HG较多(Wu et al.2016).Sujkowska-Rybkowska和Borucki (2015)使用JIM5、JIM7和2F4分别对低甲基酯化果胶、高甲基酯化果胶和钙交联果胶进行免疫标记, 检测铝处理的豌豆根瘤中果胶酯化和分布模式.Mravec等(2014)利用低聚壳聚糖(chitosan oligosaccharides, COS)与去酯化的HG果胶可以特异性结合的特点, 开发了非免疫标记分子探针.应用Alexa Fluor 488偶联COS (COS488)标记去甲基酯化HG, 可以对其分布进行高分辨率荧光成像观察; COS与纳米金粒子耦合(COSAuNP)后, 可以用于在透射电子显微镜下对HG定位进行超微结构水平的观察.与抗体免疫标记相比较, COS488具有穿透速度快和对酯化程度的变化敏感等特点, 可用于活细胞实时成像. ...
The synthesis of the rhamnogalacturonan II component 3-deoxy-D- manno-2- octulosonic acid (Kdo) is required for pollen tube growth and elongation 1 2008
... RGII结构虽然复杂, 但其结构在植物中高度保守(Matsunaga et al.2004).在植物细胞壁中, 90%以上的RGII以二聚体形式存在.如图2所示, RGII二聚体由侧链A上的芹菜糖残基之间的硼酸二酯键共价交联形成(O’Neillet al.2004; Atmodjo et al.2013), 侧链A的完整结构对RGII二聚体的形成至关重要.当RGII完整性缺失时会影响植物的生长, 导致植物矮化(O’Neillet al.2001)和花粉管伸长缺陷(Delmas et al.2008), 严重时可导致植物细胞死亡(Ahn et al.2006).在拟南芥中, MUR1 (MURUS1)编码GDP-甘露糖-4,6-脱水酶, 催化由GDP-D-mannose到GDP- L-focuse的第1步合成.GDP-L-focuse是L-岩藻糖(L- focuse, 6-deoxy-L-galactose)的活化核苷酸糖形式, L-岩藻糖是RGII侧链A的单糖组成之一, 也是其它多种结构多糖和糖蛋白的组成部分(Zhang et al.2019).MUR1功能缺失突变体mur1植株矮化, RGII交联率降低(O’Neillet al.2001), 可能是由于侧链A截断所致(Pabst et al.2013).进一步研究发现, mur1突变体侧链A中的L-focuse被L-Gal取代后, 全长侧链的比例降低, 导致RGII对硼的亲和力降低, 二聚体形成减少, 造成mur1植株地上部矮化(Reuhs et al.2004).在植物细胞壁中, 硼将2条RGII链连接在一起形成二聚体, 增强了细胞之间的黏附和机械强度.研究发现, 通过补充硼可增加RGII的交联比例, 同时减弱mur1突变体的矮化表型, 表明硼酸盐引起的RGII交联对植物正常生长至关重要(O’Neillet al.2001). ...
Plant cell wall imaging by metabolic click-mediated labelling of rhamnogalacturonan II using azido 3-deoxy-D- manno-oct-2-ulosonic acid 2 2016
Functional characterisation of a putative rhamnogalacturonan II specific xylosyltransferase 1 2008
... RGII的结构非常复杂, 由至少12种不同的单糖构成, 其主干是HG, 主干上带有4个不同的侧链(A-D), 链A和B在拟南芥中分别被描述为高度分化的八糖和七糖, 链C和D是双糖(Glushka et al.2003; Pabst et al.2013).值得注意的是, 侧链A和B的单糖组成中都含有芹菜糖残基(apiosyl, Api), 而侧链A的芹菜糖残基是形成硼酸二酯键的关键位点(Funakawa and Miwa, 2015).目前, 对RGII生物合成的研究较少, 已确定参与RGII生物合成的酶有鼠李半乳糖醛酸木糖基转移酶1-4 (rhamno galacturonan xylosyltransferase, RGXT1-4), 它们属于GT77的B亚群, 具有α-1,3-木糖基转移酶活性, 能将UDP-Xyl转移到岩藻糖(fucose, Fuc)上.RGXT1和RGXT2在拟南芥幼苗和成熟植株的营养组织中均有表达, 二者突变体的生长表型与野生型相比均无明显变化(Egelund et al.2006); RGXT3仅在成熟植株的叶片和角质层中表达(Egelund et al.2008); RGXT4在许多器官和组织中都有表达, 其功能缺失会严重影响花粉管和根的生长, 导致植株不育和幼苗死亡(Liu et al.2011). ...
Arabidopsis thaliana RGXT1 and RGXT2 encode golgi-localized (1,3)-α-D-xylosyltransferases involved in the synthesis of pectic rhamnogalacturonan-II 1 2006
... RGII的结构非常复杂, 由至少12种不同的单糖构成, 其主干是HG, 主干上带有4个不同的侧链(A-D), 链A和B在拟南芥中分别被描述为高度分化的八糖和七糖, 链C和D是双糖(Glushka et al.2003; Pabst et al.2013).值得注意的是, 侧链A和B的单糖组成中都含有芹菜糖残基(apiosyl, Api), 而侧链A的芹菜糖残基是形成硼酸二酯键的关键位点(Funakawa and Miwa, 2015).目前, 对RGII生物合成的研究较少, 已确定参与RGII生物合成的酶有鼠李半乳糖醛酸木糖基转移酶1-4 (rhamno galacturonan xylosyltransferase, RGXT1-4), 它们属于GT77的B亚群, 具有α-1,3-木糖基转移酶活性, 能将UDP-Xyl转移到岩藻糖(fucose, Fuc)上.RGXT1和RGXT2在拟南芥幼苗和成熟植株的营养组织中均有表达, 二者突变体的生长表型与野生型相比均无明显变化(Egelund et al.2006); RGXT3仅在成熟植株的叶片和角质层中表达(Egelund et al.2008); RGXT4在许多器官和组织中都有表达, 其功能缺失会严重影响花粉管和根的生长, 导致植株不育和幼苗死亡(Liu et al.2011). ...
Oligogalacturonides: plant damage-associated molecular patterns and regulators of growth and development 1 2013
... 细胞壁作为抵御病原体的屏障, 其完整性为植物防御病原体所必需(Hamann, 2015).植物细胞壁果胶被细菌和真菌分泌的果胶降解酶降解, 其中HG降解后产生去甲基酯化的果胶片段或寡聚半乳糖醛酸(oligogalacturonides, OGs) (Ferrari et al.2013; 郁有健等, 2014).OGs作为激发子可被植物体内的受体识别, 从而激活一系列防御反应, 起到保护植物的作用.研究表明, OGs的信号转导通过细胞壁关联激酶(wall-associated kinases, WAKs)发挥作用, WAKs能感知OGs, 进而激活植物的应激反应(Brutus et al.2010; Kohorn et al.2014).此外, 病原体入侵会激活PME, PME催化HG去甲基酯化过程中产生具有挥发性的甲醇, 甲醇可作为信号分子参与启动邻近植物的免疫反应, 增强邻近植物对细菌病原体的抵抗力(Komarova et al.2014). ...
Synthesis of borate cross- linked rhamnogalacturonan II 1 2015
... RGII的结构非常复杂, 由至少12种不同的单糖构成, 其主干是HG, 主干上带有4个不同的侧链(A-D), 链A和B在拟南芥中分别被描述为高度分化的八糖和七糖, 链C和D是双糖(Glushka et al.2003; Pabst et al.2013).值得注意的是, 侧链A和B的单糖组成中都含有芹菜糖残基(apiosyl, Api), 而侧链A的芹菜糖残基是形成硼酸二酯键的关键位点(Funakawa and Miwa, 2015).目前, 对RGII生物合成的研究较少, 已确定参与RGII生物合成的酶有鼠李半乳糖醛酸木糖基转移酶1-4 (rhamno galacturonan xylosyltransferase, RGXT1-4), 它们属于GT77的B亚群, 具有α-1,3-木糖基转移酶活性, 能将UDP-Xyl转移到岩藻糖(fucose, Fuc)上.RGXT1和RGXT2在拟南芥幼苗和成熟植株的营养组织中均有表达, 二者突变体的生长表型与野生型相比均无明显变化(Egelund et al.2006); RGXT3仅在成熟植株的叶片和角质层中表达(Egelund et al.2008); RGXT4在许多器官和组织中都有表达, 其功能缺失会严重影响花粉管和根的生长, 导致植株不育和幼苗死亡(Liu et al.2011). ...
Primary structure of the 2- O-methyl-α-L-fucose-containing side chain of the pectic polysaccharide, rhamnogalacturonan II 1 2003
... RGII的结构非常复杂, 由至少12种不同的单糖构成, 其主干是HG, 主干上带有4个不同的侧链(A-D), 链A和B在拟南芥中分别被描述为高度分化的八糖和七糖, 链C和D是双糖(Glushka et al.2003; Pabst et al.2013).值得注意的是, 侧链A和B的单糖组成中都含有芹菜糖残基(apiosyl, Api), 而侧链A的芹菜糖残基是形成硼酸二酯键的关键位点(Funakawa and Miwa, 2015).目前, 对RGII生物合成的研究较少, 已确定参与RGII生物合成的酶有鼠李半乳糖醛酸木糖基转移酶1-4 (rhamno galacturonan xylosyltransferase, RGXT1-4), 它们属于GT77的B亚群, 具有α-1,3-木糖基转移酶活性, 能将UDP-Xyl转移到岩藻糖(fucose, Fuc)上.RGXT1和RGXT2在拟南芥幼苗和成熟植株的营养组织中均有表达, 二者突变体的生长表型与野生型相比均无明显变化(Egelund et al.2006); RGXT3仅在成熟植株的叶片和角质层中表达(Egelund et al.2008); RGXT4在许多器官和组织中都有表达, 其功能缺失会严重影响花粉管和根的生长, 导致植株不育和幼苗死亡(Liu et al.2011). ...
The plant cell wall integrity maintenance mechanism—a case study of a cell wall plasma membrane signaling network 1 2015
... 细胞壁作为抵御病原体的屏障, 其完整性为植物防御病原体所必需(Hamann, 2015).植物细胞壁果胶被细菌和真菌分泌的果胶降解酶降解, 其中HG降解后产生去甲基酯化的果胶片段或寡聚半乳糖醛酸(oligogalacturonides, OGs) (Ferrari et al.2013; 郁有健等, 2014).OGs作为激发子可被植物体内的受体识别, 从而激活一系列防御反应, 起到保护植物的作用.研究表明, OGs的信号转导通过细胞壁关联激酶(wall-associated kinases, WAKs)发挥作用, WAKs能感知OGs, 进而激活植物的应激反应(Brutus et al.2010; Kohorn et al.2014).此外, 病原体入侵会激活PME, PME催化HG去甲基酯化过程中产生具有挥发性的甲醇, 甲醇可作为信号分子参与启动邻近植物的免疫反应, 增强邻近植物对细菌病原体的抵抗力(Komarova et al.2014). ...
ARABINAN DEFICIENT 1 is a putative arabinosyltransferase involved in biosynthesis of pectic arabinan in Arabidopsis 2 2006
... RGI占果胶成分的20%-35%, 与HG和RGII不同, 它的主链由双糖重复链[4)-α-D-GalA-(1,2)-α-L-Rha- (1, ]n组成.RGI主链中的大部分或全部GalA在O-2或O-3处被乙酰化修饰, 大多数鼠李糖(rhamnose, Rha)残基可在O-4处被线性或支链寡糖或多糖取代(Atmodjo et al.2013).RGI侧链包括β-1,4-半乳聚糖、阿拉伯聚糖或阿拉伯半乳聚糖等.RGI生物合成过程中, 其主链和侧链合成所需酶不尽相同, 主链合成需要半乳糖醛酸转移酶(galacturonosyltransferase, GalAT)和鼠李糖基转移酶(rhamnosyltransferase, RhaT)参与, 而侧链合成需要半乳糖基转移酶(galactosyltransferases, GalTs)和阿拉伯糖基转移酶(arabinosyltransferases, AraTs).目前在拟南芥中已有报道的GalTs和AraTs编码基因包括GT92家族的GALS1 (GALACTAN SYNTHASE 1) (Liwanag et al.2012)和GT47家族B亚群中的ARAD1 (ARABINAN DEFICIENT 1) (Harholt et al.2006)及其同源基因ARAD2(Harholt et al.2012).其中GALS1基因功能缺失和过表达分别导致细胞壁半乳聚糖的减少和增加(Liwanag et al.2012).对arad1突变体的生化分析显示, 突变体细胞壁中的阿拉伯糖数量明显低于野生型(Harholt et al.2006); 而对arad2突变体细胞壁的生化分析显示, 其单糖组成与野生型无明显差异, 且过表达ARAD2并不能完全恢复arad1的表型, 说明虽然ARAD1与ARAD2同源, 但是二者功能并不冗余, 推测这2个糖基转移酶可能在二硫键连接的复合物形成中发挥作用(Harholt et al.2012). ...
Requirement for pectin methyl esterase and preference for fragmented over native pectins for wall-associated kinase-activated, eds1/pad4-dependent stress response in Arabidopsis 1 2014
... 细胞壁作为抵御病原体的屏障, 其完整性为植物防御病原体所必需(Hamann, 2015).植物细胞壁果胶被细菌和真菌分泌的果胶降解酶降解, 其中HG降解后产生去甲基酯化的果胶片段或寡聚半乳糖醛酸(oligogalacturonides, OGs) (Ferrari et al.2013; 郁有健等, 2014).OGs作为激发子可被植物体内的受体识别, 从而激活一系列防御反应, 起到保护植物的作用.研究表明, OGs的信号转导通过细胞壁关联激酶(wall-associated kinases, WAKs)发挥作用, WAKs能感知OGs, 进而激活植物的应激反应(Brutus et al.2010; Kohorn et al.2014).此外, 病原体入侵会激活PME, PME催化HG去甲基酯化过程中产生具有挥发性的甲醇, 甲醇可作为信号分子参与启动邻近植物的免疫反应, 增强邻近植物对细菌病原体的抵抗力(Komarova et al.2014). ...
Cell wall methanol as a signal in plant immunity 1 2014
... 细胞壁作为抵御病原体的屏障, 其完整性为植物防御病原体所必需(Hamann, 2015).植物细胞壁果胶被细菌和真菌分泌的果胶降解酶降解, 其中HG降解后产生去甲基酯化的果胶片段或寡聚半乳糖醛酸(oligogalacturonides, OGs) (Ferrari et al.2013; 郁有健等, 2014).OGs作为激发子可被植物体内的受体识别, 从而激活一系列防御反应, 起到保护植物的作用.研究表明, OGs的信号转导通过细胞壁关联激酶(wall-associated kinases, WAKs)发挥作用, WAKs能感知OGs, 进而激活植物的应激反应(Brutus et al.2010; Kohorn et al.2014).此外, 病原体入侵会激活PME, PME催化HG去甲基酯化过程中产生具有挥发性的甲醇, 甲醇可作为信号分子参与启动邻近植物的免疫反应, 增强邻近植物对细菌病原体的抵抗力(Komarova et al.2014). ...
Multi-scale spatial heterogeneity of pectic rhamnogalacturonan I (RG-I) structural features in tobacco seed endosperm cell walls 1 2013
... 对RGI的标记研究表明, 其主链和侧链的抗体标记有所不同, 其中中性侧链可被LM5半乳糖和LM6/ LM13阿拉伯糖抗体探针标记(Wu et al.2016), 主链可被INRA-RU1和INRA-RU2标记(Ralet et al.2010).标记主链的INRA-RU2需要至少2个双糖重复序列才能被结合, INRA-RU1需要至少6个双糖重复才能被结合, 两个抗体的最佳表位均为7个双糖重复序列(Ralet et al.2010).例如, Lee等(2013)使用LM5、LM6和INRA-RU1对烟草(Nicotiana tabacum)种子胚乳细胞壁进行原位分析和色谱检测分析, 研究RGI结构在单个组织中的相对发生情况, 证明RGI聚合物在组织水平和单细胞壁水平上存在空间异质性. ...
Transition of primary to secondary cell wall synthesis 1 2016
... 细胞壁作为植物细胞的屏障, 在维持细胞膨压、完整性以及细胞间信号交流方面具有重要作用(Cosgrove, 2005).细胞壁包括初生细胞壁和次生细胞壁, 均由多糖和少量蛋白组成(Logan et al.2014), 但组成成分和功能有所不同.其中初生细胞壁主要由纤维素、半纤维素和果胶组成(解敏敏等, 2015), 具有可塑性, 可以适应植物细胞生长过程中的动态变化, 对植物细胞的形态建成具有重要作用(Sakamoto et al.2018).次生细胞壁主要由纤维素、半纤维素和木质素组成(Zhong and Ye, 2015), 刚性较强, 可赋予植物强度和硬度(Cosgrove and Jarvis, 2012; Li et al.2016; 张雨等, 2020).初生细胞壁纤维素含量较少, 果胶多糖含量较多, 在双子叶和非禾本科植物中, 果胶约占初生细胞壁的35% (Cosgrove and Jarvis, 2012).次生细胞壁的果胶含量尚不明确, 但有研究表明, 杨树(Populus trichocarpa, P. nigra × P. maximowiczii, P. trichocarpa×P. koreana)成熟木质部中有类果胶(pectin-like)组分在次生细胞壁中沉积(Arend et al.2008). ...
Membrane topology of golgi-localized probable S-adenosylmethionine- dependent methyltransferase in tobacco ( Nicotiana tabacum) BY-2 cells 1 2015
... HG是α-1,4连接的半乳糖醛酸(α-1,4-linked D-galacturonic acid, GalA)线性同聚物, 在果胶中含量最高(Mohnen,2008).HG可以在C-6羧基处进行甲基化, 参与甲基化修饰的酶包括HG半乳糖醛酸转移酶(galacturonosyltransferases, GAUT)和果胶甲基转移酶(pectin methyl-transferase, PMT) (Sebastian et al.2009).在数据库CAZy中检索发现, GAUT蛋白属于糖基转移酶(glycosyltransferases, GT)家族8, 该家族在拟南芥(Arabidopsis thaliana)中有15个成员.其中参与果胶生物合成的酶有GAUT1 (Sterling et al.2006)、GAUT7 (Atmodjo et al.2011)、GAUT4 (Biswal et al.2018)和GAUT8 (QUA1) (Bouton et al.2002).GAUT1是首个被鉴定的HG半乳糖醛酸转移酶, 它能在体外合成聚半乳糖醛酸(Sterling et al.2006); GAUT7是GAUT1的同源蛋白, 二者形成GAUT1:GAUT7复合物, 共同参与细胞壁基质多糖的生物合成(Atmodjo et al.2011); GAUT4可以合成HG, 其表达下调使细胞壁中的HG和RG-II减少(Biswal et al.2018); qua1突变体中HG含量极低, 表明QUA1可能参与果胶的生物合成(Bouton et al.2002).PMT通过催化S-腺苷-L-甲硫氨酸的甲基转移到果胶HG中α-1,4连接的半乳糖醛酸残基的羧基上, 使HG发生甲基化修饰(Liuet al.2015).PMT与GAUT可能作为异源复合体参与HG的聚合, 使HG完全甲基化(即甲基化程度达到80%).此外, HG的一些GalA残基在O-2或O-3处可发生乙酰化修饰(易建勇等, 2020).甲基化和乙酰化的酯化程度是动态可变的, 其修饰的程度影响果胶的理化性质.例如, 在Ca2+的作用下, 两条HG链之间相互作用形成二聚体.值得注意的是, 至少需要9个未甲基化修饰的GalA残基的连续延伸才能实现此同型二聚体(Scheller et al.2007). ...
MALE GAMETOPHYTE DEFECTIVE 4 encodes a rhamnogalacturonan II xylosyltransferase and is important for growth of pollen tubes and roots in Arabidopsis 1 2011
... RGII的结构非常复杂, 由至少12种不同的单糖构成, 其主干是HG, 主干上带有4个不同的侧链(A-D), 链A和B在拟南芥中分别被描述为高度分化的八糖和七糖, 链C和D是双糖(Glushka et al.2003; Pabst et al.2013).值得注意的是, 侧链A和B的单糖组成中都含有芹菜糖残基(apiosyl, Api), 而侧链A的芹菜糖残基是形成硼酸二酯键的关键位点(Funakawa and Miwa, 2015).目前, 对RGII生物合成的研究较少, 已确定参与RGII生物合成的酶有鼠李半乳糖醛酸木糖基转移酶1-4 (rhamno galacturonan xylosyltransferase, RGXT1-4), 它们属于GT77的B亚群, 具有α-1,3-木糖基转移酶活性, 能将UDP-Xyl转移到岩藻糖(fucose, Fuc)上.RGXT1和RGXT2在拟南芥幼苗和成熟植株的营养组织中均有表达, 二者突变体的生长表型与野生型相比均无明显变化(Egelund et al.2006); RGXT3仅在成熟植株的叶片和角质层中表达(Egelund et al.2008); RGXT4在许多器官和组织中都有表达, 其功能缺失会严重影响花粉管和根的生长, 导致植株不育和幼苗死亡(Liu et al.2011). ...
Pectin biosynthesis: GALS1 in Arabidopsis thaliana is a β-1,4-Galactan β-1,4-Galactosyltransferase 2 2012
... RGI占果胶成分的20%-35%, 与HG和RGII不同, 它的主链由双糖重复链[4)-α-D-GalA-(1,2)-α-L-Rha- (1, ]n组成.RGI主链中的大部分或全部GalA在O-2或O-3处被乙酰化修饰, 大多数鼠李糖(rhamnose, Rha)残基可在O-4处被线性或支链寡糖或多糖取代(Atmodjo et al.2013).RGI侧链包括β-1,4-半乳聚糖、阿拉伯聚糖或阿拉伯半乳聚糖等.RGI生物合成过程中, 其主链和侧链合成所需酶不尽相同, 主链合成需要半乳糖醛酸转移酶(galacturonosyltransferase, GalAT)和鼠李糖基转移酶(rhamnosyltransferase, RhaT)参与, 而侧链合成需要半乳糖基转移酶(galactosyltransferases, GalTs)和阿拉伯糖基转移酶(arabinosyltransferases, AraTs).目前在拟南芥中已有报道的GalTs和AraTs编码基因包括GT92家族的GALS1 (GALACTAN SYNTHASE 1) (Liwanag et al.2012)和GT47家族B亚群中的ARAD1 (ARABINAN DEFICIENT 1) (Harholt et al.2006)及其同源基因ARAD2(Harholt et al.2012).其中GALS1基因功能缺失和过表达分别导致细胞壁半乳聚糖的减少和增加(Liwanag et al.2012).对arad1突变体的生化分析显示, 突变体细胞壁中的阿拉伯糖数量明显低于野生型(Harholt et al.2006); 而对arad2突变体细胞壁的生化分析显示, 其单糖组成与野生型无明显差异, 且过表达ARAD2并不能完全恢复arad1的表型, 说明虽然ARAD1与ARAD2同源, 但是二者功能并不冗余, 推测这2个糖基转移酶可能在二硫键连接的复合物形成中发挥作用(Harholt et al.2012). ...
... 2012).对arad1突变体的生化分析显示, 突变体细胞壁中的阿拉伯糖数量明显低于野生型(Harholt et al.2006); 而对arad2突变体细胞壁的生化分析显示, 其单糖组成与野生型无明显差异, 且过表达ARAD2并不能完全恢复arad1的表型, 说明虽然ARAD1与ARAD2同源, 但是二者功能并不冗余, 推测这2个糖基转移酶可能在二硫键连接的复合物形成中发挥作用(Harholt et al.2012). ...
Cell wall, cytoskeleton, and cell expansion in higher plants 1 2014
... 细胞壁作为植物细胞的屏障, 在维持细胞膨压、完整性以及细胞间信号交流方面具有重要作用(Cosgrove, 2005).细胞壁包括初生细胞壁和次生细胞壁, 均由多糖和少量蛋白组成(Logan et al.2014), 但组成成分和功能有所不同.其中初生细胞壁主要由纤维素、半纤维素和果胶组成(解敏敏等, 2015), 具有可塑性, 可以适应植物细胞生长过程中的动态变化, 对植物细胞的形态建成具有重要作用(Sakamoto et al.2018).次生细胞壁主要由纤维素、半纤维素和木质素组成(Zhong and Ye, 2015), 刚性较强, 可赋予植物强度和硬度(Cosgrove and Jarvis, 2012; Li et al.2016; 张雨等, 2020).初生细胞壁纤维素含量较少, 果胶多糖含量较多, 在双子叶和非禾本科植物中, 果胶约占初生细胞壁的35% (Cosgrove and Jarvis, 2012).次生细胞壁的果胶含量尚不明确, 但有研究表明, 杨树(Populus trichocarpa, P. nigra × P. maximowiczii, P. trichocarpa×P. koreana)成熟木质部中有类果胶(pectin-like)组分在次生细胞壁中沉积(Arend et al.2008). ...
Occurrence of the primary cell wall polysaccharide rhamnogalacturonan II in pteridophytes, lycophytes, and bryophytes. Implications for the evolution of vascular plants 1 2004
... RGII结构虽然复杂, 但其结构在植物中高度保守(Matsunaga et al.2004).在植物细胞壁中, 90%以上的RGII以二聚体形式存在.如图2所示, RGII二聚体由侧链A上的芹菜糖残基之间的硼酸二酯键共价交联形成(O’Neillet al.2004; Atmodjo et al.2013), 侧链A的完整结构对RGII二聚体的形成至关重要.当RGII完整性缺失时会影响植物的生长, 导致植物矮化(O’Neillet al.2001)和花粉管伸长缺陷(Delmas et al.2008), 严重时可导致植物细胞死亡(Ahn et al.2006).在拟南芥中, MUR1 (MURUS1)编码GDP-甘露糖-4,6-脱水酶, 催化由GDP-D-mannose到GDP- L-focuse的第1步合成.GDP-L-focuse是L-岩藻糖(L- focuse, 6-deoxy-L-galactose)的活化核苷酸糖形式, L-岩藻糖是RGII侧链A的单糖组成之一, 也是其它多种结构多糖和糖蛋白的组成部分(Zhang et al.2019).MUR1功能缺失突变体mur1植株矮化, RGII交联率降低(O’Neillet al.2001), 可能是由于侧链A截断所致(Pabst et al.2013).进一步研究发现, mur1突变体侧链A中的L-focuse被L-Gal取代后, 全长侧链的比例降低, 导致RGII对硼的亲和力降低, 二聚体形成减少, 造成mur1植株地上部矮化(Reuhs et al.2004).在植物细胞壁中, 硼将2条RGII链连接在一起形成二聚体, 增强了细胞之间的黏附和机械强度.研究发现, 通过补充硼可增加RGII的交联比例, 同时减弱mur1突变体的矮化表型, 表明硼酸盐引起的RGII交联对植物正常生长至关重要(O’Neillet al.2001). ...
Temporal and spatial regulation of pectic (1→4)-β-D-galactan in cell walls of developing pea cotyledons: implications for mechanical properties 2 2000
... RGI侧链包括β-1,4-半乳聚糖、阿拉伯聚糖或阿拉伯半乳聚糖, 其组成和结构影响细胞壁的强度、弹性和柔韧性等机械性能.例如, 侧链阿拉伯聚糖和阿拉伯半乳聚糖的长度和数量部分决定初生细胞壁的相对灵活性, 而半乳聚糖含量增加有助于增强细胞壁的硬度和刚度(Ulvskovet al.2005; Moore et al.2008).Cankar等(2014)通过表型观察和组分分析, 对编码果胶修饰、主链降解和侧链类型等酶的多个转基因马铃薯(Solanum tuberosum)株系进行研究, 发现果胶主链被鼠李半乳糖醛酸裂解酶(rhamno galacturonanlyase, RGL)裂解为片段的转基因株系, 以及阿拉伯聚糖含量降低70%的转基因株系中, 成熟花粉粒的内壁在亲和孔的位置外翻突起, 最终导致花粉粒破裂和细胞质泄漏, 这些结果证实在花粉粒成熟和脱水过程中, 果胶阿拉伯聚糖侧链在细胞壁重塑过程中发挥了重要作用.McCartney等(2000)通过对(1→4)- β-D-半乳聚糖出现前后的豌豆(Pisum sativum cv. ‘Avola’)子叶进行机械压应力测试, 发现细胞壁富含半乳聚糖的子叶硬度明显增强.此外, RGI侧链的组成和结构与细胞和组织的发育阶段密切相关.例如, 在胡萝卜( Daucus carotacv. ‘Early Nantes’)由细胞分裂向细胞伸长过渡时, 伴随着细胞壁果胶半乳聚糖的出现和阿拉伯聚糖减少; 豌豆子叶在发育中随着果胶半乳聚糖的出现表现出组织硬度增强(Willats et al.1999; McCartney et al.2000). ...
... 2000). ...
Pectin methylesterases: cell wall enzymes with important roles in plant physiology 3 2001
... HG在高尔基体中以高甲基酯化的形式合成, 随后由果胶甲基酯酶(pectin methylesterase, PME)选择性地去甲基酯化(Caffall and Mohnen, 2009), 同时释放甲醇和质子(查笑君等, 2010), 生成带负电荷的羧基(Wormit and Usadel, 2018).去甲基酯化的HG可以增强细胞壁的刚性.例如, PMEs催化HG去甲基酯化后可形成连续的GalAs, 在Ca2+存在时, 带负电荷的羧基与Ca2+相互作用, 形成“鸡蛋盒” (Egg Box Model)结构(图1).该结构可以通过增强细胞壁刚性来维持细胞壁的完整性(Micheli, 2001; Braccini and Peréz,2001); 花粉管的顶端区域几乎只存在甲基酯化的果胶, 外源施加PME时可诱导其尖端细胞壁增厚, 从而抑制花粉管生长(Bosch et al.2005).此外, PME去甲基酯化HG的过程中会导致局部pH降低, 激活多种细胞壁降解酶, 从而促进细胞壁的扩张, 使其弹性增强.相反, 当植物体缺少PME时, 细胞壁的弹性降低.研究发现, 果胶甲基酯酶6 (PME6)在保卫细胞中表达量较高, 野生型保卫细胞富含未酯化的HG, 而突变体pme6-1的保卫细胞壁富含甲基酯化的果胶, 其气孔开启/关闭动态范围缩小, 表明异常的果胶甲基酯化改变了保卫细胞的机械特性, 导致气孔无法根据环境信号进行适度的开放与关闭(Amsbury et al.2016). ...
... 交联的“鸡蛋盒”模型(改自Micheli,2001; Braccini and Peréz,2001) 高甲基酯化的HG被果胶甲基酯酶(PME)去甲基酯化, 产生带负电荷的羧基.在Ca2+存在时, 2条HG链的羧基与Ca2+相互作用, 形成“鸡蛋盒”结构. ...
... 高甲基酯化的HG被果胶甲基酯酶(PME)去甲基酯化, 产生带负电荷的羧基.在Ca2+存在时, 2条HG链的羧基与Ca2+相互作用, 形成“鸡蛋盒”结构. An “egg-box” model of homogalacturonan (HG) polysaccharide and Ca <sup>2+</sup> cross-linking (modified from Micheli, <xref ref-type="bibr" rid="b45">2001</xref>; Braccini and Peréz,<xref ref-type="bibr" rid="b16">2001</xref>) Hypermethylated HG is de-methylated by pectin methylesterase (PME) to produce negatively charged carboxyl groups. In the presence of Ca2+, the carboxyl groups of the two HG chains interact with Ca2+ to form an “egg box” structure. ...
Pectin structure and biosynthesis 2 2008
... 果胶组分在高尔基体中合成和修饰, 然后通过共价键相互连接形成果胶网络.果胶的合成过程需要多种酶类参与.已有研究表明, 果胶的合成至少需要67种不同的转移酶, 包括糖基转移酶、甲基转移酶和乙酰转移酶(Mohnen, 2008).这些酶在顺式高尔基体、中间高尔基体和反式高尔基体中参与构建果胶多糖的复杂高级结构(Nebenfuhr and Staehelin, 2001). ...
... HG是α-1,4连接的半乳糖醛酸(α-1,4-linked D-galacturonic acid, GalA)线性同聚物, 在果胶中含量最高(Mohnen,2008).HG可以在C-6羧基处进行甲基化, 参与甲基化修饰的酶包括HG半乳糖醛酸转移酶(galacturonosyltransferases, GAUT)和果胶甲基转移酶(pectin methyl-transferase, PMT) (Sebastian et al.2009).在数据库CAZy中检索发现, GAUT蛋白属于糖基转移酶(glycosyltransferases, GT)家族8, 该家族在拟南芥(Arabidopsis thaliana)中有15个成员.其中参与果胶生物合成的酶有GAUT1 (Sterling et al.2006)、GAUT7 (Atmodjo et al.2011)、GAUT4 (Biswal et al.2018)和GAUT8 (QUA1) (Bouton et al.2002).GAUT1是首个被鉴定的HG半乳糖醛酸转移酶, 它能在体外合成聚半乳糖醛酸(Sterling et al.2006); GAUT7是GAUT1的同源蛋白, 二者形成GAUT1:GAUT7复合物, 共同参与细胞壁基质多糖的生物合成(Atmodjo et al.2011); GAUT4可以合成HG, 其表达下调使细胞壁中的HG和RG-II减少(Biswal et al.2018); qua1突变体中HG含量极低, 表明QUA1可能参与果胶的生物合成(Bouton et al.2002).PMT通过催化S-腺苷-L-甲硫氨酸的甲基转移到果胶HG中α-1,4连接的半乳糖醛酸残基的羧基上, 使HG发生甲基化修饰(Liuet al.2015).PMT与GAUT可能作为异源复合体参与HG的聚合, 使HG完全甲基化(即甲基化程度达到80%).此外, HG的一些GalA残基在O-2或O-3处可发生乙酰化修饰(易建勇等, 2020).甲基化和乙酰化的酯化程度是动态可变的, 其修饰的程度影响果胶的理化性质.例如, 在Ca2+的作用下, 两条HG链之间相互作用形成二聚体.值得注意的是, 至少需要9个未甲基化修饰的GalA残基的连续延伸才能实现此同型二聚体(Scheller et al.2007). ...
High-throughput screening of monoclonal antibodies against plant cell wall glycans by hierarchical clustering of their carbohydrate microarray binding profiles 1 2008
... 果胶多糖原位成像的方法之一是使用糖抗体, 如单克隆抗体(JIM系列和LM系列等)可以高分辨率和高灵敏度地结合特定的多糖结构(Moller et al.2008; Pattathil et al.2010; Ralet et al.2010).使用荧光素标记单克隆抗体, 当抗体与特定的果胶多糖结构结合后形成带有荧光素的复合体, 在荧光显微镜下使用合适的激发光, 即可观察多糖结构的定位与分布(Pattathil et al.2010). ...
A role for pectin-associated arabinans in maintaining the flexibility of the plant cell wall during water deficit stress 1 2008
... RGI侧链包括β-1,4-半乳聚糖、阿拉伯聚糖或阿拉伯半乳聚糖, 其组成和结构影响细胞壁的强度、弹性和柔韧性等机械性能.例如, 侧链阿拉伯聚糖和阿拉伯半乳聚糖的长度和数量部分决定初生细胞壁的相对灵活性, 而半乳聚糖含量增加有助于增强细胞壁的硬度和刚度(Ulvskovet al.2005; Moore et al.2008).Cankar等(2014)通过表型观察和组分分析, 对编码果胶修饰、主链降解和侧链类型等酶的多个转基因马铃薯(Solanum tuberosum)株系进行研究, 发现果胶主链被鼠李半乳糖醛酸裂解酶(rhamno galacturonanlyase, RGL)裂解为片段的转基因株系, 以及阿拉伯聚糖含量降低70%的转基因株系中, 成熟花粉粒的内壁在亲和孔的位置外翻突起, 最终导致花粉粒破裂和细胞质泄漏, 这些结果证实在花粉粒成熟和脱水过程中, 果胶阿拉伯聚糖侧链在细胞壁重塑过程中发挥了重要作用.McCartney等(2000)通过对(1→4)- β-D-半乳聚糖出现前后的豌豆(Pisum sativum cv. ‘Avola’)子叶进行机械压应力测试, 发现细胞壁富含半乳聚糖的子叶硬度明显增强.此外, RGI侧链的组成和结构与细胞和组织的发育阶段密切相关.例如, 在胡萝卜( Daucus carotacv. ‘Early Nantes’)由细胞分裂向细胞伸长过渡时, 伴随着细胞壁果胶半乳聚糖的出现和阿拉伯聚糖减少; 豌豆子叶在发育中随着果胶半乳聚糖的出现表现出组织硬度增强(Willats et al.1999; McCartney et al.2000). ...
Tracking developmentally regulated post-synthetic processing of homogalacturonan and chitin using reciprocal oligosaccharide probes 1 2014
... 在果胶多糖的3种组分中, 对于HG的标记成像研究较多, 用于标记HG的单克隆抗体有JIM5、JIM7和2F4等.其中, JIM5的最佳识别表位可能是4个以上相邻的未酯化GalA残基, 这些残基与甲基酯化的GalA残基相邻或在其两侧(Clausen et al.2003); JIM7识别表位的重要特征是每隔一段残基上都有甲基酯化的GalA残基(Clausen et al.2003); 2F4可结合钙离子交联的未酯化的GalA残基(Sujkowska-Rybkowska and Borucki, 2015).我们用JIM5和JIM7对青扦(Picea wilsonii)花粉管细胞壁中的果胶进行标记, 在激光共聚焦显微镜下用488 nm激发并在500-550 nm处采集发射信号, 可观察到在花粉管细胞壁中, JIM7标记的高甲基酯化的果胶聚集在生长的花粉管顶端, JIM5标记的低甲基酯化的果胶分布在花粉管伸长处(Cui et al.2015).此外, 我们利用JIM5和JIM7免疫标记, 还检测了杉木(Cunninghamia lanceolata)维管形成层细胞壁多糖在季节周期中的变化, 结果显示, 休眠期形成层细胞壁中富含低甲基酯化的HG, 而活跃期形成层细胞壁中高甲基酯化的HG较多(Wu et al.2016).Sujkowska-Rybkowska和Borucki (2015)使用JIM5、JIM7和2F4分别对低甲基酯化果胶、高甲基酯化果胶和钙交联果胶进行免疫标记, 检测铝处理的豌豆根瘤中果胶酯化和分布模式.Mravec等(2014)利用低聚壳聚糖(chitosan oligosaccharides, COS)与去酯化的HG果胶可以特异性结合的特点, 开发了非免疫标记分子探针.应用Alexa Fluor 488偶联COS (COS488)标记去甲基酯化HG, 可以对其分布进行高分辨率荧光成像观察; COS与纳米金粒子耦合(COSAuNP)后, 可以用于在透射电子显微镜下对HG定位进行超微结构水平的观察.与抗体免疫标记相比较, COS488具有穿透速度快和对酯化程度的变化敏感等特点, 可用于活细胞实时成像. ...
Mobile factories: golgi dynamics in plant cells 1 2001
... 果胶组分在高尔基体中合成和修饰, 然后通过共价键相互连接形成果胶网络.果胶的合成过程需要多种酶类参与.已有研究表明, 果胶的合成至少需要67种不同的转移酶, 包括糖基转移酶、甲基转移酶和乙酰转移酶(Mohnen, 2008).这些酶在顺式高尔基体、中间高尔基体和反式高尔基体中参与构建果胶多糖的复杂高级结构(Nebenfuhr and Staehelin, 2001). ...
Requirement of borate cross-linking of cell wall rhamnogalacturonan II for Arabidopsis growth 3 2001
... RGII结构虽然复杂, 但其结构在植物中高度保守(Matsunaga et al.2004).在植物细胞壁中, 90%以上的RGII以二聚体形式存在.如图2所示, RGII二聚体由侧链A上的芹菜糖残基之间的硼酸二酯键共价交联形成(O’Neillet al.2004; Atmodjo et al.2013), 侧链A的完整结构对RGII二聚体的形成至关重要.当RGII完整性缺失时会影响植物的生长, 导致植物矮化(O’Neillet al.2001)和花粉管伸长缺陷(Delmas et al.2008), 严重时可导致植物细胞死亡(Ahn et al.2006).在拟南芥中, MUR1 (MURUS1)编码GDP-甘露糖-4,6-脱水酶, 催化由GDP-D-mannose到GDP- L-focuse的第1步合成.GDP-L-focuse是L-岩藻糖(L- focuse, 6-deoxy-L-galactose)的活化核苷酸糖形式, L-岩藻糖是RGII侧链A的单糖组成之一, 也是其它多种结构多糖和糖蛋白的组成部分(Zhang et al.2019).MUR1功能缺失突变体mur1植株矮化, RGII交联率降低(O’Neillet al.2001), 可能是由于侧链A截断所致(Pabst et al.2013).进一步研究发现, mur1突变体侧链A中的L-focuse被L-Gal取代后, 全长侧链的比例降低, 导致RGII对硼的亲和力降低, 二聚体形成减少, 造成mur1植株地上部矮化(Reuhs et al.2004).在植物细胞壁中, 硼将2条RGII链连接在一起形成二聚体, 增强了细胞之间的黏附和机械强度.研究发现, 通过补充硼可增加RGII的交联比例, 同时减弱mur1突变体的矮化表型, 表明硼酸盐引起的RGII交联对植物正常生长至关重要(O’Neillet al.2001). ...
... 2001), 可能是由于侧链A截断所致(Pabst et al.2013).进一步研究发现, mur1突变体侧链A中的L-focuse被L-Gal取代后, 全长侧链的比例降低, 导致RGII对硼的亲和力降低, 二聚体形成减少, 造成mur1植株地上部矮化(Reuhs et al.2004).在植物细胞壁中, 硼将2条RGII链连接在一起形成二聚体, 增强了细胞之间的黏附和机械强度.研究发现, 通过补充硼可增加RGII的交联比例, 同时减弱mur1突变体的矮化表型, 表明硼酸盐引起的RGII交联对植物正常生长至关重要(O’Neillet al.2001). ...
... 2001). ...
RHAMNOGALACTURONAN II: structure and function of a borate cross-linked cell wall pectic polysaccharide 3 2004
... RGII结构虽然复杂, 但其结构在植物中高度保守(Matsunaga et al.2004).在植物细胞壁中, 90%以上的RGII以二聚体形式存在.如图2所示, RGII二聚体由侧链A上的芹菜糖残基之间的硼酸二酯键共价交联形成(O’Neillet al.2004; Atmodjo et al.2013), 侧链A的完整结构对RGII二聚体的形成至关重要.当RGII完整性缺失时会影响植物的生长, 导致植物矮化(O’Neillet al.2001)和花粉管伸长缺陷(Delmas et al.2008), 严重时可导致植物细胞死亡(Ahn et al.2006).在拟南芥中, MUR1 (MURUS1)编码GDP-甘露糖-4,6-脱水酶, 催化由GDP-D-mannose到GDP- L-focuse的第1步合成.GDP-L-focuse是L-岩藻糖(L- focuse, 6-deoxy-L-galactose)的活化核苷酸糖形式, L-岩藻糖是RGII侧链A的单糖组成之一, 也是其它多种结构多糖和糖蛋白的组成部分(Zhang et al.2019).MUR1功能缺失突变体mur1植株矮化, RGII交联率降低(O’Neillet al.2001), 可能是由于侧链A截断所致(Pabst et al.2013).进一步研究发现, mur1突变体侧链A中的L-focuse被L-Gal取代后, 全长侧链的比例降低, 导致RGII对硼的亲和力降低, 二聚体形成减少, 造成mur1植株地上部矮化(Reuhs et al.2004).在植物细胞壁中, 硼将2条RGII链连接在一起形成二聚体, 增强了细胞之间的黏附和机械强度.研究发现, 通过补充硼可增加RGII的交联比例, 同时减弱mur1突变体的矮化表型, 表明硼酸盐引起的RGII交联对植物正常生长至关重要(O’Neillet al.2001). ...
... 野生型植株中的鼠李半乳糖醛酸聚糖II二聚体由侧链A的芹菜糖残基通过硼酸二酯键共价交联而成.MUR1催化GDP-L-岩藻糖从头合成的第一步, MUR1功能缺失导致GDP-L-岩藻糖合成受阻.因此, mur1突变体中的鼠李半乳糖醛酸聚糖II侧链A被截断, 这一缺陷导致鼠李半乳糖醛酸聚糖II二聚体的形成减少.HG: 同型半乳糖醛酸聚糖; Api: 芹菜糖; Rha: L-鼠李糖; Fuc: L-岩藻糖; GlcA: D-葡萄糖醛酸; Gal: L-半乳糖; GalA: D-半乳糖醛酸; Me xyl: 2-O-甲基-D-木糖 Rhamngalacturonan II (RGII) dimer in Arabidopsis wild-type and mur1 mutants (modified from O’Neill <i>et al.</i> <xref ref-type="bibr" rid="b52">2004</xref>; Atmodjo <i>et al.</i> <xref ref-type="bibr" rid="b10">2013</xref>) In wild-type plants, RGII dimers are formed between Apiosyl residue of side chain A which are cross-linked covalently by the diester borate bonds. MUR1 catalyzes the first step in the de novo synthesis of GDP-L-fucose. The loss of MUR1 function leads to the block of GDP-L-fucose synthesis. Therefore, the RGII side chain A was truncated in mur1 mutants which resulted in reduced formation of RG-II dimers. HG: Homogalacturonan; Api: D-apiosyl; Rha: L-rhamnose; Fuc: L-fucose; GlcA: D-glucuronic acid; Gal: L-galactose; GalA: D-galacturonic acid; Me xyl: 2-O-methyl-D-xylose ...
Rhamnogalacturonan II structure shows variation in the side chains monosaccharide composition and methylation status within and across different plant species 2 2013
... RGII的结构非常复杂, 由至少12种不同的单糖构成, 其主干是HG, 主干上带有4个不同的侧链(A-D), 链A和B在拟南芥中分别被描述为高度分化的八糖和七糖, 链C和D是双糖(Glushka et al.2003; Pabst et al.2013).值得注意的是, 侧链A和B的单糖组成中都含有芹菜糖残基(apiosyl, Api), 而侧链A的芹菜糖残基是形成硼酸二酯键的关键位点(Funakawa and Miwa, 2015).目前, 对RGII生物合成的研究较少, 已确定参与RGII生物合成的酶有鼠李半乳糖醛酸木糖基转移酶1-4 (rhamno galacturonan xylosyltransferase, RGXT1-4), 它们属于GT77的B亚群, 具有α-1,3-木糖基转移酶活性, 能将UDP-Xyl转移到岩藻糖(fucose, Fuc)上.RGXT1和RGXT2在拟南芥幼苗和成熟植株的营养组织中均有表达, 二者突变体的生长表型与野生型相比均无明显变化(Egelund et al.2006); RGXT3仅在成熟植株的叶片和角质层中表达(Egelund et al.2008); RGXT4在许多器官和组织中都有表达, 其功能缺失会严重影响花粉管和根的生长, 导致植株不育和幼苗死亡(Liu et al.2011). ...
... RGII结构虽然复杂, 但其结构在植物中高度保守(Matsunaga et al.2004).在植物细胞壁中, 90%以上的RGII以二聚体形式存在.如图2所示, RGII二聚体由侧链A上的芹菜糖残基之间的硼酸二酯键共价交联形成(O’Neillet al.2004; Atmodjo et al.2013), 侧链A的完整结构对RGII二聚体的形成至关重要.当RGII完整性缺失时会影响植物的生长, 导致植物矮化(O’Neillet al.2001)和花粉管伸长缺陷(Delmas et al.2008), 严重时可导致植物细胞死亡(Ahn et al.2006).在拟南芥中, MUR1 (MURUS1)编码GDP-甘露糖-4,6-脱水酶, 催化由GDP-D-mannose到GDP- L-focuse的第1步合成.GDP-L-focuse是L-岩藻糖(L- focuse, 6-deoxy-L-galactose)的活化核苷酸糖形式, L-岩藻糖是RGII侧链A的单糖组成之一, 也是其它多种结构多糖和糖蛋白的组成部分(Zhang et al.2019).MUR1功能缺失突变体mur1植株矮化, RGII交联率降低(O’Neillet al.2001), 可能是由于侧链A截断所致(Pabst et al.2013).进一步研究发现, mur1突变体侧链A中的L-focuse被L-Gal取代后, 全长侧链的比例降低, 导致RGII对硼的亲和力降低, 二聚体形成减少, 造成mur1植株地上部矮化(Reuhs et al.2004).在植物细胞壁中, 硼将2条RGII链连接在一起形成二聚体, 增强了细胞之间的黏附和机械强度.研究发现, 通过补充硼可增加RGII的交联比例, 同时减弱mur1突变体的矮化表型, 表明硼酸盐引起的RGII交联对植物正常生长至关重要(O’Neillet al.2001). ...
MUR1-mediated cell- wall fucosylation is required for freezing tolerance in Arabidopsis thaliana 1 2019
A comprehensive toolkit of plant cell wall glycan-directed monoclonal antibodies 2 2010
... 果胶多糖原位成像的方法之一是使用糖抗体, 如单克隆抗体(JIM系列和LM系列等)可以高分辨率和高灵敏度地结合特定的多糖结构(Moller et al.2008; Pattathil et al.2010; Ralet et al.2010).使用荧光素标记单克隆抗体, 当抗体与特定的果胶多糖结构结合后形成带有荧光素的复合体, 在荧光显微镜下使用合适的激发光, 即可观察多糖结构的定位与分布(Pattathil et al.2010). ...
... 2010). ...
Monoclonal antibodies to rhamnogalacturonan I backbone 3 2010
... 果胶多糖原位成像的方法之一是使用糖抗体, 如单克隆抗体(JIM系列和LM系列等)可以高分辨率和高灵敏度地结合特定的多糖结构(Moller et al.2008; Pattathil et al.2010; Ralet et al.2010).使用荧光素标记单克隆抗体, 当抗体与特定的果胶多糖结构结合后形成带有荧光素的复合体, 在荧光显微镜下使用合适的激发光, 即可观察多糖结构的定位与分布(Pattathil et al.2010). ...
... 对RGI的标记研究表明, 其主链和侧链的抗体标记有所不同, 其中中性侧链可被LM5半乳糖和LM6/ LM13阿拉伯糖抗体探针标记(Wu et al.2016), 主链可被INRA-RU1和INRA-RU2标记(Ralet et al.2010).标记主链的INRA-RU2需要至少2个双糖重复序列才能被结合, INRA-RU1需要至少6个双糖重复才能被结合, 两个抗体的最佳表位均为7个双糖重复序列(Ralet et al.2010).例如, Lee等(2013)使用LM5、LM6和INRA-RU1对烟草(Nicotiana tabacum)种子胚乳细胞壁进行原位分析和色谱检测分析, 研究RGI结构在单个组织中的相对发生情况, 证明RGI聚合物在组织水平和单细胞壁水平上存在空间异质性. ...
Complete substitution of a secondary cell wall with a primary cell wall in Arabidopsis 1 2018
... 细胞壁作为植物细胞的屏障, 在维持细胞膨压、完整性以及细胞间信号交流方面具有重要作用(Cosgrove, 2005).细胞壁包括初生细胞壁和次生细胞壁, 均由多糖和少量蛋白组成(Logan et al.2014), 但组成成分和功能有所不同.其中初生细胞壁主要由纤维素、半纤维素和果胶组成(解敏敏等, 2015), 具有可塑性, 可以适应植物细胞生长过程中的动态变化, 对植物细胞的形态建成具有重要作用(Sakamoto et al.2018).次生细胞壁主要由纤维素、半纤维素和木质素组成(Zhong and Ye, 2015), 刚性较强, 可赋予植物强度和硬度(Cosgrove and Jarvis, 2012; Li et al.2016; 张雨等, 2020).初生细胞壁纤维素含量较少, 果胶多糖含量较多, 在双子叶和非禾本科植物中, 果胶约占初生细胞壁的35% (Cosgrove and Jarvis, 2012).次生细胞壁的果胶含量尚不明确, 但有研究表明, 杨树(Populus trichocarpa, P. nigra × P. maximowiczii, P. trichocarpa×P. koreana)成熟木质部中有类果胶(pectin-like)组分在次生细胞壁中沉积(Arend et al.2008). ...
Biosynthesis of pectin 1 2007
... HG是α-1,4连接的半乳糖醛酸(α-1,4-linked D-galacturonic acid, GalA)线性同聚物, 在果胶中含量最高(Mohnen,2008).HG可以在C-6羧基处进行甲基化, 参与甲基化修饰的酶包括HG半乳糖醛酸转移酶(galacturonosyltransferases, GAUT)和果胶甲基转移酶(pectin methyl-transferase, PMT) (Sebastian et al.2009).在数据库CAZy中检索发现, GAUT蛋白属于糖基转移酶(glycosyltransferases, GT)家族8, 该家族在拟南芥(Arabidopsis thaliana)中有15个成员.其中参与果胶生物合成的酶有GAUT1 (Sterling et al.2006)、GAUT7 (Atmodjo et al.2011)、GAUT4 (Biswal et al.2018)和GAUT8 (QUA1) (Bouton et al.2002).GAUT1是首个被鉴定的HG半乳糖醛酸转移酶, 它能在体外合成聚半乳糖醛酸(Sterling et al.2006); GAUT7是GAUT1的同源蛋白, 二者形成GAUT1:GAUT7复合物, 共同参与细胞壁基质多糖的生物合成(Atmodjo et al.2011); GAUT4可以合成HG, 其表达下调使细胞壁中的HG和RG-II减少(Biswal et al.2018); qua1突变体中HG含量极低, 表明QUA1可能参与果胶的生物合成(Bouton et al.2002).PMT通过催化S-腺苷-L-甲硫氨酸的甲基转移到果胶HG中α-1,4连接的半乳糖醛酸残基的羧基上, 使HG发生甲基化修饰(Liuet al.2015).PMT与GAUT可能作为异源复合体参与HG的聚合, 使HG完全甲基化(即甲基化程度达到80%).此外, HG的一些GalA残基在O-2或O-3处可发生乙酰化修饰(易建勇等, 2020).甲基化和乙酰化的酯化程度是动态可变的, 其修饰的程度影响果胶的理化性质.例如, 在Ca2+的作用下, 两条HG链之间相互作用形成二聚体.值得注意的是, 至少需要9个未甲基化修饰的GalA残基的连续延伸才能实现此同型二聚体(Scheller et al.2007). ...
Homogalacturonan methyl-esterification and plant development 1 2009
... HG是α-1,4连接的半乳糖醛酸(α-1,4-linked D-galacturonic acid, GalA)线性同聚物, 在果胶中含量最高(Mohnen,2008).HG可以在C-6羧基处进行甲基化, 参与甲基化修饰的酶包括HG半乳糖醛酸转移酶(galacturonosyltransferases, GAUT)和果胶甲基转移酶(pectin methyl-transferase, PMT) (Sebastian et al.2009).在数据库CAZy中检索发现, GAUT蛋白属于糖基转移酶(glycosyltransferases, GT)家族8, 该家族在拟南芥(Arabidopsis thaliana)中有15个成员.其中参与果胶生物合成的酶有GAUT1 (Sterling et al.2006)、GAUT7 (Atmodjo et al.2011)、GAUT4 (Biswal et al.2018)和GAUT8 (QUA1) (Bouton et al.2002).GAUT1是首个被鉴定的HG半乳糖醛酸转移酶, 它能在体外合成聚半乳糖醛酸(Sterling et al.2006); GAUT7是GAUT1的同源蛋白, 二者形成GAUT1:GAUT7复合物, 共同参与细胞壁基质多糖的生物合成(Atmodjo et al.2011); GAUT4可以合成HG, 其表达下调使细胞壁中的HG和RG-II减少(Biswal et al.2018); qua1突变体中HG含量极低, 表明QUA1可能参与果胶的生物合成(Bouton et al.2002).PMT通过催化S-腺苷-L-甲硫氨酸的甲基转移到果胶HG中α-1,4连接的半乳糖醛酸残基的羧基上, 使HG发生甲基化修饰(Liuet al.2015).PMT与GAUT可能作为异源复合体参与HG的聚合, 使HG完全甲基化(即甲基化程度达到80%).此外, HG的一些GalA残基在O-2或O-3处可发生乙酰化修饰(易建勇等, 2020).甲基化和乙酰化的酯化程度是动态可变的, 其修饰的程度影响果胶的理化性质.例如, 在Ca2+的作用下, 两条HG链之间相互作用形成二聚体.值得注意的是, 至少需要9个未甲基化修饰的GalA残基的连续延伸才能实现此同型二聚体(Scheller et al.2007). ...
Functional identification of an Arabidopsis pectin biosynthetic homogalacturonan galacturonosyltransferase 2 2006
... HG是α-1,4连接的半乳糖醛酸(α-1,4-linked D-galacturonic acid, GalA)线性同聚物, 在果胶中含量最高(Mohnen,2008).HG可以在C-6羧基处进行甲基化, 参与甲基化修饰的酶包括HG半乳糖醛酸转移酶(galacturonosyltransferases, GAUT)和果胶甲基转移酶(pectin methyl-transferase, PMT) (Sebastian et al.2009).在数据库CAZy中检索发现, GAUT蛋白属于糖基转移酶(glycosyltransferases, GT)家族8, 该家族在拟南芥(Arabidopsis thaliana)中有15个成员.其中参与果胶生物合成的酶有GAUT1 (Sterling et al.2006)、GAUT7 (Atmodjo et al.2011)、GAUT4 (Biswal et al.2018)和GAUT8 (QUA1) (Bouton et al.2002).GAUT1是首个被鉴定的HG半乳糖醛酸转移酶, 它能在体外合成聚半乳糖醛酸(Sterling et al.2006); GAUT7是GAUT1的同源蛋白, 二者形成GAUT1:GAUT7复合物, 共同参与细胞壁基质多糖的生物合成(Atmodjo et al.2011); GAUT4可以合成HG, 其表达下调使细胞壁中的HG和RG-II减少(Biswal et al.2018); qua1突变体中HG含量极低, 表明QUA1可能参与果胶的生物合成(Bouton et al.2002).PMT通过催化S-腺苷-L-甲硫氨酸的甲基转移到果胶HG中α-1,4连接的半乳糖醛酸残基的羧基上, 使HG发生甲基化修饰(Liuet al.2015).PMT与GAUT可能作为异源复合体参与HG的聚合, 使HG完全甲基化(即甲基化程度达到80%).此外, HG的一些GalA残基在O-2或O-3处可发生乙酰化修饰(易建勇等, 2020).甲基化和乙酰化的酯化程度是动态可变的, 其修饰的程度影响果胶的理化性质.例如, 在Ca2+的作用下, 两条HG链之间相互作用形成二聚体.值得注意的是, 至少需要9个未甲基化修饰的GalA残基的连续延伸才能实现此同型二聚体(Scheller et al.2007). ...
... 2006); GAUT7是GAUT1的同源蛋白, 二者形成GAUT1:GAUT7复合物, 共同参与细胞壁基质多糖的生物合成(Atmodjo et al.2011); GAUT4可以合成HG, 其表达下调使细胞壁中的HG和RG-II减少(Biswal et al.2018); qua1突变体中HG含量极低, 表明QUA1可能参与果胶的生物合成(Bouton et al.2002).PMT通过催化S-腺苷-L-甲硫氨酸的甲基转移到果胶HG中α-1,4连接的半乳糖醛酸残基的羧基上, 使HG发生甲基化修饰(Liuet al.2015).PMT与GAUT可能作为异源复合体参与HG的聚合, 使HG完全甲基化(即甲基化程度达到80%).此外, HG的一些GalA残基在O-2或O-3处可发生乙酰化修饰(易建勇等, 2020).甲基化和乙酰化的酯化程度是动态可变的, 其修饰的程度影响果胶的理化性质.例如, 在Ca2+的作用下, 两条HG链之间相互作用形成二聚体.值得注意的是, 至少需要9个未甲基化修饰的GalA残基的连续延伸才能实现此同型二聚体(Scheller et al.2007). ...
Pectins esterification in the apoplast of aluminum-treated pea root nodules 2 2015
... 在果胶多糖的3种组分中, 对于HG的标记成像研究较多, 用于标记HG的单克隆抗体有JIM5、JIM7和2F4等.其中, JIM5的最佳识别表位可能是4个以上相邻的未酯化GalA残基, 这些残基与甲基酯化的GalA残基相邻或在其两侧(Clausen et al.2003); JIM7识别表位的重要特征是每隔一段残基上都有甲基酯化的GalA残基(Clausen et al.2003); 2F4可结合钙离子交联的未酯化的GalA残基(Sujkowska-Rybkowska and Borucki, 2015).我们用JIM5和JIM7对青扦(Picea wilsonii)花粉管细胞壁中的果胶进行标记, 在激光共聚焦显微镜下用488 nm激发并在500-550 nm处采集发射信号, 可观察到在花粉管细胞壁中, JIM7标记的高甲基酯化的果胶聚集在生长的花粉管顶端, JIM5标记的低甲基酯化的果胶分布在花粉管伸长处(Cui et al.2015).此外, 我们利用JIM5和JIM7免疫标记, 还检测了杉木(Cunninghamia lanceolata)维管形成层细胞壁多糖在季节周期中的变化, 结果显示, 休眠期形成层细胞壁中富含低甲基酯化的HG, 而活跃期形成层细胞壁中高甲基酯化的HG较多(Wu et al.2016).Sujkowska-Rybkowska和Borucki (2015)使用JIM5、JIM7和2F4分别对低甲基酯化果胶、高甲基酯化果胶和钙交联果胶进行免疫标记, 检测铝处理的豌豆根瘤中果胶酯化和分布模式.Mravec等(2014)利用低聚壳聚糖(chitosan oligosaccharides, COS)与去酯化的HG果胶可以特异性结合的特点, 开发了非免疫标记分子探针.应用Alexa Fluor 488偶联COS (COS488)标记去甲基酯化HG, 可以对其分布进行高分辨率荧光成像观察; COS与纳米金粒子耦合(COSAuNP)后, 可以用于在透射电子显微镜下对HG定位进行超微结构水平的观察.与抗体免疫标记相比较, COS488具有穿透速度快和对酯化程度的变化敏感等特点, 可用于活细胞实时成像. ...
Peptidotriazoles on solid phase: [1,2,3]-triazoles by regiospecific copper(I)-catalyzed 1,3-dipolar cycloadditions of terminal alkynes to azides 1 2002
... 细胞壁作为植物细胞的屏障, 在维持细胞膨压、完整性以及细胞间信号交流方面具有重要作用(Cosgrove, 2005).细胞壁包括初生细胞壁和次生细胞壁, 均由多糖和少量蛋白组成(Logan et al.2014), 但组成成分和功能有所不同.其中初生细胞壁主要由纤维素、半纤维素和果胶组成(解敏敏等, 2015), 具有可塑性, 可以适应植物细胞生长过程中的动态变化, 对植物细胞的形态建成具有重要作用(Sakamoto et al.2018).次生细胞壁主要由纤维素、半纤维素和木质素组成(Zhong and Ye, 2015), 刚性较强, 可赋予植物强度和硬度(Cosgrove and Jarvis, 2012; Li et al.2016; 张雨等, 2020).初生细胞壁纤维素含量较少, 果胶多糖含量较多, 在双子叶和非禾本科植物中, 果胶约占初生细胞壁的35% (Cosgrove and Jarvis, 2012).次生细胞壁的果胶含量尚不明确, 但有研究表明, 杨树(Populus trichocarpa, P. nigra × P. maximowiczii, P. trichocarpa×P. koreana)成熟木质部中有类果胶(pectin-like)组分在次生细胞壁中沉积(Arend et al.2008). ...
 ,*
,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT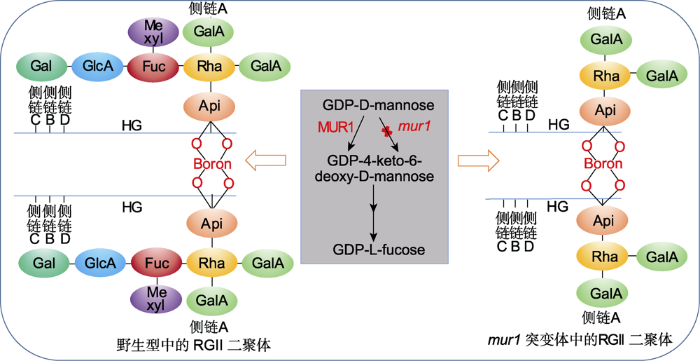 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}