郭宝生, 刘素恩, 赵存鹏, 王兆晓, 王凯辉, 李丹, 刘旭, 杜海英, 耿军义
, * 河北省农林科学院棉花研究所/农业农村部黄淮海半干旱区棉花生物学与遗传育种重点实验室/国家棉花改良中心河北分中心, 石家庄 050051 Heterosis Analysis of the Fiber Quality in Gossypium hirsutum Germplasm Jizi139 with FBP7::iaaM Baosheng Guo, Su’en Liu, Cunpeng Zhao, Zhaoxiao Wang, Kaihui Wang, Dan Li, Xu Liu, Haiying Du, Junyi Geng
, * Institute of Cotton, Hebei Academy of Agriculture and Forestry Sciences/Key Laboratory of Biology and Genetic Improvement of Cotton in Huanghuaihai Semiarid Area, Ministry of Agriculture and Rural Affairs/Hebei Branch of National Cotton Improvement Center, Shijiazhuang 050051, China 通讯作者: * E-mail:
gengjunyi66@126.com 收稿日期: 2020-11-26
接受日期: 2021-03-1
网络出版日期: 2021-03-01
基金资助: 转基因生物新品种培育科技重大专项 .2016ZX08005-005 河北省重点研发计划 .20326316D 河北省农林科学院创新工程 .2019-03-07
Corresponding authors: * E-mail:
gengjunyi66@126.com Received: 2020-11-26
Accepted: 2021-03-1
Online: 2021-03-01
摘要 提高棉花(
Gossypium hirsutum )产量兼顾改良纤维品质是棉花育种的重要目标, 而优良种质创新是品种改良的基础。
FBP7::iaaM 基因能够调控棉花胚珠表皮细胞IAA的含量, 进而促进棉纤维发育的起始。利用含有
FBP7::iaaM 基因的种质IF1-1, 通过常规杂交育种手段实现了目的基因向骨干亲本的转移, 培育了优良陆地棉种质冀资139, 并用4个不同类型的陆地棉品系对冀资139纤维品质性状进行了杂种优势分析。结果表明,
FBP7::iaaM 基因及其调控的优良性状可以在骨干育种亲本中传递, 具有较高的育种价值; 转
FBP7::iaaM 基因的冀资139具有综合性状优良、优质和高衣分等特点; 纤维长度、比强度及马克隆值的遗传主要由基因加性效应控制, 这为其作为骨干亲本的应用提供了理论依据。
关键词: 陆地棉 ;
纤维品质 ;
FBP7::iaaM ;
杂种优势 Abstract The improvement of cotton yield and fiber quality has become an important goal of cotton breeding, and the innovation of elite germplasm is the basis of variety improvement. The
FBP7::iaaM could promote the cotton fiber initiation by regulating the content of IAA in ovule epidermis. Using the transgenic line of IF1-1 (
FBP7::iaaM ) as the parent, we have transferred the target gene to backbone parents and developed an excellent upland cotton germplasm Jizi139 by conventional cross breeding, and analyzed the heterosis of fiber quality traits of Jizi139 by crossing with four different types of upland cotton lines. Our results showed that
FBP7::iaaM gene and the fine traits regulated by
FBP7::iaaM can be transferred to breeding backbone parents with high breeding value. Jizi139 with
FBP7::iaaM showed high lint percentage, high fiber quality and excellent comprehensive characteristics. Also, it was found that additive effect was the genetic basis of the fiber length, fiber strength and micronaire, which provided a theoretical basis for the selection and application of parents.
Keywords: upland cotton ;
fiber quality ;
FBP7::iaaM ;
heterosis PDF (2171KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 郭宝生, 刘素恩, 赵存鹏, 王兆晓, 王凯辉, 李丹, 刘旭, 杜海英, 耿军义 . 转
FBP7::iaaM 基因陆地棉种质冀资139纤维品质性状杂种优势分析 . 植物学报, 2021,
56 (2): 166-174 doi:10.11983/CBB20191
Guo Baosheng, Liu Su’en, Zhao Cunpeng, Wang Zhaoxiao, Wang Kaihui, Li Dan, Liu Xu, Du Haiying, Geng Junyi .
Heterosis Analysis of the Fiber Quality in Gossypium hirsutum Germplasm Jizi139 with FBP7::iaaM .
Bulletin of Botany , 2021,
56 (2): 166-174 doi:10.11983/CBB20191
提高产量并改良纤维品质是陆地棉(
Gossypium hirsutum )育种的重要目标。棉纤维由胚珠表皮细胞发育而来, 而每个胚珠上的纤维数量、长度与强度等因素决定了棉花的产量和纤维品质(毛玮和曹跃芬,
2018 )。胚珠表皮细胞发育成为纤维需经历初始分化、细胞伸长(初生壁合成)、次生壁合成和脱水成熟4个时期, 其中前3个时期主要决定棉纤维的产量和品质(Kim and Triplett,
2001 ; 潘玉欣等,
2005 )。胚珠表皮细胞的生长发育受到一系列基因的调控, 如
GhMYB (Loguercio
et al. 1999 ; Walford
et al. 2012 )、
GhHD-1 (Wan
et al. 2016 )和
GhMML3 _
A12 (李慧琴等,
2019 )。棉纤维发育受植物激素的调控, 如吲哚乙酸(IAA)和赤霉素(GA
3 )可以直接诱导纤维的发生与伸长(王芙蓉等,
2001 ; Zhang
et al. 2011 ; 王国宁等,
2014 ; 唐淑荣等,
2019 )。Zhang等(
2011 )构建了矮牵牛(
Petunia hybrida )花种皮特异启动子
FBP7 驱动的
iaaM 基因, 并导入棉花种质, 获得了转
FBP7::iaaM 基因种质。该种质胚珠上皮细胞初始分化时期的IAA水平较转化受体增高, 并且棉纤维数量显著提高, 进而衣分较受体对照提高, 同时纤维细度也得到了一定程度的改善。
近年来, 许多****围绕转
FBP7::iaaM 基因材料IF1-1在改良棉花产量和纤维品质方面的利用价值进行了探索。研究表明, 棉花转基因材料IF1-1所携带的
FBP7::iaaM 基因及所调控的性状能在杂交和回交过程中稳定遗传, 进而应用于棉花定向改良, 但IF1-1的综合性状亟待改良(丁晓艳等,
2018 )。本研究利用陆地棉与海岛棉(
G. barbadense )和野生瑟伯氏棉(
G. thurberi ) (以下简称陆海瑟)的远缘杂交后代衍生品种冀棉958与IF1-1杂交, 育成携带
FBP7::iaaM 基因、优质、高衣分的综合性状优良的棉花种质冀资139, 并评价新种质纤维品质性状的杂种优势, 旨在探索棉花骨干亲本的选育方法和应用方式。
1 材料与方法 1.1 供试材料 冀资139的母本冀棉958 (
Gossypium hirsutum L.)来源于河北省农林科学院棉花研究所, 父本为转
iaaM 基因棉花种质IF1-1, 由西南大学裴炎教授提供。用于测试冀资139纤维品质性状杂种优势的4个测试品种(系)纤维品质差异明显, 分别为: 946系(P
1 )、05-198 (P
2 )、2658系(P
3 ) (赵存鹏等,
2017 )和646系(P
4 ), 均来源于河北省农林科学院棉花研究所, 系谱及特性见
表1 。由
表1 可知, 5个亲本材料纤维长度、比强度和马克隆值都存在明显差异, 分别代表短绒棉、中绒棉以及中长绒棉纤维品种类型: 纤维长度最大值为32.2 mm, 最小值为26.3 mm; 比强度最大值为31.1 cN/tex, 最小值为24.7 cN/tex; 马克隆值的最大值为6.0, 最小值为4.2。
Table 1 表1 表1 试验材料及其纤维品质特征特性
Table 1
Test materials and their fiber quality characteristics No. Lines Source and characteristics Fiber length Fiber strength Micronaire Uniformity Elongation 1 Jimian958 (Jimian10×(HBT offspring×GK 12))×Jimian22 29.1 32.0 4.6 84.2 6.7 2 IF1-1 iaaM transgenic germplasm of Jimian14 as receptor 30.1 30.3 4.3 84.2 6.5 P1 Line946 Upland cotton variety 26.3 24.7 6.0 82.3 7.0 P2 05-198 Upland cotton variety 27.8 26.6 6.3 83.3 6.5 P3 Line2658 Jimian20×Ji1316 29.6 27.2 5.5 84.6 6.6 P4 Line646 Line571×Line7886, early mature 30.6 30.8 5.5 86.2 6.4 P5 Jizi139 Jimian958×IF1-1 32.2 31.1 4.2 82.4 6.3
新窗口打开 |
下载CSV 1.2 PCR检测 苗期选取各株系的幼嫩叶片按Paterson等(
1993 )的方法提取基因组DNA。鉴定
iaaM 基因的正向引物为: 5'-AAGGTAGCAGTTCTCTCCGC-3'; 反向引物为: 5'-TCGGCTTAGGAACATCCTCC-3'。引物由上海生工公司合成。
PCR反应条件: 94°C预变性5分钟; 94°C变性45秒, 58°C退火30秒, 72°C延伸30秒, 30个循环; 72°C延伸7分钟; 4°C保温。用1%琼脂糖凝胶对PCR产物进行检测, 电压100 V, 电泳15-20分钟。在紫外凝胶成像仪上观察结果并拍照。
1.3 冀资139纤维品质性状杂种优势分析 田间试验在河北省农林科学院小安舍试验站进行。2018年用4个材料(P
1 -P
4 )分别与冀资139 (P
5 )配置棉花杂交组合: P
1 ×P
5 、P
5 ×P
1 、P
2 ×P
5 、P
5 ×P
2 、P
3 ×P
5 、P
5 ×P
3 、P
4 ×P
5 、P
5 ×P
4 。
2019年设立杂交组合比较试验, 杂交组合P
1 ×P
5 、P
2 ×P
5 、P
3 ×P
5 和P
4 ×P
5 随机区组排列, 3次重复, 共12个小区; 亲本P
1 -P
5 、P
5 ×P
1 、P
5 ×P
2 、P
5 ×P
3 、P
5 ×P
4 均为1次重复, 共9个小区。每小区为2行, 行长8 m, 行距0.7 m, 每行植棉25株。
4 月28日播种, 棉田按照常规丰产栽培管理(马宏秀等,
2019 )。棉花采收期, 每小区混收50个正常吐絮的棉铃, 经室内轧花考种后, 皮棉样品送农业农村部棉花品质检测中心检测。检测指标包括上半部平均长度、断裂比强度、马克隆值、整齐度指数和伸长率。
1.4 数据统计分析 用Excel 2010软件整理数据, 并用SPSS v19.0软件进行计算和统计分析, 使用LSD法分析差异显著性。中亲优势(%)=(F-MP)/MP×100%, 其中, MP=(P
1 + P
2 )/2; 超亲优势(%)=(F-HP)/HP×100%, HP代表高值亲本。显著性分析采用小样本成对资料差异显著性测验,
$t=\frac{(\bar{x_{1}}-\bar{x_{2}})-(\mu_{1}-\mu_{2})}{S_{\bar{x_{1}}-\bar{x_{2}}}}$ 为检验统计量, 当
n ≤30时, 从t值的临界值表查出tα临界值, 当观察值t≤临界值tα时, 差异不显著; 当t>tα时, 差异显著。
2 结果与讨论 2.1 冀资139的选育与特征特性 棉花转基因种质IF1-1虽然衣分较高, 马克隆值性状指标较好, 但是高感枯萎病和黄萎病(刘存敬等,
2016 )、不抗鳞翅目害虫且产量低。河北省农林科学院棉花研究所培育的棉花品种冀棉958适合在黄河流域种植, 是陆海瑟远缘杂交后代衍生品种, 具有产量高、适应性广、抗棉铃虫, 并拥有海岛棉抗逆性强(阿曼古丽·买买提阿力等,
2017 )和黄萎病发病指数低等优良特点, 但是衣分偏低、马克隆值偏高。因此, 将陆海瑟远缘杂交后代衍生品种冀棉958与棉花转基因种质IF1-1进行杂交(
图1 ); 在F
2 、F
4 、F
6 和F
7 代利用PCR技术筛选携带
iaaM 基因并具有优良综合性状的单株自交进入下一世代(
图2 ); 不进行
iaaM 基因鉴定的世代混合选择优良株系; F
7 代携带
iaaM 的优良株系经进一步田间观察、选择并繁种后命名为冀资139。具体选育过程见
图1 。冀资139的系谱为: ((冀棉10号×(陆海瑟后代×GK12))×冀棉22)×IF1-1。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1冀资139棉花的选育过程 Figure 1Breeding process of Jizi139 cotton 图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2棉花F2 群体iaaM基因的检测结果 1 -12: F
2 单株; +P: 阳性对照IF1-1; -P: 阴性对照冀棉958
Figure 2The iaaM gene identification in F2 population of cotton 1 -12: F
2 individuals; +P: Positive control IF1-1; -P: Negative control Jimian958
冀资139的生育期为128天, 株型紧凑, 叶片大小中等, 单株结铃14.5个, 衣分含量为42.3%, 铃重5.8 g, 子指11.6 g。2019年经农业农村部棉花品质监督检验测试中心对纤维品质进行检测, 结果显示: 上半部平均长度32.2 mm, 整齐度指数84.0%, 断裂比强度31.1 cN/tex, 伸长率6.5%, 马克隆值4.2。该品种抗枯萎病、耐黄萎病且抗棉铃虫。冀资139比亲本冀棉958的生育期(139天)提早了11天, 铃重增加0.5 g, 衣分提高2%, 上半部平均长度增加3.1 mm, 马克隆值显著降低(
图3 )。与亲本IF1-1相比, 冀资139衣分降低3.5%, 但获得了抗虫性, 提高了黄萎病抗性, 单株成铃也接近常规品种的结铃能力。因此冀资139获得了双亲的优良性状, 成为一个优质、高衣分且综合性状较优的转
FBP7::iaaM 基因的棉花新种质。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3冀资139及其母本冀棉958的棉纤维特征 (A) 冀资139;
(B) 冀棉958
Figure 3Fiber characteristic of Jizi139 and its female parent Jimian958 (A) Jizi139;
(B) Jimian958
2.2 亲本与杂交组合纤维品质分析 每个试验小区收获50铃, 按小区分别轧花后将皮棉样品送交农业农村部棉花品质检测中心检测。设置重复的小区计算相应指标的平均值和标准差, 从检测结果看, 重复间差异不显著, 说明试验地地力均匀, 对试验结果无显著影响, 可以把3次重复的均值与反交结果(1次重复)进行比较。显著性测验采用小样本成对资料差异显著性测验, 当
n =4时, 从t值的临界值表查出t
0.05 和t
0.01 临界值, 当观察值t>临界值t
0.05 时, 差异显著, 当观察值t>临界值t
0.01 时, 差异极显著。比较正、反交杂交组合的纤维品质, 结果表明, 正、反交之间棉花纤维长度、比强度、马克隆值和伸长率均差异不显著(
表2 ), 表明这些性状由核基因控制, 不受母性遗传的影响。亲本P
1 、P
3 、P
4 与P
5 杂交, 其正、反交的纤维整齐度也无显著差异, 但P
2 与P
5 (05-198与冀资139)正、反交产生的棉花纤维整齐度存在显著差异, 说明整齐度性状可能存在母性遗传效应, 但需要进一步验证。
Table 2 表2 表2 冀资139为亲本的正反交组合纤维品质性状比较
Table 2
Comparison of fiber quality between cross-positive combinations and cross-negative combinations of Jizi139 as parent Crosses Fiber length (mm) Fiber strength (cN/tex) Micronaire Uniformity (%) Elongation (%) P1 ×P5 29.27±0.34 28.23±0.19 5.17±0.09 84.27±0.74 6.67±0.05 P5 ×P1 28.9 28.6 5.2 84.6 6.7 t-value 1.885 1.831 0.577 0.772 1.039 P2 ×P5 30.00±0.38 28.50±0.26 5.40±0.00 86.36±0.31 6.77±0.06 P5 ×P2 29.4 29.1 5.4 83.3 6.7 t-value 1.998 2.078 0 4.076* 2.020 P3 ×P5 31.47±0.83 30.27±0.40 5.03±0.15 83.60±0.56 6.80±0.10 P5 ×P3 30.9 29.6 4.8 85.1 6.7 t-value 1.189 1.901 1.655 1.665 1.732 P4 ×P5 31.57±0.45 29.73±0.40 5.00±0.17 84.40±0.89 6.77±0.06 P5 ×P4 31.5 30.7 4.9 84.2 6.8 t-value 0.269 2.100 1.019 0.389 0.866
t0.05,4 =2.776, t0.01,4 =4.604; P1 -P5 见表1 。* 在0.05水平差异显著。 0.05,4 =2.776, t0.01,4 =4.604; P1 -P5 described in Table 1 . * significant difference at 0.05 level. 新窗口打开 |
下载CSV 2.3 冀资139纤维品质性状杂种优势分析 根据冀资139与4个棉花品系配置的正、反交杂交组合结果, 分析了冀资139纤维品质相关性状的杂种优势(
表3 )。结果表明, 纤维长度和纤维比强度这2个性状在后代中既不存在超亲优势也不存在中亲优势。冀资139与任何长度的纤维种质杂交后, 杂种F
1 代纤维长度都接近于两亲本的平均值, 这表明控制纤维长度和纤维比强度的基因存在加性效应。马克隆值在杂种F
1 代没有表现出超亲优势, 只表现出微弱的中亲优势, 且偏向高值亲本。对于纤维整齐度, 在冀资139与低值亲本杂交时, 表现为超亲优势; 而与高值亲本杂交时, 则不存在中亲优势和超亲优势, 而是简单的加性效应。纤维伸长率在P
1 与P
5 杂交组合中不存在超亲优势, 但在P
2 与P
5 、P
3 与P
5 及P
4 与P
5 杂交组合中表现出超亲优势。
Table 3 表3 表3 冀资139纤维品质性状杂种优势分析
Table 3
Heterosis analysis of fiber quality traits in Jizi139 Crossing Fiber length (mm) Fiber strength (cN/tex) Micronaire Uniformity (%) Elongation (%) P1 ×P5 29.27±0.34 28.23±0.19 5.17±0.09 84.27±0.74 6.67±0.05 P1 26.3 24.7 6.0 82.3 7.0 P5 32.2 31.1 4.2 82.4 6.3 MP heterosis (%) 0.02 0.30 0.34 0.58 0.08 Over-parent heterosis (%) -9.10 -9.23 -19.76 2.27 -4.71 P2 ×P5 30.0±0.38 28.5±0.26 5.4±0.00 86.36±0.31 6.77±0.06 P2 27.8 26.6 6.3 83.3 6.5 P5 32.2 31.1 4.2 82.4 6.3 MP heterosis (%) 0.00 -0.30 0.71 1.06 1.45 Over-parent heterosis (%) -6.83 -8.36 -21.43 3.71 4.29 P3 ×P5 31.47±0.83 30.27±0.40 5.03±0.15 83.60±0.56 6.80±0.10 P3 29.6 27.2 5.5 84.6 6.6 P5 32.2 31.1 4.2 82.4 6.3 MP heterosis (%) 0.46 0.96 0.93 0.03 1.36 Over-parent heterosis (%) -2.27 -2.67 -11.19 -1.21 3.17 P4 ×P5 31.57±0.45 29.73±0.40 5.00±0.17 84.40±0.89 6.77±0.06 P4 30.6 30.8 5.5 86.2 6.4 P5 32.2 31.1 4.2 82.4 6.3 MP heterosis (%) 0.14 -0.99 0.77 0.03 1.65 Over-parent heterosis (%) -1.96 -4.41 -11.90 -2.18 5.87
新窗口打开 |
下载CSV 2.4 冀资139主要经济性状杂种优势分析 我们分析了以4个不同类型的棉花品系为母本, 冀资139为父本配置的杂交组合在衣分、铃重、子指、抗病性等经济性状上的杂种优势。结果(
表4 )表明, 以冀资139作为父本, 与其它4个品种杂交, 衣分存在一定的中亲优势, 铃重存在超亲优势; 4个杂交组合的子指都存在降低现象; 亲本及其杂交组合枯萎病指均小于10, 属于抗枯萎病水平, 但是枯萎病指值接近于父本值; 20<黄萎病指<34, 属于耐黄萎病水平, 存在一定的中亲优势, 并偏向于病指低的品种。以上结果表明, 冀资139作为育种亲本, 具有较好的增产潜力和抗病能力。
Table 4 表4 表4 冀资139主要经济性状杂种优势分析
Table 4
Heterosis analysis of main economic traits in Jizi139 Crossing Fiber lint (%) Bell weight (g) Seed index (g) Fusarium wilt index Verticillium wilt index P1 ×P5 41.5 5.8 10.9 4.3 29.0 P1 38.6 5.1 11.3 2.3 29.5 P5 42.8 5.6 11.6 4.6 28.7 MP heterosis (%) 1.97 8.41 -4.80 24.64 1.03 Over-parent heterosis (%) -3.04 3.57 -6.03 -6.52 -0.34 P2 ×P5 40.2 6.2 10.9 4.7 29.5 P2 37.1 6.3 11.1 2.1 30.6 P5 42.8 5.6 11.6 4.6 28.7 MP heterosis (%) 0.63 4.20 -3.96 40.30 4.61 Over-parent heterosis (%) -6.07 -1.59 -6.03 2.17 -1.21 P3 ×P5 42.2 6.1 10.4 10.3 24.3 P3 41.2 5.9 10.1 8.0 23.8 P5 42.8 5.6 11.6 4.6 28.7 MP heterosis (%) 0.48 6.09 -4.15 63.49 -7.43 Over-parent heterosis (%) -1.40 3.39 -10.34 28.75 -15.33 P4 ×P5 40.2 6.2 10.7 4.0 26.3 P4 39.4 5.6 10.9 3.8 21.3 P5 42.8 5.6 11.6 4.6 28.7 MP heterosis (%) -2.19 10.71 -4.89 -4.76 5.20 Over-parent heterosis (%) -6.07 10.71 -7.76 -13.04 -8.36
新窗口打开 |
下载CSV 2.5 讨论 2.5.1 FBP7::iaaM基因的育种潜力 棉花的产量和品质性状均为微效多基因控制的数量性状, 且受加性和显性效应共同控制。通常情况下, 产量性状的显性效应值大于加性效应值, 而品质性状的加性效应值大于显性效应值, 用传统育种方法同步改良棉花产量和品质比较困难。棉纤维的产量和质量直接受植物激素IAA调控, IAA在植物体内通过色氨酸途径产生。色氨酸合成途径主要包括4条支路: 吲哚丙酮途径、色胺途径、吲哚乙腈途径和吲哚乙酰胺途径(王士杰等,
2019 )。目前, 研究得比较清楚的途径是存在于根癌农杆菌(
Agrobacterium tumefaciens )和假单孢菌(
Pseudomonas savastanoi )中的吲哚乙酰胺途径, 色氨酸单加氧酶(iaaM)是催化该途径的限速酶(陈旭升等,
2015 )。Zhang等(
2011 )把
iaaM 基因导入棉花种质后获得转
FBP7::iaaM 种质, 连续4年的田间试验表明, 转
iaaM 基因种质与受体种质相比衣分提高15%, 马克隆值有所降低, 而产量和品质性状无影响。随后, 借助传统育种方法, 利用转
FBP7::iaaM 种质提高了其它棉花种质的衣分比例, 同时降低马克隆值, 而且能保持其它优异性状不变(丁晓艳等,
2018 )。例如, 肖钦之(
2015 )将转
iaaM 基因的IF11与鄂抗10选、TM-1、冀棉14、优质-1及非转基因的IF11进行正、反交, 分析了这些亲本材料和F
1 代的产量和品质等性状, 结果表明, IF11可以显著增加杂种后代的衣分含量, 并对马克隆值有明显的改善作用。刘宏伟等(
2016 )以B0011为受体亲本, 以具有
iaaM 基因的高衣分品系THL作为供体亲本, 获得了具有外源
iaaM 基因、高衣分和低马克隆值的B0011改良株系。通过B0011改良株系将
iaaM 基因导入华杂棉H318, 这个携带外源基因的杂合株系在产量构成因素中除衣分增加外, 其它主要构成因子与非转基因的华杂棉 H318无明显差异。刘存敬等(
2016 )以IF1-1为父本, 分别与16个陆地棉品系杂交, 分析了亲本及杂种后代F
1 和F
2 群体中
iaaM 基因的遗传规律, 同时选育获得了高衣分新种质, 且F
1 代皮棉产量优势值为13.4%。丁晓艳等(
2018 )通过回交将IF1-1材料中的
FBP7::iaaM 基因导入晋棉11, 获得回交后代品系JBC4, JBC4保留了晋棉11的早熟特点, 同时衣分提高了12.8%, 纤维产量增加56.3%, 马克隆值降低10.7%。本研究利用含有
FBP7::iaaM 基因的种质, 通过常规杂交育种手段实现了目的基因向骨干亲本的转移, 培育出优良陆地棉种质冀资139, 其纤维品质目标性状较突出, 综合性状实现了双亲优良特性的互补, 分子检测表明目的基因未丢失。研究结果表明,
FBP7::iaaM 基因及其所调控的优良性状可以在骨干育种亲本中传递, 具有较高的育种价值。
2.5.2 陆地棉纤维品质改良的复杂性 棉纤维由胚珠表皮细胞发育而来, 其发育受到一系列基因调控。棉纤维长度和强度的决定发生在次生壁合成期和脱水成熟期。一些在棉纤维发育过程中发挥重要作用的基因, 诸如各种转录因子基因、参与激素代谢的基因、编码细胞壁蛋白和细胞骨架蛋白的基因、活性氧代谢相关基因、以及参与糖和脂类代谢的基因已有相关报道(Fang
et al. 2017a ; 袁有禄等,
2018 )。棉花全基因组重测序及全基因组关联分析(GWAS)揭示了陆地棉进化过程中棉纤维性状演变规律, 并且发现多个候选位点与纤维品质性状相关联(Fang
et al. 2017b )。利用719份陆地棉材料作为关联分析群体, 研究发现46个显著的SNPs与5个纤维品质性状关联, 涉及612个候选基因, 有2个与纤维长度及强度有关的单倍型, 有163个及120个纤维发育基因分别与纤维长度和强度有关(Fang
et al. 2017b ; Sun
et al. 2017 )。对棉纤维发育相关基因及其调控网络进行深入研究, 发现陆地棉纤维品质改良具有复杂性和难度。
FBP7::iaaM 基因能够调控胚珠表皮细胞IAA的含量, 进而促进棉纤维的发育, 而以转育的综合性状优良、优质、高衣分的冀资139作为亲本, 其F
1 代主要纤维品质性状指标表现为两个亲本的平均值, 并不具备杂种优势, 推测是父母本调控激素的累加效应造成的。本研究在试验过程中仅考虑了
FBP7::iaaM 基因是否丢失及纤维表型数据的检测, 没有同时检测亲本和杂交种胚珠上皮细胞初始分化时期的
IAA 表达水平, 因此未能在分子机制上对不同的纤维发育性状进行阐述, 是本研究设计的缺陷, 我们将在后续的研究中筛选一些与纤维发育相关的关键基因与
FBP7::iaaM 基因一起, 评价
FBP7::iaaM 基因对不同纤维发育的影响。从育种角度看, 要注重目的基因与其它基因间的互作关系, 以及控制相关性状代谢途径潜在调控机制的遗传效应研究。仅依靠单个基因(如
FBP7::iaaM 基因)的利用实现棉花纤维产量和品质的突破, 尚有一定难度。而更应重视优化利用控制某一性状的一系列基因, 并使其在后代稳定传递。本研究仅限于杂种一代的纤维品质性状表现分析, 随后的分离重组世代是否能出现优于双亲性状的单株和株系, 还有待深入研究。
2.5.3 特异种质冀资139在陆地棉纤维品质改良中的应用 本研究利用转
FBP7::iaaM 种质培育出优良陆地棉种质冀资139。冀资139株型紧凑, 衣分为42.3%, 上半部平均长度32.2 mm, 断裂比强度31.5 cN/tex, 马克隆值4.2, 整齐度指数84.0%; 并且抗枯萎病、耐黄萎病和抗棉铃虫。杂种优势分析表明, 冀资139的纤维长度、比强度和马克隆值的遗传主要由加性效应控制。这与已有的纤维长度、比强度和马克隆值遗传规律的研究结果一致(袁有禄等,
2002 ; 殷剑美等,
2003 ; 聂新辉等,
2015 ), 但是该种质在改良纤维品质较差的品种时具有较大的加性效应, 可以根据综合性状互补原则, 提出最佳的亲本选配和后代选择策略, 同时对杂交后代抉选时要注重选择带有纯合目的基因的植株, 从而大幅度提高育种效率。而在对优质纤维品种的进一步改良时, 要充分考虑该种质对优质品种改良的微效性, 可有目的地改良个别性状。陆地棉特异种质冀资139纤维品质以加性效应为遗传基础, 这一结论为骨干亲本的构建和应用提供了理论依据。本研究重点分析了冀资139的纤维品质性状杂种优势, 而其产量性状受衣分、铃重和单株成铃等因素影响, 效应比较复杂, 还需设计更加完备的试验进行揭示。
3 结论 FBP7::iaaM 基因及其所调控的优良性状可以在骨干育种亲本中传递, 具有较高的育种价值。转
FBP7:: iaaM 基因的冀资139具有综合性状优良、优质和高衣分的特点, 这为其作为骨干亲本的应用提供了理论依据。
[1] 阿曼古丽·买买提阿力 , 拉扎提·努尔布拉提 , 高丽丽 , 张巨松 , 田立文 (2017 ). 盐胁迫对海岛棉和陆地棉幼苗生长及生理特性的影响植物学报 52 ,465 -473 . [本文引用: 1] [2] 陈旭升 , 狄佳春 , 赵亮 (2015 ). 转 iaaM 基因对陆地棉主要经济性状的相关效应分析中国棉花 42(6 ),17 -19, 24 . [本文引用: 1] [3] 丁晓艳 , 赵娟 , 钱山山 , 阎星颖 , 裴炎 (2018 ). 利用 FBP7: iaaM 转基因材料同步改良短季棉品种晋棉11纤维产量和品质作物学报 44 ,1152 -1158 . [本文引用: 3] [4] 李慧琴 , 于娅 , 王鹏 , 刘记 , 胡伟 , 鲁丽丽 , 秦文强 (2019 ). 270份陆地棉种质资源农艺性状与品质性状的遗传多样性分析植物遗传资源学报 20 ,903 -910 . [本文引用: 1] [5] 刘存敬 , 江振兴 , 张建宏 , 唐丽媛 , 张素君 , 田海燕 , 李兴河 , 师树新 , 崔瑞敏 , 张香云 (2016 ). 转 iaaM 高衣分棉花种质IF1-1杂种优势分析及育种应用河北农业科学 20(3 ),70 - 74 . [本文引用: 2] [6] 刘宏伟 , 李南南 , 苗玉焕 , 柳仕明 , 聂以春 , 朱龙付 , 张献龙 (2016 ). 利用 FBP:iaaM 改良华杂棉H318产量与纤维品质研究石河子大学学报(自然科学版) 34 ,133 -140 . [本文引用: 1] [7] 马宏秀 , 王开勇 , 张开祥 , 孟春梅 , 安梦洁 (2019 ). 棉粕对盐碱胁迫下棉花生理及生长补偿效应植物学报 54 ,208 - 216 . [本文引用: 1] [8] 毛玮 , 曹跃芬 (2018 ). 棉纤维发育的遗传特性及相关基因的研究进展浙江农林大学学报 35 ,1155 -1165 . [本文引用: 1] [9] 聂新辉 , 尤春源 , 鲍健 , 李晓方 , 惠慧 , 刘洪亮 , 秦江鸿 , 林忠旭 (2015 ). 基于关联分析的新陆早棉花品种农艺和纤维品质性状优异等位基因挖掘中国农业科学 48 ,2891 - 2910 . [本文引用: 1] [10] 潘玉欣 , 马峙英 , 方宣钧 (2005 ). 棉花纤维发育的遗传机制及分子标记河北农业大学学报 28(3 ),6-11 . [本文引用: 1] [11] 唐淑荣 , 魏守军 , 郭瑞林 , 韦京艳 , 孟俊婷 , 杨长琴 (2019 ). 不同熟性棉花品种纤维品质特征分析与评价中国生态农业学报(中英文) 27 ,1564 -1577 . [本文引用: 1] [12] 王芙蓉 , 张军 , 刘勤红 , 张传云 (2001 ). 我国棉花种质创新进展与展望棉花学报 13 ,50 -53 . [本文引用: 1] [13] 王国宁 , 张桂寅 , 吴立强 , 王省芬 , 李志坤 , 张艳 , 吴金华 , 柯会锋 , 孟成生 , 马峙英 (2014 ). 转 iaaM 基因高衣分棉花新种质材料创制中国棉花 41(7 ),27 -31 . [本文引用: 1] [14] 王士杰 , 赵红霞 , 朱继杰 , 和剑涵 , 李妙 , 王国印 , 贾晓昀 (2019 ). IaaM 基因对不同遗传背景棉花品种纤维品质及衣分的影响河北农业大学学报 42(2 ),36 -40 . [本文引用: 1] [15] 肖钦之 (2015 ). 利用转iaaM 基因种质IF-11改良棉花纤维产量与品质的研究. 硕士论文 . 杭州: 浙江大学. pp.11 -28 . [本文引用: 1] [16] 殷剑美 , 武耀廷 , 朱协飞 , 张天真 (2003 ). 陆地棉产量与品质性状的主基因与多基因遗传分析棉花学报 15 ,67 -72 . [本文引用: 1] [17] 袁有禄 , 魏晓文 , 毛树春 , 潘境涛 , 方红曼 , 吕慧颖 , 邓向东 , 葛毅强 , 魏珣 , 杨维才 (2018 ). 棉花育种行业创新与进展植物遗传资源学报 19 ,455 -463 . [本文引用: 1] [18] 袁有禄 , 张天真 , 郭旺珍 , Yu J Kohel RJ 2002 ). 棉花高品质纤维性状的主基因与多基因遗传分析遗传学报 29 ,827 -834 . [本文引用: 1] [19] 赵存鹏 , 王兆晓 , 王凯辉 , 刘素恩 , 耿军义 , 郭宝生 (2017 ). 胞质雄性不育系冀2658A细胞质对陆地棉主要性状的影响植物学报 52 ,560 -567 . [本文引用: 1] [20] Fang L Gong H Hu Y Liu CX Zhou BL Huang T Wang YK Chen SQ Fang DD Du XM Chen H Chen JD Wang S Wang Q Wan Q Liu BL Pan MQ Chang LJ Wu HT Mei GF Xiang D Li XH Cai CP Zhu XF Chen ZJ Han B Chen XY Guo WZ Zhang TZ Huang XH 2017a ). Genomic insights into divergence and dual domestication of cultivated allotetraploid cottonsGenome Biol 18 ,33 . [本文引用: 1] [21] Fang L Wang Q Hu Y Jia YH Chen JD Liu BL Zhang ZY Guan XY Chen SQ Zhou BL Mei GF Sun JL Pan ZE He SP Xiao SH Shi WJ Gong WF Liu JG Ma J Cai CP Zhu XF Guo WZ Du XM Zhang TZ 2017b ). Genomic analyses in cotton identify signatures of selection and loci associated with fiber quality and yield traitsNat Genet 49 ,1089 -1098 . [本文引用: 2] [22] Kim HJ Triplett BA 2001 ). Cotton fiber growth in planta and in vitro . Models for plant cell elongation and cell wall biogenesisPlant Physiol 127 ,1361 -1366 . [本文引用: 1] [23] Loguercio LL Zhang JQ Wilkins TA 1999 ). Differential regulation of six novel MYB-domain genes defines two distinct expression patterns in allotetraploid cotton ( Gossypium hirsutum L.)Mol Gen Genet 261 ,660 -671 . [本文引用: 1] [24] Paterson AH Brubaker CL Wendel JF 1993 ). A rapid method for extraction of cotton ( Gossypium spp.) genomic DNA suitable for RFLP or PCR analysisPlant Mol Biol Rep 11 ,122 -127 . [本文引用: 1] [25] Sun ZW Wang XF Liu ZW Gu QS Zhang Y Li ZK Ke HF Yang J Wu JH Wu LQ Zhang GY Zhang CY Ma ZY 2017 ). Genome-wide association study discovered genetic variation and candidate genes of fibre quality traits in Gossypium hirsutum LPlant Biotechnol J 15 ,982 -996 . [本文引用: 1] [26] Walford SA Wu YR Llewellyn DJ Dennis ES 2012 ). Epidermal cell differentiation in cotton mediated by the homeodomain leucine zipper gene, GhHD-1 Plant J 71 ,464 -478 . [本文引用: 1] [27] Wan Q Guan XY Yang NN Wu HT Pan MQ Liu BL Fang L Yang SP Hu Y Ye WX Zhang H Ma PY Chen JD Wang Q Mei GF Cai CP Yang DL Wang JW Guo WZ Zhang WH Chen XY Zhang TZ 2016 ). Small interfering RNAs from bidirectional transcripts of GhMML3_ A12 regulate cotton fiber developmentNew Phytol 210 ,1298 -1310 . [本文引用: 1] [28] Zhang M Zheng XL Song SQ Zeng QW Hou L Li DM Zhao J Wei Y Li XB Luo M Xiao YH Luo XY Zhang JF Xiang CB Pei Y 2011 ). Spatiotemporal manipulation of auxin biosynthesis in cotton ovule epidermal cells enhances fiber yield and qualityNat Biotechnol 29 ,453 -458 . [本文引用: 3] 盐胁迫对海岛棉和陆地棉幼苗生长及生理特性的影响
1
2017
... 棉花转基因种质IF1-1虽然衣分较高, 马克隆值性状指标较好, 但是高感枯萎病和黄萎病(刘存敬等,
2016 )、不抗鳞翅目害虫且产量低.河北省农林科学院棉花研究所培育的棉花品种冀棉958适合在黄河流域种植, 是陆海瑟远缘杂交后代衍生品种, 具有产量高、适应性广、抗棉铃虫, 并拥有海岛棉抗逆性强(阿曼古丽·买买提阿力等,
2017 )和黄萎病发病指数低等优良特点, 但是衣分偏低、马克隆值偏高.因此, 将陆海瑟远缘杂交后代衍生品种冀棉958与棉花转基因种质IF1-1进行杂交(
图1 ); 在F
2 、F
4 、F
6 和F
7 代利用PCR技术筛选携带
iaaM 基因并具有优良综合性状的单株自交进入下一世代(
图2 ); 不进行
iaaM 基因鉴定的世代混合选择优良株系; F
7 代携带
iaaM 的优良株系经进一步田间观察、选择并繁种后命名为冀资139.具体选育过程见
图1 .冀资139的系谱为: ((冀棉10号×(陆海瑟后代×GK12))×冀棉22)×IF1-1. ...
转
iaaM 基因对陆地棉主要经济性状的相关效应分析
1
2015
... 棉花的产量和品质性状均为微效多基因控制的数量性状, 且受加性和显性效应共同控制.通常情况下, 产量性状的显性效应值大于加性效应值, 而品质性状的加性效应值大于显性效应值, 用传统育种方法同步改良棉花产量和品质比较困难.棉纤维的产量和质量直接受植物激素IAA调控, IAA在植物体内通过色氨酸途径产生.色氨酸合成途径主要包括4条支路: 吲哚丙酮途径、色胺途径、吲哚乙腈途径和吲哚乙酰胺途径(王士杰等,
2019 ).目前, 研究得比较清楚的途径是存在于根癌农杆菌(
Agrobacterium tumefaciens )和假单孢菌(
Pseudomonas savastanoi )中的吲哚乙酰胺途径, 色氨酸单加氧酶(iaaM)是催化该途径的限速酶(陈旭升等,
2015 ).Zhang等(
2011 )把
iaaM 基因导入棉花种质后获得转
FBP7::iaaM 种质, 连续4年的田间试验表明, 转
iaaM 基因种质与受体种质相比衣分提高15%, 马克隆值有所降低, 而产量和品质性状无影响.随后, 借助传统育种方法, 利用转
FBP7::iaaM 种质提高了其它棉花种质的衣分比例, 同时降低马克隆值, 而且能保持其它优异性状不变(丁晓艳等,
2018 ).例如, 肖钦之(
2015 )将转
iaaM 基因的IF11与鄂抗10选、TM-1、冀棉14、优质-1及非转基因的IF11进行正、反交, 分析了这些亲本材料和F
1 代的产量和品质等性状, 结果表明, IF11可以显著增加杂种后代的衣分含量, 并对马克隆值有明显的改善作用.刘宏伟等(
2016 )以B0011为受体亲本, 以具有
iaaM 基因的高衣分品系THL作为供体亲本, 获得了具有外源
iaaM 基因、高衣分和低马克隆值的B0011改良株系.通过B0011改良株系将
iaaM 基因导入华杂棉H318, 这个携带外源基因的杂合株系在产量构成因素中除衣分增加外, 其它主要构成因子与非转基因的华杂棉 H318无明显差异.刘存敬等(
2016 )以IF1-1为父本, 分别与16个陆地棉品系杂交, 分析了亲本及杂种后代F
1 和F
2 群体中
iaaM 基因的遗传规律, 同时选育获得了高衣分新种质, 且F
1 代皮棉产量优势值为13.4%.丁晓艳等(
2018 )通过回交将IF1-1材料中的
FBP7::iaaM 基因导入晋棉11, 获得回交后代品系JBC4, JBC4保留了晋棉11的早熟特点, 同时衣分提高了12.8%, 纤维产量增加56.3%, 马克隆值降低10.7%.本研究利用含有
FBP7::iaaM 基因的种质, 通过常规杂交育种手段实现了目的基因向骨干亲本的转移, 培育出优良陆地棉种质冀资139, 其纤维品质目标性状较突出, 综合性状实现了双亲优良特性的互补, 分子检测表明目的基因未丢失.研究结果表明,
FBP7::iaaM 基因及其所调控的优良性状可以在骨干育种亲本中传递, 具有较高的育种价值. ...
利用
FBP7: iaaM 转基因材料同步改良短季棉品种晋棉11纤维产量和品质
3
2018
... 近年来, 许多****围绕转
FBP7::iaaM 基因材料IF1-1在改良棉花产量和纤维品质方面的利用价值进行了探索.研究表明, 棉花转基因材料IF1-1所携带的
FBP7::iaaM 基因及所调控的性状能在杂交和回交过程中稳定遗传, 进而应用于棉花定向改良, 但IF1-1的综合性状亟待改良(丁晓艳等,
2018 ).本研究利用陆地棉与海岛棉(
G. barbadense )和野生瑟伯氏棉(
G. thurberi ) (以下简称陆海瑟)的远缘杂交后代衍生品种冀棉958与IF1-1杂交, 育成携带
FBP7::iaaM 基因、优质、高衣分的综合性状优良的棉花种质冀资139, 并评价新种质纤维品质性状的杂种优势, 旨在探索棉花骨干亲本的选育方法和应用方式. ...
... 棉花的产量和品质性状均为微效多基因控制的数量性状, 且受加性和显性效应共同控制.通常情况下, 产量性状的显性效应值大于加性效应值, 而品质性状的加性效应值大于显性效应值, 用传统育种方法同步改良棉花产量和品质比较困难.棉纤维的产量和质量直接受植物激素IAA调控, IAA在植物体内通过色氨酸途径产生.色氨酸合成途径主要包括4条支路: 吲哚丙酮途径、色胺途径、吲哚乙腈途径和吲哚乙酰胺途径(王士杰等,
2019 ).目前, 研究得比较清楚的途径是存在于根癌农杆菌(
Agrobacterium tumefaciens )和假单孢菌(
Pseudomonas savastanoi )中的吲哚乙酰胺途径, 色氨酸单加氧酶(iaaM)是催化该途径的限速酶(陈旭升等,
2015 ).Zhang等(
2011 )把
iaaM 基因导入棉花种质后获得转
FBP7::iaaM 种质, 连续4年的田间试验表明, 转
iaaM 基因种质与受体种质相比衣分提高15%, 马克隆值有所降低, 而产量和品质性状无影响.随后, 借助传统育种方法, 利用转
FBP7::iaaM 种质提高了其它棉花种质的衣分比例, 同时降低马克隆值, 而且能保持其它优异性状不变(丁晓艳等,
2018 ).例如, 肖钦之(
2015 )将转
iaaM 基因的IF11与鄂抗10选、TM-1、冀棉14、优质-1及非转基因的IF11进行正、反交, 分析了这些亲本材料和F
1 代的产量和品质等性状, 结果表明, IF11可以显著增加杂种后代的衣分含量, 并对马克隆值有明显的改善作用.刘宏伟等(
2016 )以B0011为受体亲本, 以具有
iaaM 基因的高衣分品系THL作为供体亲本, 获得了具有外源
iaaM 基因、高衣分和低马克隆值的B0011改良株系.通过B0011改良株系将
iaaM 基因导入华杂棉H318, 这个携带外源基因的杂合株系在产量构成因素中除衣分增加外, 其它主要构成因子与非转基因的华杂棉 H318无明显差异.刘存敬等(
2016 )以IF1-1为父本, 分别与16个陆地棉品系杂交, 分析了亲本及杂种后代F
1 和F
2 群体中
iaaM 基因的遗传规律, 同时选育获得了高衣分新种质, 且F
1 代皮棉产量优势值为13.4%.丁晓艳等(
2018 )通过回交将IF1-1材料中的
FBP7::iaaM 基因导入晋棉11, 获得回交后代品系JBC4, JBC4保留了晋棉11的早熟特点, 同时衣分提高了12.8%, 纤维产量增加56.3%, 马克隆值降低10.7%.本研究利用含有
FBP7::iaaM 基因的种质, 通过常规杂交育种手段实现了目的基因向骨干亲本的转移, 培育出优良陆地棉种质冀资139, 其纤维品质目标性状较突出, 综合性状实现了双亲优良特性的互补, 分子检测表明目的基因未丢失.研究结果表明,
FBP7::iaaM 基因及其所调控的优良性状可以在骨干育种亲本中传递, 具有较高的育种价值. ...
... 代皮棉产量优势值为13.4%.丁晓艳等(
2018 )通过回交将IF1-1材料中的
FBP7::iaaM 基因导入晋棉11, 获得回交后代品系JBC4, JBC4保留了晋棉11的早熟特点, 同时衣分提高了12.8%, 纤维产量增加56.3%, 马克隆值降低10.7%.本研究利用含有
FBP7::iaaM 基因的种质, 通过常规杂交育种手段实现了目的基因向骨干亲本的转移, 培育出优良陆地棉种质冀资139, 其纤维品质目标性状较突出, 综合性状实现了双亲优良特性的互补, 分子检测表明目的基因未丢失.研究结果表明,
FBP7::iaaM 基因及其所调控的优良性状可以在骨干育种亲本中传递, 具有较高的育种价值. ...
270份陆地棉种质资源农艺性状与品质性状的遗传多样性分析
1
2019
... 提高产量并改良纤维品质是陆地棉(
Gossypium hirsutum )育种的重要目标.棉纤维由胚珠表皮细胞发育而来, 而每个胚珠上的纤维数量、长度与强度等因素决定了棉花的产量和纤维品质(毛玮和曹跃芬,
2018 ).胚珠表皮细胞发育成为纤维需经历初始分化、细胞伸长(初生壁合成)、次生壁合成和脱水成熟4个时期, 其中前3个时期主要决定棉纤维的产量和品质(Kim and Triplett,
2001 ; 潘玉欣等,
2005 ).胚珠表皮细胞的生长发育受到一系列基因的调控, 如
GhMYB (Loguercio
et al. 1999 ; Walford
et al. 2012 )、
GhHD-1 (Wan
et al. 2016 )和
GhMML3 _
A12 (李慧琴等,
2019 ).棉纤维发育受植物激素的调控, 如吲哚乙酸(IAA)和赤霉素(GA
3 )可以直接诱导纤维的发生与伸长(王芙蓉等,
2001 ; Zhang
et al. 2011 ; 王国宁等,
2014 ; 唐淑荣等,
2019 ).Zhang等(
2011 )构建了矮牵牛(
Petunia hybrida )花种皮特异启动子
FBP7 驱动的
iaaM 基因, 并导入棉花种质, 获得了转
FBP7::iaaM 基因种质.该种质胚珠上皮细胞初始分化时期的IAA水平较转化受体增高, 并且棉纤维数量显著提高, 进而衣分较受体对照提高, 同时纤维细度也得到了一定程度的改善. ...
转
iaaM 高衣分棉花种质IF1-1杂种优势分析及育种应用
2
2016
... 棉花转基因种质IF1-1虽然衣分较高, 马克隆值性状指标较好, 但是高感枯萎病和黄萎病(刘存敬等,
2016 )、不抗鳞翅目害虫且产量低.河北省农林科学院棉花研究所培育的棉花品种冀棉958适合在黄河流域种植, 是陆海瑟远缘杂交后代衍生品种, 具有产量高、适应性广、抗棉铃虫, 并拥有海岛棉抗逆性强(阿曼古丽·买买提阿力等,
2017 )和黄萎病发病指数低等优良特点, 但是衣分偏低、马克隆值偏高.因此, 将陆海瑟远缘杂交后代衍生品种冀棉958与棉花转基因种质IF1-1进行杂交(
图1 ); 在F
2 、F
4 、F
6 和F
7 代利用PCR技术筛选携带
iaaM 基因并具有优良综合性状的单株自交进入下一世代(
图2 ); 不进行
iaaM 基因鉴定的世代混合选择优良株系; F
7 代携带
iaaM 的优良株系经进一步田间观察、选择并繁种后命名为冀资139.具体选育过程见
图1 .冀资139的系谱为: ((冀棉10号×(陆海瑟后代×GK12))×冀棉22)×IF1-1. ...
... 棉花的产量和品质性状均为微效多基因控制的数量性状, 且受加性和显性效应共同控制.通常情况下, 产量性状的显性效应值大于加性效应值, 而品质性状的加性效应值大于显性效应值, 用传统育种方法同步改良棉花产量和品质比较困难.棉纤维的产量和质量直接受植物激素IAA调控, IAA在植物体内通过色氨酸途径产生.色氨酸合成途径主要包括4条支路: 吲哚丙酮途径、色胺途径、吲哚乙腈途径和吲哚乙酰胺途径(王士杰等,
2019 ).目前, 研究得比较清楚的途径是存在于根癌农杆菌(
Agrobacterium tumefaciens )和假单孢菌(
Pseudomonas savastanoi )中的吲哚乙酰胺途径, 色氨酸单加氧酶(iaaM)是催化该途径的限速酶(陈旭升等,
2015 ).Zhang等(
2011 )把
iaaM 基因导入棉花种质后获得转
FBP7::iaaM 种质, 连续4年的田间试验表明, 转
iaaM 基因种质与受体种质相比衣分提高15%, 马克隆值有所降低, 而产量和品质性状无影响.随后, 借助传统育种方法, 利用转
FBP7::iaaM 种质提高了其它棉花种质的衣分比例, 同时降低马克隆值, 而且能保持其它优异性状不变(丁晓艳等,
2018 ).例如, 肖钦之(
2015 )将转
iaaM 基因的IF11与鄂抗10选、TM-1、冀棉14、优质-1及非转基因的IF11进行正、反交, 分析了这些亲本材料和F
1 代的产量和品质等性状, 结果表明, IF11可以显著增加杂种后代的衣分含量, 并对马克隆值有明显的改善作用.刘宏伟等(
2016 )以B0011为受体亲本, 以具有
iaaM 基因的高衣分品系THL作为供体亲本, 获得了具有外源
iaaM 基因、高衣分和低马克隆值的B0011改良株系.通过B0011改良株系将
iaaM 基因导入华杂棉H318, 这个携带外源基因的杂合株系在产量构成因素中除衣分增加外, 其它主要构成因子与非转基因的华杂棉 H318无明显差异.刘存敬等(
2016 )以IF1-1为父本, 分别与16个陆地棉品系杂交, 分析了亲本及杂种后代F
1 和F
2 群体中
iaaM 基因的遗传规律, 同时选育获得了高衣分新种质, 且F
1 代皮棉产量优势值为13.4%.丁晓艳等(
2018 )通过回交将IF1-1材料中的
FBP7::iaaM 基因导入晋棉11, 获得回交后代品系JBC4, JBC4保留了晋棉11的早熟特点, 同时衣分提高了12.8%, 纤维产量增加56.3%, 马克隆值降低10.7%.本研究利用含有
FBP7::iaaM 基因的种质, 通过常规杂交育种手段实现了目的基因向骨干亲本的转移, 培育出优良陆地棉种质冀资139, 其纤维品质目标性状较突出, 综合性状实现了双亲优良特性的互补, 分子检测表明目的基因未丢失.研究结果表明,
FBP7::iaaM 基因及其所调控的优良性状可以在骨干育种亲本中传递, 具有较高的育种价值. ...
利用
FBP:iaaM 改良华杂棉H318产量与纤维品质研究
1
2016
... 棉花的产量和品质性状均为微效多基因控制的数量性状, 且受加性和显性效应共同控制.通常情况下, 产量性状的显性效应值大于加性效应值, 而品质性状的加性效应值大于显性效应值, 用传统育种方法同步改良棉花产量和品质比较困难.棉纤维的产量和质量直接受植物激素IAA调控, IAA在植物体内通过色氨酸途径产生.色氨酸合成途径主要包括4条支路: 吲哚丙酮途径、色胺途径、吲哚乙腈途径和吲哚乙酰胺途径(王士杰等,
2019 ).目前, 研究得比较清楚的途径是存在于根癌农杆菌(
Agrobacterium tumefaciens )和假单孢菌(
Pseudomonas savastanoi )中的吲哚乙酰胺途径, 色氨酸单加氧酶(iaaM)是催化该途径的限速酶(陈旭升等,
2015 ).Zhang等(
2011 )把
iaaM 基因导入棉花种质后获得转
FBP7::iaaM 种质, 连续4年的田间试验表明, 转
iaaM 基因种质与受体种质相比衣分提高15%, 马克隆值有所降低, 而产量和品质性状无影响.随后, 借助传统育种方法, 利用转
FBP7::iaaM 种质提高了其它棉花种质的衣分比例, 同时降低马克隆值, 而且能保持其它优异性状不变(丁晓艳等,
2018 ).例如, 肖钦之(
2015 )将转
iaaM 基因的IF11与鄂抗10选、TM-1、冀棉14、优质-1及非转基因的IF11进行正、反交, 分析了这些亲本材料和F
1 代的产量和品质等性状, 结果表明, IF11可以显著增加杂种后代的衣分含量, 并对马克隆值有明显的改善作用.刘宏伟等(
2016 )以B0011为受体亲本, 以具有
iaaM 基因的高衣分品系THL作为供体亲本, 获得了具有外源
iaaM 基因、高衣分和低马克隆值的B0011改良株系.通过B0011改良株系将
iaaM 基因导入华杂棉H318, 这个携带外源基因的杂合株系在产量构成因素中除衣分增加外, 其它主要构成因子与非转基因的华杂棉 H318无明显差异.刘存敬等(
2016 )以IF1-1为父本, 分别与16个陆地棉品系杂交, 分析了亲本及杂种后代F
1 和F
2 群体中
iaaM 基因的遗传规律, 同时选育获得了高衣分新种质, 且F
1 代皮棉产量优势值为13.4%.丁晓艳等(
2018 )通过回交将IF1-1材料中的
FBP7::iaaM 基因导入晋棉11, 获得回交后代品系JBC4, JBC4保留了晋棉11的早熟特点, 同时衣分提高了12.8%, 纤维产量增加56.3%, 马克隆值降低10.7%.本研究利用含有
FBP7::iaaM 基因的种质, 通过常规杂交育种手段实现了目的基因向骨干亲本的转移, 培育出优良陆地棉种质冀资139, 其纤维品质目标性状较突出, 综合性状实现了双亲优良特性的互补, 分子检测表明目的基因未丢失.研究结果表明,
FBP7::iaaM 基因及其所调控的优良性状可以在骨干育种亲本中传递, 具有较高的育种价值. ...
棉粕对盐碱胁迫下棉花生理及生长补偿效应
1
2019
... 4 月28日播种, 棉田按照常规丰产栽培管理(马宏秀等,
2019 ).棉花采收期, 每小区混收50个正常吐絮的棉铃, 经室内轧花考种后, 皮棉样品送农业农村部棉花品质检测中心检测.检测指标包括上半部平均长度、断裂比强度、马克隆值、整齐度指数和伸长率. ...
棉纤维发育的遗传特性及相关基因的研究进展
1
2018
... 提高产量并改良纤维品质是陆地棉(
Gossypium hirsutum )育种的重要目标.棉纤维由胚珠表皮细胞发育而来, 而每个胚珠上的纤维数量、长度与强度等因素决定了棉花的产量和纤维品质(毛玮和曹跃芬,
2018 ).胚珠表皮细胞发育成为纤维需经历初始分化、细胞伸长(初生壁合成)、次生壁合成和脱水成熟4个时期, 其中前3个时期主要决定棉纤维的产量和品质(Kim and Triplett,
2001 ; 潘玉欣等,
2005 ).胚珠表皮细胞的生长发育受到一系列基因的调控, 如
GhMYB (Loguercio
et al. 1999 ; Walford
et al. 2012 )、
GhHD-1 (Wan
et al. 2016 )和
GhMML3 _
A12 (李慧琴等,
2019 ).棉纤维发育受植物激素的调控, 如吲哚乙酸(IAA)和赤霉素(GA
3 )可以直接诱导纤维的发生与伸长(王芙蓉等,
2001 ; Zhang
et al. 2011 ; 王国宁等,
2014 ; 唐淑荣等,
2019 ).Zhang等(
2011 )构建了矮牵牛(
Petunia hybrida )花种皮特异启动子
FBP7 驱动的
iaaM 基因, 并导入棉花种质, 获得了转
FBP7::iaaM 基因种质.该种质胚珠上皮细胞初始分化时期的IAA水平较转化受体增高, 并且棉纤维数量显著提高, 进而衣分较受体对照提高, 同时纤维细度也得到了一定程度的改善. ...
基于关联分析的新陆早棉花品种农艺和纤维品质性状优异等位基因挖掘
1
2015
... 本研究利用转
FBP7::iaaM 种质培育出优良陆地棉种质冀资139.冀资139株型紧凑, 衣分为42.3%, 上半部平均长度32.2 mm, 断裂比强度31.5 cN/tex, 马克隆值4.2, 整齐度指数84.0%; 并且抗枯萎病、耐黄萎病和抗棉铃虫.杂种优势分析表明, 冀资139的纤维长度、比强度和马克隆值的遗传主要由加性效应控制.这与已有的纤维长度、比强度和马克隆值遗传规律的研究结果一致(袁有禄等,
2002 ; 殷剑美等,
2003 ; 聂新辉等,
2015 ), 但是该种质在改良纤维品质较差的品种时具有较大的加性效应, 可以根据综合性状互补原则, 提出最佳的亲本选配和后代选择策略, 同时对杂交后代抉选时要注重选择带有纯合目的基因的植株, 从而大幅度提高育种效率.而在对优质纤维品种的进一步改良时, 要充分考虑该种质对优质品种改良的微效性, 可有目的地改良个别性状.陆地棉特异种质冀资139纤维品质以加性效应为遗传基础, 这一结论为骨干亲本的构建和应用提供了理论依据.本研究重点分析了冀资139的纤维品质性状杂种优势, 而其产量性状受衣分、铃重和单株成铃等因素影响, 效应比较复杂, 还需设计更加完备的试验进行揭示. ...
棉花纤维发育的遗传机制及分子标记
1
2005
... 提高产量并改良纤维品质是陆地棉(
Gossypium hirsutum )育种的重要目标.棉纤维由胚珠表皮细胞发育而来, 而每个胚珠上的纤维数量、长度与强度等因素决定了棉花的产量和纤维品质(毛玮和曹跃芬,
2018 ).胚珠表皮细胞发育成为纤维需经历初始分化、细胞伸长(初生壁合成)、次生壁合成和脱水成熟4个时期, 其中前3个时期主要决定棉纤维的产量和品质(Kim and Triplett,
2001 ; 潘玉欣等,
2005 ).胚珠表皮细胞的生长发育受到一系列基因的调控, 如
GhMYB (Loguercio
et al. 1999 ; Walford
et al. 2012 )、
GhHD-1 (Wan
et al. 2016 )和
GhMML3 _
A12 (李慧琴等,
2019 ).棉纤维发育受植物激素的调控, 如吲哚乙酸(IAA)和赤霉素(GA
3 )可以直接诱导纤维的发生与伸长(王芙蓉等,
2001 ; Zhang
et al. 2011 ; 王国宁等,
2014 ; 唐淑荣等,
2019 ).Zhang等(
2011 )构建了矮牵牛(
Petunia hybrida )花种皮特异启动子
FBP7 驱动的
iaaM 基因, 并导入棉花种质, 获得了转
FBP7::iaaM 基因种质.该种质胚珠上皮细胞初始分化时期的IAA水平较转化受体增高, 并且棉纤维数量显著提高, 进而衣分较受体对照提高, 同时纤维细度也得到了一定程度的改善. ...
不同熟性棉花品种纤维品质特征分析与评价
1
2019
... 提高产量并改良纤维品质是陆地棉(
Gossypium hirsutum )育种的重要目标.棉纤维由胚珠表皮细胞发育而来, 而每个胚珠上的纤维数量、长度与强度等因素决定了棉花的产量和纤维品质(毛玮和曹跃芬,
2018 ).胚珠表皮细胞发育成为纤维需经历初始分化、细胞伸长(初生壁合成)、次生壁合成和脱水成熟4个时期, 其中前3个时期主要决定棉纤维的产量和品质(Kim and Triplett,
2001 ; 潘玉欣等,
2005 ).胚珠表皮细胞的生长发育受到一系列基因的调控, 如
GhMYB (Loguercio
et al. 1999 ; Walford
et al. 2012 )、
GhHD-1 (Wan
et al. 2016 )和
GhMML3 _
A12 (李慧琴等,
2019 ).棉纤维发育受植物激素的调控, 如吲哚乙酸(IAA)和赤霉素(GA
3 )可以直接诱导纤维的发生与伸长(王芙蓉等,
2001 ; Zhang
et al. 2011 ; 王国宁等,
2014 ; 唐淑荣等,
2019 ).Zhang等(
2011 )构建了矮牵牛(
Petunia hybrida )花种皮特异启动子
FBP7 驱动的
iaaM 基因, 并导入棉花种质, 获得了转
FBP7::iaaM 基因种质.该种质胚珠上皮细胞初始分化时期的IAA水平较转化受体增高, 并且棉纤维数量显著提高, 进而衣分较受体对照提高, 同时纤维细度也得到了一定程度的改善. ...
我国棉花种质创新进展与展望
1
2001
... 提高产量并改良纤维品质是陆地棉(
Gossypium hirsutum )育种的重要目标.棉纤维由胚珠表皮细胞发育而来, 而每个胚珠上的纤维数量、长度与强度等因素决定了棉花的产量和纤维品质(毛玮和曹跃芬,
2018 ).胚珠表皮细胞发育成为纤维需经历初始分化、细胞伸长(初生壁合成)、次生壁合成和脱水成熟4个时期, 其中前3个时期主要决定棉纤维的产量和品质(Kim and Triplett,
2001 ; 潘玉欣等,
2005 ).胚珠表皮细胞的生长发育受到一系列基因的调控, 如
GhMYB (Loguercio
et al. 1999 ; Walford
et al. 2012 )、
GhHD-1 (Wan
et al. 2016 )和
GhMML3 _
A12 (李慧琴等,
2019 ).棉纤维发育受植物激素的调控, 如吲哚乙酸(IAA)和赤霉素(GA
3 )可以直接诱导纤维的发生与伸长(王芙蓉等,
2001 ; Zhang
et al. 2011 ; 王国宁等,
2014 ; 唐淑荣等,
2019 ).Zhang等(
2011 )构建了矮牵牛(
Petunia hybrida )花种皮特异启动子
FBP7 驱动的
iaaM 基因, 并导入棉花种质, 获得了转
FBP7::iaaM 基因种质.该种质胚珠上皮细胞初始分化时期的IAA水平较转化受体增高, 并且棉纤维数量显著提高, 进而衣分较受体对照提高, 同时纤维细度也得到了一定程度的改善. ...
转
iaaM 基因高衣分棉花新种质材料创制
1
2014
... 提高产量并改良纤维品质是陆地棉(
Gossypium hirsutum )育种的重要目标.棉纤维由胚珠表皮细胞发育而来, 而每个胚珠上的纤维数量、长度与强度等因素决定了棉花的产量和纤维品质(毛玮和曹跃芬,
2018 ).胚珠表皮细胞发育成为纤维需经历初始分化、细胞伸长(初生壁合成)、次生壁合成和脱水成熟4个时期, 其中前3个时期主要决定棉纤维的产量和品质(Kim and Triplett,
2001 ; 潘玉欣等,
2005 ).胚珠表皮细胞的生长发育受到一系列基因的调控, 如
GhMYB (Loguercio
et al. 1999 ; Walford
et al. 2012 )、
GhHD-1 (Wan
et al. 2016 )和
GhMML3 _
A12 (李慧琴等,
2019 ).棉纤维发育受植物激素的调控, 如吲哚乙酸(IAA)和赤霉素(GA
3 )可以直接诱导纤维的发生与伸长(王芙蓉等,
2001 ; Zhang
et al. 2011 ; 王国宁等,
2014 ; 唐淑荣等,
2019 ).Zhang等(
2011 )构建了矮牵牛(
Petunia hybrida )花种皮特异启动子
FBP7 驱动的
iaaM 基因, 并导入棉花种质, 获得了转
FBP7::iaaM 基因种质.该种质胚珠上皮细胞初始分化时期的IAA水平较转化受体增高, 并且棉纤维数量显著提高, 进而衣分较受体对照提高, 同时纤维细度也得到了一定程度的改善. ...
IaaM 基因对不同遗传背景棉花品种纤维品质及衣分的影响
1
2019
... 棉花的产量和品质性状均为微效多基因控制的数量性状, 且受加性和显性效应共同控制.通常情况下, 产量性状的显性效应值大于加性效应值, 而品质性状的加性效应值大于显性效应值, 用传统育种方法同步改良棉花产量和品质比较困难.棉纤维的产量和质量直接受植物激素IAA调控, IAA在植物体内通过色氨酸途径产生.色氨酸合成途径主要包括4条支路: 吲哚丙酮途径、色胺途径、吲哚乙腈途径和吲哚乙酰胺途径(王士杰等,
2019 ).目前, 研究得比较清楚的途径是存在于根癌农杆菌(
Agrobacterium tumefaciens )和假单孢菌(
Pseudomonas savastanoi )中的吲哚乙酰胺途径, 色氨酸单加氧酶(iaaM)是催化该途径的限速酶(陈旭升等,
2015 ).Zhang等(
2011 )把
iaaM 基因导入棉花种质后获得转
FBP7::iaaM 种质, 连续4年的田间试验表明, 转
iaaM 基因种质与受体种质相比衣分提高15%, 马克隆值有所降低, 而产量和品质性状无影响.随后, 借助传统育种方法, 利用转
FBP7::iaaM 种质提高了其它棉花种质的衣分比例, 同时降低马克隆值, 而且能保持其它优异性状不变(丁晓艳等,
2018 ).例如, 肖钦之(
2015 )将转
iaaM 基因的IF11与鄂抗10选、TM-1、冀棉14、优质-1及非转基因的IF11进行正、反交, 分析了这些亲本材料和F
1 代的产量和品质等性状, 结果表明, IF11可以显著增加杂种后代的衣分含量, 并对马克隆值有明显的改善作用.刘宏伟等(
2016 )以B0011为受体亲本, 以具有
iaaM 基因的高衣分品系THL作为供体亲本, 获得了具有外源
iaaM 基因、高衣分和低马克隆值的B0011改良株系.通过B0011改良株系将
iaaM 基因导入华杂棉H318, 这个携带外源基因的杂合株系在产量构成因素中除衣分增加外, 其它主要构成因子与非转基因的华杂棉 H318无明显差异.刘存敬等(
2016 )以IF1-1为父本, 分别与16个陆地棉品系杂交, 分析了亲本及杂种后代F
1 和F
2 群体中
iaaM 基因的遗传规律, 同时选育获得了高衣分新种质, 且F
1 代皮棉产量优势值为13.4%.丁晓艳等(
2018 )通过回交将IF1-1材料中的
FBP7::iaaM 基因导入晋棉11, 获得回交后代品系JBC4, JBC4保留了晋棉11的早熟特点, 同时衣分提高了12.8%, 纤维产量增加56.3%, 马克隆值降低10.7%.本研究利用含有
FBP7::iaaM 基因的种质, 通过常规杂交育种手段实现了目的基因向骨干亲本的转移, 培育出优良陆地棉种质冀资139, 其纤维品质目标性状较突出, 综合性状实现了双亲优良特性的互补, 分子检测表明目的基因未丢失.研究结果表明,
FBP7::iaaM 基因及其所调控的优良性状可以在骨干育种亲本中传递, 具有较高的育种价值. ...
1
2015
... 棉花的产量和品质性状均为微效多基因控制的数量性状, 且受加性和显性效应共同控制.通常情况下, 产量性状的显性效应值大于加性效应值, 而品质性状的加性效应值大于显性效应值, 用传统育种方法同步改良棉花产量和品质比较困难.棉纤维的产量和质量直接受植物激素IAA调控, IAA在植物体内通过色氨酸途径产生.色氨酸合成途径主要包括4条支路: 吲哚丙酮途径、色胺途径、吲哚乙腈途径和吲哚乙酰胺途径(王士杰等,
2019 ).目前, 研究得比较清楚的途径是存在于根癌农杆菌(
Agrobacterium tumefaciens )和假单孢菌(
Pseudomonas savastanoi )中的吲哚乙酰胺途径, 色氨酸单加氧酶(iaaM)是催化该途径的限速酶(陈旭升等,
2015 ).Zhang等(
2011 )把
iaaM 基因导入棉花种质后获得转
FBP7::iaaM 种质, 连续4年的田间试验表明, 转
iaaM 基因种质与受体种质相比衣分提高15%, 马克隆值有所降低, 而产量和品质性状无影响.随后, 借助传统育种方法, 利用转
FBP7::iaaM 种质提高了其它棉花种质的衣分比例, 同时降低马克隆值, 而且能保持其它优异性状不变(丁晓艳等,
2018 ).例如, 肖钦之(
2015 )将转
iaaM 基因的IF11与鄂抗10选、TM-1、冀棉14、优质-1及非转基因的IF11进行正、反交, 分析了这些亲本材料和F
1 代的产量和品质等性状, 结果表明, IF11可以显著增加杂种后代的衣分含量, 并对马克隆值有明显的改善作用.刘宏伟等(
2016 )以B0011为受体亲本, 以具有
iaaM 基因的高衣分品系THL作为供体亲本, 获得了具有外源
iaaM 基因、高衣分和低马克隆值的B0011改良株系.通过B0011改良株系将
iaaM 基因导入华杂棉H318, 这个携带外源基因的杂合株系在产量构成因素中除衣分增加外, 其它主要构成因子与非转基因的华杂棉 H318无明显差异.刘存敬等(
2016 )以IF1-1为父本, 分别与16个陆地棉品系杂交, 分析了亲本及杂种后代F
1 和F
2 群体中
iaaM 基因的遗传规律, 同时选育获得了高衣分新种质, 且F
1 代皮棉产量优势值为13.4%.丁晓艳等(
2018 )通过回交将IF1-1材料中的
FBP7::iaaM 基因导入晋棉11, 获得回交后代品系JBC4, JBC4保留了晋棉11的早熟特点, 同时衣分提高了12.8%, 纤维产量增加56.3%, 马克隆值降低10.7%.本研究利用含有
FBP7::iaaM 基因的种质, 通过常规杂交育种手段实现了目的基因向骨干亲本的转移, 培育出优良陆地棉种质冀资139, 其纤维品质目标性状较突出, 综合性状实现了双亲优良特性的互补, 分子检测表明目的基因未丢失.研究结果表明,
FBP7::iaaM 基因及其所调控的优良性状可以在骨干育种亲本中传递, 具有较高的育种价值. ...
陆地棉产量与品质性状的主基因与多基因遗传分析
1
2003
... 本研究利用转
FBP7::iaaM 种质培育出优良陆地棉种质冀资139.冀资139株型紧凑, 衣分为42.3%, 上半部平均长度32.2 mm, 断裂比强度31.5 cN/tex, 马克隆值4.2, 整齐度指数84.0%; 并且抗枯萎病、耐黄萎病和抗棉铃虫.杂种优势分析表明, 冀资139的纤维长度、比强度和马克隆值的遗传主要由加性效应控制.这与已有的纤维长度、比强度和马克隆值遗传规律的研究结果一致(袁有禄等,
2002 ; 殷剑美等,
2003 ; 聂新辉等,
2015 ), 但是该种质在改良纤维品质较差的品种时具有较大的加性效应, 可以根据综合性状互补原则, 提出最佳的亲本选配和后代选择策略, 同时对杂交后代抉选时要注重选择带有纯合目的基因的植株, 从而大幅度提高育种效率.而在对优质纤维品种的进一步改良时, 要充分考虑该种质对优质品种改良的微效性, 可有目的地改良个别性状.陆地棉特异种质冀资139纤维品质以加性效应为遗传基础, 这一结论为骨干亲本的构建和应用提供了理论依据.本研究重点分析了冀资139的纤维品质性状杂种优势, 而其产量性状受衣分、铃重和单株成铃等因素影响, 效应比较复杂, 还需设计更加完备的试验进行揭示. ...
棉花育种行业创新与进展
1
2018
... 棉纤维由胚珠表皮细胞发育而来, 其发育受到一系列基因调控.棉纤维长度和强度的决定发生在次生壁合成期和脱水成熟期.一些在棉纤维发育过程中发挥重要作用的基因, 诸如各种转录因子基因、参与激素代谢的基因、编码细胞壁蛋白和细胞骨架蛋白的基因、活性氧代谢相关基因、以及参与糖和脂类代谢的基因已有相关报道(Fang
et al. 2017a ; 袁有禄等,
2018 ).棉花全基因组重测序及全基因组关联分析(GWAS)揭示了陆地棉进化过程中棉纤维性状演变规律, 并且发现多个候选位点与纤维品质性状相关联(Fang
et al. 2017b ).利用719份陆地棉材料作为关联分析群体, 研究发现46个显著的SNPs与5个纤维品质性状关联, 涉及612个候选基因, 有2个与纤维长度及强度有关的单倍型, 有163个及120个纤维发育基因分别与纤维长度和强度有关(Fang
et al. 2017b ; Sun
et al. 2017 ).对棉纤维发育相关基因及其调控网络进行深入研究, 发现陆地棉纤维品质改良具有复杂性和难度.
FBP7::iaaM 基因能够调控胚珠表皮细胞IAA的含量, 进而促进棉纤维的发育, 而以转育的综合性状优良、优质、高衣分的冀资139作为亲本, 其F
1 代主要纤维品质性状指标表现为两个亲本的平均值, 并不具备杂种优势, 推测是父母本调控激素的累加效应造成的.本研究在试验过程中仅考虑了
FBP7::iaaM 基因是否丢失及纤维表型数据的检测, 没有同时检测亲本和杂交种胚珠上皮细胞初始分化时期的
IAA 表达水平, 因此未能在分子机制上对不同的纤维发育性状进行阐述, 是本研究设计的缺陷, 我们将在后续的研究中筛选一些与纤维发育相关的关键基因与
FBP7::iaaM 基因一起, 评价
FBP7::iaaM 基因对不同纤维发育的影响.从育种角度看, 要注重目的基因与其它基因间的互作关系, 以及控制相关性状代谢途径潜在调控机制的遗传效应研究.仅依靠单个基因(如
FBP7::iaaM 基因)的利用实现棉花纤维产量和品质的突破, 尚有一定难度.而更应重视优化利用控制某一性状的一系列基因, 并使其在后代稳定传递.本研究仅限于杂种一代的纤维品质性状表现分析, 随后的分离重组世代是否能出现优于双亲性状的单株和株系, 还有待深入研究. ...
棉花高品质纤维性状的主基因与多基因遗传分析
1
2002
... 本研究利用转
FBP7::iaaM 种质培育出优良陆地棉种质冀资139.冀资139株型紧凑, 衣分为42.3%, 上半部平均长度32.2 mm, 断裂比强度31.5 cN/tex, 马克隆值4.2, 整齐度指数84.0%; 并且抗枯萎病、耐黄萎病和抗棉铃虫.杂种优势分析表明, 冀资139的纤维长度、比强度和马克隆值的遗传主要由加性效应控制.这与已有的纤维长度、比强度和马克隆值遗传规律的研究结果一致(袁有禄等,
2002 ; 殷剑美等,
2003 ; 聂新辉等,
2015 ), 但是该种质在改良纤维品质较差的品种时具有较大的加性效应, 可以根据综合性状互补原则, 提出最佳的亲本选配和后代选择策略, 同时对杂交后代抉选时要注重选择带有纯合目的基因的植株, 从而大幅度提高育种效率.而在对优质纤维品种的进一步改良时, 要充分考虑该种质对优质品种改良的微效性, 可有目的地改良个别性状.陆地棉特异种质冀资139纤维品质以加性效应为遗传基础, 这一结论为骨干亲本的构建和应用提供了理论依据.本研究重点分析了冀资139的纤维品质性状杂种优势, 而其产量性状受衣分、铃重和单株成铃等因素影响, 效应比较复杂, 还需设计更加完备的试验进行揭示. ...
胞质雄性不育系冀2658A细胞质对陆地棉主要性状的影响
1
2017
... 冀资139的母本冀棉958 (
Gossypium hirsutum L.)来源于河北省农林科学院棉花研究所, 父本为转
iaaM 基因棉花种质IF1-1, 由西南大学裴炎教授提供.用于测试冀资139纤维品质性状杂种优势的4个测试品种(系)纤维品质差异明显, 分别为: 946系(P
1 )、05-198 (P
2 )、2658系(P
3 ) (赵存鹏等,
2017 )和646系(P
4 ), 均来源于河北省农林科学院棉花研究所, 系谱及特性见
表1 .由
表1 可知, 5个亲本材料纤维长度、比强度和马克隆值都存在明显差异, 分别代表短绒棉、中绒棉以及中长绒棉纤维品种类型: 纤维长度最大值为32.2 mm, 最小值为26.3 mm; 比强度最大值为31.1 cN/tex, 最小值为24.7 cN/tex; 马克隆值的最大值为6.0, 最小值为4.2. ...
Genomic insights into divergence and dual domestication of cultivated allotetraploid cottons
1
2017a
... 棉纤维由胚珠表皮细胞发育而来, 其发育受到一系列基因调控.棉纤维长度和强度的决定发生在次生壁合成期和脱水成熟期.一些在棉纤维发育过程中发挥重要作用的基因, 诸如各种转录因子基因、参与激素代谢的基因、编码细胞壁蛋白和细胞骨架蛋白的基因、活性氧代谢相关基因、以及参与糖和脂类代谢的基因已有相关报道(Fang
et al. 2017a ; 袁有禄等,
2018 ).棉花全基因组重测序及全基因组关联分析(GWAS)揭示了陆地棉进化过程中棉纤维性状演变规律, 并且发现多个候选位点与纤维品质性状相关联(Fang
et al. 2017b ).利用719份陆地棉材料作为关联分析群体, 研究发现46个显著的SNPs与5个纤维品质性状关联, 涉及612个候选基因, 有2个与纤维长度及强度有关的单倍型, 有163个及120个纤维发育基因分别与纤维长度和强度有关(Fang
et al. 2017b ; Sun
et al. 2017 ).对棉纤维发育相关基因及其调控网络进行深入研究, 发现陆地棉纤维品质改良具有复杂性和难度.
FBP7::iaaM 基因能够调控胚珠表皮细胞IAA的含量, 进而促进棉纤维的发育, 而以转育的综合性状优良、优质、高衣分的冀资139作为亲本, 其F
1 代主要纤维品质性状指标表现为两个亲本的平均值, 并不具备杂种优势, 推测是父母本调控激素的累加效应造成的.本研究在试验过程中仅考虑了
FBP7::iaaM 基因是否丢失及纤维表型数据的检测, 没有同时检测亲本和杂交种胚珠上皮细胞初始分化时期的
IAA 表达水平, 因此未能在分子机制上对不同的纤维发育性状进行阐述, 是本研究设计的缺陷, 我们将在后续的研究中筛选一些与纤维发育相关的关键基因与
FBP7::iaaM 基因一起, 评价
FBP7::iaaM 基因对不同纤维发育的影响.从育种角度看, 要注重目的基因与其它基因间的互作关系, 以及控制相关性状代谢途径潜在调控机制的遗传效应研究.仅依靠单个基因(如
FBP7::iaaM 基因)的利用实现棉花纤维产量和品质的突破, 尚有一定难度.而更应重视优化利用控制某一性状的一系列基因, 并使其在后代稳定传递.本研究仅限于杂种一代的纤维品质性状表现分析, 随后的分离重组世代是否能出现优于双亲性状的单株和株系, 还有待深入研究. ...
Genomic analyses in cotton identify signatures of selection and loci associated with fiber quality and yield traits
2
2017b
... 棉纤维由胚珠表皮细胞发育而来, 其发育受到一系列基因调控.棉纤维长度和强度的决定发生在次生壁合成期和脱水成熟期.一些在棉纤维发育过程中发挥重要作用的基因, 诸如各种转录因子基因、参与激素代谢的基因、编码细胞壁蛋白和细胞骨架蛋白的基因、活性氧代谢相关基因、以及参与糖和脂类代谢的基因已有相关报道(Fang
et al. 2017a ; 袁有禄等,
2018 ).棉花全基因组重测序及全基因组关联分析(GWAS)揭示了陆地棉进化过程中棉纤维性状演变规律, 并且发现多个候选位点与纤维品质性状相关联(Fang
et al. 2017b ).利用719份陆地棉材料作为关联分析群体, 研究发现46个显著的SNPs与5个纤维品质性状关联, 涉及612个候选基因, 有2个与纤维长度及强度有关的单倍型, 有163个及120个纤维发育基因分别与纤维长度和强度有关(Fang
et al. 2017b ; Sun
et al. 2017 ).对棉纤维发育相关基因及其调控网络进行深入研究, 发现陆地棉纤维品质改良具有复杂性和难度.
FBP7::iaaM 基因能够调控胚珠表皮细胞IAA的含量, 进而促进棉纤维的发育, 而以转育的综合性状优良、优质、高衣分的冀资139作为亲本, 其F
1 代主要纤维品质性状指标表现为两个亲本的平均值, 并不具备杂种优势, 推测是父母本调控激素的累加效应造成的.本研究在试验过程中仅考虑了
FBP7::iaaM 基因是否丢失及纤维表型数据的检测, 没有同时检测亲本和杂交种胚珠上皮细胞初始分化时期的
IAA 表达水平, 因此未能在分子机制上对不同的纤维发育性状进行阐述, 是本研究设计的缺陷, 我们将在后续的研究中筛选一些与纤维发育相关的关键基因与
FBP7::iaaM 基因一起, 评价
FBP7::iaaM 基因对不同纤维发育的影响.从育种角度看, 要注重目的基因与其它基因间的互作关系, 以及控制相关性状代谢途径潜在调控机制的遗传效应研究.仅依靠单个基因(如
FBP7::iaaM 基因)的利用实现棉花纤维产量和品质的突破, 尚有一定难度.而更应重视优化利用控制某一性状的一系列基因, 并使其在后代稳定传递.本研究仅限于杂种一代的纤维品质性状表现分析, 随后的分离重组世代是否能出现优于双亲性状的单株和株系, 还有待深入研究. ...
...
2017b ; Sun
et al. 2017 ).对棉纤维发育相关基因及其调控网络进行深入研究, 发现陆地棉纤维品质改良具有复杂性和难度.
FBP7::iaaM 基因能够调控胚珠表皮细胞IAA的含量, 进而促进棉纤维的发育, 而以转育的综合性状优良、优质、高衣分的冀资139作为亲本, 其F
1 代主要纤维品质性状指标表现为两个亲本的平均值, 并不具备杂种优势, 推测是父母本调控激素的累加效应造成的.本研究在试验过程中仅考虑了
FBP7::iaaM 基因是否丢失及纤维表型数据的检测, 没有同时检测亲本和杂交种胚珠上皮细胞初始分化时期的
IAA 表达水平, 因此未能在分子机制上对不同的纤维发育性状进行阐述, 是本研究设计的缺陷, 我们将在后续的研究中筛选一些与纤维发育相关的关键基因与
FBP7::iaaM 基因一起, 评价
FBP7::iaaM 基因对不同纤维发育的影响.从育种角度看, 要注重目的基因与其它基因间的互作关系, 以及控制相关性状代谢途径潜在调控机制的遗传效应研究.仅依靠单个基因(如
FBP7::iaaM 基因)的利用实现棉花纤维产量和品质的突破, 尚有一定难度.而更应重视优化利用控制某一性状的一系列基因, 并使其在后代稳定传递.本研究仅限于杂种一代的纤维品质性状表现分析, 随后的分离重组世代是否能出现优于双亲性状的单株和株系, 还有待深入研究. ...
Cotton fiber growth in planta and
in vitro . Models for plant cell elongation and cell wall biogenesis
1
2001
... 提高产量并改良纤维品质是陆地棉(
Gossypium hirsutum )育种的重要目标.棉纤维由胚珠表皮细胞发育而来, 而每个胚珠上的纤维数量、长度与强度等因素决定了棉花的产量和纤维品质(毛玮和曹跃芬,
2018 ).胚珠表皮细胞发育成为纤维需经历初始分化、细胞伸长(初生壁合成)、次生壁合成和脱水成熟4个时期, 其中前3个时期主要决定棉纤维的产量和品质(Kim and Triplett,
2001 ; 潘玉欣等,
2005 ).胚珠表皮细胞的生长发育受到一系列基因的调控, 如
GhMYB (Loguercio
et al. 1999 ; Walford
et al. 2012 )、
GhHD-1 (Wan
et al. 2016 )和
GhMML3 _
A12 (李慧琴等,
2019 ).棉纤维发育受植物激素的调控, 如吲哚乙酸(IAA)和赤霉素(GA
3 )可以直接诱导纤维的发生与伸长(王芙蓉等,
2001 ; Zhang
et al. 2011 ; 王国宁等,
2014 ; 唐淑荣等,
2019 ).Zhang等(
2011 )构建了矮牵牛(
Petunia hybrida )花种皮特异启动子
FBP7 驱动的
iaaM 基因, 并导入棉花种质, 获得了转
FBP7::iaaM 基因种质.该种质胚珠上皮细胞初始分化时期的IAA水平较转化受体增高, 并且棉纤维数量显著提高, 进而衣分较受体对照提高, 同时纤维细度也得到了一定程度的改善. ...
Differential regulation of six novel MYB-domain genes defines two distinct expression patterns in allotetraploid cotton (
Gossypium hirsutum L.)
1
1999
... 提高产量并改良纤维品质是陆地棉(
Gossypium hirsutum )育种的重要目标.棉纤维由胚珠表皮细胞发育而来, 而每个胚珠上的纤维数量、长度与强度等因素决定了棉花的产量和纤维品质(毛玮和曹跃芬,
2018 ).胚珠表皮细胞发育成为纤维需经历初始分化、细胞伸长(初生壁合成)、次生壁合成和脱水成熟4个时期, 其中前3个时期主要决定棉纤维的产量和品质(Kim and Triplett,
2001 ; 潘玉欣等,
2005 ).胚珠表皮细胞的生长发育受到一系列基因的调控, 如
GhMYB (Loguercio
et al. 1999 ; Walford
et al. 2012 )、
GhHD-1 (Wan
et al. 2016 )和
GhMML3 _
A12 (李慧琴等,
2019 ).棉纤维发育受植物激素的调控, 如吲哚乙酸(IAA)和赤霉素(GA
3 )可以直接诱导纤维的发生与伸长(王芙蓉等,
2001 ; Zhang
et al. 2011 ; 王国宁等,
2014 ; 唐淑荣等,
2019 ).Zhang等(
2011 )构建了矮牵牛(
Petunia hybrida )花种皮特异启动子
FBP7 驱动的
iaaM 基因, 并导入棉花种质, 获得了转
FBP7::iaaM 基因种质.该种质胚珠上皮细胞初始分化时期的IAA水平较转化受体增高, 并且棉纤维数量显著提高, 进而衣分较受体对照提高, 同时纤维细度也得到了一定程度的改善. ...
A rapid method for extraction of cotton (
Gossypium spp.) genomic DNA suitable for RFLP or PCR analysis
1
1993
... 苗期选取各株系的幼嫩叶片按Paterson等(
1993 )的方法提取基因组DNA.鉴定
iaaM 基因的正向引物为: 5'-AAGGTAGCAGTTCTCTCCGC-3'; 反向引物为: 5'-TCGGCTTAGGAACATCCTCC-3'.引物由上海生工公司合成. ...
Genome-wide association study discovered genetic variation and candidate genes of fibre quality traits in
Gossypium hirsutum L
1
2017
... 棉纤维由胚珠表皮细胞发育而来, 其发育受到一系列基因调控.棉纤维长度和强度的决定发生在次生壁合成期和脱水成熟期.一些在棉纤维发育过程中发挥重要作用的基因, 诸如各种转录因子基因、参与激素代谢的基因、编码细胞壁蛋白和细胞骨架蛋白的基因、活性氧代谢相关基因、以及参与糖和脂类代谢的基因已有相关报道(Fang
et al. 2017a ; 袁有禄等,
2018 ).棉花全基因组重测序及全基因组关联分析(GWAS)揭示了陆地棉进化过程中棉纤维性状演变规律, 并且发现多个候选位点与纤维品质性状相关联(Fang
et al. 2017b ).利用719份陆地棉材料作为关联分析群体, 研究发现46个显著的SNPs与5个纤维品质性状关联, 涉及612个候选基因, 有2个与纤维长度及强度有关的单倍型, 有163个及120个纤维发育基因分别与纤维长度和强度有关(Fang
et al. 2017b ; Sun
et al. 2017 ).对棉纤维发育相关基因及其调控网络进行深入研究, 发现陆地棉纤维品质改良具有复杂性和难度.
FBP7::iaaM 基因能够调控胚珠表皮细胞IAA的含量, 进而促进棉纤维的发育, 而以转育的综合性状优良、优质、高衣分的冀资139作为亲本, 其F
1 代主要纤维品质性状指标表现为两个亲本的平均值, 并不具备杂种优势, 推测是父母本调控激素的累加效应造成的.本研究在试验过程中仅考虑了
FBP7::iaaM 基因是否丢失及纤维表型数据的检测, 没有同时检测亲本和杂交种胚珠上皮细胞初始分化时期的
IAA 表达水平, 因此未能在分子机制上对不同的纤维发育性状进行阐述, 是本研究设计的缺陷, 我们将在后续的研究中筛选一些与纤维发育相关的关键基因与
FBP7::iaaM 基因一起, 评价
FBP7::iaaM 基因对不同纤维发育的影响.从育种角度看, 要注重目的基因与其它基因间的互作关系, 以及控制相关性状代谢途径潜在调控机制的遗传效应研究.仅依靠单个基因(如
FBP7::iaaM 基因)的利用实现棉花纤维产量和品质的突破, 尚有一定难度.而更应重视优化利用控制某一性状的一系列基因, 并使其在后代稳定传递.本研究仅限于杂种一代的纤维品质性状表现分析, 随后的分离重组世代是否能出现优于双亲性状的单株和株系, 还有待深入研究. ...
Epidermal cell differentiation in cotton mediated by the homeodomain leucine zipper gene,
GhHD-1 1
2012
... 提高产量并改良纤维品质是陆地棉(
Gossypium hirsutum )育种的重要目标.棉纤维由胚珠表皮细胞发育而来, 而每个胚珠上的纤维数量、长度与强度等因素决定了棉花的产量和纤维品质(毛玮和曹跃芬,
2018 ).胚珠表皮细胞发育成为纤维需经历初始分化、细胞伸长(初生壁合成)、次生壁合成和脱水成熟4个时期, 其中前3个时期主要决定棉纤维的产量和品质(Kim and Triplett,
2001 ; 潘玉欣等,
2005 ).胚珠表皮细胞的生长发育受到一系列基因的调控, 如
GhMYB (Loguercio
et al. 1999 ; Walford
et al. 2012 )、
GhHD-1 (Wan
et al. 2016 )和
GhMML3 _
A12 (李慧琴等,
2019 ).棉纤维发育受植物激素的调控, 如吲哚乙酸(IAA)和赤霉素(GA
3 )可以直接诱导纤维的发生与伸长(王芙蓉等,
2001 ; Zhang
et al. 2011 ; 王国宁等,
2014 ; 唐淑荣等,
2019 ).Zhang等(
2011 )构建了矮牵牛(
Petunia hybrida )花种皮特异启动子
FBP7 驱动的
iaaM 基因, 并导入棉花种质, 获得了转
FBP7::iaaM 基因种质.该种质胚珠上皮细胞初始分化时期的IAA水平较转化受体增高, 并且棉纤维数量显著提高, 进而衣分较受体对照提高, 同时纤维细度也得到了一定程度的改善. ...
Small interfering RNAs from bidirectional transcripts of
GhMML3_ A12 regulate cotton fiber development
1
2016
... 提高产量并改良纤维品质是陆地棉(
Gossypium hirsutum )育种的重要目标.棉纤维由胚珠表皮细胞发育而来, 而每个胚珠上的纤维数量、长度与强度等因素决定了棉花的产量和纤维品质(毛玮和曹跃芬,
2018 ).胚珠表皮细胞发育成为纤维需经历初始分化、细胞伸长(初生壁合成)、次生壁合成和脱水成熟4个时期, 其中前3个时期主要决定棉纤维的产量和品质(Kim and Triplett,
2001 ; 潘玉欣等,
2005 ).胚珠表皮细胞的生长发育受到一系列基因的调控, 如
GhMYB (Loguercio
et al. 1999 ; Walford
et al. 2012 )、
GhHD-1 (Wan
et al. 2016 )和
GhMML3 _
A12 (李慧琴等,
2019 ).棉纤维发育受植物激素的调控, 如吲哚乙酸(IAA)和赤霉素(GA
3 )可以直接诱导纤维的发生与伸长(王芙蓉等,
2001 ; Zhang
et al. 2011 ; 王国宁等,
2014 ; 唐淑荣等,
2019 ).Zhang等(
2011 )构建了矮牵牛(
Petunia hybrida )花种皮特异启动子
FBP7 驱动的
iaaM 基因, 并导入棉花种质, 获得了转
FBP7::iaaM 基因种质.该种质胚珠上皮细胞初始分化时期的IAA水平较转化受体增高, 并且棉纤维数量显著提高, 进而衣分较受体对照提高, 同时纤维细度也得到了一定程度的改善. ...
Spatiotemporal manipulation of auxin biosynthesis in cotton ovule epidermal cells enhances fiber yield and quality
3
2011
... 提高产量并改良纤维品质是陆地棉(
Gossypium hirsutum )育种的重要目标.棉纤维由胚珠表皮细胞发育而来, 而每个胚珠上的纤维数量、长度与强度等因素决定了棉花的产量和纤维品质(毛玮和曹跃芬,
2018 ).胚珠表皮细胞发育成为纤维需经历初始分化、细胞伸长(初生壁合成)、次生壁合成和脱水成熟4个时期, 其中前3个时期主要决定棉纤维的产量和品质(Kim and Triplett,
2001 ; 潘玉欣等,
2005 ).胚珠表皮细胞的生长发育受到一系列基因的调控, 如
GhMYB (Loguercio
et al. 1999 ; Walford
et al. 2012 )、
GhHD-1 (Wan
et al. 2016 )和
GhMML3 _
A12 (李慧琴等,
2019 ).棉纤维发育受植物激素的调控, 如吲哚乙酸(IAA)和赤霉素(GA
3 )可以直接诱导纤维的发生与伸长(王芙蓉等,
2001 ; Zhang
et al. 2011 ; 王国宁等,
2014 ; 唐淑荣等,
2019 ).Zhang等(
2011 )构建了矮牵牛(
Petunia hybrida )花种皮特异启动子
FBP7 驱动的
iaaM 基因, 并导入棉花种质, 获得了转
FBP7::iaaM 基因种质.该种质胚珠上皮细胞初始分化时期的IAA水平较转化受体增高, 并且棉纤维数量显著提高, 进而衣分较受体对照提高, 同时纤维细度也得到了一定程度的改善. ...
... ).Zhang等(
2011 )构建了矮牵牛(
Petunia hybrida )花种皮特异启动子
FBP7 驱动的
iaaM 基因, 并导入棉花种质, 获得了转
FBP7::iaaM 基因种质.该种质胚珠上皮细胞初始分化时期的IAA水平较转化受体增高, 并且棉纤维数量显著提高, 进而衣分较受体对照提高, 同时纤维细度也得到了一定程度的改善. ...
... 棉花的产量和品质性状均为微效多基因控制的数量性状, 且受加性和显性效应共同控制.通常情况下, 产量性状的显性效应值大于加性效应值, 而品质性状的加性效应值大于显性效应值, 用传统育种方法同步改良棉花产量和品质比较困难.棉纤维的产量和质量直接受植物激素IAA调控, IAA在植物体内通过色氨酸途径产生.色氨酸合成途径主要包括4条支路: 吲哚丙酮途径、色胺途径、吲哚乙腈途径和吲哚乙酰胺途径(王士杰等,
2019 ).目前, 研究得比较清楚的途径是存在于根癌农杆菌(
Agrobacterium tumefaciens )和假单孢菌(
Pseudomonas savastanoi )中的吲哚乙酰胺途径, 色氨酸单加氧酶(iaaM)是催化该途径的限速酶(陈旭升等,
2015 ).Zhang等(
2011 )把
iaaM 基因导入棉花种质后获得转
FBP7::iaaM 种质, 连续4年的田间试验表明, 转
iaaM 基因种质与受体种质相比衣分提高15%, 马克隆值有所降低, 而产量和品质性状无影响.随后, 借助传统育种方法, 利用转
FBP7::iaaM 种质提高了其它棉花种质的衣分比例, 同时降低马克隆值, 而且能保持其它优异性状不变(丁晓艳等,
2018 ).例如, 肖钦之(
2015 )将转
iaaM 基因的IF11与鄂抗10选、TM-1、冀棉14、优质-1及非转基因的IF11进行正、反交, 分析了这些亲本材料和F
1 代的产量和品质等性状, 结果表明, IF11可以显著增加杂种后代的衣分含量, 并对马克隆值有明显的改善作用.刘宏伟等(
2016 )以B0011为受体亲本, 以具有
iaaM 基因的高衣分品系THL作为供体亲本, 获得了具有外源
iaaM 基因、高衣分和低马克隆值的B0011改良株系.通过B0011改良株系将
iaaM 基因导入华杂棉H318, 这个携带外源基因的杂合株系在产量构成因素中除衣分增加外, 其它主要构成因子与非转基因的华杂棉 H318无明显差异.刘存敬等(
2016 )以IF1-1为父本, 分别与16个陆地棉品系杂交, 分析了亲本及杂种后代F
1 和F
2 群体中
iaaM 基因的遗传规律, 同时选育获得了高衣分新种质, 且F
1 代皮棉产量优势值为13.4%.丁晓艳等(
2018 )通过回交将IF1-1材料中的
FBP7::iaaM 基因导入晋棉11, 获得回交后代品系JBC4, JBC4保留了晋棉11的早熟特点, 同时衣分提高了12.8%, 纤维产量增加56.3%, 马克隆值降低10.7%.本研究利用含有
FBP7::iaaM 基因的种质, 通过常规杂交育种手段实现了目的基因向骨干亲本的转移, 培育出优良陆地棉种质冀资139, 其纤维品质目标性状较突出, 综合性状实现了双亲优良特性的互补, 分子检测表明目的基因未丢失.研究结果表明,
FBP7::iaaM 基因及其所调控的优良性状可以在骨干育种亲本中传递, 具有较高的育种价值. ...
 ,*
,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT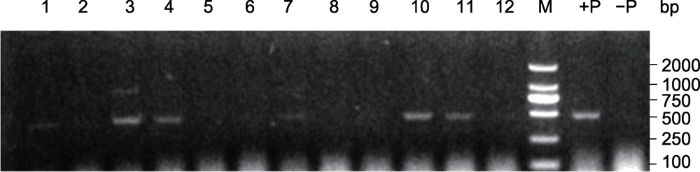 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}