Research Advances in Plant Physiological and Biochemical Mechanisms in Response to Cadmium Stress
Tingting An1,2,3, Di Huang1,2,3, Hao Wang1,2, Yi Zhang3, Yinglong Chen,1,2,4,*1State Key Laboratory of Soil Erosion and Dryland Farming on the Loess Plateau, Northwest A&F University, Yangling 712100, China 2Institute of Soil and Water Conservation, Chinese Academy of Sciences and Ministry of Water Resources, Yangling 712100, China 3College of Forestry, Northwest A&F University, Yangling 712100, China 4Institute of Agriculture, and School of Agriculture and Environment, the University of Western Australia, Perth 6009, Australia
Abstract Cadmium (Cd) is one of the widely distributed and polluted heavy metals. With serious toxicity, Cd not only affects the growth and development of plants, but also endangers human health. This review summarizes recent research advances and underlying mechanisms of physiological and biochemical responses to Cd stress in plants. We also review and discuss plant responses in photosynthetic systems, reactive oxygen species and reactive nitrogen species, antioxidant defense systems, hormones, calcium signals, and their underpinning proteomics and genomics mechanisms. We aim to form a theoretical basis for future research on understanding the physiological and biochemical mechanisms of plant responses to Cd stress. Keywords:cadmium;plant;physiological and biochemical characteristics;stress;mechanism
PDF (1229KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 安婷婷, 黄帝, 王浩, 张一, 陈应龙. 植物响应镉胁迫的生理生化机制研究进展. 植物学报, 2021, 56(3): 347-362 doi:10.11983/CBB20160 An Tingting, Huang Di, Wang Hao, Zhang Yi, Chen Yinglong. Research Advances in Plant Physiological and Biochemical Mechanisms in Response to Cadmium Stress. Chinese Bulletin of Botany, 2021, 56(3): 347-362 doi:10.11983/CBB20160
Cd处理的植物初级根尖和子叶中IAA浓度及分布有较大差异。Cd处理后, IAA含量显著降低(Guo et al., 2019), IAA合成基因表达下调(Bashir et al., 2019), IAA氧化酶活性增强, IAA生物合成和分解代谢基因表达改变(Hu et al., 2013)。此外, Cd胁迫下, 外源IAA能显著提高玉米幼苗根和地上部生物量, 降低地上部Cd含量, 但对根Cd含量的影响不显著, 显著降低叶片的MDA和GSH含量, 使玉米幼苗叶片SOD、POD和CAT活性显著增强(陈晶等, 2016)。当IAA合成受到抑制时, 如施加生长素信号抑制剂氯苯氧基异丁酸, 可有效抑制10 μmol·L-1 Cd诱导的根系生长和H2O2产生, 但不抑制60 μmol·L-1 Cd诱导的根系生长和H2O2产生(Tamás et al., 2012)。Cd胁迫下施加IAA, 可激活谷胱甘肽-S-转移酶(glutathione- S-transferase, GST) (在植物解毒过程中起关键作用)活性, 使植物更好地发生应激反应(Bo?ová et al., 2013)。Cd胁迫下, 施加外源IAA还可显著提高SODs基因的表达, 降低H2O2含量, 从而减少Cd对植物的毒害(Pan et al., 2020)。重金属污染条件下, 外源IAA可促进天然抗性相关巨噬细胞蛋白基因NRAMP (NATURAL RESISTANCE-ASSOCIATED MACROPHAGE PROTEIN)、IRT、重金属ATP酶家族基因HMA (heavy metal ATPase)和锌铁转运蛋白基因ZIP (zinc iron transporter protein)的表达, 提高植物的抗Cd能力(Korshunova et al., 1999; 罗莎, 2017)。
外源SA可改变ROS的积累和抗氧化酶系统活性, 缓解Cd胁迫带来的不利影响(Zawoznik et al., 2007)。施加外源SA可使油菜(Brassica campestris)叶片的可溶性糖和游离脯氨酸含量升高、MDA积累减少, 质膜透性降低, CAT、POD和SOD活性升高, 明显缓解Cd毒害(胡春霞和王秀芹, 2010)。SA处理显著增强玉米叶片中SOD活性, 降低CAT和POD活性, 并减轻Cd对APX活性的抑制作用(Krantev et al., 2008)。此外, SA通过抑制ROS的过量产生缓解Cd引发的氧化损伤(Zawoznik et al., 2007); 施用外源SA可减少根系中ROS和MDA的积累, 进而减轻Cd毒性(Liu et al., 2016); 外源SA也可介导NADPH氧化酶产生H2O2, 以应对Cd胁迫引起的氧化应激(Chao et al., 2010)。赵新月等(2013)研究表明, 与仅用Cd处理的玉米相比, 用50-250 μmol·L-1 SA处理玉米其叶绿素含量、硝酸还原酶(nitrite reductase, NR)、谷氨酰胺合成酶(glutamine synthetase, GS)和谷氨酸合成酶(glutamate synthase, GOGAT)活性显著升高, 氮素代谢过程良好, 玉米根系的POD活性降低(但POD和SOD活性与无Cd处理相比无显著差异), 且降低了Cd胁迫对质膜氧化系统的伤害。但外源SA对Cd胁迫修复的缓解作用受浓度影响较大, 低浓度的SA具有良好的修复效果, 高浓度的SA反而影响了光合色素的合成及酶活性, 抑制玉米的生长(赵新月等, 2013)。重金属胁迫下, 外源SA提高了植物SOD、CAT和APX合成基因的表达(Gill, 2015; Song et al., 2019); 及水稻叶片和稻穗中低亲和离子转运子基因(OsLCT1)、水稻低Cd基因(OsLCD)和ZIP的时空表达(Liu et al., 2016)。
Cd胁迫通常促进JA合成基因(Bashir et al., 2019)和抗氧化酶合成基因的转录(Keunen et al., 2013)。低浓度JA可缓解Cd胁迫。例如, 用20 μmol·L-1 JA处理可有效改善Cd诱导的氧化应激, 使硫代巴比妥酸反应物质、H2O2和O2-.的积累减少, 使GSH、CAT、SOD及血红素氧合酶(HO)活性增强(Noriega et al., 2012; Adhikari et al., 2018)。研究发现, 番茄(Solanum lycopersicum) JA缺失突变体对Cd的敏感性增强, 与光合作用相关参数(包括总叶绿素、光系统II实际效率、可变叶绿素荧光与最大荧光比及净光合速率)值降低; 相对水含量、可溶性糖和脯氨酸含量及淀粉的积累减少; 氧化应激和抗氧化防御作用(包括H2O2与MDA生成、电解质渗漏、SOD、APX和CAT活性)减弱(Zhao et al., 2016)。外源茉莉酸甲酯(MeJA)通过降低拟南芥根系和茎叶中的Cd浓度, 下调参与Cd吸收和运输的AtIRT1、AtHMA2和AtHMA4基因表达, 缓解Cd引起的新叶毒害。JA合成关键基因AtAOS突变上调AtIRT1、AtHMA2和AtHMA4等转运蛋白基因表达, 增加根和芽中的Cd浓度, 提高拟南芥对Cd的敏感性。此外, MeJA降低Cd胁迫的拟南芥根尖中的NO水平(Lei et al., 2020)。镉胁迫下植物激素ABA、IAA、SA和JA之间的相互作用值得深入探索。
1.1.5 植物钙信号
Ca2+作为细胞功能的第二信使, 参与调节植物各种逆境下的重要生理生化过程。钙信使系统包括Ca2+、钙调蛋白(CaM)和钙泵(Ca2+-ATPase)。CaM是Ca2+的感受器, Ca2+-ATPase决定着Ca2+的跨膜运输(Snedden and Fromm, 2001)。正常情况下三者的含量保持稳定, 可维持细胞正常的生理功能。在外界生物和非生物胁迫下, 细胞质中的Ca2+浓度会瞬间升高(Ca2+浓度的波动被认为是植物对外界刺激的反应信号) (Nürnberger and Scheel, 2001; Liu et al., 2017)。Cd胁迫扰动植物细胞中的Ca2+水平, 从而扰乱植物叶片的水分状况(Perfus-Barbeoch et al., 2002)。胞质Ca2+浓度增加对H2O2的生成有动态调控作用, Ca2+可调控NADH氧化酶活性, 上调植物中H2O2的含量; 也可通过刺激H2O2酶活性下调植物中的H2O2水平(Yang and Poovaiah, 2002)。当ROS含量超过Ca2+调控的浓度范围时, 会对植物造成伤害, 细胞膜和细胞器膜发生膜质过氧化, 膜的通透性被破坏, 致使细胞死亡(赵士诚等, 2008)。Cd与Ca2+的化学性质相似, Cd被植物吸收后通过金属转运体或钙通道运输至根部, 也可通过电压依赖的钙通道进入细胞, 一旦进入胞质, 它就会模拟钙离子的活性, 扰乱钙信号调控; 导致根系吸收大量的Cd, 并显著减少根长、根表面积、根体积和根系生物量, 使叶片黄化并干燥坏死(Yang and Poovaiah, 2002; Wang et al., 2016; Ismael et al., 2019)。Cd进入植物根细胞也诱导一些对重金属胁迫有反应的特定信号分子, 如CaMs、钙依赖蛋白激酶(calcium-dependent protein kinases, CDPKs)和丝裂原活化蛋白激酶(mitogen-activated protein kinases, MAPKs), 调控植物对逆境的抗性(Danquah et al., 2014; Xu et al., 2017)。
AdhikariS, GhoshS, AzaharI, AdhikariA, ShawAK, KonarS, RoyS, HossainZ (2018). Sulfate improves cadmium tolerance by limiting cadmium accumulation, modulation of sulfur metabolism and antioxidant defense system in maize Environ Exp Bot 153, 143-162. DOI:10.1016/j.envexpbot.2018.05.008URL [本文引用: 1]
AgarwalP, MitraM, BanerjeeS, RoyS (2020). MYB4 transcription factor, a member of R2R3-subfamily of MYB domain protein, regulates cadmium tolerance via enhanced protection against oxidative damage and increases expression of PCS1 and MT1C in Arabidopsis Plant Sci 297, 110501. DOI:10.1016/j.plantsci.2020.110501URL [本文引用: 1]
AnjumSA, TanveerM, HussainS, UllahE, WangLC, KhanI, SamadRA, TungSA, AnamM, ShahzadB (2016). Morpho-physiological growth and yield responses of two contrasting maize cultivars to cadmium exposure Clean-Soil Air Water 44, 29-36. DOI:10.1002/clen.v44.1URL [本文引用: 2]
Arasimowicz-JelonekM, Floryszak-WieczorekJ, DeckertJ, Rucińska-SobkowiakR, GzylJ, Pawlak-SpradaS, AbramowskiD, JelonekT, Gwó?d?EA (2012). Nitric oxide implication in cadmium-induced programmed cell death in roots and signaling response of yellow lupine plants Plant Physiol Biochem 58, 124-134. DOI:10.1016/j.plaphy.2012.06.018URL [本文引用: 1]
BashirW, AnwarS, ZhaoQ, HussainI, XieFT (2019). Interactive effect of drought and cadmium stress on soybean root morphology and gene expression Ecotoxicol Environ Saf 175, 90-101. DOI:10.1016/j.ecoenv.2019.03.042URL [本文引用: 8]
BeaupereC, LabunskyyVM (2019). ( Un)folding mechanisms of adaptation to ER stress: lessons from aneuploidy Curr Genet 65, 467-471. DOI:10.1007/s00294-018-0914-9 [本文引用: 1] During stress, accumulation of misfolded proteins in the endoplasmic reticulum (ER) triggers activation of the adaptive mechanisms that restore protein homeostasis. One mechanism that eukaryotic cells use to respond to ER stress is through activation of the unfolded protein response (UPR) signaling pathway, which initiates degradation of misfolded proteins and leads to inhibition of translation and increased expression of chaperones and oxidative folding components that enhance ER protein folding capacity. However, the mechanisms of adaptation to ER stress are not limited to the UPR. Using yeast Saccharomyces cerevisiae, we recently discovered that the protein folding burden in the ER can be alleviated in a UPR-independent manner through duplication of whole chromosomes containing ER stress-protective genes. Here we discuss our findings and their implication to our understanding of the mechanisms by which cells respond to protein misfolding in the ER.
BlaudezD, KohlerA, MartinF, SandersD, ChalotM (2003). Poplar metal tolerance protein 1 confers zinc tolerance and is an oligomeric vacuolar zinc transporter with an essential leucine zipper motif Plant Cell 15, 2911-2928. PMID:14630973 [本文引用: 1] Cation diffusion facilitator (CDF) proteins are a recently discovered family of cation efflux transporters that might play an essential role in metal homeostasis and tolerance. Here, we describe the identification, characterization, and localization of PtdMTP1, a member of the CDF family from the hybrid poplar Populus trichocarpa x Populus deltoides. PtdMTP1 is expressed constitutively and ubiquitously, although at low levels. Heterologous expression in yeast showed that PtdMTP1 was able to complement the hypersensitivity of mutant strains to Zn but not to other metals, including Cd, Co, Mn, and Ni. PtdMTP1 fused to green fluorescent protein localized to the vacuolar membrane both in yeast and in plant cells, consistent with a function of PtdMTP1 in zinc sequestration. Overexpression of PtdMTP1 in Arabidopsis confers Zn tolerance. We show that PtdMTP1, when expressed in yeast and Arabidopsis, forms homooligomers, a novel feature of CDF members. Oligomer formation is disrupted by reducing agents, indicating possible disulfide bridge formation. PtdMTP1 also contains a conserved Leu zipper motif. Although not necessary for oligomer formation, Leu residues within this motif are required for PtdMTP1 functional activity.
Bo?ováB, HuttováJ, MistríkI, TamásL (2013). Auxin signaling is involved in cadmium-induced glutathione-S-transferase activity in barley root Acta Physiol Plant 35, 2685-2690. DOI:10.1007/s11738-013-1300-3URL [本文引用: 2]
CaiZ, XianP, WangH, LinR, HaiN (2020). Transcription factor GmWRKY142 confers cadmium resistance by up- regulating the cadmium tolerance 1-like genes Front Plant Sci 11, 742. DOI:10.3389/fpls.2020.00742URL [本文引用: 2]
ChaoYY, ChenCY, HuangWD, KaoCH (2010). Salicylic acid-mediated hydrogen peroxide accumulation and protection against Cd toxicity in rice leaves Plant Soil 329, 327-337. DOI:10.1007/s11104-009-0161-4URL [本文引用: 2]
ChenSS, YuM, LiH, WangY, LuZC, ZhangYX, LiuMY, QiaoGR, WuLH, HanXJ, ZhuoRY (2020). SaHsfA4c from Sedum alfredii hance enhances cadmium tolerance by regulating ROS-scavenger activities and heat shock proteins expression Front Plant Sci 11, 142. DOI:10.3389/fpls.2020.00142URL [本文引用: 2]
CurieC, CassinG, CouchD, DivolF, HiguchiK, Le JeanM, MissonJ, SchikoraA, CzernicP, MariS (2009). Metal movement within the plant: contribution of nicotianamine and yellow stripe 1-like transporters Ann Bot 103, 1-11. DOI:10.1093/aob/mcn207URL [本文引用: 1]
DalCorsoG, FarinatiS, MaistriS, FuriniA (2008). How plants cope with cadmium: staking all on metabolism and gene expression J Integr Plant Biol 50, 1268-1280. DOI:10.1111/jipb.2008.50.issue-10URL [本文引用: 2]
DanquahA, de ZelicourtA, ColcombetJ, HirtH (2014). The role of ABA and MAPK signaling pathways in plant abiotic stress responses Biotechnol Adv 32, 40-52. DOI:10.1016/j.biotechadv.2013.09.006URL [本文引用: 1]
DasK, RoychoudhuryA (2014). Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants Front Environ Sci 2, 53. [本文引用: 2]
DelledonneM, ZeierJ, MaroccoA, LambC (2001). Signal interactions between nitric oxide and reactive oxygen intermediates in the plant hypersensitive disease resistance response Proc Natl Acad Sci USA 98, 13454-13459. DOI:10.1073/pnas.231178298URL [本文引用: 1]
DengY, SrivastavaR, HowellSH (2013). Protein kinase and ribonuclease domains of IRE1 confer stress tolerance, vegetative growth, and reproductive development in Arabidopsis Proc Natl Acad Sci USA 110, 19633-19638. DOI:10.1073/pnas.1314749110URL [本文引用: 1]
Ekmek?iY, Tanyola?D, AyhanB (2008). Effects of cadmium on antioxidant enzyme and photosynthetic activities in leaves of two maize cultivars J Plant Physiol 165, 600-611. DOI:10.1016/j.jplph.2007.01.017URL [本文引用: 3]
ErpenL, DeviHS, GrosserJW, DuttM (2018). Potential use of the DREB/ERF, MYB, NAC and WRKY transcription factors to improve abiotic and biotic stress in transgenic plants Plant Cell Tissue Organ Cult 132, 1-25. DOI:10.1007/s11240-017-1320-6URL [本文引用: 2]
EtesamiH, JeongBR (2018). Silicon (Si): review and future prospects on the action mechanisms in alleviating biotic and abiotic stresses in plants Ecotoxicol Environ Saf 147, 881-896. DOI:10.1016/j.ecoenv.2017.09.063URL [本文引用: 2]
FanSK, FangXZ, GuanMY, YeYQ, LinXY, DuST, JinCW (2014). Exogenous abscisic acid application decreases cadmium accumulation in Arabidopsis plants, which is associated with the inhibition of IRT1-mediated cadmium uptake Front Plant Sci 5, 721. [本文引用: 2]
FengSS, TanJJ, ZhangYX, LiangS, XiangSQ, WangH, ChaiTY (2017). Isolation and characterization of a novel cadmium-regulated Yellow Stripe-Like transporter (SnYSL3) in Solanum nigrum Plant Cell Rep 36, 281-296. DOI:10.1007/s00299-016-2079-7URL [本文引用: 2]
GuoJJ, QinSY, RengelZ, GaoW, NieZJ, LiuHG, LiC, ZhaoP (2019). Cadmium stress increases antioxidant enzyme activities and decreases endogenous hormone concentrations more in Cd-tolerant than Cd-sensitive wheat varieties Ecotoxicol Environ Saf 172, 380-387. DOI:10.1016/j.ecoenv.2019.01.069URL [本文引用: 3]
GuoJK, WuJ, JiQ, WangC, LuoL, YuanY, WangYH, WangJ (2008). Genome-wide analysis of heat shock transcription factor families in rice and Arabidopsis J Genet Genomics 35, 105-118. DOI:10.1016/S1673-8527(08)60016-8URL [本文引用: 1]
HartlFU, Hayer-HartlM (2002). Molecular chaperones in the cytosol: from nascent chain to folded protein Science 295, 1852-1858. DOI:10.1126/science.1068408URL [本文引用: 1]
HongCY, ChengD, ZhangGQ, ZhuDD, ChenYH, TanMP (2017). The role of ZmWRKY4 in regulating maize antioxidant defense under cadmium stress Biochem Biophys Res Commun 482, 1504-1510. DOI:10.1016/j.bbrc.2016.12.064URL [本文引用: 2]
HuXL, JiangMY, ZhangAY, LuJ (2005). Abscisic acid-induced apoplastic H2O2 accumulation up-regulates the activities of chloroplastic and cytosolic antioxidant enzymes in maize leaves Planta 223, 57-68. DOI:10.1007/s00425-005-0068-0URL [本文引用: 3]
HuXL, JiangMY, ZhangJH, ZhangAY, LinF, TanMP (2007). Calcium-calmodulin is required for abscisic acid- induced antioxidant defense and functions both upstream and downstream of H2O2 production in leaves of maize ( Zea mays) plants New Phytol 173, 27-38. DOI:10.1111/nph.2007.173.issue-1URL [本文引用: 1]
HuYF, ZhouGY, NaXF, YangLJ, NanWB, LiuX, ZhangYQ, LiJL, BiYR (2013). Cadmium interferes with maintenance of auxin homeostasis in Arabidopsis seedlings J Plant Physiol 170, 965-975. DOI:10.1016/j.jplph.2013.02.008URL [本文引用: 2]
HuangXY, DengFL, YamajiN, PinsonSRM, Fujii-KashinoM, DankuJ, DouglasA, GuerinotML, SaltDE, MaJF (2016). A heavy metal P-type ATPase OsHMA4 prevents copper accumulation in rice grain Nat Commun 7, 12138. DOI:10.1038/ncomms12138URL [本文引用: 2]
IsmaelMA, ElyamineAM, MoussaMG, CaiMM, ZhaoXH, HuCX (2019). Cadmium in plants: uptake, toxicity, and its interactions with selenium fertilizers Metallomics 11, 255-277. DOI:10.1039/c8mt00247aPMID:30632600 [本文引用: 1] Cd is the third major contaminant of greatest hazard to the environment after mercury and lead and is considered as the only metal that poses health risks to both humans and animals at plant tissue concentrations that are generally not phytotoxic. Cd accumulation in plant shoots depends on Cd entry through the roots, sequestration within root vacuoles, translocation in the xylem and phloem, and Cd dilution within the plant shoot throughout its growth. Several metal transporters, processes, and channels are involved from the first step of Cd reaching the root cells and until its final accumulation in the edible parts of the plant. It is hard to demonstrate one step as the pivotal factor to decide the Cd tolerance or accumulation ability of plants since the role of a specific transporter/process varies among plant species and even cultivars. In this review, we discuss the sources of Cd pollutants, Cd toxicity to plants, and mechanisms of Cd uptake and redistribution in plant tissues. The metal transporters involved in Cd transport within plant tissues are also discussed and how their manipulation can control Cd uptake and/or translocation. Finally, we discuss the beneficial effects of Se on plants under Cd stress, and how it can minimize or mitigate Cd toxicity in plants.
JiaHL, WangXH, WeiT, ZhouR, MuhammadH, HuaL, RenXH, GuoJK, DingYZ (2019). Accumulation and fixation of Cd by tomato cell wall pectin under Cd stress Environ Exp Bot 167, 103829. DOI:10.1016/j.envexpbot.2019.103829URL [本文引用: 2]
KeunenE, RemansT, OpdenakkerK, JozefczakM, GielenH, GuisezY, VangronsveldJ, CuypersA (2013). A mutant of the Arabidopsis thaliana LIPOXYGENASE1 gene shows altered signaling and oxidative stress related responses after cadmium exposure Plant Physiol Biochem 63, 272-280. DOI:10.1016/j.plaphy.2012.12.005URL [本文引用: 2]
KhanN, YouFM, DatlaR, RavichandranS, JiaBS, CloutierS (2020). Genome-wide identification of ATP binding cassette (ABC) transporter and heavy metal associated (HMA) gene families in flax ( Linum usitatissimum L.) BMC Genomics 21, 722. DOI:10.1186/s12864-020-07121-9URL [本文引用: 2]
KleizenB, BraakmanI (2004). Protein folding and quality control in the endoplasmic reticulum Curr Opin Cell Biol 16, 343-349. DOI:10.1016/j.ceb.2004.06.012URL [本文引用: 2]
KorenkovV, KingB, HirschiK, WagnerGJ (2009). Root-selective expression of AtCAX4 and AtCAX2 results in reduced lamina cadmium in field-grown Nicotiana tabacum L Plant Biotechnol J 7, 219-226. DOI:10.1111/pbi.2009.7.issue-3URL [本文引用: 1]
KorshunovaYO, EideD, ClarkWG, GuerinotML, PakrasiHB (1999). The IRT1 protein from Arabidopsis thaliana is a metal transporter with a broad substrate range Plant Mol Biol 40, 37-44. PMID:10394943 [本文引用: 2] The molecular basis for the transport of manganese across membranes in plant cells is poorly understood. We have found that IRT1, an Arabidopsis thaliana metal ion transporter, can complement a mutant Saccharomyces cerevisiae strain defective in high-affinity manganese uptake (smf1 delta). The IRT1 protein has previously been identified as an iron transporter. The current studies demonstrated that IRT1, when expressed in yeast, can transport manganese as well. This manganese uptake activity was inhibited by cadmium, iron(II) and zinc, suggesting that IRT1 can transport these metals. The IRT1 cDNA also complements a zinc uptake-deficient yeast mutant strain (zrt1zrt2), and IRT1-dependent zinc transport in yeast cells is inhibited by cadmium, copper, cobalt and iron(III). However, IRT1 did not complement a copper uptake-deficient yeast mutant (ctr1), implying that this transporter is not involved in the uptake of copper in plant cells. The expression of IRT1 is enhanced in A. thaliana plants grown under iron deficiency. Under these conditions, there were increased levels of root-associated manganese, zinc and cobalt, suggesting that, in addition to iron, IRT1 mediates uptake of these metals into plant cells. Taken together, these data indicate that the IRT1 protein is a broad-range metal ion transporter in plants.
Kr?merU, TalkeIN, HanikenneM (2007). Transition metal transport FEBS Lett 581, 2263-2272. PMID:17462635 [本文引用: 1] Transition metal transporters are of central importance in the plant metal homeostasis network which maintains internal metal concentrations within physiological limits. An overview is given of the functions of known transition metal transporters in the context of the unique chemical properties of their substrates. The modifications of the metal homeostasis network associated with the adaptation to an extreme metalliferous environment are illustrated in two Brassicaceae metal hyperaccumulator model plants based on cross-species transcriptomics studies. In a comparison between higher plants and unicellular algae, hypotheses are generated for evolutionary changes in metal transporter complements associated with the transition to multicellularity.
KrantevA, YordanovaR, JandaT, SzalaiG, PopovaL (2008). Treatment with salicylic acid decreases the effect of cadmium on photosynthesis in maize plants J Plant Physiol 165, 920-931. DOI:10.1016/j.jplph.2006.11.014URL [本文引用: 2]
LalorGC (2008). Review of cadmium transfers from soil to humans and its health effects in the Jamaican environment Sci Total Environ 400, 162-172. DOI:10.1016/j.scitotenv.2008.07.011URL [本文引用: 1]
LeiGJ, SunL, SunY, ZhuXF, LiGX, ZhengSJ (2020). Jasmonic acid alleviates cadmium toxicity in Arabidopsis via suppression of cadmium uptake and translocation J Integr Plant Biol 62, 218-227. DOI:10.1111/jipb.v62.2URL [本文引用: 2]
LiSW, LengY, FengL, ZengXY (2014). Involvement of abscisic acid in regulating antioxidative defense systems and IAA-oxidase activity and improving adventitious rooting in mung bean (Vigna radiata(L.) Wilczek) seedlings under cadmium stress Environ Sci Pollut Res Int 21, 525-537. DOI:10.1007/s11356-013-1942-0URL [本文引用: 1]
LiZR, MeiXY, LiT, YangS, QinL, LiB, ZuYQ (2021). Effects of calcium application on activities of membrane transporters in Panax notoginseng under cadmium stress Chemosphere 262, 127905. DOI:10.1016/j.chemosphere.2020.127905URL [本文引用: 1]
LiuSL, YangRJ, TripathiDK, LiX, HeW, WuMX, AliS, MaMD, ChengQS, PanYZ (2018). RETRACTED: the interplay between reactive oxygen and nitrogen species contributes in the regulatory mechanism of the nitro-oxidative stress induced by cadmium in Arabidopsis J Hazard Mater 344, 1007-1024. DOI:10.1016/j.jhazmat.2017.12.004URL [本文引用: 1]
LiuZP, DingYF, WangFJ, YeYY, ZhuC (2016). Role of salicylic acid in resistance to cadmium stress in plants Plant Cell Rep 35, 719-731. DOI:10.1007/s00299-015-1925-3URL [本文引用: 4]
MikkelsenMD, PedasP, SchillerM, VinczeE, MillsRF, BorgS, M?llerA, SchjoerringJK, WilliamsLE, BaekgaardL, HolmPB, PalmgrenMG (2012). Barley HvHMA1 is a heavy metal pump involved in mobilizing organellar Zn and Cu and plays a role in metal loading into grains PLoS One 7, e49027. DOI:10.1371/journal.pone.0049027URL [本文引用: 2]
MittlerR, VanderauweraS, SuzukiN, MillerG, TognettiVB, VandepoeleK, GolleryM, ShulaevV, Van BreusegemF (2011). ROS signaling: the new wave? Trends Plant Sci 16, 300-309. DOI:10.1016/j.tplants.2011.03.007URL [本文引用: 1]
MiyadateH, AdachiS, HiraizumiA, TezukaK, NakazawaN, KawamotoT, KatouK, KodamaI, SakuraiK, TakahashiH, Satoh-NagasawaN, WatanabeA, FujimuraT, AkagiH (2011). OsHMA3, a P1B-type of ATPase affects root-to-shoot cadmium translocation in rice by mediating efflux into vacuoles New Phytol 189, 190-199. DOI:10.1111/nph.2010.189.issue-1URL [本文引用: 1]
NagajyotiPC, LeeKD, SreekanthTVM (2010). Heavy metals, occurrence and toxicity for plants: a review Environ Chem Lett 8, 199-216. DOI:10.1007/s10311-010-0297-8URL [本文引用: 2]
Nieves-CordonesM, López-DelacalleM, RódenasR, MartínezV, RubioF, RiveroRM (2019). Critical responses to nutrient deprivation: a comprehensive review on the role of ROS and RNS Environ Exp Bot 161, 74-85. DOI:10.1016/j.envexpbot.2018.10.039 [本文引用: 3] Reactive Oxygen Species (ROS) and Reactive Nitrogen Species (RNS) are central regulators of plant performance and adaptation to a changing environment. In this sense, increasing evidence points to a prominent role of ROS and RNS in coping with changes in nutrient availability (either deficiency or excess). However, decoding of ROS and RNS signatures into specific nutrient responses still remains a mystery. Thus far, it is known that nutrient deficiencies result in high levels of ROS and exert profound effects on the antioxidant cell machinery. ROS signal propagation relies, in part, on the activation of an array of transmembrane transport systems at the plasma membrane. The intracellular ROS signal leads to the upregulation of adaptation mechanisms (involving activation or de novo synthesis of proteins). Unlike ROS, RNS production under nutrient stress is a much less understood process. Nevertheless, RNS, and NO in particular, intensify the antioxidant responses through the stimulation of their respective scavengers which is helpful for minimizing oxidative damage that can occur during nutrient stress. In the present review, we summarize the current state in the field of ROS and RNS signaling in plant nutrition and identify the main gaps in our knowledge, which should be filled in the forthcoming years. Further understanding of the ROS and RNS signaling cascades is pivotal for improving crop function within a context of limited nutrient resources and harsher climatological conditions.
NoriegaG, CruzDS, BatlleA, TomaroM, BalestrasseK (2012). Heme oxygenase is involved in the protection exerted by jasmonic acid against cadmium stress in soybean roots J Plant Growth Regul 31, 79-89. DOI:10.1007/s00344-011-9221-0URL [本文引用: 3]
NürnbergerT, ScheelD (2001). Signal transmission in the plant immune response Trends Plant Sci 6, 372-379. PMID:11495791 [本文引用: 1] Genetic and biochemical dissection of signaling pathways regulating plant pathogen defense has revealed remarkable similarities with the innate immune system of mammals and Drosophila. Numerous plant proteins resembling eukaryotic receptors have been implicated in the perception of pathogen-derived signal molecules. Receptor-mediated changes in levels of free calcium in the cytoplasm and production of reactive oxygen species and nitric oxide constitute early events generally observed in plant-pathogen interactions. Positive and negative regulation of plant pathogen defense responses has been attributed to mitogen-activated protein kinase cascades. In addition, salicylic acid, jasmonic acid and ethylene are components of signaling networks that provide the molecular basis for specificity of plant defense responses. This article reviews recent advances in our understanding of early signaling events involved in the establishment of plant disease resistance.
PanCL, LuHL, LiuJC, YuJY, WangQ, LiJW, YangJJ, HongHL, YanCL (2020). SODs involved in the hormone mediated regulation of H2O2 content in Kandelia obovata root tissues under cadmium stress Environ Pollut 256, 113272. DOI:10.1016/j.envpol.2019.113272URL [本文引用: 2]
ParkJ, SongWY, KoD, EomY, HansenTH, SchillerM, LeeTG, MartinoiaE, LeeY (2012). The phytochelatin transporters AtABCC1 and AtABCC2 mediate tolerance to cadmium and mercury Plant J 69, 278-288. DOI:10.1111/tpj.2011.69.issue-2URL [本文引用: 2]
Perfus-BarbeochL, LeonhardtN, VavasseurA, ForestierC (2002). Heavy metal toxicity: cadmium permeates through calcium channels and disturbs the plant water status Plant J 32, 539-548. PMID:12445125 [本文引用: 1] Because plant wilting has been described as a consequence of cadmium (Cd2+) toxicity, we investigate Cd2+ effects on plant water losses, gas exchanges and stomatal behaviour in Arabidopsis thaliana L. Effects of 1-week Cd2+ application in hydroponic condition (CdCl2 10-100 micro m) were analyzed. A 10- micro m Cd2+ concentration had no significant effect on the plant-water relationship and carbon assimilation. At higher Cd2+ concentrations, a Cd2+ -dependent decrease in leaf conductance and CO2 uptake was observed despite the photosynthetic apparatus appeared not to be affected as probed by fluorescence measurements. In epidermal strip bioassays, nanomolar Cd2+ concentrations reduced stomatal opening under light in A. thaliana, Vicia faba and Commelina communis. Application of 5 micro m ABA limited the root-to-shoot translocation of cadmium. However, the Cd2+-induced stomatal closure was likely ABA-independent, since a 5-day treatment with 50 micro m Cd2+ did not affect the plant relative water content. Additionally, a similar Cd2+-induced stomatal closure was observed in the ABA insensitive mutant abi1-1. Interestingly, this mutant displayed a higher transpiration rate than the wild type but did not accumulate more Cd2+, arguing that Cd2+ uptake is not dependent only on the transpiration flow. Application of putative calcium channels inhibitors suppressed the inhibitory effect of Cd2+ in epidermal strip experiments, suggesting that Cd2+ could enter the guard cell through calcium channels. Patch-clamp studies with V. faba guard cell protoplasts showed that plasma membrane K+ channels were insensitive to external Cd2+ application whereas Ca2+ channels were found permeable to Cd2+. In conclusion, we propose that Cd2+ affects guard cell regulation in an ABA-independent manner by entering the cytosol via Ca2+ channels.
RajakumarS, BhanupriyaN, RaviC, NachiappanV (2016). Endoplasmic reticulum stress and calcium imbalance are involved in cadmium-induced lipid aberrancy in Saccharomyces cerevisiae Cell Stress Chaperones 21, 895-906. DOI:10.1007/s12192-016-0714-4PMID:27344570 [本文引用: 1] The endoplasmic reticulum is the key organelle which controls protein folding, lipid biogenesis, and calcium (Ca(2+)) homeostasis. Cd exposure in Saccharomyces cerevisiae activated the unfolded protein response and was confirmed by the increased Kar2p expression. Cd exposure in wild-type (WT) cells increased PC levels and the PC biosynthetic genes. Deletion of the two phospholipid methyltransferases CHO2 and OPI3 modulated PC, TAG levels and the lipid droplets with cadmium exposure. Interestingly, we noticed an increase in the calcium levels upon Cd exposure in the mutant cells. This study concluded that Cd interrupted calcium homeostasis-induced lipid dysregulation leading to ER stress.
Romero-PuertasMC, SandalioLM (2016). Nitric oxide level is self-regulating and also regulates its ROS partners Front Plant Sci 7, 316. DOI:10.3389/fpls.2016.00316PMID:27014332 [本文引用: 4]
Romero-PuertasMC, Terrón-CameroLC, Peláez-VicoMá, OlmedillaA, SandalioLM (2019). Reactive oxygen and nitrogen species as key indicators of plant responses to Cd stress Environ Exp Bot 161, 107-119. DOI:10.1016/j.envexpbot.2018.10.012 [本文引用: 2] Although cadmium (Cd), an extremely toxic non-essential heavy metal, has no biological function, it is capable of entering plant roots. Cd not only presents a problem for plants, which have developed specific Cd detection, transport and detoxification mechanisms, but also for humans as it can enter the food chain. After entering the root, Cd can be loaded into the xylem and then into the leaves and fruits, thus constituting a major environmental and health hazard worldwide. Understanding the mechanisms involved in plant responses to Cd stress would facilitate the production of crops with a lower Cd uptake and accumulation capacity as well as plants with greater Cd uptake potential for phytoremediation. One of the most common symptoms of Cd toxicity is the induction of oxidative stress in plants, which have developed various strategies to avoid this toxicity, including the early production of reactive oxygen and nitrogen species (ROS and RNS) with signalling functions. This review focuses on the dual role of ROS and RNS in plant responses to Cd stress: in low concentrations, as signalling molecules capable of orchestrating plant responses on the one hand, and at higher concentrations, as oxidative stress inducers on the other.
SamiF, FaizanM, FarazA, SiddiquiH, YusufM, HayatS (2018). Nitric oxide-mediated integrative alterations in plant metabolism to confer abiotic stress tolerance, NO crosstalk with phytohormones and NO-mediated post translational modifications in modulating diverse plant stress Nitric Oxide 73, 22-38. DOI:10.1016/j.niox.2017.12.005URL [本文引用: 2]
SofoA, VittiA, NuzzaciM, TataranniG, ScopaA, VangronsveldJ, RemansT, FalascaG, AltamuraMM, DegolaF, Sanità di ToppiL (2013). Correlation between hormonal homeostasis and morphogenic responses in Arabidopsis thaliana seedlings growing in a Cd/Cu/Zn multi-pollution context Physiol Plant 149, 487-498. [本文引用: 1]
SongG, YuanSX, WenXH, XieZN, LouLQ, HuBY, CaiQS, XuB (2018). Transcriptome analysis of Cd-treated switchgrass root revealed novel transcripts and the importance of HSF/HSP network in switchgrass Cd tolerance Plant Cell Rep 37, 1485-1497. DOI:10.1007/s00299-018-2318-1URL [本文引用: 2]
SongJY, FinneganPM, LiuWH, LiX, YongJWH, XuJT, ZhangQ, WenYX, QinKX, GuoJZ, LiT, ZhaoC, ZhangY (2019). Mechanisms underlying enhanced Cd translocation and tolerance in roots of Populus euramericana in response to nitrogen fertilization Plant Sci 287, 110206. DOI:10.1016/j.plantsci.2019.110206URL [本文引用: 4]
StroińskiA, ChadzinikolauT, Gi?ewskaK, ZielezińskaM (2010). ABA or cadmium induced phytochelatin synthesis in potato tubers Biol Plant 54, 117-120. DOI:10.1007/s10535-010-0017-zURL [本文引用: 3]
TalukdarD (2012). Exogenous calcium alleviates the impact of cadmium-induced oxidative stress in Lens culinaris Medic. Seedlings through modulation of antioxidant enzyme activities J Crop Sci Biotechnol 15, 325-334. DOI:10.1007/s12892-012-0065-3URL [本文引用: 1]
TamásL, Bo?ováB, HuttováJ, LiptákováL, MistríkI, Valentovi?ováK, ZelinováV (2012). Impact of the auxin signaling inhibitor p-chlorophenoxyisobutyric acid on short- term Cd-induced hydrogen peroxide production and growth response in barley root tip J Plant Physiol 169, 1375-1381. DOI:10.1016/j.jplph.2012.05.023URL [本文引用: 1]
TamásL, DudíkováJ, ?Ur?kováK, Halu?KováLU, HuttováJ, MistríkI, OlléM (2008). Alterations of the gene expression, lipid peroxidation, proline and thiol content along the barley root exposed to cadmium J Plant Physiol 165, 1193-1203. DOI:10.1016/j.jplph.2007.08.013URL [本文引用: 1]
UenoD, MilnerMJ, YamajiN, YokoshoK, KoyamaE, ZambranoMC, KaskieM, EbbsS, KochianLV, MaJF (2011). Elevated expression of TcHMA3 plays a key role in the extreme Cd tolerance in a Cd-hyperaccumulating ecotype of Thlaspi caerulescens Plant J 66, 852-862. DOI:10.1111/j.1365-313X.2011.04548.xURL [本文引用: 1]
ülkerB, SomssichIE (2004). WRKY transcription factors: from DNA binding towards biological function Curr Opin Plant Biol 7, 491-498. DOI:10.1016/j.pbi.2004.07.012URL [本文引用: 2]
UNEP (2008). Final review of scientific information on cadmium .https://www.unep.org/resources/report/final-review-scientific-information-cadmium. URL [本文引用: 1]
VaculíkM, PavlovicA, LuxA (2015). Silicon alleviates cadmium toxicity by enhanced photosynthetic rate and modified bundle sheath’s cell chloroplasts ultrastructure in maize Ecotoxicol Environ Saf 120, 66-73. DOI:10.1016/j.ecoenv.2015.05.026URL [本文引用: 2]
VermaS, DubeyRS (2001). Effect of cadmium on soluble sugars and enzymes of their metabolism in rice Biol Plant 44, 117-123. DOI:10.1023/A:1017938809311URL [本文引用: 1]
WangH, ChenWY, SinumvayaboN, LiYF, HanZX, TianJ, MaQ, PanZZ, GengZJ, YangSQ, KangMM, RahmanSU, YangGJ, ZhangY (2020). Phosphorus deficiency induces root proliferation and Cd absorption but inhibits Cd tolerance and Cd translocation in roots of Populus × euramericana Ecotoxicol Environ Saf 204, 111148. DOI:10.1016/j.ecoenv.2020.111148URL [本文引用: 1]
WangP, DengXJ, HuangY, FangXL, ZhangJ, WanHB, YangCY (2016). Root morphological responses of five soybean (Glycine max(L.) Merr) cultivars to cadmium stress at young seedlings Environ Sci Pollut Res Int 23, 1860-1872. DOI:10.1007/s11356-015-5424-4URL [本文引用: 1]
WangY, WangC, LiuYJ, YuKF, ZhouYH (2018). GmHMA3 sequesters Cd to the root endoplasmic reticulum to limit translocation to the stems in soybean Plant Sci 270, 23-29. DOI:10.1016/j.plantsci.2018.02.007URL
WuC (1995). Heat shock transcription factors: structure and regulation Annu Rev Cell Dev Biol 11, 441-469. PMID:8689565 [本文引用: 1] Organisms respond to elevated temperatures and to chemical and physiological stresses by an increase in the synthesis of heat shock proteins. The regulation of heat shock gene expression in eukaryotes is mediated by the conserved heat shock transcription factor (HSF). HSF is present in a latent state under normal conditions; it is activated upon heat stress by induction of trimerization and high-affinity binding to DNA and by exposure of domains for transcriptional activity. Analysis of HSF cDNA clones from many species has defined structural and regulatory regions responsible for the inducible activities. The heat stress signal is thought to be transduced to HSF by changes in the physical environment, in the activity of HSF-modifying enzymes, or by changes in the intracellular level of heat shock proteins.
WuQ, ZhuXF, ZhaoXS, ShenRF (2020). Potassium affects cadmium resistance in Arabidopsis through facilitating root cell wall Cd retention in a nitric oxide dependent manner Environ Exp Bot 178, 104175. DOI:10.1016/j.envexpbot.2020.104175URL [本文引用: 1]
WuX, ChenJH, YueXM, WeiXN, ZouJW, ChenYH, SuNN, CuiJ (2019). The zinc-regulated protein (ZIP) family genes and glutathione S-transferase (GST) family genes play roles in Cd resistance and accumulation of pak choi ( Brassica campestris ssp. chinensis) Ecotoxicol Environ Saf 183, 109571. DOI:10.1016/j.ecoenv.2019.109571URL [本文引用: 2]
XuH, XuWZ, XiHM, MaWW, HeZY, MaM (2013). The ER luminal binding protein (BiP) alleviates Cd 2+-induced programmed cell death through endoplasmic reticulum stress- cell death signaling pathway in tobacco cells J Plant Physiol 170, 1434-1475. DOI:10.1016/j.jplph.2013.05.017URL [本文引用: 2]
XuL, WangY, ZhangF, TangMJ, ChenYL, WangJ, KaranjaBK, LuoXB, ZhangW, LiuLW (2017). Dissecting root proteome changes reveals new insight into cadmium stress response in radish ( Raphanus sativus L.) Plant Cell Physiol 58, 1901-1913. DOI:10.1093/pcp/pcx131URL [本文引用: 4]
XuL, ZhangF, TangMJ, WangY, DongJH, YingJL, ChenYL, HuB, LiC, LiuLW (2020). Melatonin confers cadmium tolerance by modulating critical heavy metal chelators and transporters in radish plants J Pineal Res 69, e12659. [本文引用: 2]
YadavSK (2010). Heavy metals toxicity in plants: an overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants S Afr J Bot 76, 167-179. DOI:10.1016/j.sajb.2009.10.007URL [本文引用: 3]
YamajiN, XiaJX, Mitani-UenoN, YokoshoK, FengMJ (2013). Preferential delivery of zinc to developing tissues in rice is mediated by P-type heavy metal ATPase OsHMA2 Plant Physiol 162, 927-939. DOI:10.1104/pp.113.216564URL [本文引用: 1]
YangT, PoovaiahBW (2002). Hydrogen peroxide homeostasis: activation of plant catalase by calcium/calmodulin Proc Natl Acad Sci USA 99, 4097-4102. DOI:10.1073/pnas.052564899URL [本文引用: 2]
YuanLY, YangSG, LiuBX, ZhangM, WuKQ (2012). Molecular characterization of a rice metal tolerance protein, OsMTP1 Plant Cell Rep 31, 67-79. DOI:10.1007/s00299-011-1140-9URL [本文引用: 2]
ZhangJ, ZhangM, SongHY, ZhaoJQ, ShabalaS, TianSK, YangXE (2020a). A novel plasma membrane-based NRAMP transporter contributes to Cd and Zn hyperac-cumulation in Sedum alfredii Hance Environ Exp Bot 176, 104121. DOI:10.1016/j.envexpbot.2020.104121URL [本文引用: 2]
ZhangLY, ZhangHY, GuoW, TianYL, ChenZS, WeiXF (2011). Photosynthetic responses of energy plant maize under cadmium contamination stress Adv Mat Res 356-360, 283-286. [本文引用: 1]
ZhangP, WangRL, JuQ, LiWQ, TranLSP, XuJ (2019a). The R2R3-MYB transcription factor MYB49 regulates cadmium accumulation Plant Physiol 180, 529-542. DOI:10.1104/pp.18.01380 [本文引用: 2] Abscisic acid (ABA) reduces accumulation of potentially toxic cadmium (Cd) in plants. How the ABA signal is transmitted to modulate Cd uptake remains largely unclear. Here, we report that the basic region/Leu zipper transcription factor ABSCISIC ACID-INSENSITIVE5 (ABI5), a central ABA signaling molecule, is involved in ABA-repressed Cd accumulation in plants by physically interacting with a previously uncharacterized R2R3-type MYB transcription factor, MYB49. Overexpression of the Cd-induced MYB49 gene in Arabidopsis (Arabidopsis thaliana) resulted in a significant increase in Cd accumulation, whereas myb49 knockout plants and plants expressing chimeric repressors of MYB49:ERF-associated amphiphilic repression motif repression domain (SRDX49) exhibited reduced accumulation of Cd. Further investigations revealed that MYB49 positively regulates the expression of the basic helix-loop-helix transcription factors bHLH38 and bHLH101 by directly binding to their promoters, leading to activation of IRON-REGULATED TRANSPORTER1, which encodes a metal transporter involved in Cd uptake. MYB49 also binds to the promoter regions of the heavy metal-associated isoprenylated plant proteins (HIPP22) and HIPP44, resulting in up-regulation of their expression and subsequent Cd accumulation. On the other hand, as a feedback mechanism to control Cd uptake and accumulation in plant cells, Cd-induced ABA up-regulates the expression of ABI5, whose protein product interacts with MYB49 and prevents its binding to the promoters of downstream genes, thereby reducing Cd accumulation. Our results provide new insights into the molecular feedback mechanisms underlying ABA signaling-controlled Cd uptake and accumulation in plants.
ZhangSM, YangC, ChenMM, ChenJ, PanYH, ChenYL, RahmanSU, FanJF, ZhangY (2019b). Influence of nitrogen availability on Cd accumulation and acclimation strategy of Populus leaves under Cd exposure Ecotoxicol Environ Saf 180, 439-448. DOI:10.1016/j.ecoenv.2019.05.031URL [本文引用: 3]
ZhangWW, YueSQ, SongJF, XunM, HanMY, YangHQ (2020b). MhNRAMP1 from Malus hupehensis exacerbates cell death by accelerating Cd uptake in tobacco and apple calli Front Plant Sci 11, 957. DOI:10.3389/fpls.2020.00957URL [本文引用: 2]
ZhouM, ZhengSG, LiuR, LuJ, LuL, ZhangCH, LiuZH, LuoCP, ZhangL, YantL, WuY (2019). Genome-wide identification, phylogenetic and expression analysis of the heat shock transcription factor family in bread wheat ( Triticum aestivum L.) BMC Genomics 20, 505. DOI:10.1186/s12864-019-5876-xPMID:31215411 [本文引用: 1] Environmental toxicity from non-essential heavy metals such as cadmium (Cd), which is released from human activities and other environmental causes, is rapidly increasing. Wheat can accumulate high levels of Cd in edible tissues, which poses a major hazard to human health. It has been reported that heat shock transcription factor A 4a (HsfA4a) of wheat and rice conferred Cd tolerance by upregulating metallothionein gene expression. However, genome-wide identification, classification, and comparative analysis of the Hsf family in wheat is lacking. Further, because of the promising role of Hsf genes in Cd tolerance, there is need for an understanding of the expression of this family and their functions on wheat under Cd stress. Therefore, here we identify the wheat TaHsf family and to begin to understand the molecular mechanisms mediated by the Hsf family under Cd stress.We first identified 78 putative Hsf homologs using the latest available wheat genome information, of which 38 belonged to class A, 16 to class B and 24 to class C subfamily. Then, we determined chromosome localizations, gene structures, conserved protein motifs, and phylogenetic relationships of these TaHsfs. Using RNA sequencing data over the course of development, we surveyed expression profiles of these TaHsfs during development and under different abiotic stresses to characterise the regulatory network of this family. Finally, we selected 13 TaHsf genes for expression level verification under Cd stress using qRT-PCR.To our knowledge, this is the first report of the genome organization, evolutionary features and expression profiles of the wheat Hsf gene family. This work therefore lays the foundation for targeted functional analysis of wheat Hsf genes, and contributes to a better understanding of the roles and regulatory mechanism of wheat Hsfs under Cd stress. 土壤镉污染对四种阔叶树苗木叶绿素荧光特性和生长的影响 1 2006
... 外源SA可改变ROS的积累和抗氧化酶系统活性, 缓解Cd胁迫带来的不利影响(Zawoznik et al., 2007).施加外源SA可使油菜(Brassica campestris)叶片的可溶性糖和游离脯氨酸含量升高、MDA积累减少, 质膜透性降低, CAT、POD和SOD活性升高, 明显缓解Cd毒害(胡春霞和王秀芹, 2010).SA处理显著增强玉米叶片中SOD活性, 降低CAT和POD活性, 并减轻Cd对APX活性的抑制作用(Krantev et al., 2008).此外, SA通过抑制ROS的过量产生缓解Cd引发的氧化损伤(Zawoznik et al., 2007); 施用外源SA可减少根系中ROS和MDA的积累, 进而减轻Cd毒性(Liu et al., 2016); 外源SA也可介导NADPH氧化酶产生H2O2, 以应对Cd胁迫引起的氧化应激(Chao et al., 2010).赵新月等(2013)研究表明, 与仅用Cd处理的玉米相比, 用50-250 μmol·L-1 SA处理玉米其叶绿素含量、硝酸还原酶(nitrite reductase, NR)、谷氨酰胺合成酶(glutamine synthetase, GS)和谷氨酸合成酶(glutamate synthase, GOGAT)活性显著升高, 氮素代谢过程良好, 玉米根系的POD活性降低(但POD和SOD活性与无Cd处理相比无显著差异), 且降低了Cd胁迫对质膜氧化系统的伤害.但外源SA对Cd胁迫修复的缓解作用受浓度影响较大, 低浓度的SA具有良好的修复效果, 高浓度的SA反而影响了光合色素的合成及酶活性, 抑制玉米的生长(赵新月等, 2013).重金属胁迫下, 外源SA提高了植物SOD、CAT和APX合成基因的表达(Gill, 2015; Song et al., 2019); 及水稻叶片和稻穗中低亲和离子转运子基因(OsLCT1)、水稻低Cd基因(OsLCD)和ZIP的时空表达(Liu et al., 2016). ...
植物防御素调控水稻镉积累的新机制 1 2018
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
... Cd处理的植物初级根尖和子叶中IAA浓度及分布有较大差异.Cd处理后, IAA含量显著降低(Guo et al., 2019), IAA合成基因表达下调(Bashir et al., 2019), IAA氧化酶活性增强, IAA生物合成和分解代谢基因表达改变(Hu et al., 2013).此外, Cd胁迫下, 外源IAA能显著提高玉米幼苗根和地上部生物量, 降低地上部Cd含量, 但对根Cd含量的影响不显著, 显著降低叶片的MDA和GSH含量, 使玉米幼苗叶片SOD、POD和CAT活性显著增强(陈晶等, 2016).当IAA合成受到抑制时, 如施加生长素信号抑制剂氯苯氧基异丁酸, 可有效抑制10 μmol·L-1 Cd诱导的根系生长和H2O2产生, 但不抑制60 μmol·L-1 Cd诱导的根系生长和H2O2产生(Tamás et al., 2012).Cd胁迫下施加IAA, 可激活谷胱甘肽-S-转移酶(glutathione- S-transferase, GST) (在植物解毒过程中起关键作用)活性, 使植物更好地发生应激反应(Bo?ová et al., 2013).Cd胁迫下, 施加外源IAA还可显著提高SODs基因的表达, 降低H2O2含量, 从而减少Cd对植物的毒害(Pan et al., 2020).重金属污染条件下, 外源IAA可促进天然抗性相关巨噬细胞蛋白基因NRAMP (NATURAL RESISTANCE-ASSOCIATED MACROPHAGE PROTEIN)、IRT、重金属ATP酶家族基因HMA (heavy metal ATPase)和锌铁转运蛋白基因ZIP (zinc iron transporter protein)的表达, 提高植物的抗Cd能力(Korshunova et al., 1999; 罗莎, 2017). ...
小麦根系特征对干旱胁迫的响应 1 2019
... 镉(Cd)因其在土壤中移动性强、毒性大、污染范围广和检出频率高等原因, 被联合国环境计划署(United Nations Environment Programme, UNEP)列入可引起人类癌症的致癌物质(UNEP, 2008; DalCorso et al., 2008; 付铄岚等, 2017).土壤中的Cd被植物吸收后, 会随着食物链在高营养级生物体内富集, 诱发人体骨质疏松并损坏肾脏, 导致人体慢性中毒, 甚至出现免疫系统、神经系统及生殖系统损伤等疾病, 对人类健康造成严重的威胁(Lalor, 2008; Clemens et al., 2013).由于Cd对各种环境及生物体的长期毒性作用, 其已成为全社会关注的焦点(Nagajyoti et al., 2010; Etesami and Jeong, 2018).有研究表明, 植物对Cd污染会产生一系列的应激反应, 如氧化应激、酶活性和植物信号物质(激素和钙离子)失衡, 导致光合系统受损、质膜过氧化、细胞损伤、酶活性改变、内质网胁迫、蛋白质降解、DNA损伤或突变, 从而影响其生理生化代谢过程, 最终使植物生长受到抑制, 甚至死亡(Sanità di Toppi and Gabbrielli, 1999; DalCorso et al., 2008; Nagajyoti et al., 2010; 杨正婷和刘建祥, 2016; Shahid et al., 2019; 张瑛等, 2019) (图1).此外, 植物对胁迫的响应是一个联合的信号通路和网络调控(郭倩倩和周文彬, 2019; 苗青霞等, 2019).目前, 国内外在植物对镉的吸收、积累和转运或镉对植物产生的毒害和抗镉生理及分子机制等方面已有大量研究, 而有关植物对重金属镉胁迫的主要代谢过程的应答机制报道较少.本文综述了植物响应镉胁迫的生理生化代谢变化及其应答机制, 为深入理解镉胁迫下植物的代谢差异、认识植物的抗镉策略, 及未来作物育种和环境修复提供理论依据. ...
高等植物重金属耐性与超积累特性及其分子机理研究 2 2005
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
... 镉(Cd)因其在土壤中移动性强、毒性大、污染范围广和检出频率高等原因, 被联合国环境计划署(United Nations Environment Programme, UNEP)列入可引起人类癌症的致癌物质(UNEP, 2008; DalCorso et al., 2008; 付铄岚等, 2017).土壤中的Cd被植物吸收后, 会随着食物链在高营养级生物体内富集, 诱发人体骨质疏松并损坏肾脏, 导致人体慢性中毒, 甚至出现免疫系统、神经系统及生殖系统损伤等疾病, 对人类健康造成严重的威胁(Lalor, 2008; Clemens et al., 2013).由于Cd对各种环境及生物体的长期毒性作用, 其已成为全社会关注的焦点(Nagajyoti et al., 2010; Etesami and Jeong, 2018).有研究表明, 植物对Cd污染会产生一系列的应激反应, 如氧化应激、酶活性和植物信号物质(激素和钙离子)失衡, 导致光合系统受损、质膜过氧化、细胞损伤、酶活性改变、内质网胁迫、蛋白质降解、DNA损伤或突变, 从而影响其生理生化代谢过程, 最终使植物生长受到抑制, 甚至死亡(Sanità di Toppi and Gabbrielli, 1999; DalCorso et al., 2008; Nagajyoti et al., 2010; 杨正婷和刘建祥, 2016; Shahid et al., 2019; 张瑛等, 2019) (图1).此外, 植物对胁迫的响应是一个联合的信号通路和网络调控(郭倩倩和周文彬, 2019; 苗青霞等, 2019).目前, 国内外在植物对镉的吸收、积累和转运或镉对植物产生的毒害和抗镉生理及分子机制等方面已有大量研究, 而有关植物对重金属镉胁迫的主要代谢过程的应答机制报道较少.本文综述了植物响应镉胁迫的生理生化代谢变化及其应答机制, 为深入理解镉胁迫下植物的代谢差异、认识植物的抗镉策略, 及未来作物育种和环境修复提供理论依据. ...
茶树金属耐受蛋白基因CsMTP11的克隆及功能分析 2 2017
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
... 镉(Cd)因其在土壤中移动性强、毒性大、污染范围广和检出频率高等原因, 被联合国环境计划署(United Nations Environment Programme, UNEP)列入可引起人类癌症的致癌物质(UNEP, 2008; DalCorso et al., 2008; 付铄岚等, 2017).土壤中的Cd被植物吸收后, 会随着食物链在高营养级生物体内富集, 诱发人体骨质疏松并损坏肾脏, 导致人体慢性中毒, 甚至出现免疫系统、神经系统及生殖系统损伤等疾病, 对人类健康造成严重的威胁(Lalor, 2008; Clemens et al., 2013).由于Cd对各种环境及生物体的长期毒性作用, 其已成为全社会关注的焦点(Nagajyoti et al., 2010; Etesami and Jeong, 2018).有研究表明, 植物对Cd污染会产生一系列的应激反应, 如氧化应激、酶活性和植物信号物质(激素和钙离子)失衡, 导致光合系统受损、质膜过氧化、细胞损伤、酶活性改变、内质网胁迫、蛋白质降解、DNA损伤或突变, 从而影响其生理生化代谢过程, 最终使植物生长受到抑制, 甚至死亡(Sanità di Toppi and Gabbrielli, 1999; DalCorso et al., 2008; Nagajyoti et al., 2010; 杨正婷和刘建祥, 2016; Shahid et al., 2019; 张瑛等, 2019) (图1).此外, 植物对胁迫的响应是一个联合的信号通路和网络调控(郭倩倩和周文彬, 2019; 苗青霞等, 2019).目前, 国内外在植物对镉的吸收、积累和转运或镉对植物产生的毒害和抗镉生理及分子机制等方面已有大量研究, 而有关植物对重金属镉胁迫的主要代谢过程的应答机制报道较少.本文综述了植物响应镉胁迫的生理生化代谢变化及其应答机制, 为深入理解镉胁迫下植物的代谢差异、认识植物的抗镉策略, 及未来作物育种和环境修复提供理论依据. ...
镉对玉米苗中钙调蛋白含量和Ca 2+-ATPase活性的影响 3 2008
... Ca2+作为细胞功能的第二信使, 参与调节植物各种逆境下的重要生理生化过程.钙信使系统包括Ca2+、钙调蛋白(CaM)和钙泵(Ca2+-ATPase).CaM是Ca2+的感受器, Ca2+-ATPase决定着Ca2+的跨膜运输(Snedden and Fromm, 2001).正常情况下三者的含量保持稳定, 可维持细胞正常的生理功能.在外界生物和非生物胁迫下, 细胞质中的Ca2+浓度会瞬间升高(Ca2+浓度的波动被认为是植物对外界刺激的反应信号) (Nürnberger and Scheel, 2001; Liu et al., 2017).Cd胁迫扰动植物细胞中的Ca2+水平, 从而扰乱植物叶片的水分状况(Perfus-Barbeoch et al., 2002).胞质Ca2+浓度增加对H2O2的生成有动态调控作用, Ca2+可调控NADH氧化酶活性, 上调植物中H2O2的含量; 也可通过刺激H2O2酶活性下调植物中的H2O2水平(Yang and Poovaiah, 2002).当ROS含量超过Ca2+调控的浓度范围时, 会对植物造成伤害, 细胞膜和细胞器膜发生膜质过氧化, 膜的通透性被破坏, 致使细胞死亡(赵士诚等, 2008).Cd与Ca2+的化学性质相似, Cd被植物吸收后通过金属转运体或钙通道运输至根部, 也可通过电压依赖的钙通道进入细胞, 一旦进入胞质, 它就会模拟钙离子的活性, 扰乱钙信号调控; 导致根系吸收大量的Cd, 并显著减少根长、根表面积、根体积和根系生物量, 使叶片黄化并干燥坏死(Yang and Poovaiah, 2002; Wang et al., 2016; Ismael et al., 2019).Cd进入植物根细胞也诱导一些对重金属胁迫有反应的特定信号分子, 如CaMs、钙依赖蛋白激酶(calcium-dependent protein kinases, CDPKs)和丝裂原活化蛋白激酶(mitogen-activated protein kinases, MAPKs), 调控植物对逆境的抗性(Danquah et al., 2014; Xu et al., 2017). ...
... 质膜上的Ca2+-ATPase能使高浓度的Ca2+进入非原生质体空间, 调节胁迫下细胞内的Ca2+平衡和维持细胞相对稳定的内环境(Snedden and Fromm, 2001; Hu et al., 2007).赵士诚等(2008)用不同浓度的Cd处理玉米叶片, 其Ca2+-ATPase活性均表现为细胞质膜>液泡膜、内质网膜>线粒体膜.并且低浓度Cd处理下, 为保持细胞内的Ca2+平衡, 玉米叶片生物膜系统的Ca2+-ATPase活性增强; 高浓度Cd处理下, Ca2+-ATPase活性下降, 出现代谢紊乱, 玉米中下部叶变黄且生长停滞.Ca预处理可显著降低扁豆(Lablab purpureus)幼苗的Cd积累, 并通过调节H2O2酶活性和降低膜的氧化损伤减轻Cd处理的不利影响(Talukdar, 2012).林啸等(2014)发现外源Ca处理可降低白菜(Brassica pekinensis)叶片的Cd含量, 并通过Ca2+-CaM信号途径调节酶类抗氧化剂APX、CAT和POD以及非酶类抗氧化剂GSH含量, 维持叶片ROS的平衡, 减少Cd诱导的细胞程序性死亡, 从而增强植物对Cd的耐受性.植物钙信号与抗氧化系统的关联机制值得进一步研究. ...
... Cd显著影响抗氧化酶类物质和非酶抗氧化物质的基因表达.不同浓度Cd处理的玉米幼苗, SOD基因的表达随着Cd浓度的升高先上升, 后随着时间的延长而下降; CAT基因的表达随着Cd浓度的升高和处理时间的延长逐渐增强(赵士诚等, 2008).高浓度Cd处理下, 抗氧化酶基因GR-chl、CAT3、APX-cyt、APX-per和MTLa的表达上调(Zhang et al., 2019b).当植物遭受Cd胁迫时, 金属硫蛋白(metallothionein)基因MTs、APX、GPX和DHAR等表达上调(Cobbett and Goldsbrough, 2002; Tamás et al., 2008).γ-谷氨酰半胱氨酸合成酶(γ-glutamylcysteine synthetase, γ-GCS)、谷胱甘肽合成酶(glutathione synthetase, GSHs)、胱硫醚β-合酶(cystathionine beta-synthase, CBS)、ATP硫酸化酶(ATP sulfurylase, ATPS)、GR、PCS和乙二醛酶(glyoxalase) I和II等合成基因是调节重金属耐受性的潜在候选基因(Yadav, 2010; Sofoet al., 2013).细胞壁果胶(pectin)在重金属结合中起重要作用, 根、茎和叶细胞壁的酯化及果胶酶修饰减少了Cd的积累.番茄经Cd处理后, 其细胞壁果胶相关基因SlQUA1和SlPME1表达上调, 果胶甲基脂酶活性明显升高(Jia et al., 2019). ...
外源水杨酸对镉胁迫下玉米幼苗的叶氮素代谢和根系抗氧化酶的影响 3 2013
... Responses of plant hormones to Cd stress Table 1
激素
激素合成基因
非酶物质
抗氧化酶
抗氧化物质基因
吸收和运输蛋白基因
ABA
NCED3和Glyma17G242200 (Bashir et al., 2019; Song et al., 2019)
AsA和GSH (Hu et al., 2005; Stroiński et al., 2010; Li et al., 2014)
APX、POD、SOD和CAT (Hu et al., 2005)
StPCS 1 (Stroiń- ski et al., 2010)
IRT1 (Fan et al., 2014)
IAA
Glyma02G037600.1 (Bashir et al., 2019)
GSH (Hu et al., 2013)
SOD、POD、CAT和GST (Bo?ová et al., 2013; 陈晶等, 2016)
SODs (Pan et al., 2020)
Nramp、IRT、HMA和ZIP (Korshunova et al., 1999; 罗莎, 2017)
SA
Glyma02G063400 (Bashir et al., 2019)
脯氨酸(Krantev et al., 2008; 胡春霞和王秀芹, 2010)
NR、GS和GOGAT (Zawoznik et al., 2007; Chao et al., 2010; 赵新月等, 2013)
SODS、CATS和APXS (Gill, 2015)
OsLCT1、OsLCD和ZIP (Liu et al., 2016)
JA
PtJMT1和Glyma11G007600. 1 (Bashir et al., 2019; Song et al., 2019)
GSH (Noriega et al., 2012; Zhao et al., 2016)
CAT、SOD和APX (Noriega et al., 2012; Zhao et al., 2016)
... 外源SA可改变ROS的积累和抗氧化酶系统活性, 缓解Cd胁迫带来的不利影响(Zawoznik et al., 2007).施加外源SA可使油菜(Brassica campestris)叶片的可溶性糖和游离脯氨酸含量升高、MDA积累减少, 质膜透性降低, CAT、POD和SOD活性升高, 明显缓解Cd毒害(胡春霞和王秀芹, 2010).SA处理显著增强玉米叶片中SOD活性, 降低CAT和POD活性, 并减轻Cd对APX活性的抑制作用(Krantev et al., 2008).此外, SA通过抑制ROS的过量产生缓解Cd引发的氧化损伤(Zawoznik et al., 2007); 施用外源SA可减少根系中ROS和MDA的积累, 进而减轻Cd毒性(Liu et al., 2016); 外源SA也可介导NADPH氧化酶产生H2O2, 以应对Cd胁迫引起的氧化应激(Chao et al., 2010).赵新月等(2013)研究表明, 与仅用Cd处理的玉米相比, 用50-250 μmol·L-1 SA处理玉米其叶绿素含量、硝酸还原酶(nitrite reductase, NR)、谷氨酰胺合成酶(glutamine synthetase, GS)和谷氨酸合成酶(glutamate synthase, GOGAT)活性显著升高, 氮素代谢过程良好, 玉米根系的POD活性降低(但POD和SOD活性与无Cd处理相比无显著差异), 且降低了Cd胁迫对质膜氧化系统的伤害.但外源SA对Cd胁迫修复的缓解作用受浓度影响较大, 低浓度的SA具有良好的修复效果, 高浓度的SA反而影响了光合色素的合成及酶活性, 抑制玉米的生长(赵新月等, 2013).重金属胁迫下, 外源SA提高了植物SOD、CAT和APX合成基因的表达(Gill, 2015; Song et al., 2019); 及水稻叶片和稻穗中低亲和离子转运子基因(OsLCT1)、水稻低Cd基因(OsLCD)和ZIP的时空表达(Liu et al., 2016). ...
... SA处理玉米其叶绿素含量、硝酸还原酶(nitrite reductase, NR)、谷氨酰胺合成酶(glutamine synthetase, GS)和谷氨酸合成酶(glutamate synthase, GOGAT)活性显著升高, 氮素代谢过程良好, 玉米根系的POD活性降低(但POD和SOD活性与无Cd处理相比无显著差异), 且降低了Cd胁迫对质膜氧化系统的伤害.但外源SA对Cd胁迫修复的缓解作用受浓度影响较大, 低浓度的SA具有良好的修复效果, 高浓度的SA反而影响了光合色素的合成及酶活性, 抑制玉米的生长(赵新月等, 2013).重金属胁迫下, 外源SA提高了植物SOD、CAT和APX合成基因的表达(Gill, 2015; Song et al., 2019); 及水稻叶片和稻穗中低亲和离子转运子基因(OsLCT1)、水稻低Cd基因(OsLCD)和ZIP的时空表达(Liu et al., 2016). ...
Sulfate improves cadmium tolerance by limiting cadmium accumulation, modulation of sulfur metabolism and antioxidant defense system in maize 1 2018
... Cd胁迫通常促进JA合成基因(Bashir et al., 2019)和抗氧化酶合成基因的转录(Keunen et al., 2013).低浓度JA可缓解Cd胁迫.例如, 用20 μmol·L-1 JA处理可有效改善Cd诱导的氧化应激, 使硫代巴比妥酸反应物质、H2O2和O2-.的积累减少, 使GSH、CAT、SOD及血红素氧合酶(HO)活性增强(Noriega et al., 2012; Adhikari et al., 2018).研究发现, 番茄(Solanum lycopersicum) JA缺失突变体对Cd的敏感性增强, 与光合作用相关参数(包括总叶绿素、光系统II实际效率、可变叶绿素荧光与最大荧光比及净光合速率)值降低; 相对水含量、可溶性糖和脯氨酸含量及淀粉的积累减少; 氧化应激和抗氧化防御作用(包括H2O2与MDA生成、电解质渗漏、SOD、APX和CAT活性)减弱(Zhao et al., 2016).外源茉莉酸甲酯(MeJA)通过降低拟南芥根系和茎叶中的Cd浓度, 下调参与Cd吸收和运输的AtIRT1、AtHMA2和AtHMA4基因表达, 缓解Cd引起的新叶毒害.JA合成关键基因AtAOS突变上调AtIRT1、AtHMA2和AtHMA4等转运蛋白基因表达, 增加根和芽中的Cd浓度, 提高拟南芥对Cd的敏感性.此外, MeJA降低Cd胁迫的拟南芥根尖中的NO水平(Lei et al., 2020).镉胁迫下植物激素ABA、IAA、SA和JA之间的相互作用值得深入探索. ...
MYB4 transcription factor, a member of R2R3-subfamily of MYB domain protein, regulates cadmium tolerance via enhanced protection against oxidative damage and increases expression of PCS1 and MT1C in Arabidopsis 1 2020
... 内质网(endoplasmic reticulum, ER)是一个复杂的代谢场所, 是调控蛋白质合成、信号转导和钙稳态的重要细胞器(Xu et al., 2013).所有分泌蛋白和大部分膜蛋白的合成和折叠及蛋白质的修饰和加工(主要包括糖基化、羟基化、酰基化和二硫键形成等)均在内质网进行(陈倩和谢旗, 2018).当植物受到干旱、高温、盐和重金属等胁迫后, 细胞蛋白质异常糖基化、Ca2+平衡紊乱及氧化还原状态改变会使细胞内积累大量未折叠或错误折叠的蛋白, 导致内质网胁迫(Kleizen and Braakman, 2004; Beaupere and Labunskyy, 2019).Cd诱导内质网应激, 改变内质网钙和脂质组成、ROS种类和错误折叠/未折叠蛋白(Rajakumar et al., 2016).分子伴侣参与内质网Cd胁迫的调控.BiP (binding protein)属于热休克蛋白(heat shock protein, HSP) HSP70家族, 是内质网腔内含量很高的分子伴侣, 参与内质网胁迫(Kleizen and Braakman, 2004).植物中, 程序性细胞死亡可通过ER调控应激-细胞死亡信号通路, BiP通过内质网应激-细胞死亡信号通路缓解Cd诱导的BY-2 (bright yellow-2)程序性死亡(Xu et al., 2013).同时, 内质网也是基因表达的重要场所.Wang等(2018)发现GmHMA3w主要在根内质网表达, 可将Cd从细胞壁运输至根内质网, 最终限制Cd从根向茎叶的转运.bZIP是位于根系质膜上的锌铁转运基因, 在拟南芥根细胞内质网应激信号转导中起重要作用(Deng et al., 2013). ...
Nitric oxide implication in cadmium-induced programmed cell death in roots and signaling response of yellow lupine plants 1 2012
... 活性氮(RNS)主要包括一氧化氮(NO)、S-亚硝基硫醇(SNOs)、高氮氧化物(NOx)、二硝基铁配合物和过氧亚硝酸盐(ONOO) 5种.过量的RNS会与脂类、蛋白质和核酸发生反应, 导致细胞损害(Neill et al., 2008a; Romero-Puertas and Sandalio, 2016).然而, 细胞内NO一旦产生, 就会与ROS迅速作用, 产生大量的RNS (如NO2), 然后降解为亚硝酸盐和硝酸钠(Neill et al., 2008b).低水平的NO可调节植物的发育, 而在胁迫条件下植物体内会产生大量的NO (Romero-Puertas and Sandalio, 2016; Liu et al., 2018); NO浓度过高则抑制植物的生长, 甚至引起细胞程序性死亡(Arasimowicz-Jelonek et al., 2012; Sami et al., 2018).低浓度NO促进植物生长, 高浓度则抑制植物生长, 其与生长素(indole-3-acetic acid, IAA)、细胞分裂素、赤霉素、脱落酸(abscisic acid, ABA)、乙烯、茉莉酸(jasmonic acid, JA)和水杨酸(salicylic acid, SA)也有一定的交互作用(Sami et al., 2018).Cd能通过破坏呼吸和光合作用的电子传递链使电子被传递到非靶向分子, 从而导致ROS和RNS的产生, 且Cd能通过抑制自由基降低相关酶活性(Ekmek?i et al., 2008; Romero-Puertas and Sandalio, 2016), 导致ROS和RNS不能够被及时清除而过量累积, 最终引起植物营养应激反应(Nieves-Cordones et al., 2019).外施氮(Zhang et al., 2019b)、磷(Wang et al., 2020)、钾(Wu et al., 2020)、钙(Li et al., 2021)和硅(Vaculík et al., 2015)等植物营养元素, 可起到调控植物激素含量、非酶物质和抗氧化酶活性及转运基因表达等作用, 减轻Cd对植物的毒害, 但外施营养物质对ROS和RNS有怎样的影响有待后续探索. ...
Interactive effect of drought and cadmium stress on soybean root morphology and gene expression 8 2019
... Cd胁迫下, 抗逆相关的植物激素(如ABA、IAA、SA和JA)均会发生显著变化, 且影响抗氧化物质合成相关基因和运输蛋白基因的表达(表1).多数情况下Cd胁迫可引起植物体内IAA、SA、ABA和JA含量变化(Liu et al., 2016; Bashir et al., 2019; Song et al., 2019).添加适量外源ABA、IAA、SA和JA等可缓解Cd胁迫对植物的危害. ...
... Responses of plant hormones to Cd stress Table 1
激素
激素合成基因
非酶物质
抗氧化酶
抗氧化物质基因
吸收和运输蛋白基因
ABA
NCED3和Glyma17G242200 (Bashir et al., 2019; Song et al., 2019)
AsA和GSH (Hu et al., 2005; Stroiński et al., 2010; Li et al., 2014)
APX、POD、SOD和CAT (Hu et al., 2005)
StPCS 1 (Stroiń- ski et al., 2010)
IRT1 (Fan et al., 2014)
IAA
Glyma02G037600.1 (Bashir et al., 2019)
GSH (Hu et al., 2013)
SOD、POD、CAT和GST (Bo?ová et al., 2013; 陈晶等, 2016)
SODs (Pan et al., 2020)
Nramp、IRT、HMA和ZIP (Korshunova et al., 1999; 罗莎, 2017)
SA
Glyma02G063400 (Bashir et al., 2019)
脯氨酸(Krantev et al., 2008; 胡春霞和王秀芹, 2010)
NR、GS和GOGAT (Zawoznik et al., 2007; Chao et al., 2010; 赵新月等, 2013)
SODS、CATS和APXS (Gill, 2015)
OsLCT1、OsLCD和ZIP (Liu et al., 2016)
JA
PtJMT1和Glyma11G007600. 1 (Bashir et al., 2019; Song et al., 2019)
GSH (Noriega et al., 2012; Zhao et al., 2016)
CAT、SOD和APX (Noriega et al., 2012; Zhao et al., 2016)
... Cd处理的植物初级根尖和子叶中IAA浓度及分布有较大差异.Cd处理后, IAA含量显著降低(Guo et al., 2019), IAA合成基因表达下调(Bashir et al., 2019), IAA氧化酶活性增强, IAA生物合成和分解代谢基因表达改变(Hu et al., 2013).此外, Cd胁迫下, 外源IAA能显著提高玉米幼苗根和地上部生物量, 降低地上部Cd含量, 但对根Cd含量的影响不显著, 显著降低叶片的MDA和GSH含量, 使玉米幼苗叶片SOD、POD和CAT活性显著增强(陈晶等, 2016).当IAA合成受到抑制时, 如施加生长素信号抑制剂氯苯氧基异丁酸, 可有效抑制10 μmol·L-1 Cd诱导的根系生长和H2O2产生, 但不抑制60 μmol·L-1 Cd诱导的根系生长和H2O2产生(Tamás et al., 2012).Cd胁迫下施加IAA, 可激活谷胱甘肽-S-转移酶(glutathione- S-transferase, GST) (在植物解毒过程中起关键作用)活性, 使植物更好地发生应激反应(Bo?ová et al., 2013).Cd胁迫下, 施加外源IAA还可显著提高SODs基因的表达, 降低H2O2含量, 从而减少Cd对植物的毒害(Pan et al., 2020).重金属污染条件下, 外源IAA可促进天然抗性相关巨噬细胞蛋白基因NRAMP (NATURAL RESISTANCE-ASSOCIATED MACROPHAGE PROTEIN)、IRT、重金属ATP酶家族基因HMA (heavy metal ATPase)和锌铁转运蛋白基因ZIP (zinc iron transporter protein)的表达, 提高植物的抗Cd能力(Korshunova et al., 1999; 罗莎, 2017). ...
Transcription factor GmWRKY142 confers cadmium resistance by up- regulating the cadmium tolerance 1-like genes 2 2020
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
... 外源SA可改变ROS的积累和抗氧化酶系统活性, 缓解Cd胁迫带来的不利影响(Zawoznik et al., 2007).施加外源SA可使油菜(Brassica campestris)叶片的可溶性糖和游离脯氨酸含量升高、MDA积累减少, 质膜透性降低, CAT、POD和SOD活性升高, 明显缓解Cd毒害(胡春霞和王秀芹, 2010).SA处理显著增强玉米叶片中SOD活性, 降低CAT和POD活性, 并减轻Cd对APX活性的抑制作用(Krantev et al., 2008).此外, SA通过抑制ROS的过量产生缓解Cd引发的氧化损伤(Zawoznik et al., 2007); 施用外源SA可减少根系中ROS和MDA的积累, 进而减轻Cd毒性(Liu et al., 2016); 外源SA也可介导NADPH氧化酶产生H2O2, 以应对Cd胁迫引起的氧化应激(Chao et al., 2010).赵新月等(2013)研究表明, 与仅用Cd处理的玉米相比, 用50-250 μmol·L-1 SA处理玉米其叶绿素含量、硝酸还原酶(nitrite reductase, NR)、谷氨酰胺合成酶(glutamine synthetase, GS)和谷氨酸合成酶(glutamate synthase, GOGAT)活性显著升高, 氮素代谢过程良好, 玉米根系的POD活性降低(但POD和SOD活性与无Cd处理相比无显著差异), 且降低了Cd胁迫对质膜氧化系统的伤害.但外源SA对Cd胁迫修复的缓解作用受浓度影响较大, 低浓度的SA具有良好的修复效果, 高浓度的SA反而影响了光合色素的合成及酶活性, 抑制玉米的生长(赵新月等, 2013).重金属胁迫下, 外源SA提高了植物SOD、CAT和APX合成基因的表达(Gill, 2015; Song et al., 2019); 及水稻叶片和稻穗中低亲和离子转运子基因(OsLCT1)、水稻低Cd基因(OsLCD)和ZIP的时空表达(Liu et al., 2016). ...
SaHsfA4c from Sedum alfredii hance enhances cadmium tolerance by regulating ROS-scavenger activities and heat shock proteins expression 2 2020
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
How plants cope with cadmium: staking all on metabolism and gene expression 2 2008
... 镉(Cd)因其在土壤中移动性强、毒性大、污染范围广和检出频率高等原因, 被联合国环境计划署(United Nations Environment Programme, UNEP)列入可引起人类癌症的致癌物质(UNEP, 2008; DalCorso et al., 2008; 付铄岚等, 2017).土壤中的Cd被植物吸收后, 会随着食物链在高营养级生物体内富集, 诱发人体骨质疏松并损坏肾脏, 导致人体慢性中毒, 甚至出现免疫系统、神经系统及生殖系统损伤等疾病, 对人类健康造成严重的威胁(Lalor, 2008; Clemens et al., 2013).由于Cd对各种环境及生物体的长期毒性作用, 其已成为全社会关注的焦点(Nagajyoti et al., 2010; Etesami and Jeong, 2018).有研究表明, 植物对Cd污染会产生一系列的应激反应, 如氧化应激、酶活性和植物信号物质(激素和钙离子)失衡, 导致光合系统受损、质膜过氧化、细胞损伤、酶活性改变、内质网胁迫、蛋白质降解、DNA损伤或突变, 从而影响其生理生化代谢过程, 最终使植物生长受到抑制, 甚至死亡(Sanità di Toppi and Gabbrielli, 1999; DalCorso et al., 2008; Nagajyoti et al., 2010; 杨正婷和刘建祥, 2016; Shahid et al., 2019; 张瑛等, 2019) (图1).此外, 植物对胁迫的响应是一个联合的信号通路和网络调控(郭倩倩和周文彬, 2019; 苗青霞等, 2019).目前, 国内外在植物对镉的吸收、积累和转运或镉对植物产生的毒害和抗镉生理及分子机制等方面已有大量研究, 而有关植物对重金属镉胁迫的主要代谢过程的应答机制报道较少.本文综述了植物响应镉胁迫的生理生化代谢变化及其应答机制, 为深入理解镉胁迫下植物的代谢差异、认识植物的抗镉策略, 及未来作物育种和环境修复提供理论依据. ...
... ; DalCorso et al., 2008; Nagajyoti et al., 2010; 杨正婷和刘建祥, 2016; Shahid et al., 2019; 张瑛等, 2019) (图1).此外, 植物对胁迫的响应是一个联合的信号通路和网络调控(郭倩倩和周文彬, 2019; 苗青霞等, 2019).目前, 国内外在植物对镉的吸收、积累和转运或镉对植物产生的毒害和抗镉生理及分子机制等方面已有大量研究, 而有关植物对重金属镉胁迫的主要代谢过程的应答机制报道较少.本文综述了植物响应镉胁迫的生理生化代谢变化及其应答机制, 为深入理解镉胁迫下植物的代谢差异、认识植物的抗镉策略, 及未来作物育种和环境修复提供理论依据. ...
The role of ABA and MAPK signaling pathways in plant abiotic stress responses 1 2014
... Ca2+作为细胞功能的第二信使, 参与调节植物各种逆境下的重要生理生化过程.钙信使系统包括Ca2+、钙调蛋白(CaM)和钙泵(Ca2+-ATPase).CaM是Ca2+的感受器, Ca2+-ATPase决定着Ca2+的跨膜运输(Snedden and Fromm, 2001).正常情况下三者的含量保持稳定, 可维持细胞正常的生理功能.在外界生物和非生物胁迫下, 细胞质中的Ca2+浓度会瞬间升高(Ca2+浓度的波动被认为是植物对外界刺激的反应信号) (Nürnberger and Scheel, 2001; Liu et al., 2017).Cd胁迫扰动植物细胞中的Ca2+水平, 从而扰乱植物叶片的水分状况(Perfus-Barbeoch et al., 2002).胞质Ca2+浓度增加对H2O2的生成有动态调控作用, Ca2+可调控NADH氧化酶活性, 上调植物中H2O2的含量; 也可通过刺激H2O2酶活性下调植物中的H2O2水平(Yang and Poovaiah, 2002).当ROS含量超过Ca2+调控的浓度范围时, 会对植物造成伤害, 细胞膜和细胞器膜发生膜质过氧化, 膜的通透性被破坏, 致使细胞死亡(赵士诚等, 2008).Cd与Ca2+的化学性质相似, Cd被植物吸收后通过金属转运体或钙通道运输至根部, 也可通过电压依赖的钙通道进入细胞, 一旦进入胞质, 它就会模拟钙离子的活性, 扰乱钙信号调控; 导致根系吸收大量的Cd, 并显著减少根长、根表面积、根体积和根系生物量, 使叶片黄化并干燥坏死(Yang and Poovaiah, 2002; Wang et al., 2016; Ismael et al., 2019).Cd进入植物根细胞也诱导一些对重金属胁迫有反应的特定信号分子, 如CaMs、钙依赖蛋白激酶(calcium-dependent protein kinases, CDPKs)和丝裂原活化蛋白激酶(mitogen-activated protein kinases, MAPKs), 调控植物对逆境的抗性(Danquah et al., 2014; Xu et al., 2017). ...
Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants 2 2014
... 活性氧(reactive oxygen species, ROS)和活性氮(reactive nitrogen species, RNS)是植物产生的具有一定功能的活性分子化合物, 是植物适应环境变化的主要调节因子.低含量水平下, ROS和RNS是信号分子(Romero-Puertas and Sandalio, 2016); 过多积累后, 会诱导植物氧化损伤及细胞程序性死亡, 是植物响应胁迫的关键指标(Das and Roychoudhury, 2014; Nieves-Cordones et al., 2019).Cd毒害最常见的表现是诱导植物出现氧化应激反应, 如产生ROS和RNS (Romero-Puertas et al., 2019).ROS和RNS之间存在动态平衡(Delledonne et al., 2001).ROS通过多种反应产生, 包括呼吸和光合电子传递链及酶(如过氧化物酶)的副反应等; Cd胁迫下, 植物细胞中产生ROS的部位有线粒体、叶绿体、过氧化物酶体、细胞质以及外质体等(Romero-Puertas et al., 2019).ROS包括单线态氧(single oxygen, 1O2)、超氧自由基阴离子(superoxide radical, O2-.)、过氧化氢(hydrogen peroxide, H2O2)和羟基自由基(hydroxyl radical, ·OH) (Nieves-Cordones et al., 2019).清除ROS的机制涉及酶促抗氧化剂和非酶类抗氧化剂(1.1.3节).ROS会对脂质、蛋白质和DNA等生物分子造成损害, 破坏细胞的完整性, 最终导致细胞死亡(Das and Roychoudhury, 2014).ROS作为信号分子在多种生命过程中扮演重要角色, 包括趋向性、细胞死亡和细胞对生物和非生物胁迫的反应(Mittler et al., 2011). ...
... ).清除ROS的机制涉及酶促抗氧化剂和非酶类抗氧化剂(1.1.3节).ROS会对脂质、蛋白质和DNA等生物分子造成损害, 破坏细胞的完整性, 最终导致细胞死亡(Das and Roychoudhury, 2014).ROS作为信号分子在多种生命过程中扮演重要角色, 包括趋向性、细胞死亡和细胞对生物和非生物胁迫的反应(Mittler et al., 2011). ...
Signal interactions between nitric oxide and reactive oxygen intermediates in the plant hypersensitive disease resistance response 1 2001
... 活性氧(reactive oxygen species, ROS)和活性氮(reactive nitrogen species, RNS)是植物产生的具有一定功能的活性分子化合物, 是植物适应环境变化的主要调节因子.低含量水平下, ROS和RNS是信号分子(Romero-Puertas and Sandalio, 2016); 过多积累后, 会诱导植物氧化损伤及细胞程序性死亡, 是植物响应胁迫的关键指标(Das and Roychoudhury, 2014; Nieves-Cordones et al., 2019).Cd毒害最常见的表现是诱导植物出现氧化应激反应, 如产生ROS和RNS (Romero-Puertas et al., 2019).ROS和RNS之间存在动态平衡(Delledonne et al., 2001).ROS通过多种反应产生, 包括呼吸和光合电子传递链及酶(如过氧化物酶)的副反应等; Cd胁迫下, 植物细胞中产生ROS的部位有线粒体、叶绿体、过氧化物酶体、细胞质以及外质体等(Romero-Puertas et al., 2019).ROS包括单线态氧(single oxygen, 1O2)、超氧自由基阴离子(superoxide radical, O2-.)、过氧化氢(hydrogen peroxide, H2O2)和羟基自由基(hydroxyl radical, ·OH) (Nieves-Cordones et al., 2019).清除ROS的机制涉及酶促抗氧化剂和非酶类抗氧化剂(1.1.3节).ROS会对脂质、蛋白质和DNA等生物分子造成损害, 破坏细胞的完整性, 最终导致细胞死亡(Das and Roychoudhury, 2014).ROS作为信号分子在多种生命过程中扮演重要角色, 包括趋向性、细胞死亡和细胞对生物和非生物胁迫的反应(Mittler et al., 2011). ...
Protein kinase and ribonuclease domains of IRE1 confer stress tolerance, vegetative growth, and reproductive development in Arabidopsis 1 2013
... 内质网(endoplasmic reticulum, ER)是一个复杂的代谢场所, 是调控蛋白质合成、信号转导和钙稳态的重要细胞器(Xu et al., 2013).所有分泌蛋白和大部分膜蛋白的合成和折叠及蛋白质的修饰和加工(主要包括糖基化、羟基化、酰基化和二硫键形成等)均在内质网进行(陈倩和谢旗, 2018).当植物受到干旱、高温、盐和重金属等胁迫后, 细胞蛋白质异常糖基化、Ca2+平衡紊乱及氧化还原状态改变会使细胞内积累大量未折叠或错误折叠的蛋白, 导致内质网胁迫(Kleizen and Braakman, 2004; Beaupere and Labunskyy, 2019).Cd诱导内质网应激, 改变内质网钙和脂质组成、ROS种类和错误折叠/未折叠蛋白(Rajakumar et al., 2016).分子伴侣参与内质网Cd胁迫的调控.BiP (binding protein)属于热休克蛋白(heat shock protein, HSP) HSP70家族, 是内质网腔内含量很高的分子伴侣, 参与内质网胁迫(Kleizen and Braakman, 2004).植物中, 程序性细胞死亡可通过ER调控应激-细胞死亡信号通路, BiP通过内质网应激-细胞死亡信号通路缓解Cd诱导的BY-2 (bright yellow-2)程序性死亡(Xu et al., 2013).同时, 内质网也是基因表达的重要场所.Wang等(2018)发现GmHMA3w主要在根内质网表达, 可将Cd从细胞壁运输至根内质网, 最终限制Cd从根向茎叶的转运.bZIP是位于根系质膜上的锌铁转运基因, 在拟南芥根细胞内质网应激信号转导中起重要作用(Deng et al., 2013). ...
MYB transcription factors in Arabidopsis 2 2010
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
... 活性氮(RNS)主要包括一氧化氮(NO)、S-亚硝基硫醇(SNOs)、高氮氧化物(NOx)、二硝基铁配合物和过氧亚硝酸盐(ONOO) 5种.过量的RNS会与脂类、蛋白质和核酸发生反应, 导致细胞损害(Neill et al., 2008a; Romero-Puertas and Sandalio, 2016).然而, 细胞内NO一旦产生, 就会与ROS迅速作用, 产生大量的RNS (如NO2), 然后降解为亚硝酸盐和硝酸钠(Neill et al., 2008b).低水平的NO可调节植物的发育, 而在胁迫条件下植物体内会产生大量的NO (Romero-Puertas and Sandalio, 2016; Liu et al., 2018); NO浓度过高则抑制植物的生长, 甚至引起细胞程序性死亡(Arasimowicz-Jelonek et al., 2012; Sami et al., 2018).低浓度NO促进植物生长, 高浓度则抑制植物生长, 其与生长素(indole-3-acetic acid, IAA)、细胞分裂素、赤霉素、脱落酸(abscisic acid, ABA)、乙烯、茉莉酸(jasmonic acid, JA)和水杨酸(salicylic acid, SA)也有一定的交互作用(Sami et al., 2018).Cd能通过破坏呼吸和光合作用的电子传递链使电子被传递到非靶向分子, 从而导致ROS和RNS的产生, 且Cd能通过抑制自由基降低相关酶活性(Ekmek?i et al., 2008; Romero-Puertas and Sandalio, 2016), 导致ROS和RNS不能够被及时清除而过量累积, 最终引起植物营养应激反应(Nieves-Cordones et al., 2019).外施氮(Zhang et al., 2019b)、磷(Wang et al., 2020)、钾(Wu et al., 2020)、钙(Li et al., 2021)和硅(Vaculík et al., 2015)等植物营养元素, 可起到调控植物激素含量、非酶物质和抗氧化酶活性及转运基因表达等作用, 减轻Cd对植物的毒害, 但外施营养物质对ROS和RNS有怎样的影响有待后续探索. ...
Exogenous abscisic acid application decreases cadmium accumulation in Arabidopsis plants, which is associated with the inhibition of IRT1-mediated cadmium uptake 2 2014
... Responses of plant hormones to Cd stress Table 1
激素
激素合成基因
非酶物质
抗氧化酶
抗氧化物质基因
吸收和运输蛋白基因
ABA
NCED3和Glyma17G242200 (Bashir et al., 2019; Song et al., 2019)
AsA和GSH (Hu et al., 2005; Stroiński et al., 2010; Li et al., 2014)
APX、POD、SOD和CAT (Hu et al., 2005)
StPCS 1 (Stroiń- ski et al., 2010)
IRT1 (Fan et al., 2014)
IAA
Glyma02G037600.1 (Bashir et al., 2019)
GSH (Hu et al., 2013)
SOD、POD、CAT和GST (Bo?ová et al., 2013; 陈晶等, 2016)
SODs (Pan et al., 2020)
Nramp、IRT、HMA和ZIP (Korshunova et al., 1999; 罗莎, 2017)
SA
Glyma02G063400 (Bashir et al., 2019)
脯氨酸(Krantev et al., 2008; 胡春霞和王秀芹, 2010)
NR、GS和GOGAT (Zawoznik et al., 2007; Chao et al., 2010; 赵新月等, 2013)
SODS、CATS和APXS (Gill, 2015)
OsLCT1、OsLCD和ZIP (Liu et al., 2016)
JA
PtJMT1和Glyma11G007600. 1 (Bashir et al., 2019; Song et al., 2019)
GSH (Noriega et al., 2012; Zhao et al., 2016)
CAT、SOD和APX (Noriega et al., 2012; Zhao et al., 2016)
Isolation and characterization of a novel cadmium-regulated Yellow Stripe-Like transporter (SnYSL3) in Solanum nigrum 2 2017
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
Cadmium stress increases antioxidant enzyme activities and decreases endogenous hormone concentrations more in Cd-tolerant than Cd-sensitive wheat varieties 3 2019
... Cd处理的植物初级根尖和子叶中IAA浓度及分布有较大差异.Cd处理后, IAA含量显著降低(Guo et al., 2019), IAA合成基因表达下调(Bashir et al., 2019), IAA氧化酶活性增强, IAA生物合成和分解代谢基因表达改变(Hu et al., 2013).此外, Cd胁迫下, 外源IAA能显著提高玉米幼苗根和地上部生物量, 降低地上部Cd含量, 但对根Cd含量的影响不显著, 显著降低叶片的MDA和GSH含量, 使玉米幼苗叶片SOD、POD和CAT活性显著增强(陈晶等, 2016).当IAA合成受到抑制时, 如施加生长素信号抑制剂氯苯氧基异丁酸, 可有效抑制10 μmol·L-1 Cd诱导的根系生长和H2O2产生, 但不抑制60 μmol·L-1 Cd诱导的根系生长和H2O2产生(Tamás et al., 2012).Cd胁迫下施加IAA, 可激活谷胱甘肽-S-转移酶(glutathione- S-transferase, GST) (在植物解毒过程中起关键作用)活性, 使植物更好地发生应激反应(Bo?ová et al., 2013).Cd胁迫下, 施加外源IAA还可显著提高SODs基因的表达, 降低H2O2含量, 从而减少Cd对植物的毒害(Pan et al., 2020).重金属污染条件下, 外源IAA可促进天然抗性相关巨噬细胞蛋白基因NRAMP (NATURAL RESISTANCE-ASSOCIATED MACROPHAGE PROTEIN)、IRT、重金属ATP酶家族基因HMA (heavy metal ATPase)和锌铁转运蛋白基因ZIP (zinc iron transporter protein)的表达, 提高植物的抗Cd能力(Korshunova et al., 1999; 罗莎, 2017). ...
Genome-wide analysis of heat shock transcription factor families in rice and Arabidopsis 1 2008
... HSP不仅能对抗应激相关损伤与保护细胞, 而且在蛋白质规则折叠、细胞内分布和降解中也发挥重要作用.这些功能有利于生物在压力条件下生存(Hartl and Hayer-Hart, 2002; Zhou et al., 2019).热休克转录因子HSFs (heat shock transcription factors)调节HSP的表达, 参与蛋白稳态的各个方面(如受损蛋白的重新折叠、组装和运输), 维持蛋白的稳定性(Wu, 1995; Guo et al., 2008).TaHsfA4a上调表达的水稻对Cd的耐受性增强, OsHsfA4a下调表达的水稻对Cd的耐受性减弱, 在小麦和水稻根系中, Cd胁迫导致HsfA4a以及金属硫蛋白基因的表达上调(Shim et al., 2009).此外, Chen等(2020)研究结果也表明, SaHsfA4c通过上调HSP的表达和提高ROS清除酶活性, 增强植物的抗逆性.Cd胁迫下, HSF/HSP参与柳枝稷(Panicum virgatum)正常蛋白质构象的重建以及细胞内稳态调节(Song et al., 2018).这些HSF/HSP表达基因可作为进一步研究的候选基因, 以提高植物对Cd的耐受性. ...
Molecular chaperones in the cytosol: from nascent chain to folded protein 1 2002
... HSP不仅能对抗应激相关损伤与保护细胞, 而且在蛋白质规则折叠、细胞内分布和降解中也发挥重要作用.这些功能有利于生物在压力条件下生存(Hartl and Hayer-Hart, 2002; Zhou et al., 2019).热休克转录因子HSFs (heat shock transcription factors)调节HSP的表达, 参与蛋白稳态的各个方面(如受损蛋白的重新折叠、组装和运输), 维持蛋白的稳定性(Wu, 1995; Guo et al., 2008).TaHsfA4a上调表达的水稻对Cd的耐受性增强, OsHsfA4a下调表达的水稻对Cd的耐受性减弱, 在小麦和水稻根系中, Cd胁迫导致HsfA4a以及金属硫蛋白基因的表达上调(Shim et al., 2009).此外, Chen等(2020)研究结果也表明, SaHsfA4c通过上调HSP的表达和提高ROS清除酶活性, 增强植物的抗逆性.Cd胁迫下, HSF/HSP参与柳枝稷(Panicum virgatum)正常蛋白质构象的重建以及细胞内稳态调节(Song et al., 2018).这些HSF/HSP表达基因可作为进一步研究的候选基因, 以提高植物对Cd的耐受性. ...
The role of ZmWRKY4 in regulating maize antioxidant defense under cadmium stress 2 2017
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
... 转录因子(transcription factors, TFs)是一类DNA结合蛋白, 参与植物的各种生理过程, 涉及生长、发育、自我应激信号转导或与不同的基因和转录因子交叉调节(Erpen et al., 2018).转录因子可通过控制下游基因的表达调控Cd胁迫反应, 是Cd解毒和耐受调控网络中的核心成分.目前, 已发现植物中大多数类型的Cd解毒和耐受转录因子.其中, WRKY、MYB和HSF等转录因子在调节植物Cd胁迫响应方面具有重要作用.WRKY TFs是植物中最大的转录因子家族之一, 拥有强大的转录调控功能(ülker and Somssich, 2004; Erpen et al., 2018).WRKY TFs不仅可通过调节植物激素信号转导途径, 还可结合其靶基因启动子通过激活或抑制下游基因的表达来调节它们的应激反应(Rushton et al., 2010).Hong等(2017)研究发现, Cd胁迫下玉米ZmWRKY4基因的表达上调, SOD和APX活性升高.ZmWRKY4在玉米叶肉细胞原生质体中的瞬时表达和RNA干扰(RNAi)进一步表明, ZmWRKY4是ABA诱导SOD和APX表达及活性增强所必需; ZmWRKY4在原生质体中的过表达上调了抗氧化酶的表达和活性, 而ABA诱导的抗氧化酶的表达和活性增强被ZmWRKY4的RNAi基因沉默所阻断.Cai等(2020)对大豆(Glycine max)的研究表明, GmWRKY4直接作用于ATCDT1、GmCDT1-1和GmCDT1-2, 从而减少Cd的摄取并积极调节对Cd胁迫的耐受性.GmWRKY142-GmCDT1-1/2级联模块为降低Cd在大豆中积累提供了潜在策略. ...
Abscisic acid-induced apoplastic H2O2 accumulation up-regulates the activities of chloroplastic and cytosolic antioxidant enzymes in maize leaves 3 2005
... Responses of plant hormones to Cd stress Table 1
激素
激素合成基因
非酶物质
抗氧化酶
抗氧化物质基因
吸收和运输蛋白基因
ABA
NCED3和Glyma17G242200 (Bashir et al., 2019; Song et al., 2019)
AsA和GSH (Hu et al., 2005; Stroiński et al., 2010; Li et al., 2014)
APX、POD、SOD和CAT (Hu et al., 2005)
StPCS 1 (Stroiń- ski et al., 2010)
IRT1 (Fan et al., 2014)
IAA
Glyma02G037600.1 (Bashir et al., 2019)
GSH (Hu et al., 2013)
SOD、POD、CAT和GST (Bo?ová et al., 2013; 陈晶等, 2016)
SODs (Pan et al., 2020)
Nramp、IRT、HMA和ZIP (Korshunova et al., 1999; 罗莎, 2017)
SA
Glyma02G063400 (Bashir et al., 2019)
脯氨酸(Krantev et al., 2008; 胡春霞和王秀芹, 2010)
NR、GS和GOGAT (Zawoznik et al., 2007; Chao et al., 2010; 赵新月等, 2013)
SODS、CATS和APXS (Gill, 2015)
OsLCT1、OsLCD和ZIP (Liu et al., 2016)
JA
PtJMT1和Glyma11G007600. 1 (Bashir et al., 2019; Song et al., 2019)
GSH (Noriega et al., 2012; Zhao et al., 2016)
CAT、SOD和APX (Noriega et al., 2012; Zhao et al., 2016)
Calcium-calmodulin is required for abscisic acid- induced antioxidant defense and functions both upstream and downstream of H2O2 production in leaves of maize ( Zea mays) plants 1 2007
... 质膜上的Ca2+-ATPase能使高浓度的Ca2+进入非原生质体空间, 调节胁迫下细胞内的Ca2+平衡和维持细胞相对稳定的内环境(Snedden and Fromm, 2001; Hu et al., 2007).赵士诚等(2008)用不同浓度的Cd处理玉米叶片, 其Ca2+-ATPase活性均表现为细胞质膜>液泡膜、内质网膜>线粒体膜.并且低浓度Cd处理下, 为保持细胞内的Ca2+平衡, 玉米叶片生物膜系统的Ca2+-ATPase活性增强; 高浓度Cd处理下, Ca2+-ATPase活性下降, 出现代谢紊乱, 玉米中下部叶变黄且生长停滞.Ca预处理可显著降低扁豆(Lablab purpureus)幼苗的Cd积累, 并通过调节H2O2酶活性和降低膜的氧化损伤减轻Cd处理的不利影响(Talukdar, 2012).林啸等(2014)发现外源Ca处理可降低白菜(Brassica pekinensis)叶片的Cd含量, 并通过Ca2+-CaM信号途径调节酶类抗氧化剂APX、CAT和POD以及非酶类抗氧化剂GSH含量, 维持叶片ROS的平衡, 减少Cd诱导的细胞程序性死亡, 从而增强植物对Cd的耐受性.植物钙信号与抗氧化系统的关联机制值得进一步研究. ...
Cadmium interferes with maintenance of auxin homeostasis in Arabidopsis seedlings 2 2013
... Responses of plant hormones to Cd stress Table 1
激素
激素合成基因
非酶物质
抗氧化酶
抗氧化物质基因
吸收和运输蛋白基因
ABA
NCED3和Glyma17G242200 (Bashir et al., 2019; Song et al., 2019)
AsA和GSH (Hu et al., 2005; Stroiński et al., 2010; Li et al., 2014)
APX、POD、SOD和CAT (Hu et al., 2005)
StPCS 1 (Stroiń- ski et al., 2010)
IRT1 (Fan et al., 2014)
IAA
Glyma02G037600.1 (Bashir et al., 2019)
GSH (Hu et al., 2013)
SOD、POD、CAT和GST (Bo?ová et al., 2013; 陈晶等, 2016)
SODs (Pan et al., 2020)
Nramp、IRT、HMA和ZIP (Korshunova et al., 1999; 罗莎, 2017)
SA
Glyma02G063400 (Bashir et al., 2019)
脯氨酸(Krantev et al., 2008; 胡春霞和王秀芹, 2010)
NR、GS和GOGAT (Zawoznik et al., 2007; Chao et al., 2010; 赵新月等, 2013)
SODS、CATS和APXS (Gill, 2015)
OsLCT1、OsLCD和ZIP (Liu et al., 2016)
JA
PtJMT1和Glyma11G007600. 1 (Bashir et al., 2019; Song et al., 2019)
GSH (Noriega et al., 2012; Zhao et al., 2016)
CAT、SOD和APX (Noriega et al., 2012; Zhao et al., 2016)
... Cd处理的植物初级根尖和子叶中IAA浓度及分布有较大差异.Cd处理后, IAA含量显著降低(Guo et al., 2019), IAA合成基因表达下调(Bashir et al., 2019), IAA氧化酶活性增强, IAA生物合成和分解代谢基因表达改变(Hu et al., 2013).此外, Cd胁迫下, 外源IAA能显著提高玉米幼苗根和地上部生物量, 降低地上部Cd含量, 但对根Cd含量的影响不显著, 显著降低叶片的MDA和GSH含量, 使玉米幼苗叶片SOD、POD和CAT活性显著增强(陈晶等, 2016).当IAA合成受到抑制时, 如施加生长素信号抑制剂氯苯氧基异丁酸, 可有效抑制10 μmol·L-1 Cd诱导的根系生长和H2O2产生, 但不抑制60 μmol·L-1 Cd诱导的根系生长和H2O2产生(Tamás et al., 2012).Cd胁迫下施加IAA, 可激活谷胱甘肽-S-转移酶(glutathione- S-transferase, GST) (在植物解毒过程中起关键作用)活性, 使植物更好地发生应激反应(Bo?ová et al., 2013).Cd胁迫下, 施加外源IAA还可显著提高SODs基因的表达, 降低H2O2含量, 从而减少Cd对植物的毒害(Pan et al., 2020).重金属污染条件下, 外源IAA可促进天然抗性相关巨噬细胞蛋白基因NRAMP (NATURAL RESISTANCE-ASSOCIATED MACROPHAGE PROTEIN)、IRT、重金属ATP酶家族基因HMA (heavy metal ATPase)和锌铁转运蛋白基因ZIP (zinc iron transporter protein)的表达, 提高植物的抗Cd能力(Korshunova et al., 1999; 罗莎, 2017). ...
A heavy metal P-type ATPase OsHMA4 prevents copper accumulation in rice grain 2 2016
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
... 重金属ATP酶基因家族(HMA)已被证明在不同的植物种类中行使特定的功能, 不同的HMA在不同组织中表达, 最终导致功能的多样性.HvHMA1参与Zn以及Cd向大麦(Hordeum vulgare)籽粒的转运(Mikkelsen et al., 2012).HMA2和HMA4可将Cd装载到木质部, 以便从根运输至茎和叶, OsHMA2与水稻维管组织中的锌装载及液泡膜定位有关(Yamaji et al., 2013).液泡膜上的OsHMA3将Cd转移至根系液泡, OsHMA4则将Cu转运至根系液泡, 限制Cd从根向茎以及叶的转移, 从而减少Cd在种子中的积累(Miyadate et al., 2011; Huang et al., 2016).TcHMA3在叶片的液泡中表达, 将Cd隔离在根和叶的液泡中, 最终导致Cd的超积累, 增强对Cd的耐受性(Ueno et al., 2011).AtHMA3在拟南芥中过表达导致Cd的积累量比野生型高2-3倍(Morel et al., 2009).这些转运蛋白为植物的基因工程提供了有用的工具, 增强了植物对金属的耐受性和积累能力, 继续推进它们的协同作用研究具有重要意义. ...
Cadmium in plants: uptake, toxicity, and its interactions with selenium fertilizers 1 2019
... Ca2+作为细胞功能的第二信使, 参与调节植物各种逆境下的重要生理生化过程.钙信使系统包括Ca2+、钙调蛋白(CaM)和钙泵(Ca2+-ATPase).CaM是Ca2+的感受器, Ca2+-ATPase决定着Ca2+的跨膜运输(Snedden and Fromm, 2001).正常情况下三者的含量保持稳定, 可维持细胞正常的生理功能.在外界生物和非生物胁迫下, 细胞质中的Ca2+浓度会瞬间升高(Ca2+浓度的波动被认为是植物对外界刺激的反应信号) (Nürnberger and Scheel, 2001; Liu et al., 2017).Cd胁迫扰动植物细胞中的Ca2+水平, 从而扰乱植物叶片的水分状况(Perfus-Barbeoch et al., 2002).胞质Ca2+浓度增加对H2O2的生成有动态调控作用, Ca2+可调控NADH氧化酶活性, 上调植物中H2O2的含量; 也可通过刺激H2O2酶活性下调植物中的H2O2水平(Yang and Poovaiah, 2002).当ROS含量超过Ca2+调控的浓度范围时, 会对植物造成伤害, 细胞膜和细胞器膜发生膜质过氧化, 膜的通透性被破坏, 致使细胞死亡(赵士诚等, 2008).Cd与Ca2+的化学性质相似, Cd被植物吸收后通过金属转运体或钙通道运输至根部, 也可通过电压依赖的钙通道进入细胞, 一旦进入胞质, 它就会模拟钙离子的活性, 扰乱钙信号调控; 导致根系吸收大量的Cd, 并显著减少根长、根表面积、根体积和根系生物量, 使叶片黄化并干燥坏死(Yang and Poovaiah, 2002; Wang et al., 2016; Ismael et al., 2019).Cd进入植物根细胞也诱导一些对重金属胁迫有反应的特定信号分子, 如CaMs、钙依赖蛋白激酶(calcium-dependent protein kinases, CDPKs)和丝裂原活化蛋白激酶(mitogen-activated protein kinases, MAPKs), 调控植物对逆境的抗性(Danquah et al., 2014; Xu et al., 2017). ...
Accumulation and fixation of Cd by tomato cell wall pectin under Cd stress 2 2019
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
... Cd胁迫通常促进JA合成基因(Bashir et al., 2019)和抗氧化酶合成基因的转录(Keunen et al., 2013).低浓度JA可缓解Cd胁迫.例如, 用20 μmol·L-1 JA处理可有效改善Cd诱导的氧化应激, 使硫代巴比妥酸反应物质、H2O2和O2-.的积累减少, 使GSH、CAT、SOD及血红素氧合酶(HO)活性增强(Noriega et al., 2012; Adhikari et al., 2018).研究发现, 番茄(Solanum lycopersicum) JA缺失突变体对Cd的敏感性增强, 与光合作用相关参数(包括总叶绿素、光系统II实际效率、可变叶绿素荧光与最大荧光比及净光合速率)值降低; 相对水含量、可溶性糖和脯氨酸含量及淀粉的积累减少; 氧化应激和抗氧化防御作用(包括H2O2与MDA生成、电解质渗漏、SOD、APX和CAT活性)减弱(Zhao et al., 2016).外源茉莉酸甲酯(MeJA)通过降低拟南芥根系和茎叶中的Cd浓度, 下调参与Cd吸收和运输的AtIRT1、AtHMA2和AtHMA4基因表达, 缓解Cd引起的新叶毒害.JA合成关键基因AtAOS突变上调AtIRT1、AtHMA2和AtHMA4等转运蛋白基因表达, 增加根和芽中的Cd浓度, 提高拟南芥对Cd的敏感性.此外, MeJA降低Cd胁迫的拟南芥根尖中的NO水平(Lei et al., 2020).镉胁迫下植物激素ABA、IAA、SA和JA之间的相互作用值得深入探索. ...
Genome-wide identification of ATP binding cassette (ABC) transporter and heavy metal associated (HMA) gene families in flax ( Linum usitatissimum L.) 2 2020
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
... Cd胁迫通常促进JA合成基因(Bashir et al., 2019)和抗氧化酶合成基因的转录(Keunen et al., 2013).低浓度JA可缓解Cd胁迫.例如, 用20 μmol·L-1 JA处理可有效改善Cd诱导的氧化应激, 使硫代巴比妥酸反应物质、H2O2和O2-.的积累减少, 使GSH、CAT、SOD及血红素氧合酶(HO)活性增强(Noriega et al., 2012; Adhikari et al., 2018).研究发现, 番茄(Solanum lycopersicum) JA缺失突变体对Cd的敏感性增强, 与光合作用相关参数(包括总叶绿素、光系统II实际效率、可变叶绿素荧光与最大荧光比及净光合速率)值降低; 相对水含量、可溶性糖和脯氨酸含量及淀粉的积累减少; 氧化应激和抗氧化防御作用(包括H2O2与MDA生成、电解质渗漏、SOD、APX和CAT活性)减弱(Zhao et al., 2016).外源茉莉酸甲酯(MeJA)通过降低拟南芥根系和茎叶中的Cd浓度, 下调参与Cd吸收和运输的AtIRT1、AtHMA2和AtHMA4基因表达, 缓解Cd引起的新叶毒害.JA合成关键基因AtAOS突变上调AtIRT1、AtHMA2和AtHMA4等转运蛋白基因表达, 增加根和芽中的Cd浓度, 提高拟南芥对Cd的敏感性.此外, MeJA降低Cd胁迫的拟南芥根尖中的NO水平(Lei et al., 2020).镉胁迫下植物激素ABA、IAA、SA和JA之间的相互作用值得深入探索. ...
Involvement of abscisic acid in regulating antioxidative defense systems and IAA-oxidase activity and improving adventitious rooting in mung bean (Vigna radiata(L.) Wilczek) seedlings under cadmium stress 1 2014
... Responses of plant hormones to Cd stress Table 1
激素
激素合成基因
非酶物质
抗氧化酶
抗氧化物质基因
吸收和运输蛋白基因
ABA
NCED3和Glyma17G242200 (Bashir et al., 2019; Song et al., 2019)
AsA和GSH (Hu et al., 2005; Stroiński et al., 2010; Li et al., 2014)
APX、POD、SOD和CAT (Hu et al., 2005)
StPCS 1 (Stroiń- ski et al., 2010)
IRT1 (Fan et al., 2014)
IAA
Glyma02G037600.1 (Bashir et al., 2019)
GSH (Hu et al., 2013)
SOD、POD、CAT和GST (Bo?ová et al., 2013; 陈晶等, 2016)
SODs (Pan et al., 2020)
Nramp、IRT、HMA和ZIP (Korshunova et al., 1999; 罗莎, 2017)
SA
Glyma02G063400 (Bashir et al., 2019)
脯氨酸(Krantev et al., 2008; 胡春霞和王秀芹, 2010)
NR、GS和GOGAT (Zawoznik et al., 2007; Chao et al., 2010; 赵新月等, 2013)
SODS、CATS和APXS (Gill, 2015)
OsLCT1、OsLCD和ZIP (Liu et al., 2016)
JA
PtJMT1和Glyma11G007600. 1 (Bashir et al., 2019; Song et al., 2019)
GSH (Noriega et al., 2012; Zhao et al., 2016)
CAT、SOD和APX (Noriega et al., 2012; Zhao et al., 2016)
Effects of calcium application on activities of membrane transporters in Panax notoginseng under cadmium stress 1 2021
... 活性氮(RNS)主要包括一氧化氮(NO)、S-亚硝基硫醇(SNOs)、高氮氧化物(NOx)、二硝基铁配合物和过氧亚硝酸盐(ONOO) 5种.过量的RNS会与脂类、蛋白质和核酸发生反应, 导致细胞损害(Neill et al., 2008a; Romero-Puertas and Sandalio, 2016).然而, 细胞内NO一旦产生, 就会与ROS迅速作用, 产生大量的RNS (如NO2), 然后降解为亚硝酸盐和硝酸钠(Neill et al., 2008b).低水平的NO可调节植物的发育, 而在胁迫条件下植物体内会产生大量的NO (Romero-Puertas and Sandalio, 2016; Liu et al., 2018); NO浓度过高则抑制植物的生长, 甚至引起细胞程序性死亡(Arasimowicz-Jelonek et al., 2012; Sami et al., 2018).低浓度NO促进植物生长, 高浓度则抑制植物生长, 其与生长素(indole-3-acetic acid, IAA)、细胞分裂素、赤霉素、脱落酸(abscisic acid, ABA)、乙烯、茉莉酸(jasmonic acid, JA)和水杨酸(salicylic acid, SA)也有一定的交互作用(Sami et al., 2018).Cd能通过破坏呼吸和光合作用的电子传递链使电子被传递到非靶向分子, 从而导致ROS和RNS的产生, 且Cd能通过抑制自由基降低相关酶活性(Ekmek?i et al., 2008; Romero-Puertas and Sandalio, 2016), 导致ROS和RNS不能够被及时清除而过量累积, 最终引起植物营养应激反应(Nieves-Cordones et al., 2019).外施氮(Zhang et al., 2019b)、磷(Wang et al., 2020)、钾(Wu et al., 2020)、钙(Li et al., 2021)和硅(Vaculík et al., 2015)等植物营养元素, 可起到调控植物激素含量、非酶物质和抗氧化酶活性及转运基因表达等作用, 减轻Cd对植物的毒害, 但外施营养物质对ROS和RNS有怎样的影响有待后续探索. ...
RETRACTED: the interplay between reactive oxygen and nitrogen species contributes in the regulatory mechanism of the nitro-oxidative stress induced by cadmium in Arabidopsis 1 2018
... 活性氮(RNS)主要包括一氧化氮(NO)、S-亚硝基硫醇(SNOs)、高氮氧化物(NOx)、二硝基铁配合物和过氧亚硝酸盐(ONOO) 5种.过量的RNS会与脂类、蛋白质和核酸发生反应, 导致细胞损害(Neill et al., 2008a; Romero-Puertas and Sandalio, 2016).然而, 细胞内NO一旦产生, 就会与ROS迅速作用, 产生大量的RNS (如NO2), 然后降解为亚硝酸盐和硝酸钠(Neill et al., 2008b).低水平的NO可调节植物的发育, 而在胁迫条件下植物体内会产生大量的NO (Romero-Puertas and Sandalio, 2016; Liu et al., 2018); NO浓度过高则抑制植物的生长, 甚至引起细胞程序性死亡(Arasimowicz-Jelonek et al., 2012; Sami et al., 2018).低浓度NO促进植物生长, 高浓度则抑制植物生长, 其与生长素(indole-3-acetic acid, IAA)、细胞分裂素、赤霉素、脱落酸(abscisic acid, ABA)、乙烯、茉莉酸(jasmonic acid, JA)和水杨酸(salicylic acid, SA)也有一定的交互作用(Sami et al., 2018).Cd能通过破坏呼吸和光合作用的电子传递链使电子被传递到非靶向分子, 从而导致ROS和RNS的产生, 且Cd能通过抑制自由基降低相关酶活性(Ekmek?i et al., 2008; Romero-Puertas and Sandalio, 2016), 导致ROS和RNS不能够被及时清除而过量累积, 最终引起植物营养应激反应(Nieves-Cordones et al., 2019).外施氮(Zhang et al., 2019b)、磷(Wang et al., 2020)、钾(Wu et al., 2020)、钙(Li et al., 2021)和硅(Vaculík et al., 2015)等植物营养元素, 可起到调控植物激素含量、非酶物质和抗氧化酶活性及转运基因表达等作用, 减轻Cd对植物的毒害, 但外施营养物质对ROS和RNS有怎样的影响有待后续探索. ...
Improved short-term drought response of transgenic rice over-expressing maize C4 phosphoenolpyruvate carboxylase via calcium signal cascade 1 2017
... Ca2+作为细胞功能的第二信使, 参与调节植物各种逆境下的重要生理生化过程.钙信使系统包括Ca2+、钙调蛋白(CaM)和钙泵(Ca2+-ATPase).CaM是Ca2+的感受器, Ca2+-ATPase决定着Ca2+的跨膜运输(Snedden and Fromm, 2001).正常情况下三者的含量保持稳定, 可维持细胞正常的生理功能.在外界生物和非生物胁迫下, 细胞质中的Ca2+浓度会瞬间升高(Ca2+浓度的波动被认为是植物对外界刺激的反应信号) (Nürnberger and Scheel, 2001; Liu et al., 2017).Cd胁迫扰动植物细胞中的Ca2+水平, 从而扰乱植物叶片的水分状况(Perfus-Barbeoch et al., 2002).胞质Ca2+浓度增加对H2O2的生成有动态调控作用, Ca2+可调控NADH氧化酶活性, 上调植物中H2O2的含量; 也可通过刺激H2O2酶活性下调植物中的H2O2水平(Yang and Poovaiah, 2002).当ROS含量超过Ca2+调控的浓度范围时, 会对植物造成伤害, 细胞膜和细胞器膜发生膜质过氧化, 膜的通透性被破坏, 致使细胞死亡(赵士诚等, 2008).Cd与Ca2+的化学性质相似, Cd被植物吸收后通过金属转运体或钙通道运输至根部, 也可通过电压依赖的钙通道进入细胞, 一旦进入胞质, 它就会模拟钙离子的活性, 扰乱钙信号调控; 导致根系吸收大量的Cd, 并显著减少根长、根表面积、根体积和根系生物量, 使叶片黄化并干燥坏死(Yang and Poovaiah, 2002; Wang et al., 2016; Ismael et al., 2019).Cd进入植物根细胞也诱导一些对重金属胁迫有反应的特定信号分子, 如CaMs、钙依赖蛋白激酶(calcium-dependent protein kinases, CDPKs)和丝裂原活化蛋白激酶(mitogen-activated protein kinases, MAPKs), 调控植物对逆境的抗性(Danquah et al., 2014; Xu et al., 2017). ...
Role of salicylic acid in resistance to cadmium stress in plants 4 2016
... Cd胁迫下, 抗逆相关的植物激素(如ABA、IAA、SA和JA)均会发生显著变化, 且影响抗氧化物质合成相关基因和运输蛋白基因的表达(表1).多数情况下Cd胁迫可引起植物体内IAA、SA、ABA和JA含量变化(Liu et al., 2016; Bashir et al., 2019; Song et al., 2019).添加适量外源ABA、IAA、SA和JA等可缓解Cd胁迫对植物的危害. ...
... Responses of plant hormones to Cd stress Table 1
激素
激素合成基因
非酶物质
抗氧化酶
抗氧化物质基因
吸收和运输蛋白基因
ABA
NCED3和Glyma17G242200 (Bashir et al., 2019; Song et al., 2019)
AsA和GSH (Hu et al., 2005; Stroiński et al., 2010; Li et al., 2014)
APX、POD、SOD和CAT (Hu et al., 2005)
StPCS 1 (Stroiń- ski et al., 2010)
IRT1 (Fan et al., 2014)
IAA
Glyma02G037600.1 (Bashir et al., 2019)
GSH (Hu et al., 2013)
SOD、POD、CAT和GST (Bo?ová et al., 2013; 陈晶等, 2016)
SODs (Pan et al., 2020)
Nramp、IRT、HMA和ZIP (Korshunova et al., 1999; 罗莎, 2017)
SA
Glyma02G063400 (Bashir et al., 2019)
脯氨酸(Krantev et al., 2008; 胡春霞和王秀芹, 2010)
NR、GS和GOGAT (Zawoznik et al., 2007; Chao et al., 2010; 赵新月等, 2013)
SODS、CATS和APXS (Gill, 2015)
OsLCT1、OsLCD和ZIP (Liu et al., 2016)
JA
PtJMT1和Glyma11G007600. 1 (Bashir et al., 2019; Song et al., 2019)
GSH (Noriega et al., 2012; Zhao et al., 2016)
CAT、SOD和APX (Noriega et al., 2012; Zhao et al., 2016)
... 外源SA可改变ROS的积累和抗氧化酶系统活性, 缓解Cd胁迫带来的不利影响(Zawoznik et al., 2007).施加外源SA可使油菜(Brassica campestris)叶片的可溶性糖和游离脯氨酸含量升高、MDA积累减少, 质膜透性降低, CAT、POD和SOD活性升高, 明显缓解Cd毒害(胡春霞和王秀芹, 2010).SA处理显著增强玉米叶片中SOD活性, 降低CAT和POD活性, 并减轻Cd对APX活性的抑制作用(Krantev et al., 2008).此外, SA通过抑制ROS的过量产生缓解Cd引发的氧化损伤(Zawoznik et al., 2007); 施用外源SA可减少根系中ROS和MDA的积累, 进而减轻Cd毒性(Liu et al., 2016); 外源SA也可介导NADPH氧化酶产生H2O2, 以应对Cd胁迫引起的氧化应激(Chao et al., 2010).赵新月等(2013)研究表明, 与仅用Cd处理的玉米相比, 用50-250 μmol·L-1 SA处理玉米其叶绿素含量、硝酸还原酶(nitrite reductase, NR)、谷氨酰胺合成酶(glutamine synthetase, GS)和谷氨酸合成酶(glutamate synthase, GOGAT)活性显著升高, 氮素代谢过程良好, 玉米根系的POD活性降低(但POD和SOD活性与无Cd处理相比无显著差异), 且降低了Cd胁迫对质膜氧化系统的伤害.但外源SA对Cd胁迫修复的缓解作用受浓度影响较大, 低浓度的SA具有良好的修复效果, 高浓度的SA反而影响了光合色素的合成及酶活性, 抑制玉米的生长(赵新月等, 2013).重金属胁迫下, 外源SA提高了植物SOD、CAT和APX合成基因的表达(Gill, 2015; Song et al., 2019); 及水稻叶片和稻穗中低亲和离子转运子基因(OsLCT1)、水稻低Cd基因(OsLCD)和ZIP的时空表达(Liu et al., 2016). ...
... 的时空表达(Liu et al., 2016). ...
Barley HvHMA1 is a heavy metal pump involved in mobilizing organellar Zn and Cu and plays a role in metal loading into grains 2 2012
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
Nitric oxide, stomatal closure, and abiotic stress 1 2008
... 活性氮(RNS)主要包括一氧化氮(NO)、S-亚硝基硫醇(SNOs)、高氮氧化物(NOx)、二硝基铁配合物和过氧亚硝酸盐(ONOO) 5种.过量的RNS会与脂类、蛋白质和核酸发生反应, 导致细胞损害(Neill et al., 2008a; Romero-Puertas and Sandalio, 2016).然而, 细胞内NO一旦产生, 就会与ROS迅速作用, 产生大量的RNS (如NO2), 然后降解为亚硝酸盐和硝酸钠(Neill et al., 2008b).低水平的NO可调节植物的发育, 而在胁迫条件下植物体内会产生大量的NO (Romero-Puertas and Sandalio, 2016; Liu et al., 2018); NO浓度过高则抑制植物的生长, 甚至引起细胞程序性死亡(Arasimowicz-Jelonek et al., 2012; Sami et al., 2018).低浓度NO促进植物生长, 高浓度则抑制植物生长, 其与生长素(indole-3-acetic acid, IAA)、细胞分裂素、赤霉素、脱落酸(abscisic acid, ABA)、乙烯、茉莉酸(jasmonic acid, JA)和水杨酸(salicylic acid, SA)也有一定的交互作用(Sami et al., 2018).Cd能通过破坏呼吸和光合作用的电子传递链使电子被传递到非靶向分子, 从而导致ROS和RNS的产生, 且Cd能通过抑制自由基降低相关酶活性(Ekmek?i et al., 2008; Romero-Puertas and Sandalio, 2016), 导致ROS和RNS不能够被及时清除而过量累积, 最终引起植物营养应激反应(Nieves-Cordones et al., 2019).外施氮(Zhang et al., 2019b)、磷(Wang et al., 2020)、钾(Wu et al., 2020)、钙(Li et al., 2021)和硅(Vaculík et al., 2015)等植物营养元素, 可起到调控植物激素含量、非酶物质和抗氧化酶活性及转运基因表达等作用, 减轻Cd对植物的毒害, 但外施营养物质对ROS和RNS有怎样的影响有待后续探索. ...
Nitric oxide evolution and perception 1 2008
... 活性氮(RNS)主要包括一氧化氮(NO)、S-亚硝基硫醇(SNOs)、高氮氧化物(NOx)、二硝基铁配合物和过氧亚硝酸盐(ONOO) 5种.过量的RNS会与脂类、蛋白质和核酸发生反应, 导致细胞损害(Neill et al., 2008a; Romero-Puertas and Sandalio, 2016).然而, 细胞内NO一旦产生, 就会与ROS迅速作用, 产生大量的RNS (如NO2), 然后降解为亚硝酸盐和硝酸钠(Neill et al., 2008b).低水平的NO可调节植物的发育, 而在胁迫条件下植物体内会产生大量的NO (Romero-Puertas and Sandalio, 2016; Liu et al., 2018); NO浓度过高则抑制植物的生长, 甚至引起细胞程序性死亡(Arasimowicz-Jelonek et al., 2012; Sami et al., 2018).低浓度NO促进植物生长, 高浓度则抑制植物生长, 其与生长素(indole-3-acetic acid, IAA)、细胞分裂素、赤霉素、脱落酸(abscisic acid, ABA)、乙烯、茉莉酸(jasmonic acid, JA)和水杨酸(salicylic acid, SA)也有一定的交互作用(Sami et al., 2018).Cd能通过破坏呼吸和光合作用的电子传递链使电子被传递到非靶向分子, 从而导致ROS和RNS的产生, 且Cd能通过抑制自由基降低相关酶活性(Ekmek?i et al., 2008; Romero-Puertas and Sandalio, 2016), 导致ROS和RNS不能够被及时清除而过量累积, 最终引起植物营养应激反应(Nieves-Cordones et al., 2019).外施氮(Zhang et al., 2019b)、磷(Wang et al., 2020)、钾(Wu et al., 2020)、钙(Li et al., 2021)和硅(Vaculík et al., 2015)等植物营养元素, 可起到调控植物激素含量、非酶物质和抗氧化酶活性及转运基因表达等作用, 减轻Cd对植物的毒害, 但外施营养物质对ROS和RNS有怎样的影响有待后续探索. ...
Critical responses to nutrient deprivation: a comprehensive review on the role of ROS and RNS 3 2019
... 活性氧(reactive oxygen species, ROS)和活性氮(reactive nitrogen species, RNS)是植物产生的具有一定功能的活性分子化合物, 是植物适应环境变化的主要调节因子.低含量水平下, ROS和RNS是信号分子(Romero-Puertas and Sandalio, 2016); 过多积累后, 会诱导植物氧化损伤及细胞程序性死亡, 是植物响应胁迫的关键指标(Das and Roychoudhury, 2014; Nieves-Cordones et al., 2019).Cd毒害最常见的表现是诱导植物出现氧化应激反应, 如产生ROS和RNS (Romero-Puertas et al., 2019).ROS和RNS之间存在动态平衡(Delledonne et al., 2001).ROS通过多种反应产生, 包括呼吸和光合电子传递链及酶(如过氧化物酶)的副反应等; Cd胁迫下, 植物细胞中产生ROS的部位有线粒体、叶绿体、过氧化物酶体、细胞质以及外质体等(Romero-Puertas et al., 2019).ROS包括单线态氧(single oxygen, 1O2)、超氧自由基阴离子(superoxide radical, O2-.)、过氧化氢(hydrogen peroxide, H2O2)和羟基自由基(hydroxyl radical, ·OH) (Nieves-Cordones et al., 2019).清除ROS的机制涉及酶促抗氧化剂和非酶类抗氧化剂(1.1.3节).ROS会对脂质、蛋白质和DNA等生物分子造成损害, 破坏细胞的完整性, 最终导致细胞死亡(Das and Roychoudhury, 2014).ROS作为信号分子在多种生命过程中扮演重要角色, 包括趋向性、细胞死亡和细胞对生物和非生物胁迫的反应(Mittler et al., 2011). ...
... )和羟基自由基(hydroxyl radical, ·OH) (Nieves-Cordones et al., 2019).清除ROS的机制涉及酶促抗氧化剂和非酶类抗氧化剂(1.1.3节).ROS会对脂质、蛋白质和DNA等生物分子造成损害, 破坏细胞的完整性, 最终导致细胞死亡(Das and Roychoudhury, 2014).ROS作为信号分子在多种生命过程中扮演重要角色, 包括趋向性、细胞死亡和细胞对生物和非生物胁迫的反应(Mittler et al., 2011). ...
... 活性氮(RNS)主要包括一氧化氮(NO)、S-亚硝基硫醇(SNOs)、高氮氧化物(NOx)、二硝基铁配合物和过氧亚硝酸盐(ONOO) 5种.过量的RNS会与脂类、蛋白质和核酸发生反应, 导致细胞损害(Neill et al., 2008a; Romero-Puertas and Sandalio, 2016).然而, 细胞内NO一旦产生, 就会与ROS迅速作用, 产生大量的RNS (如NO2), 然后降解为亚硝酸盐和硝酸钠(Neill et al., 2008b).低水平的NO可调节植物的发育, 而在胁迫条件下植物体内会产生大量的NO (Romero-Puertas and Sandalio, 2016; Liu et al., 2018); NO浓度过高则抑制植物的生长, 甚至引起细胞程序性死亡(Arasimowicz-Jelonek et al., 2012; Sami et al., 2018).低浓度NO促进植物生长, 高浓度则抑制植物生长, 其与生长素(indole-3-acetic acid, IAA)、细胞分裂素、赤霉素、脱落酸(abscisic acid, ABA)、乙烯、茉莉酸(jasmonic acid, JA)和水杨酸(salicylic acid, SA)也有一定的交互作用(Sami et al., 2018).Cd能通过破坏呼吸和光合作用的电子传递链使电子被传递到非靶向分子, 从而导致ROS和RNS的产生, 且Cd能通过抑制自由基降低相关酶活性(Ekmek?i et al., 2008; Romero-Puertas and Sandalio, 2016), 导致ROS和RNS不能够被及时清除而过量累积, 最终引起植物营养应激反应(Nieves-Cordones et al., 2019).外施氮(Zhang et al., 2019b)、磷(Wang et al., 2020)、钾(Wu et al., 2020)、钙(Li et al., 2021)和硅(Vaculík et al., 2015)等植物营养元素, 可起到调控植物激素含量、非酶物质和抗氧化酶活性及转运基因表达等作用, 减轻Cd对植物的毒害, 但外施营养物质对ROS和RNS有怎样的影响有待后续探索. ...
Heme oxygenase is involved in the protection exerted by jasmonic acid against cadmium stress in soybean roots 3 2012
... Responses of plant hormones to Cd stress Table 1
激素
激素合成基因
非酶物质
抗氧化酶
抗氧化物质基因
吸收和运输蛋白基因
ABA
NCED3和Glyma17G242200 (Bashir et al., 2019; Song et al., 2019)
AsA和GSH (Hu et al., 2005; Stroiński et al., 2010; Li et al., 2014)
APX、POD、SOD和CAT (Hu et al., 2005)
StPCS 1 (Stroiń- ski et al., 2010)
IRT1 (Fan et al., 2014)
IAA
Glyma02G037600.1 (Bashir et al., 2019)
GSH (Hu et al., 2013)
SOD、POD、CAT和GST (Bo?ová et al., 2013; 陈晶等, 2016)
SODs (Pan et al., 2020)
Nramp、IRT、HMA和ZIP (Korshunova et al., 1999; 罗莎, 2017)
SA
Glyma02G063400 (Bashir et al., 2019)
脯氨酸(Krantev et al., 2008; 胡春霞和王秀芹, 2010)
NR、GS和GOGAT (Zawoznik et al., 2007; Chao et al., 2010; 赵新月等, 2013)
SODS、CATS和APXS (Gill, 2015)
OsLCT1、OsLCD和ZIP (Liu et al., 2016)
JA
PtJMT1和Glyma11G007600. 1 (Bashir et al., 2019; Song et al., 2019)
GSH (Noriega et al., 2012; Zhao et al., 2016)
CAT、SOD和APX (Noriega et al., 2012; Zhao et al., 2016)
... Cd胁迫通常促进JA合成基因(Bashir et al., 2019)和抗氧化酶合成基因的转录(Keunen et al., 2013).低浓度JA可缓解Cd胁迫.例如, 用20 μmol·L-1 JA处理可有效改善Cd诱导的氧化应激, 使硫代巴比妥酸反应物质、H2O2和O2-.的积累减少, 使GSH、CAT、SOD及血红素氧合酶(HO)活性增强(Noriega et al., 2012; Adhikari et al., 2018).研究发现, 番茄(Solanum lycopersicum) JA缺失突变体对Cd的敏感性增强, 与光合作用相关参数(包括总叶绿素、光系统II实际效率、可变叶绿素荧光与最大荧光比及净光合速率)值降低; 相对水含量、可溶性糖和脯氨酸含量及淀粉的积累减少; 氧化应激和抗氧化防御作用(包括H2O2与MDA生成、电解质渗漏、SOD、APX和CAT活性)减弱(Zhao et al., 2016).外源茉莉酸甲酯(MeJA)通过降低拟南芥根系和茎叶中的Cd浓度, 下调参与Cd吸收和运输的AtIRT1、AtHMA2和AtHMA4基因表达, 缓解Cd引起的新叶毒害.JA合成关键基因AtAOS突变上调AtIRT1、AtHMA2和AtHMA4等转运蛋白基因表达, 增加根和芽中的Cd浓度, 提高拟南芥对Cd的敏感性.此外, MeJA降低Cd胁迫的拟南芥根尖中的NO水平(Lei et al., 2020).镉胁迫下植物激素ABA、IAA、SA和JA之间的相互作用值得深入探索. ...
Signal transmission in the plant immune response 1 2001
... Ca2+作为细胞功能的第二信使, 参与调节植物各种逆境下的重要生理生化过程.钙信使系统包括Ca2+、钙调蛋白(CaM)和钙泵(Ca2+-ATPase).CaM是Ca2+的感受器, Ca2+-ATPase决定着Ca2+的跨膜运输(Snedden and Fromm, 2001).正常情况下三者的含量保持稳定, 可维持细胞正常的生理功能.在外界生物和非生物胁迫下, 细胞质中的Ca2+浓度会瞬间升高(Ca2+浓度的波动被认为是植物对外界刺激的反应信号) (Nürnberger and Scheel, 2001; Liu et al., 2017).Cd胁迫扰动植物细胞中的Ca2+水平, 从而扰乱植物叶片的水分状况(Perfus-Barbeoch et al., 2002).胞质Ca2+浓度增加对H2O2的生成有动态调控作用, Ca2+可调控NADH氧化酶活性, 上调植物中H2O2的含量; 也可通过刺激H2O2酶活性下调植物中的H2O2水平(Yang and Poovaiah, 2002).当ROS含量超过Ca2+调控的浓度范围时, 会对植物造成伤害, 细胞膜和细胞器膜发生膜质过氧化, 膜的通透性被破坏, 致使细胞死亡(赵士诚等, 2008).Cd与Ca2+的化学性质相似, Cd被植物吸收后通过金属转运体或钙通道运输至根部, 也可通过电压依赖的钙通道进入细胞, 一旦进入胞质, 它就会模拟钙离子的活性, 扰乱钙信号调控; 导致根系吸收大量的Cd, 并显著减少根长、根表面积、根体积和根系生物量, 使叶片黄化并干燥坏死(Yang and Poovaiah, 2002; Wang et al., 2016; Ismael et al., 2019).Cd进入植物根细胞也诱导一些对重金属胁迫有反应的特定信号分子, 如CaMs、钙依赖蛋白激酶(calcium-dependent protein kinases, CDPKs)和丝裂原活化蛋白激酶(mitogen-activated protein kinases, MAPKs), 调控植物对逆境的抗性(Danquah et al., 2014; Xu et al., 2017). ...
SODs involved in the hormone mediated regulation of H2O2 content in Kandelia obovata root tissues under cadmium stress 2 2020
... Responses of plant hormones to Cd stress Table 1
激素
激素合成基因
非酶物质
抗氧化酶
抗氧化物质基因
吸收和运输蛋白基因
ABA
NCED3和Glyma17G242200 (Bashir et al., 2019; Song et al., 2019)
AsA和GSH (Hu et al., 2005; Stroiński et al., 2010; Li et al., 2014)
APX、POD、SOD和CAT (Hu et al., 2005)
StPCS 1 (Stroiń- ski et al., 2010)
IRT1 (Fan et al., 2014)
IAA
Glyma02G037600.1 (Bashir et al., 2019)
GSH (Hu et al., 2013)
SOD、POD、CAT和GST (Bo?ová et al., 2013; 陈晶等, 2016)
SODs (Pan et al., 2020)
Nramp、IRT、HMA和ZIP (Korshunova et al., 1999; 罗莎, 2017)
SA
Glyma02G063400 (Bashir et al., 2019)
脯氨酸(Krantev et al., 2008; 胡春霞和王秀芹, 2010)
NR、GS和GOGAT (Zawoznik et al., 2007; Chao et al., 2010; 赵新月等, 2013)
SODS、CATS和APXS (Gill, 2015)
OsLCT1、OsLCD和ZIP (Liu et al., 2016)
JA
PtJMT1和Glyma11G007600. 1 (Bashir et al., 2019; Song et al., 2019)
GSH (Noriega et al., 2012; Zhao et al., 2016)
CAT、SOD和APX (Noriega et al., 2012; Zhao et al., 2016)
... Cd处理的植物初级根尖和子叶中IAA浓度及分布有较大差异.Cd处理后, IAA含量显著降低(Guo et al., 2019), IAA合成基因表达下调(Bashir et al., 2019), IAA氧化酶活性增强, IAA生物合成和分解代谢基因表达改变(Hu et al., 2013).此外, Cd胁迫下, 外源IAA能显著提高玉米幼苗根和地上部生物量, 降低地上部Cd含量, 但对根Cd含量的影响不显著, 显著降低叶片的MDA和GSH含量, 使玉米幼苗叶片SOD、POD和CAT活性显著增强(陈晶等, 2016).当IAA合成受到抑制时, 如施加生长素信号抑制剂氯苯氧基异丁酸, 可有效抑制10 μmol·L-1 Cd诱导的根系生长和H2O2产生, 但不抑制60 μmol·L-1 Cd诱导的根系生长和H2O2产生(Tamás et al., 2012).Cd胁迫下施加IAA, 可激活谷胱甘肽-S-转移酶(glutathione- S-transferase, GST) (在植物解毒过程中起关键作用)活性, 使植物更好地发生应激反应(Bo?ová et al., 2013).Cd胁迫下, 施加外源IAA还可显著提高SODs基因的表达, 降低H2O2含量, 从而减少Cd对植物的毒害(Pan et al., 2020).重金属污染条件下, 外源IAA可促进天然抗性相关巨噬细胞蛋白基因NRAMP (NATURAL RESISTANCE-ASSOCIATED MACROPHAGE PROTEIN)、IRT、重金属ATP酶家族基因HMA (heavy metal ATPase)和锌铁转运蛋白基因ZIP (zinc iron transporter protein)的表达, 提高植物的抗Cd能力(Korshunova et al., 1999; 罗莎, 2017). ...
The phytochelatin transporters AtABCC1 and AtABCC2 mediate tolerance to cadmium and mercury 2 2012
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
... ABC (ATP-binding cassette)转运体家族广泛参与细胞中离子、碳水化合物、脂类、药物和重金属的运输(Khan et al., 2020). 其中, 两种ABCC转运蛋白(AtABCC1和AtABCC2)是重要的液泡转运体, 通过液泡隔离介导对Cd的耐受性(Park et al., 2012).ZIP包括锌调控转运体(zinc-regulated transporter, ZRT)和铁调控转运体(iron-regulated transporter, IRT)家族.拟南芥中, 定位于根系细胞质膜的AtIRT1是主要的根部铁摄取载体, 其也可运输大量的Cd (Wu et al., 2019). ...
Heavy metal toxicity: cadmium permeates through calcium channels and disturbs the plant water status 1 2002
... Ca2+作为细胞功能的第二信使, 参与调节植物各种逆境下的重要生理生化过程.钙信使系统包括Ca2+、钙调蛋白(CaM)和钙泵(Ca2+-ATPase).CaM是Ca2+的感受器, Ca2+-ATPase决定着Ca2+的跨膜运输(Snedden and Fromm, 2001).正常情况下三者的含量保持稳定, 可维持细胞正常的生理功能.在外界生物和非生物胁迫下, 细胞质中的Ca2+浓度会瞬间升高(Ca2+浓度的波动被认为是植物对外界刺激的反应信号) (Nürnberger and Scheel, 2001; Liu et al., 2017).Cd胁迫扰动植物细胞中的Ca2+水平, 从而扰乱植物叶片的水分状况(Perfus-Barbeoch et al., 2002).胞质Ca2+浓度增加对H2O2的生成有动态调控作用, Ca2+可调控NADH氧化酶活性, 上调植物中H2O2的含量; 也可通过刺激H2O2酶活性下调植物中的H2O2水平(Yang and Poovaiah, 2002).当ROS含量超过Ca2+调控的浓度范围时, 会对植物造成伤害, 细胞膜和细胞器膜发生膜质过氧化, 膜的通透性被破坏, 致使细胞死亡(赵士诚等, 2008).Cd与Ca2+的化学性质相似, Cd被植物吸收后通过金属转运体或钙通道运输至根部, 也可通过电压依赖的钙通道进入细胞, 一旦进入胞质, 它就会模拟钙离子的活性, 扰乱钙信号调控; 导致根系吸收大量的Cd, 并显著减少根长、根表面积、根体积和根系生物量, 使叶片黄化并干燥坏死(Yang and Poovaiah, 2002; Wang et al., 2016; Ismael et al., 2019).Cd进入植物根细胞也诱导一些对重金属胁迫有反应的特定信号分子, 如CaMs、钙依赖蛋白激酶(calcium-dependent protein kinases, CDPKs)和丝裂原活化蛋白激酶(mitogen-activated protein kinases, MAPKs), 调控植物对逆境的抗性(Danquah et al., 2014; Xu et al., 2017). ...
Cadmium toxicity and tolerance in vascular plants 1 1995
Endoplasmic reticulum stress and calcium imbalance are involved in cadmium-induced lipid aberrancy in Saccharomyces cerevisiae 1 2016
... 内质网(endoplasmic reticulum, ER)是一个复杂的代谢场所, 是调控蛋白质合成、信号转导和钙稳态的重要细胞器(Xu et al., 2013).所有分泌蛋白和大部分膜蛋白的合成和折叠及蛋白质的修饰和加工(主要包括糖基化、羟基化、酰基化和二硫键形成等)均在内质网进行(陈倩和谢旗, 2018).当植物受到干旱、高温、盐和重金属等胁迫后, 细胞蛋白质异常糖基化、Ca2+平衡紊乱及氧化还原状态改变会使细胞内积累大量未折叠或错误折叠的蛋白, 导致内质网胁迫(Kleizen and Braakman, 2004; Beaupere and Labunskyy, 2019).Cd诱导内质网应激, 改变内质网钙和脂质组成、ROS种类和错误折叠/未折叠蛋白(Rajakumar et al., 2016).分子伴侣参与内质网Cd胁迫的调控.BiP (binding protein)属于热休克蛋白(heat shock protein, HSP) HSP70家族, 是内质网腔内含量很高的分子伴侣, 参与内质网胁迫(Kleizen and Braakman, 2004).植物中, 程序性细胞死亡可通过ER调控应激-细胞死亡信号通路, BiP通过内质网应激-细胞死亡信号通路缓解Cd诱导的BY-2 (bright yellow-2)程序性死亡(Xu et al., 2013).同时, 内质网也是基因表达的重要场所.Wang等(2018)发现GmHMA3w主要在根内质网表达, 可将Cd从细胞壁运输至根内质网, 最终限制Cd从根向茎叶的转运.bZIP是位于根系质膜上的锌铁转运基因, 在拟南芥根细胞内质网应激信号转导中起重要作用(Deng et al., 2013). ...
Nitric oxide level is self-regulating and also regulates its ROS partners 4 2016
... 活性氧(reactive oxygen species, ROS)和活性氮(reactive nitrogen species, RNS)是植物产生的具有一定功能的活性分子化合物, 是植物适应环境变化的主要调节因子.低含量水平下, ROS和RNS是信号分子(Romero-Puertas and Sandalio, 2016); 过多积累后, 会诱导植物氧化损伤及细胞程序性死亡, 是植物响应胁迫的关键指标(Das and Roychoudhury, 2014; Nieves-Cordones et al., 2019).Cd毒害最常见的表现是诱导植物出现氧化应激反应, 如产生ROS和RNS (Romero-Puertas et al., 2019).ROS和RNS之间存在动态平衡(Delledonne et al., 2001).ROS通过多种反应产生, 包括呼吸和光合电子传递链及酶(如过氧化物酶)的副反应等; Cd胁迫下, 植物细胞中产生ROS的部位有线粒体、叶绿体、过氧化物酶体、细胞质以及外质体等(Romero-Puertas et al., 2019).ROS包括单线态氧(single oxygen, 1O2)、超氧自由基阴离子(superoxide radical, O2-.)、过氧化氢(hydrogen peroxide, H2O2)和羟基自由基(hydroxyl radical, ·OH) (Nieves-Cordones et al., 2019).清除ROS的机制涉及酶促抗氧化剂和非酶类抗氧化剂(1.1.3节).ROS会对脂质、蛋白质和DNA等生物分子造成损害, 破坏细胞的完整性, 最终导致细胞死亡(Das and Roychoudhury, 2014).ROS作为信号分子在多种生命过程中扮演重要角色, 包括趋向性、细胞死亡和细胞对生物和非生物胁迫的反应(Mittler et al., 2011). ...
... 活性氮(RNS)主要包括一氧化氮(NO)、S-亚硝基硫醇(SNOs)、高氮氧化物(NOx)、二硝基铁配合物和过氧亚硝酸盐(ONOO) 5种.过量的RNS会与脂类、蛋白质和核酸发生反应, 导致细胞损害(Neill et al., 2008a; Romero-Puertas and Sandalio, 2016).然而, 细胞内NO一旦产生, 就会与ROS迅速作用, 产生大量的RNS (如NO2), 然后降解为亚硝酸盐和硝酸钠(Neill et al., 2008b).低水平的NO可调节植物的发育, 而在胁迫条件下植物体内会产生大量的NO (Romero-Puertas and Sandalio, 2016; Liu et al., 2018); NO浓度过高则抑制植物的生长, 甚至引起细胞程序性死亡(Arasimowicz-Jelonek et al., 2012; Sami et al., 2018).低浓度NO促进植物生长, 高浓度则抑制植物生长, 其与生长素(indole-3-acetic acid, IAA)、细胞分裂素、赤霉素、脱落酸(abscisic acid, ABA)、乙烯、茉莉酸(jasmonic acid, JA)和水杨酸(salicylic acid, SA)也有一定的交互作用(Sami et al., 2018).Cd能通过破坏呼吸和光合作用的电子传递链使电子被传递到非靶向分子, 从而导致ROS和RNS的产生, 且Cd能通过抑制自由基降低相关酶活性(Ekmek?i et al., 2008; Romero-Puertas and Sandalio, 2016), 导致ROS和RNS不能够被及时清除而过量累积, 最终引起植物营养应激反应(Nieves-Cordones et al., 2019).外施氮(Zhang et al., 2019b)、磷(Wang et al., 2020)、钾(Wu et al., 2020)、钙(Li et al., 2021)和硅(Vaculík et al., 2015)等植物营养元素, 可起到调控植物激素含量、非酶物质和抗氧化酶活性及转运基因表达等作用, 减轻Cd对植物的毒害, 但外施营养物质对ROS和RNS有怎样的影响有待后续探索. ...
... ).低水平的NO可调节植物的发育, 而在胁迫条件下植物体内会产生大量的NO (Romero-Puertas and Sandalio, 2016; Liu et al., 2018); NO浓度过高则抑制植物的生长, 甚至引起细胞程序性死亡(Arasimowicz-Jelonek et al., 2012; Sami et al., 2018).低浓度NO促进植物生长, 高浓度则抑制植物生长, 其与生长素(indole-3-acetic acid, IAA)、细胞分裂素、赤霉素、脱落酸(abscisic acid, ABA)、乙烯、茉莉酸(jasmonic acid, JA)和水杨酸(salicylic acid, SA)也有一定的交互作用(Sami et al., 2018).Cd能通过破坏呼吸和光合作用的电子传递链使电子被传递到非靶向分子, 从而导致ROS和RNS的产生, 且Cd能通过抑制自由基降低相关酶活性(Ekmek?i et al., 2008; Romero-Puertas and Sandalio, 2016), 导致ROS和RNS不能够被及时清除而过量累积, 最终引起植物营养应激反应(Nieves-Cordones et al., 2019).外施氮(Zhang et al., 2019b)、磷(Wang et al., 2020)、钾(Wu et al., 2020)、钙(Li et al., 2021)和硅(Vaculík et al., 2015)等植物营养元素, 可起到调控植物激素含量、非酶物质和抗氧化酶活性及转运基因表达等作用, 减轻Cd对植物的毒害, 但外施营养物质对ROS和RNS有怎样的影响有待后续探索. ...
... ; Romero-Puertas and Sandalio, 2016), 导致ROS和RNS不能够被及时清除而过量累积, 最终引起植物营养应激反应(Nieves-Cordones et al., 2019).外施氮(Zhang et al., 2019b)、磷(Wang et al., 2020)、钾(Wu et al., 2020)、钙(Li et al., 2021)和硅(Vaculík et al., 2015)等植物营养元素, 可起到调控植物激素含量、非酶物质和抗氧化酶活性及转运基因表达等作用, 减轻Cd对植物的毒害, 但外施营养物质对ROS和RNS有怎样的影响有待后续探索. ...
Reactive oxygen and nitrogen species as key indicators of plant responses to Cd stress 2 2019
... 活性氧(reactive oxygen species, ROS)和活性氮(reactive nitrogen species, RNS)是植物产生的具有一定功能的活性分子化合物, 是植物适应环境变化的主要调节因子.低含量水平下, ROS和RNS是信号分子(Romero-Puertas and Sandalio, 2016); 过多积累后, 会诱导植物氧化损伤及细胞程序性死亡, 是植物响应胁迫的关键指标(Das and Roychoudhury, 2014; Nieves-Cordones et al., 2019).Cd毒害最常见的表现是诱导植物出现氧化应激反应, 如产生ROS和RNS (Romero-Puertas et al., 2019).ROS和RNS之间存在动态平衡(Delledonne et al., 2001).ROS通过多种反应产生, 包括呼吸和光合电子传递链及酶(如过氧化物酶)的副反应等; Cd胁迫下, 植物细胞中产生ROS的部位有线粒体、叶绿体、过氧化物酶体、细胞质以及外质体等(Romero-Puertas et al., 2019).ROS包括单线态氧(single oxygen, 1O2)、超氧自由基阴离子(superoxide radical, O2-.)、过氧化氢(hydrogen peroxide, H2O2)和羟基自由基(hydroxyl radical, ·OH) (Nieves-Cordones et al., 2019).清除ROS的机制涉及酶促抗氧化剂和非酶类抗氧化剂(1.1.3节).ROS会对脂质、蛋白质和DNA等生物分子造成损害, 破坏细胞的完整性, 最终导致细胞死亡(Das and Roychoudhury, 2014).ROS作为信号分子在多种生命过程中扮演重要角色, 包括趋向性、细胞死亡和细胞对生物和非生物胁迫的反应(Mittler et al., 2011). ...
... ).ROS通过多种反应产生, 包括呼吸和光合电子传递链及酶(如过氧化物酶)的副反应等; Cd胁迫下, 植物细胞中产生ROS的部位有线粒体、叶绿体、过氧化物酶体、细胞质以及外质体等(Romero-Puertas et al., 2019).ROS包括单线态氧(single oxygen, 1O2)、超氧自由基阴离子(superoxide radical, O2-.)、过氧化氢(hydrogen peroxide, H2O2)和羟基自由基(hydroxyl radical, ·OH) (Nieves-Cordones et al., 2019).清除ROS的机制涉及酶促抗氧化剂和非酶类抗氧化剂(1.1.3节).ROS会对脂质、蛋白质和DNA等生物分子造成损害, 破坏细胞的完整性, 最终导致细胞死亡(Das and Roychoudhury, 2014).ROS作为信号分子在多种生命过程中扮演重要角色, 包括趋向性、细胞死亡和细胞对生物和非生物胁迫的反应(Mittler et al., 2011). ...
WRKY transcription factors 2 2010
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
... 转录因子(transcription factors, TFs)是一类DNA结合蛋白, 参与植物的各种生理过程, 涉及生长、发育、自我应激信号转导或与不同的基因和转录因子交叉调节(Erpen et al., 2018).转录因子可通过控制下游基因的表达调控Cd胁迫反应, 是Cd解毒和耐受调控网络中的核心成分.目前, 已发现植物中大多数类型的Cd解毒和耐受转录因子.其中, WRKY、MYB和HSF等转录因子在调节植物Cd胁迫响应方面具有重要作用.WRKY TFs是植物中最大的转录因子家族之一, 拥有强大的转录调控功能(ülker and Somssich, 2004; Erpen et al., 2018).WRKY TFs不仅可通过调节植物激素信号转导途径, 还可结合其靶基因启动子通过激活或抑制下游基因的表达来调节它们的应激反应(Rushton et al., 2010).Hong等(2017)研究发现, Cd胁迫下玉米ZmWRKY4基因的表达上调, SOD和APX活性升高.ZmWRKY4在玉米叶肉细胞原生质体中的瞬时表达和RNA干扰(RNAi)进一步表明, ZmWRKY4是ABA诱导SOD和APX表达及活性增强所必需; ZmWRKY4在原生质体中的过表达上调了抗氧化酶的表达和活性, 而ABA诱导的抗氧化酶的表达和活性增强被ZmWRKY4的RNAi基因沉默所阻断.Cai等(2020)对大豆(Glycine max)的研究表明, GmWRKY4直接作用于ATCDT1、GmCDT1-1和GmCDT1-2, 从而减少Cd的摄取并积极调节对Cd胁迫的耐受性.GmWRKY142-GmCDT1-1/2级联模块为降低Cd在大豆中积累提供了潜在策略. ...
Nitric oxide-mediated integrative alterations in plant metabolism to confer abiotic stress tolerance, NO crosstalk with phytohormones and NO-mediated post translational modifications in modulating diverse plant stress 2 2018
... 活性氮(RNS)主要包括一氧化氮(NO)、S-亚硝基硫醇(SNOs)、高氮氧化物(NOx)、二硝基铁配合物和过氧亚硝酸盐(ONOO) 5种.过量的RNS会与脂类、蛋白质和核酸发生反应, 导致细胞损害(Neill et al., 2008a; Romero-Puertas and Sandalio, 2016).然而, 细胞内NO一旦产生, 就会与ROS迅速作用, 产生大量的RNS (如NO2), 然后降解为亚硝酸盐和硝酸钠(Neill et al., 2008b).低水平的NO可调节植物的发育, 而在胁迫条件下植物体内会产生大量的NO (Romero-Puertas and Sandalio, 2016; Liu et al., 2018); NO浓度过高则抑制植物的生长, 甚至引起细胞程序性死亡(Arasimowicz-Jelonek et al., 2012; Sami et al., 2018).低浓度NO促进植物生长, 高浓度则抑制植物生长, 其与生长素(indole-3-acetic acid, IAA)、细胞分裂素、赤霉素、脱落酸(abscisic acid, ABA)、乙烯、茉莉酸(jasmonic acid, JA)和水杨酸(salicylic acid, SA)也有一定的交互作用(Sami et al., 2018).Cd能通过破坏呼吸和光合作用的电子传递链使电子被传递到非靶向分子, 从而导致ROS和RNS的产生, 且Cd能通过抑制自由基降低相关酶活性(Ekmek?i et al., 2008; Romero-Puertas and Sandalio, 2016), 导致ROS和RNS不能够被及时清除而过量累积, 最终引起植物营养应激反应(Nieves-Cordones et al., 2019).外施氮(Zhang et al., 2019b)、磷(Wang et al., 2020)、钾(Wu et al., 2020)、钙(Li et al., 2021)和硅(Vaculík et al., 2015)等植物营养元素, 可起到调控植物激素含量、非酶物质和抗氧化酶活性及转运基因表达等作用, 减轻Cd对植物的毒害, 但外施营养物质对ROS和RNS有怎样的影响有待后续探索. ...
... ).低浓度NO促进植物生长, 高浓度则抑制植物生长, 其与生长素(indole-3-acetic acid, IAA)、细胞分裂素、赤霉素、脱落酸(abscisic acid, ABA)、乙烯、茉莉酸(jasmonic acid, JA)和水杨酸(salicylic acid, SA)也有一定的交互作用(Sami et al., 2018).Cd能通过破坏呼吸和光合作用的电子传递链使电子被传递到非靶向分子, 从而导致ROS和RNS的产生, 且Cd能通过抑制自由基降低相关酶活性(Ekmek?i et al., 2008; Romero-Puertas and Sandalio, 2016), 导致ROS和RNS不能够被及时清除而过量累积, 最终引起植物营养应激反应(Nieves-Cordones et al., 2019).外施氮(Zhang et al., 2019b)、磷(Wang et al., 2020)、钾(Wu et al., 2020)、钙(Li et al., 2021)和硅(Vaculík et al., 2015)等植物营养元素, 可起到调控植物激素含量、非酶物质和抗氧化酶活性及转运基因表达等作用, 减轻Cd对植物的毒害, 但外施营养物质对ROS和RNS有怎样的影响有待后续探索. ...
Response to cadmium in higher plants 1 1999
... 镉(Cd)因其在土壤中移动性强、毒性大、污染范围广和检出频率高等原因, 被联合国环境计划署(United Nations Environment Programme, UNEP)列入可引起人类癌症的致癌物质(UNEP, 2008; DalCorso et al., 2008; 付铄岚等, 2017).土壤中的Cd被植物吸收后, 会随着食物链在高营养级生物体内富集, 诱发人体骨质疏松并损坏肾脏, 导致人体慢性中毒, 甚至出现免疫系统、神经系统及生殖系统损伤等疾病, 对人类健康造成严重的威胁(Lalor, 2008; Clemens et al., 2013).由于Cd对各种环境及生物体的长期毒性作用, 其已成为全社会关注的焦点(Nagajyoti et al., 2010; Etesami and Jeong, 2018).有研究表明, 植物对Cd污染会产生一系列的应激反应, 如氧化应激、酶活性和植物信号物质(激素和钙离子)失衡, 导致光合系统受损、质膜过氧化、细胞损伤、酶活性改变、内质网胁迫、蛋白质降解、DNA损伤或突变, 从而影响其生理生化代谢过程, 最终使植物生长受到抑制, 甚至死亡(Sanità di Toppi and Gabbrielli, 1999; DalCorso et al., 2008; Nagajyoti et al., 2010; 杨正婷和刘建祥, 2016; Shahid et al., 2019; 张瑛等, 2019) (图1).此外, 植物对胁迫的响应是一个联合的信号通路和网络调控(郭倩倩和周文彬, 2019; 苗青霞等, 2019).目前, 国内外在植物对镉的吸收、积累和转运或镉对植物产生的毒害和抗镉生理及分子机制等方面已有大量研究, 而有关植物对重金属镉胁迫的主要代谢过程的应答机制报道较少.本文综述了植物响应镉胁迫的生理生化代谢变化及其应答机制, 为深入理解镉胁迫下植物的代谢差异、认识植物的抗镉策略, 及未来作物育种和环境修复提供理论依据. ...
Selenium impedes cadmium and arsenic toxicity in potato by modulating carbohydrate and nitrogen metabolism 2 2019
... 镉(Cd)因其在土壤中移动性强、毒性大、污染范围广和检出频率高等原因, 被联合国环境计划署(United Nations Environment Programme, UNEP)列入可引起人类癌症的致癌物质(UNEP, 2008; DalCorso et al., 2008; 付铄岚等, 2017).土壤中的Cd被植物吸收后, 会随着食物链在高营养级生物体内富集, 诱发人体骨质疏松并损坏肾脏, 导致人体慢性中毒, 甚至出现免疫系统、神经系统及生殖系统损伤等疾病, 对人类健康造成严重的威胁(Lalor, 2008; Clemens et al., 2013).由于Cd对各种环境及生物体的长期毒性作用, 其已成为全社会关注的焦点(Nagajyoti et al., 2010; Etesami and Jeong, 2018).有研究表明, 植物对Cd污染会产生一系列的应激反应, 如氧化应激、酶活性和植物信号物质(激素和钙离子)失衡, 导致光合系统受损、质膜过氧化、细胞损伤、酶活性改变、内质网胁迫、蛋白质降解、DNA损伤或突变, 从而影响其生理生化代谢过程, 最终使植物生长受到抑制, 甚至死亡(Sanità di Toppi and Gabbrielli, 1999; DalCorso et al., 2008; Nagajyoti et al., 2010; 杨正婷和刘建祥, 2016; Shahid et al., 2019; 张瑛等, 2019) (图1).此外, 植物对胁迫的响应是一个联合的信号通路和网络调控(郭倩倩和周文彬, 2019; 苗青霞等, 2019).目前, 国内外在植物对镉的吸收、积累和转运或镉对植物产生的毒害和抗镉生理及分子机制等方面已有大量研究, 而有关植物对重金属镉胁迫的主要代谢过程的应答机制报道较少.本文综述了植物响应镉胁迫的生理生化代谢变化及其应答机制, 为深入理解镉胁迫下植物的代谢差异、认识植物的抗镉策略, 及未来作物育种和环境修复提供理论依据. ...
Calmodulin as a versatile calcium signal transducer in plants 2 2001
... Ca2+作为细胞功能的第二信使, 参与调节植物各种逆境下的重要生理生化过程.钙信使系统包括Ca2+、钙调蛋白(CaM)和钙泵(Ca2+-ATPase).CaM是Ca2+的感受器, Ca2+-ATPase决定着Ca2+的跨膜运输(Snedden and Fromm, 2001).正常情况下三者的含量保持稳定, 可维持细胞正常的生理功能.在外界生物和非生物胁迫下, 细胞质中的Ca2+浓度会瞬间升高(Ca2+浓度的波动被认为是植物对外界刺激的反应信号) (Nürnberger and Scheel, 2001; Liu et al., 2017).Cd胁迫扰动植物细胞中的Ca2+水平, 从而扰乱植物叶片的水分状况(Perfus-Barbeoch et al., 2002).胞质Ca2+浓度增加对H2O2的生成有动态调控作用, Ca2+可调控NADH氧化酶活性, 上调植物中H2O2的含量; 也可通过刺激H2O2酶活性下调植物中的H2O2水平(Yang and Poovaiah, 2002).当ROS含量超过Ca2+调控的浓度范围时, 会对植物造成伤害, 细胞膜和细胞器膜发生膜质过氧化, 膜的通透性被破坏, 致使细胞死亡(赵士诚等, 2008).Cd与Ca2+的化学性质相似, Cd被植物吸收后通过金属转运体或钙通道运输至根部, 也可通过电压依赖的钙通道进入细胞, 一旦进入胞质, 它就会模拟钙离子的活性, 扰乱钙信号调控; 导致根系吸收大量的Cd, 并显著减少根长、根表面积、根体积和根系生物量, 使叶片黄化并干燥坏死(Yang and Poovaiah, 2002; Wang et al., 2016; Ismael et al., 2019).Cd进入植物根细胞也诱导一些对重金属胁迫有反应的特定信号分子, 如CaMs、钙依赖蛋白激酶(calcium-dependent protein kinases, CDPKs)和丝裂原活化蛋白激酶(mitogen-activated protein kinases, MAPKs), 调控植物对逆境的抗性(Danquah et al., 2014; Xu et al., 2017). ...
... 质膜上的Ca2+-ATPase能使高浓度的Ca2+进入非原生质体空间, 调节胁迫下细胞内的Ca2+平衡和维持细胞相对稳定的内环境(Snedden and Fromm, 2001; Hu et al., 2007).赵士诚等(2008)用不同浓度的Cd处理玉米叶片, 其Ca2+-ATPase活性均表现为细胞质膜>液泡膜、内质网膜>线粒体膜.并且低浓度Cd处理下, 为保持细胞内的Ca2+平衡, 玉米叶片生物膜系统的Ca2+-ATPase活性增强; 高浓度Cd处理下, Ca2+-ATPase活性下降, 出现代谢紊乱, 玉米中下部叶变黄且生长停滞.Ca预处理可显著降低扁豆(Lablab purpureus)幼苗的Cd积累, 并通过调节H2O2酶活性和降低膜的氧化损伤减轻Cd处理的不利影响(Talukdar, 2012).林啸等(2014)发现外源Ca处理可降低白菜(Brassica pekinensis)叶片的Cd含量, 并通过Ca2+-CaM信号途径调节酶类抗氧化剂APX、CAT和POD以及非酶类抗氧化剂GSH含量, 维持叶片ROS的平衡, 减少Cd诱导的细胞程序性死亡, 从而增强植物对Cd的耐受性.植物钙信号与抗氧化系统的关联机制值得进一步研究. ...
Correlation between hormonal homeostasis and morphogenic responses in Arabidopsis thaliana seedlings growing in a Cd/Cu/Zn multi-pollution context 1 2013
... Cd显著影响抗氧化酶类物质和非酶抗氧化物质的基因表达.不同浓度Cd处理的玉米幼苗, SOD基因的表达随着Cd浓度的升高先上升, 后随着时间的延长而下降; CAT基因的表达随着Cd浓度的升高和处理时间的延长逐渐增强(赵士诚等, 2008).高浓度Cd处理下, 抗氧化酶基因GR-chl、CAT3、APX-cyt、APX-per和MTLa的表达上调(Zhang et al., 2019b).当植物遭受Cd胁迫时, 金属硫蛋白(metallothionein)基因MTs、APX、GPX和DHAR等表达上调(Cobbett and Goldsbrough, 2002; Tamás et al., 2008).γ-谷氨酰半胱氨酸合成酶(γ-glutamylcysteine synthetase, γ-GCS)、谷胱甘肽合成酶(glutathione synthetase, GSHs)、胱硫醚β-合酶(cystathionine beta-synthase, CBS)、ATP硫酸化酶(ATP sulfurylase, ATPS)、GR、PCS和乙二醛酶(glyoxalase) I和II等合成基因是调节重金属耐受性的潜在候选基因(Yadav, 2010; Sofoet al., 2013).细胞壁果胶(pectin)在重金属结合中起重要作用, 根、茎和叶细胞壁的酯化及果胶酶修饰减少了Cd的积累.番茄经Cd处理后, 其细胞壁果胶相关基因SlQUA1和SlPME1表达上调, 果胶甲基脂酶活性明显升高(Jia et al., 2019). ...
Transcriptome analysis of Cd-treated switchgrass root revealed novel transcripts and the importance of HSF/HSP network in switchgrass Cd tolerance 2 2018
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
... HSP不仅能对抗应激相关损伤与保护细胞, 而且在蛋白质规则折叠、细胞内分布和降解中也发挥重要作用.这些功能有利于生物在压力条件下生存(Hartl and Hayer-Hart, 2002; Zhou et al., 2019).热休克转录因子HSFs (heat shock transcription factors)调节HSP的表达, 参与蛋白稳态的各个方面(如受损蛋白的重新折叠、组装和运输), 维持蛋白的稳定性(Wu, 1995; Guo et al., 2008).TaHsfA4a上调表达的水稻对Cd的耐受性增强, OsHsfA4a下调表达的水稻对Cd的耐受性减弱, 在小麦和水稻根系中, Cd胁迫导致HsfA4a以及金属硫蛋白基因的表达上调(Shim et al., 2009).此外, Chen等(2020)研究结果也表明, SaHsfA4c通过上调HSP的表达和提高ROS清除酶活性, 增强植物的抗逆性.Cd胁迫下, HSF/HSP参与柳枝稷(Panicum virgatum)正常蛋白质构象的重建以及细胞内稳态调节(Song et al., 2018).这些HSF/HSP表达基因可作为进一步研究的候选基因, 以提高植物对Cd的耐受性. ...
Mechanisms underlying enhanced Cd translocation and tolerance in roots of Populus euramericana in response to nitrogen fertilization 4 2019
... Cd胁迫下, 抗逆相关的植物激素(如ABA、IAA、SA和JA)均会发生显著变化, 且影响抗氧化物质合成相关基因和运输蛋白基因的表达(表1).多数情况下Cd胁迫可引起植物体内IAA、SA、ABA和JA含量变化(Liu et al., 2016; Bashir et al., 2019; Song et al., 2019).添加适量外源ABA、IAA、SA和JA等可缓解Cd胁迫对植物的危害. ...
... Responses of plant hormones to Cd stress Table 1
激素
激素合成基因
非酶物质
抗氧化酶
抗氧化物质基因
吸收和运输蛋白基因
ABA
NCED3和Glyma17G242200 (Bashir et al., 2019; Song et al., 2019)
AsA和GSH (Hu et al., 2005; Stroiński et al., 2010; Li et al., 2014)
APX、POD、SOD和CAT (Hu et al., 2005)
StPCS 1 (Stroiń- ski et al., 2010)
IRT1 (Fan et al., 2014)
IAA
Glyma02G037600.1 (Bashir et al., 2019)
GSH (Hu et al., 2013)
SOD、POD、CAT和GST (Bo?ová et al., 2013; 陈晶等, 2016)
SODs (Pan et al., 2020)
Nramp、IRT、HMA和ZIP (Korshunova et al., 1999; 罗莎, 2017)
SA
Glyma02G063400 (Bashir et al., 2019)
脯氨酸(Krantev et al., 2008; 胡春霞和王秀芹, 2010)
NR、GS和GOGAT (Zawoznik et al., 2007; Chao et al., 2010; 赵新月等, 2013)
SODS、CATS和APXS (Gill, 2015)
OsLCT1、OsLCD和ZIP (Liu et al., 2016)
JA
PtJMT1和Glyma11G007600. 1 (Bashir et al., 2019; Song et al., 2019)
GSH (Noriega et al., 2012; Zhao et al., 2016)
CAT、SOD和APX (Noriega et al., 2012; Zhao et al., 2016)
Exogenous calcium alleviates the impact of cadmium-induced oxidative stress in Lens culinaris Medic. Seedlings through modulation of antioxidant enzyme activities 1 2012
... 质膜上的Ca2+-ATPase能使高浓度的Ca2+进入非原生质体空间, 调节胁迫下细胞内的Ca2+平衡和维持细胞相对稳定的内环境(Snedden and Fromm, 2001; Hu et al., 2007).赵士诚等(2008)用不同浓度的Cd处理玉米叶片, 其Ca2+-ATPase活性均表现为细胞质膜>液泡膜、内质网膜>线粒体膜.并且低浓度Cd处理下, 为保持细胞内的Ca2+平衡, 玉米叶片生物膜系统的Ca2+-ATPase活性增强; 高浓度Cd处理下, Ca2+-ATPase活性下降, 出现代谢紊乱, 玉米中下部叶变黄且生长停滞.Ca预处理可显著降低扁豆(Lablab purpureus)幼苗的Cd积累, 并通过调节H2O2酶活性和降低膜的氧化损伤减轻Cd处理的不利影响(Talukdar, 2012).林啸等(2014)发现外源Ca处理可降低白菜(Brassica pekinensis)叶片的Cd含量, 并通过Ca2+-CaM信号途径调节酶类抗氧化剂APX、CAT和POD以及非酶类抗氧化剂GSH含量, 维持叶片ROS的平衡, 减少Cd诱导的细胞程序性死亡, 从而增强植物对Cd的耐受性.植物钙信号与抗氧化系统的关联机制值得进一步研究. ...
Impact of the auxin signaling inhibitor p-chlorophenoxyisobutyric acid on short- term Cd-induced hydrogen peroxide production and growth response in barley root tip 1 2012
... Cd处理的植物初级根尖和子叶中IAA浓度及分布有较大差异.Cd处理后, IAA含量显著降低(Guo et al., 2019), IAA合成基因表达下调(Bashir et al., 2019), IAA氧化酶活性增强, IAA生物合成和分解代谢基因表达改变(Hu et al., 2013).此外, Cd胁迫下, 外源IAA能显著提高玉米幼苗根和地上部生物量, 降低地上部Cd含量, 但对根Cd含量的影响不显著, 显著降低叶片的MDA和GSH含量, 使玉米幼苗叶片SOD、POD和CAT活性显著增强(陈晶等, 2016).当IAA合成受到抑制时, 如施加生长素信号抑制剂氯苯氧基异丁酸, 可有效抑制10 μmol·L-1 Cd诱导的根系生长和H2O2产生, 但不抑制60 μmol·L-1 Cd诱导的根系生长和H2O2产生(Tamás et al., 2012).Cd胁迫下施加IAA, 可激活谷胱甘肽-S-转移酶(glutathione- S-transferase, GST) (在植物解毒过程中起关键作用)活性, 使植物更好地发生应激反应(Bo?ová et al., 2013).Cd胁迫下, 施加外源IAA还可显著提高SODs基因的表达, 降低H2O2含量, 从而减少Cd对植物的毒害(Pan et al., 2020).重金属污染条件下, 外源IAA可促进天然抗性相关巨噬细胞蛋白基因NRAMP (NATURAL RESISTANCE-ASSOCIATED MACROPHAGE PROTEIN)、IRT、重金属ATP酶家族基因HMA (heavy metal ATPase)和锌铁转运蛋白基因ZIP (zinc iron transporter protein)的表达, 提高植物的抗Cd能力(Korshunova et al., 1999; 罗莎, 2017). ...
Alterations of the gene expression, lipid peroxidation, proline and thiol content along the barley root exposed to cadmium 1 2008
... Cd显著影响抗氧化酶类物质和非酶抗氧化物质的基因表达.不同浓度Cd处理的玉米幼苗, SOD基因的表达随着Cd浓度的升高先上升, 后随着时间的延长而下降; CAT基因的表达随着Cd浓度的升高和处理时间的延长逐渐增强(赵士诚等, 2008).高浓度Cd处理下, 抗氧化酶基因GR-chl、CAT3、APX-cyt、APX-per和MTLa的表达上调(Zhang et al., 2019b).当植物遭受Cd胁迫时, 金属硫蛋白(metallothionein)基因MTs、APX、GPX和DHAR等表达上调(Cobbett and Goldsbrough, 2002; Tamás et al., 2008).γ-谷氨酰半胱氨酸合成酶(γ-glutamylcysteine synthetase, γ-GCS)、谷胱甘肽合成酶(glutathione synthetase, GSHs)、胱硫醚β-合酶(cystathionine beta-synthase, CBS)、ATP硫酸化酶(ATP sulfurylase, ATPS)、GR、PCS和乙二醛酶(glyoxalase) I和II等合成基因是调节重金属耐受性的潜在候选基因(Yadav, 2010; Sofoet al., 2013).细胞壁果胶(pectin)在重金属结合中起重要作用, 根、茎和叶细胞壁的酯化及果胶酶修饰减少了Cd的积累.番茄经Cd处理后, 其细胞壁果胶相关基因SlQUA1和SlPME1表达上调, 果胶甲基脂酶活性明显升高(Jia et al., 2019). ...
Elevated expression of TcHMA3 plays a key role in the extreme Cd tolerance in a Cd-hyperaccumulating ecotype of Thlaspi caerulescens 1 2011
... 重金属ATP酶基因家族(HMA)已被证明在不同的植物种类中行使特定的功能, 不同的HMA在不同组织中表达, 最终导致功能的多样性.HvHMA1参与Zn以及Cd向大麦(Hordeum vulgare)籽粒的转运(Mikkelsen et al., 2012).HMA2和HMA4可将Cd装载到木质部, 以便从根运输至茎和叶, OsHMA2与水稻维管组织中的锌装载及液泡膜定位有关(Yamaji et al., 2013).液泡膜上的OsHMA3将Cd转移至根系液泡, OsHMA4则将Cu转运至根系液泡, 限制Cd从根向茎以及叶的转移, 从而减少Cd在种子中的积累(Miyadate et al., 2011; Huang et al., 2016).TcHMA3在叶片的液泡中表达, 将Cd隔离在根和叶的液泡中, 最终导致Cd的超积累, 增强对Cd的耐受性(Ueno et al., 2011).AtHMA3在拟南芥中过表达导致Cd的积累量比野生型高2-3倍(Morel et al., 2009).这些转运蛋白为植物的基因工程提供了有用的工具, 增强了植物对金属的耐受性和积累能力, 继续推进它们的协同作用研究具有重要意义. ...
WRKY transcription factors: from DNA binding towards biological function 2 2004
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
... 活性氮(RNS)主要包括一氧化氮(NO)、S-亚硝基硫醇(SNOs)、高氮氧化物(NOx)、二硝基铁配合物和过氧亚硝酸盐(ONOO) 5种.过量的RNS会与脂类、蛋白质和核酸发生反应, 导致细胞损害(Neill et al., 2008a; Romero-Puertas and Sandalio, 2016).然而, 细胞内NO一旦产生, 就会与ROS迅速作用, 产生大量的RNS (如NO2), 然后降解为亚硝酸盐和硝酸钠(Neill et al., 2008b).低水平的NO可调节植物的发育, 而在胁迫条件下植物体内会产生大量的NO (Romero-Puertas and Sandalio, 2016; Liu et al., 2018); NO浓度过高则抑制植物的生长, 甚至引起细胞程序性死亡(Arasimowicz-Jelonek et al., 2012; Sami et al., 2018).低浓度NO促进植物生长, 高浓度则抑制植物生长, 其与生长素(indole-3-acetic acid, IAA)、细胞分裂素、赤霉素、脱落酸(abscisic acid, ABA)、乙烯、茉莉酸(jasmonic acid, JA)和水杨酸(salicylic acid, SA)也有一定的交互作用(Sami et al., 2018).Cd能通过破坏呼吸和光合作用的电子传递链使电子被传递到非靶向分子, 从而导致ROS和RNS的产生, 且Cd能通过抑制自由基降低相关酶活性(Ekmek?i et al., 2008; Romero-Puertas and Sandalio, 2016), 导致ROS和RNS不能够被及时清除而过量累积, 最终引起植物营养应激反应(Nieves-Cordones et al., 2019).外施氮(Zhang et al., 2019b)、磷(Wang et al., 2020)、钾(Wu et al., 2020)、钙(Li et al., 2021)和硅(Vaculík et al., 2015)等植物营养元素, 可起到调控植物激素含量、非酶物质和抗氧化酶活性及转运基因表达等作用, 减轻Cd对植物的毒害, 但外施营养物质对ROS和RNS有怎样的影响有待后续探索. ...
Effect of cadmium on soluble sugars and enzymes of their metabolism in rice 1 2001
Phosphorus deficiency induces root proliferation and Cd absorption but inhibits Cd tolerance and Cd translocation in roots of Populus × euramericana 1 2020
... 活性氮(RNS)主要包括一氧化氮(NO)、S-亚硝基硫醇(SNOs)、高氮氧化物(NOx)、二硝基铁配合物和过氧亚硝酸盐(ONOO) 5种.过量的RNS会与脂类、蛋白质和核酸发生反应, 导致细胞损害(Neill et al., 2008a; Romero-Puertas and Sandalio, 2016).然而, 细胞内NO一旦产生, 就会与ROS迅速作用, 产生大量的RNS (如NO2), 然后降解为亚硝酸盐和硝酸钠(Neill et al., 2008b).低水平的NO可调节植物的发育, 而在胁迫条件下植物体内会产生大量的NO (Romero-Puertas and Sandalio, 2016; Liu et al., 2018); NO浓度过高则抑制植物的生长, 甚至引起细胞程序性死亡(Arasimowicz-Jelonek et al., 2012; Sami et al., 2018).低浓度NO促进植物生长, 高浓度则抑制植物生长, 其与生长素(indole-3-acetic acid, IAA)、细胞分裂素、赤霉素、脱落酸(abscisic acid, ABA)、乙烯、茉莉酸(jasmonic acid, JA)和水杨酸(salicylic acid, SA)也有一定的交互作用(Sami et al., 2018).Cd能通过破坏呼吸和光合作用的电子传递链使电子被传递到非靶向分子, 从而导致ROS和RNS的产生, 且Cd能通过抑制自由基降低相关酶活性(Ekmek?i et al., 2008; Romero-Puertas and Sandalio, 2016), 导致ROS和RNS不能够被及时清除而过量累积, 最终引起植物营养应激反应(Nieves-Cordones et al., 2019).外施氮(Zhang et al., 2019b)、磷(Wang et al., 2020)、钾(Wu et al., 2020)、钙(Li et al., 2021)和硅(Vaculík et al., 2015)等植物营养元素, 可起到调控植物激素含量、非酶物质和抗氧化酶活性及转运基因表达等作用, 减轻Cd对植物的毒害, 但外施营养物质对ROS和RNS有怎样的影响有待后续探索. ...
Root morphological responses of five soybean (Glycine max(L.) Merr) cultivars to cadmium stress at young seedlings 1 2016
... Ca2+作为细胞功能的第二信使, 参与调节植物各种逆境下的重要生理生化过程.钙信使系统包括Ca2+、钙调蛋白(CaM)和钙泵(Ca2+-ATPase).CaM是Ca2+的感受器, Ca2+-ATPase决定着Ca2+的跨膜运输(Snedden and Fromm, 2001).正常情况下三者的含量保持稳定, 可维持细胞正常的生理功能.在外界生物和非生物胁迫下, 细胞质中的Ca2+浓度会瞬间升高(Ca2+浓度的波动被认为是植物对外界刺激的反应信号) (Nürnberger and Scheel, 2001; Liu et al., 2017).Cd胁迫扰动植物细胞中的Ca2+水平, 从而扰乱植物叶片的水分状况(Perfus-Barbeoch et al., 2002).胞质Ca2+浓度增加对H2O2的生成有动态调控作用, Ca2+可调控NADH氧化酶活性, 上调植物中H2O2的含量; 也可通过刺激H2O2酶活性下调植物中的H2O2水平(Yang and Poovaiah, 2002).当ROS含量超过Ca2+调控的浓度范围时, 会对植物造成伤害, 细胞膜和细胞器膜发生膜质过氧化, 膜的通透性被破坏, 致使细胞死亡(赵士诚等, 2008).Cd与Ca2+的化学性质相似, Cd被植物吸收后通过金属转运体或钙通道运输至根部, 也可通过电压依赖的钙通道进入细胞, 一旦进入胞质, 它就会模拟钙离子的活性, 扰乱钙信号调控; 导致根系吸收大量的Cd, 并显著减少根长、根表面积、根体积和根系生物量, 使叶片黄化并干燥坏死(Yang and Poovaiah, 2002; Wang et al., 2016; Ismael et al., 2019).Cd进入植物根细胞也诱导一些对重金属胁迫有反应的特定信号分子, 如CaMs、钙依赖蛋白激酶(calcium-dependent protein kinases, CDPKs)和丝裂原活化蛋白激酶(mitogen-activated protein kinases, MAPKs), 调控植物对逆境的抗性(Danquah et al., 2014; Xu et al., 2017). ...
GmHMA3 sequesters Cd to the root endoplasmic reticulum to limit translocation to the stems in soybean 0 2018
Heat shock transcription factors: structure and regulation 1 1995
... HSP不仅能对抗应激相关损伤与保护细胞, 而且在蛋白质规则折叠、细胞内分布和降解中也发挥重要作用.这些功能有利于生物在压力条件下生存(Hartl and Hayer-Hart, 2002; Zhou et al., 2019).热休克转录因子HSFs (heat shock transcription factors)调节HSP的表达, 参与蛋白稳态的各个方面(如受损蛋白的重新折叠、组装和运输), 维持蛋白的稳定性(Wu, 1995; Guo et al., 2008).TaHsfA4a上调表达的水稻对Cd的耐受性增强, OsHsfA4a下调表达的水稻对Cd的耐受性减弱, 在小麦和水稻根系中, Cd胁迫导致HsfA4a以及金属硫蛋白基因的表达上调(Shim et al., 2009).此外, Chen等(2020)研究结果也表明, SaHsfA4c通过上调HSP的表达和提高ROS清除酶活性, 增强植物的抗逆性.Cd胁迫下, HSF/HSP参与柳枝稷(Panicum virgatum)正常蛋白质构象的重建以及细胞内稳态调节(Song et al., 2018).这些HSF/HSP表达基因可作为进一步研究的候选基因, 以提高植物对Cd的耐受性. ...
Potassium affects cadmium resistance in Arabidopsis through facilitating root cell wall Cd retention in a nitric oxide dependent manner 1 2020
... 活性氮(RNS)主要包括一氧化氮(NO)、S-亚硝基硫醇(SNOs)、高氮氧化物(NOx)、二硝基铁配合物和过氧亚硝酸盐(ONOO) 5种.过量的RNS会与脂类、蛋白质和核酸发生反应, 导致细胞损害(Neill et al., 2008a; Romero-Puertas and Sandalio, 2016).然而, 细胞内NO一旦产生, 就会与ROS迅速作用, 产生大量的RNS (如NO2), 然后降解为亚硝酸盐和硝酸钠(Neill et al., 2008b).低水平的NO可调节植物的发育, 而在胁迫条件下植物体内会产生大量的NO (Romero-Puertas and Sandalio, 2016; Liu et al., 2018); NO浓度过高则抑制植物的生长, 甚至引起细胞程序性死亡(Arasimowicz-Jelonek et al., 2012; Sami et al., 2018).低浓度NO促进植物生长, 高浓度则抑制植物生长, 其与生长素(indole-3-acetic acid, IAA)、细胞分裂素、赤霉素、脱落酸(abscisic acid, ABA)、乙烯、茉莉酸(jasmonic acid, JA)和水杨酸(salicylic acid, SA)也有一定的交互作用(Sami et al., 2018).Cd能通过破坏呼吸和光合作用的电子传递链使电子被传递到非靶向分子, 从而导致ROS和RNS的产生, 且Cd能通过抑制自由基降低相关酶活性(Ekmek?i et al., 2008; Romero-Puertas and Sandalio, 2016), 导致ROS和RNS不能够被及时清除而过量累积, 最终引起植物营养应激反应(Nieves-Cordones et al., 2019).外施氮(Zhang et al., 2019b)、磷(Wang et al., 2020)、钾(Wu et al., 2020)、钙(Li et al., 2021)和硅(Vaculík et al., 2015)等植物营养元素, 可起到调控植物激素含量、非酶物质和抗氧化酶活性及转运基因表达等作用, 减轻Cd对植物的毒害, 但外施营养物质对ROS和RNS有怎样的影响有待后续探索. ...
The zinc-regulated protein (ZIP) family genes and glutathione S-transferase (GST) family genes play roles in Cd resistance and accumulation of pak choi ( Brassica campestris ssp. chinensis) 2 2019
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
... ABC (ATP-binding cassette)转运体家族广泛参与细胞中离子、碳水化合物、脂类、药物和重金属的运输(Khan et al., 2020). 其中, 两种ABCC转运蛋白(AtABCC1和AtABCC2)是重要的液泡转运体, 通过液泡隔离介导对Cd的耐受性(Park et al., 2012).ZIP包括锌调控转运体(zinc-regulated transporter, ZRT)和铁调控转运体(iron-regulated transporter, IRT)家族.拟南芥中, 定位于根系细胞质膜的AtIRT1是主要的根部铁摄取载体, 其也可运输大量的Cd (Wu et al., 2019). ...
The ER luminal binding protein (BiP) alleviates Cd 2+-induced programmed cell death through endoplasmic reticulum stress- cell death signaling pathway in tobacco cells 2 2013
... 内质网(endoplasmic reticulum, ER)是一个复杂的代谢场所, 是调控蛋白质合成、信号转导和钙稳态的重要细胞器(Xu et al., 2013).所有分泌蛋白和大部分膜蛋白的合成和折叠及蛋白质的修饰和加工(主要包括糖基化、羟基化、酰基化和二硫键形成等)均在内质网进行(陈倩和谢旗, 2018).当植物受到干旱、高温、盐和重金属等胁迫后, 细胞蛋白质异常糖基化、Ca2+平衡紊乱及氧化还原状态改变会使细胞内积累大量未折叠或错误折叠的蛋白, 导致内质网胁迫(Kleizen and Braakman, 2004; Beaupere and Labunskyy, 2019).Cd诱导内质网应激, 改变内质网钙和脂质组成、ROS种类和错误折叠/未折叠蛋白(Rajakumar et al., 2016).分子伴侣参与内质网Cd胁迫的调控.BiP (binding protein)属于热休克蛋白(heat shock protein, HSP) HSP70家族, 是内质网腔内含量很高的分子伴侣, 参与内质网胁迫(Kleizen and Braakman, 2004).植物中, 程序性细胞死亡可通过ER调控应激-细胞死亡信号通路, BiP通过内质网应激-细胞死亡信号通路缓解Cd诱导的BY-2 (bright yellow-2)程序性死亡(Xu et al., 2013).同时, 内质网也是基因表达的重要场所.Wang等(2018)发现GmHMA3w主要在根内质网表达, 可将Cd从细胞壁运输至根内质网, 最终限制Cd从根向茎叶的转运.bZIP是位于根系质膜上的锌铁转运基因, 在拟南芥根细胞内质网应激信号转导中起重要作用(Deng et al., 2013). ...
... ).植物中, 程序性细胞死亡可通过ER调控应激-细胞死亡信号通路, BiP通过内质网应激-细胞死亡信号通路缓解Cd诱导的BY-2 (bright yellow-2)程序性死亡(Xu et al., 2013).同时, 内质网也是基因表达的重要场所.Wang等(2018)发现GmHMA3w主要在根内质网表达, 可将Cd从细胞壁运输至根内质网, 最终限制Cd从根向茎叶的转运.bZIP是位于根系质膜上的锌铁转运基因, 在拟南芥根细胞内质网应激信号转导中起重要作用(Deng et al., 2013). ...
Dissecting root proteome changes reveals new insight into cadmium stress response in radish ( Raphanus sativus L.) 4 2017
... Ca2+作为细胞功能的第二信使, 参与调节植物各种逆境下的重要生理生化过程.钙信使系统包括Ca2+、钙调蛋白(CaM)和钙泵(Ca2+-ATPase).CaM是Ca2+的感受器, Ca2+-ATPase决定着Ca2+的跨膜运输(Snedden and Fromm, 2001).正常情况下三者的含量保持稳定, 可维持细胞正常的生理功能.在外界生物和非生物胁迫下, 细胞质中的Ca2+浓度会瞬间升高(Ca2+浓度的波动被认为是植物对外界刺激的反应信号) (Nürnberger and Scheel, 2001; Liu et al., 2017).Cd胁迫扰动植物细胞中的Ca2+水平, 从而扰乱植物叶片的水分状况(Perfus-Barbeoch et al., 2002).胞质Ca2+浓度增加对H2O2的生成有动态调控作用, Ca2+可调控NADH氧化酶活性, 上调植物中H2O2的含量; 也可通过刺激H2O2酶活性下调植物中的H2O2水平(Yang and Poovaiah, 2002).当ROS含量超过Ca2+调控的浓度范围时, 会对植物造成伤害, 细胞膜和细胞器膜发生膜质过氧化, 膜的通透性被破坏, 致使细胞死亡(赵士诚等, 2008).Cd与Ca2+的化学性质相似, Cd被植物吸收后通过金属转运体或钙通道运输至根部, 也可通过电压依赖的钙通道进入细胞, 一旦进入胞质, 它就会模拟钙离子的活性, 扰乱钙信号调控; 导致根系吸收大量的Cd, 并显著减少根长、根表面积、根体积和根系生物量, 使叶片黄化并干燥坏死(Yang and Poovaiah, 2002; Wang et al., 2016; Ismael et al., 2019).Cd进入植物根细胞也诱导一些对重金属胁迫有反应的特定信号分子, 如CaMs、钙依赖蛋白激酶(calcium-dependent protein kinases, CDPKs)和丝裂原活化蛋白激酶(mitogen-activated protein kinases, MAPKs), 调控植物对逆境的抗性(Danquah et al., 2014; Xu et al., 2017). ...
... )的表达, 减轻Cd胁迫并重建萝卜体内氧化还原稳态(Xu et al., 2017; 2020). ...
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
... Cd显著影响抗氧化酶类物质和非酶抗氧化物质的基因表达.不同浓度Cd处理的玉米幼苗, SOD基因的表达随着Cd浓度的升高先上升, 后随着时间的延长而下降; CAT基因的表达随着Cd浓度的升高和处理时间的延长逐渐增强(赵士诚等, 2008).高浓度Cd处理下, 抗氧化酶基因GR-chl、CAT3、APX-cyt、APX-per和MTLa的表达上调(Zhang et al., 2019b).当植物遭受Cd胁迫时, 金属硫蛋白(metallothionein)基因MTs、APX、GPX和DHAR等表达上调(Cobbett and Goldsbrough, 2002; Tamás et al., 2008).γ-谷氨酰半胱氨酸合成酶(γ-glutamylcysteine synthetase, γ-GCS)、谷胱甘肽合成酶(glutathione synthetase, GSHs)、胱硫醚β-合酶(cystathionine beta-synthase, CBS)、ATP硫酸化酶(ATP sulfurylase, ATPS)、GR、PCS和乙二醛酶(glyoxalase) I和II等合成基因是调节重金属耐受性的潜在候选基因(Yadav, 2010; Sofoet al., 2013).细胞壁果胶(pectin)在重金属结合中起重要作用, 根、茎和叶细胞壁的酯化及果胶酶修饰减少了Cd的积累.番茄经Cd处理后, 其细胞壁果胶相关基因SlQUA1和SlPME1表达上调, 果胶甲基脂酶活性明显升高(Jia et al., 2019). ...
Preferential delivery of zinc to developing tissues in rice is mediated by P-type heavy metal ATPase OsHMA2 1 2013
... 重金属ATP酶基因家族(HMA)已被证明在不同的植物种类中行使特定的功能, 不同的HMA在不同组织中表达, 最终导致功能的多样性.HvHMA1参与Zn以及Cd向大麦(Hordeum vulgare)籽粒的转运(Mikkelsen et al., 2012).HMA2和HMA4可将Cd装载到木质部, 以便从根运输至茎和叶, OsHMA2与水稻维管组织中的锌装载及液泡膜定位有关(Yamaji et al., 2013).液泡膜上的OsHMA3将Cd转移至根系液泡, OsHMA4则将Cu转运至根系液泡, 限制Cd从根向茎以及叶的转移, 从而减少Cd在种子中的积累(Miyadate et al., 2011; Huang et al., 2016).TcHMA3在叶片的液泡中表达, 将Cd隔离在根和叶的液泡中, 最终导致Cd的超积累, 增强对Cd的耐受性(Ueno et al., 2011).AtHMA3在拟南芥中过表达导致Cd的积累量比野生型高2-3倍(Morel et al., 2009).这些转运蛋白为植物的基因工程提供了有用的工具, 增强了植物对金属的耐受性和积累能力, 继续推进它们的协同作用研究具有重要意义. ...
Hydrogen peroxide homeostasis: activation of plant catalase by calcium/calmodulin 2 2002
... Ca2+作为细胞功能的第二信使, 参与调节植物各种逆境下的重要生理生化过程.钙信使系统包括Ca2+、钙调蛋白(CaM)和钙泵(Ca2+-ATPase).CaM是Ca2+的感受器, Ca2+-ATPase决定着Ca2+的跨膜运输(Snedden and Fromm, 2001).正常情况下三者的含量保持稳定, 可维持细胞正常的生理功能.在外界生物和非生物胁迫下, 细胞质中的Ca2+浓度会瞬间升高(Ca2+浓度的波动被认为是植物对外界刺激的反应信号) (Nürnberger and Scheel, 2001; Liu et al., 2017).Cd胁迫扰动植物细胞中的Ca2+水平, 从而扰乱植物叶片的水分状况(Perfus-Barbeoch et al., 2002).胞质Ca2+浓度增加对H2O2的生成有动态调控作用, Ca2+可调控NADH氧化酶活性, 上调植物中H2O2的含量; 也可通过刺激H2O2酶活性下调植物中的H2O2水平(Yang and Poovaiah, 2002).当ROS含量超过Ca2+调控的浓度范围时, 会对植物造成伤害, 细胞膜和细胞器膜发生膜质过氧化, 膜的通透性被破坏, 致使细胞死亡(赵士诚等, 2008).Cd与Ca2+的化学性质相似, Cd被植物吸收后通过金属转运体或钙通道运输至根部, 也可通过电压依赖的钙通道进入细胞, 一旦进入胞质, 它就会模拟钙离子的活性, 扰乱钙信号调控; 导致根系吸收大量的Cd, 并显著减少根长、根表面积、根体积和根系生物量, 使叶片黄化并干燥坏死(Yang and Poovaiah, 2002; Wang et al., 2016; Ismael et al., 2019).Cd进入植物根细胞也诱导一些对重金属胁迫有反应的特定信号分子, 如CaMs、钙依赖蛋白激酶(calcium-dependent protein kinases, CDPKs)和丝裂原活化蛋白激酶(mitogen-activated protein kinases, MAPKs), 调控植物对逆境的抗性(Danquah et al., 2014; Xu et al., 2017). ...
... 的化学性质相似, Cd被植物吸收后通过金属转运体或钙通道运输至根部, 也可通过电压依赖的钙通道进入细胞, 一旦进入胞质, 它就会模拟钙离子的活性, 扰乱钙信号调控; 导致根系吸收大量的Cd, 并显著减少根长、根表面积、根体积和根系生物量, 使叶片黄化并干燥坏死(Yang and Poovaiah, 2002; Wang et al., 2016; Ismael et al., 2019).Cd进入植物根细胞也诱导一些对重金属胁迫有反应的特定信号分子, 如CaMs、钙依赖蛋白激酶(calcium-dependent protein kinases, CDPKs)和丝裂原活化蛋白激酶(mitogen-activated protein kinases, MAPKs), 调控植物对逆境的抗性(Danquah et al., 2014; Xu et al., 2017). ...
Molecular characterization of a rice metal tolerance protein, OsMTP1 2 2012
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
... 外源SA可改变ROS的积累和抗氧化酶系统活性, 缓解Cd胁迫带来的不利影响(Zawoznik et al., 2007).施加外源SA可使油菜(Brassica campestris)叶片的可溶性糖和游离脯氨酸含量升高、MDA积累减少, 质膜透性降低, CAT、POD和SOD活性升高, 明显缓解Cd毒害(胡春霞和王秀芹, 2010).SA处理显著增强玉米叶片中SOD活性, 降低CAT和POD活性, 并减轻Cd对APX活性的抑制作用(Krantev et al., 2008).此外, SA通过抑制ROS的过量产生缓解Cd引发的氧化损伤(Zawoznik et al., 2007); 施用外源SA可减少根系中ROS和MDA的积累, 进而减轻Cd毒性(Liu et al., 2016); 外源SA也可介导NADPH氧化酶产生H2O2, 以应对Cd胁迫引起的氧化应激(Chao et al., 2010).赵新月等(2013)研究表明, 与仅用Cd处理的玉米相比, 用50-250 μmol·L-1 SA处理玉米其叶绿素含量、硝酸还原酶(nitrite reductase, NR)、谷氨酰胺合成酶(glutamine synthetase, GS)和谷氨酸合成酶(glutamate synthase, GOGAT)活性显著升高, 氮素代谢过程良好, 玉米根系的POD活性降低(但POD和SOD活性与无Cd处理相比无显著差异), 且降低了Cd胁迫对质膜氧化系统的伤害.但外源SA对Cd胁迫修复的缓解作用受浓度影响较大, 低浓度的SA具有良好的修复效果, 高浓度的SA反而影响了光合色素的合成及酶活性, 抑制玉米的生长(赵新月等, 2013).重金属胁迫下, 外源SA提高了植物SOD、CAT和APX合成基因的表达(Gill, 2015; Song et al., 2019); 及水稻叶片和稻穗中低亲和离子转运子基因(OsLCT1)、水稻低Cd基因(OsLCD)和ZIP的时空表达(Liu et al., 2016). ...
... ).此外, SA通过抑制ROS的过量产生缓解Cd引发的氧化损伤(Zawoznik et al., 2007); 施用外源SA可减少根系中ROS和MDA的积累, 进而减轻Cd毒性(Liu et al., 2016); 外源SA也可介导NADPH氧化酶产生H2O2, 以应对Cd胁迫引起的氧化应激(Chao et al., 2010).赵新月等(2013)研究表明, 与仅用Cd处理的玉米相比, 用50-250 μmol·L-1 SA处理玉米其叶绿素含量、硝酸还原酶(nitrite reductase, NR)、谷氨酰胺合成酶(glutamine synthetase, GS)和谷氨酸合成酶(glutamate synthase, GOGAT)活性显著升高, 氮素代谢过程良好, 玉米根系的POD活性降低(但POD和SOD活性与无Cd处理相比无显著差异), 且降低了Cd胁迫对质膜氧化系统的伤害.但外源SA对Cd胁迫修复的缓解作用受浓度影响较大, 低浓度的SA具有良好的修复效果, 高浓度的SA反而影响了光合色素的合成及酶活性, 抑制玉米的生长(赵新月等, 2013).重金属胁迫下, 外源SA提高了植物SOD、CAT和APX合成基因的表达(Gill, 2015; Song et al., 2019); 及水稻叶片和稻穗中低亲和离子转运子基因(OsLCT1)、水稻低Cd基因(OsLCD)和ZIP的时空表达(Liu et al., 2016). ...
A novel plasma membrane-based NRAMP transporter contributes to Cd and Zn hyperac-cumulation in Sedum alfredii Hance 2 2020
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
The R2R3-MYB transcription factor MYB49 regulates cadmium accumulation 2 2019
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
Influence of nitrogen availability on Cd accumulation and acclimation strategy of Populus leaves under Cd exposure 3 2019
... 活性氮(RNS)主要包括一氧化氮(NO)、S-亚硝基硫醇(SNOs)、高氮氧化物(NOx)、二硝基铁配合物和过氧亚硝酸盐(ONOO) 5种.过量的RNS会与脂类、蛋白质和核酸发生反应, 导致细胞损害(Neill et al., 2008a; Romero-Puertas and Sandalio, 2016).然而, 细胞内NO一旦产生, 就会与ROS迅速作用, 产生大量的RNS (如NO2), 然后降解为亚硝酸盐和硝酸钠(Neill et al., 2008b).低水平的NO可调节植物的发育, 而在胁迫条件下植物体内会产生大量的NO (Romero-Puertas and Sandalio, 2016; Liu et al., 2018); NO浓度过高则抑制植物的生长, 甚至引起细胞程序性死亡(Arasimowicz-Jelonek et al., 2012; Sami et al., 2018).低浓度NO促进植物生长, 高浓度则抑制植物生长, 其与生长素(indole-3-acetic acid, IAA)、细胞分裂素、赤霉素、脱落酸(abscisic acid, ABA)、乙烯、茉莉酸(jasmonic acid, JA)和水杨酸(salicylic acid, SA)也有一定的交互作用(Sami et al., 2018).Cd能通过破坏呼吸和光合作用的电子传递链使电子被传递到非靶向分子, 从而导致ROS和RNS的产生, 且Cd能通过抑制自由基降低相关酶活性(Ekmek?i et al., 2008; Romero-Puertas and Sandalio, 2016), 导致ROS和RNS不能够被及时清除而过量累积, 最终引起植物营养应激反应(Nieves-Cordones et al., 2019).外施氮(Zhang et al., 2019b)、磷(Wang et al., 2020)、钾(Wu et al., 2020)、钙(Li et al., 2021)和硅(Vaculík et al., 2015)等植物营养元素, 可起到调控植物激素含量、非酶物质和抗氧化酶活性及转运基因表达等作用, 减轻Cd对植物的毒害, 但外施营养物质对ROS和RNS有怎样的影响有待后续探索. ...
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
... Cd显著影响抗氧化酶类物质和非酶抗氧化物质的基因表达.不同浓度Cd处理的玉米幼苗, SOD基因的表达随着Cd浓度的升高先上升, 后随着时间的延长而下降; CAT基因的表达随着Cd浓度的升高和处理时间的延长逐渐增强(赵士诚等, 2008).高浓度Cd处理下, 抗氧化酶基因GR-chl、CAT3、APX-cyt、APX-per和MTLa的表达上调(Zhang et al., 2019b).当植物遭受Cd胁迫时, 金属硫蛋白(metallothionein)基因MTs、APX、GPX和DHAR等表达上调(Cobbett and Goldsbrough, 2002; Tamás et al., 2008).γ-谷氨酰半胱氨酸合成酶(γ-glutamylcysteine synthetase, γ-GCS)、谷胱甘肽合成酶(glutathione synthetase, GSHs)、胱硫醚β-合酶(cystathionine beta-synthase, CBS)、ATP硫酸化酶(ATP sulfurylase, ATPS)、GR、PCS和乙二醛酶(glyoxalase) I和II等合成基因是调节重金属耐受性的潜在候选基因(Yadav, 2010; Sofoet al., 2013).细胞壁果胶(pectin)在重金属结合中起重要作用, 根、茎和叶细胞壁的酯化及果胶酶修饰减少了Cd的积累.番茄经Cd处理后, 其细胞壁果胶相关基因SlQUA1和SlPME1表达上调, 果胶甲基脂酶活性明显升高(Jia et al., 2019). ...
MhNRAMP1 from Malus hupehensis exacerbates cell death by accelerating Cd uptake in tobacco and apple calli 2 2020
... Responses of plant proteins to Cd stress Table 2
物质
主要因子
功能
参考文献
非酶物质
MT、PC、GSH和果胶
与Cd螯合, 固定在液泡或细胞壁中
Xu et al., 2017, 2020; Jia et al., 2019
抗氧化酶
SOD、CAT、APX、GPX、DHAR、GR、GSH、CBS和ATPS
减少ROS和RNS等物质的积累
Yadav, 2010; Zhang et al., 2019b
金属转运蛋白基因
LCT、CDF、CAX、NRAMP、ABC、ZIP、IRT、HMA和LCD
吸收或运输Cd离子, 提高植物的Cd积累和对Cd的耐受性
孙瑞莲和周启星, 2005; Yuan et al., 2012; Park et al., 2012; Mikkelsen et al., 2012; 张标金等, 2013; Huang et al., 2016; Feng et al., 2017; 袁连玉等, 2017; 黄新元和赵方杰, 2018; Wu et al., 2019; Zhang et al., 2020a, 2020b; Khan et al., 2020
转录因子调节蛋白基因
WRKY、MYB和HSF
与DNA结合, 参与激素和抗氧化酶等, 从而调控Cd的吸收、运输和积累
ülker and Somssich, 2004; Rushton et al., 2010; Dubos et al., 2010; Hong et al., 2017; Song et al., 2018; Zhang et al., 2019a; Cai et al., 2020; Chen et al., 2020
... Cd胁迫通常促进JA合成基因(Bashir et al., 2019)和抗氧化酶合成基因的转录(Keunen et al., 2013).低浓度JA可缓解Cd胁迫.例如, 用20 μmol·L-1 JA处理可有效改善Cd诱导的氧化应激, 使硫代巴比妥酸反应物质、H2O2和O2-.的积累减少, 使GSH、CAT、SOD及血红素氧合酶(HO)活性增强(Noriega et al., 2012; Adhikari et al., 2018).研究发现, 番茄(Solanum lycopersicum) JA缺失突变体对Cd的敏感性增强, 与光合作用相关参数(包括总叶绿素、光系统II实际效率、可变叶绿素荧光与最大荧光比及净光合速率)值降低; 相对水含量、可溶性糖和脯氨酸含量及淀粉的积累减少; 氧化应激和抗氧化防御作用(包括H2O2与MDA生成、电解质渗漏、SOD、APX和CAT活性)减弱(Zhao et al., 2016).外源茉莉酸甲酯(MeJA)通过降低拟南芥根系和茎叶中的Cd浓度, 下调参与Cd吸收和运输的AtIRT1、AtHMA2和AtHMA4基因表达, 缓解Cd引起的新叶毒害.JA合成关键基因AtAOS突变上调AtIRT1、AtHMA2和AtHMA4等转运蛋白基因表达, 增加根和芽中的Cd浓度, 提高拟南芥对Cd的敏感性.此外, MeJA降低Cd胁迫的拟南芥根尖中的NO水平(Lei et al., 2020).镉胁迫下植物激素ABA、IAA、SA和JA之间的相互作用值得深入探索. ...
Genome-wide identification, phylogenetic and expression analysis of the heat shock transcription factor family in bread wheat ( Triticum aestivum L.) 1 2019
... HSP不仅能对抗应激相关损伤与保护细胞, 而且在蛋白质规则折叠、细胞内分布和降解中也发挥重要作用.这些功能有利于生物在压力条件下生存(Hartl and Hayer-Hart, 2002; Zhou et al., 2019).热休克转录因子HSFs (heat shock transcription factors)调节HSP的表达, 参与蛋白稳态的各个方面(如受损蛋白的重新折叠、组装和运输), 维持蛋白的稳定性(Wu, 1995; Guo et al., 2008).TaHsfA4a上调表达的水稻对Cd的耐受性增强, OsHsfA4a下调表达的水稻对Cd的耐受性减弱, 在小麦和水稻根系中, Cd胁迫导致HsfA4a以及金属硫蛋白基因的表达上调(Shim et al., 2009).此外, Chen等(2020)研究结果也表明, SaHsfA4c通过上调HSP的表达和提高ROS清除酶活性, 增强植物的抗逆性.Cd胁迫下, HSF/HSP参与柳枝稷(Panicum virgatum)正常蛋白质构象的重建以及细胞内稳态调节(Song et al., 2018).这些HSF/HSP表达基因可作为进一步研究的候选基因, 以提高植物对Cd的耐受性. ...
 ,1,2,4,*
,1,2,4,*
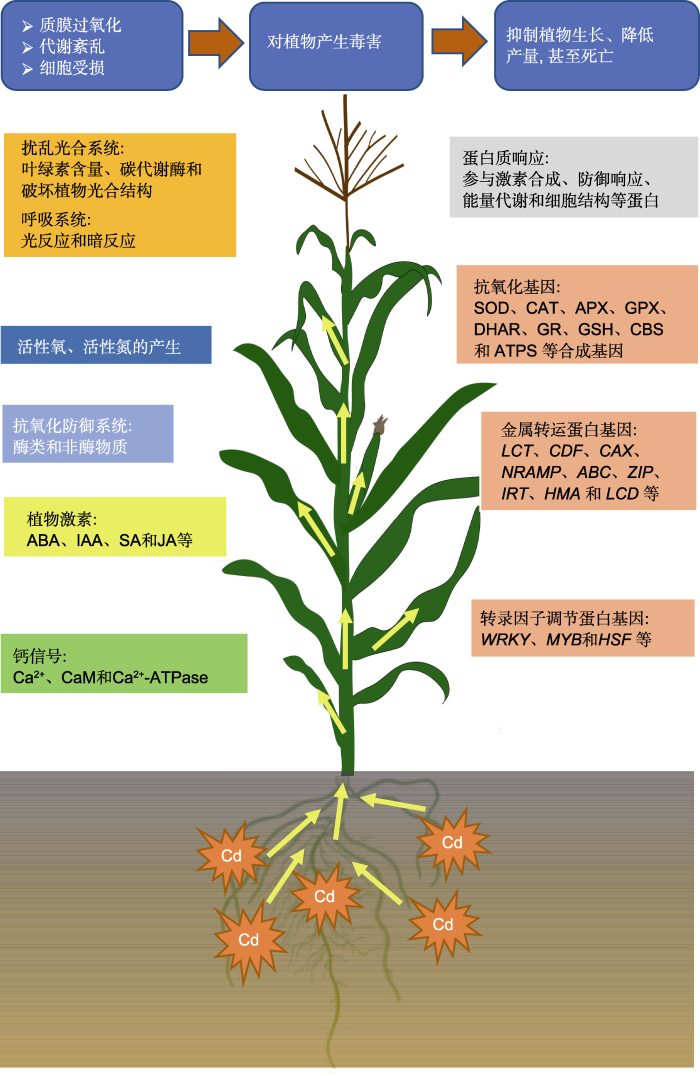 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}