Binbin Hu1,2, Zhihui Xue1, Cui Zhang,1,2,*1Key Laboratory of Plant Molecular Physiology, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 2University of Chinese Academy of Sciences, Beijing 100049, China
Abstract Small RNAs are a type of small nucleotide molecules that are essential for plant growth and development, playing a key role in variety of life processes and in response to stresses. Research on the location of small RNAs could discover their functions in plants. Small RNA FISH is a qualitative or semi-quantitative analysis of small RNA in organisms by fluorescence detection technology. At present, this technology has been widely used in animals, but it is still less applied in plants. This article introduces the specific operation procedures and attentions based on ultra-high resolution microscopy that combines locked nucleic acid (LNA) probe in situ hybridization with immunofluorescence. This protocol can be used to detect the expression and localization of small RNA in plant tissues. Keywords:fluorescent in situ hybridization;small RNA;plants
PDF (1549KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 胡滨滨, 薛治慧, 张翠. 植物小RNA荧光原位杂交实验方法. 植物学报, 2021, 56(3): 330-338 doi:10.11983/CBB21057 Hu Binbin, Xue Zhihui, Zhang Cui. Protocols for Small RNA FISH in Plants. Chinese Bulletin of Botany, 2021, 56(3): 330-338 doi:10.11983/CBB21057
小RNA是动植物中一类非常重要的18-30 nt小分子核苷酸(Chen, 2009; Kim et al., 2009), 成熟的小RNA通过对靶基因进行转录或转录后水平的调节参与多种植物的生长发育与胁迫响应过程。目前植物中主要有3种小RNA,分别为miRNA (microRNA)、siRNA (Small interfering RNA)和phasiRNA (phased secondary small interfering RNA) (Yu et al., 2019)。在植物细胞中, miRNA的生物合成过程已有报道, miRNA编码基因在RNA聚合酶II的作用下被转录至细胞核中, 形成初级miRNA (Primary miRNA, Pri- miRNA) (Lee et al., 2004; Jones-Rhoades et al., 2006)。Pri-miRNA在DCL1 (DICER-LIKE 1)、HYL1 (HYPONASTIC LEAVES 1)以及SE (SERRATE)等蛋白因子的作用下被切割成具有互补茎环结构的前体miRNA (Precursor miRNA, Pre-miRNA), Pre- miRNA又被加工成miRNA/miRNA*双链体。在HEN1 (HUA ENHANCER 1)的作用下, miRNA/miRNA*双链聚合体的3'端被甲基化修饰, 以免被降解, 稳定的miRNA/miRNA*双链聚合体在HASTY蛋白的作用下被运输至细胞质中(Kurihara and Watanabe, 2004; Li et al., 2005; Park et al., 2005; Jones-Rhoades et al., 2006)。在细胞质中, miRNA/miRNA*双链聚合物
BaumbergerN, BaulcombeDC (2005). Arabidopsis ARGONAUTE1 is an RNA slicer that selectively recruits microRNAs and short interfering RNAs Proc Natl Acad Sci USA 102, 11928-11933. DOI:10.1073/pnas.0505461102URL [本文引用: 1]
BhogaleS, MahajanAS, NatarajanB, RajabhojM, ThulasiramHV, BanerjeeAK (2014). MicroRNA156: a potential graft-transmissible microRNA that modulates plant architecture and tuberization in Solanum tuberosum ssp. andigena Plant Physiol 164, 1011-1027. DOI:10.1104/pp.113.230714URL [本文引用: 1]
BleckmannA, DresselhausT (2016). Fluorescent whole- mount RNA in situ hybridization (F-WISH) in plant germ cells and the fertilized ovule Methods 98, 66-73. DOI:S1046-2023(15)30137-7PMID:26521978 [本文引用: 1] First evidence on gene function and regulation is provided by the cellular expression pattern in complex tissues. However, to understand the activity of a specific gene, it is essential to analyze the regulatory network, which controls the spatio-temporal translation pattern during the entire life span of the transcribed mRNA. To explore mechanisms which control mRNA abundance and localization in space and time, it is necessary to visualize mRNAs quantitatively with a subcellular resolution, without sectioning the tissues. We have adapted and optimized a protocol for colorimetric whole-mount RNA in situ hybridization (WISH) using egg cell-specific digoxigenin (DIG) labeled probes (Hejátko et al., 2006) [1] on ovules and early seeds of Arabidopsis. Furthermore, we established a fluorescent whole-mount RNA in situ hybridization (F-WISH) protocol, which allows mRNA visualization on a subcellular level. The polar localized mRNA of SBT4.13, encoding a subtilase, was identified using this protocol. Both methods are described and discussed in detail. Additionally a (F)-WISH flow-chart is provided along with a troubleshooting table. Copyright ? 2015 Elsevier Inc. All rights reserved.
BrunoL, MutoA, SpadaforaND, IariaD, ChiappettaA, Van LijsebettensM, BitontiMB (2011). Multi-probe in situ hybridization to whole mount Arabidopsis seedlings Int J Dev Biol 55, 197-203. DOI:10.1387/ijdb.103132lbURL [本文引用: 1]
BrunoL, RonchiniM, GagliardiO, CorintiT, ChiappettaA, GerolaP, BitontiMB (2015). Analysis of ATGUS1 and ATGUS2 in Arabidopsis root apex by a highly sensitive TSA-MISH method Int J Dev Biol 59, 221-228. DOI:10.1387/ijdb.140195LBURL [本文引用: 1]
BuhtzA, SpringerF, ChappellL, BaulcombeDC, KehrJ (2008). Identification and characterization of small RNAs from the phloem of Brassica napus Plant J 53, 739-749. DOI:10.1111/tpj.2008.53.issue-5URL [本文引用: 1]
CarlsbeckerA, LeeJY, RobertsCJ, DettmerJ, LehesrantaS, ZhouJ, LindgrenO, Moreno-RisuenoMA, VaténA, ThitamadeeS, CampilhoA, SebastianJ, BowmanJL, HelariuttaY, BenfeyPN (2010). Cell signaling by microRNA165/6 directs gene dose-dependent root cell fate Nature 465, 316-321. DOI:10.1038/nature08977PMID:20410882 [本文引用: 1] A key question in developmental biology is how cells exchange positional information for proper patterning during organ development. In plant roots the radial tissue organization is highly conserved with a central vascular cylinder in which two water conducting cell types, protoxylem and metaxylem, are patterned centripetally. We show that this patterning occurs through crosstalk between the vascular cylinder and the surrounding endodermis mediated by cell-to-cell movement of a transcription factor in one direction and microRNAs in the other. SHORT ROOT, produced in the vascular cylinder, moves into the endodermis to activate SCARECROW. Together these transcription factors activate MIR165a and MIR166b. Endodermally produced microRNA165/6 then acts to degrade its target mRNAs encoding class III homeodomain-leucine zipper transcription factors in the endodermis and stele periphery. The resulting differential distribution of target mRNA in the vascular cylinder determines xylem cell types in a dosage-dependent manner.
ChenXM (2004). A microRNA as a translational repressor of APETALA2 in Arabidopsis flower development Science 303, 2022-2025. DOI:10.1126/science.1088060URL [本文引用: 1]
HimanenK, WoloszynskaM, BoccardiTM, De GroeveS, NelissenH, BrunoL, VuylstekeM, Van LijsebettensM (2012). Histone H2b monoubiquitination is required to reach maximal transcript levels of circadian clock genes in Arabidopsis Plant J 72, 249-260. DOI:10.1111/j.1365-313X.2012.05071.xURL [本文引用: 1]
HowellMD, FahlgrenN, ChapmanEJ, CumbieJS, SullivanCM, GivanSA, KasschauKD, CarringtonJC (2007). Genome-wide analysis of the RNA-DEPENDENT RNA POLYMERASE6/DICER-LIKE4 pathway in Arabidop- sis reveals dependency on miRNA- and tasiRNA-directed targeting Plant Cell 19, 926-942. PMID:17400893 [本文引用: 1] Posttranscriptional RNA silencing of many endogenous transcripts, viruses, and transgenes involves the RNA-DEPENDENT RNA POLYMERASE6/DICER-LIKE4 (RDR6/DCL4)-dependent short interfering RNA (siRNA) biogenesis pathway. Arabidopsis thaliana contains several families of trans-acting siRNAs (tasiRNAs) that form in 21-nucleotide phased arrays through the RDR6/DCL4-dependent pathway and that negatively regulate target transcripts. Using deep sequencing technology and computational approaches, the phasing patterns of known tasiRNAs and tasiRNA-like loci from across the Arabidopsis genome were analyzed in wild-type plants and silencing-defective mutants. Several gene transcripts were found to be routed through the RDR6/DCL4-dependent pathway after initial targeting by one or multiple miRNAs or tasiRNAs, the most conspicuous example of which was an expanding clade of genes encoding pentatricopeptide repeat (PPR) proteins. Interestingly, phylogenetic analysis using Populus trichocarpa revealed evidence for small RNA-mediated regulatory mechanisms within a similarly expanded group of PPR genes. We suggest that posttranscriptional silencing mechanisms operate on an evolutionary scale to buffer the effects of rapidly expanding gene families.
HuangK, BaldrichP, MeyersBC, CaplanJL (2019). sRNA-FISH: versatile fluorescent in situ detection of small RNAs in plants Plant J 98, 359-369. DOI:10.1111/tpj.14210 [本文引用: 1] Localization of mRNA and small RNAs (sRNAs) is important for understanding their function. Fluorescent in situ hybridization (FISH) has been used extensively in animal systems to study the localization and expression of sRNAs. However, current methods for fluorescent in situ detection of sRNA in plant tissues are less developed. Here we report a protocol (sRNA-FISH) for efficient fluorescent detection of sRNAs in plants. This protocol is suitable for application in diverse plant species and tissue types. The use of locked nucleic acid probes and antibodies conjugated with different fluorophores allows the detection of two sRNAs in the same sample. Using this method, we have successfully detected the co-localization of miR2275 and a 24-nucleotide phased small interfering RNA in maize anther tapetal and archesporial cells. We describe how to overcome the common problem of the wide range of autofluorescence in embedded plant tissue using linear spectral unmixing on a laser scanning confocal microscope. For highly autofluorescent samples, we show that multi-photon fluorescence excitation microscopy can be used to separate the target sRNA-FISH signal from background autofluorescence. In contrast to colorimetric in situ hybridization, sRNA-FISH signals can be imaged using super-resolution microscopy to examine the subcellular localization of sRNAs. We detected maize miR2275 by super-resolution structured illumination microscopy and direct stochastic optical reconstruction microscopy. In this study, we describe how we overcame the challenges of adapting FISH for imaging in plant tissue and provide a step-by-step sRNA-FISH protocol for studying sRNAs at the cellular and even subcellular level.
HuenAK, Rodriguez-MedinaC, HoAYY, AtkinsCA, SmithPMC (2017). Long-distance movement of phospha- te starvation-responsive microRNAs in Arabidopsis Plant Biol 19, 643-649. DOI:10.1111/plb.2017.19.issue-4URL [本文引用: 1]
JavelleM, TimmermansMCP (2012). In situ localization of small RNAs in plants by using LNA probes Nat Protoc 7, 533-541. DOI:10.1038/nprot.2012.006PMID:22362159 [本文引用: 1] Small RNAs have crucial roles in numerous aspects of plant biology. Despite our current understanding of their biogenesis and mechanisms of action, the biological function of small RNAs, particularly miRNAs, remains largely unknown. To decipher small RNA function, knowledge about their spatiotemporal patterns of expression is essential. Here we report an in situ hybridization method for the precise localization of small RNAs in plants by using locked nucleic acid (LNA) oligonucleotide probes. This method has been adapted from protocols used to detect messenger RNAs in formaldehyde-fixed and paraffin-embedded tissue sections, but it includes essential optimizations in key prehybridization, hybridization and posthybridization steps. Most importantly, optimization of probe concentration and hybridization temperature is required for each unique LNA probe. We present the detailed protocol starting from sectioned tissues, and we include troubleshooting tips and recommended controls. This method has been used successfully in several plant species and can be completed within 2-6 d.
JiangJM (2019). Fluorescence in situ hybridization in plants: recent developments and future applications Chromoso- me Res 27, 153-165. [本文引用: 1]
Jones-RhoadesMW, BartelDP, BartelB (2006). Micro- RNAs and their regulatory roles in plants Annu Rev Plant Biol 57, 19-53. PMID:16669754 [本文引用: 2] MicroRNAs (miRNAs) are small, endogenous RNAs that regulate gene expression in plants and animals. In plants, these approximately 21-nucleotide RNAs are processed from stem-loop regions of long primary transcripts by a Dicer-like enzyme and are loaded into silencing complexes, where they generally direct cleavage of complementary mRNAs. Although plant miRNAs have some conserved functions extending beyond development, the importance of miRNA-directed gene regulation during plant development is now particularly clear. Identified in plants less than four years ago, miRNAs are already known to play numerous crucial roles at each major stage of development-typically at the cores of gene regulatory networks, targeting genes that are themselves regulators, such as those encoding transcription factors and F-box proteins.
JungJH, ParkCM (2007). MIR166/ 165 genes exhibit dynamic expression patterns in regulating shoot apical meris- tem and floral development in Arabidopsis Planta 225, 1327-1338. DOI:10.1007/s00425-006-0439-1URL [本文引用: 1]
KidnerC, TimmermansM (2006). In situ hybridization as a tool to study the role of microRNAs in plant development Methods Mol Biol 342, 159-179. [本文引用: 1]
KimVN, HanJJ, SiomiMC (2009). Biogenesis of small RNAs in animals Nat Rev Mol Cell Biol 10, 126-139. [本文引用: 1]
KnauerS, HoltAL, Rubio-SomozaI, TuckerEJ, HinzeA, PischM, JavelleM, TimmermansMC, TuckerMR, LauxT (2013). A protodermal miR394 signal defines a region of stem cell competence in the Arabidopsis shoot meristem Dev Cell 24, 125-132. DOI:10.1016/j.devcel.2012.12.009URL [本文引用: 1]
KuriharaY, WatanabeY (2004). Arabidopsis micro-RNA biogenesis through dicer-like 1 protein functions Proc Natl Acad Sci USA 101, 12753-12758. DOI:10.1073/pnas.0403115101URL [本文引用: 1]
LambJC, DanilovaT, BauerMJ, MeyerJM, HollandJJ, JensenMD, BirchlerJA (2007). Single-gene detection and karyotyping using small-target fluorescence in situ hybridization on maize somatic chromosomes Genetics 175, 1047-1058. DOI:10.1534/genetics.106.065573URL [本文引用: 1]
LeeY, KimM, HanJJ, YeomKH, LeeS, BaekSH, KimVN (2004). MicroRNA genes are transcribed by RNA polymerase II EMBO J 23, 4051-4060. DOI:10.1038/sj.emboj.7600385URL [本文引用: 1]
LevskyJM, SingerRH (2003). Fluorescence in situ hybridization: past, present and future J Cell Sci 116, 2833-2838. DOI:10.1242/jcs.00633URL [本文引用: 1]
LiJJ, YangZY, YuB, LiuJ, ChenXM (2005). Methylation protects miRNAs and siRNAs from a 3'-end uridylation activity in Arabidopsis Curr Biol 15, 1501-1507. DOI:10.1016/j.cub.2005.07.029URL [本文引用: 1]
LiS, WangXT, XuWY, LiuT, CaiCM, ChenLY, ClarkCB, MaJX (2021). Unidirectional movement of small RNAs from shoots to roots in interspecific heterografts Nat Plants 7, 50-59. DOI:10.1038/s41477-020-00829-2URL [本文引用: 1]
LinSI, ChiangSF, LinWY, ChenJW, TsengCY, WuPC, ChiouTJ (2008). Regulatory network of microRNA399 and PHO2 by systemic signaling Plant Physiol 147, 732-746. DOI:10.1104/pp.108.116269URL [本文引用: 1]
LiuQL, YaoXZ, PiLM, WangH, CuiXF, HuangH (2009). The ARGONAUTE10 gene modulates shoot apical meris- tem maintenance and establishment of leaf polarity by repressing miR165/166 in Arabidopsis Plant J 58, 27-40. DOI:10.1111/j.1365-313X.2008.03757.xURL [本文引用: 1]
LuJ, TsourkasA (2009). Imaging individual microRNAs in single mammalian cells in situ Nucleic Acids Res 37, e100. DOI:10.1093/nar/gkp482URL [本文引用: 1]
MoissiardG, ParizottoEA, HimberC, VoinnetO (2007). Transitivity in Arabidopsis can be primed, requires the redundant action of the antiviral Dicer-like 4 and Dicer-like 2, and is compromised by viral-encoded suppressor proteins RNA 13, 1268-1278. DOI:10.1261/rna.541307URL [本文引用: 1]
NodineMD, BartelDP (2010). MicroRNAs prevent precocious gene expression and enable pattern formation during plant embryogenesis Gene Dev 24, 2678-2692. DOI:10.1101/gad.1986710URL [本文引用: 1]
NogueiraFTS, ChitwoodDH, MadiS, OhtsuK, SchnablePS, ScanlonMJ, TimmermansMCP (2009). Regulation of small RNA accumulation in the maize shoot apex PLoS Genet 5, e1000320. DOI:10.1371/journal.pgen.1000320URL [本文引用: 1]
NogueiraFTS, MadiS, ChitwoodDH, JuarezMT, Timmer- mansMCP (2007). Two small regulatory RNAs establish opposing fates of a developmental axis Gene Dev 21, 750-755. PMID:17403777 [本文引用: 1] Small RNAs are important regulators of gene expression. In maize, adaxial/abaxial (dorsoventral) leaf polarity is established by an abaxial gradient of microRNA166 (miR166), which spatially restricts the expression domain of class III homeodomain leucine zipper (HD-ZIPIII) transcription factors that specify adaxial/upper fate. Here, we show that leafbladeless1 encodes a key component in the trans-acting small interfering RNA (ta-siRNA) biogenesis pathway that acts on the adaxial side of developing leaves and demarcates the domains of hd-zipIII and miR166 accumulation. Our findings indicate that tasiR-ARF, a ta-siRNA, and miR166 establish opposing domains along the adaxial-abaxial axis, thus revealing a novel mechanism of pattern formation.
OriN, CohenAR, EtzioniA, BrandA, YanaiO, ShleizerS, MendaN, AmsellemZ, EfroniI, PekkerI, AlvarezJP, BlumE, ZamirD, EshedY (2007). Regulation of LANCEOLATE by miR319 is required for compound-leaf development in tomato Nat Genet 39, 787-791. DOI:10.1038/ng2036URL [本文引用: 1]
PagliaraniC, GambinoG (2019). Small RNA mobility: spread of RNA silencing effectors and its effect on developmental processes and stress adaptation in plants Int J Mol Sci 20, 4306. DOI:10.3390/ijms20174306URL [本文引用: 1]
PantBD, BuhtzA, KehrJ, ScheibleWR (2008). MicroRNA399 is a long-distance signal for the regulation of plant phosphate homeostasis Plant J 53, 731-738. DOI:10.1111/tpj.2008.53.issue-5URL [本文引用: 1]
PantBD, Musialak-LangeM, NucP, MayP, BuhtzA, KehrJ, WaltherD, ScheibleWR (2009). Identification of nutrient-responsive Arabidopsis and rapeseed microRNAs by comprehensive real-time polymerase chain reaction profiling and small RNA sequencing Plant Physiol 150, 1541-1555. DOI:10.1104/pp.109.139139URL [本文引用: 1]
ParizottoEA, DunoyerP, RahmN, HimberC, VoinnetO (2004). In vivo investigation of the transcription, processing, endonucleolytic activity, and functional relevance of the spatial distribution of a plant miRNA Gene Dev 18, 2237-2242. PMID:15371337 [本文引用: 1] We show, with miR171, that plant miRNA genes are modular independent transcription units in which the fold-back pre-miRNA is sufficient for miRNA processing, and that the upstream region contains highly specific promoter elements. Processing depends on flanking sequences within the miRNA stem-loop precursor rather than the miRNA sequence itself, and mutations affecting target pairing at the center and 5' but not 3' region of the miRNA compromise its function in vivo. Inactivation of the SDE1 RNA-dependent-RNA-polymerase was mandatory for accurate representation of miRNA activity by sensor constructs in Arabidopsis. Work in sde1 background revealed a near-perfect spatial overlap between the patterns of miR171 transcription and activity, supporting the idea that plant miRNAs enable cell differentiation.
ParkMY, WuG, Gonzalez-SulserA, VaucheretH, PoethigRS (2005). Nuclear processing and export of microRNAs in Arabidopsis Proc Natl Acad Sci USA 102, 3691-3696. DOI:10.1073/pnas.0405570102URL [本文引用: 1]
RamanS, GrebT, PeaucelleA, BleinT, LaufsP, TheresK (2008). Interplay of miR164, CUP-SHAPED COTYLEDON genes and LATERAL SUPPRESSOR controls axillary meristem formation in Arabidopsis thaliana Plant J 55, 65-76. DOI:10.1111/tpj.2008.55.issue-1URL [本文引用: 1]
RozierF, MirabetV, VernouxT, DasP (2014). Analysis of 3D gene expression patterns in plants using whole-mount RNA in situ hybridization Nat Protoc 9, 2464-2475. DOI:10.1038/nprot.2014.162URL [本文引用: 1]
SkopelitisDS, HillK, KlesenS, MarcoCF, von BornP, ChitwoodDH, TimmermansMCP (2018). Gating of miRNA movement at defined cell-cell interfaces governs their impact as positional signals Nat Commun 9, 3107. DOI:10.1038/s41467-018-05571-0PMID:30082703 [本文引用: 1] Mobile small RNAs serve as local positional signals in development and coordinate stress responses across the plant. Despite its central importance, an understanding of how the cell-to-cell movement of small RNAs is governed is lacking. Here, we show that miRNA mobility is precisely regulated through a gating mechanism polarised at defined cell-cell interfaces. This generates directional movement between neighbouring cells that limits long-distance shoot-to-root trafficking, and underpins domain-autonomous behaviours of small RNAs within stem cell niches. We further show that the gating of miRNA mobility occurs independent of mechanisms controlling protein movement, identifying the small RNA as the mobile unit. These findings reveal gate-keepers of cell-to-cell small RNA mobility generate selectivity in long-distance signalling, and help safeguard functional domains within dynamic stem cell niches while mitigating a 'signalling gridlock' in contexts where developmental patterning events occur in close spatial and temporal vicinity.
TirichineL, AndreyP, BiotE, MaurinY, GaudinV (2009). 3D fluorescent in situ hybridization using Arabidopsis leaf cryosections and isolated nuclei Plant Methods 5, 11. DOI:10.1186/1746-4811-5-11PMID:19650905 [本文引用: 1] Fluorescent hybridization techniques are widely used to study the functional organization of different compartments within the mammalian nucleus. However, few examples of such studies are known in the plant kingdom. Indeed, preservation of nuclei 3D structure, which is required for nuclear organization studies, is difficult to fulfill.We report a rapid protocol for fluorescent in situ hybridization (FISH) performed on 3D isolated nuclei and thin cryosectioned leaves of Arabidopsis thaliana. The use of direct labeling minimized treatment steps, shortening the overall procedure. Using image analysis, we measured different parameters related to nucleus morphology and overall 3D structure.Our work describes a 3D-FISH protocol that preserves the 3D structure of Arabidopsis interphase nuclei. Moreover, we report for the first time FISH using cryosections of Arabidopsis leaves. This protocol is a valuable tool to investigate nuclear architecture and chromatin organization.
VaucheretH, VazquezF, CrétéP, BartelDP (2004). The action of ARGONAUTE1 in the miRNA pathway and its regulation by the miRNA pathway are crucial for plant development Gene Dev 18, 1187-1197. PMID:15131082 [本文引用: 1] MicroRNAs (miRNAs) are endogenous 21-24-nt RNAs that can down-regulate gene expression by pairing to the messages of protein-coding genes to specify mRNA cleavage or repression of productive translation. They act within the RNA-induced silencing complex (RISC), which in animals contains a member of the Argonaute family of proteins. In the present study, we show that Arabidopsis ago1 mutants have increased accumulation of mRNAs known to be targeted for cleavage by miRNAs. In hypomorphic ago1 alleles, this compromised miRNA function occurs without a substantial change in miRNA accumulation, whereas in null alleles it is accompanied by a drop in some of the miRNAs. Therefore, AGO1 acts within the Arabidopsis miRNA pathway, probably within the miRNA-programmed RISC, such that the absence of AGO1 destabilizes some of the miRNAs. We also show that targeting of AGO1 mRNA by miR168 is needed for proper plant development, illustrating the importance of feedback control by this miRNA. Transgenic plants expressing a mutant AGO1 mRNA with decreased complementarity to miR168 overaccumulate AGO1 mRNA and exhibit developmental defects partially overlapping with those of dcl1, hen1, and hyl1 mutants showing a decrease in miRNA accumulation. miRNA targets overaccumulate in miR168-resistant plants, suggesting that a large excess of AGO1 protein interferes with the function of RISC or sequesters miRNAs or other RISC components. Developmental defects induced by a miR168-resistant AGO1 mRNA can be rescued by a compensatory miRNA that is complementary to the mutant AGO1 mRNA, proving the regulatory relationship between miR168 and its target and opening the way for engineering artificial miRNAs in plants.
WollmannH, MicaE, TodescoM, LongJA, WeigelD (2010). On reconciling the interactions between APETA- LA2, miR172 and AGAMOUS with the ABC model of flower development Development 137, 3633-3642. DOI:10.1242/dev.036673PMID:20876650 [本文引用: 1] The ABC model of flower development explains how three classes of homeotic genes confer identity to the four types of floral organs. In Arabidopsis thaliana, APETALA2 (AP2) and AGAMOUS (AG) represent A- and C-class genes that act in an antagonistic fashion to specify perianth and reproductive organs, respectively. An apparent paradox was the finding that AP2 mRNA is supposedly uniformly distributed throughout young floral primordia. Although miR172 has a role in preventing AP2 protein accumulation, miR172 was reported to disappear from the periphery only several days after AG activation in the center of the flower. Here, we resolve the enigmatic behavior of AP2 and its negative regulator miR172 through careful expression analyses. We find that AP2 mRNA accumulates predominantly in the outer floral whorls, as expected for an A-class homeotic gene. Its pattern overlaps only transiently with that of miR172, which we find to be restricted to the center of young floral primordia from early stages on. MiR172 also accumulates in the shoot meristem upon floral induction, compatible with its known role in regulating AP2-related genes with a role in flowering. Furthermore, we show that AP2 can cause striking organ proliferation defects that are not limited to the center of the floral meristem, where its antagonist AG is required for terminating stem cell proliferation. Moreover, AP2 never expands uniformly into the center of ag mutant flowers, while miR172 is largely unaffected by loss of AG activity. We present a model in which the decision whether stamens or petals develop is based on the balance between AP2 and AG activities, rather than the two being mutually exclusive.
WoloszynskaM, Le GallS, NeytP, BoccardiTM, GrasserM, L?ngstG, AesaertS, CoussensG, DhondtS, Van De SlijkeE, BrunoL, Fung-UcedaJ, MasP, Van MontaguM, InzéD, HimanenK, De JaegerG, GrasserKD, Van LijsebettensM (2019). Histone 2B monoubiquitination complex integrates transcript elongation with RNA processing at circadian clock and flowering regulators Proc Natl Acad Sci USA 116, 8060-8069. DOI:10.1073/pnas.1806541116URL [本文引用: 1]
XueZH, LiuLY, ZhangC (2020). Regulation of shoot apical meristem and axillary meristem development in plants Int J Mol Sci 21, 2917. DOI:10.3390/ijms21082917URL [本文引用: 1]
YangWB, SchusterC, PrunetN, DongQK, LandreinB, WightmanR, MeyerowitzEM (2020). Visualization of protein coding, long noncoding, and nuclear RNAs by fluorescence in situ hybridization in sections of shoot apical meristems and developing flowers Plant Physiol 182, 147-158. DOI:10.1104/pp.19.00980URL [本文引用: 1]
YangWB, WightmanR, MeyerowitzEM (2017). Cell cycle control by nuclear sequestration of CDC20 and CDH1 mRNA in plant stem cells Mol Cell 68, 1108-1119. DOI:10.1016/j.molcel.2017.11.008URL [本文引用: 1]
YuY, ZhangYC, ChenXM, ChenYQ (2019). Plant noncoding RNAs: hidden players in development and stress responses Annu Rev Cell Dev Biol 35, 407-431. DOI:10.1146/annurev-cellbio-100818-125218 [本文引用: 2] A large and significant portion of eukaryotic transcriptomes consists of non-coding RNAs (ncRNAs) that have minimal or no protein-coding capacity but are functional. Diverse ncRNAs, including both small RNAs and long ncRNAs (lncRNAs), play essential regulatory roles in almost all biological processes by modulating gene expression at the transcriptional and post-transcriptional levels. In this review, we summarize the current knowledge of plant small RNAs and lncRNAs, with a focus on their biogenesis, modes of action, local and systemic movement, and functions at the nexus of plant development and environmental responses. The complex connections among small RNAs, lncRNAs, and small peptides in plants are also discussed, along with the challenges of identifying and investigating new classes of ncRNAs.
ZhangC, FanLS, LeBH, YePY, MoBX, ChenXM (2020). Regulation of ARGONAUTE10 expression enables temporal and spatial precision in axillary meristem initiation in Arabidopsis Dev Cell 55, 603-616. DOI:10.1016/j.devcel.2020.10.019URL [本文引用: 1]
ZhangC, WangJ, WenkelS, ChandlerJW, WerrW, JiaoYL (2018). Spatiotemporal control of axillary meristem formation by interacting transcriptional regulators Development 145, dev158352. [本文引用: 1]
Arabidopsis ARGONAUTE1 is an RNA slicer that selectively recruits microRNAs and short interfering RNAs 1 2005
... 解旋, 反义链被降解, 成熟miRNA单链与含有RNA解旋酶和AGO (ARGONAUTE)蛋白的RNA诱导沉默复合体(RNA-induced silencing complex, RISC)结合, 控制其相关靶基因的表达(Vaucheret et al., 2004; Baumberger and Baulcombe, 2005). ...
MicroRNA156: a potential graft-transmissible microRNA that modulates plant architecture and tuberization in Solanum tuberosum ssp. andigena 1 2014
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
Fluorescent whole- mount RNA in situ hybridization (F-WISH) in plant germ cells and the fertilized ovule 1 2016
... 基于RNA FISH改进的LNA探针原位杂交技术与免疫荧光相结合的小RNA荧光原位杂交(small RNA fluorescent in situ hybridization, sRNA-FISH)实验技术能够很好地解决上述问题.RNA-FISH属于荧光原位杂交技术的一种, 用于检测和定位细胞中特定的RNA分子.该方法使用与靶标RNA特异性结合的RNA分子作为探针, 探针上带有半抗原或者荧光素, 利用碱基互补配对原则, 可实现间接或者直接的荧光显色.RNA FISH最初被用于检测动物以及酵母(Saccharomyces spp.)中RNA的表达模式与细胞定位(Levsky and Singer, 2003).而在植物中, FISH技术最先被用于染色体以及染色体中特定基因座位的成像(Lamb et al., 2007; Tirichine et al., 2009; Jiang, 2019), 后来又被研究人员用于指示mRNA在植物体内的组织成像以及亚细胞定位(Bruno et al., 2011, 2015; Himanen et al., 2012; Rozier et al., 2014; Bleckmann and Dresselhaus, 2016; Yang et al., 2017, 2020; Woloszynska et al., 2019).针对小RNA的FISH技术也已广泛应用于动物系统中(Lu and Tsourkas, 2009), 但在植物中的应用还比较少.本文介绍一种地高辛标记的LNA探针与靶标RNA相结合, 再通过免疫反应, 用偶联荧光基团的抗体检测地高辛半抗原, 最后通过超高分辨率激光共聚焦显微镜进行荧光成像, 对生物体内小RNA进行定位、定性或半定量分析的实验技术(Huang et al., 2019).本课题组前期通过大量研究证明, 小RNA能够参与调控植物侧生分生组织的起始发育(Zhang et al., 2018, 2020; Xue et al., 2020), 因此我们以模式植物拟南芥为实验材料, 详细介绍该项实验技术的具体操作流程及注意事项. ...
Multi-probe in situ hybridization to whole mount Arabidopsis seedlings 1 2011
... 基于RNA FISH改进的LNA探针原位杂交技术与免疫荧光相结合的小RNA荧光原位杂交(small RNA fluorescent in situ hybridization, sRNA-FISH)实验技术能够很好地解决上述问题.RNA-FISH属于荧光原位杂交技术的一种, 用于检测和定位细胞中特定的RNA分子.该方法使用与靶标RNA特异性结合的RNA分子作为探针, 探针上带有半抗原或者荧光素, 利用碱基互补配对原则, 可实现间接或者直接的荧光显色.RNA FISH最初被用于检测动物以及酵母(Saccharomyces spp.)中RNA的表达模式与细胞定位(Levsky and Singer, 2003).而在植物中, FISH技术最先被用于染色体以及染色体中特定基因座位的成像(Lamb et al., 2007; Tirichine et al., 2009; Jiang, 2019), 后来又被研究人员用于指示mRNA在植物体内的组织成像以及亚细胞定位(Bruno et al., 2011, 2015; Himanen et al., 2012; Rozier et al., 2014; Bleckmann and Dresselhaus, 2016; Yang et al., 2017, 2020; Woloszynska et al., 2019).针对小RNA的FISH技术也已广泛应用于动物系统中(Lu and Tsourkas, 2009), 但在植物中的应用还比较少.本文介绍一种地高辛标记的LNA探针与靶标RNA相结合, 再通过免疫反应, 用偶联荧光基团的抗体检测地高辛半抗原, 最后通过超高分辨率激光共聚焦显微镜进行荧光成像, 对生物体内小RNA进行定位、定性或半定量分析的实验技术(Huang et al., 2019).本课题组前期通过大量研究证明, 小RNA能够参与调控植物侧生分生组织的起始发育(Zhang et al., 2018, 2020; Xue et al., 2020), 因此我们以模式植物拟南芥为实验材料, 详细介绍该项实验技术的具体操作流程及注意事项. ...
Analysis of ATGUS1 and ATGUS2 in Arabidopsis root apex by a highly sensitive TSA-MISH method 1 2015
... 基于RNA FISH改进的LNA探针原位杂交技术与免疫荧光相结合的小RNA荧光原位杂交(small RNA fluorescent in situ hybridization, sRNA-FISH)实验技术能够很好地解决上述问题.RNA-FISH属于荧光原位杂交技术的一种, 用于检测和定位细胞中特定的RNA分子.该方法使用与靶标RNA特异性结合的RNA分子作为探针, 探针上带有半抗原或者荧光素, 利用碱基互补配对原则, 可实现间接或者直接的荧光显色.RNA FISH最初被用于检测动物以及酵母(Saccharomyces spp.)中RNA的表达模式与细胞定位(Levsky and Singer, 2003).而在植物中, FISH技术最先被用于染色体以及染色体中特定基因座位的成像(Lamb et al., 2007; Tirichine et al., 2009; Jiang, 2019), 后来又被研究人员用于指示mRNA在植物体内的组织成像以及亚细胞定位(Bruno et al., 2011, 2015; Himanen et al., 2012; Rozier et al., 2014; Bleckmann and Dresselhaus, 2016; Yang et al., 2017, 2020; Woloszynska et al., 2019).针对小RNA的FISH技术也已广泛应用于动物系统中(Lu and Tsourkas, 2009), 但在植物中的应用还比较少.本文介绍一种地高辛标记的LNA探针与靶标RNA相结合, 再通过免疫反应, 用偶联荧光基团的抗体检测地高辛半抗原, 最后通过超高分辨率激光共聚焦显微镜进行荧光成像, 对生物体内小RNA进行定位、定性或半定量分析的实验技术(Huang et al., 2019).本课题组前期通过大量研究证明, 小RNA能够参与调控植物侧生分生组织的起始发育(Zhang et al., 2018, 2020; Xue et al., 2020), 因此我们以模式植物拟南芥为实验材料, 详细介绍该项实验技术的具体操作流程及注意事项. ...
Identification and characterization of small RNAs from the phloem of Brassica napus 1 2008
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
A microRNA as a translational repressor of APETALA2 in Arabidopsis flower development 1 2004
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
Small RNAs and their roles in plant development 1 2009
... 小RNA是动植物中一类非常重要的18-30 nt小分子核苷酸(Chen, 2009; Kim et al., 2009), 成熟的小RNA通过对靶基因进行转录或转录后水平的调节参与多种植物的生长发育与胁迫响应过程.目前植物中主要有3种小RNA,分别为miRNA (microRNA)、siRNA (Small interfering RNA)和phasiRNA (phased secondary small interfering RNA) (Yu et al., 2019).在植物细胞中, miRNA的生物合成过程已有报道, miRNA编码基因在RNA聚合酶II的作用下被转录至细胞核中, 形成初级miRNA (Primary miRNA, Pri- miRNA) (Lee et al., 2004; Jones-Rhoades et al., 2006).Pri-miRNA在DCL1 (DICER-LIKE 1)、HYL1 (HYPONASTIC LEAVES 1)以及SE (SERRATE)等蛋白因子的作用下被切割成具有互补茎环结构的前体miRNA (Precursor miRNA, Pre-miRNA), Pre- miRNA又被加工成miRNA/miRNA*双链体.在HEN1 (HUA ENHANCER 1)的作用下, miRNA/miRNA*双链聚合体的3'端被甲基化修饰, 以免被降解, 稳定的miRNA/miRNA*双链聚合体在HASTY蛋白的作用下被运输至细胞质中(Kurihara and Watanabe, 2004; Li et al., 2005; Park et al., 2005; Jones-Rhoades et al., 2006).在细胞质中, miRNA/miRNA*双链聚合物 ...
Pattern formation in leaves via small RNA mobility 1 2008
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
Pattern formation via small RNA mobility 2 2009
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
... ).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
Paraffin embedding tissue samples for sectioning 1 2008
... 固定完成后, 可以根据提供的方法(Fischer et al., 2008)进行石蜡包埋.包埋后的样品可以在4°C冰箱保存6个月. ...
Histone H2b monoubiquitination is required to reach maximal transcript levels of circadian clock genes in Arabidopsis 1 2012
... 基于RNA FISH改进的LNA探针原位杂交技术与免疫荧光相结合的小RNA荧光原位杂交(small RNA fluorescent in situ hybridization, sRNA-FISH)实验技术能够很好地解决上述问题.RNA-FISH属于荧光原位杂交技术的一种, 用于检测和定位细胞中特定的RNA分子.该方法使用与靶标RNA特异性结合的RNA分子作为探针, 探针上带有半抗原或者荧光素, 利用碱基互补配对原则, 可实现间接或者直接的荧光显色.RNA FISH最初被用于检测动物以及酵母(Saccharomyces spp.)中RNA的表达模式与细胞定位(Levsky and Singer, 2003).而在植物中, FISH技术最先被用于染色体以及染色体中特定基因座位的成像(Lamb et al., 2007; Tirichine et al., 2009; Jiang, 2019), 后来又被研究人员用于指示mRNA在植物体内的组织成像以及亚细胞定位(Bruno et al., 2011, 2015; Himanen et al., 2012; Rozier et al., 2014; Bleckmann and Dresselhaus, 2016; Yang et al., 2017, 2020; Woloszynska et al., 2019).针对小RNA的FISH技术也已广泛应用于动物系统中(Lu and Tsourkas, 2009), 但在植物中的应用还比较少.本文介绍一种地高辛标记的LNA探针与靶标RNA相结合, 再通过免疫反应, 用偶联荧光基团的抗体检测地高辛半抗原, 最后通过超高分辨率激光共聚焦显微镜进行荧光成像, 对生物体内小RNA进行定位、定性或半定量分析的实验技术(Huang et al., 2019).本课题组前期通过大量研究证明, 小RNA能够参与调控植物侧生分生组织的起始发育(Zhang et al., 2018, 2020; Xue et al., 2020), 因此我们以模式植物拟南芥为实验材料, 详细介绍该项实验技术的具体操作流程及注意事项. ...
Genome-wide analysis of the RNA-DEPENDENT RNA POLYMERASE6/DICER-LIKE4 pathway in Arabidop- sis reveals dependency on miRNA- and tasiRNA-directed targeting 1 2007
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
sRNA-FISH: versatile fluorescent in situ detection of small RNAs in plants 1 2019
... 基于RNA FISH改进的LNA探针原位杂交技术与免疫荧光相结合的小RNA荧光原位杂交(small RNA fluorescent in situ hybridization, sRNA-FISH)实验技术能够很好地解决上述问题.RNA-FISH属于荧光原位杂交技术的一种, 用于检测和定位细胞中特定的RNA分子.该方法使用与靶标RNA特异性结合的RNA分子作为探针, 探针上带有半抗原或者荧光素, 利用碱基互补配对原则, 可实现间接或者直接的荧光显色.RNA FISH最初被用于检测动物以及酵母(Saccharomyces spp.)中RNA的表达模式与细胞定位(Levsky and Singer, 2003).而在植物中, FISH技术最先被用于染色体以及染色体中特定基因座位的成像(Lamb et al., 2007; Tirichine et al., 2009; Jiang, 2019), 后来又被研究人员用于指示mRNA在植物体内的组织成像以及亚细胞定位(Bruno et al., 2011, 2015; Himanen et al., 2012; Rozier et al., 2014; Bleckmann and Dresselhaus, 2016; Yang et al., 2017, 2020; Woloszynska et al., 2019).针对小RNA的FISH技术也已广泛应用于动物系统中(Lu and Tsourkas, 2009), 但在植物中的应用还比较少.本文介绍一种地高辛标记的LNA探针与靶标RNA相结合, 再通过免疫反应, 用偶联荧光基团的抗体检测地高辛半抗原, 最后通过超高分辨率激光共聚焦显微镜进行荧光成像, 对生物体内小RNA进行定位、定性或半定量分析的实验技术(Huang et al., 2019).本课题组前期通过大量研究证明, 小RNA能够参与调控植物侧生分生组织的起始发育(Zhang et al., 2018, 2020; Xue et al., 2020), 因此我们以模式植物拟南芥为实验材料, 详细介绍该项实验技术的具体操作流程及注意事项. ...
Long-distance movement of phospha- te starvation-responsive microRNAs in Arabidopsis 1 2017
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
In situ localization of small RNAs in plants by using LNA probes 1 2012
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
Fluorescence in situ hybridization in plants: recent developments and future applications 1 2019
... 基于RNA FISH改进的LNA探针原位杂交技术与免疫荧光相结合的小RNA荧光原位杂交(small RNA fluorescent in situ hybridization, sRNA-FISH)实验技术能够很好地解决上述问题.RNA-FISH属于荧光原位杂交技术的一种, 用于检测和定位细胞中特定的RNA分子.该方法使用与靶标RNA特异性结合的RNA分子作为探针, 探针上带有半抗原或者荧光素, 利用碱基互补配对原则, 可实现间接或者直接的荧光显色.RNA FISH最初被用于检测动物以及酵母(Saccharomyces spp.)中RNA的表达模式与细胞定位(Levsky and Singer, 2003).而在植物中, FISH技术最先被用于染色体以及染色体中特定基因座位的成像(Lamb et al., 2007; Tirichine et al., 2009; Jiang, 2019), 后来又被研究人员用于指示mRNA在植物体内的组织成像以及亚细胞定位(Bruno et al., 2011, 2015; Himanen et al., 2012; Rozier et al., 2014; Bleckmann and Dresselhaus, 2016; Yang et al., 2017, 2020; Woloszynska et al., 2019).针对小RNA的FISH技术也已广泛应用于动物系统中(Lu and Tsourkas, 2009), 但在植物中的应用还比较少.本文介绍一种地高辛标记的LNA探针与靶标RNA相结合, 再通过免疫反应, 用偶联荧光基团的抗体检测地高辛半抗原, 最后通过超高分辨率激光共聚焦显微镜进行荧光成像, 对生物体内小RNA进行定位、定性或半定量分析的实验技术(Huang et al., 2019).本课题组前期通过大量研究证明, 小RNA能够参与调控植物侧生分生组织的起始发育(Zhang et al., 2018, 2020; Xue et al., 2020), 因此我们以模式植物拟南芥为实验材料, 详细介绍该项实验技术的具体操作流程及注意事项. ...
Micro- RNAs and their regulatory roles in plants 2 2006
... 小RNA是动植物中一类非常重要的18-30 nt小分子核苷酸(Chen, 2009; Kim et al., 2009), 成熟的小RNA通过对靶基因进行转录或转录后水平的调节参与多种植物的生长发育与胁迫响应过程.目前植物中主要有3种小RNA,分别为miRNA (microRNA)、siRNA (Small interfering RNA)和phasiRNA (phased secondary small interfering RNA) (Yu et al., 2019).在植物细胞中, miRNA的生物合成过程已有报道, miRNA编码基因在RNA聚合酶II的作用下被转录至细胞核中, 形成初级miRNA (Primary miRNA, Pri- miRNA) (Lee et al., 2004; Jones-Rhoades et al., 2006).Pri-miRNA在DCL1 (DICER-LIKE 1)、HYL1 (HYPONASTIC LEAVES 1)以及SE (SERRATE)等蛋白因子的作用下被切割成具有互补茎环结构的前体miRNA (Precursor miRNA, Pre-miRNA), Pre- miRNA又被加工成miRNA/miRNA*双链体.在HEN1 (HUA ENHANCER 1)的作用下, miRNA/miRNA*双链聚合体的3'端被甲基化修饰, 以免被降解, 稳定的miRNA/miRNA*双链聚合体在HASTY蛋白的作用下被运输至细胞质中(Kurihara and Watanabe, 2004; Li et al., 2005; Park et al., 2005; Jones-Rhoades et al., 2006).在细胞质中, miRNA/miRNA*双链聚合物 ...
... ; Jones-Rhoades et al., 2006).在细胞质中, miRNA/miRNA*双链聚合物 ...
MIR166/ 165 genes exhibit dynamic expression patterns in regulating shoot apical meris- tem and floral development in Arabidopsis 1 2007
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
In situ hybridization as a tool to study the role of microRNAs in plant development 1 2006
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
Biogenesis of small RNAs in animals 1 2009
... 小RNA是动植物中一类非常重要的18-30 nt小分子核苷酸(Chen, 2009; Kim et al., 2009), 成熟的小RNA通过对靶基因进行转录或转录后水平的调节参与多种植物的生长发育与胁迫响应过程.目前植物中主要有3种小RNA,分别为miRNA (microRNA)、siRNA (Small interfering RNA)和phasiRNA (phased secondary small interfering RNA) (Yu et al., 2019).在植物细胞中, miRNA的生物合成过程已有报道, miRNA编码基因在RNA聚合酶II的作用下被转录至细胞核中, 形成初级miRNA (Primary miRNA, Pri- miRNA) (Lee et al., 2004; Jones-Rhoades et al., 2006).Pri-miRNA在DCL1 (DICER-LIKE 1)、HYL1 (HYPONASTIC LEAVES 1)以及SE (SERRATE)等蛋白因子的作用下被切割成具有互补茎环结构的前体miRNA (Precursor miRNA, Pre-miRNA), Pre- miRNA又被加工成miRNA/miRNA*双链体.在HEN1 (HUA ENHANCER 1)的作用下, miRNA/miRNA*双链聚合体的3'端被甲基化修饰, 以免被降解, 稳定的miRNA/miRNA*双链聚合体在HASTY蛋白的作用下被运输至细胞质中(Kurihara and Watanabe, 2004; Li et al., 2005; Park et al., 2005; Jones-Rhoades et al., 2006).在细胞质中, miRNA/miRNA*双链聚合物 ...
A protodermal miR394 signal defines a region of stem cell competence in the Arabidopsis shoot meristem 1 2013
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
Arabidopsis micro-RNA biogenesis through dicer-like 1 protein functions 1 2004
... 小RNA是动植物中一类非常重要的18-30 nt小分子核苷酸(Chen, 2009; Kim et al., 2009), 成熟的小RNA通过对靶基因进行转录或转录后水平的调节参与多种植物的生长发育与胁迫响应过程.目前植物中主要有3种小RNA,分别为miRNA (microRNA)、siRNA (Small interfering RNA)和phasiRNA (phased secondary small interfering RNA) (Yu et al., 2019).在植物细胞中, miRNA的生物合成过程已有报道, miRNA编码基因在RNA聚合酶II的作用下被转录至细胞核中, 形成初级miRNA (Primary miRNA, Pri- miRNA) (Lee et al., 2004; Jones-Rhoades et al., 2006).Pri-miRNA在DCL1 (DICER-LIKE 1)、HYL1 (HYPONASTIC LEAVES 1)以及SE (SERRATE)等蛋白因子的作用下被切割成具有互补茎环结构的前体miRNA (Precursor miRNA, Pre-miRNA), Pre- miRNA又被加工成miRNA/miRNA*双链体.在HEN1 (HUA ENHANCER 1)的作用下, miRNA/miRNA*双链聚合体的3'端被甲基化修饰, 以免被降解, 稳定的miRNA/miRNA*双链聚合体在HASTY蛋白的作用下被运输至细胞质中(Kurihara and Watanabe, 2004; Li et al., 2005; Park et al., 2005; Jones-Rhoades et al., 2006).在细胞质中, miRNA/miRNA*双链聚合物 ...
Single-gene detection and karyotyping using small-target fluorescence in situ hybridization on maize somatic chromosomes 1 2007
... 基于RNA FISH改进的LNA探针原位杂交技术与免疫荧光相结合的小RNA荧光原位杂交(small RNA fluorescent in situ hybridization, sRNA-FISH)实验技术能够很好地解决上述问题.RNA-FISH属于荧光原位杂交技术的一种, 用于检测和定位细胞中特定的RNA分子.该方法使用与靶标RNA特异性结合的RNA分子作为探针, 探针上带有半抗原或者荧光素, 利用碱基互补配对原则, 可实现间接或者直接的荧光显色.RNA FISH最初被用于检测动物以及酵母(Saccharomyces spp.)中RNA的表达模式与细胞定位(Levsky and Singer, 2003).而在植物中, FISH技术最先被用于染色体以及染色体中特定基因座位的成像(Lamb et al., 2007; Tirichine et al., 2009; Jiang, 2019), 后来又被研究人员用于指示mRNA在植物体内的组织成像以及亚细胞定位(Bruno et al., 2011, 2015; Himanen et al., 2012; Rozier et al., 2014; Bleckmann and Dresselhaus, 2016; Yang et al., 2017, 2020; Woloszynska et al., 2019).针对小RNA的FISH技术也已广泛应用于动物系统中(Lu and Tsourkas, 2009), 但在植物中的应用还比较少.本文介绍一种地高辛标记的LNA探针与靶标RNA相结合, 再通过免疫反应, 用偶联荧光基团的抗体检测地高辛半抗原, 最后通过超高分辨率激光共聚焦显微镜进行荧光成像, 对生物体内小RNA进行定位、定性或半定量分析的实验技术(Huang et al., 2019).本课题组前期通过大量研究证明, 小RNA能够参与调控植物侧生分生组织的起始发育(Zhang et al., 2018, 2020; Xue et al., 2020), 因此我们以模式植物拟南芥为实验材料, 详细介绍该项实验技术的具体操作流程及注意事项. ...
MicroRNA genes are transcribed by RNA polymerase II 1 2004
... 小RNA是动植物中一类非常重要的18-30 nt小分子核苷酸(Chen, 2009; Kim et al., 2009), 成熟的小RNA通过对靶基因进行转录或转录后水平的调节参与多种植物的生长发育与胁迫响应过程.目前植物中主要有3种小RNA,分别为miRNA (microRNA)、siRNA (Small interfering RNA)和phasiRNA (phased secondary small interfering RNA) (Yu et al., 2019).在植物细胞中, miRNA的生物合成过程已有报道, miRNA编码基因在RNA聚合酶II的作用下被转录至细胞核中, 形成初级miRNA (Primary miRNA, Pri- miRNA) (Lee et al., 2004; Jones-Rhoades et al., 2006).Pri-miRNA在DCL1 (DICER-LIKE 1)、HYL1 (HYPONASTIC LEAVES 1)以及SE (SERRATE)等蛋白因子的作用下被切割成具有互补茎环结构的前体miRNA (Precursor miRNA, Pre-miRNA), Pre- miRNA又被加工成miRNA/miRNA*双链体.在HEN1 (HUA ENHANCER 1)的作用下, miRNA/miRNA*双链聚合体的3'端被甲基化修饰, 以免被降解, 稳定的miRNA/miRNA*双链聚合体在HASTY蛋白的作用下被运输至细胞质中(Kurihara and Watanabe, 2004; Li et al., 2005; Park et al., 2005; Jones-Rhoades et al., 2006).在细胞质中, miRNA/miRNA*双链聚合物 ...
Fluorescence in situ hybridization: past, present and future 1 2003
... 基于RNA FISH改进的LNA探针原位杂交技术与免疫荧光相结合的小RNA荧光原位杂交(small RNA fluorescent in situ hybridization, sRNA-FISH)实验技术能够很好地解决上述问题.RNA-FISH属于荧光原位杂交技术的一种, 用于检测和定位细胞中特定的RNA分子.该方法使用与靶标RNA特异性结合的RNA分子作为探针, 探针上带有半抗原或者荧光素, 利用碱基互补配对原则, 可实现间接或者直接的荧光显色.RNA FISH最初被用于检测动物以及酵母(Saccharomyces spp.)中RNA的表达模式与细胞定位(Levsky and Singer, 2003).而在植物中, FISH技术最先被用于染色体以及染色体中特定基因座位的成像(Lamb et al., 2007; Tirichine et al., 2009; Jiang, 2019), 后来又被研究人员用于指示mRNA在植物体内的组织成像以及亚细胞定位(Bruno et al., 2011, 2015; Himanen et al., 2012; Rozier et al., 2014; Bleckmann and Dresselhaus, 2016; Yang et al., 2017, 2020; Woloszynska et al., 2019).针对小RNA的FISH技术也已广泛应用于动物系统中(Lu and Tsourkas, 2009), 但在植物中的应用还比较少.本文介绍一种地高辛标记的LNA探针与靶标RNA相结合, 再通过免疫反应, 用偶联荧光基团的抗体检测地高辛半抗原, 最后通过超高分辨率激光共聚焦显微镜进行荧光成像, 对生物体内小RNA进行定位、定性或半定量分析的实验技术(Huang et al., 2019).本课题组前期通过大量研究证明, 小RNA能够参与调控植物侧生分生组织的起始发育(Zhang et al., 2018, 2020; Xue et al., 2020), 因此我们以模式植物拟南芥为实验材料, 详细介绍该项实验技术的具体操作流程及注意事项. ...
Methylation protects miRNAs and siRNAs from a 3'-end uridylation activity in Arabidopsis 1 2005
... 小RNA是动植物中一类非常重要的18-30 nt小分子核苷酸(Chen, 2009; Kim et al., 2009), 成熟的小RNA通过对靶基因进行转录或转录后水平的调节参与多种植物的生长发育与胁迫响应过程.目前植物中主要有3种小RNA,分别为miRNA (microRNA)、siRNA (Small interfering RNA)和phasiRNA (phased secondary small interfering RNA) (Yu et al., 2019).在植物细胞中, miRNA的生物合成过程已有报道, miRNA编码基因在RNA聚合酶II的作用下被转录至细胞核中, 形成初级miRNA (Primary miRNA, Pri- miRNA) (Lee et al., 2004; Jones-Rhoades et al., 2006).Pri-miRNA在DCL1 (DICER-LIKE 1)、HYL1 (HYPONASTIC LEAVES 1)以及SE (SERRATE)等蛋白因子的作用下被切割成具有互补茎环结构的前体miRNA (Precursor miRNA, Pre-miRNA), Pre- miRNA又被加工成miRNA/miRNA*双链体.在HEN1 (HUA ENHANCER 1)的作用下, miRNA/miRNA*双链聚合体的3'端被甲基化修饰, 以免被降解, 稳定的miRNA/miRNA*双链聚合体在HASTY蛋白的作用下被运输至细胞质中(Kurihara and Watanabe, 2004; Li et al., 2005; Park et al., 2005; Jones-Rhoades et al., 2006).在细胞质中, miRNA/miRNA*双链聚合物 ...
Unidirectional movement of small RNAs from shoots to roots in interspecific heterografts 1 2021
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
Regulatory network of microRNA399 and PHO2 by systemic signaling 1 2008
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
The ARGONAUTE10 gene modulates shoot apical meris- tem maintenance and establishment of leaf polarity by repressing miR165/166 in Arabidopsis 1 2009
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
Imaging individual microRNAs in single mammalian cells in situ 1 2009
... 基于RNA FISH改进的LNA探针原位杂交技术与免疫荧光相结合的小RNA荧光原位杂交(small RNA fluorescent in situ hybridization, sRNA-FISH)实验技术能够很好地解决上述问题.RNA-FISH属于荧光原位杂交技术的一种, 用于检测和定位细胞中特定的RNA分子.该方法使用与靶标RNA特异性结合的RNA分子作为探针, 探针上带有半抗原或者荧光素, 利用碱基互补配对原则, 可实现间接或者直接的荧光显色.RNA FISH最初被用于检测动物以及酵母(Saccharomyces spp.)中RNA的表达模式与细胞定位(Levsky and Singer, 2003).而在植物中, FISH技术最先被用于染色体以及染色体中特定基因座位的成像(Lamb et al., 2007; Tirichine et al., 2009; Jiang, 2019), 后来又被研究人员用于指示mRNA在植物体内的组织成像以及亚细胞定位(Bruno et al., 2011, 2015; Himanen et al., 2012; Rozier et al., 2014; Bleckmann and Dresselhaus, 2016; Yang et al., 2017, 2020; Woloszynska et al., 2019).针对小RNA的FISH技术也已广泛应用于动物系统中(Lu and Tsourkas, 2009), 但在植物中的应用还比较少.本文介绍一种地高辛标记的LNA探针与靶标RNA相结合, 再通过免疫反应, 用偶联荧光基团的抗体检测地高辛半抗原, 最后通过超高分辨率激光共聚焦显微镜进行荧光成像, 对生物体内小RNA进行定位、定性或半定量分析的实验技术(Huang et al., 2019).本课题组前期通过大量研究证明, 小RNA能够参与调控植物侧生分生组织的起始发育(Zhang et al., 2018, 2020; Xue et al., 2020), 因此我们以模式植物拟南芥为实验材料, 详细介绍该项实验技术的具体操作流程及注意事项. ...
Transitivity in Arabidopsis can be primed, requires the redundant action of the antiviral Dicer-like 4 and Dicer-like 2, and is compromised by viral-encoded suppressor proteins 1 2007
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
MicroRNAs prevent precocious gene expression and enable pattern formation during plant embryogenesis 1 2010
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
Regulation of small RNA accumulation in the maize shoot apex 1 2009
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
Two small regulatory RNAs establish opposing fates of a developmental axis 1 2007
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
Regulation of LANCEOLATE by miR319 is required for compound-leaf development in tomato 1 2007
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
Small RNA mobility: spread of RNA silencing effectors and its effect on developmental processes and stress adaptation in plants 1 2019
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
MicroRNA399 is a long-distance signal for the regulation of plant phosphate homeostasis 1 2008
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
Identification of nutrient-responsive Arabidopsis and rapeseed microRNAs by comprehensive real-time polymerase chain reaction profiling and small RNA sequencing 1 2009
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
In vivo investigation of the transcription, processing, endonucleolytic activity, and functional relevance of the spatial distribution of a plant miRNA 1 2004
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
Nuclear processing and export of microRNAs in Arabidopsis 1 2005
... 小RNA是动植物中一类非常重要的18-30 nt小分子核苷酸(Chen, 2009; Kim et al., 2009), 成熟的小RNA通过对靶基因进行转录或转录后水平的调节参与多种植物的生长发育与胁迫响应过程.目前植物中主要有3种小RNA,分别为miRNA (microRNA)、siRNA (Small interfering RNA)和phasiRNA (phased secondary small interfering RNA) (Yu et al., 2019).在植物细胞中, miRNA的生物合成过程已有报道, miRNA编码基因在RNA聚合酶II的作用下被转录至细胞核中, 形成初级miRNA (Primary miRNA, Pri- miRNA) (Lee et al., 2004; Jones-Rhoades et al., 2006).Pri-miRNA在DCL1 (DICER-LIKE 1)、HYL1 (HYPONASTIC LEAVES 1)以及SE (SERRATE)等蛋白因子的作用下被切割成具有互补茎环结构的前体miRNA (Precursor miRNA, Pre-miRNA), Pre- miRNA又被加工成miRNA/miRNA*双链体.在HEN1 (HUA ENHANCER 1)的作用下, miRNA/miRNA*双链聚合体的3'端被甲基化修饰, 以免被降解, 稳定的miRNA/miRNA*双链聚合体在HASTY蛋白的作用下被运输至细胞质中(Kurihara and Watanabe, 2004; Li et al., 2005; Park et al., 2005; Jones-Rhoades et al., 2006).在细胞质中, miRNA/miRNA*双链聚合物 ...
Interplay of miR164, CUP-SHAPED COTYLEDON genes and LATERAL SUPPRESSOR controls axillary meristem formation in Arabidopsis thaliana 1 2008
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
Analysis of 3D gene expression patterns in plants using whole-mount RNA in situ hybridization 1 2014
... 基于RNA FISH改进的LNA探针原位杂交技术与免疫荧光相结合的小RNA荧光原位杂交(small RNA fluorescent in situ hybridization, sRNA-FISH)实验技术能够很好地解决上述问题.RNA-FISH属于荧光原位杂交技术的一种, 用于检测和定位细胞中特定的RNA分子.该方法使用与靶标RNA特异性结合的RNA分子作为探针, 探针上带有半抗原或者荧光素, 利用碱基互补配对原则, 可实现间接或者直接的荧光显色.RNA FISH最初被用于检测动物以及酵母(Saccharomyces spp.)中RNA的表达模式与细胞定位(Levsky and Singer, 2003).而在植物中, FISH技术最先被用于染色体以及染色体中特定基因座位的成像(Lamb et al., 2007; Tirichine et al., 2009; Jiang, 2019), 后来又被研究人员用于指示mRNA在植物体内的组织成像以及亚细胞定位(Bruno et al., 2011, 2015; Himanen et al., 2012; Rozier et al., 2014; Bleckmann and Dresselhaus, 2016; Yang et al., 2017, 2020; Woloszynska et al., 2019).针对小RNA的FISH技术也已广泛应用于动物系统中(Lu and Tsourkas, 2009), 但在植物中的应用还比较少.本文介绍一种地高辛标记的LNA探针与靶标RNA相结合, 再通过免疫反应, 用偶联荧光基团的抗体检测地高辛半抗原, 最后通过超高分辨率激光共聚焦显微镜进行荧光成像, 对生物体内小RNA进行定位、定性或半定量分析的实验技术(Huang et al., 2019).本课题组前期通过大量研究证明, 小RNA能够参与调控植物侧生分生组织的起始发育(Zhang et al., 2018, 2020; Xue et al., 2020), 因此我们以模式植物拟南芥为实验材料, 详细介绍该项实验技术的具体操作流程及注意事项. ...
Gating of miRNA movement at defined cell-cell interfaces governs their impact as positional signals 1 2018
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
3D fluorescent in situ hybridization using Arabidopsis leaf cryosections and isolated nuclei 1 2009
... 基于RNA FISH改进的LNA探针原位杂交技术与免疫荧光相结合的小RNA荧光原位杂交(small RNA fluorescent in situ hybridization, sRNA-FISH)实验技术能够很好地解决上述问题.RNA-FISH属于荧光原位杂交技术的一种, 用于检测和定位细胞中特定的RNA分子.该方法使用与靶标RNA特异性结合的RNA分子作为探针, 探针上带有半抗原或者荧光素, 利用碱基互补配对原则, 可实现间接或者直接的荧光显色.RNA FISH最初被用于检测动物以及酵母(Saccharomyces spp.)中RNA的表达模式与细胞定位(Levsky and Singer, 2003).而在植物中, FISH技术最先被用于染色体以及染色体中特定基因座位的成像(Lamb et al., 2007; Tirichine et al., 2009; Jiang, 2019), 后来又被研究人员用于指示mRNA在植物体内的组织成像以及亚细胞定位(Bruno et al., 2011, 2015; Himanen et al., 2012; Rozier et al., 2014; Bleckmann and Dresselhaus, 2016; Yang et al., 2017, 2020; Woloszynska et al., 2019).针对小RNA的FISH技术也已广泛应用于动物系统中(Lu and Tsourkas, 2009), 但在植物中的应用还比较少.本文介绍一种地高辛标记的LNA探针与靶标RNA相结合, 再通过免疫反应, 用偶联荧光基团的抗体检测地高辛半抗原, 最后通过超高分辨率激光共聚焦显微镜进行荧光成像, 对生物体内小RNA进行定位、定性或半定量分析的实验技术(Huang et al., 2019).本课题组前期通过大量研究证明, 小RNA能够参与调控植物侧生分生组织的起始发育(Zhang et al., 2018, 2020; Xue et al., 2020), 因此我们以模式植物拟南芥为实验材料, 详细介绍该项实验技术的具体操作流程及注意事项. ...
The action of ARGONAUTE1 in the miRNA pathway and its regulation by the miRNA pathway are crucial for plant development 1 2004
... 解旋, 反义链被降解, 成熟miRNA单链与含有RNA解旋酶和AGO (ARGONAUTE)蛋白的RNA诱导沉默复合体(RNA-induced silencing complex, RISC)结合, 控制其相关靶基因的表达(Vaucheret et al., 2004; Baumberger and Baulcombe, 2005). ...
On reconciling the interactions between APETA- LA2, miR172 and AGAMOUS with the ABC model of flower development 1 2010
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
Histone 2B monoubiquitination complex integrates transcript elongation with RNA processing at circadian clock and flowering regulators 1 2019
... 基于RNA FISH改进的LNA探针原位杂交技术与免疫荧光相结合的小RNA荧光原位杂交(small RNA fluorescent in situ hybridization, sRNA-FISH)实验技术能够很好地解决上述问题.RNA-FISH属于荧光原位杂交技术的一种, 用于检测和定位细胞中特定的RNA分子.该方法使用与靶标RNA特异性结合的RNA分子作为探针, 探针上带有半抗原或者荧光素, 利用碱基互补配对原则, 可实现间接或者直接的荧光显色.RNA FISH最初被用于检测动物以及酵母(Saccharomyces spp.)中RNA的表达模式与细胞定位(Levsky and Singer, 2003).而在植物中, FISH技术最先被用于染色体以及染色体中特定基因座位的成像(Lamb et al., 2007; Tirichine et al., 2009; Jiang, 2019), 后来又被研究人员用于指示mRNA在植物体内的组织成像以及亚细胞定位(Bruno et al., 2011, 2015; Himanen et al., 2012; Rozier et al., 2014; Bleckmann and Dresselhaus, 2016; Yang et al., 2017, 2020; Woloszynska et al., 2019).针对小RNA的FISH技术也已广泛应用于动物系统中(Lu and Tsourkas, 2009), 但在植物中的应用还比较少.本文介绍一种地高辛标记的LNA探针与靶标RNA相结合, 再通过免疫反应, 用偶联荧光基团的抗体检测地高辛半抗原, 最后通过超高分辨率激光共聚焦显微镜进行荧光成像, 对生物体内小RNA进行定位、定性或半定量分析的实验技术(Huang et al., 2019).本课题组前期通过大量研究证明, 小RNA能够参与调控植物侧生分生组织的起始发育(Zhang et al., 2018, 2020; Xue et al., 2020), 因此我们以模式植物拟南芥为实验材料, 详细介绍该项实验技术的具体操作流程及注意事项. ...
Regulation of shoot apical meristem and axillary meristem development in plants 1 2020
... 基于RNA FISH改进的LNA探针原位杂交技术与免疫荧光相结合的小RNA荧光原位杂交(small RNA fluorescent in situ hybridization, sRNA-FISH)实验技术能够很好地解决上述问题.RNA-FISH属于荧光原位杂交技术的一种, 用于检测和定位细胞中特定的RNA分子.该方法使用与靶标RNA特异性结合的RNA分子作为探针, 探针上带有半抗原或者荧光素, 利用碱基互补配对原则, 可实现间接或者直接的荧光显色.RNA FISH最初被用于检测动物以及酵母(Saccharomyces spp.)中RNA的表达模式与细胞定位(Levsky and Singer, 2003).而在植物中, FISH技术最先被用于染色体以及染色体中特定基因座位的成像(Lamb et al., 2007; Tirichine et al., 2009; Jiang, 2019), 后来又被研究人员用于指示mRNA在植物体内的组织成像以及亚细胞定位(Bruno et al., 2011, 2015; Himanen et al., 2012; Rozier et al., 2014; Bleckmann and Dresselhaus, 2016; Yang et al., 2017, 2020; Woloszynska et al., 2019).针对小RNA的FISH技术也已广泛应用于动物系统中(Lu and Tsourkas, 2009), 但在植物中的应用还比较少.本文介绍一种地高辛标记的LNA探针与靶标RNA相结合, 再通过免疫反应, 用偶联荧光基团的抗体检测地高辛半抗原, 最后通过超高分辨率激光共聚焦显微镜进行荧光成像, 对生物体内小RNA进行定位、定性或半定量分析的实验技术(Huang et al., 2019).本课题组前期通过大量研究证明, 小RNA能够参与调控植物侧生分生组织的起始发育(Zhang et al., 2018, 2020; Xue et al., 2020), 因此我们以模式植物拟南芥为实验材料, 详细介绍该项实验技术的具体操作流程及注意事项. ...
Visualization of protein coding, long noncoding, and nuclear RNAs by fluorescence in situ hybridization in sections of shoot apical meristems and developing flowers 1 2020
... 基于RNA FISH改进的LNA探针原位杂交技术与免疫荧光相结合的小RNA荧光原位杂交(small RNA fluorescent in situ hybridization, sRNA-FISH)实验技术能够很好地解决上述问题.RNA-FISH属于荧光原位杂交技术的一种, 用于检测和定位细胞中特定的RNA分子.该方法使用与靶标RNA特异性结合的RNA分子作为探针, 探针上带有半抗原或者荧光素, 利用碱基互补配对原则, 可实现间接或者直接的荧光显色.RNA FISH最初被用于检测动物以及酵母(Saccharomyces spp.)中RNA的表达模式与细胞定位(Levsky and Singer, 2003).而在植物中, FISH技术最先被用于染色体以及染色体中特定基因座位的成像(Lamb et al., 2007; Tirichine et al., 2009; Jiang, 2019), 后来又被研究人员用于指示mRNA在植物体内的组织成像以及亚细胞定位(Bruno et al., 2011, 2015; Himanen et al., 2012; Rozier et al., 2014; Bleckmann and Dresselhaus, 2016; Yang et al., 2017, 2020; Woloszynska et al., 2019).针对小RNA的FISH技术也已广泛应用于动物系统中(Lu and Tsourkas, 2009), 但在植物中的应用还比较少.本文介绍一种地高辛标记的LNA探针与靶标RNA相结合, 再通过免疫反应, 用偶联荧光基团的抗体检测地高辛半抗原, 最后通过超高分辨率激光共聚焦显微镜进行荧光成像, 对生物体内小RNA进行定位、定性或半定量分析的实验技术(Huang et al., 2019).本课题组前期通过大量研究证明, 小RNA能够参与调控植物侧生分生组织的起始发育(Zhang et al., 2018, 2020; Xue et al., 2020), 因此我们以模式植物拟南芥为实验材料, 详细介绍该项实验技术的具体操作流程及注意事项. ...
Cell cycle control by nuclear sequestration of CDC20 and CDH1 mRNA in plant stem cells 1 2017
... 基于RNA FISH改进的LNA探针原位杂交技术与免疫荧光相结合的小RNA荧光原位杂交(small RNA fluorescent in situ hybridization, sRNA-FISH)实验技术能够很好地解决上述问题.RNA-FISH属于荧光原位杂交技术的一种, 用于检测和定位细胞中特定的RNA分子.该方法使用与靶标RNA特异性结合的RNA分子作为探针, 探针上带有半抗原或者荧光素, 利用碱基互补配对原则, 可实现间接或者直接的荧光显色.RNA FISH最初被用于检测动物以及酵母(Saccharomyces spp.)中RNA的表达模式与细胞定位(Levsky and Singer, 2003).而在植物中, FISH技术最先被用于染色体以及染色体中特定基因座位的成像(Lamb et al., 2007; Tirichine et al., 2009; Jiang, 2019), 后来又被研究人员用于指示mRNA在植物体内的组织成像以及亚细胞定位(Bruno et al., 2011, 2015; Himanen et al., 2012; Rozier et al., 2014; Bleckmann and Dresselhaus, 2016; Yang et al., 2017, 2020; Woloszynska et al., 2019).针对小RNA的FISH技术也已广泛应用于动物系统中(Lu and Tsourkas, 2009), 但在植物中的应用还比较少.本文介绍一种地高辛标记的LNA探针与靶标RNA相结合, 再通过免疫反应, 用偶联荧光基团的抗体检测地高辛半抗原, 最后通过超高分辨率激光共聚焦显微镜进行荧光成像, 对生物体内小RNA进行定位、定性或半定量分析的实验技术(Huang et al., 2019).本课题组前期通过大量研究证明, 小RNA能够参与调控植物侧生分生组织的起始发育(Zhang et al., 2018, 2020; Xue et al., 2020), 因此我们以模式植物拟南芥为实验材料, 详细介绍该项实验技术的具体操作流程及注意事项. ...
Plant noncoding RNAs: hidden players in development and stress responses 2 2019
... 小RNA是动植物中一类非常重要的18-30 nt小分子核苷酸(Chen, 2009; Kim et al., 2009), 成熟的小RNA通过对靶基因进行转录或转录后水平的调节参与多种植物的生长发育与胁迫响应过程.目前植物中主要有3种小RNA,分别为miRNA (microRNA)、siRNA (Small interfering RNA)和phasiRNA (phased secondary small interfering RNA) (Yu et al., 2019).在植物细胞中, miRNA的生物合成过程已有报道, miRNA编码基因在RNA聚合酶II的作用下被转录至细胞核中, 形成初级miRNA (Primary miRNA, Pri- miRNA) (Lee et al., 2004; Jones-Rhoades et al., 2006).Pri-miRNA在DCL1 (DICER-LIKE 1)、HYL1 (HYPONASTIC LEAVES 1)以及SE (SERRATE)等蛋白因子的作用下被切割成具有互补茎环结构的前体miRNA (Precursor miRNA, Pre-miRNA), Pre- miRNA又被加工成miRNA/miRNA*双链体.在HEN1 (HUA ENHANCER 1)的作用下, miRNA/miRNA*双链聚合体的3'端被甲基化修饰, 以免被降解, 稳定的miRNA/miRNA*双链聚合体在HASTY蛋白的作用下被运输至细胞质中(Kurihara and Watanabe, 2004; Li et al., 2005; Park et al., 2005; Jones-Rhoades et al., 2006).在细胞质中, miRNA/miRNA*双链聚合物 ...
... 小RNA作为一种重要的转录后水平调控因子, 已被证明具有很强的组织特异性及细胞特异性(Nogueira et al., 2007, 2009; Ori et al., 2007; Chitwood et al., 2008, 2009; Liu et al., 2009; Wollmann et al., 2010), 并且小RNA还被发现能够在植物中移动并参与植物发育的调控(Pagliarani and Gambino, 2019; Yu et al., 2019).为了探究小RNA在植物体内的组织定位以及运输, 研究人员开发了一系列实验方法.最初是对植物体内Pre-miRNA的组织以及细胞定位进行检测.研究者利用地高辛(Digoxin, DIG)标记的反义RNA探针进行原位杂交(Chen, 2004; Kidner and Timmermans, 2006).该技术也被用于检测植物组织内的成熟miRNA及其靶基因mRNA的表达模式, 从而探究小RNA在植物组织中的运输过程(Chitwood et al., 2009; Carlsbecker et al., 2010), 但传统的体外转录合成探针对低表达量的小RNA灵敏度不高.为了探究低丰度成熟的小RNA在植物体内的表达模式与组织定位, 研究者开发了基于类寡核苷酸衍生物(locked nucleic acid, LNA)修饰探针的体内原位杂交技术(Javelle and Timmermans, 2012).针对Pri- miRNA, 研究者通过MIRNA自身启动子驱动报告基因(如GUS和GFP)来检测Pri-miRNA的转录表达模式(Jung and Park, 2007; Raman et al., 2008).一种基于荧光蛋白(如绿色荧光蛋白GFP)的小RNA传感器系统(通常在荧光蛋白DNA序列的3′非编码区加上小RNA的结合位点, 这样荧光蛋白不表达的地方也就是小RNA表达的地方)也通常被用于检测小RNA在植物中的表达(Nodine and Bartel, 2010).小RNA传感器系统也被用于小RNA运输研究.例如, 研究者利用该技术与原位杂交相结合, 发现拟南芥(Arabidopsis thaliana) miR394能够进行细胞间的短距离运输, Pri-miR394在茎尖分生组织的L1层细胞表达, 成熟的miR394能够运输至L2与L3层细胞, 抑制其靶基因LCR (LEAF CURLING RESPONSIVENESS)的表达, 从而维持干细胞的稳态(Knauer et al., 2013).还有研究者利用该系统发现, 植物中小RNA在细胞间的短距离运输与蛋白质的运输机制不同, 其运输不受胞间连丝状态性质变化的影响(Skopelitis et al., 2018).此外, 除了能在细胞间进行短距离运输, 小RNA也能够在植物组织间进行长距离运输.研究者利用嫁接实验以及对植物的韧皮部汁液进行小RNA测序, 证实大量的小RNA能够从植物地上部运输至地下部并抑制其地下部靶基因的表达, 进而参与生物发育过程以及胁迫响应(Buhtz et al., 2008; Lin et al., 2008; Pant et al., 2008, 2009; Bhogale et al., 2014; Huen et al., 2017).最近, 研究者利用嫁接实验、小RNA测序、降解组测序和5'-RACE实验证明植物中小RNA能够从茎端到根部进行单向运输, 并且运输至根部后能够在转录水平调控基因在根部的表达(Li et al., 2021).利用上述实验技术研究小RNA的运输以及表达模式均存在一定的缺点.例如, miRGFP等依赖荧光蛋白的传感器系统极易在实验过程中出现信号丢失, 无法对小RNA的表达进行精确定量且空间分辨率相对较低; 也有研究证明植物miRNA会诱发形成大量的次级小RNA, 这些次级小RNA能够在细胞间进行移动, 从而极大地影响miRGFP系统实验结果的准确性(Parizotto et al., 2004; Howell et al., 2007; Moissiard et al., 2007).而传统的基于化学发光法的原位杂交技术也存在对于低表达丰度的小RNA检测效果不佳的缺点, 而基于LNA探针的原位杂交虽然解决了这一问题, 但也存在分辨率不高, 无法一次检测多个靶标等缺点.RNA测序这种生物信息学方法则无法精确检测植物miRNA准确的组织定位与细胞定位, 且测序存在一定的偏好性与错误率等. ...
Regulation of ARGONAUTE10 expression enables temporal and spatial precision in axillary meristem initiation in Arabidopsis 1 2020
... 基于RNA FISH改进的LNA探针原位杂交技术与免疫荧光相结合的小RNA荧光原位杂交(small RNA fluorescent in situ hybridization, sRNA-FISH)实验技术能够很好地解决上述问题.RNA-FISH属于荧光原位杂交技术的一种, 用于检测和定位细胞中特定的RNA分子.该方法使用与靶标RNA特异性结合的RNA分子作为探针, 探针上带有半抗原或者荧光素, 利用碱基互补配对原则, 可实现间接或者直接的荧光显色.RNA FISH最初被用于检测动物以及酵母(Saccharomyces spp.)中RNA的表达模式与细胞定位(Levsky and Singer, 2003).而在植物中, FISH技术最先被用于染色体以及染色体中特定基因座位的成像(Lamb et al., 2007; Tirichine et al., 2009; Jiang, 2019), 后来又被研究人员用于指示mRNA在植物体内的组织成像以及亚细胞定位(Bruno et al., 2011, 2015; Himanen et al., 2012; Rozier et al., 2014; Bleckmann and Dresselhaus, 2016; Yang et al., 2017, 2020; Woloszynska et al., 2019).针对小RNA的FISH技术也已广泛应用于动物系统中(Lu and Tsourkas, 2009), 但在植物中的应用还比较少.本文介绍一种地高辛标记的LNA探针与靶标RNA相结合, 再通过免疫反应, 用偶联荧光基团的抗体检测地高辛半抗原, 最后通过超高分辨率激光共聚焦显微镜进行荧光成像, 对生物体内小RNA进行定位、定性或半定量分析的实验技术(Huang et al., 2019).本课题组前期通过大量研究证明, 小RNA能够参与调控植物侧生分生组织的起始发育(Zhang et al., 2018, 2020; Xue et al., 2020), 因此我们以模式植物拟南芥为实验材料, 详细介绍该项实验技术的具体操作流程及注意事项. ...
Spatiotemporal control of axillary meristem formation by interacting transcriptional regulators 1 2018
... 基于RNA FISH改进的LNA探针原位杂交技术与免疫荧光相结合的小RNA荧光原位杂交(small RNA fluorescent in situ hybridization, sRNA-FISH)实验技术能够很好地解决上述问题.RNA-FISH属于荧光原位杂交技术的一种, 用于检测和定位细胞中特定的RNA分子.该方法使用与靶标RNA特异性结合的RNA分子作为探针, 探针上带有半抗原或者荧光素, 利用碱基互补配对原则, 可实现间接或者直接的荧光显色.RNA FISH最初被用于检测动物以及酵母(Saccharomyces spp.)中RNA的表达模式与细胞定位(Levsky and Singer, 2003).而在植物中, FISH技术最先被用于染色体以及染色体中特定基因座位的成像(Lamb et al., 2007; Tirichine et al., 2009; Jiang, 2019), 后来又被研究人员用于指示mRNA在植物体内的组织成像以及亚细胞定位(Bruno et al., 2011, 2015; Himanen et al., 2012; Rozier et al., 2014; Bleckmann and Dresselhaus, 2016; Yang et al., 2017, 2020; Woloszynska et al., 2019).针对小RNA的FISH技术也已广泛应用于动物系统中(Lu and Tsourkas, 2009), 但在植物中的应用还比较少.本文介绍一种地高辛标记的LNA探针与靶标RNA相结合, 再通过免疫反应, 用偶联荧光基团的抗体检测地高辛半抗原, 最后通过超高分辨率激光共聚焦显微镜进行荧光成像, 对生物体内小RNA进行定位、定性或半定量分析的实验技术(Huang et al., 2019).本课题组前期通过大量研究证明, 小RNA能够参与调控植物侧生分生组织的起始发育(Zhang et al., 2018, 2020; Xue et al., 2020), 因此我们以模式植物拟南芥为实验材料, 详细介绍该项实验技术的具体操作流程及注意事项. ...
 ,1,2,*
,1,2,*
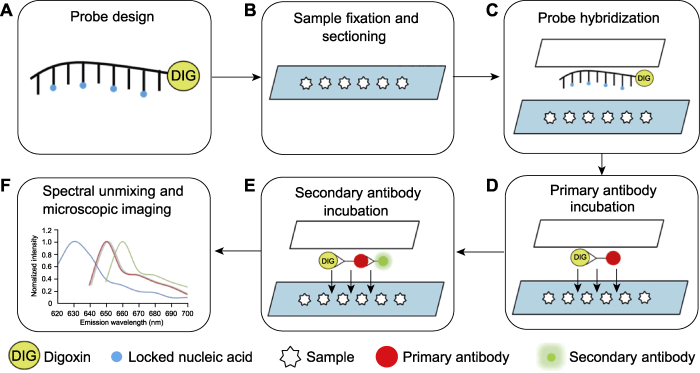 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT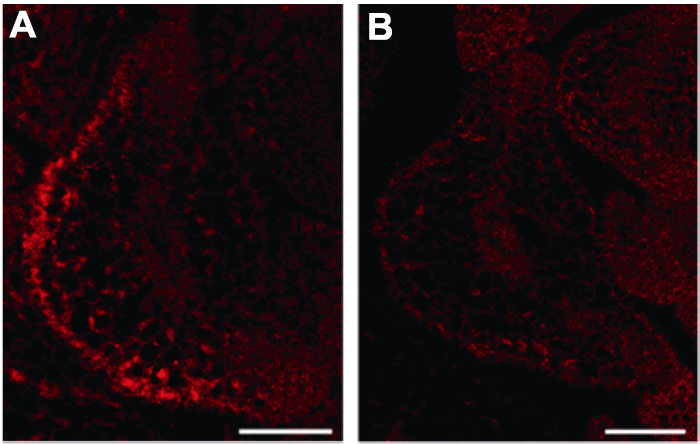 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}