崔晓敏
1 , 2 , 季东超
1 , 2 , 陈彤
, 1 , * , 田世平
1 , 2 1 中国科学院植物研究所, 中国科学院北方资源植物重点实验室, 北京 1000932 中国科学院大学, 北京 100049Advances in the Studies on Molecular Mechanism of Receptor-like Protein Kinase FER Regulating Host Plant-pathogen Interaction Xiaomin Cui
1 , 2 , Dongchao Ji
1 , 2 , Tong Chen
, 1 , * , Shiping Tian
1 , 2 1 Key Laboratory of Plant Resources, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 2 University of Chinese Academy of Sciences, Beijing 100049, China 通讯作者: * E-mail:
chentong@ibcas.ac.cn 责任编辑: 孙冬花
收稿日期: 2020-11-11
接受日期: 2021-02-25
网络出版日期: 2021-05-01
基金资助: 北京市自然科学基金 .6212025 国家重点研发计划No .2017YFD0401301 国家自然科学基金No .31930086 国家自然科学基金No .32072637
Received: 2020-11-11
Accepted: 2021-02-25
Online: 2021-05-01
摘要 植物细胞依赖细胞质膜上的受体感知并传递环境信号, 而受体通过与配体特异结合启动一系列下游信号转导途径, 维持植物正常的生命活动及其对外界环境变化的适应。类受体激酶是其中一类重要受体, 通常由胞外结合结构域、跨膜结构域和胞内激酶结构域3部分组成, 是植物适应外界环境变化的重要调节枢纽。FER属于
Cr RLK1L类受体蛋白激酶家族, 最早被发现在高等植物雌雄配子体识别过程中发挥作用。随后, 众多研究表明, FER在植物生长发育、激素间交互作用、植物与病原菌互作和逆境响应等多种生物学过程中扮演重要角色, 是近年来植物信号通路研究领域的“明星蛋白”。随着植物病理学研究的不断深入, FER在植物与病原菌互作过程中的功能备受关注。该文主要综述FER调节植物与病原菌互作的研究进展, 旨在为进一步解析类受体蛋白激酶在植物细胞响应病原菌侵染过程中的信号转导机制提供参考。
关键词: 类受体激酶 ;
病原菌 ;
信号转导 ;
寄主植物 Abstract Plant cells rely on the receptors on the plasma membrane to sense and transmit environmental signals that mediate by the specific binding of ligands, thus initiating a series of downstream signaling pathways to maintain normal activities of plants and their responses to environmental stimuli. Receptor-like kinases (RLKs) are a group of important receptors composed of extracellular binding domain, transmembrane domain and intracellular kinase domain, which are the important regulatory hubs of plants to respond to environmental stimuli. FERONIA (FER) is a member of
Cr RLK1L receptor protein kinase family, which was firstly identified to play a role in the recognition between male and female game- tes in higher plants. Subsequently, numerous studies have shown that FER plays an important role in various biological processes, such as plant growth and development, hormonal cross-talk, responses to biotic and abiotic stress. FER has become a “star protein” in the studies of plants signaling transduction in recent years. With the intensification of the studies on plant pathology, the functions of FER in the interaction between plants and pathogens have attracted more attention. In this paper, we summarized the advances in understanding the functions of FER in the plants-pathogen interaction. This review will provide a reference for further understanding the mechanisms of receptor-like protein kinases of plant cells in response to pathogen infection.
Keywords: receptor-like kinase ;
pathogen ;
signal transduction ;
host plant PDF (913KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 崔晓敏, 季东超, 陈彤, 田世平 . 类受体激酶FER调节植物与病原菌相互作用的分子机制. 植物学报, 2021,
56 (3): 339-346 doi:10.11983/CBB20180
Cui Xiaomin, Ji Dongchao, Chen Tong, Tian Shiping .
Advances in the Studies on Molecular Mechanism of Receptor-like Protein Kinase FER Regulating Host Plant-pathogen Interaction .
Chinese Bulletin of Botany , 2021,
56 (3): 339-346 doi:10.11983/CBB20180
植物在与外界环境长期互作过程中, 逐渐进化出一套精细的信号响应机制以趋利避害, 适应外界环境的变化。类受体激酶是其中一类非常重要的受体蛋白, 主要通过胞外结合结构域识别信号, 然后将信号传递至细胞质一侧激活胞质激酶结构域, 活化的激酶结构域与下游靶蛋白互作, 启动磷酸化等生化反应, 最终将信号传递到细胞核内, 从而调控下游基因的表达, 实现对环境变化的快速适应。本文主要综述类受体激酶FER (FERONIA)的相关研究进展, 重点关注FER在调节植物与病原菌互作过程中的功能及分子机制, 以期加深我们对受体激酶/类受体激酶调节植物与病原菌互作机制的理解, 并为农作物品质的分子改良提供参考。
1 类受体蛋白激酶简介 蛋白激酶(protein kinases)通过催化蛋白磷酸化反应调节蛋白活性, 广泛参与植物的生长发育、逆境胁迫响应和抗性应答等重要生命过程(
Shiu et al., 2004 ;
Jose et al., 2020 )。植物受体蛋白激酶(receptor protein kinases, RPKs)大多为丝氨酸/苏氨酸蛋白激酶, 能够识别胞外信号, 并将信号传递到胞内, 通过丝/苏氨酸残基的磷酸化/去磷酸化向下游传递(
Tang et al., 2017 ;
朱巍巍等, 2018 )。基因组测序结果显示, 水稻(
Oryza sativa )和拟南芥(
Arabidopsis thaliana )基因组中含有数量众多的受体激酶编码基因, 但其中大多数基因编码产物的天然配体尚未找到, 受体功能也未确定, 因此这些基因编码产物常被称为类受体激酶(receptor-like kinase, RLKs) (
朱巍巍等, 2018 )。RLKs通常由胞外结合结构域、跨膜结构域和胞内激酶结构域3部分组成。不同类受体激酶的胞外结合结构域存在显著差异, 根据其胞外域的差异主要分为6类, 即PR5型(pathogenesis-related protein-5 like receptor kinases)、S-结构域型(S-domain)、类表皮生长因子型(epidermal growth factor like)、类肿瘤坏死因子受体型(tumor-necrosis factor receptor-like)、类凝集素型(lectin-like)和富含亮氨酸重复序列型(leucine-rich repeat) (
朱巍巍等, 2018 )。最新研究表明, 根据受体激酶胞外域的特征基序, 拟南芥跨膜RLKs可进一步分为富含亮氨酸重复序列型(leucine- rich repeat, LRR)、凝集素型(C-lectin和L-lectin)、细胞壁相关激酶(wall-associated kinase, WAK)、类伸展蛋白型(extensin like)、富含脯氨酸类伸展蛋白型(proline-rich extensin like, PERK)、细胞溶解酶基序型(lysin motif, LysM)、类长春花(
Catharanthus rose- like,
Cr RLK)和S-结构域型(self-incompatibility domain, S-domain)等14种(
Jose et al., 2020 )。近年来, 一类胞外结构域中含有Malectin-like基序的类受体激酶(主要包括FER、ANX1 (ANXUR1)、ANX2、THE1 (THESEUS1)和HERK1 (HERCULES1)等)受到广泛关注。其中, FER在植物生长发育、激素互作网络、逆境响应以及植物与病原菌互作等众多生物学过程中发挥重要作用。
2 类受体激酶FER的功能 FER属于长春花类受体激酶家族(
Catharanthus roseus RLK1-like kinases,
Cr RLK1Ls) (
Escobar-Restrepo et al., 2007 ), 最早被发现能够介导雄配子体和雌配子体之间的识别(
Huck et al., 2003 ;
Escobar- Restrepo et al., 2007 )。拟南芥中的FER与该家族的其它16个蛋白在类受体激酶超家族中形成1个分支, 其胞外结构域中的Malectin-like蛋白基序具有结合二聚及寡聚葡萄糖的能力(
Li and Zhang, 2014 ), 暗示该类受体激酶在植物细胞间互作及植物-环境互作过程中发挥重要作用。近年研究表明, FER广泛参与调节高等植物的受精、细胞伸长、生物及非生物胁迫响应等不同的生物学过程(
Chen et al., 2020 ;
Ji et al., 2020b ;
Zhang et al., 2020b )。FER是胚囊接受花粉管所必需, 并且是雌配子体育性的重要调节因子(
Huck et al., 2003 )。拟南芥
fer 突变体中雌配子体对花粉管接收发生功能缺陷, 雌雄配子之间的相互作用被破坏, 导致花粉管生长失控, 不能破裂释放精细胞, 最终导致育性下降(
Huck et al., 2003 )。进一步研究发现, FER定位于丝状器(filiform apparatus), 通过介导雌配子体对花粉管的接收, 触发助细胞内的信号级联反应, 其缺失导致植株生长矮小和精细胞释放受阻(
Escobar-Restrepo et al., 2007 )。
Duan等(2020) 研究表明, FER能控制丝状器中的去酯化果胶水平, 防止多根花粉管进入胚囊。花粉管到达胚珠时会诱导丝状器处产生一氧化氮(nitric oxide, NO), 而这一过程受FER和去酯化果胶共同调节。NO能使前体及其成熟形式的LURE小肽发生硝基化, 从而抑制助细胞分泌LURE。有研究表明, 正常情况下, FER进行依赖网格蛋白和不依赖网格蛋白的内吞作用, 而小肽RALF1 (rapid alkalinization factor 1)处理能刺激CME (clathrin-mediated endocytosis)途径的内吞。阻断CME途径可减弱RALF1对根系生长的抑制, 表明RALF1- FER互作调节植物的生长发育, RALF1则可通过CME途径发挥作用(
Yu et al., 2020 )。
此外, FER还可调节植物许多其它生物学过程。FER通过调节RHO GTPase信号调控根毛发育。例如,
Duan等(2010) 研究表明, 拟南芥
fer-4 突变体植株矮小, 生长缓慢且根毛发育严重缺陷。鸟苷酸交换因子RopGEF4和RopGEF10 (guanine nucleotide exchange factors for ROPs, RopGEF)也参与了该过程的调节(
Huang et al., 2013 )。此外, FER还通过激活蛋白磷酸酶ABI2 (ABA insensitive 2)抑制脱落酸(abscisic acid, ABA)信号通路(
Yu et al., 2012 ); FER、HERCULES1和THESEUS1与油菜素甾醇(brassino- steroids, BRs)途径协同控制细胞伸长(
Guo et al., 2009 )。在暗形态建成的拟南芥幼苗中, FER与乙烯和油菜素甾醇信号通路平行或协同促进下胚轴伸长(
Guo et al., 2009 ;
Deslauriers and Larsen, 2010 ); 且FER能够调节乙烯的生物合成途径, 拟南芥
fer 突变体中积累较高水平的S-腺苷甲硫氨酸(S-adenosyl methionine, SAM)和乙烯, FER通过与S-腺苷甲硫氨酸合成酶互作, 抑制S-腺苷甲硫氨酸和乙烯的生物合成, 导致植株出现矮化表型(
Mao et al., 2015 )。
Ngo等(2014) 和
Shih等(2014) 研究表明, FER还通过调节胞质Ca
2+ 信号响应细胞内外压力信号和盐胁迫。有研究表明, 拟南芥细胞壁中富含亮氨酸重复的细胞伸展蛋白LRXs3/4/5 (leucine-rich repeat extensins3/4/5), 该类蛋白对植物的耐盐性至关重要, 而LRXs、RALFs和FER作为1个功能模块, 通过感受细胞壁完整信号调节植物生长及对盐胁迫的响应(
Zhao et al., 2018 )。FER也可作为细胞壁传感器参与调节细胞间的互作, 控制RAC/ROP GTPase信号, 进而激活RAC/ROPs蛋白, 刺激ROS产生(
Cheung and Wu, 2011 )。
另外, FER在果实成熟过程中也发挥重要作用。在草莓(
Fragaria × ananassa )果实中过表达FER- like蛋白FaMRLK47能延迟草莓果实的成熟, 下调
FaMRLK47 表达则加速果实成熟(
Jia et al., 2017a )。异源表达苹果(
Malus × domestica ) FER-like蛋白MdFERL1/MdFERL6可抑制番茄(
Solanum lycoper-sicum )果实产生乙烯, 并延缓果实成熟, 而MdFERL6在苹果愈伤组织中过表达抑制乙烯产生(
Jia et al., 2017b )。番茄
SlFERL 在果实中表达量较高, 受果实成熟相关转录因子RIN和TAGL1共调控;
Ji等(2020a) 通过表面等离子共振结合质谱分析技术鉴定到若干可能与SlFERL互作的蛋白, SlFERL能与乙烯合成途径的关键酶SlSAMS1互作, 调节乙烯的生物合成, 进而影响果实的成熟进程(
Ji et al., 2020a )。
3 FER在植物与病原菌互作中的作用机制 在病原菌与植物互作过程中, 除植物表面的细胞壁, 病原菌还要突破植物先天免疫系统中的两道屏障——病原相关分子模式诱导的免疫反应(PAMP-triggered immunity, PTI)和效应蛋白诱导的免疫反应(effector- triggered immunity, ETI)。PTI是由模式识别受体(pattern recognition receptors, PRRs)识别病原相关分子模式(pathogen-associated molecular patterns, PAMPs)所引起的防卫反应, 而ETI是由寄主分泌的专化性抗病蛋白(R蛋白)识别病原菌分泌的效应蛋白(effector)引起的特异性免疫反应(
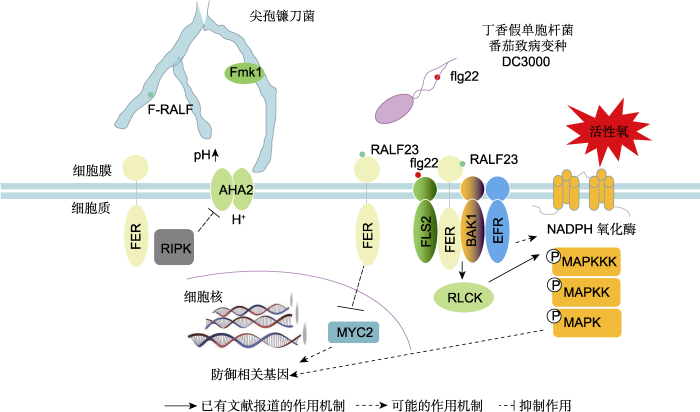
季东超等, 2015 )。FER作为细胞质膜上一类重要的类受体激酶, 可能作为免疫受体复合物中的重要成员参与调节植物与病原菌间的互作(
图1 )。
近年来, FER调节植物与病原菌间互作的报道不断涌现(
表1 )。
Keinath等(2010) 以FLS2的配体flg22处理
15 N/
14 N标记的拟南芥悬浮细胞, 采用定量蛋白质组学技术, 量化flg22刺激后
15 N/
14 N标记的拟南芥细胞质膜的早期反应, 重点分析了DRM (detergent- resistant membranes)组分中蛋白表达水平的变化。 结果表明, 经flg22刺激后, 细胞质膜上DRM组分富集的蛋白种类发生显著变化, 15分钟内64个蛋白在DRM组分中显著富集, 其中转运体和RLKs的富集程度最为显著(包括FLS2 (FLAGELLIN-SENSING 2)和FER)。随后该研究组采用反向遗传学和药理干预方法分析了这些蛋白在flg22诱导的防御反应中的作用, 结果表明, 3个候选突变体(
det3 、
aha1 和
fer4 )在flg22 诱发的活性氧积累、丝裂原活化蛋白激酶(mitogen- activated protein kinase, MAPK)活性和气孔关闭等方面表现出功能缺陷, 暗示这3种蛋白可能是调节植物先天免疫的新组分(
Keinath et al., 2010 )。
Kessler等(2010) 研究发现, FER负调节拟南芥对高氏白粉病菌(
Golovinomyces orontii )的抗性,
fer 纯合突变体对白粉病菌表现出更高的抗性。通过分析高氏白粉病菌孢子穿透细胞表面的比率和每个菌落分生孢子的产量,
Kessler等(2010) 发现, 与野生型植株相比,
fer 纯合突变体对高氏白粉病菌侵染表现出显著抗性, 且高氏白粉病菌的侵入率更低, 每个真菌菌落产生的分生孢子更少。NTA (NORTIA)是最初在对白粉病菌抗性中发现的MLO家族成员,
Kessler等(2010) 研究表明, NTA和FER协同调控白粉病菌的入侵和雌配子体对花粉管的识别, 且乙烯/茉莉酸防御途径可能参与二者介导的抗性。
此外, FER也负调控寄主植物对尖孢镰刀菌(
Fu- sarium oxysporium )的免疫应答。
Masachis等(2016) 将尖孢镰刀菌接种到番茄根部, 分别供给水、pH5.0和pH7.0的缓冲液, 发现在pH5.0条件下, 植物的死亡率相比水和pH7.0缓冲液条件下更低, 说明胞外碱化促进尖孢镰刀菌对寄主植物的侵染。有研究表明, 尖孢镰刀菌中的丝裂原活化蛋白激酶Fmk1对尖孢镰刀菌的侵染至关重要, 胞外pH值从5.0上升至7.0可触发Fmk1快速磷酸化, 激活与致病性相关的MAPK信号级联反应, 从而促进侵入性菌丝生长和病原菌致病(
Masachis et al., 2016 )。进一步研究发现, 尖孢镰刀菌会产生一种类似于植物分泌的小肽F-RALF, 这种小肽与RALF1类似, 可诱导宿主组织碱化, 从而增强尖孢镰刀菌对寄主植物的侵染效率(
Masachis et al., 2016 )。拟南芥突变体
fer4 对尖孢镰刀菌产生的F-RALF不敏感, 与野生型相比其对尖孢镰刀菌菌株表现出更强的抗性, 暗示FER负调控寄主植物对尖孢镰刀菌的免疫应答。该研究揭示了一种植物病原真菌干扰植物正常生理功能的新机制, 即F-RALF与FER结合, 阻断AHA2 (H
+ -ATPase 2)介导的H
+ 外流, 提高根系细胞外pH值, 促进病原菌侵染, 从而负调控免疫反应(
Masachis et al., 2016 )。
质外体区域酸化是引起细胞壁松弛和细胞延展的重要因素。PSY1是一种酪氨酸磺基化的分泌性糖肽, 其受体直接磷酸化并且激活质膜H
+ -ATPase。RALF能抑制质膜H
+ -ATPase活性, 导致质外体区域碱化, 并抑制细胞伸长。PSY1处理能诱导拟南芥根系
RALF33 和
RALFL36 的转录, 并伴随特异性的胞内Ca
2+ 快速爆发, 而Ca
2+ 通道活性的抑制消除了RALF诱导的碱化, 表明Ca
2+ 信号为胞外空间碱化所必需。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1FER介导的抗病相关途径示意图 FER定位于质膜微区, 能够识别尖孢镰刀菌(
Fusarium oxysporium )分泌的F-RALF, 阻断AHA2介导的H
+ 外流, 从而诱导根系胞外环境碱化, 增强真菌对植物的致病力。在此过程中, 尖孢镰刀菌细胞中丝裂原活化蛋白激酶(MAPK)途径中的Fmk1对其侵染至关重要。FER与RALF23相互作用, 负向调控茉莉酸和冠菌素信号, 正向促进植物免疫; 拟南芥(
Arabidopsis thaliana ) SITE-1蛋白酶(S1P)剪切内源快速碱化因子(RALF)前体肽, 而RALF与拟南芥FER相互作用抑制FLS2、BAK1和EFR免疫复合物的形成, 抑制植物免疫。FER可能调节细胞内活性氧的积累和MAPK活性。F-RALF: 尖孢镰刀菌分泌的快速碱化因子; AHA2: H
+ -ATPase 2; RIPK: RESISTANCE TO
Pseudomonas syringae pv.
maculicola 1-INDUCED PROTEIN KINASE; Fmk1: 一种保守的真菌丝裂原活化蛋白激酶(MAPK); flg22: 鞭毛蛋白抗原表位22; RALF23: 内源性肽快速碱化因子23; MYC2: MYELOCYTOMATOSIS PRO- TEINS 2; EFR: ELONGATION FACTOR THERMO UNSTABLE RECEPTOR; FLS2: FLAGELLIN-SENSING 2; BAK1: BRAS- SINOSTEROID INSENSITIVE 1-ASSOCIATED KINASE 1; RLCK: 类受体胞质激酶; MAPK: 丝裂原活化蛋白激酶; NADPH: 烟酰胺腺嘌呤二核苷酸磷酸
Figure 1A schematic diagram of the disease resistance pathway mediated by FER The microdomain-localized FER in the plasma membrane can recognize F-RALF secreted by
Fusarium oxysporium and further block AHA2-mediated H
+ outflow, thus inducing the alkalinization of extracellular environment and increasing the pathogenicity of
F. oxysporium . During this process, Fmk1 in the mitogen-activated protein kinase (MAPK) pathway in
F. oxysporium cells is critical for infection. FER interacts with RALF23, negatively regulating JA and COR signals and positively promoting plant immunity; the SITE-1 protease (S1P) can digest endogenous fast alkalization factor (RALF) precursor peptide and the mature RALF further interacts with FER to inhibit the formation of immune complexes (mainly FLS2, BAK1, and EFR), inhibits plant immunity. FER may regulate intracellular ROS accumulation and MAPK signaling. F-RALF:
F. oxysporium RALF; AHA2: H
+ -ATPase 2; RIPK: RESISTANCE TO
Pseudomonas syringae pv.
maculicola 1-INDUCED PROTEIN KINASE; Fmk1: A con- served fungal MAPK; flg22: Flagellin epitope 22; RALF23: The endogenous peptide RALF 23; MYC2: MYELOCYTOMATOSIS PROTEINS 2; EFR: ELONGATION FACTOR THERMO UNSTABLE RECEPTOR; FLS2: FLAGELLIN-SENSING 2; BAK1: BRASSINOSTEROID INSENSITIVE 1-ASSOCIATED KINASE 1; RLCK: Receptor-like cytoplasmic kinase; MAPK: Mitogenactivated protein kinase; NADPH: Nicotinamide adenine dinucleotide phosphate
Table 1 表1 表1 FER调节植物细胞与病原菌间的相互作用
Table 1
FER regulates the interaction between host plant cells and pathogens 互作蛋白 涉及物种 配体 主要功能/结论 参考文献 FER与病原菌分泌的 拟南芥(Arabidopsis thaliana )和尖孢镰刀菌(Fusarium oxysporium ) F-RALF FER识别尖孢镰刀菌分泌的F- RALF, 阻断质膜H+ -ATPase2诱导胞外环境碱化, 从而负调控拟南芥对尖孢镰刀菌的抗性 Masachis et al., 2016 FER与转录因子MYC2 拟南芥和丁香假单胞杆菌番茄致病变种(Pseudomonas syrin- pv. tomato DC3000) RALF23 FER正调控拟南芥对Pst DC3000 Guo et al., 2018 FER与FLS2、EFR和BAK1形成免疫复合体 拟南芥 RALF23 拟南芥SITE-1蛋白酶剪切内源性快速碱化因子(RALF)前肽, 抑制植物免疫 Stegmann et al., 2017 FER与MLO家族成员NORTIA协同作用 拟南芥和高氏白粉病(Golovinomyces orontii ) FER负调节拟南芥对高氏白粉病的抗性 Kessler et al., 2010 FER与FLS2和BAK1互作 拟南芥和丁香假单胞杆菌番茄致病变种 RALF23 fer 突变体出现氧化爆发、MAPK活性、气孔开合和细胞死亡等方面的功能缺陷Keinath et al., 2010
F-RALF, MYC2, FLS2, EFR, BAK1, RALF23 and MAPK see Figure 1 . F-RALF、MYC2、FLS2、EFR、BAK1、RALF23和MAPK同图1 。 新窗口打开 |
下载CSV 另外, FER也起到类似支架蛋白的作用, 介导植物免疫应答反应。
Stegmann等(2017) 研究发现, 拟南芥SITE-1蛋白酶(S1P)剪切内源性RALF前体肽, FER抑制植物免疫反应。研究表明, 在elf18、flg22和几丁质的作用下,
s1p-3 和
s1p-6 单突变体产生更多的ROS, 对低致病力的丁香假单胞菌番茄致病变种(
Pseudomonas syringae pv.
tomato ) (
Pto ) DC3000
COR- (
coronatine-minus )表现出更高抗性, 因此证明S1P是免疫负调控因子(
Stegmann et al., 2017 )。过表达RALF23能够抑制elf18触发的ROS生成, 增强植株对
Pto DC3000
COR - 和黄瓜枯萎菌
Plectosp- haerella cucumerina 的敏感性, 而RALF23缺失促进elf18触发的ROS产生, 增强植物对
Pto DC3000的抗性, 表明RALF23同样负调控免疫。进一步研究显示,
fer2 和
fer4 突变体对elf18、flg22和几丁质的敏感性较低, 对
Pto DC3000
COR - 更敏感, 说明FER能正向调节免疫过程。flg22处理后, FER与FLS2和BAK1之间的互作增强, 而在
fer4 突变体中, flg22诱导的FLS2- BAK1复合物组装减少,
fer4 /
FER-GFP 转基因株系中复合物的水平恢复正常。
RALF23 过表达对flg22诱导的FLS2-BAK1复合物形成也有类似影响。值得注意的是, RALF23或FER的缺失并不影响FLS2、EFR或BAK1的积累。上述结果表明, FER促进配体诱导的FLS2/EFR与其共受体BAK1之间形成复合物, 而RALF23抑制这一过程, 进一步证实FER是调节免疫受体复合物形成的重要支架组分, 与其受体和共受体共同定位于质膜微区中(
Stegmann et al., 2017 )。这种FER-RALF模块可能通过调节不同的受体激酶复合物, 在生长发育或环境感知等过程中发挥重要作用(
Stegmann et al., 2017 )。
Huang等(2020a) 研究表明, 水稻中类FER受体FLR (FER-like receptor)家族成员能够调节水稻与稻瘟病菌(
Magnaporthe oryzae )互作。他们利用CRISPR/Cas9基因编辑技术成功敲除了FLR家族的14个成员, 结果显示
FLR1 和
FLR13 的突变体对稻瘟病的敏感性增强, 而
FLR2 和
FLR11 的功能缺失增强了水稻对稻瘟病的抗性, 且几乎未对水稻生长造成影响(
Huang et al., 2020a )。
冠菌素(coronatine, COR)是由丁香假单胞菌致病种产生的一种细菌毒素, 在结构和功能上与茉莉酸(JA)-异亮氨酸复合物相似(
Xin and He, 2013 ), 能通过茉莉酸信号途径的关键因子MYC2激活NAC转录因子, 抑制水杨酸积累, 从而破坏植物的免疫应答(
Zheng et al., 2012 )。对
fer 突变体全基因组水平的基因表达分析显示,
fer 突变体中JA和COR信号途径相关的基因表达显著上调,
fer 突变体更易被病原菌侵染。遗传学研究表明, FER通过磷酸化MYC2降低其稳定性, 从而抑制JA和COR信号, 正调控植物的免疫应答。与此相反, RALF23通过与FER互作增强MYC2的稳定性, 从而激活JA信号途径, 实现对植物免疫应答的负调控(
Guo et al., 2018 )。上述结果表明, RALF23-FER-MYC2模块是JA和COR信号途径的关键调控组件, 寄主植物可以利用FER信号途径拮抗COR介导的寄主植物对病原菌的易感性(
Guo et al., 2018 )。
值得注意的是,
Cr RLK1L家族的其它成员在植物与病原菌互作过程中也具有类似功能。LET1 (LETUM- 1)和LET2均是定位于细胞质膜的
Cr RLK1L家族成员, 其中LET1与MEKK2和NLR蛋白SUMM2形成异源三聚体, MEKK2负责维持LET1和SUMM2的稳定性。SUMM2降解依赖CPR1介导的泛素-蛋白酶体降解途径, 而MEKK2能够抑制这一过程并稳定SUMM2蛋白, 从而诱导免疫自激活和细胞死亡; LET2具有类似的功能, 且能够磷酸化LET1。LET1/2蛋白的质膜定位依赖LLG1, 三者共同参与调控SUMM2介导的免疫自激活和细胞死亡途径, 暗示LLG1-
Cr RLK1L模块在植物免疫过程中的功能可能广泛存在且很保守(
Liu et al., 2020 ;
Huang et al., 2020b )。
4 展望 植物与病原菌相互作用的分子机制一直是植物病理学研究的热点和难点。作为细胞膜上重要的类受体激酶, FER在植物信号转导过程中起着关键作用。尽管已有研究对FER的功能及其调控植物免疫应答的机制进行了解析, 但目前仍然存在许多问题有待解决。例如, FER如何向下游组分传递信号? 磷酸化等翻译后修饰过程有何作用? FER的激酶活性是否与其生物功能紧密相关? FER如何平衡生长发育与病原菌侵染响应之间的关系? FER作为支架蛋白招募并维持免疫复合物的形成, 其在不同营养型病原菌中的具体作用机制是否相同? 这些问题的解决将有助于加深我们对受体激酶/类受体激酶调节植物与病原菌互作分子机制的理解, 并为农作物品质的分子改良提供重要参考。
(责任编辑: 孙冬花)
[1] 季东超 , 宋凯 , 邢晶晶 , 陈彤 , 田世平 (2015 ). LysM蛋白介导植物免疫防卫反应及其信号激发的研究进展植物学报 50, 628 -636 . [本文引用: 1] [2] 朱巍巍 , 马天意 , 张梅娟 , 沙伟 (2018 ). 类受体蛋白激酶在植物中的研究进展基因组学与应用生物学 37, 451 -458 . [本文引用: 3] [3] Chen J Liu SR Ming ZH Liu XM Yu F 2020 ). FERONIA cytoplasmic domain: node of varied signal outputsaBIO- TECH 1, 135 -146 . [本文引用: 1] [4] Cheung AY Wu HM 2011 ). Theseus 1, FERONIA and relatives: a family of cell wall-sensing receptor kinases?Curr Opin Plant Biol 14, 632 -641 . DOI:
10.1016/j.pbi.2011.09.001 URL [本文引用: 1] [5] Deslauriers SD Larsen PB 2010 ). FERONIA is a key modulator of brassinosteroid and ethylene responsiveness in Arabidopsis hypocotylsMol Plant 626 -640 . DOI:
10.1093/mp/ssq015 PMID:
20400488 [本文引用: 1] Ethylene signaling is a complex pathway that has been intensively analyzed partly due to its importance to the manifestation of horticultural phenomena, including fruit ripening and tissue senescence. In order to further our understanding of how this pathway is regulated, a screen for Arabidopsis mutants with increased ethylene response was conducted. From this, a mutant was identified as having a dark-grown hypocotyl that is indistinguishable from Col-0 wt in the presence of the ethylene perception inhibitor AgNO?, yet has extreme responsiveness to even low levels of ethylene. Map-based cloning of the mutation revealed a T-DNA insertion in the coding sequence of the receptor-like kinase FERONIA, which is required for normal pollen tube reception and cell elongation in a currently unknown capacity. In contrast to a previous report, analysis of our feronia knockout mutant shows it also has altered responsiveness to brassinosteroids, with etiolated fer-2 seedlings being partially brassinosteroid insensitive with regard to promotion of hypocotyl elongation. Our results indicate that FERONIA-dependent brassinosteroid response serves to antagonize the effect of ethylene on hypocotyl growth of etiolated seedlings, with loss of proper brassinosteroid signaling disrupting this balance and leading to a greater impact of ethylene on hypocotyl shortening.
[6] Duan QH Kita D Li C Cheung AY Wu HM 2010 ). FERONIA receptor-like kinase regulates RHO GTPase signaling of root hair developmentProc Natl Acad Sci USA 107, 17821 -17826 . DOI:
10.1073/pnas.1005366107 URL [本文引用: 1] [7] Duan QL Liu MCJ Kita D Jordan SS Yeh FLJ Yvon R Carpenter H Federico AN Garcia-Valencia LE Eyles SJ Wang CS Wu HM Cheung AY 2020 ). FERONIA controls pectin- and nitric oxide-mediated male-female interactionNature 579, 561 -566 . DOI:
10.1038/s41586-020-2106-2 URL [本文引用: 1] [8] Escobar-Restrepo JM Huck N Kessler S Gagliardini V Gheyselinck J Yang WC Grossniklaus U 2007 ). The FERONIA receptor-like kinase mediates male-female interactions during pollen tube receptionScience 317, 656 -660 . DOI:
10.1126/science.1143562 URL [本文引用: 3] [9] Gjetting SK Mahmood K Shabala L Kristensen A Shabala S Palmgren M Fuglsang AT 2020 ). Evidence for multiple receptors mediating RALF-triggered Ca 2 + sig- naling and proton pump inhibitionPlant J 104, 433 -446 . DOI:
10.1111/tpj.v104.2 URL [10] Guo HQ Li L Ye HX Yu XF Algreen A Yin YH 2009 ). Three related receptor-like kinases are required for optimal cell elongation in Arabidopsis thaliana Proc Natl Acad Sci USA 7648 -7653 . DOI:
10.1073/pnas.0812346106 URL [本文引用: 2] [11] Guo HQ Nolan TM Song GY Liu SZ Xie ZL Chen JN Schnable PS Walley JW Yin YH 2018 ). FERONIA receptor kinase contributes to plant immunity by suppres- sing jasmonic acid signaling in Arabidopsis thaliana Curr Biol 3316 -3324 . DOI:
10.1016/j.cub.2018.07.078 URL [本文引用: 3] [12] Huang GQ Li E Ge FR Li S Wang Q Zhang CQ Zhang Y 2013 ). Arabidopsis RopGEF4 and RopGEF10 are important for FERONIA-mediated developmental but not environmental regulation of root hair growthNew Phytol 200, 1089 -1101 . DOI:
10.1111/nph.2013.200.issue-4 URL [本文引用: 1] [13] Huang YY Liu XX Xie Y Lin XY Hu ZJ Wang H Wang LF Dang WQ Zhang LL Zhu Y Feng H Pu M Zhao JQ Zhang JW Li Y Fan J Wang WM 2020 a). Identification of FERONIA-like receptor genes involved in rice- Magnaporthe oryzae interactionPhytopathol Res 14 . DOI:
10.1186/s42483-020-00052-z URL [本文引用: 2] [14] Huang YY Yin CC Liu J Feng BM Ge DD Kong L Ortiz-Morea FA Richter J Hauser MT Wang WM Shan LB He P 2020 b). A trimeric CrRLK1L-LLG1 complex genetically modulates SUMM2-mediated autoimmunityNat Commun 11, 4859 . DOI:
10.1038/s41467-020-18600-8 URL [本文引用: 1] [15] Huck N Moore JM Federer M Grossniklaus U 2003 ). The Arabidopsis mutant feronia disrupts the female gametophytic control of pollen tube receptionDevelopment 2149 -2159 . DOI:
10.1242/dev.00458 URL [本文引用: 3] [16] Ji DC Chen T Zhang ZQ Li BQ Tian SP 2020 b). Versatile roles of the receptor-like kinase feronia in plant growth, development and host-pathogen interactionInt J Mol Sci 21, 7881 . DOI:
10.3390/ijms21217881 URL [本文引用: 2] [17] Ji DC Cui XM Qin GZ Chen T Tian SP 2020 a). SlFERL interacts with S -adenosylmethionine synthetase to regulate fruit ripeningPlant Physiol 2168 -2181 . DOI:
10.1104/pp.20.01203 URL [本文引用: 1] [18] Jia MR Ding N Zhang Q Xing SN Wei LZ Zhao YY Du P Mao WW Li JZ Li BB Jia WS 2017 a). A FERONIA-like receptor kinase regulates strawberry (Fragaria × ananassa ) fruit ripening and quality formationFront Plant Sci 1099 . DOI:
10.3389/fpls.2017.01099 URL [本文引用: 1] [19] Jia MR Du P Ding N Zhang Q Xing SN Wei LZ Zhao YY Mao WW Li JZ Li BB Jia WS 2017 b). Two FERONIA-like receptor kinases regulate apple fruit ripe- ning by modulating ethylene productionFront Plant Sci 8, 1406 . DOI:
10.3389/fpls.2017.01406 URL [本文引用: 1] [20] Jose J Ghantasala S Choudhury SR 2020 ). Arabidopsis transmembrane receptor-like kinases (RLKs): a bridge between extracellular signal and intracellular regulatory machineryInt J Mol Sci 21, 4000 . DOI:
10.3390/ijms21114000 URL [本文引用: 2] [21] Keinath NF Kierszniowska S Lorek J Bourdais G Kessler SA Shimosato-Asano H Grossniklaus U Schulze WX Robatzek S Panstruga R 2010 ). PAMP (pathogen-associated molecular pattern)-induced changes in plasma membrane compartmentalization reveal novel components of plant immunityJ Biol Chem 285, 39140 -39149 . DOI:
10.1074/jbc.M110.160531 URL [本文引用: 3] [22] Kessler SA Shimosato-Asano H Keinath NF Wuest SE Ingram G Panstruga R Grossniklaus U 2010 ). Conserved molecular components for pollen tube reception and fungal invasionScience 330, 968 -971 . DOI:
10.1126/science.1195211 URL [本文引用: 4] [23] Li S Zhang Y 2014 ). To grow or not to grow: FERONIA has her sayMol Plant 7, 1261 -1263 . DOI:
10.1093/mp/ssu031 URL [本文引用: 1] [24] Liu J Huang YY Kong L Yu X Feng BM Liu DR Zhao BY Mendes GC Yuan PG Ge DD Wang WM Fontes EPB Li PW Shan LB He P 2020 ). The malectin-like receptor-like kinase LETUM1 modulates NLR protein SUMM2 activation via MEKK2 scaffoldingNat Plants 6, 1106 -1115 . DOI:
10.1038/s41477-020-0748-6 URL [本文引用: 1] [25] Mao DD Yu F Li J Van de Poel B Tan D Li JL Liu YQ Li XS Dong MQ Chen LB Li DP Luan S 2015 ). FERONIA receptor kinase interacts with S -adenosylme- thionine synthetase and suppresses S -adenosylme-thio- nine production and ethylene biosynthesis in Arabidopsis Plant Cell Environ 2566 -2574 . DOI:
10.1111/pce.12570 URL [本文引用: 1] [26] Masachis S Segorbe D Turrà D Leon-Ruiz M Fürst U El Ghalid M Leonard G López-Berges MS Richards TA Felix G Di Pietro A 2016 ). A fungal pathogen secretes plant alkalinizing peptides to increase infectionNat Microbiol 1, 16043 . DOI:
10.1038/nmicrobiol.2016.43 PMID:
27572834 [本文引用: 5] Plant infections caused by fungi are often associated with an increase in the pH of the surrounding host tissue(1). Extracellular alkalinization is thought to contribute to fungal pathogenesis, but the underlying mechanisms are poorly understood. Here, we show that the root-infecting fungus Fusarium oxysporum uses a functional homologue of the plant regulatory peptide RALF (rapid alkalinization factor)(2,3) to induce alkalinization and cause disease in plants. An upshift in extracellular pH promotes infectious growth of Fusarium by stimulating phosphorylation of a conserved mitogen-activated protein kinase essential for pathogenicity(4,5). Fungal mutants lacking a functional Fusarium (F)-RALF peptide failed to induce host alkalinization and showed markedly reduced virulence in tomato plants, while eliciting a strong host immune response. Arabidopsis plants lacking the receptor-like kinase FERONIA, which mediates the RALF-triggered alkalinization response(6), displayed enhanced resistance against Fusarium. RALF homologues are found across a number of phylogenetically distant groups of fungi, many of which infect plants. We propose that fungal pathogens use functional homologues of alkalinizing peptides found in their host plants to increase their infectious potential and suppress host immunity.
[27] Ngo QA Vogler H Lituiev DS Nestorova A Grossniklaus U 2014 ). A calcium dialog mediated by the FERONIA signal transduction pathway controls plant sperm deliveryDev Cell 29, 491 -500 . DOI:
10.1016/j.devcel.2014.04.008 URL [本文引用: 1] [28] Shih HW Miller ND Dai C Spalding EP Monshausen GB 2014 ). The receptor-like kinase FERONIA is required for mechanical signal transduction in Arabidopsis seedlingsCurr Biol 1887 -1892 . DOI:
10.1016/j.cub.2014.06.064 URL [本文引用: 1] [29] Shiu SH Karlowski WM Pan RS Tzeng YH Mayer KFX Li WH 2004 ). Comparative analysis of the receptor-like kinase family in Arabidopsis and ricePlant Cell 1220 -1234 . DOI:
10.1105/tpc.020834 URL [本文引用: 1] [30] Stegmann M Monaghan J Smakowska-Luzan E Rove- nich H Lehner A Holton N Bellkhadir Y Zipfel C 2017 ). The receptor kinase FER is a RALF-regulated scaffold controlling plant immune signalingScience 355, 287 -289 . DOI:
10.1126/science.aal2541 URL [本文引用: 5] [31] Tang DZ Wang GX Zhou JM 2017 ). Receptor kinases in plant-pathogen interactions: more than pattern recognitionPlant Cell 29, 618 -637 . DOI:
10.1105/tpc.16.00891 URL [本文引用: 1] [32] Xin XF He SY 2013 ). Pseudomonas syringae pv. tomato DC3000: a model pathogen for probing disease susceptibility and hormone signaling in plantsAnnu Rev Phytopathol 473 -498 . DOI:
10.1146/annurev-phyto-082712-102321 URL [本文引用: 1] [33] Yu F Qian LC Nibau C Duan QH Kita D Levasseur K Li XQ Lu CQ Li H Hou CC Li LG Buchanan BB Chen LB Cheung AY Li DP Luan S 2012 ). FERONIA receptor kinase pathway suppresses abscisic acid signaling in Arabidopsis by activating ABI2 phosphataseProc Natl Acad Sci USA 14693 -14698 . DOI:
10.1073/pnas.1212547109 URL [本文引用: 1] [34] Yu M Li RL Cui YN Chen WJ Li B Zhang X Bu YF Cao YY Xing JJ Jewaria PK Li XJ Bhalerao RP Yu F Lin JX 2020 ). The RALF1-FERONIA interaction modulates endocytosis to mediate control of root growth in Arabidopsis Development dev189902 . [本文引用: 1] [35] Zhang X Peng H Zhu SR Xing JJ Li X Zhu ZZ Zheng JY Wang L Wang BQ Chen J Ming ZH Yao K Jian JZ Luan S Coleman-Derr D Liao HD Peng YS Peng DL Yu F 2020 a). Nematode-encoded RALF peptide mi- mics facilitate parasitism of plants through the FERONIA receptor kinaseMol Plant 13, 1434 -1454 . DOI:
10.1016/j.molp.2020.08.014 URL [本文引用: 1] [36] Zhang X Yang ZH Wu DS Yu F 2020 b). RALF-FERONIA signaling: linking plant immune response with cell growthPlant Commun 1, 100084 . DOI:
10.1016/j.xplc.2020.100084 URL [37] Zhao CZ Zayed O Yu ZP Jiang W Zhu PP Hsu CC Zhang LR Tao WA Lozano-Durán R Zhu JK 2018 ). Leucine-rich repeat extensin proteins regulate plant salt tolerance in Arabidopsis Proc Natl Acad Sci USA 13123 -13128 . DOI:
10.1073/pnas.1816991115 URL [本文引用: 1] [38] Zheng XY Spivey NW Zeng WQ Liu PP Fu ZQ Klessig DF He SY Dong XN 2012 ). Coronatine promotes Pseu- domonas syringae virulence in plants by activating a signaling cascade that inhibits salicylic acid accumulationCell Host Microbe 587 -596 . DOI:
10.1016/j.chom.2012.04.014 URL [本文引用: 1] LysM蛋白介导植物免疫防卫反应及其信号激发的研究进展
1
2015
... 在病原菌与植物互作过程中, 除植物表面的细胞壁, 病原菌还要突破植物先天免疫系统中的两道屏障——病原相关分子模式诱导的免疫反应(PAMP-triggered immunity, PTI)和效应蛋白诱导的免疫反应(effector- triggered immunity, ETI).PTI是由模式识别受体(pattern recognition receptors, PRRs)识别病原相关分子模式(pathogen-associated molecular patterns, PAMPs)所引起的防卫反应, 而ETI是由寄主分泌的专化性抗病蛋白(R蛋白)识别病原菌分泌的效应蛋白(effector)引起的特异性免疫反应(
季东超等, 2015 ).FER作为细胞质膜上一类重要的类受体激酶, 可能作为免疫受体复合物中的重要成员参与调节植物与病原菌间的互作(
图1 ). ...
类受体蛋白激酶在植物中的研究进展
3
2018
... 蛋白激酶(protein kinases)通过催化蛋白磷酸化反应调节蛋白活性, 广泛参与植物的生长发育、逆境胁迫响应和抗性应答等重要生命过程(
Shiu et al., 2004 ;
Jose et al., 2020 ).植物受体蛋白激酶(receptor protein kinases, RPKs)大多为丝氨酸/苏氨酸蛋白激酶, 能够识别胞外信号, 并将信号传递到胞内, 通过丝/苏氨酸残基的磷酸化/去磷酸化向下游传递(
Tang et al., 2017 ;
朱巍巍等, 2018 ).基因组测序结果显示, 水稻(
Oryza sativa )和拟南芥(
Arabidopsis thaliana )基因组中含有数量众多的受体激酶编码基因, 但其中大多数基因编码产物的天然配体尚未找到, 受体功能也未确定, 因此这些基因编码产物常被称为类受体激酶(receptor-like kinase, RLKs) (
朱巍巍等, 2018 ).RLKs通常由胞外结合结构域、跨膜结构域和胞内激酶结构域3部分组成.不同类受体激酶的胞外结合结构域存在显著差异, 根据其胞外域的差异主要分为6类, 即PR5型(pathogenesis-related protein-5 like receptor kinases)、S-结构域型(S-domain)、类表皮生长因子型(epidermal growth factor like)、类肿瘤坏死因子受体型(tumor-necrosis factor receptor-like)、类凝集素型(lectin-like)和富含亮氨酸重复序列型(leucine-rich repeat) (
朱巍巍等, 2018 ).最新研究表明, 根据受体激酶胞外域的特征基序, 拟南芥跨膜RLKs可进一步分为富含亮氨酸重复序列型(leucine- rich repeat, LRR)、凝集素型(C-lectin和L-lectin)、细胞壁相关激酶(wall-associated kinase, WAK)、类伸展蛋白型(extensin like)、富含脯氨酸类伸展蛋白型(proline-rich extensin like, PERK)、细胞溶解酶基序型(lysin motif, LysM)、类长春花(
Catharanthus rose- like,
Cr RLK)和S-结构域型(self-incompatibility domain, S-domain)等14种(
Jose et al., 2020 ).近年来, 一类胞外结构域中含有Malectin-like基序的类受体激酶(主要包括FER、ANX1 (ANXUR1)、ANX2、THE1 (THESEUS1)和HERK1 (HERCULES1)等)受到广泛关注.其中, FER在植物生长发育、激素互作网络、逆境响应以及植物与病原菌互作等众多生物学过程中发挥重要作用. ...
... )基因组中含有数量众多的受体激酶编码基因, 但其中大多数基因编码产物的天然配体尚未找到, 受体功能也未确定, 因此这些基因编码产物常被称为类受体激酶(receptor-like kinase, RLKs) (
朱巍巍等, 2018 ).RLKs通常由胞外结合结构域、跨膜结构域和胞内激酶结构域3部分组成.不同类受体激酶的胞外结合结构域存在显著差异, 根据其胞外域的差异主要分为6类, 即PR5型(pathogenesis-related protein-5 like receptor kinases)、S-结构域型(S-domain)、类表皮生长因子型(epidermal growth factor like)、类肿瘤坏死因子受体型(tumor-necrosis factor receptor-like)、类凝集素型(lectin-like)和富含亮氨酸重复序列型(leucine-rich repeat) (
朱巍巍等, 2018 ).最新研究表明, 根据受体激酶胞外域的特征基序, 拟南芥跨膜RLKs可进一步分为富含亮氨酸重复序列型(leucine- rich repeat, LRR)、凝集素型(C-lectin和L-lectin)、细胞壁相关激酶(wall-associated kinase, WAK)、类伸展蛋白型(extensin like)、富含脯氨酸类伸展蛋白型(proline-rich extensin like, PERK)、细胞溶解酶基序型(lysin motif, LysM)、类长春花(
Catharanthus rose- like,
Cr RLK)和S-结构域型(self-incompatibility domain, S-domain)等14种(
Jose et al., 2020 ).近年来, 一类胞外结构域中含有Malectin-like基序的类受体激酶(主要包括FER、ANX1 (ANXUR1)、ANX2、THE1 (THESEUS1)和HERK1 (HERCULES1)等)受到广泛关注.其中, FER在植物生长发育、激素互作网络、逆境响应以及植物与病原菌互作等众多生物学过程中发挥重要作用. ...
... ).RLKs通常由胞外结合结构域、跨膜结构域和胞内激酶结构域3部分组成.不同类受体激酶的胞外结合结构域存在显著差异, 根据其胞外域的差异主要分为6类, 即PR5型(pathogenesis-related protein-5 like receptor kinases)、S-结构域型(S-domain)、类表皮生长因子型(epidermal growth factor like)、类肿瘤坏死因子受体型(tumor-necrosis factor receptor-like)、类凝集素型(lectin-like)和富含亮氨酸重复序列型(leucine-rich repeat) (
朱巍巍等, 2018 ).最新研究表明, 根据受体激酶胞外域的特征基序, 拟南芥跨膜RLKs可进一步分为富含亮氨酸重复序列型(leucine- rich repeat, LRR)、凝集素型(C-lectin和L-lectin)、细胞壁相关激酶(wall-associated kinase, WAK)、类伸展蛋白型(extensin like)、富含脯氨酸类伸展蛋白型(proline-rich extensin like, PERK)、细胞溶解酶基序型(lysin motif, LysM)、类长春花(
Catharanthus rose- like,
Cr RLK)和S-结构域型(self-incompatibility domain, S-domain)等14种(
Jose et al., 2020 ).近年来, 一类胞外结构域中含有Malectin-like基序的类受体激酶(主要包括FER、ANX1 (ANXUR1)、ANX2、THE1 (THESEUS1)和HERK1 (HERCULES1)等)受到广泛关注.其中, FER在植物生长发育、激素互作网络、逆境响应以及植物与病原菌互作等众多生物学过程中发挥重要作用. ...
FERONIA cytoplasmic domain: node of varied signal outputs
1
2020
... FER属于长春花类受体激酶家族(
Catharanthus roseus RLK1-like kinases,
Cr RLK1Ls) (
Escobar-Restrepo et al., 2007 ), 最早被发现能够介导雄配子体和雌配子体之间的识别(
Huck et al., 2003 ;
Escobar- Restrepo et al., 2007 ).拟南芥中的FER与该家族的其它16个蛋白在类受体激酶超家族中形成1个分支, 其胞外结构域中的Malectin-like蛋白基序具有结合二聚及寡聚葡萄糖的能力(
Li and Zhang, 2014 ), 暗示该类受体激酶在植物细胞间互作及植物-环境互作过程中发挥重要作用.近年研究表明, FER广泛参与调节高等植物的受精、细胞伸长、生物及非生物胁迫响应等不同的生物学过程(
Chen et al., 2020 ;
Ji et al., 2020b ;
Zhang et al., 2020b ).FER是胚囊接受花粉管所必需, 并且是雌配子体育性的重要调节因子(
Huck et al., 2003 ).拟南芥
fer 突变体中雌配子体对花粉管接收发生功能缺陷, 雌雄配子之间的相互作用被破坏, 导致花粉管生长失控, 不能破裂释放精细胞, 最终导致育性下降(
Huck et al., 2003 ).进一步研究发现, FER定位于丝状器(filiform apparatus), 通过介导雌配子体对花粉管的接收, 触发助细胞内的信号级联反应, 其缺失导致植株生长矮小和精细胞释放受阻(
Escobar-Restrepo et al., 2007 ).
Duan等(2020) 研究表明, FER能控制丝状器中的去酯化果胶水平, 防止多根花粉管进入胚囊.花粉管到达胚珠时会诱导丝状器处产生一氧化氮(nitric oxide, NO), 而这一过程受FER和去酯化果胶共同调节.NO能使前体及其成熟形式的LURE小肽发生硝基化, 从而抑制助细胞分泌LURE.有研究表明, 正常情况下, FER进行依赖网格蛋白和不依赖网格蛋白的内吞作用, 而小肽RALF1 (rapid alkalinization factor 1)处理能刺激CME (clathrin-mediated endocytosis)途径的内吞.阻断CME途径可减弱RALF1对根系生长的抑制, 表明RALF1- FER互作调节植物的生长发育, RALF1则可通过CME途径发挥作用(
Yu et al., 2020 ). ...
Theseus 1, FERONIA and relatives: a family of cell wall-sensing receptor kinases?
1
2011
...
Ngo等(2014) 和
Shih等(2014) 研究表明, FER还通过调节胞质Ca
2+ 信号响应细胞内外压力信号和盐胁迫.有研究表明, 拟南芥细胞壁中富含亮氨酸重复的细胞伸展蛋白LRXs3/4/5 (leucine-rich repeat extensins3/4/5), 该类蛋白对植物的耐盐性至关重要, 而LRXs、RALFs和FER作为1个功能模块, 通过感受细胞壁完整信号调节植物生长及对盐胁迫的响应(
Zhao et al., 2018 ).FER也可作为细胞壁传感器参与调节细胞间的互作, 控制RAC/ROP GTPase信号, 进而激活RAC/ROPs蛋白, 刺激ROS产生(
Cheung and Wu, 2011 ). ...
FERONIA is a key modulator of brassinosteroid and ethylene responsiveness in
Arabidopsis hypocotyls
1
2010
... 此外, FER还可调节植物许多其它生物学过程.FER通过调节RHO GTPase信号调控根毛发育.例如,
Duan等(2010) 研究表明, 拟南芥
fer-4 突变体植株矮小, 生长缓慢且根毛发育严重缺陷.鸟苷酸交换因子RopGEF4和RopGEF10 (guanine nucleotide exchange factors for ROPs, RopGEF)也参与了该过程的调节(
Huang et al., 2013 ).此外, FER还通过激活蛋白磷酸酶ABI2 (ABA insensitive 2)抑制脱落酸(abscisic acid, ABA)信号通路(
Yu et al., 2012 ); FER、HERCULES1和THESEUS1与油菜素甾醇(brassino- steroids, BRs)途径协同控制细胞伸长(
Guo et al., 2009 ).在暗形态建成的拟南芥幼苗中, FER与乙烯和油菜素甾醇信号通路平行或协同促进下胚轴伸长(
Guo et al., 2009 ;
Deslauriers and Larsen, 2010 ); 且FER能够调节乙烯的生物合成途径, 拟南芥
fer 突变体中积累较高水平的S-腺苷甲硫氨酸(S-adenosyl methionine, SAM)和乙烯, FER通过与S-腺苷甲硫氨酸合成酶互作, 抑制S-腺苷甲硫氨酸和乙烯的生物合成, 导致植株出现矮化表型(
Mao et al., 2015 ). ...
FERONIA receptor-like kinase regulates RHO GTPase signaling of root hair development
1
2010
... 此外, FER还可调节植物许多其它生物学过程.FER通过调节RHO GTPase信号调控根毛发育.例如,
Duan等(2010) 研究表明, 拟南芥
fer-4 突变体植株矮小, 生长缓慢且根毛发育严重缺陷.鸟苷酸交换因子RopGEF4和RopGEF10 (guanine nucleotide exchange factors for ROPs, RopGEF)也参与了该过程的调节(
Huang et al., 2013 ).此外, FER还通过激活蛋白磷酸酶ABI2 (ABA insensitive 2)抑制脱落酸(abscisic acid, ABA)信号通路(
Yu et al., 2012 ); FER、HERCULES1和THESEUS1与油菜素甾醇(brassino- steroids, BRs)途径协同控制细胞伸长(
Guo et al., 2009 ).在暗形态建成的拟南芥幼苗中, FER与乙烯和油菜素甾醇信号通路平行或协同促进下胚轴伸长(
Guo et al., 2009 ;
Deslauriers and Larsen, 2010 ); 且FER能够调节乙烯的生物合成途径, 拟南芥
fer 突变体中积累较高水平的S-腺苷甲硫氨酸(S-adenosyl methionine, SAM)和乙烯, FER通过与S-腺苷甲硫氨酸合成酶互作, 抑制S-腺苷甲硫氨酸和乙烯的生物合成, 导致植株出现矮化表型(
Mao et al., 2015 ). ...
FERONIA controls pectin- and nitric oxide-mediated male-female interaction
1
2020
... FER属于长春花类受体激酶家族(
Catharanthus roseus RLK1-like kinases,
Cr RLK1Ls) (
Escobar-Restrepo et al., 2007 ), 最早被发现能够介导雄配子体和雌配子体之间的识别(
Huck et al., 2003 ;
Escobar- Restrepo et al., 2007 ).拟南芥中的FER与该家族的其它16个蛋白在类受体激酶超家族中形成1个分支, 其胞外结构域中的Malectin-like蛋白基序具有结合二聚及寡聚葡萄糖的能力(
Li and Zhang, 2014 ), 暗示该类受体激酶在植物细胞间互作及植物-环境互作过程中发挥重要作用.近年研究表明, FER广泛参与调节高等植物的受精、细胞伸长、生物及非生物胁迫响应等不同的生物学过程(
Chen et al., 2020 ;
Ji et al., 2020b ;
Zhang et al., 2020b ).FER是胚囊接受花粉管所必需, 并且是雌配子体育性的重要调节因子(
Huck et al., 2003 ).拟南芥
fer 突变体中雌配子体对花粉管接收发生功能缺陷, 雌雄配子之间的相互作用被破坏, 导致花粉管生长失控, 不能破裂释放精细胞, 最终导致育性下降(
Huck et al., 2003 ).进一步研究发现, FER定位于丝状器(filiform apparatus), 通过介导雌配子体对花粉管的接收, 触发助细胞内的信号级联反应, 其缺失导致植株生长矮小和精细胞释放受阻(
Escobar-Restrepo et al., 2007 ).
Duan等(2020) 研究表明, FER能控制丝状器中的去酯化果胶水平, 防止多根花粉管进入胚囊.花粉管到达胚珠时会诱导丝状器处产生一氧化氮(nitric oxide, NO), 而这一过程受FER和去酯化果胶共同调节.NO能使前体及其成熟形式的LURE小肽发生硝基化, 从而抑制助细胞分泌LURE.有研究表明, 正常情况下, FER进行依赖网格蛋白和不依赖网格蛋白的内吞作用, 而小肽RALF1 (rapid alkalinization factor 1)处理能刺激CME (clathrin-mediated endocytosis)途径的内吞.阻断CME途径可减弱RALF1对根系生长的抑制, 表明RALF1- FER互作调节植物的生长发育, RALF1则可通过CME途径发挥作用(
Yu et al., 2020 ). ...
The FERONIA receptor-like kinase mediates male-female interactions during pollen tube reception
3
2007
... FER属于长春花类受体激酶家族(
Catharanthus roseus RLK1-like kinases,
Cr RLK1Ls) (
Escobar-Restrepo et al., 2007 ), 最早被发现能够介导雄配子体和雌配子体之间的识别(
Huck et al., 2003 ;
Escobar- Restrepo et al., 2007 ).拟南芥中的FER与该家族的其它16个蛋白在类受体激酶超家族中形成1个分支, 其胞外结构域中的Malectin-like蛋白基序具有结合二聚及寡聚葡萄糖的能力(
Li and Zhang, 2014 ), 暗示该类受体激酶在植物细胞间互作及植物-环境互作过程中发挥重要作用.近年研究表明, FER广泛参与调节高等植物的受精、细胞伸长、生物及非生物胁迫响应等不同的生物学过程(
Chen et al., 2020 ;
Ji et al., 2020b ;
Zhang et al., 2020b ).FER是胚囊接受花粉管所必需, 并且是雌配子体育性的重要调节因子(
Huck et al., 2003 ).拟南芥
fer 突变体中雌配子体对花粉管接收发生功能缺陷, 雌雄配子之间的相互作用被破坏, 导致花粉管生长失控, 不能破裂释放精细胞, 最终导致育性下降(
Huck et al., 2003 ).进一步研究发现, FER定位于丝状器(filiform apparatus), 通过介导雌配子体对花粉管的接收, 触发助细胞内的信号级联反应, 其缺失导致植株生长矮小和精细胞释放受阻(
Escobar-Restrepo et al., 2007 ).
Duan等(2020) 研究表明, FER能控制丝状器中的去酯化果胶水平, 防止多根花粉管进入胚囊.花粉管到达胚珠时会诱导丝状器处产生一氧化氮(nitric oxide, NO), 而这一过程受FER和去酯化果胶共同调节.NO能使前体及其成熟形式的LURE小肽发生硝基化, 从而抑制助细胞分泌LURE.有研究表明, 正常情况下, FER进行依赖网格蛋白和不依赖网格蛋白的内吞作用, 而小肽RALF1 (rapid alkalinization factor 1)处理能刺激CME (clathrin-mediated endocytosis)途径的内吞.阻断CME途径可减弱RALF1对根系生长的抑制, 表明RALF1- FER互作调节植物的生长发育, RALF1则可通过CME途径发挥作用(
Yu et al., 2020 ). ...
... ;
Escobar- Restrepo et al., 2007 ).拟南芥中的FER与该家族的其它16个蛋白在类受体激酶超家族中形成1个分支, 其胞外结构域中的Malectin-like蛋白基序具有结合二聚及寡聚葡萄糖的能力(
Li and Zhang, 2014 ), 暗示该类受体激酶在植物细胞间互作及植物-环境互作过程中发挥重要作用.近年研究表明, FER广泛参与调节高等植物的受精、细胞伸长、生物及非生物胁迫响应等不同的生物学过程(
Chen et al., 2020 ;
Ji et al., 2020b ;
Zhang et al., 2020b ).FER是胚囊接受花粉管所必需, 并且是雌配子体育性的重要调节因子(
Huck et al., 2003 ).拟南芥
fer 突变体中雌配子体对花粉管接收发生功能缺陷, 雌雄配子之间的相互作用被破坏, 导致花粉管生长失控, 不能破裂释放精细胞, 最终导致育性下降(
Huck et al., 2003 ).进一步研究发现, FER定位于丝状器(filiform apparatus), 通过介导雌配子体对花粉管的接收, 触发助细胞内的信号级联反应, 其缺失导致植株生长矮小和精细胞释放受阻(
Escobar-Restrepo et al., 2007 ).
Duan等(2020) 研究表明, FER能控制丝状器中的去酯化果胶水平, 防止多根花粉管进入胚囊.花粉管到达胚珠时会诱导丝状器处产生一氧化氮(nitric oxide, NO), 而这一过程受FER和去酯化果胶共同调节.NO能使前体及其成熟形式的LURE小肽发生硝基化, 从而抑制助细胞分泌LURE.有研究表明, 正常情况下, FER进行依赖网格蛋白和不依赖网格蛋白的内吞作用, 而小肽RALF1 (rapid alkalinization factor 1)处理能刺激CME (clathrin-mediated endocytosis)途径的内吞.阻断CME途径可减弱RALF1对根系生长的抑制, 表明RALF1- FER互作调节植物的生长发育, RALF1则可通过CME途径发挥作用(
Yu et al., 2020 ). ...
... ).进一步研究发现, FER定位于丝状器(filiform apparatus), 通过介导雌配子体对花粉管的接收, 触发助细胞内的信号级联反应, 其缺失导致植株生长矮小和精细胞释放受阻(
Escobar-Restrepo et al., 2007 ).
Duan等(2020) 研究表明, FER能控制丝状器中的去酯化果胶水平, 防止多根花粉管进入胚囊.花粉管到达胚珠时会诱导丝状器处产生一氧化氮(nitric oxide, NO), 而这一过程受FER和去酯化果胶共同调节.NO能使前体及其成熟形式的LURE小肽发生硝基化, 从而抑制助细胞分泌LURE.有研究表明, 正常情况下, FER进行依赖网格蛋白和不依赖网格蛋白的内吞作用, 而小肽RALF1 (rapid alkalinization factor 1)处理能刺激CME (clathrin-mediated endocytosis)途径的内吞.阻断CME途径可减弱RALF1对根系生长的抑制, 表明RALF1- FER互作调节植物的生长发育, RALF1则可通过CME途径发挥作用(
Yu et al., 2020 ). ...
Evidence for multiple receptors mediating RALF-triggered Ca
2 + sig- naling and proton pump inhibition
0
2020
Three related receptor-like kinases are required for optimal cell elongation in
Arabidopsis thaliana 2
2009
... 此外, FER还可调节植物许多其它生物学过程.FER通过调节RHO GTPase信号调控根毛发育.例如,
Duan等(2010) 研究表明, 拟南芥
fer-4 突变体植株矮小, 生长缓慢且根毛发育严重缺陷.鸟苷酸交换因子RopGEF4和RopGEF10 (guanine nucleotide exchange factors for ROPs, RopGEF)也参与了该过程的调节(
Huang et al., 2013 ).此外, FER还通过激活蛋白磷酸酶ABI2 (ABA insensitive 2)抑制脱落酸(abscisic acid, ABA)信号通路(
Yu et al., 2012 ); FER、HERCULES1和THESEUS1与油菜素甾醇(brassino- steroids, BRs)途径协同控制细胞伸长(
Guo et al., 2009 ).在暗形态建成的拟南芥幼苗中, FER与乙烯和油菜素甾醇信号通路平行或协同促进下胚轴伸长(
Guo et al., 2009 ;
Deslauriers and Larsen, 2010 ); 且FER能够调节乙烯的生物合成途径, 拟南芥
fer 突变体中积累较高水平的S-腺苷甲硫氨酸(S-adenosyl methionine, SAM)和乙烯, FER通过与S-腺苷甲硫氨酸合成酶互作, 抑制S-腺苷甲硫氨酸和乙烯的生物合成, 导致植株出现矮化表型(
Mao et al., 2015 ). ...
... ).在暗形态建成的拟南芥幼苗中, FER与乙烯和油菜素甾醇信号通路平行或协同促进下胚轴伸长(
Guo et al., 2009 ;
Deslauriers and Larsen, 2010 ); 且FER能够调节乙烯的生物合成途径, 拟南芥
fer 突变体中积累较高水平的S-腺苷甲硫氨酸(S-adenosyl methionine, SAM)和乙烯, FER通过与S-腺苷甲硫氨酸合成酶互作, 抑制S-腺苷甲硫氨酸和乙烯的生物合成, 导致植株出现矮化表型(
Mao et al., 2015 ). ...
FERONIA receptor kinase contributes to plant immunity by suppres- sing jasmonic acid signaling in
Arabidopsis thaliana 3
2018
... FER regulates the interaction between host plant cells and pathogens
Table 1 互作蛋白 涉及物种 配体 主要功能/结论 参考文献 FER与病原菌分泌的 拟南芥(Arabidopsis thaliana )和尖孢镰刀菌(Fusarium oxysporium ) F-RALF FER识别尖孢镰刀菌分泌的F- RALF, 阻断质膜H+ -ATPase2诱导胞外环境碱化, 从而负调控拟南芥对尖孢镰刀菌的抗性 Masachis et al., 2016 FER与转录因子MYC2 拟南芥和丁香假单胞杆菌番茄致病变种(Pseudomonas syrin- pv. tomato DC3000) RALF23 FER正调控拟南芥对Pst DC3000 Guo et al., 2018 FER与FLS2、EFR和BAK1形成免疫复合体 拟南芥 RALF23 拟南芥SITE-1蛋白酶剪切内源性快速碱化因子(RALF)前肽, 抑制植物免疫 Stegmann et al., 2017 FER与MLO家族成员NORTIA协同作用 拟南芥和高氏白粉病(Golovinomyces orontii ) FER负调节拟南芥对高氏白粉病的抗性 Kessler et al., 2010 FER与FLS2和BAK1互作 拟南芥和丁香假单胞杆菌番茄致病变种 RALF23 fer 突变体出现氧化爆发、MAPK活性、气孔开合和细胞死亡等方面的功能缺陷Keinath et al., 2010
F-RALF, MYC2, FLS2, EFR, BAK1, RALF23 and MAPK see Figure 1 . ...Xin and He, 2013 ), 能通过茉莉酸信号途径的关键因子MYC2激活NAC转录因子, 抑制水杨酸积累, 从而破坏植物的免疫应答(Zheng et al., 2012 ).对fer 突变体全基因组水平的基因表达分析显示, fer 突变体中JA和COR信号途径相关的基因表达显著上调, fer 突变体更易被病原菌侵染.遗传学研究表明, FER通过磷酸化MYC2降低其稳定性, 从而抑制JA和COR信号, 正调控植物的免疫应答.与此相反, RALF23通过与FER互作增强MYC2的稳定性, 从而激活JA信号途径, 实现对植物免疫应答的负调控(Guo et al., 2018 ).上述结果表明, RALF23-FER-MYC2模块是JA和COR信号途径的关键调控组件, 寄主植物可以利用FER信号途径拮抗COR介导的寄主植物对病原菌的易感性(Guo et al., 2018 ). ...Guo et al., 2018 ). ...Arabidopsis RopGEF4 and RopGEF10 are important for FERONIA-mediated developmental but not environmental regulation of root hair growthDuan等(2010) 研究表明, 拟南芥fer-4 突变体植株矮小, 生长缓慢且根毛发育严重缺陷.鸟苷酸交换因子RopGEF4和RopGEF10 (guanine nucleotide exchange factors for ROPs, RopGEF)也参与了该过程的调节(Huang et al., 2013 ).此外, FER还通过激活蛋白磷酸酶ABI2 (ABA insensitive 2)抑制脱落酸(abscisic acid, ABA)信号通路(Yu et al., 2012 ); FER、HERCULES1和THESEUS1与油菜素甾醇(brassino- steroids, BRs)途径协同控制细胞伸长(Guo et al., 2009 ).在暗形态建成的拟南芥幼苗中, FER与乙烯和油菜素甾醇信号通路平行或协同促进下胚轴伸长(Guo et al., 2009 ; Deslauriers and Larsen, 2010 ); 且FER能够调节乙烯的生物合成途径, 拟南芥fer 突变体中积累较高水平的S-腺苷甲硫氨酸(S-adenosyl methionine, SAM)和乙烯, FER通过与S-腺苷甲硫氨酸合成酶互作, 抑制S-腺苷甲硫氨酸和乙烯的生物合成, 导致植株出现矮化表型(Mao et al., 2015 ). ...FERONIA-like receptor genes involved in rice- Magnaporthe oryzae interactionStegmann等(2017) 研究发现, 拟南芥SITE-1蛋白酶(S1P)剪切内源性RALF前体肽, FER抑制植物免疫反应.研究表明, 在elf18、flg22和几丁质的作用下, s1p-3 和s1p-6 单突变体产生更多的ROS, 对低致病力的丁香假单胞菌番茄致病变种(Pseudomonas syringae pv. tomato ) (Pto ) DC3000 COR- (coronatine-minus )表现出更高抗性, 因此证明S1P是免疫负调控因子(Stegmann et al., 2017 ).过表达RALF23能够抑制elf18触发的ROS生成, 增强植株对Pto DC3000 COR - 和黄瓜枯萎菌Plectosp- haerella cucumerina 的敏感性, 而RALF23缺失促进elf18触发的ROS产生, 增强植物对Pto DC3000的抗性, 表明RALF23同样负调控免疫.进一步研究显示, fer2 和fer4 突变体对elf18、flg22和几丁质的敏感性较低, 对Pto DC3000 COR - 更敏感, 说明FER能正向调节免疫过程.flg22处理后, FER与FLS2和BAK1之间的互作增强, 而在fer4 突变体中, flg22诱导的FLS2- BAK1复合物组装减少, fer4 /FER-GFP 转基因株系中复合物的水平恢复正常.RALF23 过表达对flg22诱导的FLS2-BAK1复合物形成也有类似影响.值得注意的是, RALF23或FER的缺失并不影响FLS2、EFR或BAK1的积累.上述结果表明, FER促进配体诱导的FLS2/EFR与其共受体BAK1之间形成复合物, 而RALF23抑制这一过程, 进一步证实FER是调节免疫受体复合物形成的重要支架组分, 与其受体和共受体共同定位于质膜微区中(Stegmann et al., 2017 ).这种FER-RALF模块可能通过调节不同的受体激酶复合物, 在生长发育或环境感知等过程中发挥重要作用(Stegmann et al., 2017 ).Huang等(2020a) 研究表明, 水稻中类FER受体FLR (FER-like receptor)家族成员能够调节水稻与稻瘟病菌(Magnaporthe oryzae )互作.他们利用CRISPR/Cas9基因编辑技术成功敲除了FLR家族的14个成员, 结果显示FLR1 和FLR13 的突变体对稻瘟病的敏感性增强, 而FLR2 和FLR11 的功能缺失增强了水稻对稻瘟病的抗性, 且几乎未对水稻生长造成影响(Huang et al., 2020a ). ...Huang et al., 2020a ). ...Cr RLK1L家族的其它成员在植物与病原菌互作过程中也具有类似功能.LET1 (LETUM- 1)和LET2均是定位于细胞质膜的Cr RLK1L家族成员, 其中LET1与MEKK2和NLR蛋白SUMM2形成异源三聚体, MEKK2负责维持LET1和SUMM2的稳定性.SUMM2降解依赖CPR1介导的泛素-蛋白酶体降解途径, 而MEKK2能够抑制这一过程并稳定SUMM2蛋白, 从而诱导免疫自激活和细胞死亡; LET2具有类似的功能, 且能够磷酸化LET1.LET1/2蛋白的质膜定位依赖LLG1, 三者共同参与调控SUMM2介导的免疫自激活和细胞死亡途径, 暗示LLG1-Cr RLK1L模块在植物免疫过程中的功能可能广泛存在且很保守(Liu et al., 2020 ; Huang et al., 2020b ). ...Arabidopsis mutant feronia disrupts the female gametophytic control of pollen tube receptionCatharanthus roseus RLK1-like kinases, Cr RLK1Ls) (Escobar-Restrepo et al., 2007 ), 最早被发现能够介导雄配子体和雌配子体之间的识别(Huck et al., 2003 ; Escobar- Restrepo et al., 2007 ).拟南芥中的FER与该家族的其它16个蛋白在类受体激酶超家族中形成1个分支, 其胞外结构域中的Malectin-like蛋白基序具有结合二聚及寡聚葡萄糖的能力(Li and Zhang, 2014 ), 暗示该类受体激酶在植物细胞间互作及植物-环境互作过程中发挥重要作用.近年研究表明, FER广泛参与调节高等植物的受精、细胞伸长、生物及非生物胁迫响应等不同的生物学过程(Chen et al., 2020 ; Ji et al., 2020b ; Zhang et al., 2020b ).FER是胚囊接受花粉管所必需, 并且是雌配子体育性的重要调节因子(Huck et al., 2003 ).拟南芥fer 突变体中雌配子体对花粉管接收发生功能缺陷, 雌雄配子之间的相互作用被破坏, 导致花粉管生长失控, 不能破裂释放精细胞, 最终导致育性下降(Huck et al., 2003 ).进一步研究发现, FER定位于丝状器(filiform apparatus), 通过介导雌配子体对花粉管的接收, 触发助细胞内的信号级联反应, 其缺失导致植株生长矮小和精细胞释放受阻(Escobar-Restrepo et al., 2007 ).Duan等(2020) 研究表明, FER能控制丝状器中的去酯化果胶水平, 防止多根花粉管进入胚囊.花粉管到达胚珠时会诱导丝状器处产生一氧化氮(nitric oxide, NO), 而这一过程受FER和去酯化果胶共同调节.NO能使前体及其成熟形式的LURE小肽发生硝基化, 从而抑制助细胞分泌LURE.有研究表明, 正常情况下, FER进行依赖网格蛋白和不依赖网格蛋白的内吞作用, 而小肽RALF1 (rapid alkalinization factor 1)处理能刺激CME (clathrin-mediated endocytosis)途径的内吞.阻断CME途径可减弱RALF1对根系生长的抑制, 表明RALF1- FER互作调节植物的生长发育, RALF1则可通过CME途径发挥作用(Yu et al., 2020 ). ...Huck et al., 2003 ).拟南芥fer 突变体中雌配子体对花粉管接收发生功能缺陷, 雌雄配子之间的相互作用被破坏, 导致花粉管生长失控, 不能破裂释放精细胞, 最终导致育性下降(Huck et al., 2003 ).进一步研究发现, FER定位于丝状器(filiform apparatus), 通过介导雌配子体对花粉管的接收, 触发助细胞内的信号级联反应, 其缺失导致植株生长矮小和精细胞释放受阻(Escobar-Restrepo et al., 2007 ).Duan等(2020) 研究表明, FER能控制丝状器中的去酯化果胶水平, 防止多根花粉管进入胚囊.花粉管到达胚珠时会诱导丝状器处产生一氧化氮(nitric oxide, NO), 而这一过程受FER和去酯化果胶共同调节.NO能使前体及其成熟形式的LURE小肽发生硝基化, 从而抑制助细胞分泌LURE.有研究表明, 正常情况下, FER进行依赖网格蛋白和不依赖网格蛋白的内吞作用, 而小肽RALF1 (rapid alkalinization factor 1)处理能刺激CME (clathrin-mediated endocytosis)途径的内吞.阻断CME途径可减弱RALF1对根系生长的抑制, 表明RALF1- FER互作调节植物的生长发育, RALF1则可通过CME途径发挥作用(Yu et al., 2020 ). ...Huck et al., 2003 ).进一步研究发现, FER定位于丝状器(filiform apparatus), 通过介导雌配子体对花粉管的接收, 触发助细胞内的信号级联反应, 其缺失导致植株生长矮小和精细胞释放受阻(Escobar-Restrepo et al., 2007 ).Duan等(2020) 研究表明, FER能控制丝状器中的去酯化果胶水平, 防止多根花粉管进入胚囊.花粉管到达胚珠时会诱导丝状器处产生一氧化氮(nitric oxide, NO), 而这一过程受FER和去酯化果胶共同调节.NO能使前体及其成熟形式的LURE小肽发生硝基化, 从而抑制助细胞分泌LURE.有研究表明, 正常情况下, FER进行依赖网格蛋白和不依赖网格蛋白的内吞作用, 而小肽RALF1 (rapid alkalinization factor 1)处理能刺激CME (clathrin-mediated endocytosis)途径的内吞.阻断CME途径可减弱RALF1对根系生长的抑制, 表明RALF1- FER互作调节植物的生长发育, RALF1则可通过CME途径发挥作用(Yu et al., 2020 ). ...Fragaria × ananassa )果实中过表达FER- like蛋白FaMRLK47能延迟草莓果实的成熟, 下调FaMRLK47 表达则加速果实成熟(Jia et al., 2017a ).异源表达苹果(Malus × domestica ) FER-like蛋白MdFERL1/MdFERL6可抑制番茄(Solanum lycoper-sicum )果实产生乙烯, 并延缓果实成熟, 而MdFERL6在苹果愈伤组织中过表达抑制乙烯产生(Jia et al., 2017b ).番茄SlFERL 在果实中表达量较高, 受果实成熟相关转录因子RIN和TAGL1共调控; Ji等(2020a) 通过表面等离子共振结合质谱分析技术鉴定到若干可能与SlFERL互作的蛋白, SlFERL能与乙烯合成途径的关键酶SlSAMS1互作, 调节乙烯的生物合成, 进而影响果实的成熟进程(Ji et al., 2020a ). ...Ji et al., 2020a ). ...S -adenosylmethionine synthetase to regulate fruit ripeningCatharanthus roseus RLK1-like kinases, Cr RLK1Ls) (Escobar-Restrepo et al., 2007 ), 最早被发现能够介导雄配子体和雌配子体之间的识别(Huck et al., 2003 ; Escobar- Restrepo et al., 2007 ).拟南芥中的FER与该家族的其它16个蛋白在类受体激酶超家族中形成1个分支, 其胞外结构域中的Malectin-like蛋白基序具有结合二聚及寡聚葡萄糖的能力(Li and Zhang, 2014 ), 暗示该类受体激酶在植物细胞间互作及植物-环境互作过程中发挥重要作用.近年研究表明, FER广泛参与调节高等植物的受精、细胞伸长、生物及非生物胁迫响应等不同的生物学过程(Chen et al., 2020 ; Ji et al., 2020b ; Zhang et al., 2020b ).FER是胚囊接受花粉管所必需, 并且是雌配子体育性的重要调节因子(Huck et al., 2003 ).拟南芥fer 突变体中雌配子体对花粉管接收发生功能缺陷, 雌雄配子之间的相互作用被破坏, 导致花粉管生长失控, 不能破裂释放精细胞, 最终导致育性下降(Huck et al., 2003 ).进一步研究发现, FER定位于丝状器(filiform apparatus), 通过介导雌配子体对花粉管的接收, 触发助细胞内的信号级联反应, 其缺失导致植株生长矮小和精细胞释放受阻(Escobar-Restrepo et al., 2007 ).Duan等(2020) 研究表明, FER能控制丝状器中的去酯化果胶水平, 防止多根花粉管进入胚囊.花粉管到达胚珠时会诱导丝状器处产生一氧化氮(nitric oxide, NO), 而这一过程受FER和去酯化果胶共同调节.NO能使前体及其成熟形式的LURE小肽发生硝基化, 从而抑制助细胞分泌LURE.有研究表明, 正常情况下, FER进行依赖网格蛋白和不依赖网格蛋白的内吞作用, 而小肽RALF1 (rapid alkalinization factor 1)处理能刺激CME (clathrin-mediated endocytosis)途径的内吞.阻断CME途径可减弱RALF1对根系生长的抑制, 表明RALF1- FER互作调节植物的生长发育, RALF1则可通过CME途径发挥作用(Yu et al., 2020 ). ...Fragaria × ananassa ) fruit ripening and quality formationFragaria × ananassa )果实中过表达FER- like蛋白FaMRLK47能延迟草莓果实的成熟, 下调FaMRLK47 表达则加速果实成熟(Jia et al., 2017a ).异源表达苹果(Malus × domestica ) FER-like蛋白MdFERL1/MdFERL6可抑制番茄(Solanum lycoper-sicum )果实产生乙烯, 并延缓果实成熟, 而MdFERL6在苹果愈伤组织中过表达抑制乙烯产生(Jia et al., 2017b ).番茄SlFERL 在果实中表达量较高, 受果实成熟相关转录因子RIN和TAGL1共调控; Ji等(2020a) 通过表面等离子共振结合质谱分析技术鉴定到若干可能与SlFERL互作的蛋白, SlFERL能与乙烯合成途径的关键酶SlSAMS1互作, 调节乙烯的生物合成, 进而影响果实的成熟进程(Ji et al., 2020a ). ...Fragaria × ananassa )果实中过表达FER- like蛋白FaMRLK47能延迟草莓果实的成熟, 下调FaMRLK47 表达则加速果实成熟(Jia et al., 2017a ).异源表达苹果(Malus × domestica ) FER-like蛋白MdFERL1/MdFERL6可抑制番茄(Solanum lycoper-sicum )果实产生乙烯, 并延缓果实成熟, 而MdFERL6在苹果愈伤组织中过表达抑制乙烯产生(Jia et al., 2017b ).番茄SlFERL 在果实中表达量较高, 受果实成熟相关转录因子RIN和TAGL1共调控; Ji等(2020a) 通过表面等离子共振结合质谱分析技术鉴定到若干可能与SlFERL互作的蛋白, SlFERL能与乙烯合成途径的关键酶SlSAMS1互作, 调节乙烯的生物合成, 进而影响果实的成熟进程(Ji et al., 2020a ). ...Arabidopsis transmembrane receptor-like kinases (RLKs): a bridge between extracellular signal and intracellular regulatory machineryShiu et al., 2004 ; Jose et al., 2020 ).植物受体蛋白激酶(receptor protein kinases, RPKs)大多为丝氨酸/苏氨酸蛋白激酶, 能够识别胞外信号, 并将信号传递到胞内, 通过丝/苏氨酸残基的磷酸化/去磷酸化向下游传递(Tang et al., 2017 ; 朱巍巍等, 2018 ).基因组测序结果显示, 水稻(Oryza sativa )和拟南芥(Arabidopsis thaliana )基因组中含有数量众多的受体激酶编码基因, 但其中大多数基因编码产物的天然配体尚未找到, 受体功能也未确定, 因此这些基因编码产物常被称为类受体激酶(receptor-like kinase, RLKs) (朱巍巍等, 2018 ).RLKs通常由胞外结合结构域、跨膜结构域和胞内激酶结构域3部分组成.不同类受体激酶的胞外结合结构域存在显著差异, 根据其胞外域的差异主要分为6类, 即PR5型(pathogenesis-related protein-5 like receptor kinases)、S-结构域型(S-domain)、类表皮生长因子型(epidermal growth factor like)、类肿瘤坏死因子受体型(tumor-necrosis factor receptor-like)、类凝集素型(lectin-like)和富含亮氨酸重复序列型(leucine-rich repeat) (朱巍巍等, 2018 ).最新研究表明, 根据受体激酶胞外域的特征基序, 拟南芥跨膜RLKs可进一步分为富含亮氨酸重复序列型(leucine- rich repeat, LRR)、凝集素型(C-lectin和L-lectin)、细胞壁相关激酶(wall-associated kinase, WAK)、类伸展蛋白型(extensin like)、富含脯氨酸类伸展蛋白型(proline-rich extensin like, PERK)、细胞溶解酶基序型(lysin motif, LysM)、类长春花(Catharanthus rose- like, Cr RLK)和S-结构域型(self-incompatibility domain, S-domain)等14种(Jose et al., 2020 ).近年来, 一类胞外结构域中含有Malectin-like基序的类受体激酶(主要包括FER、ANX1 (ANXUR1)、ANX2、THE1 (THESEUS1)和HERK1 (HERCULES1)等)受到广泛关注.其中, FER在植物生长发育、激素互作网络、逆境响应以及植物与病原菌互作等众多生物学过程中发挥重要作用. ...Jose et al., 2020 ).近年来, 一类胞外结构域中含有Malectin-like基序的类受体激酶(主要包括FER、ANX1 (ANXUR1)、ANX2、THE1 (THESEUS1)和HERK1 (HERCULES1)等)受到广泛关注.其中, FER在植物生长发育、激素互作网络、逆境响应以及植物与病原菌互作等众多生物学过程中发挥重要作用. ...表1 ).Keinath等(2010) 以FLS2的配体flg22处理15 N/14 N标记的拟南芥悬浮细胞, 采用定量蛋白质组学技术, 量化flg22刺激后15 N/14 N标记的拟南芥细胞质膜的早期反应, 重点分析了DRM (detergent- resistant membranes)组分中蛋白表达水平的变化. 结果表明, 经flg22刺激后, 细胞质膜上DRM组分富集的蛋白种类发生显著变化, 15分钟内64个蛋白在DRM组分中显著富集, 其中转运体和RLKs的富集程度最为显著(包括FLS2 (FLAGELLIN-SENSING 2)和FER).随后该研究组采用反向遗传学和药理干预方法分析了这些蛋白在flg22诱导的防御反应中的作用, 结果表明, 3个候选突变体(det3 、aha1 和fer4 )在flg22 诱发的活性氧积累、丝裂原活化蛋白激酶(mitogen- activated protein kinase, MAPK)活性和气孔关闭等方面表现出功能缺陷, 暗示这3种蛋白可能是调节植物先天免疫的新组分(Keinath et al., 2010 ). ...Keinath et al., 2010 ). ...Table 1 互作蛋白 涉及物种 配体 主要功能/结论 参考文献 FER与病原菌分泌的 拟南芥(Arabidopsis thaliana )和尖孢镰刀菌(Fusarium oxysporium ) F-RALF FER识别尖孢镰刀菌分泌的F- RALF, 阻断质膜H+ -ATPase2诱导胞外环境碱化, 从而负调控拟南芥对尖孢镰刀菌的抗性 Masachis et al., 2016 FER与转录因子MYC2 拟南芥和丁香假单胞杆菌番茄致病变种(Pseudomonas syrin- pv. tomato DC3000) RALF23 FER正调控拟南芥对Pst DC3000 Guo et al., 2018 FER与FLS2、EFR和BAK1形成免疫复合体 拟南芥 RALF23 拟南芥SITE-1蛋白酶剪切内源性快速碱化因子(RALF)前肽, 抑制植物免疫 Stegmann et al., 2017 FER与MLO家族成员NORTIA协同作用 拟南芥和高氏白粉病(Golovinomyces orontii ) FER负调节拟南芥对高氏白粉病的抗性 Kessler et al., 2010 FER与FLS2和BAK1互作 拟南芥和丁香假单胞杆菌番茄致病变种 RALF23 fer 突变体出现氧化爆发、MAPK活性、气孔开合和细胞死亡等方面的功能缺陷Keinath et al., 2010
F-RALF, MYC2, FLS2, EFR, BAK1, RALF23 and MAPK see Figure 1 . ...Kessler等(2010) 研究发现, FER负调节拟南芥对高氏白粉病菌(Golovinomyces orontii )的抗性, fer 纯合突变体对白粉病菌表现出更高的抗性.通过分析高氏白粉病菌孢子穿透细胞表面的比率和每个菌落分生孢子的产量, Kessler等(2010) 发现, 与野生型植株相比, fer 纯合突变体对高氏白粉病菌侵染表现出显著抗性, 且高氏白粉病菌的侵入率更低, 每个真菌菌落产生的分生孢子更少.NTA (NORTIA)是最初在对白粉病菌抗性中发现的MLO家族成员, Kessler等(2010) 研究表明, NTA和FER协同调控白粉病菌的入侵和雌配子体对花粉管的识别, 且乙烯/茉莉酸防御途径可能参与二者介导的抗性. ...Kessler等(2010) 发现, 与野生型植株相比, fer 纯合突变体对高氏白粉病菌侵染表现出显著抗性, 且高氏白粉病菌的侵入率更低, 每个真菌菌落产生的分生孢子更少.NTA (NORTIA)是最初在对白粉病菌抗性中发现的MLO家族成员, Kessler等(2010) 研究表明, NTA和FER协同调控白粉病菌的入侵和雌配子体对花粉管的识别, 且乙烯/茉莉酸防御途径可能参与二者介导的抗性. ...Kessler等(2010) 研究表明, NTA和FER协同调控白粉病菌的入侵和雌配子体对花粉管的识别, 且乙烯/茉莉酸防御途径可能参与二者介导的抗性. ...Table 1 互作蛋白 涉及物种 配体 主要功能/结论 参考文献 FER与病原菌分泌的 拟南芥(Arabidopsis thaliana )和尖孢镰刀菌(Fusarium oxysporium ) F-RALF FER识别尖孢镰刀菌分泌的F- RALF, 阻断质膜H+ -ATPase2诱导胞外环境碱化, 从而负调控拟南芥对尖孢镰刀菌的抗性 Masachis et al., 2016 FER与转录因子MYC2 拟南芥和丁香假单胞杆菌番茄致病变种(Pseudomonas syrin- pv. tomato DC3000) RALF23 FER正调控拟南芥对Pst DC3000 Guo et al., 2018 FER与FLS2、EFR和BAK1形成免疫复合体 拟南芥 RALF23 拟南芥SITE-1蛋白酶剪切内源性快速碱化因子(RALF)前肽, 抑制植物免疫 Stegmann et al., 2017 FER与MLO家族成员NORTIA协同作用 拟南芥和高氏白粉病(Golovinomyces orontii ) FER负调节拟南芥对高氏白粉病的抗性 Kessler et al., 2010 FER与FLS2和BAK1互作 拟南芥和丁香假单胞杆菌番茄致病变种 RALF23 fer 突变体出现氧化爆发、MAPK活性、气孔开合和细胞死亡等方面的功能缺陷Keinath et al., 2010
F-RALF, MYC2, FLS2, EFR, BAK1, RALF23 and MAPK see Figure 1 . ...Catharanthus roseus RLK1-like kinases, Cr RLK1Ls) (Escobar-Restrepo et al., 2007 ), 最早被发现能够介导雄配子体和雌配子体之间的识别(Huck et al., 2003 ; Escobar- Restrepo et al., 2007 ).拟南芥中的FER与该家族的其它16个蛋白在类受体激酶超家族中形成1个分支, 其胞外结构域中的Malectin-like蛋白基序具有结合二聚及寡聚葡萄糖的能力(Li and Zhang, 2014 ), 暗示该类受体激酶在植物细胞间互作及植物-环境互作过程中发挥重要作用.近年研究表明, FER广泛参与调节高等植物的受精、细胞伸长、生物及非生物胁迫响应等不同的生物学过程(Chen et al., 2020 ; Ji et al., 2020b ; Zhang et al., 2020b ).FER是胚囊接受花粉管所必需, 并且是雌配子体育性的重要调节因子(Huck et al., 2003 ).拟南芥fer 突变体中雌配子体对花粉管接收发生功能缺陷, 雌雄配子之间的相互作用被破坏, 导致花粉管生长失控, 不能破裂释放精细胞, 最终导致育性下降(Huck et al., 2003 ).进一步研究发现, FER定位于丝状器(filiform apparatus), 通过介导雌配子体对花粉管的接收, 触发助细胞内的信号级联反应, 其缺失导致植株生长矮小和精细胞释放受阻(Escobar-Restrepo et al., 2007 ).Duan等(2020) 研究表明, FER能控制丝状器中的去酯化果胶水平, 防止多根花粉管进入胚囊.花粉管到达胚珠时会诱导丝状器处产生一氧化氮(nitric oxide, NO), 而这一过程受FER和去酯化果胶共同调节.NO能使前体及其成熟形式的LURE小肽发生硝基化, 从而抑制助细胞分泌LURE.有研究表明, 正常情况下, FER进行依赖网格蛋白和不依赖网格蛋白的内吞作用, 而小肽RALF1 (rapid alkalinization factor 1)处理能刺激CME (clathrin-mediated endocytosis)途径的内吞.阻断CME途径可减弱RALF1对根系生长的抑制, 表明RALF1- FER互作调节植物的生长发育, RALF1则可通过CME途径发挥作用(Yu et al., 2020 ). ...Cr RLK1L家族的其它成员在植物与病原菌互作过程中也具有类似功能.LET1 (LETUM- 1)和LET2均是定位于细胞质膜的Cr RLK1L家族成员, 其中LET1与MEKK2和NLR蛋白SUMM2形成异源三聚体, MEKK2负责维持LET1和SUMM2的稳定性.SUMM2降解依赖CPR1介导的泛素-蛋白酶体降解途径, 而MEKK2能够抑制这一过程并稳定SUMM2蛋白, 从而诱导免疫自激活和细胞死亡; LET2具有类似的功能, 且能够磷酸化LET1.LET1/2蛋白的质膜定位依赖LLG1, 三者共同参与调控SUMM2介导的免疫自激活和细胞死亡途径, 暗示LLG1-Cr RLK1L模块在植物免疫过程中的功能可能广泛存在且很保守(Liu et al., 2020 ; Huang et al., 2020b ). ...S -adenosylme- thionine synthetase and suppresses S -adenosylme-thio- nine production and ethylene biosynthesis in Arabidopsis Duan等(2010) 研究表明, 拟南芥fer-4 突变体植株矮小, 生长缓慢且根毛发育严重缺陷.鸟苷酸交换因子RopGEF4和RopGEF10 (guanine nucleotide exchange factors for ROPs, RopGEF)也参与了该过程的调节(Huang et al., 2013 ).此外, FER还通过激活蛋白磷酸酶ABI2 (ABA insensitive 2)抑制脱落酸(abscisic acid, ABA)信号通路(Yu et al., 2012 ); FER、HERCULES1和THESEUS1与油菜素甾醇(brassino- steroids, BRs)途径协同控制细胞伸长(Guo et al., 2009 ).在暗形态建成的拟南芥幼苗中, FER与乙烯和油菜素甾醇信号通路平行或协同促进下胚轴伸长(Guo et al., 2009 ; Deslauriers and Larsen, 2010 ); 且FER能够调节乙烯的生物合成途径, 拟南芥fer 突变体中积累较高水平的S-腺苷甲硫氨酸(S-adenosyl methionine, SAM)和乙烯, FER通过与S-腺苷甲硫氨酸合成酶互作, 抑制S-腺苷甲硫氨酸和乙烯的生物合成, 导致植株出现矮化表型(Mao et al., 2015 ). ...Fu- sarium oxysporium )的免疫应答.Masachis等(2016) 将尖孢镰刀菌接种到番茄根部, 分别供给水、pH5.0和pH7.0的缓冲液, 发现在pH5.0条件下, 植物的死亡率相比水和pH7.0缓冲液条件下更低, 说明胞外碱化促进尖孢镰刀菌对寄主植物的侵染.有研究表明, 尖孢镰刀菌中的丝裂原活化蛋白激酶Fmk1对尖孢镰刀菌的侵染至关重要, 胞外pH值从5.0上升至7.0可触发Fmk1快速磷酸化, 激活与致病性相关的MAPK信号级联反应, 从而促进侵入性菌丝生长和病原菌致病(Masachis et al., 2016 ).进一步研究发现, 尖孢镰刀菌会产生一种类似于植物分泌的小肽F-RALF, 这种小肽与RALF1类似, 可诱导宿主组织碱化, 从而增强尖孢镰刀菌对寄主植物的侵染效率(Masachis et al., 2016 ).拟南芥突变体fer4 对尖孢镰刀菌产生的F-RALF不敏感, 与野生型相比其对尖孢镰刀菌菌株表现出更强的抗性, 暗示FER负调控寄主植物对尖孢镰刀菌的免疫应答.该研究揭示了一种植物病原真菌干扰植物正常生理功能的新机制, 即F-RALF与FER结合, 阻断AHA2 (H+ -ATPase 2)介导的H+ 外流, 提高根系细胞外pH值, 促进病原菌侵染, 从而负调控免疫反应(Masachis et al., 2016 ). ...Masachis et al., 2016 ).进一步研究发现, 尖孢镰刀菌会产生一种类似于植物分泌的小肽F-RALF, 这种小肽与RALF1类似, 可诱导宿主组织碱化, 从而增强尖孢镰刀菌对寄主植物的侵染效率(Masachis et al., 2016 ).拟南芥突变体fer4 对尖孢镰刀菌产生的F-RALF不敏感, 与野生型相比其对尖孢镰刀菌菌株表现出更强的抗性, 暗示FER负调控寄主植物对尖孢镰刀菌的免疫应答.该研究揭示了一种植物病原真菌干扰植物正常生理功能的新机制, 即F-RALF与FER结合, 阻断AHA2 (H+ -ATPase 2)介导的H+ 外流, 提高根系细胞外pH值, 促进病原菌侵染, 从而负调控免疫反应(Masachis et al., 2016 ). ...Masachis et al., 2016 ).拟南芥突变体fer4 对尖孢镰刀菌产生的F-RALF不敏感, 与野生型相比其对尖孢镰刀菌菌株表现出更强的抗性, 暗示FER负调控寄主植物对尖孢镰刀菌的免疫应答.该研究揭示了一种植物病原真菌干扰植物正常生理功能的新机制, 即F-RALF与FER结合, 阻断AHA2 (H+ -ATPase 2)介导的H+ 外流, 提高根系细胞外pH值, 促进病原菌侵染, 从而负调控免疫反应(Masachis et al., 2016 ). ...Masachis et al., 2016 ). ...Table 1 互作蛋白 涉及物种 配体 主要功能/结论 参考文献 FER与病原菌分泌的 拟南芥(Arabidopsis thaliana )和尖孢镰刀菌(Fusarium oxysporium ) F-RALF FER识别尖孢镰刀菌分泌的F- RALF, 阻断质膜H+ -ATPase2诱导胞外环境碱化, 从而负调控拟南芥对尖孢镰刀菌的抗性 Masachis et al., 2016 FER与转录因子MYC2 拟南芥和丁香假单胞杆菌番茄致病变种(Pseudomonas syrin- pv. tomato DC3000) RALF23 FER正调控拟南芥对Pst DC3000 Guo et al., 2018 FER与FLS2、EFR和BAK1形成免疫复合体 拟南芥 RALF23 拟南芥SITE-1蛋白酶剪切内源性快速碱化因子(RALF)前肽, 抑制植物免疫 Stegmann et al., 2017 FER与MLO家族成员NORTIA协同作用 拟南芥和高氏白粉病(Golovinomyces orontii ) FER负调节拟南芥对高氏白粉病的抗性 Kessler et al., 2010 FER与FLS2和BAK1互作 拟南芥和丁香假单胞杆菌番茄致病变种 RALF23 fer 突变体出现氧化爆发、MAPK活性、气孔开合和细胞死亡等方面的功能缺陷Keinath et al., 2010
F-RALF, MYC2, FLS2, EFR, BAK1, RALF23 and MAPK see Figure 1 . ...Ngo等(2014) 和Shih等(2014) 研究表明, FER还通过调节胞质Ca2+ 信号响应细胞内外压力信号和盐胁迫.有研究表明, 拟南芥细胞壁中富含亮氨酸重复的细胞伸展蛋白LRXs3/4/5 (leucine-rich repeat extensins3/4/5), 该类蛋白对植物的耐盐性至关重要, 而LRXs、RALFs和FER作为1个功能模块, 通过感受细胞壁完整信号调节植物生长及对盐胁迫的响应(Zhao et al., 2018 ).FER也可作为细胞壁传感器参与调节细胞间的互作, 控制RAC/ROP GTPase信号, 进而激活RAC/ROPs蛋白, 刺激ROS产生(Cheung and Wu, 2011 ). ...Arabidopsis seedlingsNgo等(2014) 和Shih等(2014) 研究表明, FER还通过调节胞质Ca2+ 信号响应细胞内外压力信号和盐胁迫.有研究表明, 拟南芥细胞壁中富含亮氨酸重复的细胞伸展蛋白LRXs3/4/5 (leucine-rich repeat extensins3/4/5), 该类蛋白对植物的耐盐性至关重要, 而LRXs、RALFs和FER作为1个功能模块, 通过感受细胞壁完整信号调节植物生长及对盐胁迫的响应(Zhao et al., 2018 ).FER也可作为细胞壁传感器参与调节细胞间的互作, 控制RAC/ROP GTPase信号, 进而激活RAC/ROPs蛋白, 刺激ROS产生(Cheung and Wu, 2011 ). ...Arabidopsis and riceShiu et al., 2004 ; Jose et al., 2020 ).植物受体蛋白激酶(receptor protein kinases, RPKs)大多为丝氨酸/苏氨酸蛋白激酶, 能够识别胞外信号, 并将信号传递到胞内, 通过丝/苏氨酸残基的磷酸化/去磷酸化向下游传递(Tang et al., 2017 ; 朱巍巍等, 2018 ).基因组测序结果显示, 水稻(Oryza sativa )和拟南芥(Arabidopsis thaliana )基因组中含有数量众多的受体激酶编码基因, 但其中大多数基因编码产物的天然配体尚未找到, 受体功能也未确定, 因此这些基因编码产物常被称为类受体激酶(receptor-like kinase, RLKs) (朱巍巍等, 2018 ).RLKs通常由胞外结合结构域、跨膜结构域和胞内激酶结构域3部分组成.不同类受体激酶的胞外结合结构域存在显著差异, 根据其胞外域的差异主要分为6类, 即PR5型(pathogenesis-related protein-5 like receptor kinases)、S-结构域型(S-domain)、类表皮生长因子型(epidermal growth factor like)、类肿瘤坏死因子受体型(tumor-necrosis factor receptor-like)、类凝集素型(lectin-like)和富含亮氨酸重复序列型(leucine-rich repeat) (朱巍巍等, 2018 ).最新研究表明, 根据受体激酶胞外域的特征基序, 拟南芥跨膜RLKs可进一步分为富含亮氨酸重复序列型(leucine- rich repeat, LRR)、凝集素型(C-lectin和L-lectin)、细胞壁相关激酶(wall-associated kinase, WAK)、类伸展蛋白型(extensin like)、富含脯氨酸类伸展蛋白型(proline-rich extensin like, PERK)、细胞溶解酶基序型(lysin motif, LysM)、类长春花(Catharanthus rose- like, Cr RLK)和S-结构域型(self-incompatibility domain, S-domain)等14种(Jose et al., 2020 ).近年来, 一类胞外结构域中含有Malectin-like基序的类受体激酶(主要包括FER、ANX1 (ANXUR1)、ANX2、THE1 (THESEUS1)和HERK1 (HERCULES1)等)受到广泛关注.其中, FER在植物生长发育、激素互作网络、逆境响应以及植物与病原菌互作等众多生物学过程中发挥重要作用. ...Table 1 互作蛋白 涉及物种 配体 主要功能/结论 参考文献 FER与病原菌分泌的 拟南芥(Arabidopsis thaliana )和尖孢镰刀菌(Fusarium oxysporium ) F-RALF FER识别尖孢镰刀菌分泌的F- RALF, 阻断质膜H+ -ATPase2诱导胞外环境碱化, 从而负调控拟南芥对尖孢镰刀菌的抗性 Masachis et al., 2016 FER与转录因子MYC2 拟南芥和丁香假单胞杆菌番茄致病变种(Pseudomonas syrin- pv. tomato DC3000) RALF23 FER正调控拟南芥对Pst DC3000 Guo et al., 2018 FER与FLS2、EFR和BAK1形成免疫复合体 拟南芥 RALF23 拟南芥SITE-1蛋白酶剪切内源性快速碱化因子(RALF)前肽, 抑制植物免疫 Stegmann et al., 2017 FER与MLO家族成员NORTIA协同作用 拟南芥和高氏白粉病(Golovinomyces orontii ) FER负调节拟南芥对高氏白粉病的抗性 Kessler et al., 2010 FER与FLS2和BAK1互作 拟南芥和丁香假单胞杆菌番茄致病变种 RALF23 fer 突变体出现氧化爆发、MAPK活性、气孔开合和细胞死亡等方面的功能缺陷Keinath et al., 2010
F-RALF, MYC2, FLS2, EFR, BAK1, RALF23 and MAPK see Figure 1 . ...Stegmann等(2017) 研究发现, 拟南芥SITE-1蛋白酶(S1P)剪切内源性RALF前体肽, FER抑制植物免疫反应.研究表明, 在elf18、flg22和几丁质的作用下, s1p-3 和s1p-6 单突变体产生更多的ROS, 对低致病力的丁香假单胞菌番茄致病变种(Pseudomonas syringae pv. tomato ) (Pto ) DC3000 COR- (coronatine-minus )表现出更高抗性, 因此证明S1P是免疫负调控因子(Stegmann et al., 2017 ).过表达RALF23能够抑制elf18触发的ROS生成, 增强植株对Pto DC3000 COR - 和黄瓜枯萎菌Plectosp- haerella cucumerina 的敏感性, 而RALF23缺失促进elf18触发的ROS产生, 增强植物对Pto DC3000的抗性, 表明RALF23同样负调控免疫.进一步研究显示, fer2 和fer4 突变体对elf18、flg22和几丁质的敏感性较低, 对Pto DC3000 COR - 更敏感, 说明FER能正向调节免疫过程.flg22处理后, FER与FLS2和BAK1之间的互作增强, 而在fer4 突变体中, flg22诱导的FLS2- BAK1复合物组装减少, fer4 /FER-GFP 转基因株系中复合物的水平恢复正常.RALF23 过表达对flg22诱导的FLS2-BAK1复合物形成也有类似影响.值得注意的是, RALF23或FER的缺失并不影响FLS2、EFR或BAK1的积累.上述结果表明, FER促进配体诱导的FLS2/EFR与其共受体BAK1之间形成复合物, 而RALF23抑制这一过程, 进一步证实FER是调节免疫受体复合物形成的重要支架组分, 与其受体和共受体共同定位于质膜微区中(Stegmann et al., 2017 ).这种FER-RALF模块可能通过调节不同的受体激酶复合物, 在生长发育或环境感知等过程中发挥重要作用(Stegmann et al., 2017 ).Huang等(2020a) 研究表明, 水稻中类FER受体FLR (FER-like receptor)家族成员能够调节水稻与稻瘟病菌(Magnaporthe oryzae )互作.他们利用CRISPR/Cas9基因编辑技术成功敲除了FLR家族的14个成员, 结果显示FLR1 和FLR13 的突变体对稻瘟病的敏感性增强, 而FLR2 和FLR11 的功能缺失增强了水稻对稻瘟病的抗性, 且几乎未对水稻生长造成影响(Huang et al., 2020a ). ...Stegmann et al., 2017 ).过表达RALF23能够抑制elf18触发的ROS生成, 增强植株对Pto DC3000 COR - 和黄瓜枯萎菌Plectosp- haerella cucumerina 的敏感性, 而RALF23缺失促进elf18触发的ROS产生, 增强植物对Pto DC3000的抗性, 表明RALF23同样负调控免疫.进一步研究显示, fer2 和fer4 突变体对elf18、flg22和几丁质的敏感性较低, 对Pto DC3000 COR - 更敏感, 说明FER能正向调节免疫过程.flg22处理后, FER与FLS2和BAK1之间的互作增强, 而在fer4 突变体中, flg22诱导的FLS2- BAK1复合物组装减少, fer4 /FER-GFP 转基因株系中复合物的水平恢复正常.RALF23 过表达对flg22诱导的FLS2-BAK1复合物形成也有类似影响.值得注意的是, RALF23或FER的缺失并不影响FLS2、EFR或BAK1的积累.上述结果表明, FER促进配体诱导的FLS2/EFR与其共受体BAK1之间形成复合物, 而RALF23抑制这一过程, 进一步证实FER是调节免疫受体复合物形成的重要支架组分, 与其受体和共受体共同定位于质膜微区中(Stegmann et al., 2017 ).这种FER-RALF模块可能通过调节不同的受体激酶复合物, 在生长发育或环境感知等过程中发挥重要作用(Stegmann et al., 2017 ).Huang等(2020a) 研究表明, 水稻中类FER受体FLR (FER-like receptor)家族成员能够调节水稻与稻瘟病菌(Magnaporthe oryzae )互作.他们利用CRISPR/Cas9基因编辑技术成功敲除了FLR家族的14个成员, 结果显示FLR1 和FLR13 的突变体对稻瘟病的敏感性增强, 而FLR2 和FLR11 的功能缺失增强了水稻对稻瘟病的抗性, 且几乎未对水稻生长造成影响(Huang et al., 2020a ). ...Stegmann et al., 2017 ).这种FER-RALF模块可能通过调节不同的受体激酶复合物, 在生长发育或环境感知等过程中发挥重要作用(Stegmann et al., 2017 ).Huang等(2020a) 研究表明, 水稻中类FER受体FLR (FER-like receptor)家族成员能够调节水稻与稻瘟病菌(Magnaporthe oryzae )互作.他们利用CRISPR/Cas9基因编辑技术成功敲除了FLR家族的14个成员, 结果显示FLR1 和FLR13 的突变体对稻瘟病的敏感性增强, 而FLR2 和FLR11 的功能缺失增强了水稻对稻瘟病的抗性, 且几乎未对水稻生长造成影响(Huang et al., 2020a ). ...Stegmann et al., 2017 ).Huang等(2020a) 研究表明, 水稻中类FER受体FLR (FER-like receptor)家族成员能够调节水稻与稻瘟病菌(Magnaporthe oryzae )互作.他们利用CRISPR/Cas9基因编辑技术成功敲除了FLR家族的14个成员, 结果显示FLR1 和FLR13 的突变体对稻瘟病的敏感性增强, 而FLR2 和FLR11 的功能缺失增强了水稻对稻瘟病的抗性, 且几乎未对水稻生长造成影响(Huang et al., 2020a ). ...Shiu et al., 2004 ; Jose et al., 2020 ).植物受体蛋白激酶(receptor protein kinases, RPKs)大多为丝氨酸/苏氨酸蛋白激酶, 能够识别胞外信号, 并将信号传递到胞内, 通过丝/苏氨酸残基的磷酸化/去磷酸化向下游传递(Tang et al., 2017 ; 朱巍巍等, 2018 ).基因组测序结果显示, 水稻(Oryza sativa )和拟南芥(Arabidopsis thaliana )基因组中含有数量众多的受体激酶编码基因, 但其中大多数基因编码产物的天然配体尚未找到, 受体功能也未确定, 因此这些基因编码产物常被称为类受体激酶(receptor-like kinase, RLKs) (朱巍巍等, 2018 ).RLKs通常由胞外结合结构域、跨膜结构域和胞内激酶结构域3部分组成.不同类受体激酶的胞外结合结构域存在显著差异, 根据其胞外域的差异主要分为6类, 即PR5型(pathogenesis-related protein-5 like receptor kinases)、S-结构域型(S-domain)、类表皮生长因子型(epidermal growth factor like)、类肿瘤坏死因子受体型(tumor-necrosis factor receptor-like)、类凝集素型(lectin-like)和富含亮氨酸重复序列型(leucine-rich repeat) (朱巍巍等, 2018 ).最新研究表明, 根据受体激酶胞外域的特征基序, 拟南芥跨膜RLKs可进一步分为富含亮氨酸重复序列型(leucine- rich repeat, LRR)、凝集素型(C-lectin和L-lectin)、细胞壁相关激酶(wall-associated kinase, WAK)、类伸展蛋白型(extensin like)、富含脯氨酸类伸展蛋白型(proline-rich extensin like, PERK)、细胞溶解酶基序型(lysin motif, LysM)、类长春花(Catharanthus rose- like, Cr RLK)和S-结构域型(self-incompatibility domain, S-domain)等14种(Jose et al., 2020 ).近年来, 一类胞外结构域中含有Malectin-like基序的类受体激酶(主要包括FER、ANX1 (ANXUR1)、ANX2、THE1 (THESEUS1)和HERK1 (HERCULES1)等)受到广泛关注.其中, FER在植物生长发育、激素互作网络、逆境响应以及植物与病原菌互作等众多生物学过程中发挥重要作用. ...Pseudomonas syringae pv. tomato DC3000: a model pathogen for probing disease susceptibility and hormone signaling in plantsXin and He, 2013 ), 能通过茉莉酸信号途径的关键因子MYC2激活NAC转录因子, 抑制水杨酸积累, 从而破坏植物的免疫应答(Zheng et al., 2012 ).对fer 突变体全基因组水平的基因表达分析显示, fer 突变体中JA和COR信号途径相关的基因表达显著上调, fer 突变体更易被病原菌侵染.遗传学研究表明, FER通过磷酸化MYC2降低其稳定性, 从而抑制JA和COR信号, 正调控植物的免疫应答.与此相反, RALF23通过与FER互作增强MYC2的稳定性, 从而激活JA信号途径, 实现对植物免疫应答的负调控(Guo et al., 2018 ).上述结果表明, RALF23-FER-MYC2模块是JA和COR信号途径的关键调控组件, 寄主植物可以利用FER信号途径拮抗COR介导的寄主植物对病原菌的易感性(Guo et al., 2018 ). ...Arabidopsis by activating ABI2 phosphataseDuan等(2010) 研究表明, 拟南芥fer-4 突变体植株矮小, 生长缓慢且根毛发育严重缺陷.鸟苷酸交换因子RopGEF4和RopGEF10 (guanine nucleotide exchange factors for ROPs, RopGEF)也参与了该过程的调节(Huang et al., 2013 ).此外, FER还通过激活蛋白磷酸酶ABI2 (ABA insensitive 2)抑制脱落酸(abscisic acid, ABA)信号通路(Yu et al., 2012 ); FER、HERCULES1和THESEUS1与油菜素甾醇(brassino- steroids, BRs)途径协同控制细胞伸长(Guo et al., 2009 ).在暗形态建成的拟南芥幼苗中, FER与乙烯和油菜素甾醇信号通路平行或协同促进下胚轴伸长(Guo et al., 2009 ; Deslauriers and Larsen, 2010 ); 且FER能够调节乙烯的生物合成途径, 拟南芥fer 突变体中积累较高水平的S-腺苷甲硫氨酸(S-adenosyl methionine, SAM)和乙烯, FER通过与S-腺苷甲硫氨酸合成酶互作, 抑制S-腺苷甲硫氨酸和乙烯的生物合成, 导致植株出现矮化表型(Mao et al., 2015 ). ...Arabidopsis Catharanthus roseus RLK1-like kinases, Cr RLK1Ls) (Escobar-Restrepo et al., 2007 ), 最早被发现能够介导雄配子体和雌配子体之间的识别(Huck et al., 2003 ; Escobar- Restrepo et al., 2007 ).拟南芥中的FER与该家族的其它16个蛋白在类受体激酶超家族中形成1个分支, 其胞外结构域中的Malectin-like蛋白基序具有结合二聚及寡聚葡萄糖的能力(Li and Zhang, 2014 ), 暗示该类受体激酶在植物细胞间互作及植物-环境互作过程中发挥重要作用.近年研究表明, FER广泛参与调节高等植物的受精、细胞伸长、生物及非生物胁迫响应等不同的生物学过程(Chen et al., 2020 ; Ji et al., 2020b ; Zhang et al., 2020b ).FER是胚囊接受花粉管所必需, 并且是雌配子体育性的重要调节因子(Huck et al., 2003 ).拟南芥fer 突变体中雌配子体对花粉管接收发生功能缺陷, 雌雄配子之间的相互作用被破坏, 导致花粉管生长失控, 不能破裂释放精细胞, 最终导致育性下降(Huck et al., 2003 ).进一步研究发现, FER定位于丝状器(filiform apparatus), 通过介导雌配子体对花粉管的接收, 触发助细胞内的信号级联反应, 其缺失导致植株生长矮小和精细胞释放受阻(Escobar-Restrepo et al., 2007 ).Duan等(2020) 研究表明, FER能控制丝状器中的去酯化果胶水平, 防止多根花粉管进入胚囊.花粉管到达胚珠时会诱导丝状器处产生一氧化氮(nitric oxide, NO), 而这一过程受FER和去酯化果胶共同调节.NO能使前体及其成熟形式的LURE小肽发生硝基化, 从而抑制助细胞分泌LURE.有研究表明, 正常情况下, FER进行依赖网格蛋白和不依赖网格蛋白的内吞作用, 而小肽RALF1 (rapid alkalinization factor 1)处理能刺激CME (clathrin-mediated endocytosis)途径的内吞.阻断CME途径可减弱RALF1对根系生长的抑制, 表明RALF1- FER互作调节植物的生长发育, RALF1则可通过CME途径发挥作用(Yu et al., 2020 ). ...Catharanthus roseus RLK1-like kinases, Cr RLK1Ls) (Escobar-Restrepo et al., 2007 ), 最早被发现能够介导雄配子体和雌配子体之间的识别(Huck et al., 2003 ; Escobar- Restrepo et al., 2007 ).拟南芥中的FER与该家族的其它16个蛋白在类受体激酶超家族中形成1个分支, 其胞外结构域中的Malectin-like蛋白基序具有结合二聚及寡聚葡萄糖的能力(Li and Zhang, 2014 ), 暗示该类受体激酶在植物细胞间互作及植物-环境互作过程中发挥重要作用.近年研究表明, FER广泛参与调节高等植物的受精、细胞伸长、生物及非生物胁迫响应等不同的生物学过程(Chen et al., 2020 ; Ji et al., 2020b ; Zhang et al., 2020b ).FER是胚囊接受花粉管所必需, 并且是雌配子体育性的重要调节因子(Huck et al., 2003 ).拟南芥fer 突变体中雌配子体对花粉管接收发生功能缺陷, 雌雄配子之间的相互作用被破坏, 导致花粉管生长失控, 不能破裂释放精细胞, 最终导致育性下降(Huck et al., 2003 ).进一步研究发现, FER定位于丝状器(filiform apparatus), 通过介导雌配子体对花粉管的接收, 触发助细胞内的信号级联反应, 其缺失导致植株生长矮小和精细胞释放受阻(Escobar-Restrepo et al., 2007 ).Duan等(2020) 研究表明, FER能控制丝状器中的去酯化果胶水平, 防止多根花粉管进入胚囊.花粉管到达胚珠时会诱导丝状器处产生一氧化氮(nitric oxide, NO), 而这一过程受FER和去酯化果胶共同调节.NO能使前体及其成熟形式的LURE小肽发生硝基化, 从而抑制助细胞分泌LURE.有研究表明, 正常情况下, FER进行依赖网格蛋白和不依赖网格蛋白的内吞作用, 而小肽RALF1 (rapid alkalinization factor 1)处理能刺激CME (clathrin-mediated endocytosis)途径的内吞.阻断CME途径可减弱RALF1对根系生长的抑制, 表明RALF1- FER互作调节植物的生长发育, RALF1则可通过CME途径发挥作用(Yu et al., 2020 ). ...Arabidopsis Ngo等(2014) 和Shih等(2014) 研究表明, FER还通过调节胞质Ca2+ 信号响应细胞内外压力信号和盐胁迫.有研究表明, 拟南芥细胞壁中富含亮氨酸重复的细胞伸展蛋白LRXs3/4/5 (leucine-rich repeat extensins3/4/5), 该类蛋白对植物的耐盐性至关重要, 而LRXs、RALFs和FER作为1个功能模块, 通过感受细胞壁完整信号调节植物生长及对盐胁迫的响应(Zhao et al., 2018 ).FER也可作为细胞壁传感器参与调节细胞间的互作, 控制RAC/ROP GTPase信号, 进而激活RAC/ROPs蛋白, 刺激ROS产生(Cheung and Wu, 2011 ). ...Pseu- domonas syringae virulence in plants by activating a signaling cascade that inhibits salicylic acid accumulationXin and He, 2013 ), 能通过茉莉酸信号途径的关键因子MYC2激活NAC转录因子, 抑制水杨酸积累, 从而破坏植物的免疫应答(Zheng et al., 2012 ).对fer 突变体全基因组水平的基因表达分析显示, fer 突变体中JA和COR信号途径相关的基因表达显著上调, fer 突变体更易被病原菌侵染.遗传学研究表明, FER通过磷酸化MYC2降低其稳定性, 从而抑制JA和COR信号, 正调控植物的免疫应答.与此相反, RALF23通过与FER互作增强MYC2的稳定性, 从而激活JA信号途径, 实现对植物免疫应答的负调控(Guo et al., 2018 ).上述结果表明, RALF23-FER-MYC2模块是JA和COR信号途径的关键调控组件, 寄主植物可以利用FER信号途径拮抗COR介导的寄主植物对病原菌的易感性(Guo et al., 2018 ). ...  ,1,*, 田世平1,2
,1,*, 田世平1,2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}