Comparative Genomics of the Small Heat Shock Protein Family in Angiosperms
Kai Fan, Fangting Ye, Zhijun Mao, Xinfeng Pan, Zhaowei Li, Wenxiong Lin,*Key Laboratory of Ministry of Education for Genetics, Breeding and Multiple Utilization of Crops, College of Agriculture, Fujian Agriculture and Forestry University, Fuzhou 350002
Abstract Small heat shock protein (sHSP) can respond to environmental stresses and regulate plant growth and development. In this study, 33 NcsHSPs, 24 OssHSPs, 17 AtsHSPs and 47 VvsHSPs were found in waterlily, rice, Arabidopsis, and grape, respectively. The sHSP family can be divided into 12 subfamilies, and different subfamily had different number of sHSP members, conserved motifs, gene structures and number of duplicated genes. Then, 12 gene duplication events in tle sHSP fanily were identified from 4 model angiosperms of sHSP family. The segmental and tandem duplication events were related to the gene expansion in the sHSP family, and the segmental duplication events occurred earlier than the tandem duplication events. Moreover, sHSP members in Arabidopsis and grape had the highest orthologous similarity, followed by sHSP members in waterlily and grape. Furthermore, sHSP family in angiosperms evolved towards sHSP members with smaller length of amino acid, lower molecular weight, simpler gene structure, and concentrated chromosomal localization. Besides, some sHSP members may be related to plant growth and development in waterlily, rice, Arabidopsis, and grape. Above all, these results could lay vital foundations on comparative genomics of the sHSP family in four model angiosperms and advance the sHSP family in other angiosperms. Keywords:sHSP family;comparative genomics;expansion;gene expression
PDF (1709KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 范凯, 叶方婷, 毛志君, 潘鑫峰, 李兆伟, 林文雄. 被子植物小热激蛋白家族的比较基因组学分析. 植物学报, 2021, 56(3): 245-261 doi:10.11983/CBB20209 Fan Kai, Ye Fangting, Mao Zhijun, Pan Xinfeng, Li Zhaowei, Lin Wenxiong. Comparative Genomics of the Small Heat Shock Protein Family in Angiosperms. Chinese Bulletin of Botany, 2021, 56(3): 245-261 doi:10.11983/CBB20209
Table 1 表1 表1睡莲、水稻、拟南芥和葡萄中sHSP成员的鉴定及结构分析 Table 1The identification and structural analysis of the sHSP members in waterlily, rice, Arabidopsis and grape
通过IQ-tree软件构建系统进化树(左), 分支上的数字代表bootstrap值, sHSP亚家族用红色字母表示。保守基序(中)通过MEME程序识别, 不同颜色代表不同的保守基序, 保守基序的位置使用下方标尺进行估计。根据内含子和外显子的位置构建基因结构示意图(右), 绿色方框表示外显子, 黑色线表示内含子, 内含子和外显子的位置使用下方标尺进行估计。 Figure 2The phylogenetic analysis (left), conserved motif (middle), and gene structure (right) of the sHSF family in waterlily, rice, Arabidopsis and grape
The phylogenetic tree (left) was constructed by IQ-tree software. The numbers in the clades stood for the bootstrap values, and sHSP subfamilies were indicated by red letters. The conserved motifs (middle) were identified by MEME software. Each conserved motif was marked by a specific color, and the location of motifs can be estimated using the scale at the bottom. The gene structures (right) were visualized according to the location of exons and introns. The exons and introns were represented by green box and black line, respectively. The location of exons and introns can be estimated using the scale at the bottom.
实线连线和虚线连线分别代表片段复制事件和串联复制事件。 Figure 3Chromosomal locations of NcsHSP (A), OssHSP (B), AtsHSP (C) and VvsHSP (D) on the waterlily, rice, Arabidopsis and grape chromosomes
The solid lines and dot lines represented the segmental and tandem duplication events, respectively.
Table 2 表2 表2在睡莲、水稻、拟南芥和葡萄中复制的sHSP成员的Ka和Ks分析 Table 2Ka and Ks analysis for the duplicated sHSP members in waterlily, rice, Arabidopsis and grape
(A) 睡莲与水稻、拟南芥和葡萄中sHSP成员的共线性分析; (B) 水稻与拟南芥和葡萄sHSP成员的共线性分析; (C) 拟南芥与葡萄sHSP成员的共线性分析; (D) 在不同物种中sHSP同源基因对的Ka值分析。(A)-(C) 灰线代表所有的同源基因, 红线代表sHSP同源基因。 Figure 4Synteny analysis of sHSP members in waterlily, rice, Arabidopsis and grape
(A) Synteny analysis of sHSP members between waterlily and rice, Arabidopsis and grape; (B) Synteny analysis of sHSP members between rice and Arabidopsis and grape; (C) Synteny analysis of sHSP members between Arabidopsis and grape; (D) Comparison of Ka values of orthologous sHSPs between different species. (A)-(C) Gray lines connected orthologous genes, and orthologous sHSPs were marked with red lines.
之前的研究表明, sHSP家族可以参与许多植物生长与发育过程(Zhang et al., 2018; Ma et al., 2019)。本研究在睡莲、水稻、拟南芥和葡萄中新发现一些与植物生长发育过程相关的sHSP成员(图5)。CI亚家族的NcsHSP19、OssHSP02、OssHSP14和VvsHSP40以及CIV亚家族的AtsHSP09和OssHSP23在几乎所有组织中均高度表达, 其在棉花和马铃薯中的同源基因也具有类似的表达模式(Ma et al., 2016; Zhao et al., 2018), 这表明sHSP家族在植物中的功能可能具有一定的保守性。此外, 本研究还发现一些与水稻花器官发育相关的基因(OssHSP03、OssHSP04、OssHSP06、OssHSP12、OssHSP16和OssHSP21)。有报道显示, 在玫瑰(Rosa hybrida)中与CI亚家族OssHSP03、OssHSP04和OssHSP16相同亚家族的sHSP17.5-CI可能与开花过程中保护细胞蛋白有关(Dafny-Yelin et al., 2008)。
CannonSB, MitraA, BaumgartenA, YoungND, MayG (2004). The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana BMC Plant Biol 4, 10. DOI:10.1186/1471-2229-4-10URL [本文引用: 1]
ChenCJ, ChenH, ZhangY, ThomasHR, FrankMH, HeYH, XiaR (2020). TBtools: an integrative toolkit developed for interactive analyses of big biological data Mol Plant 13, 1194-1202. DOI:10.1016/j.molp.2020.06.009URL [本文引用: 2]
Dafny-YelinM, TzfiraT, VainsteinA, AdamZ (2008). Non-redundant functions of sHSP-CIs in acquired thermotolerance and their role in early seed development in Arabidopsis Plant Mol Biol 67, 363-373. DOI:10.1007/s11103-008-9326-4PMID:18379884 [本文引用: 1] Heat-shock proteins (HSPs) are a group of evolutionarily conserved polypeptides whose expression is induced in all organisms in response to environmental stresses and during various developmental processes. In this work, we show that the rose (Rosa hybrida) cytoplasmic 17.5-kDa Class I small HSP (sHSP17.5-CI, accession number: BQ103946) increases dramatically during flower development, and accumulates in closed bud petals and leaves only in response to heat stress. mRNA for a putative ortholog of this protein is also found in petals, but not leaves, of Arabidopsis (Arabidopsis thaliana) plants grown under optimal conditions, and it accumulates in leaves in response to heat stress. Analysis of Arabidopsis T-DNA insertion lines affected at three homologous genes revealed that their acquired thermotolerance, as measured by hypocotyl-elongation assay, is impaired. The correlation between sHSP-CI accumulation and expansion of rose petal cells, impairment of acquired thermotolerance, and defects in early embryogenesis of the double mutants (hsp17.4/hsp17.6A), all suggest that sHSP-CI proteins play a role in protecting cell proteins at various developmental stages, whereas in hypocotyl elongation they have a non-redundant function in acquired thermotolerance but have a redundant function in early embryogenesis.
FanK, ChenYR, MaoZJ, FangY, LiZW, LinWW, ZhangYQ, LiuJP, HuangJW, LinWX (2020a). Pervasive duplication, biased molecular evolution and comprehensive functional analysis of the PP2C family in Glycine max BMC Genomics 21, 465. DOI:10.1186/s12864-020-06877-4URL [本文引用: 2]
FanK, MaoZJ, ZhengJX, ChenYR, LiZW, LinWW, ZhangYQ, HuangJW, LinWX (2020b). Molecular evolution and expansion of the KUP family in the allopolyploid cotton species Gossypium hirsutum and Gossypium barbadense Front Plant Sci 11, 545042. DOI:10.3389/fpls.2020.545042URL [本文引用: 2]
FanK, ShenH, BibiN, LiF, YuanSN, WangM, WangXD (2015). Molecular evolution and species-specific expansion of the NAP members in plants J Integr Plant Biol 57, 673-687. DOI:10.1111/jipb.v57.8URL [本文引用: 1]
FanK, WangM, MiaoY, NiM, BibiN, YuanSN, LiF, WangXD (2014). Molecular evolution and expansion analysis of the NAC transcription factor in Zea mays PLoS One 9, e111837. DOI:10.1371/journal.pone.0111837URL [本文引用: 1]
GuoCL, GuoRR, XuXZ, GaoM, LiXQ, SongJY, ZhengY, WangXP (2014). Evolution and expression analysis of the grape (Vitis vinifera L.) WRKY gene family J Exp Bot 6, 1513-1528. [本文引用: 1]
GuoLM, LiJ, HeJ, LiuH, ZhangHM (2020). A class I cytosolic HSP20 of rice enhances heat and salt tolerance in different organisms Sci Rep 10, 1383. DOI:10.1038/s41598-020-58395-8URL [本文引用: 1]
GuoM, LiuJH, LuJP, ZhaiYF, WangH, GongZH, WangSB, LuMH (2015). Genome-wide analysis of the CaHsp20 gene family in pepper: comprehensive sequence and expression profile analysis under heat stress Front Plant Sci 6, 806. [本文引用: 3]
JiXR, YuYH, NiPY, ZhangGH, GuoDL (2019). Genome-wide identification of small heat-shock protein (HSP20) gene family in grape and expression profile during berry development BMC Plant Biol 19, 433. DOI:10.1186/s12870-019-2031-4URL [本文引用: 1]
JiaoYN, Leebens-MackJ, AyyampalayamS, BowersJE, McKainM, McNealJ, RolfM, RuzickaDR, WafulaE, WickettNJ, WuXL, ZhangY, WangJ, ZhangYT, CarpenterEJ, DeyholosMK, KutchanTM, ChanderbaliAS, SoltisPS, StevensonDW, McCombieR, PiresJC, WongGKS, SoltisDE, DePamphilisDW (2012). A genome triplication associated with early diversification of the core eudicots Genome Biol 13, R3. DOI:10.1186/gb-2012-13-1-r3URL [本文引用: 1]
KeyJL, LinCY, ChenYM (1981). Heat shock proteins of higher plants Proc Natl Acad Sci USA 78, 3526-3530. DOI:10.1073/pnas.78.6.3526URL [本文引用: 1]
LameschP, BerardiniTZ, LiDH, SwarbreckD, WilksC, SasidharanR, MullerR, DreherK, AlexanderDL, Garcia-HernandezM, KarthikeyanAS, LeeCH, NelsonWD, PloetzL, SinghS, WenselA, HualaE (2012). The Arabidopsis Information Resource (TAIR): improved gene annotation and new tools Nucleic Acids Res 40, D1202-D1210. DOI:10.1093/nar/gkr1090URL [本文引用: 3]
LiJB, ZhangJ, JiaHX, LiY, XuXD, WangLJ, LuMZ (2016). The Populus trichocarpa PtHSP17.8 involved in heat and salt stress tolerances Plant Cell Rep 35, 1587-1599. DOI:10.1007/s00299-016-1973-3URL [本文引用: 1]
LiJB, ZhangJ, JiaHX, YueZQ, LuMZ, XinXB, HuJJ (2018). Genome-wide characterization of the sHsp gene family in Salix suchowensis reveals its functions under different abiotic stresses Int J Mol Sci 19, 3246. DOI:10.3390/ijms19103246URL [本文引用: 3]
LiQ, ZhangN, ZhangLS, MaH (2015). Differential evolution of members of the rhomboid gene family with conservative and divergent patterns New Phytol 206, 368-380. DOI:10.1111/nph.2015.206.issue-1URL [本文引用: 1]
MaW, GuanXY, LiJ, PanRH, WangLY, LiuFJ, MaHY, ZhuSJ, HuJ, RuanYL, ChenXY, ZhangTZ (2019). Mitochondrial small heat shock protein mediates seed germination via thermal sensing Proc Natl Acad Sci USA 116, 4716-4721. DOI:10.1073/pnas.1815790116URL [本文引用: 2]
MaW, ZhaoT, LiJ, LiuBL, FangL, HuY, ZhangTZ (2016). Identification and characterization of the GhHsp20 gene family in Gossypium hirsutum Sci Rep 6, 32517. DOI:10.1038/srep32517URL [本文引用: 4]
MaereS, de BodtS, RaesJ, CasneufT, van MontaguM, KuiperM, van de PeerY (2005). Modeling gene and genome duplications in eukaryotes Proc Natl Acad Sci USA 102, 5454-5459. DOI:10.1073/pnas.0501102102URL [本文引用: 1]
OuyangS, ZhuW, HamiltonJ, LinHN, CampbellM, ChildsK, Thibaud-NissenF, MalekRL, LeeY, ZhengL, OrvisJ, HaasB, WortmanJ, BuellCR (2007). The TIGR Rice Genome Annotation Resource: improvements and new features Nucleic Acids Res 35, D883-D887. DOI:10.1093/nar/gkl976URL [本文引用: 3]
ParkKC, KimNS (2010). Intron loss mediated structural dynamics and functional differentiation of the polygalacturonase gene family in land plants Genes Genom 32, 570-577. DOI:10.1007/s13258-010-0076-8URL [本文引用: 1]
PerteaM, KimD, PerteaGM, LeekJT, SalzbergSL (2016). Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown Nat Protoc 11, 1650-1667. DOI:10.1038/nprot.2016.095URL [本文引用: 1]
SarkarNK, KimYK, GroverA (2009). Rice sHsp genes: genomic organization and expression profiling under stress and development BMC Genomics 10, 393. DOI:10.1186/1471-2164-10-393URL [本文引用: 1]
ScharfKD, SiddiqueM, VierlingE (2001). The expanding family of Arabidopsis thaliana small heat stress proteins and a new family of proteins containing α-crystallin domains (Acd proteins) Cell Stress Chaperones 6, 225-237. DOI:10.1379/1466-1268(2001)006<0225:TEFOAT>2.0.CO;2URL [本文引用: 1]
ShazadeeH, KhanN, WangJJ, WangCC, ZengJG, HuangZY, WangXY (2019). Identification and expression profiling of protein phosphatases (PP2C) gene family in Gossypium hirsutum L Int J Mol Sci 20, 1395. DOI:10.3390/ijms20061395URL [本文引用: 1]
SunWN, BernardC, Van De CotteB, Van MontaguM, VerbruggenN (2010). At-HSP17.6A, encoding a small heat-shock protein in Arabidopsis, can enhance osmotolerance upon overexpression Plant J 27, 407-415. DOI:10.1046/j.1365-313X.2001.01107.xURL [本文引用: 1]
The French-Italian Public Consortium for Grapevine Genome Characterization (2007). The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla Nature 449, 463-467. DOI:10.1038/nature06148URL [本文引用: 5]
van MontfortRLM, BashaE, FriedrichKL, SlingsbyC, VierlingE (2001). Crystal structure and assembly of a eukaryotic small heat shock protein Nat Struct Biol 8, 1025-1030. PMID:11702068 [本文引用: 1] The 2.7 A structure of wheat HSP16.9, a member of the small heat shock proteins (sHSPs), indicates how its alpha-crystallin domain and flanking extensions assemble into a dodecameric double disk. The folding of the monomer and assembly of the oligomer are mutually interdependent, involving strand exchange, helix swapping, loose knots and hinged extensions. In support of the chaperone mechanism, the substrate-bound dimers, in temperature-dependent equilibrium with higher assembly forms, have unfolded N-terminal arms and exposed conserved hydrophobic binding sites on the alpha-crystallin domain. The structure also provides a model by which members of the sHSP protein family bind unfolded substrates, which are involved in a variety of neurodegenerative diseases and cataract formation.
WangL, ZhangSL, ZhangXM, HuXY, GuoCL, WangXP, SongJY (2018). Evolutionary and expression analysis of Vitis vinifera OFP gene family Plant Syst Evol 304, 995-1008. DOI:10.1007/s00606-018-1528-xURL [本文引用: 1]
WangWX, VinocurB, ShoseyovO, AltmanA (2004). Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response Trends Plant Sci 9, 244-252. DOI:10.1016/j.tplants.2004.03.006URL [本文引用: 1]
WatersER, VierlingE (2020). Plant small heat shock proteins—evolutionary and functional diversity New Phytol 227, 24-37. DOI:10.1111/nph.v227.1URL [本文引用: 5]
ZhaiMZ, SunYD, JiaCX, PengSB, LiuZX, YangGY (2016). Over-expression of JrsHSP17.3 gene from Juglans regia confer the tolerance to abnormal temperature and NaCl stresses J Plant Biol 59, 549-558. DOI:10.1007/s12374-015-0507-9URL [本文引用: 1]
ZhangK, HanYT, ZhaoFL, HuY, GaoYR, MaYF, ZhengY, WangYJ, WenYQ (2015). Genome-wide identification and expression analysis of the CDPK gene family in grape, Vitis spp BMC Plant Biol 15, 164. DOI:10.1186/s12870-015-0552-zPMID:26122404 [本文引用: 1] Background: Calcium-dependent protein kinases (CDPKs) play vital roles in plant growth and development, biotic and abiotic stress responses, and hormone signaling. Little is known about the CDPK gene family in grapevine. Results: In this study, we performed a genome-wide analysis of the 12X grape genome (Vitis vinifera) and identified nineteen CDPK genes. Comparison of the structures of grape CDPK genes allowed us to examine their functional conservation and differentiation. Segmentally duplicated grape CDPK genes showed high structural conservation and contributed to gene family expansion. Additional comparisons between grape and Arabidopsis thaliana demonstrated that several grape CDPK genes occured in the corresponding syntenic blocks of Arabidopsis, suggesting that these genes arose before the divergence of grapevine and Arabidopsis. Phylogenetic analysis divided the grape CDPK genes into four groups. Furthermore, we examined the expression of the corresponding nineteen homologous CDPK genes in the Chinese wild grape (Vitis pseudoreticulata) under various conditions, including biotic stress, abiotic stress, and hormone treatments. The expression profiles derived from reverse transcription and quantitative PCR suggested that a large number of VpCDPKs responded to various stimuli on the transcriptional level, indicating their versatile roles in the responses to biotic and abiotic stresses. Moreover, we examined the subcellular localization of VpCDPKs by transiently expressing six VpCDPK-GFP fusion proteins in Arabidopsis mesophyll protoplasts; this revealed high variability consistent with potential functional differences. Conclusions: Taken as a whole, our data provide significant insights into the evolution and function of grape CDPKs and a framework for future investigation of grape CDPK genes.
ZhangLS, ChenF, ZhangXT, LiZ, ZhaoYY, LohausR, ChangXJ, DongW, HoSYW, LiuX, SongAX, ChenJH, GuoWL, WangZJ, ZhuangYY, WangHF, ChenXQ, HuJ, LiuYH, QinY, WangK, DongSS, LiuY, ZhangSZ, YuXX, WuQ, WangLS, YanXQ, JiaoYN, KongHZ, ZhouXF, YuCW, ChenYC, LiF, WangJH, ChenW, ChenXL, JiaQD, ZhangC, JiangYF, ZhangWB, LiuGH, FuJY, ChenF, MaH, van de PeerY, TangHB (2020a). The water lily genome and the early evolution of flowering plants Nature 577, 79-84. DOI:10.1038/s41586-019-1852-5URL [本文引用: 5]
ZhangLS, WuSD, ChangXJ, WangXJ, ZhaoYP, XiaYP, TrigianoRN, JiaoYN, ChenF (2020b). The ancient wave of polyploidization events in flowering plants and their facilitated adaptation to environmental stress Plant Cell Environ 43, 2847-2856. DOI:10.1111/pce.v43.12URL [本文引用: 1]
ZhangN, ShiJW, ZhaoHY, JiangJ (2018). Activation of small heat shock protein (SlHSP17.7) gene by cell wall invertase inhibitor (SlCIF1) gene involved in sugar metabolism in tomato Gene 679, 90-99. DOI:S0378-1119(18)30939-9PMID:30176314 [本文引用: 2] Fruit quality formation involves a series of physiological and biochemical changes during fruit ripening. Sucrose metabolism plays not only important roles in fruit ripening to establish energy status and nutritional quality but also a non-nutritive role in gene expression. In carbon metabolism and fruit ripening, cell wall invertases (CWINs) perform essential regulatory functions. Knowledge regarding the gene expression changes that occur following the repression of CWIN activity in fruit through the overexpression of a cell-wall inhibitor of β-fructosidase (CIF) under a fruit-specific promoter is limited. To further explore the molecular mechanism of sucrose regulation, global expression profiling of the fruits of transgenic tomato (Solanum lycopersicum) plants carrying a cell wall invertase inhibitor (SlCIF1) gene was performed using a microarray. In total, 622 and 833 differentially expressed genes (DEGs) were identified. The expression of the SlHSP17.7 gene was increased by thousands of times in the transgenic-SlCIF1 tomato. Then, SlHSP17.7-RNA interference (RNAi) lines were generated by introducing pB7GWIWG2 (I)-SlHSP17.7 into wild-type chmielewskii tomatoes (WT). The sucrose and fructose contents significantly decreased in the RNAi fruits compared with those in the WT. Furthermore, 14 sugar metabolism related genes were also decreased synergistically by silencing SlHSP17.7 gene. Our data indicate that the posttranslational modulation of CWIN activity by SlCIF1 contributes to earlier bloom times. SlHSP17.7 and sugar can interact to regulate the development of tomato fruit and affect the quality of tomato, providing a different insight into improving the quality of tomato.Copyright ? 2018 Elsevier B.V. All rights reserved.
ZhaoP, WangDD, WangRQ, KongNN, ZhangC, YangCH, WuWT, MaHL, ChenQ (2018). Genome-wide analysis of the potato Hsp20 gene family: identification, genomic organization and expression profiles in response to heat stress BMC Genomics 19, 61. DOI:10.1186/s12864-018-4443-1URL [本文引用: 4]
ZhuW, LuMH, GongZH, ChenRG (2011). Cloning and expression of a small heat shock protein gene CaHSP24 from pepper under abiotic stress Afr J Biotechnol 10, 4968-4975. [本文引用: 1]
ZhuYX, YanHW, WangYY, FengL, ChenZ, XiangY (2016). Genome duplication and evolution of heat shock transcription factor (HSF) gene family in four model angiosperms J Plant Growth Regul 35, 903-920. DOI:10.1007/s00344-016-9590-5URL [本文引用: 1]
ZhuoXK, ZhengTX, ZhangZY, ZhangYC, JiangLB, AhmadS, SunLD, WangJ, ChengTR, ZhangQX (2018). Genome-wide analysis of the NAC transcription factor gene family reveals differential expression patterns and cold-stress responses in the woody plant Prunus mume Genes (Basel) 9, 494. DOI:10.3390/genes9100494URL [本文引用: 2]
银杏(Ginkgo biloba) GbHsp20基因家族的鉴定及系统进化分析 2 2019
... 在外界环境发生变化时, 植物在生理和分子水平上发生一系列改变以适应环境.热激蛋白(heat shock protein, HSP)是一类重要的响应外界环境变化的蛋白家族, 小热激蛋白(small heat shock protein, sHSP)是其中一类分子量较小的热激蛋白(刘德立, 1996; 黄祥富等, 1999; Wang et al., 2004; 栗振义等, 2016).目前已在古生菌、细菌和真核生物中发现sHSP成员(Waters and Vierling, 2020).sHSP成员具有保守的α-晶体结构域, 该结构域由80-100个氨基酸残基构成的结构紧凑的β链组成(van Montfort et al., 2001).在植物中, 首先在烟草(Nicotiana tabacum)和大豆(Glycine max)中鉴定出sHSP成员(Key et al., 1981), 随后相继在拟南芥(Scharf et al., 2001)、水稻(Sarkar et al., 2009)、葡萄(Ji et al., 2019)、番茄(Solanum lycopersicum) (张宁等, 2017)、马铃薯(S. tuberosum) (Zhao et al., 2018)、棉花(Gossypium hirsutum) (Ma et al., 2016)、银杏(Ginkgo biloba) (何福林和张斌, 2019)、辣椒(Capsicum annuum) (Guo et al., 2015)和簸箕柳 ...
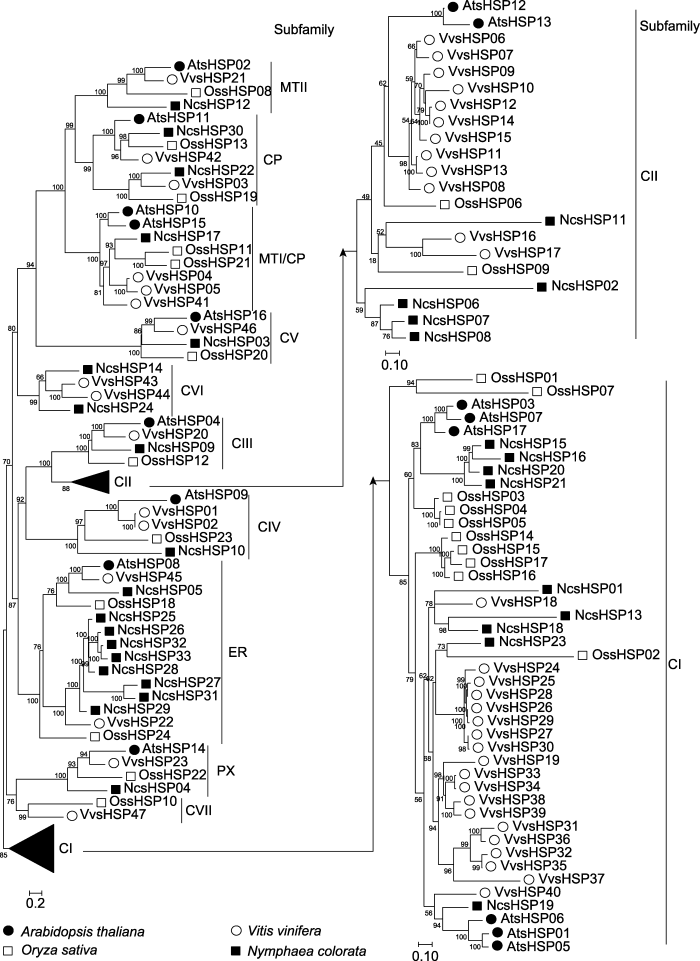
... 本研究在睡莲、水稻、拟南芥和葡萄中分别鉴定到33个NcsHSPs、24个OssHSPs、17个AtsHSPs和47个VvsHSPs (表1).4种植物中, 在葡萄中鉴定到的sHSP成员最多, 而在拟南芥中鉴定到的sHSP成员最少, 这主要与4种植物基因组大小有关(The French-Italian Public Consortium for Grapevine Genome Characterization, 2007; Ouyang et al., 2007; Lamesch et al., 2012; Zhang et al., 2020a).通过系统进化分析, 我们将sHSP家族进一步分为12个亚家族(图1).不同的亚家族含有不同数目的sHSP成员, 其中CI亚家族包含最多的sHSP成员, 这可能与CI亚家族包含最多的基因复制事件(4个)有关(表2).相似的分布也出现在番茄(张宁等, 2017)、马铃薯(Zhao et al., 2018)、棉花(Ma et al., 2016)、银杏(何福林和张斌, 2019)和辣椒(Guo et al., 2015)等植物中.因此, 在植物中sHSP家族具有保守的亚家族分布.在拟南芥中未发现任何属于CVI和CVII亚家族的成员, 而在葡萄中CI和CII亚家族的成员数目远多于另外3个物种, 这直接导致拟南芥和葡萄中含有不同数目的sHSP成员.类似现象也出现在棉花扩展蛋白基因家族(张奇艳等, 2019)、玉米(Zea mays) NAC家族(Fan et al., 2014)及大豆PP2C家族(Fan et al., 2020a).sHSP家族的保守基序分布和基因结构分布也与通过系统进化分析得出的亚家族分布一致(图2). ...
植物热激蛋白的功能及其基因表达的调控 1 1999
... 在外界环境发生变化时, 植物在生理和分子水平上发生一系列改变以适应环境.热激蛋白(heat shock protein, HSP)是一类重要的响应外界环境变化的蛋白家族, 小热激蛋白(small heat shock protein, sHSP)是其中一类分子量较小的热激蛋白(刘德立, 1996; 黄祥富等, 1999; Wang et al., 2004; 栗振义等, 2016).目前已在古生菌、细菌和真核生物中发现sHSP成员(Waters and Vierling, 2020).sHSP成员具有保守的α-晶体结构域, 该结构域由80-100个氨基酸残基构成的结构紧凑的β链组成(van Montfort et al., 2001).在植物中, 首先在烟草(Nicotiana tabacum)和大豆(Glycine max)中鉴定出sHSP成员(Key et al., 1981), 随后相继在拟南芥(Scharf et al., 2001)、水稻(Sarkar et al., 2009)、葡萄(Ji et al., 2019)、番茄(Solanum lycopersicum) (张宁等, 2017)、马铃薯(S. tuberosum) (Zhao et al., 2018)、棉花(Gossypium hirsutum) (Ma et al., 2016)、银杏(Ginkgo biloba) (何福林和张斌, 2019)、辣椒(Capsicum annuum) (Guo et al., 2015)和簸箕柳 ...
植物热激蛋白研究进展 1 2016
... 在外界环境发生变化时, 植物在生理和分子水平上发生一系列改变以适应环境.热激蛋白(heat shock protein, HSP)是一类重要的响应外界环境变化的蛋白家族, 小热激蛋白(small heat shock protein, sHSP)是其中一类分子量较小的热激蛋白(刘德立, 1996; 黄祥富等, 1999; Wang et al., 2004; 栗振义等, 2016).目前已在古生菌、细菌和真核生物中发现sHSP成员(Waters and Vierling, 2020).sHSP成员具有保守的α-晶体结构域, 该结构域由80-100个氨基酸残基构成的结构紧凑的β链组成(van Montfort et al., 2001).在植物中, 首先在烟草(Nicotiana tabacum)和大豆(Glycine max)中鉴定出sHSP成员(Key et al., 1981), 随后相继在拟南芥(Scharf et al., 2001)、水稻(Sarkar et al., 2009)、葡萄(Ji et al., 2019)、番茄(Solanum lycopersicum) (张宁等, 2017)、马铃薯(S. tuberosum) (Zhao et al., 2018)、棉花(Gossypium hirsutum) (Ma et al., 2016)、银杏(Ginkgo biloba) (何福林和张斌, 2019)、辣椒(Capsicum annuum) (Guo et al., 2015)和簸箕柳 ...
植物热激蛋白及其功能 1 1996
... 在外界环境发生变化时, 植物在生理和分子水平上发生一系列改变以适应环境.热激蛋白(heat shock protein, HSP)是一类重要的响应外界环境变化的蛋白家族, 小热激蛋白(small heat shock protein, sHSP)是其中一类分子量较小的热激蛋白(刘德立, 1996; 黄祥富等, 1999; Wang et al., 2004; 栗振义等, 2016).目前已在古生菌、细菌和真核生物中发现sHSP成员(Waters and Vierling, 2020).sHSP成员具有保守的α-晶体结构域, 该结构域由80-100个氨基酸残基构成的结构紧凑的β链组成(van Montfort et al., 2001).在植物中, 首先在烟草(Nicotiana tabacum)和大豆(Glycine max)中鉴定出sHSP成员(Key et al., 1981), 随后相继在拟南芥(Scharf et al., 2001)、水稻(Sarkar et al., 2009)、葡萄(Ji et al., 2019)、番茄(Solanum lycopersicum) (张宁等, 2017)、马铃薯(S. tuberosum) (Zhao et al., 2018)、棉花(Gossypium hirsutum) (Ma et al., 2016)、银杏(Ginkgo biloba) (何福林和张斌, 2019)、辣椒(Capsicum annuum) (Guo et al., 2015)和簸箕柳 ...
番茄HSP20基因家族的全基因组鉴定、系统进化及表达分析 2 2017
... 在外界环境发生变化时, 植物在生理和分子水平上发生一系列改变以适应环境.热激蛋白(heat shock protein, HSP)是一类重要的响应外界环境变化的蛋白家族, 小热激蛋白(small heat shock protein, sHSP)是其中一类分子量较小的热激蛋白(刘德立, 1996; 黄祥富等, 1999; Wang et al., 2004; 栗振义等, 2016).目前已在古生菌、细菌和真核生物中发现sHSP成员(Waters and Vierling, 2020).sHSP成员具有保守的α-晶体结构域, 该结构域由80-100个氨基酸残基构成的结构紧凑的β链组成(van Montfort et al., 2001).在植物中, 首先在烟草(Nicotiana tabacum)和大豆(Glycine max)中鉴定出sHSP成员(Key et al., 1981), 随后相继在拟南芥(Scharf et al., 2001)、水稻(Sarkar et al., 2009)、葡萄(Ji et al., 2019)、番茄(Solanum lycopersicum) (张宁等, 2017)、马铃薯(S. tuberosum) (Zhao et al., 2018)、棉花(Gossypium hirsutum) (Ma et al., 2016)、银杏(Ginkgo biloba) (何福林和张斌, 2019)、辣椒(Capsicum annuum) (Guo et al., 2015)和簸箕柳 ...
... 本研究在睡莲、水稻、拟南芥和葡萄中分别鉴定到33个NcsHSPs、24个OssHSPs、17个AtsHSPs和47个VvsHSPs (表1).4种植物中, 在葡萄中鉴定到的sHSP成员最多, 而在拟南芥中鉴定到的sHSP成员最少, 这主要与4种植物基因组大小有关(The French-Italian Public Consortium for Grapevine Genome Characterization, 2007; Ouyang et al., 2007; Lamesch et al., 2012; Zhang et al., 2020a).通过系统进化分析, 我们将sHSP家族进一步分为12个亚家族(图1).不同的亚家族含有不同数目的sHSP成员, 其中CI亚家族包含最多的sHSP成员, 这可能与CI亚家族包含最多的基因复制事件(4个)有关(表2).相似的分布也出现在番茄(张宁等, 2017)、马铃薯(Zhao et al., 2018)、棉花(Ma et al., 2016)、银杏(何福林和张斌, 2019)和辣椒(Guo et al., 2015)等植物中.因此, 在植物中sHSP家族具有保守的亚家族分布.在拟南芥中未发现任何属于CVI和CVII亚家族的成员, 而在葡萄中CI和CII亚家族的成员数目远多于另外3个物种, 这直接导致拟南芥和葡萄中含有不同数目的sHSP成员.类似现象也出现在棉花扩展蛋白基因家族(张奇艳等, 2019)、玉米(Zea mays) NAC家族(Fan et al., 2014)及大豆PP2C家族(Fan et al., 2020a).sHSP家族的保守基序分布和基因结构分布也与通过系统进化分析得出的亚家族分布一致(图2). ...
陆地棉扩展蛋白基因的鉴定与特征分析 1 2019
... 本研究在睡莲、水稻、拟南芥和葡萄中分别鉴定到33个NcsHSPs、24个OssHSPs、17个AtsHSPs和47个VvsHSPs (表1).4种植物中, 在葡萄中鉴定到的sHSP成员最多, 而在拟南芥中鉴定到的sHSP成员最少, 这主要与4种植物基因组大小有关(The French-Italian Public Consortium for Grapevine Genome Characterization, 2007; Ouyang et al., 2007; Lamesch et al., 2012; Zhang et al., 2020a).通过系统进化分析, 我们将sHSP家族进一步分为12个亚家族(图1).不同的亚家族含有不同数目的sHSP成员, 其中CI亚家族包含最多的sHSP成员, 这可能与CI亚家族包含最多的基因复制事件(4个)有关(表2).相似的分布也出现在番茄(张宁等, 2017)、马铃薯(Zhao et al., 2018)、棉花(Ma et al., 2016)、银杏(何福林和张斌, 2019)和辣椒(Guo et al., 2015)等植物中.因此, 在植物中sHSP家族具有保守的亚家族分布.在拟南芥中未发现任何属于CVI和CVII亚家族的成员, 而在葡萄中CI和CII亚家族的成员数目远多于另外3个物种, 这直接导致拟南芥和葡萄中含有不同数目的sHSP成员.类似现象也出现在棉花扩展蛋白基因家族(张奇艳等, 2019)、玉米(Zea mays) NAC家族(Fan et al., 2014)及大豆PP2C家族(Fan et al., 2020a).sHSP家族的保守基序分布和基因结构分布也与通过系统进化分析得出的亚家族分布一致(图2). ...
The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana 1 2004
... 在睡莲、水稻、拟南芥和葡萄中共鉴定到12个与sHSP家族有关的基因复制事件(表2).相比其它2种植物, 在葡萄和拟南芥中发生的基因复制事件最多(各4个).葡萄中sHSP成员数量最多可能与葡萄发生的基因复制事件最多有关, 相似的研究结果也出现在葡萄的CDPL家族(Zhang et al., 2015)、卵形家族蛋白(Wang et al., 2018)以及WRKY家族中(Guo et al., 2014).在拟南芥中, 虽然与AtsHSP成员相关的基因复制事件最多, 但成员数量较少, 这可能与拟南芥基因复制事件后大量基因丢失有关(Cannon et al., 2004; Maere et al., 2005).结合染色体定位结果, 可以将12个与sHSP家族有关的基因复制事件进一步分为6个片段复制事件和6个串联复制事件, 并且每个物种中均含有这2种基因复制事件, 这表明sHSP家族的扩增不仅与大规模的片段复制事件有关, 还与小规模的串联复制事件有关(图3).相似的sHSP家族扩增模式也出现在辣椒(Guo et al., 2015)、簸箕柳(Li et al., 2018)和马铃薯(Zhao et al., 2018)等植物中.在每个物种中, 片段复制事件的Ks值均大于串联复制事件的Ks值, 这表明在sHSP家族中片段复制事件发生的时间均早于串联复制事件, 相似的现象也出现在梅花(Prunus mume) NAC家族(Zhuo et al., 2018)和棉花PP2C家族(Shazadee et al., 2019)中.基于复制事件的Ka/Ks比值表明, sHSP成员在发生复制事件以后会发生强烈的纯化选择作用, 类似的Ka/Ks比值分布也出现在其它植物的sHSP家族中(Ma et al., 2016; Li et al., 2018).与此同时, 与其它基因家族相比(Zhuo et al., 2018; Fan et al., 2020b), sHSP家族发生的基因复制事件较少(表2), 这也直接导致sHSP家族成员数相对较少, 表明在植物的进化过程中, sHSP成员可能具有相对保守的功能(如响应高温胁迫) (Waters and Vierling, 2020).此外, 本研究发现4个片段复制事件的Ks值大于1 (表2), 其中在拟南芥中有1个片段复制事件(AtsHSP01/AtsHSP06), 在另外3种植物中均发现对应的同源基因(图4), 该片段复制事件发生的时间大约为1-1.2亿年前温暖干旱的白垩纪(Zhang et al., 2020b), 这表明AtsHSP01/AtsHSP06及其在植物界的同源基因可能与植物适应高温环境有关, 可以作为今后研究高温胁迫的重要候选基因. ...
TBtools: an integrative toolkit developed for interactive analyses of big biological data 2 2020
... 采用MCScan软件使用默认参数分别对睡莲、水稻、拟南芥和葡萄中sHSP成员的基因复制事件进行识别.在睡莲、水稻、拟南芥和葡萄中, 同源基因的识别主要基于MCScan软件获得的共线性结果进行确定, 同源基因之间的共线性关系主要采用MCScan软件进行可视化分析.此外, 复制基因之间以及同源基因之间的非同义替换率(Ka)、同义替换率(Ks)以及它们的比值(Ka/Ks)使用TBtools软件计算获得(Chen et al., 2020). ...
Non-redundant functions of sHSP-CIs in acquired thermotolerance and their role in early seed development in Arabidopsis 1 2008
... 之前的研究表明, sHSP家族可以参与许多植物生长与发育过程(Zhang et al., 2018; Ma et al., 2019).本研究在睡莲、水稻、拟南芥和葡萄中新发现一些与植物生长发育过程相关的sHSP成员(图5).CI亚家族的NcsHSP19、OssHSP02、OssHSP14和VvsHSP40以及CIV亚家族的AtsHSP09和OssHSP23在几乎所有组织中均高度表达, 其在棉花和马铃薯中的同源基因也具有类似的表达模式(Ma et al., 2016; Zhao et al., 2018), 这表明sHSP家族在植物中的功能可能具有一定的保守性.此外, 本研究还发现一些与水稻花器官发育相关的基因(OssHSP03、OssHSP04、OssHSP06、OssHSP12、OssHSP16和OssHSP21).有报道显示, 在玫瑰(Rosa hybrida)中与CI亚家族OssHSP03、OssHSP04和OssHSP16相同亚家族的sHSP17.5-CI可能与开花过程中保护细胞蛋白有关(Dafny-Yelin et al., 2008). ...
Pervasive duplication, biased molecular evolution and comprehensive functional analysis of the PP2C family in Glycine max 2 2020a
... 本研究在睡莲、水稻、拟南芥和葡萄中分别鉴定到33个NcsHSPs、24个OssHSPs、17个AtsHSPs和47个VvsHSPs (表1).4种植物中, 在葡萄中鉴定到的sHSP成员最多, 而在拟南芥中鉴定到的sHSP成员最少, 这主要与4种植物基因组大小有关(The French-Italian Public Consortium for Grapevine Genome Characterization, 2007; Ouyang et al., 2007; Lamesch et al., 2012; Zhang et al., 2020a).通过系统进化分析, 我们将sHSP家族进一步分为12个亚家族(图1).不同的亚家族含有不同数目的sHSP成员, 其中CI亚家族包含最多的sHSP成员, 这可能与CI亚家族包含最多的基因复制事件(4个)有关(表2).相似的分布也出现在番茄(张宁等, 2017)、马铃薯(Zhao et al., 2018)、棉花(Ma et al., 2016)、银杏(何福林和张斌, 2019)和辣椒(Guo et al., 2015)等植物中.因此, 在植物中sHSP家族具有保守的亚家族分布.在拟南芥中未发现任何属于CVI和CVII亚家族的成员, 而在葡萄中CI和CII亚家族的成员数目远多于另外3个物种, 这直接导致拟南芥和葡萄中含有不同数目的sHSP成员.类似现象也出现在棉花扩展蛋白基因家族(张奇艳等, 2019)、玉米(Zea mays) NAC家族(Fan et al., 2014)及大豆PP2C家族(Fan et al., 2020a).sHSP家族的保守基序分布和基因结构分布也与通过系统进化分析得出的亚家族分布一致(图2). ...
Molecular evolution and expansion of the KUP family in the allopolyploid cotton species Gossypium hirsutum and Gossypium barbadense 2 2020b
... 通过MAFFT软件对睡莲、水稻、拟南芥和葡萄中sHSP成员的氨基酸序列进行序列连配分析.MAFFT软件参数设置见本实验室之前的研究(Fan et al., 2020b).采用IQ-tree软件构建所有sHSP成员的系统进化树, 其中最适模型主要利用ModelFinder程序进行选择, bootstrap值设定为1 000. ...
... 在睡莲、水稻、拟南芥和葡萄中共鉴定到12个与sHSP家族有关的基因复制事件(表2).相比其它2种植物, 在葡萄和拟南芥中发生的基因复制事件最多(各4个).葡萄中sHSP成员数量最多可能与葡萄发生的基因复制事件最多有关, 相似的研究结果也出现在葡萄的CDPL家族(Zhang et al., 2015)、卵形家族蛋白(Wang et al., 2018)以及WRKY家族中(Guo et al., 2014).在拟南芥中, 虽然与AtsHSP成员相关的基因复制事件最多, 但成员数量较少, 这可能与拟南芥基因复制事件后大量基因丢失有关(Cannon et al., 2004; Maere et al., 2005).结合染色体定位结果, 可以将12个与sHSP家族有关的基因复制事件进一步分为6个片段复制事件和6个串联复制事件, 并且每个物种中均含有这2种基因复制事件, 这表明sHSP家族的扩增不仅与大规模的片段复制事件有关, 还与小规模的串联复制事件有关(图3).相似的sHSP家族扩增模式也出现在辣椒(Guo et al., 2015)、簸箕柳(Li et al., 2018)和马铃薯(Zhao et al., 2018)等植物中.在每个物种中, 片段复制事件的Ks值均大于串联复制事件的Ks值, 这表明在sHSP家族中片段复制事件发生的时间均早于串联复制事件, 相似的现象也出现在梅花(Prunus mume) NAC家族(Zhuo et al., 2018)和棉花PP2C家族(Shazadee et al., 2019)中.基于复制事件的Ka/Ks比值表明, sHSP成员在发生复制事件以后会发生强烈的纯化选择作用, 类似的Ka/Ks比值分布也出现在其它植物的sHSP家族中(Ma et al., 2016; Li et al., 2018).与此同时, 与其它基因家族相比(Zhuo et al., 2018; Fan et al., 2020b), sHSP家族发生的基因复制事件较少(表2), 这也直接导致sHSP家族成员数相对较少, 表明在植物的进化过程中, sHSP成员可能具有相对保守的功能(如响应高温胁迫) (Waters and Vierling, 2020).此外, 本研究发现4个片段复制事件的Ks值大于1 (表2), 其中在拟南芥中有1个片段复制事件(AtsHSP01/AtsHSP06), 在另外3种植物中均发现对应的同源基因(图4), 该片段复制事件发生的时间大约为1-1.2亿年前温暖干旱的白垩纪(Zhang et al., 2020b), 这表明AtsHSP01/AtsHSP06及其在植物界的同源基因可能与植物适应高温环境有关, 可以作为今后研究高温胁迫的重要候选基因. ...
Molecular evolution and species-specific expansion of the NAP members in plants 1 2015
... 睡莲是现存的早期被子植物之一(Zhang et al., 2020a), 通过与另外3种模式被子植物的sHSP家族进行对比, 发现睡莲sHSP家族中氨基酸长度较长以及分子量较大的成员较多(表1), 其次为具有很多古代植物基因组特征的葡萄sHSP成员(The French- Italian Public Consortium for Grapevine Genome Characterization, 2007), 表明sHSP家族在被子植物中可能向更短的氨基酸长度和更小的分子量进化, 这种进化趋势与已报道的rhomboid家族不同(Li et al., 2015), 但与PG家族类似(Park and Kim, 2010).其次, 水稻、拟南芥和葡萄大部分sHSP成员的内含子不多于1个(图2), 而睡莲含有较多基因结构较为复杂的sHSP成员, 这表明sHSP家族在被子植物中可能向更简单的基因结构进化, 这种趋势与已报道的NAP亚家族不同(Fan et al., 2015).此外, 睡莲sHSP成员分布在12条染色体上(图3), 葡萄sHSP成员分布在11条染色体上, 而拟南芥和水稻的sHSP成员分别分布在5条和8条染色体上, 表明sHSP成员在被子植物中可能更集中分布于某几条染色体上, 这种集中分布模式可能更有利于sHSP成员协同发挥作用. ...
Molecular evolution and expansion analysis of the NAC transcription factor in Zea mays 1 2014
... 本研究在睡莲、水稻、拟南芥和葡萄中分别鉴定到33个NcsHSPs、24个OssHSPs、17个AtsHSPs和47个VvsHSPs (表1).4种植物中, 在葡萄中鉴定到的sHSP成员最多, 而在拟南芥中鉴定到的sHSP成员最少, 这主要与4种植物基因组大小有关(The French-Italian Public Consortium for Grapevine Genome Characterization, 2007; Ouyang et al., 2007; Lamesch et al., 2012; Zhang et al., 2020a).通过系统进化分析, 我们将sHSP家族进一步分为12个亚家族(图1).不同的亚家族含有不同数目的sHSP成员, 其中CI亚家族包含最多的sHSP成员, 这可能与CI亚家族包含最多的基因复制事件(4个)有关(表2).相似的分布也出现在番茄(张宁等, 2017)、马铃薯(Zhao et al., 2018)、棉花(Ma et al., 2016)、银杏(何福林和张斌, 2019)和辣椒(Guo et al., 2015)等植物中.因此, 在植物中sHSP家族具有保守的亚家族分布.在拟南芥中未发现任何属于CVI和CVII亚家族的成员, 而在葡萄中CI和CII亚家族的成员数目远多于另外3个物种, 这直接导致拟南芥和葡萄中含有不同数目的sHSP成员.类似现象也出现在棉花扩展蛋白基因家族(张奇艳等, 2019)、玉米(Zea mays) NAC家族(Fan et al., 2014)及大豆PP2C家族(Fan et al., 2020a).sHSP家族的保守基序分布和基因结构分布也与通过系统进化分析得出的亚家族分布一致(图2). ...
Evolution and expression analysis of the grape (Vitis vinifera L.) WRKY gene family 1 2014
... 在睡莲、水稻、拟南芥和葡萄中共鉴定到12个与sHSP家族有关的基因复制事件(表2).相比其它2种植物, 在葡萄和拟南芥中发生的基因复制事件最多(各4个).葡萄中sHSP成员数量最多可能与葡萄发生的基因复制事件最多有关, 相似的研究结果也出现在葡萄的CDPL家族(Zhang et al., 2015)、卵形家族蛋白(Wang et al., 2018)以及WRKY家族中(Guo et al., 2014).在拟南芥中, 虽然与AtsHSP成员相关的基因复制事件最多, 但成员数量较少, 这可能与拟南芥基因复制事件后大量基因丢失有关(Cannon et al., 2004; Maere et al., 2005).结合染色体定位结果, 可以将12个与sHSP家族有关的基因复制事件进一步分为6个片段复制事件和6个串联复制事件, 并且每个物种中均含有这2种基因复制事件, 这表明sHSP家族的扩增不仅与大规模的片段复制事件有关, 还与小规模的串联复制事件有关(图3).相似的sHSP家族扩增模式也出现在辣椒(Guo et al., 2015)、簸箕柳(Li et al., 2018)和马铃薯(Zhao et al., 2018)等植物中.在每个物种中, 片段复制事件的Ks值均大于串联复制事件的Ks值, 这表明在sHSP家族中片段复制事件发生的时间均早于串联复制事件, 相似的现象也出现在梅花(Prunus mume) NAC家族(Zhuo et al., 2018)和棉花PP2C家族(Shazadee et al., 2019)中.基于复制事件的Ka/Ks比值表明, sHSP成员在发生复制事件以后会发生强烈的纯化选择作用, 类似的Ka/Ks比值分布也出现在其它植物的sHSP家族中(Ma et al., 2016; Li et al., 2018).与此同时, 与其它基因家族相比(Zhuo et al., 2018; Fan et al., 2020b), sHSP家族发生的基因复制事件较少(表2), 这也直接导致sHSP家族成员数相对较少, 表明在植物的进化过程中, sHSP成员可能具有相对保守的功能(如响应高温胁迫) (Waters and Vierling, 2020).此外, 本研究发现4个片段复制事件的Ks值大于1 (表2), 其中在拟南芥中有1个片段复制事件(AtsHSP01/AtsHSP06), 在另外3种植物中均发现对应的同源基因(图4), 该片段复制事件发生的时间大约为1-1.2亿年前温暖干旱的白垩纪(Zhang et al., 2020b), 这表明AtsHSP01/AtsHSP06及其在植物界的同源基因可能与植物适应高温环境有关, 可以作为今后研究高温胁迫的重要候选基因. ...
A class I cytosolic HSP20 of rice enhances heat and salt tolerance in different organisms 1 2020
... (Salix suchowensis) (Li et al., 2018)等植物中发现了大量的sHSP成员.sHSP家族主要作为分子伴侣, 以独立于ATP的形式保护植物免受外界环境的影响(Waters and Vierling, 2020).大量研究表明, sHSP成员可以响应外界多种环境胁迫(Sun et al., 2010; Zhu et al., 2011; Li et al., 2016; Zhai et al., 2016; Guo et al., 2020).此外, sHSP家族也参与果实品质形成和植株发芽等植物生长发育过程(Zhang et al., 2018; Ma et al., 2019). ...
Genome-wide analysis of the CaHsp20 gene family in pepper: comprehensive sequence and expression profile analysis under heat stress 3 2015
... 在外界环境发生变化时, 植物在生理和分子水平上发生一系列改变以适应环境.热激蛋白(heat shock protein, HSP)是一类重要的响应外界环境变化的蛋白家族, 小热激蛋白(small heat shock protein, sHSP)是其中一类分子量较小的热激蛋白(刘德立, 1996; 黄祥富等, 1999; Wang et al., 2004; 栗振义等, 2016).目前已在古生菌、细菌和真核生物中发现sHSP成员(Waters and Vierling, 2020).sHSP成员具有保守的α-晶体结构域, 该结构域由80-100个氨基酸残基构成的结构紧凑的β链组成(van Montfort et al., 2001).在植物中, 首先在烟草(Nicotiana tabacum)和大豆(Glycine max)中鉴定出sHSP成员(Key et al., 1981), 随后相继在拟南芥(Scharf et al., 2001)、水稻(Sarkar et al., 2009)、葡萄(Ji et al., 2019)、番茄(Solanum lycopersicum) (张宁等, 2017)、马铃薯(S. tuberosum) (Zhao et al., 2018)、棉花(Gossypium hirsutum) (Ma et al., 2016)、银杏(Ginkgo biloba) (何福林和张斌, 2019)、辣椒(Capsicum annuum) (Guo et al., 2015)和簸箕柳 ...
... 本研究在睡莲、水稻、拟南芥和葡萄中分别鉴定到33个NcsHSPs、24个OssHSPs、17个AtsHSPs和47个VvsHSPs (表1).4种植物中, 在葡萄中鉴定到的sHSP成员最多, 而在拟南芥中鉴定到的sHSP成员最少, 这主要与4种植物基因组大小有关(The French-Italian Public Consortium for Grapevine Genome Characterization, 2007; Ouyang et al., 2007; Lamesch et al., 2012; Zhang et al., 2020a).通过系统进化分析, 我们将sHSP家族进一步分为12个亚家族(图1).不同的亚家族含有不同数目的sHSP成员, 其中CI亚家族包含最多的sHSP成员, 这可能与CI亚家族包含最多的基因复制事件(4个)有关(表2).相似的分布也出现在番茄(张宁等, 2017)、马铃薯(Zhao et al., 2018)、棉花(Ma et al., 2016)、银杏(何福林和张斌, 2019)和辣椒(Guo et al., 2015)等植物中.因此, 在植物中sHSP家族具有保守的亚家族分布.在拟南芥中未发现任何属于CVI和CVII亚家族的成员, 而在葡萄中CI和CII亚家族的成员数目远多于另外3个物种, 这直接导致拟南芥和葡萄中含有不同数目的sHSP成员.类似现象也出现在棉花扩展蛋白基因家族(张奇艳等, 2019)、玉米(Zea mays) NAC家族(Fan et al., 2014)及大豆PP2C家族(Fan et al., 2020a).sHSP家族的保守基序分布和基因结构分布也与通过系统进化分析得出的亚家族分布一致(图2). ...
... 在睡莲、水稻、拟南芥和葡萄中共鉴定到12个与sHSP家族有关的基因复制事件(表2).相比其它2种植物, 在葡萄和拟南芥中发生的基因复制事件最多(各4个).葡萄中sHSP成员数量最多可能与葡萄发生的基因复制事件最多有关, 相似的研究结果也出现在葡萄的CDPL家族(Zhang et al., 2015)、卵形家族蛋白(Wang et al., 2018)以及WRKY家族中(Guo et al., 2014).在拟南芥中, 虽然与AtsHSP成员相关的基因复制事件最多, 但成员数量较少, 这可能与拟南芥基因复制事件后大量基因丢失有关(Cannon et al., 2004; Maere et al., 2005).结合染色体定位结果, 可以将12个与sHSP家族有关的基因复制事件进一步分为6个片段复制事件和6个串联复制事件, 并且每个物种中均含有这2种基因复制事件, 这表明sHSP家族的扩增不仅与大规模的片段复制事件有关, 还与小规模的串联复制事件有关(图3).相似的sHSP家族扩增模式也出现在辣椒(Guo et al., 2015)、簸箕柳(Li et al., 2018)和马铃薯(Zhao et al., 2018)等植物中.在每个物种中, 片段复制事件的Ks值均大于串联复制事件的Ks值, 这表明在sHSP家族中片段复制事件发生的时间均早于串联复制事件, 相似的现象也出现在梅花(Prunus mume) NAC家族(Zhuo et al., 2018)和棉花PP2C家族(Shazadee et al., 2019)中.基于复制事件的Ka/Ks比值表明, sHSP成员在发生复制事件以后会发生强烈的纯化选择作用, 类似的Ka/Ks比值分布也出现在其它植物的sHSP家族中(Ma et al., 2016; Li et al., 2018).与此同时, 与其它基因家族相比(Zhuo et al., 2018; Fan et al., 2020b), sHSP家族发生的基因复制事件较少(表2), 这也直接导致sHSP家族成员数相对较少, 表明在植物的进化过程中, sHSP成员可能具有相对保守的功能(如响应高温胁迫) (Waters and Vierling, 2020).此外, 本研究发现4个片段复制事件的Ks值大于1 (表2), 其中在拟南芥中有1个片段复制事件(AtsHSP01/AtsHSP06), 在另外3种植物中均发现对应的同源基因(图4), 该片段复制事件发生的时间大约为1-1.2亿年前温暖干旱的白垩纪(Zhang et al., 2020b), 这表明AtsHSP01/AtsHSP06及其在植物界的同源基因可能与植物适应高温环境有关, 可以作为今后研究高温胁迫的重要候选基因. ...
Genome-wide identification of small heat-shock protein (HSP20) gene family in grape and expression profile during berry development 1 2019
... 在外界环境发生变化时, 植物在生理和分子水平上发生一系列改变以适应环境.热激蛋白(heat shock protein, HSP)是一类重要的响应外界环境变化的蛋白家族, 小热激蛋白(small heat shock protein, sHSP)是其中一类分子量较小的热激蛋白(刘德立, 1996; 黄祥富等, 1999; Wang et al., 2004; 栗振义等, 2016).目前已在古生菌、细菌和真核生物中发现sHSP成员(Waters and Vierling, 2020).sHSP成员具有保守的α-晶体结构域, 该结构域由80-100个氨基酸残基构成的结构紧凑的β链组成(van Montfort et al., 2001).在植物中, 首先在烟草(Nicotiana tabacum)和大豆(Glycine max)中鉴定出sHSP成员(Key et al., 1981), 随后相继在拟南芥(Scharf et al., 2001)、水稻(Sarkar et al., 2009)、葡萄(Ji et al., 2019)、番茄(Solanum lycopersicum) (张宁等, 2017)、马铃薯(S. tuberosum) (Zhao et al., 2018)、棉花(Gossypium hirsutum) (Ma et al., 2016)、银杏(Ginkgo biloba) (何福林和张斌, 2019)、辣椒(Capsicum annuum) (Guo et al., 2015)和簸箕柳 ...
A genome triplication associated with early diversification of the core eudicots 1 2012
... 对睡莲、水稻、拟南芥和葡萄中sHSP家族之间的共线性分析(图4)表明, 拟南芥和葡萄中的同源基因对最多(10对), 这主要与拟南芥和葡萄之间具有较近的进化关系有关(Jiao et al., 2012).虽然睡莲(基部被子植物)和葡萄(双子叶植物)具有较远的进化距离, 但其sHSP成员却有较多的同源基因对(8对), 这主要与葡萄基因组并未发生最近的基因复制事件从而导致基因组有很多古代植物基因组的特征有关(The French-Italian Public Consortium for Grapevine Genome Characterization, 2007; Zhang et al., 2020a).此外, 本研究通过Ka值计算发现同源基因之间的进化同步, 即在植物中sHSP家族的进化相对保守, 这可能与sHSP家族的功能有一定关系(Waters and Vierling, 2020). ...
Heat shock proteins of higher plants 1 1981
... 在外界环境发生变化时, 植物在生理和分子水平上发生一系列改变以适应环境.热激蛋白(heat shock protein, HSP)是一类重要的响应外界环境变化的蛋白家族, 小热激蛋白(small heat shock protein, sHSP)是其中一类分子量较小的热激蛋白(刘德立, 1996; 黄祥富等, 1999; Wang et al., 2004; 栗振义等, 2016).目前已在古生菌、细菌和真核生物中发现sHSP成员(Waters and Vierling, 2020).sHSP成员具有保守的α-晶体结构域, 该结构域由80-100个氨基酸残基构成的结构紧凑的β链组成(van Montfort et al., 2001).在植物中, 首先在烟草(Nicotiana tabacum)和大豆(Glycine max)中鉴定出sHSP成员(Key et al., 1981), 随后相继在拟南芥(Scharf et al., 2001)、水稻(Sarkar et al., 2009)、葡萄(Ji et al., 2019)、番茄(Solanum lycopersicum) (张宁等, 2017)、马铃薯(S. tuberosum) (Zhao et al., 2018)、棉花(Gossypium hirsutum) (Ma et al., 2016)、银杏(Ginkgo biloba) (何福林和张斌, 2019)、辣椒(Capsicum annuum) (Guo et al., 2015)和簸箕柳 ...
The Arabidopsis Information Resource (TAIR): improved gene annotation and new tools 3 2012
... 被子植物在日常生活中随处可见, 可为人们提供粮食、纤维、燃料和药品等.被子植物的多样性与外界环境变化息息相关.在众多被子植物中, 基部被子植物睡莲(Nymphaea colorata) (Zhang et al., 2020a)、单子叶模式植物水稻(Oryza sativa) (Ouyang et al., 2007)、双子叶模式植物拟南芥(Arabidopsis thaliana) (Lamesch et al., 2012)以及类似古代双子叶植物基因组的葡萄(Vitis vinifera) (The French-Italian Public Consortium for Grapevine Genome Characterization, 2007)在被子植物分类学中具有重要意义.目前, 睡莲、水稻、拟南芥和葡萄的基因组测序已经完成, 从与外界环境变化相关的基因家族层面上对这4类被子植物进化历程进行研究, 不仅有助于解析被子植物进化的方向, 而且可为后续的分子育种提供重要参考. ...
... 目前, 虽然已在许多植物中鉴定到sHSP成员, 但主要集中在对单个物种进行全基因组分析, 缺乏对植物整体进化趋势的认识.睡莲、水稻、拟南芥和葡萄在被子植物进化过程中具有重要意义.目前, 睡莲(Zhang et al., 2020a)、水稻(Ouyang et al., 2007)、拟南芥(Lamesch et al., 2012)和葡萄(The French- Italian Public Consortium for Grapevine Genome Characterization, 2007)的全基因组测序、组装以及注释工作已经完成, 这为在4种被子植物中进行sHSP家族的比较基因组学分析奠定了基础.本研究利用最新的睡莲、水稻、拟南芥和葡萄的全基因组测序信息, 对其sHSP家族进行鉴定, 并在此基础上进一步对其系统发育、保守基序、基因结构、染色体定位、基因复制事件、共线性和基因表达等方面进行比较基因组学分析, 旨在为从基因家族层面进一步揭示植物的进化方向提供重要参考, 同时也为分子育种提供具有潜在育种价值的候选基因. ...
... 本研究在睡莲、水稻、拟南芥和葡萄中分别鉴定到33个NcsHSPs、24个OssHSPs、17个AtsHSPs和47个VvsHSPs (表1).4种植物中, 在葡萄中鉴定到的sHSP成员最多, 而在拟南芥中鉴定到的sHSP成员最少, 这主要与4种植物基因组大小有关(The French-Italian Public Consortium for Grapevine Genome Characterization, 2007; Ouyang et al., 2007; Lamesch et al., 2012; Zhang et al., 2020a).通过系统进化分析, 我们将sHSP家族进一步分为12个亚家族(图1).不同的亚家族含有不同数目的sHSP成员, 其中CI亚家族包含最多的sHSP成员, 这可能与CI亚家族包含最多的基因复制事件(4个)有关(表2).相似的分布也出现在番茄(张宁等, 2017)、马铃薯(Zhao et al., 2018)、棉花(Ma et al., 2016)、银杏(何福林和张斌, 2019)和辣椒(Guo et al., 2015)等植物中.因此, 在植物中sHSP家族具有保守的亚家族分布.在拟南芥中未发现任何属于CVI和CVII亚家族的成员, 而在葡萄中CI和CII亚家族的成员数目远多于另外3个物种, 这直接导致拟南芥和葡萄中含有不同数目的sHSP成员.类似现象也出现在棉花扩展蛋白基因家族(张奇艳等, 2019)、玉米(Zea mays) NAC家族(Fan et al., 2014)及大豆PP2C家族(Fan et al., 2020a).sHSP家族的保守基序分布和基因结构分布也与通过系统进化分析得出的亚家族分布一致(图2). ...
The Populus trichocarpa PtHSP17.8 involved in heat and salt stress tolerances 1 2016
... (Salix suchowensis) (Li et al., 2018)等植物中发现了大量的sHSP成员.sHSP家族主要作为分子伴侣, 以独立于ATP的形式保护植物免受外界环境的影响(Waters and Vierling, 2020).大量研究表明, sHSP成员可以响应外界多种环境胁迫(Sun et al., 2010; Zhu et al., 2011; Li et al., 2016; Zhai et al., 2016; Guo et al., 2020).此外, sHSP家族也参与果实品质形成和植株发芽等植物生长发育过程(Zhang et al., 2018; Ma et al., 2019). ...
Genome-wide characterization of the sHsp gene family in Salix suchowensis reveals its functions under different abiotic stresses 3 2018
... (Salix suchowensis) (Li et al., 2018)等植物中发现了大量的sHSP成员.sHSP家族主要作为分子伴侣, 以独立于ATP的形式保护植物免受外界环境的影响(Waters and Vierling, 2020).大量研究表明, sHSP成员可以响应外界多种环境胁迫(Sun et al., 2010; Zhu et al., 2011; Li et al., 2016; Zhai et al., 2016; Guo et al., 2020).此外, sHSP家族也参与果实品质形成和植株发芽等植物生长发育过程(Zhang et al., 2018; Ma et al., 2019). ...
... 在睡莲、水稻、拟南芥和葡萄中共鉴定到12个与sHSP家族有关的基因复制事件(表2).相比其它2种植物, 在葡萄和拟南芥中发生的基因复制事件最多(各4个).葡萄中sHSP成员数量最多可能与葡萄发生的基因复制事件最多有关, 相似的研究结果也出现在葡萄的CDPL家族(Zhang et al., 2015)、卵形家族蛋白(Wang et al., 2018)以及WRKY家族中(Guo et al., 2014).在拟南芥中, 虽然与AtsHSP成员相关的基因复制事件最多, 但成员数量较少, 这可能与拟南芥基因复制事件后大量基因丢失有关(Cannon et al., 2004; Maere et al., 2005).结合染色体定位结果, 可以将12个与sHSP家族有关的基因复制事件进一步分为6个片段复制事件和6个串联复制事件, 并且每个物种中均含有这2种基因复制事件, 这表明sHSP家族的扩增不仅与大规模的片段复制事件有关, 还与小规模的串联复制事件有关(图3).相似的sHSP家族扩增模式也出现在辣椒(Guo et al., 2015)、簸箕柳(Li et al., 2018)和马铃薯(Zhao et al., 2018)等植物中.在每个物种中, 片段复制事件的Ks值均大于串联复制事件的Ks值, 这表明在sHSP家族中片段复制事件发生的时间均早于串联复制事件, 相似的现象也出现在梅花(Prunus mume) NAC家族(Zhuo et al., 2018)和棉花PP2C家族(Shazadee et al., 2019)中.基于复制事件的Ka/Ks比值表明, sHSP成员在发生复制事件以后会发生强烈的纯化选择作用, 类似的Ka/Ks比值分布也出现在其它植物的sHSP家族中(Ma et al., 2016; Li et al., 2018).与此同时, 与其它基因家族相比(Zhuo et al., 2018; Fan et al., 2020b), sHSP家族发生的基因复制事件较少(表2), 这也直接导致sHSP家族成员数相对较少, 表明在植物的进化过程中, sHSP成员可能具有相对保守的功能(如响应高温胁迫) (Waters and Vierling, 2020).此外, 本研究发现4个片段复制事件的Ks值大于1 (表2), 其中在拟南芥中有1个片段复制事件(AtsHSP01/AtsHSP06), 在另外3种植物中均发现对应的同源基因(图4), 该片段复制事件发生的时间大约为1-1.2亿年前温暖干旱的白垩纪(Zhang et al., 2020b), 这表明AtsHSP01/AtsHSP06及其在植物界的同源基因可能与植物适应高温环境有关, 可以作为今后研究高温胁迫的重要候选基因. ...
... ; Li et al., 2018).与此同时, 与其它基因家族相比(Zhuo et al., 2018; Fan et al., 2020b), sHSP家族发生的基因复制事件较少(表2), 这也直接导致sHSP家族成员数相对较少, 表明在植物的进化过程中, sHSP成员可能具有相对保守的功能(如响应高温胁迫) (Waters and Vierling, 2020).此外, 本研究发现4个片段复制事件的Ks值大于1 (表2), 其中在拟南芥中有1个片段复制事件(AtsHSP01/AtsHSP06), 在另外3种植物中均发现对应的同源基因(图4), 该片段复制事件发生的时间大约为1-1.2亿年前温暖干旱的白垩纪(Zhang et al., 2020b), 这表明AtsHSP01/AtsHSP06及其在植物界的同源基因可能与植物适应高温环境有关, 可以作为今后研究高温胁迫的重要候选基因. ...
Differential evolution of members of the rhomboid gene family with conservative and divergent patterns 1 2015
... 睡莲是现存的早期被子植物之一(Zhang et al., 2020a), 通过与另外3种模式被子植物的sHSP家族进行对比, 发现睡莲sHSP家族中氨基酸长度较长以及分子量较大的成员较多(表1), 其次为具有很多古代植物基因组特征的葡萄sHSP成员(The French- Italian Public Consortium for Grapevine Genome Characterization, 2007), 表明sHSP家族在被子植物中可能向更短的氨基酸长度和更小的分子量进化, 这种进化趋势与已报道的rhomboid家族不同(Li et al., 2015), 但与PG家族类似(Park and Kim, 2010).其次, 水稻、拟南芥和葡萄大部分sHSP成员的内含子不多于1个(图2), 而睡莲含有较多基因结构较为复杂的sHSP成员, 这表明sHSP家族在被子植物中可能向更简单的基因结构进化, 这种趋势与已报道的NAP亚家族不同(Fan et al., 2015).此外, 睡莲sHSP成员分布在12条染色体上(图3), 葡萄sHSP成员分布在11条染色体上, 而拟南芥和水稻的sHSP成员分别分布在5条和8条染色体上, 表明sHSP成员在被子植物中可能更集中分布于某几条染色体上, 这种集中分布模式可能更有利于sHSP成员协同发挥作用. ...
Mitochondrial small heat shock protein mediates seed germination via thermal sensing 2 2019
... (Salix suchowensis) (Li et al., 2018)等植物中发现了大量的sHSP成员.sHSP家族主要作为分子伴侣, 以独立于ATP的形式保护植物免受外界环境的影响(Waters and Vierling, 2020).大量研究表明, sHSP成员可以响应外界多种环境胁迫(Sun et al., 2010; Zhu et al., 2011; Li et al., 2016; Zhai et al., 2016; Guo et al., 2020).此外, sHSP家族也参与果实品质形成和植株发芽等植物生长发育过程(Zhang et al., 2018; Ma et al., 2019). ...
... 之前的研究表明, sHSP家族可以参与许多植物生长与发育过程(Zhang et al., 2018; Ma et al., 2019).本研究在睡莲、水稻、拟南芥和葡萄中新发现一些与植物生长发育过程相关的sHSP成员(图5).CI亚家族的NcsHSP19、OssHSP02、OssHSP14和VvsHSP40以及CIV亚家族的AtsHSP09和OssHSP23在几乎所有组织中均高度表达, 其在棉花和马铃薯中的同源基因也具有类似的表达模式(Ma et al., 2016; Zhao et al., 2018), 这表明sHSP家族在植物中的功能可能具有一定的保守性.此外, 本研究还发现一些与水稻花器官发育相关的基因(OssHSP03、OssHSP04、OssHSP06、OssHSP12、OssHSP16和OssHSP21).有报道显示, 在玫瑰(Rosa hybrida)中与CI亚家族OssHSP03、OssHSP04和OssHSP16相同亚家族的sHSP17.5-CI可能与开花过程中保护细胞蛋白有关(Dafny-Yelin et al., 2008). ...
Identification and characterization of the GhHsp20 gene family in Gossypium hirsutum 4 2016
... 在外界环境发生变化时, 植物在生理和分子水平上发生一系列改变以适应环境.热激蛋白(heat shock protein, HSP)是一类重要的响应外界环境变化的蛋白家族, 小热激蛋白(small heat shock protein, sHSP)是其中一类分子量较小的热激蛋白(刘德立, 1996; 黄祥富等, 1999; Wang et al., 2004; 栗振义等, 2016).目前已在古生菌、细菌和真核生物中发现sHSP成员(Waters and Vierling, 2020).sHSP成员具有保守的α-晶体结构域, 该结构域由80-100个氨基酸残基构成的结构紧凑的β链组成(van Montfort et al., 2001).在植物中, 首先在烟草(Nicotiana tabacum)和大豆(Glycine max)中鉴定出sHSP成员(Key et al., 1981), 随后相继在拟南芥(Scharf et al., 2001)、水稻(Sarkar et al., 2009)、葡萄(Ji et al., 2019)、番茄(Solanum lycopersicum) (张宁等, 2017)、马铃薯(S. tuberosum) (Zhao et al., 2018)、棉花(Gossypium hirsutum) (Ma et al., 2016)、银杏(Ginkgo biloba) (何福林和张斌, 2019)、辣椒(Capsicum annuum) (Guo et al., 2015)和簸箕柳 ...
... 本研究在睡莲、水稻、拟南芥和葡萄中分别鉴定到33个NcsHSPs、24个OssHSPs、17个AtsHSPs和47个VvsHSPs (表1).4种植物中, 在葡萄中鉴定到的sHSP成员最多, 而在拟南芥中鉴定到的sHSP成员最少, 这主要与4种植物基因组大小有关(The French-Italian Public Consortium for Grapevine Genome Characterization, 2007; Ouyang et al., 2007; Lamesch et al., 2012; Zhang et al., 2020a).通过系统进化分析, 我们将sHSP家族进一步分为12个亚家族(图1).不同的亚家族含有不同数目的sHSP成员, 其中CI亚家族包含最多的sHSP成员, 这可能与CI亚家族包含最多的基因复制事件(4个)有关(表2).相似的分布也出现在番茄(张宁等, 2017)、马铃薯(Zhao et al., 2018)、棉花(Ma et al., 2016)、银杏(何福林和张斌, 2019)和辣椒(Guo et al., 2015)等植物中.因此, 在植物中sHSP家族具有保守的亚家族分布.在拟南芥中未发现任何属于CVI和CVII亚家族的成员, 而在葡萄中CI和CII亚家族的成员数目远多于另外3个物种, 这直接导致拟南芥和葡萄中含有不同数目的sHSP成员.类似现象也出现在棉花扩展蛋白基因家族(张奇艳等, 2019)、玉米(Zea mays) NAC家族(Fan et al., 2014)及大豆PP2C家族(Fan et al., 2020a).sHSP家族的保守基序分布和基因结构分布也与通过系统进化分析得出的亚家族分布一致(图2). ...
... 在睡莲、水稻、拟南芥和葡萄中共鉴定到12个与sHSP家族有关的基因复制事件(表2).相比其它2种植物, 在葡萄和拟南芥中发生的基因复制事件最多(各4个).葡萄中sHSP成员数量最多可能与葡萄发生的基因复制事件最多有关, 相似的研究结果也出现在葡萄的CDPL家族(Zhang et al., 2015)、卵形家族蛋白(Wang et al., 2018)以及WRKY家族中(Guo et al., 2014).在拟南芥中, 虽然与AtsHSP成员相关的基因复制事件最多, 但成员数量较少, 这可能与拟南芥基因复制事件后大量基因丢失有关(Cannon et al., 2004; Maere et al., 2005).结合染色体定位结果, 可以将12个与sHSP家族有关的基因复制事件进一步分为6个片段复制事件和6个串联复制事件, 并且每个物种中均含有这2种基因复制事件, 这表明sHSP家族的扩增不仅与大规模的片段复制事件有关, 还与小规模的串联复制事件有关(图3).相似的sHSP家族扩增模式也出现在辣椒(Guo et al., 2015)、簸箕柳(Li et al., 2018)和马铃薯(Zhao et al., 2018)等植物中.在每个物种中, 片段复制事件的Ks值均大于串联复制事件的Ks值, 这表明在sHSP家族中片段复制事件发生的时间均早于串联复制事件, 相似的现象也出现在梅花(Prunus mume) NAC家族(Zhuo et al., 2018)和棉花PP2C家族(Shazadee et al., 2019)中.基于复制事件的Ka/Ks比值表明, sHSP成员在发生复制事件以后会发生强烈的纯化选择作用, 类似的Ka/Ks比值分布也出现在其它植物的sHSP家族中(Ma et al., 2016; Li et al., 2018).与此同时, 与其它基因家族相比(Zhuo et al., 2018; Fan et al., 2020b), sHSP家族发生的基因复制事件较少(表2), 这也直接导致sHSP家族成员数相对较少, 表明在植物的进化过程中, sHSP成员可能具有相对保守的功能(如响应高温胁迫) (Waters and Vierling, 2020).此外, 本研究发现4个片段复制事件的Ks值大于1 (表2), 其中在拟南芥中有1个片段复制事件(AtsHSP01/AtsHSP06), 在另外3种植物中均发现对应的同源基因(图4), 该片段复制事件发生的时间大约为1-1.2亿年前温暖干旱的白垩纪(Zhang et al., 2020b), 这表明AtsHSP01/AtsHSP06及其在植物界的同源基因可能与植物适应高温环境有关, 可以作为今后研究高温胁迫的重要候选基因. ...
... 之前的研究表明, sHSP家族可以参与许多植物生长与发育过程(Zhang et al., 2018; Ma et al., 2019).本研究在睡莲、水稻、拟南芥和葡萄中新发现一些与植物生长发育过程相关的sHSP成员(图5).CI亚家族的NcsHSP19、OssHSP02、OssHSP14和VvsHSP40以及CIV亚家族的AtsHSP09和OssHSP23在几乎所有组织中均高度表达, 其在棉花和马铃薯中的同源基因也具有类似的表达模式(Ma et al., 2016; Zhao et al., 2018), 这表明sHSP家族在植物中的功能可能具有一定的保守性.此外, 本研究还发现一些与水稻花器官发育相关的基因(OssHSP03、OssHSP04、OssHSP06、OssHSP12、OssHSP16和OssHSP21).有报道显示, 在玫瑰(Rosa hybrida)中与CI亚家族OssHSP03、OssHSP04和OssHSP16相同亚家族的sHSP17.5-CI可能与开花过程中保护细胞蛋白有关(Dafny-Yelin et al., 2008). ...
Modeling gene and genome duplications in eukaryotes 1 2005
... 在睡莲、水稻、拟南芥和葡萄中共鉴定到12个与sHSP家族有关的基因复制事件(表2).相比其它2种植物, 在葡萄和拟南芥中发生的基因复制事件最多(各4个).葡萄中sHSP成员数量最多可能与葡萄发生的基因复制事件最多有关, 相似的研究结果也出现在葡萄的CDPL家族(Zhang et al., 2015)、卵形家族蛋白(Wang et al., 2018)以及WRKY家族中(Guo et al., 2014).在拟南芥中, 虽然与AtsHSP成员相关的基因复制事件最多, 但成员数量较少, 这可能与拟南芥基因复制事件后大量基因丢失有关(Cannon et al., 2004; Maere et al., 2005).结合染色体定位结果, 可以将12个与sHSP家族有关的基因复制事件进一步分为6个片段复制事件和6个串联复制事件, 并且每个物种中均含有这2种基因复制事件, 这表明sHSP家族的扩增不仅与大规模的片段复制事件有关, 还与小规模的串联复制事件有关(图3).相似的sHSP家族扩增模式也出现在辣椒(Guo et al., 2015)、簸箕柳(Li et al., 2018)和马铃薯(Zhao et al., 2018)等植物中.在每个物种中, 片段复制事件的Ks值均大于串联复制事件的Ks值, 这表明在sHSP家族中片段复制事件发生的时间均早于串联复制事件, 相似的现象也出现在梅花(Prunus mume) NAC家族(Zhuo et al., 2018)和棉花PP2C家族(Shazadee et al., 2019)中.基于复制事件的Ka/Ks比值表明, sHSP成员在发生复制事件以后会发生强烈的纯化选择作用, 类似的Ka/Ks比值分布也出现在其它植物的sHSP家族中(Ma et al., 2016; Li et al., 2018).与此同时, 与其它基因家族相比(Zhuo et al., 2018; Fan et al., 2020b), sHSP家族发生的基因复制事件较少(表2), 这也直接导致sHSP家族成员数相对较少, 表明在植物的进化过程中, sHSP成员可能具有相对保守的功能(如响应高温胁迫) (Waters and Vierling, 2020).此外, 本研究发现4个片段复制事件的Ks值大于1 (表2), 其中在拟南芥中有1个片段复制事件(AtsHSP01/AtsHSP06), 在另外3种植物中均发现对应的同源基因(图4), 该片段复制事件发生的时间大约为1-1.2亿年前温暖干旱的白垩纪(Zhang et al., 2020b), 这表明AtsHSP01/AtsHSP06及其在植物界的同源基因可能与植物适应高温环境有关, 可以作为今后研究高温胁迫的重要候选基因. ...
The TIGR Rice Genome Annotation Resource: improvements and new features 3 2007
... 被子植物在日常生活中随处可见, 可为人们提供粮食、纤维、燃料和药品等.被子植物的多样性与外界环境变化息息相关.在众多被子植物中, 基部被子植物睡莲(Nymphaea colorata) (Zhang et al., 2020a)、单子叶模式植物水稻(Oryza sativa) (Ouyang et al., 2007)、双子叶模式植物拟南芥(Arabidopsis thaliana) (Lamesch et al., 2012)以及类似古代双子叶植物基因组的葡萄(Vitis vinifera) (The French-Italian Public Consortium for Grapevine Genome Characterization, 2007)在被子植物分类学中具有重要意义.目前, 睡莲、水稻、拟南芥和葡萄的基因组测序已经完成, 从与外界环境变化相关的基因家族层面上对这4类被子植物进化历程进行研究, 不仅有助于解析被子植物进化的方向, 而且可为后续的分子育种提供重要参考. ...
... 目前, 虽然已在许多植物中鉴定到sHSP成员, 但主要集中在对单个物种进行全基因组分析, 缺乏对植物整体进化趋势的认识.睡莲、水稻、拟南芥和葡萄在被子植物进化过程中具有重要意义.目前, 睡莲(Zhang et al., 2020a)、水稻(Ouyang et al., 2007)、拟南芥(Lamesch et al., 2012)和葡萄(The French- Italian Public Consortium for Grapevine Genome Characterization, 2007)的全基因组测序、组装以及注释工作已经完成, 这为在4种被子植物中进行sHSP家族的比较基因组学分析奠定了基础.本研究利用最新的睡莲、水稻、拟南芥和葡萄的全基因组测序信息, 对其sHSP家族进行鉴定, 并在此基础上进一步对其系统发育、保守基序、基因结构、染色体定位、基因复制事件、共线性和基因表达等方面进行比较基因组学分析, 旨在为从基因家族层面进一步揭示植物的进化方向提供重要参考, 同时也为分子育种提供具有潜在育种价值的候选基因. ...
... 本研究在睡莲、水稻、拟南芥和葡萄中分别鉴定到33个NcsHSPs、24个OssHSPs、17个AtsHSPs和47个VvsHSPs (表1).4种植物中, 在葡萄中鉴定到的sHSP成员最多, 而在拟南芥中鉴定到的sHSP成员最少, 这主要与4种植物基因组大小有关(The French-Italian Public Consortium for Grapevine Genome Characterization, 2007; Ouyang et al., 2007; Lamesch et al., 2012; Zhang et al., 2020a).通过系统进化分析, 我们将sHSP家族进一步分为12个亚家族(图1).不同的亚家族含有不同数目的sHSP成员, 其中CI亚家族包含最多的sHSP成员, 这可能与CI亚家族包含最多的基因复制事件(4个)有关(表2).相似的分布也出现在番茄(张宁等, 2017)、马铃薯(Zhao et al., 2018)、棉花(Ma et al., 2016)、银杏(何福林和张斌, 2019)和辣椒(Guo et al., 2015)等植物中.因此, 在植物中sHSP家族具有保守的亚家族分布.在拟南芥中未发现任何属于CVI和CVII亚家族的成员, 而在葡萄中CI和CII亚家族的成员数目远多于另外3个物种, 这直接导致拟南芥和葡萄中含有不同数目的sHSP成员.类似现象也出现在棉花扩展蛋白基因家族(张奇艳等, 2019)、玉米(Zea mays) NAC家族(Fan et al., 2014)及大豆PP2C家族(Fan et al., 2020a).sHSP家族的保守基序分布和基因结构分布也与通过系统进化分析得出的亚家族分布一致(图2). ...
Intron loss mediated structural dynamics and functional differentiation of the polygalacturonase gene family in land plants 1 2010
... 睡莲是现存的早期被子植物之一(Zhang et al., 2020a), 通过与另外3种模式被子植物的sHSP家族进行对比, 发现睡莲sHSP家族中氨基酸长度较长以及分子量较大的成员较多(表1), 其次为具有很多古代植物基因组特征的葡萄sHSP成员(The French- Italian Public Consortium for Grapevine Genome Characterization, 2007), 表明sHSP家族在被子植物中可能向更短的氨基酸长度和更小的分子量进化, 这种进化趋势与已报道的rhomboid家族不同(Li et al., 2015), 但与PG家族类似(Park and Kim, 2010).其次, 水稻、拟南芥和葡萄大部分sHSP成员的内含子不多于1个(图2), 而睡莲含有较多基因结构较为复杂的sHSP成员, 这表明sHSP家族在被子植物中可能向更简单的基因结构进化, 这种趋势与已报道的NAP亚家族不同(Fan et al., 2015).此外, 睡莲sHSP成员分布在12条染色体上(图3), 葡萄sHSP成员分布在11条染色体上, 而拟南芥和水稻的sHSP成员分别分布在5条和8条染色体上, 表明sHSP成员在被子植物中可能更集中分布于某几条染色体上, 这种集中分布模式可能更有利于sHSP成员协同发挥作用. ...
Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown 1 2016
... 拟南芥AtsHSP和水稻OssHSP的表达谱数据主要从Genevestigator数据库下载获得.葡萄中VvsHSP成员的表达谱数据主要从Grape RNA数据库(http://www.grapeworld.cn/gt/index.html)下载获得.睡莲中NcsHSP成员的表达谱数据主要从NCBI-SRA数据库(https://www.ncbi.nlm.nih.gov/sra)下载获得(SRA Study: SRP222853).下载的睡莲RNA-Seq数据主要通过HISAT2、StringTie和Ballgown流程进行转录组分析(Pertea et al., 2016).拟南芥AtsHSP和水稻OssHSP的表达谱数据通过基因表达潜力的百分比进行计算; 葡萄VvsHSP和睡莲NcsHSP的表达谱数据通过将FPKM值进行log2(FPKM+1)转化获得.所有NcsHSP、OssHSP、AtsHSP和VvsHSP表达谱数据均使用Rstudio中的ComplexHeatmap包进行聚类分析并进行可视化展示. ...
Rice sHsp genes: genomic organization and expression profiling under stress and development 1 2009
... 在外界环境发生变化时, 植物在生理和分子水平上发生一系列改变以适应环境.热激蛋白(heat shock protein, HSP)是一类重要的响应外界环境变化的蛋白家族, 小热激蛋白(small heat shock protein, sHSP)是其中一类分子量较小的热激蛋白(刘德立, 1996; 黄祥富等, 1999; Wang et al., 2004; 栗振义等, 2016).目前已在古生菌、细菌和真核生物中发现sHSP成员(Waters and Vierling, 2020).sHSP成员具有保守的α-晶体结构域, 该结构域由80-100个氨基酸残基构成的结构紧凑的β链组成(van Montfort et al., 2001).在植物中, 首先在烟草(Nicotiana tabacum)和大豆(Glycine max)中鉴定出sHSP成员(Key et al., 1981), 随后相继在拟南芥(Scharf et al., 2001)、水稻(Sarkar et al., 2009)、葡萄(Ji et al., 2019)、番茄(Solanum lycopersicum) (张宁等, 2017)、马铃薯(S. tuberosum) (Zhao et al., 2018)、棉花(Gossypium hirsutum) (Ma et al., 2016)、银杏(Ginkgo biloba) (何福林和张斌, 2019)、辣椒(Capsicum annuum) (Guo et al., 2015)和簸箕柳 ...
The expanding family of Arabidopsis thaliana small heat stress proteins and a new family of proteins containing α-crystallin domains (Acd proteins) 1 2001
... 在外界环境发生变化时, 植物在生理和分子水平上发生一系列改变以适应环境.热激蛋白(heat shock protein, HSP)是一类重要的响应外界环境变化的蛋白家族, 小热激蛋白(small heat shock protein, sHSP)是其中一类分子量较小的热激蛋白(刘德立, 1996; 黄祥富等, 1999; Wang et al., 2004; 栗振义等, 2016).目前已在古生菌、细菌和真核生物中发现sHSP成员(Waters and Vierling, 2020).sHSP成员具有保守的α-晶体结构域, 该结构域由80-100个氨基酸残基构成的结构紧凑的β链组成(van Montfort et al., 2001).在植物中, 首先在烟草(Nicotiana tabacum)和大豆(Glycine max)中鉴定出sHSP成员(Key et al., 1981), 随后相继在拟南芥(Scharf et al., 2001)、水稻(Sarkar et al., 2009)、葡萄(Ji et al., 2019)、番茄(Solanum lycopersicum) (张宁等, 2017)、马铃薯(S. tuberosum) (Zhao et al., 2018)、棉花(Gossypium hirsutum) (Ma et al., 2016)、银杏(Ginkgo biloba) (何福林和张斌, 2019)、辣椒(Capsicum annuum) (Guo et al., 2015)和簸箕柳 ...
Identification and expression profiling of protein phosphatases (PP2C) gene family in Gossypium hirsutum L 1 2019
... 在睡莲、水稻、拟南芥和葡萄中共鉴定到12个与sHSP家族有关的基因复制事件(表2).相比其它2种植物, 在葡萄和拟南芥中发生的基因复制事件最多(各4个).葡萄中sHSP成员数量最多可能与葡萄发生的基因复制事件最多有关, 相似的研究结果也出现在葡萄的CDPL家族(Zhang et al., 2015)、卵形家族蛋白(Wang et al., 2018)以及WRKY家族中(Guo et al., 2014).在拟南芥中, 虽然与AtsHSP成员相关的基因复制事件最多, 但成员数量较少, 这可能与拟南芥基因复制事件后大量基因丢失有关(Cannon et al., 2004; Maere et al., 2005).结合染色体定位结果, 可以将12个与sHSP家族有关的基因复制事件进一步分为6个片段复制事件和6个串联复制事件, 并且每个物种中均含有这2种基因复制事件, 这表明sHSP家族的扩增不仅与大规模的片段复制事件有关, 还与小规模的串联复制事件有关(图3).相似的sHSP家族扩增模式也出现在辣椒(Guo et al., 2015)、簸箕柳(Li et al., 2018)和马铃薯(Zhao et al., 2018)等植物中.在每个物种中, 片段复制事件的Ks值均大于串联复制事件的Ks值, 这表明在sHSP家族中片段复制事件发生的时间均早于串联复制事件, 相似的现象也出现在梅花(Prunus mume) NAC家族(Zhuo et al., 2018)和棉花PP2C家族(Shazadee et al., 2019)中.基于复制事件的Ka/Ks比值表明, sHSP成员在发生复制事件以后会发生强烈的纯化选择作用, 类似的Ka/Ks比值分布也出现在其它植物的sHSP家族中(Ma et al., 2016; Li et al., 2018).与此同时, 与其它基因家族相比(Zhuo et al., 2018; Fan et al., 2020b), sHSP家族发生的基因复制事件较少(表2), 这也直接导致sHSP家族成员数相对较少, 表明在植物的进化过程中, sHSP成员可能具有相对保守的功能(如响应高温胁迫) (Waters and Vierling, 2020).此外, 本研究发现4个片段复制事件的Ks值大于1 (表2), 其中在拟南芥中有1个片段复制事件(AtsHSP01/AtsHSP06), 在另外3种植物中均发现对应的同源基因(图4), 该片段复制事件发生的时间大约为1-1.2亿年前温暖干旱的白垩纪(Zhang et al., 2020b), 这表明AtsHSP01/AtsHSP06及其在植物界的同源基因可能与植物适应高温环境有关, 可以作为今后研究高温胁迫的重要候选基因. ...
At-HSP17.6A, encoding a small heat-shock protein in Arabidopsis, can enhance osmotolerance upon overexpression 1 2010
... (Salix suchowensis) (Li et al., 2018)等植物中发现了大量的sHSP成员.sHSP家族主要作为分子伴侣, 以独立于ATP的形式保护植物免受外界环境的影响(Waters and Vierling, 2020).大量研究表明, sHSP成员可以响应外界多种环境胁迫(Sun et al., 2010; Zhu et al., 2011; Li et al., 2016; Zhai et al., 2016; Guo et al., 2020).此外, sHSP家族也参与果实品质形成和植株发芽等植物生长发育过程(Zhang et al., 2018; Ma et al., 2019). ...
The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla 5 2007
... 被子植物在日常生活中随处可见, 可为人们提供粮食、纤维、燃料和药品等.被子植物的多样性与外界环境变化息息相关.在众多被子植物中, 基部被子植物睡莲(Nymphaea colorata) (Zhang et al., 2020a)、单子叶模式植物水稻(Oryza sativa) (Ouyang et al., 2007)、双子叶模式植物拟南芥(Arabidopsis thaliana) (Lamesch et al., 2012)以及类似古代双子叶植物基因组的葡萄(Vitis vinifera) (The French-Italian Public Consortium for Grapevine Genome Characterization, 2007)在被子植物分类学中具有重要意义.目前, 睡莲、水稻、拟南芥和葡萄的基因组测序已经完成, 从与外界环境变化相关的基因家族层面上对这4类被子植物进化历程进行研究, 不仅有助于解析被子植物进化的方向, 而且可为后续的分子育种提供重要参考. ...
... 目前, 虽然已在许多植物中鉴定到sHSP成员, 但主要集中在对单个物种进行全基因组分析, 缺乏对植物整体进化趋势的认识.睡莲、水稻、拟南芥和葡萄在被子植物进化过程中具有重要意义.目前, 睡莲(Zhang et al., 2020a)、水稻(Ouyang et al., 2007)、拟南芥(Lamesch et al., 2012)和葡萄(The French- Italian Public Consortium for Grapevine Genome Characterization, 2007)的全基因组测序、组装以及注释工作已经完成, 这为在4种被子植物中进行sHSP家族的比较基因组学分析奠定了基础.本研究利用最新的睡莲、水稻、拟南芥和葡萄的全基因组测序信息, 对其sHSP家族进行鉴定, 并在此基础上进一步对其系统发育、保守基序、基因结构、染色体定位、基因复制事件、共线性和基因表达等方面进行比较基因组学分析, 旨在为从基因家族层面进一步揭示植物的进化方向提供重要参考, 同时也为分子育种提供具有潜在育种价值的候选基因. ...
... 本研究在睡莲、水稻、拟南芥和葡萄中分别鉴定到33个NcsHSPs、24个OssHSPs、17个AtsHSPs和47个VvsHSPs (表1).4种植物中, 在葡萄中鉴定到的sHSP成员最多, 而在拟南芥中鉴定到的sHSP成员最少, 这主要与4种植物基因组大小有关(The French-Italian Public Consortium for Grapevine Genome Characterization, 2007; Ouyang et al., 2007; Lamesch et al., 2012; Zhang et al., 2020a).通过系统进化分析, 我们将sHSP家族进一步分为12个亚家族(图1).不同的亚家族含有不同数目的sHSP成员, 其中CI亚家族包含最多的sHSP成员, 这可能与CI亚家族包含最多的基因复制事件(4个)有关(表2).相似的分布也出现在番茄(张宁等, 2017)、马铃薯(Zhao et al., 2018)、棉花(Ma et al., 2016)、银杏(何福林和张斌, 2019)和辣椒(Guo et al., 2015)等植物中.因此, 在植物中sHSP家族具有保守的亚家族分布.在拟南芥中未发现任何属于CVI和CVII亚家族的成员, 而在葡萄中CI和CII亚家族的成员数目远多于另外3个物种, 这直接导致拟南芥和葡萄中含有不同数目的sHSP成员.类似现象也出现在棉花扩展蛋白基因家族(张奇艳等, 2019)、玉米(Zea mays) NAC家族(Fan et al., 2014)及大豆PP2C家族(Fan et al., 2020a).sHSP家族的保守基序分布和基因结构分布也与通过系统进化分析得出的亚家族分布一致(图2). ...
... 对睡莲、水稻、拟南芥和葡萄中sHSP家族之间的共线性分析(图4)表明, 拟南芥和葡萄中的同源基因对最多(10对), 这主要与拟南芥和葡萄之间具有较近的进化关系有关(Jiao et al., 2012).虽然睡莲(基部被子植物)和葡萄(双子叶植物)具有较远的进化距离, 但其sHSP成员却有较多的同源基因对(8对), 这主要与葡萄基因组并未发生最近的基因复制事件从而导致基因组有很多古代植物基因组的特征有关(The French-Italian Public Consortium for Grapevine Genome Characterization, 2007; Zhang et al., 2020a).此外, 本研究通过Ka值计算发现同源基因之间的进化同步, 即在植物中sHSP家族的进化相对保守, 这可能与sHSP家族的功能有一定关系(Waters and Vierling, 2020). ...
... 睡莲是现存的早期被子植物之一(Zhang et al., 2020a), 通过与另外3种模式被子植物的sHSP家族进行对比, 发现睡莲sHSP家族中氨基酸长度较长以及分子量较大的成员较多(表1), 其次为具有很多古代植物基因组特征的葡萄sHSP成员(The French- Italian Public Consortium for Grapevine Genome Characterization, 2007), 表明sHSP家族在被子植物中可能向更短的氨基酸长度和更小的分子量进化, 这种进化趋势与已报道的rhomboid家族不同(Li et al., 2015), 但与PG家族类似(Park and Kim, 2010).其次, 水稻、拟南芥和葡萄大部分sHSP成员的内含子不多于1个(图2), 而睡莲含有较多基因结构较为复杂的sHSP成员, 这表明sHSP家族在被子植物中可能向更简单的基因结构进化, 这种趋势与已报道的NAP亚家族不同(Fan et al., 2015).此外, 睡莲sHSP成员分布在12条染色体上(图3), 葡萄sHSP成员分布在11条染色体上, 而拟南芥和水稻的sHSP成员分别分布在5条和8条染色体上, 表明sHSP成员在被子植物中可能更集中分布于某几条染色体上, 这种集中分布模式可能更有利于sHSP成员协同发挥作用. ...
Crystal structure and assembly of a eukaryotic small heat shock protein 1 2001
... 在外界环境发生变化时, 植物在生理和分子水平上发生一系列改变以适应环境.热激蛋白(heat shock protein, HSP)是一类重要的响应外界环境变化的蛋白家族, 小热激蛋白(small heat shock protein, sHSP)是其中一类分子量较小的热激蛋白(刘德立, 1996; 黄祥富等, 1999; Wang et al., 2004; 栗振义等, 2016).目前已在古生菌、细菌和真核生物中发现sHSP成员(Waters and Vierling, 2020).sHSP成员具有保守的α-晶体结构域, 该结构域由80-100个氨基酸残基构成的结构紧凑的β链组成(van Montfort et al., 2001).在植物中, 首先在烟草(Nicotiana tabacum)和大豆(Glycine max)中鉴定出sHSP成员(Key et al., 1981), 随后相继在拟南芥(Scharf et al., 2001)、水稻(Sarkar et al., 2009)、葡萄(Ji et al., 2019)、番茄(Solanum lycopersicum) (张宁等, 2017)、马铃薯(S. tuberosum) (Zhao et al., 2018)、棉花(Gossypium hirsutum) (Ma et al., 2016)、银杏(Ginkgo biloba) (何福林和张斌, 2019)、辣椒(Capsicum annuum) (Guo et al., 2015)和簸箕柳 ...
Evolutionary and expression analysis of Vitis vinifera OFP gene family 1 2018
... 在睡莲、水稻、拟南芥和葡萄中共鉴定到12个与sHSP家族有关的基因复制事件(表2).相比其它2种植物, 在葡萄和拟南芥中发生的基因复制事件最多(各4个).葡萄中sHSP成员数量最多可能与葡萄发生的基因复制事件最多有关, 相似的研究结果也出现在葡萄的CDPL家族(Zhang et al., 2015)、卵形家族蛋白(Wang et al., 2018)以及WRKY家族中(Guo et al., 2014).在拟南芥中, 虽然与AtsHSP成员相关的基因复制事件最多, 但成员数量较少, 这可能与拟南芥基因复制事件后大量基因丢失有关(Cannon et al., 2004; Maere et al., 2005).结合染色体定位结果, 可以将12个与sHSP家族有关的基因复制事件进一步分为6个片段复制事件和6个串联复制事件, 并且每个物种中均含有这2种基因复制事件, 这表明sHSP家族的扩增不仅与大规模的片段复制事件有关, 还与小规模的串联复制事件有关(图3).相似的sHSP家族扩增模式也出现在辣椒(Guo et al., 2015)、簸箕柳(Li et al., 2018)和马铃薯(Zhao et al., 2018)等植物中.在每个物种中, 片段复制事件的Ks值均大于串联复制事件的Ks值, 这表明在sHSP家族中片段复制事件发生的时间均早于串联复制事件, 相似的现象也出现在梅花(Prunus mume) NAC家族(Zhuo et al., 2018)和棉花PP2C家族(Shazadee et al., 2019)中.基于复制事件的Ka/Ks比值表明, sHSP成员在发生复制事件以后会发生强烈的纯化选择作用, 类似的Ka/Ks比值分布也出现在其它植物的sHSP家族中(Ma et al., 2016; Li et al., 2018).与此同时, 与其它基因家族相比(Zhuo et al., 2018; Fan et al., 2020b), sHSP家族发生的基因复制事件较少(表2), 这也直接导致sHSP家族成员数相对较少, 表明在植物的进化过程中, sHSP成员可能具有相对保守的功能(如响应高温胁迫) (Waters and Vierling, 2020).此外, 本研究发现4个片段复制事件的Ks值大于1 (表2), 其中在拟南芥中有1个片段复制事件(AtsHSP01/AtsHSP06), 在另外3种植物中均发现对应的同源基因(图4), 该片段复制事件发生的时间大约为1-1.2亿年前温暖干旱的白垩纪(Zhang et al., 2020b), 这表明AtsHSP01/AtsHSP06及其在植物界的同源基因可能与植物适应高温环境有关, 可以作为今后研究高温胁迫的重要候选基因. ...
Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response 1 2004
... 在外界环境发生变化时, 植物在生理和分子水平上发生一系列改变以适应环境.热激蛋白(heat shock protein, HSP)是一类重要的响应外界环境变化的蛋白家族, 小热激蛋白(small heat shock protein, sHSP)是其中一类分子量较小的热激蛋白(刘德立, 1996; 黄祥富等, 1999; Wang et al., 2004; 栗振义等, 2016).目前已在古生菌、细菌和真核生物中发现sHSP成员(Waters and Vierling, 2020).sHSP成员具有保守的α-晶体结构域, 该结构域由80-100个氨基酸残基构成的结构紧凑的β链组成(van Montfort et al., 2001).在植物中, 首先在烟草(Nicotiana tabacum)和大豆(Glycine max)中鉴定出sHSP成员(Key et al., 1981), 随后相继在拟南芥(Scharf et al., 2001)、水稻(Sarkar et al., 2009)、葡萄(Ji et al., 2019)、番茄(Solanum lycopersicum) (张宁等, 2017)、马铃薯(S. tuberosum) (Zhao et al., 2018)、棉花(Gossypium hirsutum) (Ma et al., 2016)、银杏(Ginkgo biloba) (何福林和张斌, 2019)、辣椒(Capsicum annuum) (Guo et al., 2015)和簸箕柳 ...
Plant small heat shock proteins—evolutionary and functional diversity 5 2020
... 在外界环境发生变化时, 植物在生理和分子水平上发生一系列改变以适应环境.热激蛋白(heat shock protein, HSP)是一类重要的响应外界环境变化的蛋白家族, 小热激蛋白(small heat shock protein, sHSP)是其中一类分子量较小的热激蛋白(刘德立, 1996; 黄祥富等, 1999; Wang et al., 2004; 栗振义等, 2016).目前已在古生菌、细菌和真核生物中发现sHSP成员(Waters and Vierling, 2020).sHSP成员具有保守的α-晶体结构域, 该结构域由80-100个氨基酸残基构成的结构紧凑的β链组成(van Montfort et al., 2001).在植物中, 首先在烟草(Nicotiana tabacum)和大豆(Glycine max)中鉴定出sHSP成员(Key et al., 1981), 随后相继在拟南芥(Scharf et al., 2001)、水稻(Sarkar et al., 2009)、葡萄(Ji et al., 2019)、番茄(Solanum lycopersicum) (张宁等, 2017)、马铃薯(S. tuberosum) (Zhao et al., 2018)、棉花(Gossypium hirsutum) (Ma et al., 2016)、银杏(Ginkgo biloba) (何福林和张斌, 2019)、辣椒(Capsicum annuum) (Guo et al., 2015)和簸箕柳 ...
... (Salix suchowensis) (Li et al., 2018)等植物中发现了大量的sHSP成员.sHSP家族主要作为分子伴侣, 以独立于ATP的形式保护植物免受外界环境的影响(Waters and Vierling, 2020).大量研究表明, sHSP成员可以响应外界多种环境胁迫(Sun et al., 2010; Zhu et al., 2011; Li et al., 2016; Zhai et al., 2016; Guo et al., 2020).此外, sHSP家族也参与果实品质形成和植株发芽等植物生长发育过程(Zhang et al., 2018; Ma et al., 2019). ...
... 在睡莲、水稻、拟南芥和葡萄中共鉴定到12个与sHSP家族有关的基因复制事件(表2).相比其它2种植物, 在葡萄和拟南芥中发生的基因复制事件最多(各4个).葡萄中sHSP成员数量最多可能与葡萄发生的基因复制事件最多有关, 相似的研究结果也出现在葡萄的CDPL家族(Zhang et al., 2015)、卵形家族蛋白(Wang et al., 2018)以及WRKY家族中(Guo et al., 2014).在拟南芥中, 虽然与AtsHSP成员相关的基因复制事件最多, 但成员数量较少, 这可能与拟南芥基因复制事件后大量基因丢失有关(Cannon et al., 2004; Maere et al., 2005).结合染色体定位结果, 可以将12个与sHSP家族有关的基因复制事件进一步分为6个片段复制事件和6个串联复制事件, 并且每个物种中均含有这2种基因复制事件, 这表明sHSP家族的扩增不仅与大规模的片段复制事件有关, 还与小规模的串联复制事件有关(图3).相似的sHSP家族扩增模式也出现在辣椒(Guo et al., 2015)、簸箕柳(Li et al., 2018)和马铃薯(Zhao et al., 2018)等植物中.在每个物种中, 片段复制事件的Ks值均大于串联复制事件的Ks值, 这表明在sHSP家族中片段复制事件发生的时间均早于串联复制事件, 相似的现象也出现在梅花(Prunus mume) NAC家族(Zhuo et al., 2018)和棉花PP2C家族(Shazadee et al., 2019)中.基于复制事件的Ka/Ks比值表明, sHSP成员在发生复制事件以后会发生强烈的纯化选择作用, 类似的Ka/Ks比值分布也出现在其它植物的sHSP家族中(Ma et al., 2016; Li et al., 2018).与此同时, 与其它基因家族相比(Zhuo et al., 2018; Fan et al., 2020b), sHSP家族发生的基因复制事件较少(表2), 这也直接导致sHSP家族成员数相对较少, 表明在植物的进化过程中, sHSP成员可能具有相对保守的功能(如响应高温胁迫) (Waters and Vierling, 2020).此外, 本研究发现4个片段复制事件的Ks值大于1 (表2), 其中在拟南芥中有1个片段复制事件(AtsHSP01/AtsHSP06), 在另外3种植物中均发现对应的同源基因(图4), 该片段复制事件发生的时间大约为1-1.2亿年前温暖干旱的白垩纪(Zhang et al., 2020b), 这表明AtsHSP01/AtsHSP06及其在植物界的同源基因可能与植物适应高温环境有关, 可以作为今后研究高温胁迫的重要候选基因. ...
... 对睡莲、水稻、拟南芥和葡萄中sHSP家族之间的共线性分析(图4)表明, 拟南芥和葡萄中的同源基因对最多(10对), 这主要与拟南芥和葡萄之间具有较近的进化关系有关(Jiao et al., 2012).虽然睡莲(基部被子植物)和葡萄(双子叶植物)具有较远的进化距离, 但其sHSP成员却有较多的同源基因对(8对), 这主要与葡萄基因组并未发生最近的基因复制事件从而导致基因组有很多古代植物基因组的特征有关(The French-Italian Public Consortium for Grapevine Genome Characterization, 2007; Zhang et al., 2020a).此外, 本研究通过Ka值计算发现同源基因之间的进化同步, 即在植物中sHSP家族的进化相对保守, 这可能与sHSP家族的功能有一定关系(Waters and Vierling, 2020). ...
Over-expression of JrsHSP17.3 gene from Juglans regia confer the tolerance to abnormal temperature and NaCl stresses 1 2016
... (Salix suchowensis) (Li et al., 2018)等植物中发现了大量的sHSP成员.sHSP家族主要作为分子伴侣, 以独立于ATP的形式保护植物免受外界环境的影响(Waters and Vierling, 2020).大量研究表明, sHSP成员可以响应外界多种环境胁迫(Sun et al., 2010; Zhu et al., 2011; Li et al., 2016; Zhai et al., 2016; Guo et al., 2020).此外, sHSP家族也参与果实品质形成和植株发芽等植物生长发育过程(Zhang et al., 2018; Ma et al., 2019). ...
Genome-wide identification and expression analysis of the CDPK gene family in grape, Vitis spp 1 2015
... 在睡莲、水稻、拟南芥和葡萄中共鉴定到12个与sHSP家族有关的基因复制事件(表2).相比其它2种植物, 在葡萄和拟南芥中发生的基因复制事件最多(各4个).葡萄中sHSP成员数量最多可能与葡萄发生的基因复制事件最多有关, 相似的研究结果也出现在葡萄的CDPL家族(Zhang et al., 2015)、卵形家族蛋白(Wang et al., 2018)以及WRKY家族中(Guo et al., 2014).在拟南芥中, 虽然与AtsHSP成员相关的基因复制事件最多, 但成员数量较少, 这可能与拟南芥基因复制事件后大量基因丢失有关(Cannon et al., 2004; Maere et al., 2005).结合染色体定位结果, 可以将12个与sHSP家族有关的基因复制事件进一步分为6个片段复制事件和6个串联复制事件, 并且每个物种中均含有这2种基因复制事件, 这表明sHSP家族的扩增不仅与大规模的片段复制事件有关, 还与小规模的串联复制事件有关(图3).相似的sHSP家族扩增模式也出现在辣椒(Guo et al., 2015)、簸箕柳(Li et al., 2018)和马铃薯(Zhao et al., 2018)等植物中.在每个物种中, 片段复制事件的Ks值均大于串联复制事件的Ks值, 这表明在sHSP家族中片段复制事件发生的时间均早于串联复制事件, 相似的现象也出现在梅花(Prunus mume) NAC家族(Zhuo et al., 2018)和棉花PP2C家族(Shazadee et al., 2019)中.基于复制事件的Ka/Ks比值表明, sHSP成员在发生复制事件以后会发生强烈的纯化选择作用, 类似的Ka/Ks比值分布也出现在其它植物的sHSP家族中(Ma et al., 2016; Li et al., 2018).与此同时, 与其它基因家族相比(Zhuo et al., 2018; Fan et al., 2020b), sHSP家族发生的基因复制事件较少(表2), 这也直接导致sHSP家族成员数相对较少, 表明在植物的进化过程中, sHSP成员可能具有相对保守的功能(如响应高温胁迫) (Waters and Vierling, 2020).此外, 本研究发现4个片段复制事件的Ks值大于1 (表2), 其中在拟南芥中有1个片段复制事件(AtsHSP01/AtsHSP06), 在另外3种植物中均发现对应的同源基因(图4), 该片段复制事件发生的时间大约为1-1.2亿年前温暖干旱的白垩纪(Zhang et al., 2020b), 这表明AtsHSP01/AtsHSP06及其在植物界的同源基因可能与植物适应高温环境有关, 可以作为今后研究高温胁迫的重要候选基因. ...
The water lily genome and the early evolution of flowering plants 5 2020a
... 被子植物在日常生活中随处可见, 可为人们提供粮食、纤维、燃料和药品等.被子植物的多样性与外界环境变化息息相关.在众多被子植物中, 基部被子植物睡莲(Nymphaea colorata) (Zhang et al., 2020a)、单子叶模式植物水稻(Oryza sativa) (Ouyang et al., 2007)、双子叶模式植物拟南芥(Arabidopsis thaliana) (Lamesch et al., 2012)以及类似古代双子叶植物基因组的葡萄(Vitis vinifera) (The French-Italian Public Consortium for Grapevine Genome Characterization, 2007)在被子植物分类学中具有重要意义.目前, 睡莲、水稻、拟南芥和葡萄的基因组测序已经完成, 从与外界环境变化相关的基因家族层面上对这4类被子植物进化历程进行研究, 不仅有助于解析被子植物进化的方向, 而且可为后续的分子育种提供重要参考. ...
... 目前, 虽然已在许多植物中鉴定到sHSP成员, 但主要集中在对单个物种进行全基因组分析, 缺乏对植物整体进化趋势的认识.睡莲、水稻、拟南芥和葡萄在被子植物进化过程中具有重要意义.目前, 睡莲(Zhang et al., 2020a)、水稻(Ouyang et al., 2007)、拟南芥(Lamesch et al., 2012)和葡萄(The French- Italian Public Consortium for Grapevine Genome Characterization, 2007)的全基因组测序、组装以及注释工作已经完成, 这为在4种被子植物中进行sHSP家族的比较基因组学分析奠定了基础.本研究利用最新的睡莲、水稻、拟南芥和葡萄的全基因组测序信息, 对其sHSP家族进行鉴定, 并在此基础上进一步对其系统发育、保守基序、基因结构、染色体定位、基因复制事件、共线性和基因表达等方面进行比较基因组学分析, 旨在为从基因家族层面进一步揭示植物的进化方向提供重要参考, 同时也为分子育种提供具有潜在育种价值的候选基因. ...
... 本研究在睡莲、水稻、拟南芥和葡萄中分别鉴定到33个NcsHSPs、24个OssHSPs、17个AtsHSPs和47个VvsHSPs (表1).4种植物中, 在葡萄中鉴定到的sHSP成员最多, 而在拟南芥中鉴定到的sHSP成员最少, 这主要与4种植物基因组大小有关(The French-Italian Public Consortium for Grapevine Genome Characterization, 2007; Ouyang et al., 2007; Lamesch et al., 2012; Zhang et al., 2020a).通过系统进化分析, 我们将sHSP家族进一步分为12个亚家族(图1).不同的亚家族含有不同数目的sHSP成员, 其中CI亚家族包含最多的sHSP成员, 这可能与CI亚家族包含最多的基因复制事件(4个)有关(表2).相似的分布也出现在番茄(张宁等, 2017)、马铃薯(Zhao et al., 2018)、棉花(Ma et al., 2016)、银杏(何福林和张斌, 2019)和辣椒(Guo et al., 2015)等植物中.因此, 在植物中sHSP家族具有保守的亚家族分布.在拟南芥中未发现任何属于CVI和CVII亚家族的成员, 而在葡萄中CI和CII亚家族的成员数目远多于另外3个物种, 这直接导致拟南芥和葡萄中含有不同数目的sHSP成员.类似现象也出现在棉花扩展蛋白基因家族(张奇艳等, 2019)、玉米(Zea mays) NAC家族(Fan et al., 2014)及大豆PP2C家族(Fan et al., 2020a).sHSP家族的保守基序分布和基因结构分布也与通过系统进化分析得出的亚家族分布一致(图2). ...
... 对睡莲、水稻、拟南芥和葡萄中sHSP家族之间的共线性分析(图4)表明, 拟南芥和葡萄中的同源基因对最多(10对), 这主要与拟南芥和葡萄之间具有较近的进化关系有关(Jiao et al., 2012).虽然睡莲(基部被子植物)和葡萄(双子叶植物)具有较远的进化距离, 但其sHSP成员却有较多的同源基因对(8对), 这主要与葡萄基因组并未发生最近的基因复制事件从而导致基因组有很多古代植物基因组的特征有关(The French-Italian Public Consortium for Grapevine Genome Characterization, 2007; Zhang et al., 2020a).此外, 本研究通过Ka值计算发现同源基因之间的进化同步, 即在植物中sHSP家族的进化相对保守, 这可能与sHSP家族的功能有一定关系(Waters and Vierling, 2020). ...
... 睡莲是现存的早期被子植物之一(Zhang et al., 2020a), 通过与另外3种模式被子植物的sHSP家族进行对比, 发现睡莲sHSP家族中氨基酸长度较长以及分子量较大的成员较多(表1), 其次为具有很多古代植物基因组特征的葡萄sHSP成员(The French- Italian Public Consortium for Grapevine Genome Characterization, 2007), 表明sHSP家族在被子植物中可能向更短的氨基酸长度和更小的分子量进化, 这种进化趋势与已报道的rhomboid家族不同(Li et al., 2015), 但与PG家族类似(Park and Kim, 2010).其次, 水稻、拟南芥和葡萄大部分sHSP成员的内含子不多于1个(图2), 而睡莲含有较多基因结构较为复杂的sHSP成员, 这表明sHSP家族在被子植物中可能向更简单的基因结构进化, 这种趋势与已报道的NAP亚家族不同(Fan et al., 2015).此外, 睡莲sHSP成员分布在12条染色体上(图3), 葡萄sHSP成员分布在11条染色体上, 而拟南芥和水稻的sHSP成员分别分布在5条和8条染色体上, 表明sHSP成员在被子植物中可能更集中分布于某几条染色体上, 这种集中分布模式可能更有利于sHSP成员协同发挥作用. ...
The ancient wave of polyploidization events in flowering plants and their facilitated adaptation to environmental stress 1 2020b
... 在睡莲、水稻、拟南芥和葡萄中共鉴定到12个与sHSP家族有关的基因复制事件(表2).相比其它2种植物, 在葡萄和拟南芥中发生的基因复制事件最多(各4个).葡萄中sHSP成员数量最多可能与葡萄发生的基因复制事件最多有关, 相似的研究结果也出现在葡萄的CDPL家族(Zhang et al., 2015)、卵形家族蛋白(Wang et al., 2018)以及WRKY家族中(Guo et al., 2014).在拟南芥中, 虽然与AtsHSP成员相关的基因复制事件最多, 但成员数量较少, 这可能与拟南芥基因复制事件后大量基因丢失有关(Cannon et al., 2004; Maere et al., 2005).结合染色体定位结果, 可以将12个与sHSP家族有关的基因复制事件进一步分为6个片段复制事件和6个串联复制事件, 并且每个物种中均含有这2种基因复制事件, 这表明sHSP家族的扩增不仅与大规模的片段复制事件有关, 还与小规模的串联复制事件有关(图3).相似的sHSP家族扩增模式也出现在辣椒(Guo et al., 2015)、簸箕柳(Li et al., 2018)和马铃薯(Zhao et al., 2018)等植物中.在每个物种中, 片段复制事件的Ks值均大于串联复制事件的Ks值, 这表明在sHSP家族中片段复制事件发生的时间均早于串联复制事件, 相似的现象也出现在梅花(Prunus mume) NAC家族(Zhuo et al., 2018)和棉花PP2C家族(Shazadee et al., 2019)中.基于复制事件的Ka/Ks比值表明, sHSP成员在发生复制事件以后会发生强烈的纯化选择作用, 类似的Ka/Ks比值分布也出现在其它植物的sHSP家族中(Ma et al., 2016; Li et al., 2018).与此同时, 与其它基因家族相比(Zhuo et al., 2018; Fan et al., 2020b), sHSP家族发生的基因复制事件较少(表2), 这也直接导致sHSP家族成员数相对较少, 表明在植物的进化过程中, sHSP成员可能具有相对保守的功能(如响应高温胁迫) (Waters and Vierling, 2020).此外, 本研究发现4个片段复制事件的Ks值大于1 (表2), 其中在拟南芥中有1个片段复制事件(AtsHSP01/AtsHSP06), 在另外3种植物中均发现对应的同源基因(图4), 该片段复制事件发生的时间大约为1-1.2亿年前温暖干旱的白垩纪(Zhang et al., 2020b), 这表明AtsHSP01/AtsHSP06及其在植物界的同源基因可能与植物适应高温环境有关, 可以作为今后研究高温胁迫的重要候选基因. ...
Activation of small heat shock protein (SlHSP17.7) gene by cell wall invertase inhibitor (SlCIF1) gene involved in sugar metabolism in tomato 2 2018
... (Salix suchowensis) (Li et al., 2018)等植物中发现了大量的sHSP成员.sHSP家族主要作为分子伴侣, 以独立于ATP的形式保护植物免受外界环境的影响(Waters and Vierling, 2020).大量研究表明, sHSP成员可以响应外界多种环境胁迫(Sun et al., 2010; Zhu et al., 2011; Li et al., 2016; Zhai et al., 2016; Guo et al., 2020).此外, sHSP家族也参与果实品质形成和植株发芽等植物生长发育过程(Zhang et al., 2018; Ma et al., 2019). ...
... 之前的研究表明, sHSP家族可以参与许多植物生长与发育过程(Zhang et al., 2018; Ma et al., 2019).本研究在睡莲、水稻、拟南芥和葡萄中新发现一些与植物生长发育过程相关的sHSP成员(图5).CI亚家族的NcsHSP19、OssHSP02、OssHSP14和VvsHSP40以及CIV亚家族的AtsHSP09和OssHSP23在几乎所有组织中均高度表达, 其在棉花和马铃薯中的同源基因也具有类似的表达模式(Ma et al., 2016; Zhao et al., 2018), 这表明sHSP家族在植物中的功能可能具有一定的保守性.此外, 本研究还发现一些与水稻花器官发育相关的基因(OssHSP03、OssHSP04、OssHSP06、OssHSP12、OssHSP16和OssHSP21).有报道显示, 在玫瑰(Rosa hybrida)中与CI亚家族OssHSP03、OssHSP04和OssHSP16相同亚家族的sHSP17.5-CI可能与开花过程中保护细胞蛋白有关(Dafny-Yelin et al., 2008). ...
Genome-wide analysis of the potato Hsp20 gene family: identification, genomic organization and expression profiles in response to heat stress 4 2018
... 在外界环境发生变化时, 植物在生理和分子水平上发生一系列改变以适应环境.热激蛋白(heat shock protein, HSP)是一类重要的响应外界环境变化的蛋白家族, 小热激蛋白(small heat shock protein, sHSP)是其中一类分子量较小的热激蛋白(刘德立, 1996; 黄祥富等, 1999; Wang et al., 2004; 栗振义等, 2016).目前已在古生菌、细菌和真核生物中发现sHSP成员(Waters and Vierling, 2020).sHSP成员具有保守的α-晶体结构域, 该结构域由80-100个氨基酸残基构成的结构紧凑的β链组成(van Montfort et al., 2001).在植物中, 首先在烟草(Nicotiana tabacum)和大豆(Glycine max)中鉴定出sHSP成员(Key et al., 1981), 随后相继在拟南芥(Scharf et al., 2001)、水稻(Sarkar et al., 2009)、葡萄(Ji et al., 2019)、番茄(Solanum lycopersicum) (张宁等, 2017)、马铃薯(S. tuberosum) (Zhao et al., 2018)、棉花(Gossypium hirsutum) (Ma et al., 2016)、银杏(Ginkgo biloba) (何福林和张斌, 2019)、辣椒(Capsicum annuum) (Guo et al., 2015)和簸箕柳 ...
... 本研究在睡莲、水稻、拟南芥和葡萄中分别鉴定到33个NcsHSPs、24个OssHSPs、17个AtsHSPs和47个VvsHSPs (表1).4种植物中, 在葡萄中鉴定到的sHSP成员最多, 而在拟南芥中鉴定到的sHSP成员最少, 这主要与4种植物基因组大小有关(The French-Italian Public Consortium for Grapevine Genome Characterization, 2007; Ouyang et al., 2007; Lamesch et al., 2012; Zhang et al., 2020a).通过系统进化分析, 我们将sHSP家族进一步分为12个亚家族(图1).不同的亚家族含有不同数目的sHSP成员, 其中CI亚家族包含最多的sHSP成员, 这可能与CI亚家族包含最多的基因复制事件(4个)有关(表2).相似的分布也出现在番茄(张宁等, 2017)、马铃薯(Zhao et al., 2018)、棉花(Ma et al., 2016)、银杏(何福林和张斌, 2019)和辣椒(Guo et al., 2015)等植物中.因此, 在植物中sHSP家族具有保守的亚家族分布.在拟南芥中未发现任何属于CVI和CVII亚家族的成员, 而在葡萄中CI和CII亚家族的成员数目远多于另外3个物种, 这直接导致拟南芥和葡萄中含有不同数目的sHSP成员.类似现象也出现在棉花扩展蛋白基因家族(张奇艳等, 2019)、玉米(Zea mays) NAC家族(Fan et al., 2014)及大豆PP2C家族(Fan et al., 2020a).sHSP家族的保守基序分布和基因结构分布也与通过系统进化分析得出的亚家族分布一致(图2). ...
... 在睡莲、水稻、拟南芥和葡萄中共鉴定到12个与sHSP家族有关的基因复制事件(表2).相比其它2种植物, 在葡萄和拟南芥中发生的基因复制事件最多(各4个).葡萄中sHSP成员数量最多可能与葡萄发生的基因复制事件最多有关, 相似的研究结果也出现在葡萄的CDPL家族(Zhang et al., 2015)、卵形家族蛋白(Wang et al., 2018)以及WRKY家族中(Guo et al., 2014).在拟南芥中, 虽然与AtsHSP成员相关的基因复制事件最多, 但成员数量较少, 这可能与拟南芥基因复制事件后大量基因丢失有关(Cannon et al., 2004; Maere et al., 2005).结合染色体定位结果, 可以将12个与sHSP家族有关的基因复制事件进一步分为6个片段复制事件和6个串联复制事件, 并且每个物种中均含有这2种基因复制事件, 这表明sHSP家族的扩增不仅与大规模的片段复制事件有关, 还与小规模的串联复制事件有关(图3).相似的sHSP家族扩增模式也出现在辣椒(Guo et al., 2015)、簸箕柳(Li et al., 2018)和马铃薯(Zhao et al., 2018)等植物中.在每个物种中, 片段复制事件的Ks值均大于串联复制事件的Ks值, 这表明在sHSP家族中片段复制事件发生的时间均早于串联复制事件, 相似的现象也出现在梅花(Prunus mume) NAC家族(Zhuo et al., 2018)和棉花PP2C家族(Shazadee et al., 2019)中.基于复制事件的Ka/Ks比值表明, sHSP成员在发生复制事件以后会发生强烈的纯化选择作用, 类似的Ka/Ks比值分布也出现在其它植物的sHSP家族中(Ma et al., 2016; Li et al., 2018).与此同时, 与其它基因家族相比(Zhuo et al., 2018; Fan et al., 2020b), sHSP家族发生的基因复制事件较少(表2), 这也直接导致sHSP家族成员数相对较少, 表明在植物的进化过程中, sHSP成员可能具有相对保守的功能(如响应高温胁迫) (Waters and Vierling, 2020).此外, 本研究发现4个片段复制事件的Ks值大于1 (表2), 其中在拟南芥中有1个片段复制事件(AtsHSP01/AtsHSP06), 在另外3种植物中均发现对应的同源基因(图4), 该片段复制事件发生的时间大约为1-1.2亿年前温暖干旱的白垩纪(Zhang et al., 2020b), 这表明AtsHSP01/AtsHSP06及其在植物界的同源基因可能与植物适应高温环境有关, 可以作为今后研究高温胁迫的重要候选基因. ...
... 之前的研究表明, sHSP家族可以参与许多植物生长与发育过程(Zhang et al., 2018; Ma et al., 2019).本研究在睡莲、水稻、拟南芥和葡萄中新发现一些与植物生长发育过程相关的sHSP成员(图5).CI亚家族的NcsHSP19、OssHSP02、OssHSP14和VvsHSP40以及CIV亚家族的AtsHSP09和OssHSP23在几乎所有组织中均高度表达, 其在棉花和马铃薯中的同源基因也具有类似的表达模式(Ma et al., 2016; Zhao et al., 2018), 这表明sHSP家族在植物中的功能可能具有一定的保守性.此外, 本研究还发现一些与水稻花器官发育相关的基因(OssHSP03、OssHSP04、OssHSP06、OssHSP12、OssHSP16和OssHSP21).有报道显示, 在玫瑰(Rosa hybrida)中与CI亚家族OssHSP03、OssHSP04和OssHSP16相同亚家族的sHSP17.5-CI可能与开花过程中保护细胞蛋白有关(Dafny-Yelin et al., 2008). ...
Cloning and expression of a small heat shock protein gene CaHSP24 from pepper under abiotic stress 1 2011
... (Salix suchowensis) (Li et al., 2018)等植物中发现了大量的sHSP成员.sHSP家族主要作为分子伴侣, 以独立于ATP的形式保护植物免受外界环境的影响(Waters and Vierling, 2020).大量研究表明, sHSP成员可以响应外界多种环境胁迫(Sun et al., 2010; Zhu et al., 2011; Li et al., 2016; Zhai et al., 2016; Guo et al., 2020).此外, sHSP家族也参与果实品质形成和植株发芽等植物生长发育过程(Zhang et al., 2018; Ma et al., 2019). ...
Genome duplication and evolution of heat shock transcription factor (HSF) gene family in four model angiosperms 1 2016
Genome-wide analysis of the NAC transcription factor gene family reveals differential expression patterns and cold-stress responses in the woody plant Prunus mume 2 2018
... 在睡莲、水稻、拟南芥和葡萄中共鉴定到12个与sHSP家族有关的基因复制事件(表2).相比其它2种植物, 在葡萄和拟南芥中发生的基因复制事件最多(各4个).葡萄中sHSP成员数量最多可能与葡萄发生的基因复制事件最多有关, 相似的研究结果也出现在葡萄的CDPL家族(Zhang et al., 2015)、卵形家族蛋白(Wang et al., 2018)以及WRKY家族中(Guo et al., 2014).在拟南芥中, 虽然与AtsHSP成员相关的基因复制事件最多, 但成员数量较少, 这可能与拟南芥基因复制事件后大量基因丢失有关(Cannon et al., 2004; Maere et al., 2005).结合染色体定位结果, 可以将12个与sHSP家族有关的基因复制事件进一步分为6个片段复制事件和6个串联复制事件, 并且每个物种中均含有这2种基因复制事件, 这表明sHSP家族的扩增不仅与大规模的片段复制事件有关, 还与小规模的串联复制事件有关(图3).相似的sHSP家族扩增模式也出现在辣椒(Guo et al., 2015)、簸箕柳(Li et al., 2018)和马铃薯(Zhao et al., 2018)等植物中.在每个物种中, 片段复制事件的Ks值均大于串联复制事件的Ks值, 这表明在sHSP家族中片段复制事件发生的时间均早于串联复制事件, 相似的现象也出现在梅花(Prunus mume) NAC家族(Zhuo et al., 2018)和棉花PP2C家族(Shazadee et al., 2019)中.基于复制事件的Ka/Ks比值表明, sHSP成员在发生复制事件以后会发生强烈的纯化选择作用, 类似的Ka/Ks比值分布也出现在其它植物的sHSP家族中(Ma et al., 2016; Li et al., 2018).与此同时, 与其它基因家族相比(Zhuo et al., 2018; Fan et al., 2020b), sHSP家族发生的基因复制事件较少(表2), 这也直接导致sHSP家族成员数相对较少, 表明在植物的进化过程中, sHSP成员可能具有相对保守的功能(如响应高温胁迫) (Waters and Vierling, 2020).此外, 本研究发现4个片段复制事件的Ks值大于1 (表2), 其中在拟南芥中有1个片段复制事件(AtsHSP01/AtsHSP06), 在另外3种植物中均发现对应的同源基因(图4), 该片段复制事件发生的时间大约为1-1.2亿年前温暖干旱的白垩纪(Zhang et al., 2020b), 这表明AtsHSP01/AtsHSP06及其在植物界的同源基因可能与植物适应高温环境有关, 可以作为今后研究高温胁迫的重要候选基因. ...
... ).与此同时, 与其它基因家族相比(Zhuo et al., 2018; Fan et al., 2020b), sHSP家族发生的基因复制事件较少(表2), 这也直接导致sHSP家族成员数相对较少, 表明在植物的进化过程中, sHSP成员可能具有相对保守的功能(如响应高温胁迫) (Waters and Vierling, 2020).此外, 本研究发现4个片段复制事件的Ks值大于1 (表2), 其中在拟南芥中有1个片段复制事件(AtsHSP01/AtsHSP06), 在另外3种植物中均发现对应的同源基因(图4), 该片段复制事件发生的时间大约为1-1.2亿年前温暖干旱的白垩纪(Zhang et al., 2020b), 这表明AtsHSP01/AtsHSP06及其在植物界的同源基因可能与植物适应高温环境有关, 可以作为今后研究高温胁迫的重要候选基因. ...
 ,*福建农林大学农学院, 作物遗传育种与综合利用教育部重点实验室, 福州 350002
,*福建农林大学农学院, 作物遗传育种与综合利用教育部重点实验室, 福州 350002

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT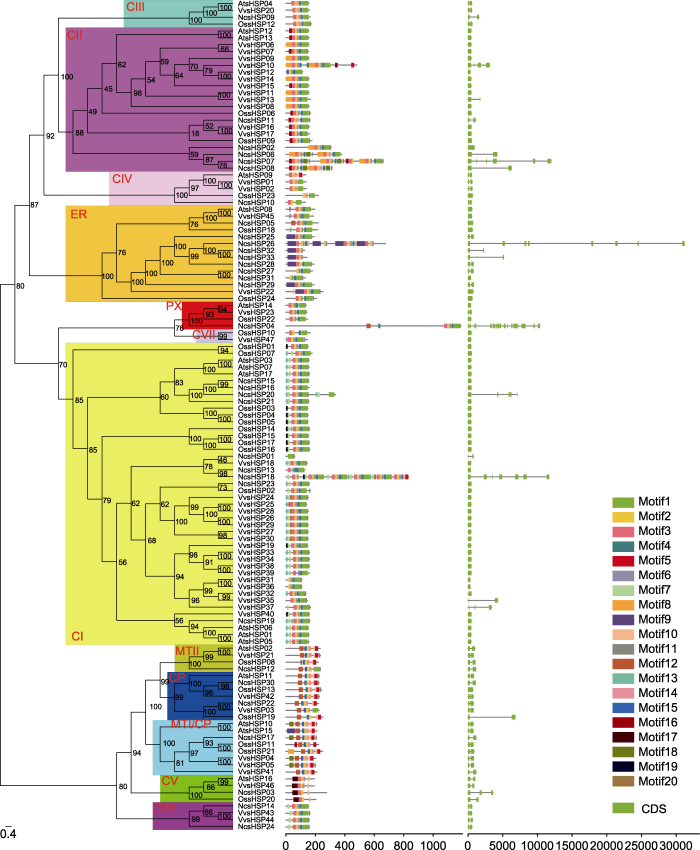 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT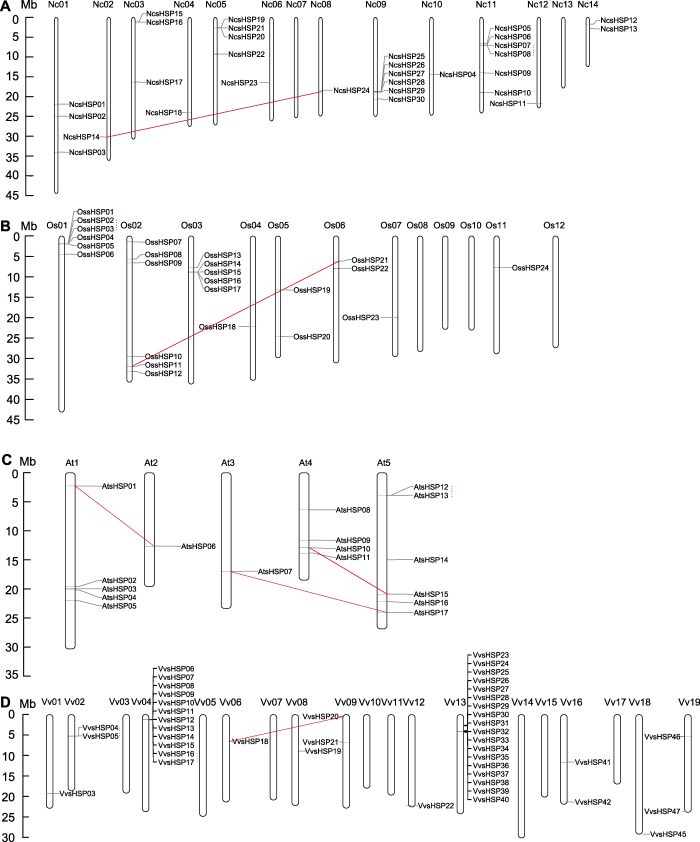 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT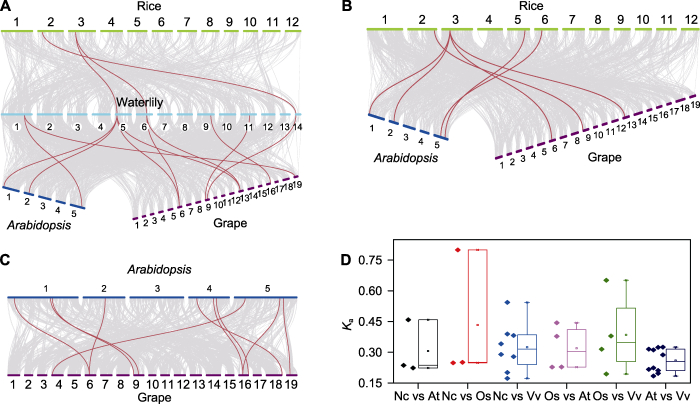 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT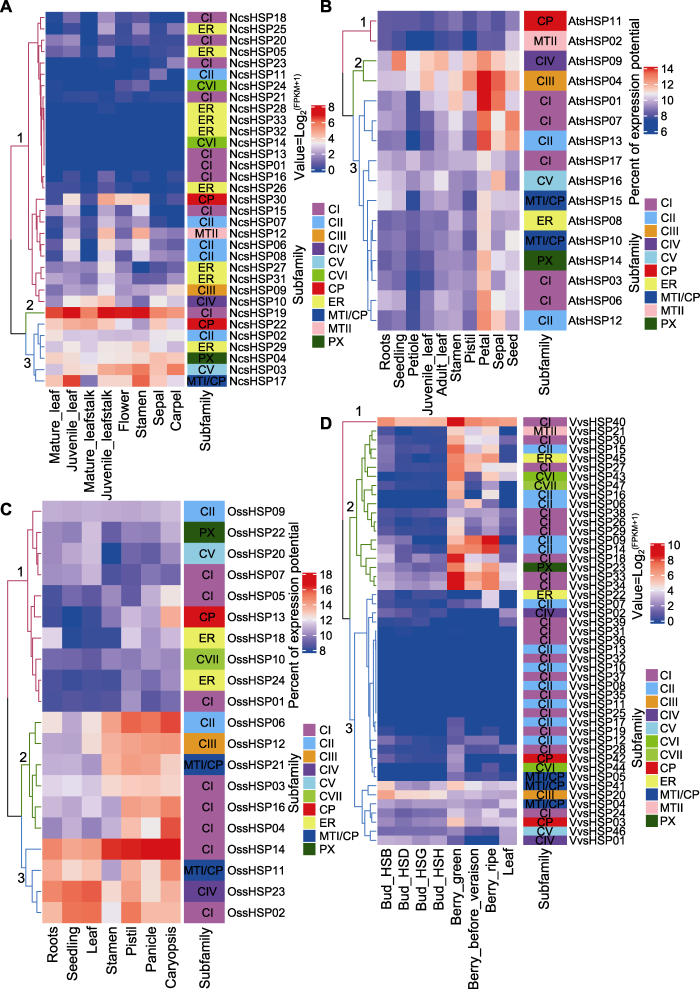 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}