Abstract As one of the largest transcription factor (TF) families in plants, MYB TFs are involved in various physiological and biochemical processes, such as plant growth, metabolism, and response to various biotic and abiotic stresses. R2R3-MYB is the main form of MYB TFs in higher plants. Pepper is a vegetable crop with important economic value, but the R2R3-MYB TF family has not been systematically studied in pepper. In this study, 94 CaMYB, 92 CbMYB, and 94 CcMYB TFs genes were identified with comparative genomic analysis in Capsicum annuum, C. baccatum, and C. chinense, respectively. These genes were categorized into 28 subfamilies. Collinearity analysis indicated that there were 73 groups of orthologous R2R3-MYB genes among three pepper species. There were five, four, and two unique R2R3-MYB genes in C. annuum, C. baccatum, and C. chinense, respectively. In addition, we identified 12 pairs of duplicated genes, and eight of which are tandemly repeated genes, which already existed before the divergence of three pepper species. Comparative genomics analysis suggested that the homologous R2R3-MYB TFs underwent functionally divergence during the evolution of pepper. Analysis on the expression profile showed that R2R3-MYB genes were expressed in three major patterns: high expression in roots, leaves, stems, and flowers, such as CaMYB13/CbMYB12/- CcMYB13; high expression in flowers, such as CaMYB93/CbMYB86/CcMYB12; high expression in roots, such as CaMYB48/CbMYB47/CcMYB51. These results lay a foundation for further study on the biological functions of R2R3-MYB TFs in the growth and development of pepper. Keywords:Capsicum annuum; C. baccatum; C. chinense;R2R3-MYB transcription factor;orthologous gene
PDF (3055KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 康凯程, 牛西强, 黄先忠, 胡能兵, 隋益虎, 张开京, 艾昊. 辣椒R2R3-MYB转录因子家族的全基因组鉴定与比较进化分析. 植物学报, 2021, 56(3): 315-329 doi:10.11983/CBB20143 Kang Kaicheng, Niu Xiqiang, Huang Xianzhong, Hu Nengbing, Sui Yihu, Zhang Kaijing, Ai Hao. Genome-wide Identification and Comparative Evolutionary Analysis of the R2R3-MYB Transcription Factor Gene Family in Pepper. Chinese Bulletin of Botany, 2021, 56(3): 315-329 doi:10.11983/CBB20143
氨基酸的得分(Bits)表示它在这个位点出现的频率。 Figure 1Sequence logos of the R2 (A) and R3 (B) MYB domains from three Capsicum species
The score of an amino acid (Bits) indicates the frequency of its occurrence at this site.
2.2 辣椒R2R3-MYB基因的分类和结构
为了研究辣椒R2R3-MYB基因的进化关系, 利用3种辣椒和拟南芥R2R3-MYB转录因子的蛋白序列构建系统进化树。与拟南芥R2R3-MYB转录因子聚集到同一分支的辣椒R2R3-MYB转录因子被认定为同一亚族, 可分为30个亚家族(S1-S30) (图2)。分类中的大多数亚家族与其它植物中的亚家族一致, 但是辣椒R2R3-MYB基因家族未发现有成员属于亚家族S10、S12和S15, 暗示辣椒基因组在长期进化过程中可能丢失了一些R2R3-MYB基因(图2, 图3A)。辣椒的近源物种马铃薯R2R3-MYB转录因子家族中不存在亚家族S10和S12也证明了这一点(Li et al., 2019; Sun et al., 2019)。
(A) 27个亚家族的系统进化树; (B) 27个亚家族的外显子-内含子结构(黑色方框表示外显子, 黑色线条表示内含子); (C) 27个亚家族的motif分布(不同颜色的方框表示不同的motif) Figure 3Phylogenetic tree, gene exon-intron structure and motif analysis of R2R3-MYB transcription factor family in three Capsicum species
(A) Phylogenetic tree of the 27 subfamilies; (B) Exon-intron structure of the 27 subfamilies (Black boxes indicate exons and black lines indicate introns); (C) The motif distribution of the 27 subfamilies (The boxes with different colors indicate different motifs)
2.3 3种辣椒R2R3-MYB转录因子的系统进化关系
在植物进化过程中经常会出现基因重复, 这也是植物基因家族发生扩张的重要原因之一(Cannon et al., 2004)。前人研究表明, 大约在170万年前浆果状辣椒和一年生辣椒、中国辣椒的祖先发生分化, 在110万年前一年生辣椒和中国辣椒也发生了分化(Kim et al., 2017)。为了确定3种辣椒间R2R3-MYB同源基因以及每种辣椒内部可能出现的基因重复现象, 本研究鉴定了3种辣椒中R2R3-MYB基因的进化关系。最终在3种辣椒中鉴定出73组直系同源R2R3-MYB基因(图4), 分别占一年生辣椒R2R3-MYB基因的77.66% (73/ 94)、浆果状辣椒R2R3-MYB基因的79.35% (73/92)和中国辣椒R2R3-MYB基因的77.66% (73/94)。这说明大多数R2R3-MYB基因在它们的祖先物种中存在, 并且在辣椒的进化过程中高度保守。
从外向内依次是3种辣椒的染色体数目、20 Mb窗口统计R2R3-MYB基因的数量(0-5)和3种辣椒中R2R3-MYB同源基因对。灰色线连接3种辣椒中都存在的同源基因对; 红色线连接只在两种辣椒中存在的同源基因对; 绿色线连接重复基因; 蓝色线连接串联重复基因。 Figure 5Homologous R2R3-MYB transcription factor gene pairs in Capsicum annuum (Ca), C. baccatum (Cb), and C. chinense (Cc)
Tracks from outside to inside are the number of chromosomes, number of R2R3-MYB genes (0-5) (20 Mb windows), and homologous gene pairs among three Capsicum species. The gray lines connect the homologous gene pairs that exist in three Capsicum species; the red lines connect the homologous gene pairs in two species; the green lines connect the replicated genes; and the blue lines connect the tandem repeated genes.
黑色点表示非同义替换率(Ka)/同义替换率(Ks)>1的基因对。 Figure 6The Ka/Ks ratios for the orthologous and duplicated gene pairs of R2R3-MYB transcription factor in Capsicum annuum, C. baccatum, and C. chinense
The black dots represent gene pairs with non-synonymous substitution rate (Ka)/synonymous substitution rate (Ks)>1.
Figure 7Expression profile of R2R3-MYB genes in different tissues from Capsicum annuum (Ca), C. baccatum (Cb), and C. chinense (Cc)
The color bars indicate the variation range of log10(FPKM+1) values of R2R3-MYB genes in different tissues. The expression of the genes in red was verified by qRT-PCR.
BlancG, WolfeKH (2004). Widespread paleopolyploidy in model plant species inferred from age distributions of duplicate genes Plant Cell 16, 1667-1678. DOI:10.1105/tpc.021345URL
CannonSB, MitraA, BaumgartenA, YoungND, MayG (2004). The roles of segmental and tandem gene duplication in the evolution of large gene families inArabidopsis thaliana BMC Plant Biol 4, 10. DOI:10.1186/1471-2229-4-10URL [本文引用: 1]
ChenYH, YangXY, HeK, LiuMH, LiJG, GaoZF, LinZQ, ZhangYF, WangXX, QiuXM, ShenYP, ZhangL, DengXH, LuoJC, DengXW, ChenZL, GuHY, QuLJ (2006). The MYB transcription factor superfamily of Arabidopsis: expression analysis and phylogenetic comparison with the rice MYB family Plant Mol Biol 60, 107-124. DOI:10.1007/s11103-005-2910-yURL [本文引用: 3]
CrooksGE, HonG, ChandoniaJM, BrennerSE (2004). WebLogo: a sequence logo generator Genome Res 14, 1188-1190. PMID:15173120 [本文引用: 1] WebLogo generates sequence logos, graphical representations of the patterns within a multiple sequence alignment. Sequence logos provide a richer and more precise description of sequence similarity than consensus sequences and can rapidly reveal significant features of the alignment otherwise difficult to perceive. Each logo consists of stacks of letters, one stack for each position in the sequence. The overall height of each stack indicates the sequence conservation at that position (measured in bits), whereas the height of symbols within the stack reflects the relative frequency of the corresponding amino or nucleic acid at that position. WebLogo has been enhanced recently with additional features and options, to provide a convenient and highly configurable sequence logo generator. A command line interface and the complete, open WebLogo source code are available for local installation and customization.Copyright 2004 Cold Spring Harbor Laboratory Press
DiasAP, BraunEL, McMullenMD, GrotewoldE (2003). Recently duplicated maize R2R3 Myb genes provide evidence for distinct mechanisms of evolutionary divergence after duplication Plant Physiol 131, 610-620. DOI:10.1104/pp.012047URL [本文引用: 1]
DossaK, MmadiMA, ZhouR, LiuAL, YangYX, DioufD, YouJ, ZhangXR (2020). Ectopic expression of the sesame MYB transcription factor SiMYB305 promotes root growth and modulates ABA-mediated tolerance to drought and salt stresses in Arabidopsis AoB Plants 12, plz081. DOI:10.1093/aobpla/plz081URL [本文引用: 1]
DuH, YangSS, LiangZ, FengBR, LiuL, HuangYB, TangYX (2012). Genome-wide analysis of the MYB transcription factor superfamily in soybean BMC Plant Biol 12, 106. DOI:10.1186/1471-2229-12-106URL [本文引用: 2]
HagaN, KatoK, MuraseM, ArakiS, KuboM, DemuraT, SuzukiK, MullerI, Vo?U, JürgensG, ItoM (2007). R1R2R3-Myb proteins positively regulate cytokinesis through activation of KNOLLE transcription in Arabidopsis thaliana Development 134, 1101-1110. DOI:10.1242/dev.02801URL [本文引用: 1]
HajiebrahimiA, OwjiH, HemmatiS (2017). Genome-wide identification, functional prediction, and evolutionary analysis of the R2R3-MYB superfamily in Brassica napus Genome 60, 797-814. DOI:10.1139/gen-2017-0059PMID:28732175 [本文引用: 2] R2R3-MYB transcription factors (TFs) have been shown to play important roles in plants, including in development and in various stress conditions. Phylogenetic analysis showed the presence of 249 R2R3-MYB TFs in Brassica napus, called BnaR2R3-MYB TFs, clustered into 38 clades. BnaR2R3-MYB TFs were distributed on 19 chromosomes of B. napus. Sixteen gene clusters were identified. BnaR2R3-MYB TFs were characterized by motif prediction, gene structure analysis, and gene ontology. Evolutionary analysis revealed that BnaR2R3-MYB TFs are mainly formed as a result of whole-genome duplication. Orthologs and paralogs of BnaR2R3-MYB TFs were identified in B. napus, B. rapa, B. oleracea, and Arabidopsis thaliana using synteny-based methods. Purifying selection was pervasive within R2R3-MYB TFs. K/K values lower than 0.3 indicated that BnaR2R3-MYB TFs are being functionally converged. The role of gene conversion in the formation of BnaR2R3-MYB TFs was significant. Cis-regulatory elements in the upstream regions of BnaR2R3-MYB genes, miRNA targeting BnaR2R3MYB TFs, and post translational modifications were identified. Digital expression data revealed that BnaR2R3-MYB genes were highly expressed in the roots and under high salinity treatment after 24 h. BnaMYB21, BnaMYB141, and BnaMYB148 have been suggested for improving salt-tolerant B. napus. BnaR2R3-MYB genes were mostly up regulated on the 14th day post inoculation with Leptosphaeria biglobosa and L. maculan. BnaMYB150 is a candidate for increased tolerance to Leptospheria in B. napus.
HeberleH, MeirellesGV, da SilvaFR, TellesGP, MinghimR (2015). InteractiVenn: a web-based tool for the analysis of sets through Venn diagrams BMC Bioinformatics 16, 169. DOI:10.1186/s12859-015-0611-3PMID:25994840 [本文引用: 1] Background: Set comparisons permeate a large number of data analysis workflows, in particular workflows in biological sciences. Venn diagrams are frequently employed for such analysis but current tools are limited. Results: We have developed InteractiVenn, a more flexible tool for interacting with Venn diagrams including up to six sets. It offers a clean interface for Venn diagram construction and enables analysis of set unions while preserving the shape of the diagram. Set unions are useful to reveal differences and similarities among sets and may be guided in our tool by a tree or by a list of set unions. The tool also allows obtaining subsets' elements, saving and loading sets for further analyses, and exporting the diagram in vector and image formats. InteractiVenn has been used to analyze two biological datasets, but it may serve set analysis in a broad range of domains. Conclusions: InteractiVenn allows set unions in Venn diagrams to be explored thoroughly, by consequence extending the ability to analyze combinations of sets with additional observations, yielded by novel interactions between joined sets. InteractiVenn is freely available online at: www.interactivenn.net.
IbizaVP, BlancaJ, Ca?izaresJ, NuezF (2012). Taxonomy and genetic diversity of domesticated Capsicum species in the Andean region Genet Resour Crop Evol 59, 1077-1088. DOI:10.1007/s10722-011-9744-zURL [本文引用: 1]
JiaL, CleggMT, JiangT (2004). Evolutionary dynamics of the DNA-binding domains in putative R2R3-MYB genes identified from rice subspecies indica and japonica genomes Plant Physiol 134, 575-585. PMID:14966247 [本文引用: 1] The molecular evolution of the R2R3-MYB gene family is of great interest because it is one of the most important transcription factor gene families in the plant kingdom. Comparative analyses of a gene family may reveal important adaptive changes at the protein level and thereby provide insights that relate structure to function. We have performed a range of comparative and bioinformatics analyses on R2R3-MYB genes identified from the rice (Oryza sativa subsp. japonica and indica) and Arabidopsis genome sequences. The study provides an initial framework to investigate how different evolutionary lineages in a gene family evolve new functions. Our results reveal a remarkable excess of non-synonymous substitutions, an indication of adaptive selection on protein structure that occurred during the evolution of both helix1 and helix2 of rice R2R3-MYB DNA-binding domains. These flexible alpha-helix regions associated with high frequencies of excess non-synonymous substitutions may play critical roles in the characteristic packing of R2R3-MYB DNA-binding domains and thereby modify the protein-DNA interaction process resulting in the recognition of novel DNA-binding sites. Furthermore, a co-evolutionary pattern is found between the second alpha-helix of the R2 domain and the second alpha-helix of the R3 domain by examining all the possible alpha-helix pairings in both the R2 and R3 domains. This points to the functional importance of pairing interactions between related secondary structures.
JiangCZ, GuJY, ChopraS, GuX, PetersonT (2004). Ordered origin of the typical two- and three-repeat MYB genes Gene 326, 13-22. DOI:10.1016/j.gene.2003.09.049URL [本文引用: 1]
KasugaM, LiuQ, MiuraS, Yamaguchi-ShinozakiK, ShinozakiK (1999). Improving plant drought, salt, and freezing tolerance by gene transfer of a single stress-inducible transcription factor Nat Biotechnol 17, 287-291. PMID:10096298 [本文引用: 1] Plant productivity is greatly affected by environmental stresses such as drought, salt loading, and freezing. We reported previously that a cis-acting promoter element, the dehydration response element (DRE), plays an important role in regulating gene expression in response to these stresses. The transcription factor DREB1A specifically interacts with the DRE and induces expression of stress tolerance genes. We show here that overexpression of the cDNA encoding DREB1A in transgenic plants activated the expression of many of these stress tolerance genes under normal growing conditions and resulted in improved tolerance to drought, salt loading, and freezing. However, use of the strong constitutive 35S cauliflower mosaic virus (CaMV) promoter to drive expression of DREB1A also resulted in severe growth retardation under normal growing conditions. In contrast, expression of DREB1A from the stress inducible rd29A promoter gave rise to minimal effects on plant growth while providing an even greater tolerance to stress conditions than did expression of the gene from the CaMV promoter.
LangmeadB, SalzbergSL (2012). Fast gapped-read alignment with Bowtie 2 Nat Methods 9, 357-359. DOI:10.1038/nmeth.1923URL [本文引用: 1]
LiB, DeweyCN (2011). RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome BMC Bioinformatics 12, 323. DOI:10.1186/1471-2105-12-323URL [本文引用: 1]
LiXX, GuoC, AhmadS, WangQ, YuJ, LiuC, GuoYF (2019). Systematic analysis of MYB family genes in potato and their multiple roles in development and stress responses Biomolecules 9, 317. DOI:10.3390/biom9080317URL [本文引用: 5]
LiuYH, KuiLW, EspleyRV, WangL, LiYM, LiuZ, ZhouP, ZengLH, ZhangXJ, ZhangJL, AllanAC (2019). StMYB44 negatively regulates anthocyanin biosynthesis at high temperatures in tuber flesh of potato J Exp Bot 70, 3809-3824. DOI:10.1093/jxb/erz194URL [本文引用: 1]
LivakKJ, SchmittgenTD (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2 ?ΔΔCT method Methods 25, 402-408. PMID:11846609 [本文引用: 1] The two most commonly used methods to analyze data from real-time, quantitative PCR experiments are absolute quantification and relative quantification. Absolute quantification determines the input copy number, usually by relating the PCR signal to a standard curve. Relative quantification relates the PCR signal of the target transcript in a treatment group to that of another sample such as an untreated control. The 2(-Delta Delta C(T)) method is a convenient way to analyze the relative changes in gene expression from real-time quantitative PCR experiments. The purpose of this report is to present the derivation, assumptions, and applications of the 2(-Delta Delta C(T)) method. In addition, we present the derivation and applications of two variations of the 2(-Delta Delta C(T)) method that may be useful in the analysis of real-time, quantitative PCR data.Copyright 2001 Elsevier Science (USA).
LotkowskaME, TohgeT, FernieAR, XueGP, BalazadehS, Mueller-RoeberB (2015). The Arabidopsis transcription factor MYB112 promotes anthocyanin formation during salinity and under high light stress Plant Physiol 169, 1862-1880. [本文引用: 1]
MatusJT, AqueaF, Arce-JohnsonP (2008). Analysis of the grape MYB R2R3 subfamily reveals expanded wine quality-related clades and conserved gene structure organization across Vitis and Arabidopsis genomes BMC Plant Biol 8, 83. DOI:10.1186/1471-2229-8-83URL [本文引用: 1]
OgataK, Kanei-IshiiC, SasakiM, HatanakaH, NagadoiA, EnariM, NakamuraH, NishimuraY, IshiiS, SaraiA (1996). The cavity in the hydrophobic core of Myb DNA- binding domain is reserved for DNA recognition and trans- activation Nat Struct Biol 3, 178-187. PMID:8564545 [本文引用: 1] The DNA-binding domain of Myb consists of three imperfect repeats, R1, R2 and R3, each containing a helix-turn-helix motif variation. Among these repeats, R2 has distinct characteristics with high thermal instability. The NMR structure analysis found a cavity inside the hydrophobic core of R2 but not in R1 or R3. Here, we show that R2 has slow conformational fluctuations, and that a cavity-filling mutation which stabilizes the R2 structure significantly reduces specific Myb DNA-binding activity and trans-activation. Structural observations of the free and DNA-complexed stages suggest that the implied inherent conformational flexibility of R2, associated with the presence of the cavity, could be important for DNA recognition by Myb.
PaboCO, SauerRT (1992). Transcription factors: structural families and principles of DNA recognition Annu Rev Biochem 61, 1053-1095. PMID:1497306 [本文引用: 1]
Paz-AresJ, GhosalD, WienandU, PetersonPA, SaedlerH (1987). The regulatory c1 locus of Zea mays encodes a protein with homology to myb proto-oncogene products and with structural similarities to transcriptional activators EMBO J 6, 3553-3558. PMID:3428265 [本文引用: 1] The structure of the wild-type c1 locus of Zea mays was determined by sequence analysis of one genomic and two cDNA clones. The coding region is composed of three exons (150 bp, 129 bp and one, at least 720 bp) and two small introns (88 bp and 145 bp). Transcription of the mRNAs corresponding to the two cDNA clones cLC6 (1.1 kb) and cLC28 (2.1 kb) starts from the same promoter. Both cDNAs are identical except that cLC28 extends further at its 3' end. A putative protein, 273 amino acids in length was deduced from the sequence of both transcripts. It contains two domains, one basic and the other acidic and might function as a transcriptional activator. The basic domain of this c1-encoded protein shows 40% sequence homology to the protein products of animal myb proto-oncogenes.
QiXW, FangHL, ChenZQ, LiuZQ, YuX, LiangCY (2019). Ectopic expression of a R2R3-MYB transcription factor gene LjaMYB12 from Lonicera japonica increases flavonoid accumulation in Arabidopsis thaliana Int J Mol Sci 20, 4494. DOI:10.3390/ijms20184494URL [本文引用: 1]
RosinskiJA, AtchleyWR (1998). Molecular evolution of the MYB family of transcription factors: evidence for polyphyletic origin J Mol Evol 46, 74-83. PMID:9419227 [本文引用: 1] The Myb family of proteins is a group of functionally diverse transcriptional activators found in both plants and animals that is characterized by a conserved DNA-binding domain of approximately 50 amino acids. Phylogenetic analyses of amino acid sequences of this family of proteins portray very disparate evolutionary histories in plants and animals. Animal Myb proteins have diverged from a common ancestor, while plants appear related only within the DNA-binding domain. Results imply a pattern of modular evolution of the Myb proteins centering on the possession of a helix-turn-helix motif. Based on this it is suggested that Myb proteins are a polyphyletic group related only by a "Myb-box" DNA-binding motif.
SchmutzJ, CannonSB, SchlueterJ, MaJX, MitrosT, NelsonW, HytenDL, SongQJ, ThelenJJ, ChengJL, XuD, HellstenU, MayGD, YuY, SakuraiT, UmezawaT, BhattacharyyaMK, SandhuD, ValliyodanB, LindquistE, PetoM, GrantD, ShuSQ, GoodsteinD, BarryK, Futrell-GriggsM, AbernathyB, DuJC, TianZX, ZhuLC, GillN, JoshiT, LibaultM, SethuramanA, ZhangXC, ShinozakiK, NguyenHT, WingRA, CreganP, SpechtJ, GrimwoodJ, RokhsarD, StaceyG, Shoema- kerRC, JacksonSA (2010). Genome sequence of the palaeopolyploid soybean Nature 463, 178-183. DOI:10.1038/nature08670PMID:20075913 [本文引用: 1] Soybean (Glycine max) is one of the most important crop plants for seed protein and oil content, and for its capacity to fix atmospheric nitrogen through symbioses with soil-borne microorganisms. We sequenced the 1.1-gigabase genome by a whole-genome shotgun approach and integrated it with physical and high-density genetic maps to create a chromosome-scale draft sequence assembly. We predict 46,430 protein-coding genes, 70% more than Arabidopsis and similar to the poplar genome which, like soybean, is an ancient polyploid (palaeopolyploid). About 78% of the predicted genes occur in chromosome ends, which comprise less than one-half of the genome but account for nearly all of the genetic recombination. Genome duplications occurred at approximately 59 and 13 million years ago, resulting in a highly duplicated genome with nearly 75% of the genes present in multiple copies. The two duplication events were followed by gene diversification and loss, and numerous chromosome rearrangements. An accurate soybean genome sequence will facilitate the identification of the genetic basis of many soybean traits, and accelerate the creation of improved soybean varieties.
StrackeR, IshiharaH, HuepG, BarschA, MehrtensF, NiehausK, WeisshaarB (2007). Differential regulation of closely related R2R3-MYB transcription factors controls flavonol accumulation in different parts of the Arabidopsis thaliana seedling Plant J 50, 660-677. PMID:17419845 [本文引用: 1] The genes MYB11, MYB12 and MYB111 share significant structural similarity and form subgroup 7 of the Arabidopsis thaliana R2R3-MYB gene family. To determine the regulatory potential of these three transcription factors, we used a combination of genetic, functional genomics and metabolite analysis approaches. MYB11, MYB12 and MYB111 show a high degree of functional similarity and display very similar target gene specificity for several genes of flavonoid biosynthesis, including CHALCONE SYNTHASE, CHALCONE ISOMERASE, FLAVANONE 3-HYDROXYLASE and FLAVONOL SYNTHASE1. Seedlings of the triple mutant myb11 myb12 myb111, which genetically lack a complete subgroup of R2R3-MYB genes, do not form flavonols while the accumulation of anthocyanins is not affected. In developing seedlings, MYB11, MYB12 and MYB111 act in an additive manner due to their differential spatial activity; MYB12 controls flavonol biosynthesis mainly in the root, while MYB111 controls flavonol biosynthesis primarily in cotyledons. We identified and confirmed additional target genes of the R2R3-MYB subgroup 7 factors, including the UDP-glycosyltransferases UGT91A1 and UGT84A1, and we demonstrate that the accumulation of distinct and structurally identified flavonol glycosides in seedlings correlates with the expression domains of the different R2R3-MYB factors. Therefore, we refer to these genes as PFG1-3 for 'PRODUCTION OF FLAVONOL GLYCOSIDES'.
StrackeR, WerberM, WeisshaarB (2001). The R2R3- MYB gene family in Arabidopsis thaliana Curr Opin Plant Biol 4, 447-456. PMID:11597504 [本文引用: 3] MYB factors represent a family of proteins that include the conserved MYB DNA-binding domain. In contrast to animals, plants contain a MYB-protein subfamily that is characterised by the R2R3-type MYB domain. 'Classical' MYB factors, which are related to c-Myb, seem to be involved in the control of the cell cycle in animals, plants and other higher eukaryotes. Systematic screens for knockout mutations in MYB genes, followed by phenotypic analyses and the dissection of mutants with interesting phenotypes, have started to unravel the functions of the 125 R2R3-MYB genes in Arabidopsis thaliana. R2R3-type MYB genes control many aspects of plant secondary metabolism, as well as the identity and fate of plant cells.
SunWJ, MaZT, ChenH, LiuMY (2019). MYB gene family in potato ( Solanum tuberosum L.): genome-wide identification of hormone-responsive reveals their potential functions in growth and development Int J Mol Sci 20, 4847. DOI:10.3390/ijms20194847URL [本文引用: 5]
The Potato Genome Sequencing Consortium (2011). Genome sequence and analysis of the tuber crop potato Nature 475, 189-195. DOI:10.1038/nature10158URL [本文引用: 1]
The Tomato GenomeConsortium (2012). The tomato genome sequence provides insights into fleshy fruit evolution Nature 485, 635-641. DOI:10.1038/nature11119URL [本文引用: 1]
UllahA, Ul QamarMT, NisarM, HazratA, RahimG, KhanAH, HayatK, AhmedS, AliW, KhanA, YangXY (2020). Characterization of a novel cotton MYB gene, GhMYB108- like responsive to abiotic stresses Mol Biol Rep 47, 1573-1581. DOI:10.1007/s11033-020-05244-6URL [本文引用: 1]
ValliyodanB, NguyenHT (2006). Understanding regulatory networks and engineering for enhanced drought tolerance in plants Curr Opin Plant Biol 9, 189-195. PMID:16483835 [本文引用: 1] Drought stress is one of the major limitations to crop productivity. To develop crop plants with enhanced tolerance of drought stress, a basic understanding of physiological, biochemical and gene regulatory networks is essential. Various functional genomics tools have helped to advance our understanding of stress signal perception and transduction, and of the associated molecular regulatory network. These tools have revealed several stress-inducible genes and various transcription factors that regulate the drought-stress-inducible systems. Translational genomics of these candidate genes using model plants provided encouraging results, but the field testing of transgenic crop plants for better performance and yield is still minimal. Better understanding of the specific roles of various metabolites in crop stress tolerance will give rise to a strategy for the metabolic engineering of crop tolerance of drought.
WangDP, WanHL, ZhangS, YuJ (2009). γ-MYN: a new algorithm for estimating Ka and Ks with consideration of variable substitution rates Biol Direct 4, 20. DOI:10.1186/1745-6150-4-20URL [本文引用: 1]
WangDP, ZhangYB, ZhangZ, ZhuJ, YuJ (2010). KaKs_Calculator 2.0: a toolkit incorporating gamma-series methods and sliding window strategies Genom Proteom Bioinf 8, 77-80. DOI:10.1016/S1672-0229(10)60008-3URL [本文引用: 1]
WangYC, LiuWJ, JiangHY, MaoZL, WangN, JiangSH, XuHF, YangGX, ZhangZY, ChenXS (2019). The R2R3-MYB transcription factor MdMYB24-like is involved in methyl jasmonate-induced anthocyanin biosynthesis in apple Plant Physiol Biochem 139, 273-282. DOI:10.1016/j.plaphy.2019.03.031URL [本文引用: 1]
WangYP, TangHB, DebarryJD, TanX, LiJP, WangXY, LeeTH, JinHZ, MarlerB, GuoH, KissingerJC, PatersonAH (2012). MCScanX: a toolkit for detection and evolutionary analysis of gene synteny and collinearity Nucleic Acids Res 40, e49. DOI:10.1093/nar/gkr1293URL [本文引用: 1]
YadavCB, BonthalaVS, MuthamilarasanM, PandeyG, KhanY, PrasadM (2015). Genome-wide development of transposable elements-based markers in foxtail millet and construction of an integrated database DNA Res 22, 79-90. DOI:10.1093/dnares/dsu039URL [本文引用: 2]
ZapataL, DingJ, WillingEM, HartwigB, BezdanD, JiaoWB, PatelV, Velikkakam JamesG, KoornneefM, OssowskiS, SchneebergerK (2016). Chromosome-level assembly of Arabidopsis thaliana Ler reveals the extent of translocation and inversion polymorphisms Proc Natl Acad Sci USA 113, E4052-E4060. DOI:10.1073/pnas.1607532113URL [本文引用: 1]
... MYB转录因子广泛分布于植物基因组中, 是植物中最大的转录因子家族之一(Riechmann et al., 2000; Dubos et al., 2010).首个植物MYB转录因子是从玉米(Zea mays)中鉴定出的COLORED1 (Paz-Ares et al., 1987).目前对MYB转录因子研究不断深入, R2R3-MYB转录因子已成为研究重点之一.据报道, R2R3-MYB转录因子参与调控许多重要的生理生化过程, 包括调节植物的初级和次生代谢、控制生长发育以及参与响应各种生物和非生物胁迫(Stracke et al., 2007).例如, 马铃薯(Solanum tuberosum) StMYB44在高温下负调控花青素的生物合成(Liu et al., 2019); 金银花(Lonicera japonica) LjaMYB12在拟南芥(Arabidopsis thaliana)中异位表达可以提高类黄酮含量, 其对类黄酮的生物合成的上下游途径具有不同的调控模式(Qi et al., 2019); 拟南芥AtMYB20、AtMYB42、AtMYB43和AtMYB85能够特异性地抑制类黄酮的生物合成(Geng et al., 2020); 棉花(Gossypium hirsutum) GhMYB108-like在干旱和盐胁迫下发挥重要作用(Ullah et al., 2020); 苹果(Malus domestica) MdMYB24-like参与茉莉酸甲酯诱导的花青素苷生物合成(Wang et al., 2019), MdMYB30在苹果角质层蜡质的积累中起重要作用, 并能增强苹果的抗病性(Zhang et al., 2019); 芝麻(Sesamum indicum) SiMYB75在拟南芥中的异位过表达显著促进根系生长, 提高植株对干旱、盐胁迫和渗透胁迫的耐受性(Dossa et al., 2020); 盐、干旱及脱落酸胁迫促进甘薯(Ipomoea batatas) IbMYB3的表达, 暗示IbMYB3在甘薯非生物胁迫应答中起重要作用(李格等, 2020). ...
辣椒属种间远缘杂交育种研究进展 1 2009
... 随着测序技术的发展, R2R3-MYB转录因子家族先后在许多高等植物的全基因组水平得到系统鉴定和分析, 包括拟南芥(Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(Brassica napus) (Hajiebrahimi et al., 2017)、大豆(Glycine max) (Du et al., 2012)、番茄(Solanum lycopersicum) (Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019).基因组测序技术促进了园艺植物基因组学和遗传学研究(唐嘉瓅等, 2020).辣椒是茄科(Solanaceae)辣椒属(Capsicum)一年生或多年生草本植物, 是具有重要经济价值的蔬菜作物, 在世界范围内广泛种植.辣椒属包含27种, 被人类驯化长期栽培的辣椒有5种, 分别为一年生辣椒(Capsicum annuum)、浆果状辣椒(C. baccatum)、中国辣椒(C. chinense)、灌木状辣椒(C. frutescens)和绒毛辣椒(C. pubescens) (Ibiza et al., 2012).我国主要栽培品种是一年生辣椒, 其它4个栽培种具有许多优良性状, 是对一年生辣椒进行品种改良的重要种质资源(隋益虎和陈劲枫, 2009; 魏家香等, 2018).本研究以一年生辣椒、浆果状辣椒和中国辣椒为材料, 对3种辣椒R2R3-MYB转录因子家族在全基因组范围内进行鉴定, 在此基础上分析了辣椒R2R3-MYB基因的序列、motif分布、外显子-内含子结构、染色体定位、比较进化和不同组织(根、茎、叶和花)中的表达情况.通过共线性分析, 发现3种辣椒都存在特有的R2R3-MYB基因, 一年生辣椒和浆果状辣椒中的R2R3-MYB转录因子家族发生了基因组重复事件.研究结果为揭示辣椒R2R3-MYB转录因子功能及辣椒分子遗传育种奠定了理论基础. ...
基因组学技术大发展助力园艺植物研究取得新进展 1 2020
... 随着测序技术的发展, R2R3-MYB转录因子家族先后在许多高等植物的全基因组水平得到系统鉴定和分析, 包括拟南芥(Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(Brassica napus) (Hajiebrahimi et al., 2017)、大豆(Glycine max) (Du et al., 2012)、番茄(Solanum lycopersicum) (Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019).基因组测序技术促进了园艺植物基因组学和遗传学研究(唐嘉瓅等, 2020).辣椒是茄科(Solanaceae)辣椒属(Capsicum)一年生或多年生草本植物, 是具有重要经济价值的蔬菜作物, 在世界范围内广泛种植.辣椒属包含27种, 被人类驯化长期栽培的辣椒有5种, 分别为一年生辣椒(Capsicum annuum)、浆果状辣椒(C. baccatum)、中国辣椒(C. chinense)、灌木状辣椒(C. frutescens)和绒毛辣椒(C. pubescens) (Ibiza et al., 2012).我国主要栽培品种是一年生辣椒, 其它4个栽培种具有许多优良性状, 是对一年生辣椒进行品种改良的重要种质资源(隋益虎和陈劲枫, 2009; 魏家香等, 2018).本研究以一年生辣椒、浆果状辣椒和中国辣椒为材料, 对3种辣椒R2R3-MYB转录因子家族在全基因组范围内进行鉴定, 在此基础上分析了辣椒R2R3-MYB基因的序列、motif分布、外显子-内含子结构、染色体定位、比较进化和不同组织(根、茎、叶和花)中的表达情况.通过共线性分析, 发现3种辣椒都存在特有的R2R3-MYB基因, 一年生辣椒和浆果状辣椒中的R2R3-MYB转录因子家族发生了基因组重复事件.研究结果为揭示辣椒R2R3-MYB转录因子功能及辣椒分子遗传育种奠定了理论基础. ...
辣椒种间杂交的现状及其研究进展 1 2018
... 随着测序技术的发展, R2R3-MYB转录因子家族先后在许多高等植物的全基因组水平得到系统鉴定和分析, 包括拟南芥(Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(Brassica napus) (Hajiebrahimi et al., 2017)、大豆(Glycine max) (Du et al., 2012)、番茄(Solanum lycopersicum) (Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019).基因组测序技术促进了园艺植物基因组学和遗传学研究(唐嘉瓅等, 2020).辣椒是茄科(Solanaceae)辣椒属(Capsicum)一年生或多年生草本植物, 是具有重要经济价值的蔬菜作物, 在世界范围内广泛种植.辣椒属包含27种, 被人类驯化长期栽培的辣椒有5种, 分别为一年生辣椒(Capsicum annuum)、浆果状辣椒(C. baccatum)、中国辣椒(C. chinense)、灌木状辣椒(C. frutescens)和绒毛辣椒(C. pubescens) (Ibiza et al., 2012).我国主要栽培品种是一年生辣椒, 其它4个栽培种具有许多优良性状, 是对一年生辣椒进行品种改良的重要种质资源(隋益虎和陈劲枫, 2009; 魏家香等, 2018).本研究以一年生辣椒、浆果状辣椒和中国辣椒为材料, 对3种辣椒R2R3-MYB转录因子家族在全基因组范围内进行鉴定, 在此基础上分析了辣椒R2R3-MYB基因的序列、motif分布、外显子-内含子结构、染色体定位、比较进化和不同组织(根、茎、叶和花)中的表达情况.通过共线性分析, 发现3种辣椒都存在特有的R2R3-MYB基因, 一年生辣椒和浆果状辣椒中的R2R3-MYB转录因子家族发生了基因组重复事件.研究结果为揭示辣椒R2R3-MYB转录因子功能及辣椒分子遗传育种奠定了理论基础. ...
Widespread paleopolyploidy in model plant species inferred from age distributions of duplicate genes 0 2004
The roles of segmental and tandem gene duplication in the evolution of large gene families inArabidopsis thaliana 1 2004
... 在植物进化过程中经常会出现基因重复, 这也是植物基因家族发生扩张的重要原因之一(Cannon et al., 2004).前人研究表明, 大约在170万年前浆果状辣椒和一年生辣椒、中国辣椒的祖先发生分化, 在110万年前一年生辣椒和中国辣椒也发生了分化(Kim et al., 2017).为了确定3种辣椒间R2R3-MYB同源基因以及每种辣椒内部可能出现的基因重复现象, 本研究鉴定了3种辣椒中R2R3-MYB基因的进化关系.最终在3种辣椒中鉴定出73组直系同源R2R3-MYB基因(图4), 分别占一年生辣椒R2R3-MYB基因的77.66% (73/ 94)、浆果状辣椒R2R3-MYB基因的79.35% (73/92)和中国辣椒R2R3-MYB基因的77.66% (73/94).这说明大多数R2R3-MYB基因在它们的祖先物种中存在, 并且在辣椒的进化过程中高度保守. ...
Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome 1 2014
... 随着基因测序技术的不断发展, 已在基因组水平上对许多植物的R2R3-MYB转录因子家族进行了鉴定, 如拟南芥(125个) (Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(249个) (Hajiebrahimi et al., 2017)、大豆(244个) (Du et al., 2012)、番茄(121个) (Zhao et al., 2014)和马铃薯(112个) (Li et al., 2019; Sun et al., 2019), 它们的基因组大小分别为125 Mb (Zapata et al., 2016)、912 Mb (Chalhoub et al., 2014)、1.03 Gb (Schmutz et al., 2010)、828 Mb (The Tomato Genome Consortium, 2012)和663 Mb (The Potato Genome Sequencing Consortium, 2011).这表明物种中R2R3-MYB基因数目与物种基因组的大小并不成正比.一年生辣椒、浆果状辣椒和中国辣椒的基因组大小分别为2.85、2.99和2.80 Gb (Kim et al., 2017).本研究从中分别鉴定出94、92和94个R2R3-MYB基因, 也支持这一观点.与茄科植物番茄(Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019)相比, 3种辣椒中R2R3-MYB基因数目明显较少, 说明R2R3-MYB转录因子家族在辣椒中发生了明显收缩. ...
Fastp: an ultra- fast all-in-one FASTQ preprocessor 1 2018
... 一年生辣椒、浆果状辣椒和中国辣椒的转录组测序数据从BioProject数据库(https://www.ncbi.nlm.nih.gov/ bioproject)中获得, 登录号分别为PRJNA223222、PRJNA308879和PRJNA331024.测序样品为生长6周植株的根、茎和叶组织和生长4个月植株盛开的花(Kim et al., 2017).使用fastp (Chen et al., 2018)对测序数据进行过滤, 参数设为默认值.使用RSEM (Li and Dewey, 2011)将过滤后的数据比对到对应物种的基因组序列上, 比对方式为bowtie2 (Langmead and Salzberg, 2012), 参数设为默认值.将比对结果进行标准化处理, 得到基因的每百万个转录片段中每千碱基片段的映射读数(fragments per kilobase of exon model per million mapped reads, FPKM).FPKM值经log10 (FPKM+1)转换后, 使用R包pheatmap (https://CRAN.R-project.org/package=pheatmap)绘制热图. ...
The MYB transcription factor superfamily of Arabidopsis: expression analysis and phylogenetic comparison with the rice MYB family 3 2006
... 转录因子在基因转录调节中起重要作用, 通过激活或抑制靶基因的转录, 调控植物的生长发育(Pabo and Sauer, 1992; Kasuga et al., 1999; Riechmann et al., 2000; Valliyodan and Nguyen, 2006).根据转录因子与其靶基因结合位点的相似性, 可将其分成不同的家族, 如AP2/EREBP、bHLH、HB、MYB和WRKY (Riechmann et al., 2000).MYB转录因子具有与DNA结合的特征性MYB结构域, 该结构域在植物中高度保守.MYB转录因子包含1-4个不完全重复的MYB结构域, 分别命名为R1 (Repeat1)、R2、R3和R4, 每个MYB结构域约有52个氨基酸残基, 形成3个α-螺旋, 其中第2和第3螺旋形成螺旋-转角-螺旋(helix-turn-helix)结构并与DNA大沟槽结合(Ogata et al., 1996; Jia et al., 2004; Dubos et al., 2010).根据包含的MYB结构域数目, MYB转录因子可分为4类, 即1R-MYB、2R-MYB (R2R3-MYB)、3R-MYB (R1R2R3- MYB)和4R-MYB (Dubos et al., 2010).1R-MYB转录因子又称MYB-related转录因子, 包含1个完整或部分MYB结构域(Stracke et al., 2001; Chen et al., 2006).3R-MYB转录因子包含3个连续的MYB结构域(R1、R2和R3), 在大多数真核生物基因组中都发现了3R-MYB转录因子, 其在调控细胞周期中发挥作用(Haga et al., 2007).4R-MYB转录因子是最小的一类, 每个基因都含有4个R1/R2 MYB结构域, 它们在植物中发挥的作用尚不明确.包含2个MYB结构域(R2和R3)的R2R3-MYB转录因子是高等植物中MYB转录因子的主要存在形式, 该家族基因可能是因3R-MYB转录因子中R1 MYB结构域缺失进化而来(Rosinski and Atchley, 1998), 也有观点认为3R-MYB转录因子是从R2R3-MYB转录因子进化来的, R1 MYB结构域从R2R3-MYB转录因子获得(Jiang et al., 2004).通常在MYB转录因子的N末端检测到高度保守的MYB结构域, C末端MYB结构域是高度分化的激活结构域, 这也决定了MYB转录因子具有多种调节作用(Dias et al., 2003; Matus et al., 2008). ...
... 随着测序技术的发展, R2R3-MYB转录因子家族先后在许多高等植物的全基因组水平得到系统鉴定和分析, 包括拟南芥(Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(Brassica napus) (Hajiebrahimi et al., 2017)、大豆(Glycine max) (Du et al., 2012)、番茄(Solanum lycopersicum) (Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019).基因组测序技术促进了园艺植物基因组学和遗传学研究(唐嘉瓅等, 2020).辣椒是茄科(Solanaceae)辣椒属(Capsicum)一年生或多年生草本植物, 是具有重要经济价值的蔬菜作物, 在世界范围内广泛种植.辣椒属包含27种, 被人类驯化长期栽培的辣椒有5种, 分别为一年生辣椒(Capsicum annuum)、浆果状辣椒(C. baccatum)、中国辣椒(C. chinense)、灌木状辣椒(C. frutescens)和绒毛辣椒(C. pubescens) (Ibiza et al., 2012).我国主要栽培品种是一年生辣椒, 其它4个栽培种具有许多优良性状, 是对一年生辣椒进行品种改良的重要种质资源(隋益虎和陈劲枫, 2009; 魏家香等, 2018).本研究以一年生辣椒、浆果状辣椒和中国辣椒为材料, 对3种辣椒R2R3-MYB转录因子家族在全基因组范围内进行鉴定, 在此基础上分析了辣椒R2R3-MYB基因的序列、motif分布、外显子-内含子结构、染色体定位、比较进化和不同组织(根、茎、叶和花)中的表达情况.通过共线性分析, 发现3种辣椒都存在特有的R2R3-MYB基因, 一年生辣椒和浆果状辣椒中的R2R3-MYB转录因子家族发生了基因组重复事件.研究结果为揭示辣椒R2R3-MYB转录因子功能及辣椒分子遗传育种奠定了理论基础. ...
... 随着基因测序技术的不断发展, 已在基因组水平上对许多植物的R2R3-MYB转录因子家族进行了鉴定, 如拟南芥(125个) (Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(249个) (Hajiebrahimi et al., 2017)、大豆(244个) (Du et al., 2012)、番茄(121个) (Zhao et al., 2014)和马铃薯(112个) (Li et al., 2019; Sun et al., 2019), 它们的基因组大小分别为125 Mb (Zapata et al., 2016)、912 Mb (Chalhoub et al., 2014)、1.03 Gb (Schmutz et al., 2010)、828 Mb (The Tomato Genome Consortium, 2012)和663 Mb (The Potato Genome Sequencing Consortium, 2011).这表明物种中R2R3-MYB基因数目与物种基因组的大小并不成正比.一年生辣椒、浆果状辣椒和中国辣椒的基因组大小分别为2.85、2.99和2.80 Gb (Kim et al., 2017).本研究从中分别鉴定出94、92和94个R2R3-MYB基因, 也支持这一观点.与茄科植物番茄(Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019)相比, 3种辣椒中R2R3-MYB基因数目明显较少, 说明R2R3-MYB转录因子家族在辣椒中发生了明显收缩. ...
WebLogo: a sequence logo generator 1 2004
... 一年生辣椒(Capsicum annuum L.) (ECW)、浆果状辣椒(Capsicum baccatum L.) (PBC81)和中国辣椒(Capsicum chinense Jacq.) (PI159236)的基因组数据从辣椒基因组平台(http://peppergenome.snu.ac.kr)中获得(Kim et al., 2017).从拟南芥(Arabidopsis thaliana L.)全基因组数据库(https://www.arabidopsis.org/)中下载拟南芥R2R3-MYB转录因子家族的氨基酸序列作为query序列, 分别与3种辣椒的蛋白序列进行BLASTP比对, 参数设置E-value≤1e-10, 每条query保留5条匹配结果, 其余参数为默认值.将得到的候选序列使用hmmer (http://www.hmmer.org/)比对到PFAM32.0数据库中, 基于Pfam的MYB模型(PF00249)进行进一步比对筛选, 参数为默认值, 从而确定辣椒的R2R3-MYB转录因子基因成员.使用WebLogo (Crooks et al., 2004)绘制R2R3-MYB结构域seqlogo图. ...
Recently duplicated maize R2R3 Myb genes provide evidence for distinct mechanisms of evolutionary divergence after duplication 1 2003
... 转录因子在基因转录调节中起重要作用, 通过激活或抑制靶基因的转录, 调控植物的生长发育(Pabo and Sauer, 1992; Kasuga et al., 1999; Riechmann et al., 2000; Valliyodan and Nguyen, 2006).根据转录因子与其靶基因结合位点的相似性, 可将其分成不同的家族, 如AP2/EREBP、bHLH、HB、MYB和WRKY (Riechmann et al., 2000).MYB转录因子具有与DNA结合的特征性MYB结构域, 该结构域在植物中高度保守.MYB转录因子包含1-4个不完全重复的MYB结构域, 分别命名为R1 (Repeat1)、R2、R3和R4, 每个MYB结构域约有52个氨基酸残基, 形成3个α-螺旋, 其中第2和第3螺旋形成螺旋-转角-螺旋(helix-turn-helix)结构并与DNA大沟槽结合(Ogata et al., 1996; Jia et al., 2004; Dubos et al., 2010).根据包含的MYB结构域数目, MYB转录因子可分为4类, 即1R-MYB、2R-MYB (R2R3-MYB)、3R-MYB (R1R2R3- MYB)和4R-MYB (Dubos et al., 2010).1R-MYB转录因子又称MYB-related转录因子, 包含1个完整或部分MYB结构域(Stracke et al., 2001; Chen et al., 2006).3R-MYB转录因子包含3个连续的MYB结构域(R1、R2和R3), 在大多数真核生物基因组中都发现了3R-MYB转录因子, 其在调控细胞周期中发挥作用(Haga et al., 2007).4R-MYB转录因子是最小的一类, 每个基因都含有4个R1/R2 MYB结构域, 它们在植物中发挥的作用尚不明确.包含2个MYB结构域(R2和R3)的R2R3-MYB转录因子是高等植物中MYB转录因子的主要存在形式, 该家族基因可能是因3R-MYB转录因子中R1 MYB结构域缺失进化而来(Rosinski and Atchley, 1998), 也有观点认为3R-MYB转录因子是从R2R3-MYB转录因子进化来的, R1 MYB结构域从R2R3-MYB转录因子获得(Jiang et al., 2004).通常在MYB转录因子的N末端检测到高度保守的MYB结构域, C末端MYB结构域是高度分化的激活结构域, 这也决定了MYB转录因子具有多种调节作用(Dias et al., 2003; Matus et al., 2008). ...
Ectopic expression of the sesame MYB transcription factor SiMYB305 promotes root growth and modulates ABA-mediated tolerance to drought and salt stresses in Arabidopsis 1 2020
... MYB转录因子广泛分布于植物基因组中, 是植物中最大的转录因子家族之一(Riechmann et al., 2000; Dubos et al., 2010).首个植物MYB转录因子是从玉米(Zea mays)中鉴定出的COLORED1 (Paz-Ares et al., 1987).目前对MYB转录因子研究不断深入, R2R3-MYB转录因子已成为研究重点之一.据报道, R2R3-MYB转录因子参与调控许多重要的生理生化过程, 包括调节植物的初级和次生代谢、控制生长发育以及参与响应各种生物和非生物胁迫(Stracke et al., 2007).例如, 马铃薯(Solanum tuberosum) StMYB44在高温下负调控花青素的生物合成(Liu et al., 2019); 金银花(Lonicera japonica) LjaMYB12在拟南芥(Arabidopsis thaliana)中异位表达可以提高类黄酮含量, 其对类黄酮的生物合成的上下游途径具有不同的调控模式(Qi et al., 2019); 拟南芥AtMYB20、AtMYB42、AtMYB43和AtMYB85能够特异性地抑制类黄酮的生物合成(Geng et al., 2020); 棉花(Gossypium hirsutum) GhMYB108-like在干旱和盐胁迫下发挥重要作用(Ullah et al., 2020); 苹果(Malus domestica) MdMYB24-like参与茉莉酸甲酯诱导的花青素苷生物合成(Wang et al., 2019), MdMYB30在苹果角质层蜡质的积累中起重要作用, 并能增强苹果的抗病性(Zhang et al., 2019); 芝麻(Sesamum indicum) SiMYB75在拟南芥中的异位过表达显著促进根系生长, 提高植株对干旱、盐胁迫和渗透胁迫的耐受性(Dossa et al., 2020); 盐、干旱及脱落酸胁迫促进甘薯(Ipomoea batatas) IbMYB3的表达, 暗示IbMYB3在甘薯非生物胁迫应答中起重要作用(李格等, 2020). ...
Genome-wide analysis of the MYB transcription factor superfamily in soybean 2 2012
... 随着测序技术的发展, R2R3-MYB转录因子家族先后在许多高等植物的全基因组水平得到系统鉴定和分析, 包括拟南芥(Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(Brassica napus) (Hajiebrahimi et al., 2017)、大豆(Glycine max) (Du et al., 2012)、番茄(Solanum lycopersicum) (Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019).基因组测序技术促进了园艺植物基因组学和遗传学研究(唐嘉瓅等, 2020).辣椒是茄科(Solanaceae)辣椒属(Capsicum)一年生或多年生草本植物, 是具有重要经济价值的蔬菜作物, 在世界范围内广泛种植.辣椒属包含27种, 被人类驯化长期栽培的辣椒有5种, 分别为一年生辣椒(Capsicum annuum)、浆果状辣椒(C. baccatum)、中国辣椒(C. chinense)、灌木状辣椒(C. frutescens)和绒毛辣椒(C. pubescens) (Ibiza et al., 2012).我国主要栽培品种是一年生辣椒, 其它4个栽培种具有许多优良性状, 是对一年生辣椒进行品种改良的重要种质资源(隋益虎和陈劲枫, 2009; 魏家香等, 2018).本研究以一年生辣椒、浆果状辣椒和中国辣椒为材料, 对3种辣椒R2R3-MYB转录因子家族在全基因组范围内进行鉴定, 在此基础上分析了辣椒R2R3-MYB基因的序列、motif分布、外显子-内含子结构、染色体定位、比较进化和不同组织(根、茎、叶和花)中的表达情况.通过共线性分析, 发现3种辣椒都存在特有的R2R3-MYB基因, 一年生辣椒和浆果状辣椒中的R2R3-MYB转录因子家族发生了基因组重复事件.研究结果为揭示辣椒R2R3-MYB转录因子功能及辣椒分子遗传育种奠定了理论基础. ...
... 随着基因测序技术的不断发展, 已在基因组水平上对许多植物的R2R3-MYB转录因子家族进行了鉴定, 如拟南芥(125个) (Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(249个) (Hajiebrahimi et al., 2017)、大豆(244个) (Du et al., 2012)、番茄(121个) (Zhao et al., 2014)和马铃薯(112个) (Li et al., 2019; Sun et al., 2019), 它们的基因组大小分别为125 Mb (Zapata et al., 2016)、912 Mb (Chalhoub et al., 2014)、1.03 Gb (Schmutz et al., 2010)、828 Mb (The Tomato Genome Consortium, 2012)和663 Mb (The Potato Genome Sequencing Consortium, 2011).这表明物种中R2R3-MYB基因数目与物种基因组的大小并不成正比.一年生辣椒、浆果状辣椒和中国辣椒的基因组大小分别为2.85、2.99和2.80 Gb (Kim et al., 2017).本研究从中分别鉴定出94、92和94个R2R3-MYB基因, 也支持这一观点.与茄科植物番茄(Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019)相比, 3种辣椒中R2R3-MYB基因数目明显较少, 说明R2R3-MYB转录因子家族在辣椒中发生了明显收缩. ...
MYB transcription factors in Arabidopsis 3 2010
... 转录因子在基因转录调节中起重要作用, 通过激活或抑制靶基因的转录, 调控植物的生长发育(Pabo and Sauer, 1992; Kasuga et al., 1999; Riechmann et al., 2000; Valliyodan and Nguyen, 2006).根据转录因子与其靶基因结合位点的相似性, 可将其分成不同的家族, 如AP2/EREBP、bHLH、HB、MYB和WRKY (Riechmann et al., 2000).MYB转录因子具有与DNA结合的特征性MYB结构域, 该结构域在植物中高度保守.MYB转录因子包含1-4个不完全重复的MYB结构域, 分别命名为R1 (Repeat1)、R2、R3和R4, 每个MYB结构域约有52个氨基酸残基, 形成3个α-螺旋, 其中第2和第3螺旋形成螺旋-转角-螺旋(helix-turn-helix)结构并与DNA大沟槽结合(Ogata et al., 1996; Jia et al., 2004; Dubos et al., 2010).根据包含的MYB结构域数目, MYB转录因子可分为4类, 即1R-MYB、2R-MYB (R2R3-MYB)、3R-MYB (R1R2R3- MYB)和4R-MYB (Dubos et al., 2010).1R-MYB转录因子又称MYB-related转录因子, 包含1个完整或部分MYB结构域(Stracke et al., 2001; Chen et al., 2006).3R-MYB转录因子包含3个连续的MYB结构域(R1、R2和R3), 在大多数真核生物基因组中都发现了3R-MYB转录因子, 其在调控细胞周期中发挥作用(Haga et al., 2007).4R-MYB转录因子是最小的一类, 每个基因都含有4个R1/R2 MYB结构域, 它们在植物中发挥的作用尚不明确.包含2个MYB结构域(R2和R3)的R2R3-MYB转录因子是高等植物中MYB转录因子的主要存在形式, 该家族基因可能是因3R-MYB转录因子中R1 MYB结构域缺失进化而来(Rosinski and Atchley, 1998), 也有观点认为3R-MYB转录因子是从R2R3-MYB转录因子进化来的, R1 MYB结构域从R2R3-MYB转录因子获得(Jiang et al., 2004).通常在MYB转录因子的N末端检测到高度保守的MYB结构域, C末端MYB结构域是高度分化的激活结构域, 这也决定了MYB转录因子具有多种调节作用(Dias et al., 2003; Matus et al., 2008). ...
... ).根据包含的MYB结构域数目, MYB转录因子可分为4类, 即1R-MYB、2R-MYB (R2R3-MYB)、3R-MYB (R1R2R3- MYB)和4R-MYB (Dubos et al., 2010).1R-MYB转录因子又称MYB-related转录因子, 包含1个完整或部分MYB结构域(Stracke et al., 2001; Chen et al., 2006).3R-MYB转录因子包含3个连续的MYB结构域(R1、R2和R3), 在大多数真核生物基因组中都发现了3R-MYB转录因子, 其在调控细胞周期中发挥作用(Haga et al., 2007).4R-MYB转录因子是最小的一类, 每个基因都含有4个R1/R2 MYB结构域, 它们在植物中发挥的作用尚不明确.包含2个MYB结构域(R2和R3)的R2R3-MYB转录因子是高等植物中MYB转录因子的主要存在形式, 该家族基因可能是因3R-MYB转录因子中R1 MYB结构域缺失进化而来(Rosinski and Atchley, 1998), 也有观点认为3R-MYB转录因子是从R2R3-MYB转录因子进化来的, R1 MYB结构域从R2R3-MYB转录因子获得(Jiang et al., 2004).通常在MYB转录因子的N末端检测到高度保守的MYB结构域, C末端MYB结构域是高度分化的激活结构域, 这也决定了MYB转录因子具有多种调节作用(Dias et al., 2003; Matus et al., 2008). ...
... MYB转录因子广泛分布于植物基因组中, 是植物中最大的转录因子家族之一(Riechmann et al., 2000; Dubos et al., 2010).首个植物MYB转录因子是从玉米(Zea mays)中鉴定出的COLORED1 (Paz-Ares et al., 1987).目前对MYB转录因子研究不断深入, R2R3-MYB转录因子已成为研究重点之一.据报道, R2R3-MYB转录因子参与调控许多重要的生理生化过程, 包括调节植物的初级和次生代谢、控制生长发育以及参与响应各种生物和非生物胁迫(Stracke et al., 2007).例如, 马铃薯(Solanum tuberosum) StMYB44在高温下负调控花青素的生物合成(Liu et al., 2019); 金银花(Lonicera japonica) LjaMYB12在拟南芥(Arabidopsis thaliana)中异位表达可以提高类黄酮含量, 其对类黄酮的生物合成的上下游途径具有不同的调控模式(Qi et al., 2019); 拟南芥AtMYB20、AtMYB42、AtMYB43和AtMYB85能够特异性地抑制类黄酮的生物合成(Geng et al., 2020); 棉花(Gossypium hirsutum) GhMYB108-like在干旱和盐胁迫下发挥重要作用(Ullah et al., 2020); 苹果(Malus domestica) MdMYB24-like参与茉莉酸甲酯诱导的花青素苷生物合成(Wang et al., 2019), MdMYB30在苹果角质层蜡质的积累中起重要作用, 并能增强苹果的抗病性(Zhang et al., 2019); 芝麻(Sesamum indicum) SiMYB75在拟南芥中的异位过表达显著促进根系生长, 提高植株对干旱、盐胁迫和渗透胁迫的耐受性(Dossa et al., 2020); 盐、干旱及脱落酸胁迫促进甘薯(Ipomoea batatas) IbMYB3的表达, 暗示IbMYB3在甘薯非生物胁迫应答中起重要作用(李格等, 2020). ...
MYB20, MYB42, MYB43, and MYB85 regulate phenylalanine and lignin biosynthesis during secondary cell wall formation 1 2020
... MYB转录因子广泛分布于植物基因组中, 是植物中最大的转录因子家族之一(Riechmann et al., 2000; Dubos et al., 2010).首个植物MYB转录因子是从玉米(Zea mays)中鉴定出的COLORED1 (Paz-Ares et al., 1987).目前对MYB转录因子研究不断深入, R2R3-MYB转录因子已成为研究重点之一.据报道, R2R3-MYB转录因子参与调控许多重要的生理生化过程, 包括调节植物的初级和次生代谢、控制生长发育以及参与响应各种生物和非生物胁迫(Stracke et al., 2007).例如, 马铃薯(Solanum tuberosum) StMYB44在高温下负调控花青素的生物合成(Liu et al., 2019); 金银花(Lonicera japonica) LjaMYB12在拟南芥(Arabidopsis thaliana)中异位表达可以提高类黄酮含量, 其对类黄酮的生物合成的上下游途径具有不同的调控模式(Qi et al., 2019); 拟南芥AtMYB20、AtMYB42、AtMYB43和AtMYB85能够特异性地抑制类黄酮的生物合成(Geng et al., 2020); 棉花(Gossypium hirsutum) GhMYB108-like在干旱和盐胁迫下发挥重要作用(Ullah et al., 2020); 苹果(Malus domestica) MdMYB24-like参与茉莉酸甲酯诱导的花青素苷生物合成(Wang et al., 2019), MdMYB30在苹果角质层蜡质的积累中起重要作用, 并能增强苹果的抗病性(Zhang et al., 2019); 芝麻(Sesamum indicum) SiMYB75在拟南芥中的异位过表达显著促进根系生长, 提高植株对干旱、盐胁迫和渗透胁迫的耐受性(Dossa et al., 2020); 盐、干旱及脱落酸胁迫促进甘薯(Ipomoea batatas) IbMYB3的表达, 暗示IbMYB3在甘薯非生物胁迫应答中起重要作用(李格等, 2020). ...
R1R2R3-Myb proteins positively regulate cytokinesis through activation of KNOLLE transcription in Arabidopsis thaliana 1 2007
... 转录因子在基因转录调节中起重要作用, 通过激活或抑制靶基因的转录, 调控植物的生长发育(Pabo and Sauer, 1992; Kasuga et al., 1999; Riechmann et al., 2000; Valliyodan and Nguyen, 2006).根据转录因子与其靶基因结合位点的相似性, 可将其分成不同的家族, 如AP2/EREBP、bHLH、HB、MYB和WRKY (Riechmann et al., 2000).MYB转录因子具有与DNA结合的特征性MYB结构域, 该结构域在植物中高度保守.MYB转录因子包含1-4个不完全重复的MYB结构域, 分别命名为R1 (Repeat1)、R2、R3和R4, 每个MYB结构域约有52个氨基酸残基, 形成3个α-螺旋, 其中第2和第3螺旋形成螺旋-转角-螺旋(helix-turn-helix)结构并与DNA大沟槽结合(Ogata et al., 1996; Jia et al., 2004; Dubos et al., 2010).根据包含的MYB结构域数目, MYB转录因子可分为4类, 即1R-MYB、2R-MYB (R2R3-MYB)、3R-MYB (R1R2R3- MYB)和4R-MYB (Dubos et al., 2010).1R-MYB转录因子又称MYB-related转录因子, 包含1个完整或部分MYB结构域(Stracke et al., 2001; Chen et al., 2006).3R-MYB转录因子包含3个连续的MYB结构域(R1、R2和R3), 在大多数真核生物基因组中都发现了3R-MYB转录因子, 其在调控细胞周期中发挥作用(Haga et al., 2007).4R-MYB转录因子是最小的一类, 每个基因都含有4个R1/R2 MYB结构域, 它们在植物中发挥的作用尚不明确.包含2个MYB结构域(R2和R3)的R2R3-MYB转录因子是高等植物中MYB转录因子的主要存在形式, 该家族基因可能是因3R-MYB转录因子中R1 MYB结构域缺失进化而来(Rosinski and Atchley, 1998), 也有观点认为3R-MYB转录因子是从R2R3-MYB转录因子进化来的, R1 MYB结构域从R2R3-MYB转录因子获得(Jiang et al., 2004).通常在MYB转录因子的N末端检测到高度保守的MYB结构域, C末端MYB结构域是高度分化的激活结构域, 这也决定了MYB转录因子具有多种调节作用(Dias et al., 2003; Matus et al., 2008). ...
Genome-wide identification, functional prediction, and evolutionary analysis of the R2R3-MYB superfamily in Brassica napus 2 2017
... 随着测序技术的发展, R2R3-MYB转录因子家族先后在许多高等植物的全基因组水平得到系统鉴定和分析, 包括拟南芥(Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(Brassica napus) (Hajiebrahimi et al., 2017)、大豆(Glycine max) (Du et al., 2012)、番茄(Solanum lycopersicum) (Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019).基因组测序技术促进了园艺植物基因组学和遗传学研究(唐嘉瓅等, 2020).辣椒是茄科(Solanaceae)辣椒属(Capsicum)一年生或多年生草本植物, 是具有重要经济价值的蔬菜作物, 在世界范围内广泛种植.辣椒属包含27种, 被人类驯化长期栽培的辣椒有5种, 分别为一年生辣椒(Capsicum annuum)、浆果状辣椒(C. baccatum)、中国辣椒(C. chinense)、灌木状辣椒(C. frutescens)和绒毛辣椒(C. pubescens) (Ibiza et al., 2012).我国主要栽培品种是一年生辣椒, 其它4个栽培种具有许多优良性状, 是对一年生辣椒进行品种改良的重要种质资源(隋益虎和陈劲枫, 2009; 魏家香等, 2018).本研究以一年生辣椒、浆果状辣椒和中国辣椒为材料, 对3种辣椒R2R3-MYB转录因子家族在全基因组范围内进行鉴定, 在此基础上分析了辣椒R2R3-MYB基因的序列、motif分布、外显子-内含子结构、染色体定位、比较进化和不同组织(根、茎、叶和花)中的表达情况.通过共线性分析, 发现3种辣椒都存在特有的R2R3-MYB基因, 一年生辣椒和浆果状辣椒中的R2R3-MYB转录因子家族发生了基因组重复事件.研究结果为揭示辣椒R2R3-MYB转录因子功能及辣椒分子遗传育种奠定了理论基础. ...
... 随着基因测序技术的不断发展, 已在基因组水平上对许多植物的R2R3-MYB转录因子家族进行了鉴定, 如拟南芥(125个) (Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(249个) (Hajiebrahimi et al., 2017)、大豆(244个) (Du et al., 2012)、番茄(121个) (Zhao et al., 2014)和马铃薯(112个) (Li et al., 2019; Sun et al., 2019), 它们的基因组大小分别为125 Mb (Zapata et al., 2016)、912 Mb (Chalhoub et al., 2014)、1.03 Gb (Schmutz et al., 2010)、828 Mb (The Tomato Genome Consortium, 2012)和663 Mb (The Potato Genome Sequencing Consortium, 2011).这表明物种中R2R3-MYB基因数目与物种基因组的大小并不成正比.一年生辣椒、浆果状辣椒和中国辣椒的基因组大小分别为2.85、2.99和2.80 Gb (Kim et al., 2017).本研究从中分别鉴定出94、92和94个R2R3-MYB基因, 也支持这一观点.与茄科植物番茄(Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019)相比, 3种辣椒中R2R3-MYB基因数目明显较少, 说明R2R3-MYB转录因子家族在辣椒中发生了明显收缩. ...
InteractiVenn: a web-based tool for the analysis of sets through Venn diagrams 1 2015
... 共线性分析使用MCScanX (Wang et al., 2012)预测物种间和物种内存在的同源基因, 参数设为默认值.将3种辣椒的基因组进行两两比较, 设置E-value≤ 1e-10, 提取R2R3-MYB基因家族中的基因重复对信息, 可筛选出3种辣椒间的直系同源基因; 分别将3种辣椒的基因组与自身基因组进行比较, 设置E-value≤ 1e-10, 根据结果筛选出3种辣椒内的旁系同源基因, 即重复基因; 如果1对重复基因位于染色体上的邻近位置(<100 kb), 就可以认定为串联重复基因.使用InteractiVenn (Heberle et al., 2015)绘制Venn图.使用Circos (Krzywinski et al., 2009)显示一年生辣椒、浆果状辣椒和中国辣椒同源基因的共线性关系.使用ParaAT2.0 (Zhang et al., 2012)对同源基因对的CDS序列和ORF序列进行比对, 参数设为默认值; 使用KaKs_Calculator2.0 (Wang et al., 2010)读取比对结果, 采用γ-MYN法(Wang et al., 2009)计算同源基因对的非同义替换率(Ka)和同义替换率(Ks), 通过Ka/Ks值来评估同源基因对中同源基因在进化过程中受到的选择压力.Ka/Ks>1, 基因受到正选择; Ka/Ks<1, 基因受到纯化选择; Ka/Ks=1, 基因中性进化(Yadav et al., 2015). ...
GSDS 2.0: an upgraded gene feature visualization server 1 2015
Taxonomy and genetic diversity of domesticated Capsicum species in the Andean region 1 2012
... 随着测序技术的发展, R2R3-MYB转录因子家族先后在许多高等植物的全基因组水平得到系统鉴定和分析, 包括拟南芥(Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(Brassica napus) (Hajiebrahimi et al., 2017)、大豆(Glycine max) (Du et al., 2012)、番茄(Solanum lycopersicum) (Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019).基因组测序技术促进了园艺植物基因组学和遗传学研究(唐嘉瓅等, 2020).辣椒是茄科(Solanaceae)辣椒属(Capsicum)一年生或多年生草本植物, 是具有重要经济价值的蔬菜作物, 在世界范围内广泛种植.辣椒属包含27种, 被人类驯化长期栽培的辣椒有5种, 分别为一年生辣椒(Capsicum annuum)、浆果状辣椒(C. baccatum)、中国辣椒(C. chinense)、灌木状辣椒(C. frutescens)和绒毛辣椒(C. pubescens) (Ibiza et al., 2012).我国主要栽培品种是一年生辣椒, 其它4个栽培种具有许多优良性状, 是对一年生辣椒进行品种改良的重要种质资源(隋益虎和陈劲枫, 2009; 魏家香等, 2018).本研究以一年生辣椒、浆果状辣椒和中国辣椒为材料, 对3种辣椒R2R3-MYB转录因子家族在全基因组范围内进行鉴定, 在此基础上分析了辣椒R2R3-MYB基因的序列、motif分布、外显子-内含子结构、染色体定位、比较进化和不同组织(根、茎、叶和花)中的表达情况.通过共线性分析, 发现3种辣椒都存在特有的R2R3-MYB基因, 一年生辣椒和浆果状辣椒中的R2R3-MYB转录因子家族发生了基因组重复事件.研究结果为揭示辣椒R2R3-MYB转录因子功能及辣椒分子遗传育种奠定了理论基础. ...
Evolutionary dynamics of the DNA-binding domains in putative R2R3-MYB genes identified from rice subspecies indica and japonica genomes 1 2004
... 转录因子在基因转录调节中起重要作用, 通过激活或抑制靶基因的转录, 调控植物的生长发育(Pabo and Sauer, 1992; Kasuga et al., 1999; Riechmann et al., 2000; Valliyodan and Nguyen, 2006).根据转录因子与其靶基因结合位点的相似性, 可将其分成不同的家族, 如AP2/EREBP、bHLH、HB、MYB和WRKY (Riechmann et al., 2000).MYB转录因子具有与DNA结合的特征性MYB结构域, 该结构域在植物中高度保守.MYB转录因子包含1-4个不完全重复的MYB结构域, 分别命名为R1 (Repeat1)、R2、R3和R4, 每个MYB结构域约有52个氨基酸残基, 形成3个α-螺旋, 其中第2和第3螺旋形成螺旋-转角-螺旋(helix-turn-helix)结构并与DNA大沟槽结合(Ogata et al., 1996; Jia et al., 2004; Dubos et al., 2010).根据包含的MYB结构域数目, MYB转录因子可分为4类, 即1R-MYB、2R-MYB (R2R3-MYB)、3R-MYB (R1R2R3- MYB)和4R-MYB (Dubos et al., 2010).1R-MYB转录因子又称MYB-related转录因子, 包含1个完整或部分MYB结构域(Stracke et al., 2001; Chen et al., 2006).3R-MYB转录因子包含3个连续的MYB结构域(R1、R2和R3), 在大多数真核生物基因组中都发现了3R-MYB转录因子, 其在调控细胞周期中发挥作用(Haga et al., 2007).4R-MYB转录因子是最小的一类, 每个基因都含有4个R1/R2 MYB结构域, 它们在植物中发挥的作用尚不明确.包含2个MYB结构域(R2和R3)的R2R3-MYB转录因子是高等植物中MYB转录因子的主要存在形式, 该家族基因可能是因3R-MYB转录因子中R1 MYB结构域缺失进化而来(Rosinski and Atchley, 1998), 也有观点认为3R-MYB转录因子是从R2R3-MYB转录因子进化来的, R1 MYB结构域从R2R3-MYB转录因子获得(Jiang et al., 2004).通常在MYB转录因子的N末端检测到高度保守的MYB结构域, C末端MYB结构域是高度分化的激活结构域, 这也决定了MYB转录因子具有多种调节作用(Dias et al., 2003; Matus et al., 2008). ...
Ordered origin of the typical two- and three-repeat MYB genes 1 2004
... 转录因子在基因转录调节中起重要作用, 通过激活或抑制靶基因的转录, 调控植物的生长发育(Pabo and Sauer, 1992; Kasuga et al., 1999; Riechmann et al., 2000; Valliyodan and Nguyen, 2006).根据转录因子与其靶基因结合位点的相似性, 可将其分成不同的家族, 如AP2/EREBP、bHLH、HB、MYB和WRKY (Riechmann et al., 2000).MYB转录因子具有与DNA结合的特征性MYB结构域, 该结构域在植物中高度保守.MYB转录因子包含1-4个不完全重复的MYB结构域, 分别命名为R1 (Repeat1)、R2、R3和R4, 每个MYB结构域约有52个氨基酸残基, 形成3个α-螺旋, 其中第2和第3螺旋形成螺旋-转角-螺旋(helix-turn-helix)结构并与DNA大沟槽结合(Ogata et al., 1996; Jia et al., 2004; Dubos et al., 2010).根据包含的MYB结构域数目, MYB转录因子可分为4类, 即1R-MYB、2R-MYB (R2R3-MYB)、3R-MYB (R1R2R3- MYB)和4R-MYB (Dubos et al., 2010).1R-MYB转录因子又称MYB-related转录因子, 包含1个完整或部分MYB结构域(Stracke et al., 2001; Chen et al., 2006).3R-MYB转录因子包含3个连续的MYB结构域(R1、R2和R3), 在大多数真核生物基因组中都发现了3R-MYB转录因子, 其在调控细胞周期中发挥作用(Haga et al., 2007).4R-MYB转录因子是最小的一类, 每个基因都含有4个R1/R2 MYB结构域, 它们在植物中发挥的作用尚不明确.包含2个MYB结构域(R2和R3)的R2R3-MYB转录因子是高等植物中MYB转录因子的主要存在形式, 该家族基因可能是因3R-MYB转录因子中R1 MYB结构域缺失进化而来(Rosinski and Atchley, 1998), 也有观点认为3R-MYB转录因子是从R2R3-MYB转录因子进化来的, R1 MYB结构域从R2R3-MYB转录因子获得(Jiang et al., 2004).通常在MYB转录因子的N末端检测到高度保守的MYB结构域, C末端MYB结构域是高度分化的激活结构域, 这也决定了MYB转录因子具有多种调节作用(Dias et al., 2003; Matus et al., 2008). ...
Improving plant drought, salt, and freezing tolerance by gene transfer of a single stress-inducible transcription factor 1 1999
... 转录因子在基因转录调节中起重要作用, 通过激活或抑制靶基因的转录, 调控植物的生长发育(Pabo and Sauer, 1992; Kasuga et al., 1999; Riechmann et al., 2000; Valliyodan and Nguyen, 2006).根据转录因子与其靶基因结合位点的相似性, 可将其分成不同的家族, 如AP2/EREBP、bHLH、HB、MYB和WRKY (Riechmann et al., 2000).MYB转录因子具有与DNA结合的特征性MYB结构域, 该结构域在植物中高度保守.MYB转录因子包含1-4个不完全重复的MYB结构域, 分别命名为R1 (Repeat1)、R2、R3和R4, 每个MYB结构域约有52个氨基酸残基, 形成3个α-螺旋, 其中第2和第3螺旋形成螺旋-转角-螺旋(helix-turn-helix)结构并与DNA大沟槽结合(Ogata et al., 1996; Jia et al., 2004; Dubos et al., 2010).根据包含的MYB结构域数目, MYB转录因子可分为4类, 即1R-MYB、2R-MYB (R2R3-MYB)、3R-MYB (R1R2R3- MYB)和4R-MYB (Dubos et al., 2010).1R-MYB转录因子又称MYB-related转录因子, 包含1个完整或部分MYB结构域(Stracke et al., 2001; Chen et al., 2006).3R-MYB转录因子包含3个连续的MYB结构域(R1、R2和R3), 在大多数真核生物基因组中都发现了3R-MYB转录因子, 其在调控细胞周期中发挥作用(Haga et al., 2007).4R-MYB转录因子是最小的一类, 每个基因都含有4个R1/R2 MYB结构域, 它们在植物中发挥的作用尚不明确.包含2个MYB结构域(R2和R3)的R2R3-MYB转录因子是高等植物中MYB转录因子的主要存在形式, 该家族基因可能是因3R-MYB转录因子中R1 MYB结构域缺失进化而来(Rosinski and Atchley, 1998), 也有观点认为3R-MYB转录因子是从R2R3-MYB转录因子进化来的, R1 MYB结构域从R2R3-MYB转录因子获得(Jiang et al., 2004).通常在MYB转录因子的N末端检测到高度保守的MYB结构域, C末端MYB结构域是高度分化的激活结构域, 这也决定了MYB转录因子具有多种调节作用(Dias et al., 2003; Matus et al., 2008). ...
New reference genome sequences of hot pepper reveal the massive evolution of plant disease-resistance genes by retroduplication 6 2017
... 一年生辣椒(Capsicum annuum L.) (ECW)、浆果状辣椒(Capsicum baccatum L.) (PBC81)和中国辣椒(Capsicum chinense Jacq.) (PI159236)的基因组数据从辣椒基因组平台(http://peppergenome.snu.ac.kr)中获得(Kim et al., 2017).从拟南芥(Arabidopsis thaliana L.)全基因组数据库(https://www.arabidopsis.org/)中下载拟南芥R2R3-MYB转录因子家族的氨基酸序列作为query序列, 分别与3种辣椒的蛋白序列进行BLASTP比对, 参数设置E-value≤1e-10, 每条query保留5条匹配结果, 其余参数为默认值.将得到的候选序列使用hmmer (http://www.hmmer.org/)比对到PFAM32.0数据库中, 基于Pfam的MYB模型(PF00249)进行进一步比对筛选, 参数为默认值, 从而确定辣椒的R2R3-MYB转录因子基因成员.使用WebLogo (Crooks et al., 2004)绘制R2R3-MYB结构域seqlogo图. ...
... 一年生辣椒、浆果状辣椒和中国辣椒的转录组测序数据从BioProject数据库(https://www.ncbi.nlm.nih.gov/ bioproject)中获得, 登录号分别为PRJNA223222、PRJNA308879和PRJNA331024.测序样品为生长6周植株的根、茎和叶组织和生长4个月植株盛开的花(Kim et al., 2017).使用fastp (Chen et al., 2018)对测序数据进行过滤, 参数设为默认值.使用RSEM (Li and Dewey, 2011)将过滤后的数据比对到对应物种的基因组序列上, 比对方式为bowtie2 (Langmead and Salzberg, 2012), 参数设为默认值.将比对结果进行标准化处理, 得到基因的每百万个转录片段中每千碱基片段的映射读数(fragments per kilobase of exon model per million mapped reads, FPKM).FPKM值经log10 (FPKM+1)转换后, 使用R包pheatmap (https://CRAN.R-project.org/package=pheatmap)绘制热图. ...
... 在植物进化过程中经常会出现基因重复, 这也是植物基因家族发生扩张的重要原因之一(Cannon et al., 2004).前人研究表明, 大约在170万年前浆果状辣椒和一年生辣椒、中国辣椒的祖先发生分化, 在110万年前一年生辣椒和中国辣椒也发生了分化(Kim et al., 2017).为了确定3种辣椒间R2R3-MYB同源基因以及每种辣椒内部可能出现的基因重复现象, 本研究鉴定了3种辣椒中R2R3-MYB基因的进化关系.最终在3种辣椒中鉴定出73组直系同源R2R3-MYB基因(图4), 分别占一年生辣椒R2R3-MYB基因的77.66% (73/ 94)、浆果状辣椒R2R3-MYB基因的79.35% (73/92)和中国辣椒R2R3-MYB基因的77.66% (73/94).这说明大多数R2R3-MYB基因在它们的祖先物种中存在, 并且在辣椒的进化过程中高度保守. ...
... 随着基因测序技术的不断发展, 已在基因组水平上对许多植物的R2R3-MYB转录因子家族进行了鉴定, 如拟南芥(125个) (Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(249个) (Hajiebrahimi et al., 2017)、大豆(244个) (Du et al., 2012)、番茄(121个) (Zhao et al., 2014)和马铃薯(112个) (Li et al., 2019; Sun et al., 2019), 它们的基因组大小分别为125 Mb (Zapata et al., 2016)、912 Mb (Chalhoub et al., 2014)、1.03 Gb (Schmutz et al., 2010)、828 Mb (The Tomato Genome Consortium, 2012)和663 Mb (The Potato Genome Sequencing Consortium, 2011).这表明物种中R2R3-MYB基因数目与物种基因组的大小并不成正比.一年生辣椒、浆果状辣椒和中国辣椒的基因组大小分别为2.85、2.99和2.80 Gb (Kim et al., 2017).本研究从中分别鉴定出94、92和94个R2R3-MYB基因, 也支持这一观点.与茄科植物番茄(Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019)相比, 3种辣椒中R2R3-MYB基因数目明显较少, 说明R2R3-MYB转录因子家族在辣椒中发生了明显收缩. ...
... 一年生辣椒、浆果状辣椒和中国辣椒的转录组测序数据从BioProject数据库(https://www.ncbi.nlm.nih.gov/ bioproject)中获得, 登录号分别为PRJNA223222、PRJNA308879和PRJNA331024.测序样品为生长6周植株的根、茎和叶组织和生长4个月植株盛开的花(Kim et al., 2017).使用fastp (Chen et al., 2018)对测序数据进行过滤, 参数设为默认值.使用RSEM (Li and Dewey, 2011)将过滤后的数据比对到对应物种的基因组序列上, 比对方式为bowtie2 (Langmead and Salzberg, 2012), 参数设为默认值.将比对结果进行标准化处理, 得到基因的每百万个转录片段中每千碱基片段的映射读数(fragments per kilobase of exon model per million mapped reads, FPKM).FPKM值经log10 (FPKM+1)转换后, 使用R包pheatmap (https://CRAN.R-project.org/package=pheatmap)绘制热图. ...
RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome 1 2011
... 一年生辣椒、浆果状辣椒和中国辣椒的转录组测序数据从BioProject数据库(https://www.ncbi.nlm.nih.gov/ bioproject)中获得, 登录号分别为PRJNA223222、PRJNA308879和PRJNA331024.测序样品为生长6周植株的根、茎和叶组织和生长4个月植株盛开的花(Kim et al., 2017).使用fastp (Chen et al., 2018)对测序数据进行过滤, 参数设为默认值.使用RSEM (Li and Dewey, 2011)将过滤后的数据比对到对应物种的基因组序列上, 比对方式为bowtie2 (Langmead and Salzberg, 2012), 参数设为默认值.将比对结果进行标准化处理, 得到基因的每百万个转录片段中每千碱基片段的映射读数(fragments per kilobase of exon model per million mapped reads, FPKM).FPKM值经log10 (FPKM+1)转换后, 使用R包pheatmap (https://CRAN.R-project.org/package=pheatmap)绘制热图. ...
Systematic analysis of MYB family genes in potato and their multiple roles in development and stress responses 5 2019
... 随着测序技术的发展, R2R3-MYB转录因子家族先后在许多高等植物的全基因组水平得到系统鉴定和分析, 包括拟南芥(Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(Brassica napus) (Hajiebrahimi et al., 2017)、大豆(Glycine max) (Du et al., 2012)、番茄(Solanum lycopersicum) (Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019).基因组测序技术促进了园艺植物基因组学和遗传学研究(唐嘉瓅等, 2020).辣椒是茄科(Solanaceae)辣椒属(Capsicum)一年生或多年生草本植物, 是具有重要经济价值的蔬菜作物, 在世界范围内广泛种植.辣椒属包含27种, 被人类驯化长期栽培的辣椒有5种, 分别为一年生辣椒(Capsicum annuum)、浆果状辣椒(C. baccatum)、中国辣椒(C. chinense)、灌木状辣椒(C. frutescens)和绒毛辣椒(C. pubescens) (Ibiza et al., 2012).我国主要栽培品种是一年生辣椒, 其它4个栽培种具有许多优良性状, 是对一年生辣椒进行品种改良的重要种质资源(隋益虎和陈劲枫, 2009; 魏家香等, 2018).本研究以一年生辣椒、浆果状辣椒和中国辣椒为材料, 对3种辣椒R2R3-MYB转录因子家族在全基因组范围内进行鉴定, 在此基础上分析了辣椒R2R3-MYB基因的序列、motif分布、外显子-内含子结构、染色体定位、比较进化和不同组织(根、茎、叶和花)中的表达情况.通过共线性分析, 发现3种辣椒都存在特有的R2R3-MYB基因, 一年生辣椒和浆果状辣椒中的R2R3-MYB转录因子家族发生了基因组重复事件.研究结果为揭示辣椒R2R3-MYB转录因子功能及辣椒分子遗传育种奠定了理论基础. ...
... 为了研究辣椒R2R3-MYB基因的进化关系, 利用3种辣椒和拟南芥R2R3-MYB转录因子的蛋白序列构建系统进化树.与拟南芥R2R3-MYB转录因子聚集到同一分支的辣椒R2R3-MYB转录因子被认定为同一亚族, 可分为30个亚家族(S1-S30) (图2).分类中的大多数亚家族与其它植物中的亚家族一致, 但是辣椒R2R3-MYB基因家族未发现有成员属于亚家族S10、S12和S15, 暗示辣椒基因组在长期进化过程中可能丢失了一些R2R3-MYB基因(图2, 图3A).辣椒的近源物种马铃薯R2R3-MYB转录因子家族中不存在亚家族S10和S12也证明了这一点(Li et al., 2019; Sun et al., 2019). ...
... 随着基因测序技术的不断发展, 已在基因组水平上对许多植物的R2R3-MYB转录因子家族进行了鉴定, 如拟南芥(125个) (Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(249个) (Hajiebrahimi et al., 2017)、大豆(244个) (Du et al., 2012)、番茄(121个) (Zhao et al., 2014)和马铃薯(112个) (Li et al., 2019; Sun et al., 2019), 它们的基因组大小分别为125 Mb (Zapata et al., 2016)、912 Mb (Chalhoub et al., 2014)、1.03 Gb (Schmutz et al., 2010)、828 Mb (The Tomato Genome Consortium, 2012)和663 Mb (The Potato Genome Sequencing Consortium, 2011).这表明物种中R2R3-MYB基因数目与物种基因组的大小并不成正比.一年生辣椒、浆果状辣椒和中国辣椒的基因组大小分别为2.85、2.99和2.80 Gb (Kim et al., 2017).本研究从中分别鉴定出94、92和94个R2R3-MYB基因, 也支持这一观点.与茄科植物番茄(Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019)相比, 3种辣椒中R2R3-MYB基因数目明显较少, 说明R2R3-MYB转录因子家族在辣椒中发生了明显收缩. ...
... )和马铃薯(Li et al., 2019; Sun et al., 2019)相比, 3种辣椒中R2R3-MYB基因数目明显较少, 说明R2R3-MYB转录因子家族在辣椒中发生了明显收缩. ...
... 基于3种辣椒和拟南芥R2R3-MYB基因序列的相似性, 可将其划分为30个不同的亚家族(图2).与马铃薯(Li et al., 2019; Sun et al., 2019)类似, 辣椒R2R3-MYB转录因子家族中存在部分亚家族成员丢失, 也获得了一些新的亚家族成员, 表明辣椒R2R3- MYB转录因子家族也有扩张.一年生辣椒、浆果状辣椒和中国辣椒中R2R3-MYB基因的结构分析(图3)显示, 它们主要由2个内含子、3个外显子和1个内含子、2个外显子这两类基因结构组成, 相同亚家族成员也普遍具有相同的基因结构.辣椒R2R3-MYB转录因子中包含2个MYB结构域与所有转录因子共有的motif 相对应, 位于同一亚家族的R2R3-MYB基因motif也都相同, 这说明R2R3-MYB基因的亚家族分类主要由C端序列决定. ...
StMYB44 negatively regulates anthocyanin biosynthesis at high temperatures in tuber flesh of potato 1 2019
... MYB转录因子广泛分布于植物基因组中, 是植物中最大的转录因子家族之一(Riechmann et al., 2000; Dubos et al., 2010).首个植物MYB转录因子是从玉米(Zea mays)中鉴定出的COLORED1 (Paz-Ares et al., 1987).目前对MYB转录因子研究不断深入, R2R3-MYB转录因子已成为研究重点之一.据报道, R2R3-MYB转录因子参与调控许多重要的生理生化过程, 包括调节植物的初级和次生代谢、控制生长发育以及参与响应各种生物和非生物胁迫(Stracke et al., 2007).例如, 马铃薯(Solanum tuberosum) StMYB44在高温下负调控花青素的生物合成(Liu et al., 2019); 金银花(Lonicera japonica) LjaMYB12在拟南芥(Arabidopsis thaliana)中异位表达可以提高类黄酮含量, 其对类黄酮的生物合成的上下游途径具有不同的调控模式(Qi et al., 2019); 拟南芥AtMYB20、AtMYB42、AtMYB43和AtMYB85能够特异性地抑制类黄酮的生物合成(Geng et al., 2020); 棉花(Gossypium hirsutum) GhMYB108-like在干旱和盐胁迫下发挥重要作用(Ullah et al., 2020); 苹果(Malus domestica) MdMYB24-like参与茉莉酸甲酯诱导的花青素苷生物合成(Wang et al., 2019), MdMYB30在苹果角质层蜡质的积累中起重要作用, 并能增强苹果的抗病性(Zhang et al., 2019); 芝麻(Sesamum indicum) SiMYB75在拟南芥中的异位过表达显著促进根系生长, 提高植株对干旱、盐胁迫和渗透胁迫的耐受性(Dossa et al., 2020); 盐、干旱及脱落酸胁迫促进甘薯(Ipomoea batatas) IbMYB3的表达, 暗示IbMYB3在甘薯非生物胁迫应答中起重要作用(李格等, 2020). ...
Analysis of relative gene expression data using real-time quantitative PCR and the 2 ?ΔΔCT method 1 2001
Analysis of the grape MYB R2R3 subfamily reveals expanded wine quality-related clades and conserved gene structure organization across Vitis and Arabidopsis genomes 1 2008
... 转录因子在基因转录调节中起重要作用, 通过激活或抑制靶基因的转录, 调控植物的生长发育(Pabo and Sauer, 1992; Kasuga et al., 1999; Riechmann et al., 2000; Valliyodan and Nguyen, 2006).根据转录因子与其靶基因结合位点的相似性, 可将其分成不同的家族, 如AP2/EREBP、bHLH、HB、MYB和WRKY (Riechmann et al., 2000).MYB转录因子具有与DNA结合的特征性MYB结构域, 该结构域在植物中高度保守.MYB转录因子包含1-4个不完全重复的MYB结构域, 分别命名为R1 (Repeat1)、R2、R3和R4, 每个MYB结构域约有52个氨基酸残基, 形成3个α-螺旋, 其中第2和第3螺旋形成螺旋-转角-螺旋(helix-turn-helix)结构并与DNA大沟槽结合(Ogata et al., 1996; Jia et al., 2004; Dubos et al., 2010).根据包含的MYB结构域数目, MYB转录因子可分为4类, 即1R-MYB、2R-MYB (R2R3-MYB)、3R-MYB (R1R2R3- MYB)和4R-MYB (Dubos et al., 2010).1R-MYB转录因子又称MYB-related转录因子, 包含1个完整或部分MYB结构域(Stracke et al., 2001; Chen et al., 2006).3R-MYB转录因子包含3个连续的MYB结构域(R1、R2和R3), 在大多数真核生物基因组中都发现了3R-MYB转录因子, 其在调控细胞周期中发挥作用(Haga et al., 2007).4R-MYB转录因子是最小的一类, 每个基因都含有4个R1/R2 MYB结构域, 它们在植物中发挥的作用尚不明确.包含2个MYB结构域(R2和R3)的R2R3-MYB转录因子是高等植物中MYB转录因子的主要存在形式, 该家族基因可能是因3R-MYB转录因子中R1 MYB结构域缺失进化而来(Rosinski and Atchley, 1998), 也有观点认为3R-MYB转录因子是从R2R3-MYB转录因子进化来的, R1 MYB结构域从R2R3-MYB转录因子获得(Jiang et al., 2004).通常在MYB转录因子的N末端检测到高度保守的MYB结构域, C末端MYB结构域是高度分化的激活结构域, 这也决定了MYB转录因子具有多种调节作用(Dias et al., 2003; Matus et al., 2008). ...
The cavity in the hydrophobic core of Myb DNA- binding domain is reserved for DNA recognition and trans- activation 1 1996
... 转录因子在基因转录调节中起重要作用, 通过激活或抑制靶基因的转录, 调控植物的生长发育(Pabo and Sauer, 1992; Kasuga et al., 1999; Riechmann et al., 2000; Valliyodan and Nguyen, 2006).根据转录因子与其靶基因结合位点的相似性, 可将其分成不同的家族, 如AP2/EREBP、bHLH、HB、MYB和WRKY (Riechmann et al., 2000).MYB转录因子具有与DNA结合的特征性MYB结构域, 该结构域在植物中高度保守.MYB转录因子包含1-4个不完全重复的MYB结构域, 分别命名为R1 (Repeat1)、R2、R3和R4, 每个MYB结构域约有52个氨基酸残基, 形成3个α-螺旋, 其中第2和第3螺旋形成螺旋-转角-螺旋(helix-turn-helix)结构并与DNA大沟槽结合(Ogata et al., 1996; Jia et al., 2004; Dubos et al., 2010).根据包含的MYB结构域数目, MYB转录因子可分为4类, 即1R-MYB、2R-MYB (R2R3-MYB)、3R-MYB (R1R2R3- MYB)和4R-MYB (Dubos et al., 2010).1R-MYB转录因子又称MYB-related转录因子, 包含1个完整或部分MYB结构域(Stracke et al., 2001; Chen et al., 2006).3R-MYB转录因子包含3个连续的MYB结构域(R1、R2和R3), 在大多数真核生物基因组中都发现了3R-MYB转录因子, 其在调控细胞周期中发挥作用(Haga et al., 2007).4R-MYB转录因子是最小的一类, 每个基因都含有4个R1/R2 MYB结构域, 它们在植物中发挥的作用尚不明确.包含2个MYB结构域(R2和R3)的R2R3-MYB转录因子是高等植物中MYB转录因子的主要存在形式, 该家族基因可能是因3R-MYB转录因子中R1 MYB结构域缺失进化而来(Rosinski and Atchley, 1998), 也有观点认为3R-MYB转录因子是从R2R3-MYB转录因子进化来的, R1 MYB结构域从R2R3-MYB转录因子获得(Jiang et al., 2004).通常在MYB转录因子的N末端检测到高度保守的MYB结构域, C末端MYB结构域是高度分化的激活结构域, 这也决定了MYB转录因子具有多种调节作用(Dias et al., 2003; Matus et al., 2008). ...
Transcription factors: structural families and principles of DNA recognition 1 1992
... 转录因子在基因转录调节中起重要作用, 通过激活或抑制靶基因的转录, 调控植物的生长发育(Pabo and Sauer, 1992; Kasuga et al., 1999; Riechmann et al., 2000; Valliyodan and Nguyen, 2006).根据转录因子与其靶基因结合位点的相似性, 可将其分成不同的家族, 如AP2/EREBP、bHLH、HB、MYB和WRKY (Riechmann et al., 2000).MYB转录因子具有与DNA结合的特征性MYB结构域, 该结构域在植物中高度保守.MYB转录因子包含1-4个不完全重复的MYB结构域, 分别命名为R1 (Repeat1)、R2、R3和R4, 每个MYB结构域约有52个氨基酸残基, 形成3个α-螺旋, 其中第2和第3螺旋形成螺旋-转角-螺旋(helix-turn-helix)结构并与DNA大沟槽结合(Ogata et al., 1996; Jia et al., 2004; Dubos et al., 2010).根据包含的MYB结构域数目, MYB转录因子可分为4类, 即1R-MYB、2R-MYB (R2R3-MYB)、3R-MYB (R1R2R3- MYB)和4R-MYB (Dubos et al., 2010).1R-MYB转录因子又称MYB-related转录因子, 包含1个完整或部分MYB结构域(Stracke et al., 2001; Chen et al., 2006).3R-MYB转录因子包含3个连续的MYB结构域(R1、R2和R3), 在大多数真核生物基因组中都发现了3R-MYB转录因子, 其在调控细胞周期中发挥作用(Haga et al., 2007).4R-MYB转录因子是最小的一类, 每个基因都含有4个R1/R2 MYB结构域, 它们在植物中发挥的作用尚不明确.包含2个MYB结构域(R2和R3)的R2R3-MYB转录因子是高等植物中MYB转录因子的主要存在形式, 该家族基因可能是因3R-MYB转录因子中R1 MYB结构域缺失进化而来(Rosinski and Atchley, 1998), 也有观点认为3R-MYB转录因子是从R2R3-MYB转录因子进化来的, R1 MYB结构域从R2R3-MYB转录因子获得(Jiang et al., 2004).通常在MYB转录因子的N末端检测到高度保守的MYB结构域, C末端MYB结构域是高度分化的激活结构域, 这也决定了MYB转录因子具有多种调节作用(Dias et al., 2003; Matus et al., 2008). ...
The regulatory c1 locus of Zea mays encodes a protein with homology to myb proto-oncogene products and with structural similarities to transcriptional activators 1 1987
... MYB转录因子广泛分布于植物基因组中, 是植物中最大的转录因子家族之一(Riechmann et al., 2000; Dubos et al., 2010).首个植物MYB转录因子是从玉米(Zea mays)中鉴定出的COLORED1 (Paz-Ares et al., 1987).目前对MYB转录因子研究不断深入, R2R3-MYB转录因子已成为研究重点之一.据报道, R2R3-MYB转录因子参与调控许多重要的生理生化过程, 包括调节植物的初级和次生代谢、控制生长发育以及参与响应各种生物和非生物胁迫(Stracke et al., 2007).例如, 马铃薯(Solanum tuberosum) StMYB44在高温下负调控花青素的生物合成(Liu et al., 2019); 金银花(Lonicera japonica) LjaMYB12在拟南芥(Arabidopsis thaliana)中异位表达可以提高类黄酮含量, 其对类黄酮的生物合成的上下游途径具有不同的调控模式(Qi et al., 2019); 拟南芥AtMYB20、AtMYB42、AtMYB43和AtMYB85能够特异性地抑制类黄酮的生物合成(Geng et al., 2020); 棉花(Gossypium hirsutum) GhMYB108-like在干旱和盐胁迫下发挥重要作用(Ullah et al., 2020); 苹果(Malus domestica) MdMYB24-like参与茉莉酸甲酯诱导的花青素苷生物合成(Wang et al., 2019), MdMYB30在苹果角质层蜡质的积累中起重要作用, 并能增强苹果的抗病性(Zhang et al., 2019); 芝麻(Sesamum indicum) SiMYB75在拟南芥中的异位过表达显著促进根系生长, 提高植株对干旱、盐胁迫和渗透胁迫的耐受性(Dossa et al., 2020); 盐、干旱及脱落酸胁迫促进甘薯(Ipomoea batatas) IbMYB3的表达, 暗示IbMYB3在甘薯非生物胁迫应答中起重要作用(李格等, 2020). ...
Ectopic expression of a R2R3-MYB transcription factor gene LjaMYB12 from Lonicera japonica increases flavonoid accumulation in Arabidopsis thaliana 1 2019
... MYB转录因子广泛分布于植物基因组中, 是植物中最大的转录因子家族之一(Riechmann et al., 2000; Dubos et al., 2010).首个植物MYB转录因子是从玉米(Zea mays)中鉴定出的COLORED1 (Paz-Ares et al., 1987).目前对MYB转录因子研究不断深入, R2R3-MYB转录因子已成为研究重点之一.据报道, R2R3-MYB转录因子参与调控许多重要的生理生化过程, 包括调节植物的初级和次生代谢、控制生长发育以及参与响应各种生物和非生物胁迫(Stracke et al., 2007).例如, 马铃薯(Solanum tuberosum) StMYB44在高温下负调控花青素的生物合成(Liu et al., 2019); 金银花(Lonicera japonica) LjaMYB12在拟南芥(Arabidopsis thaliana)中异位表达可以提高类黄酮含量, 其对类黄酮的生物合成的上下游途径具有不同的调控模式(Qi et al., 2019); 拟南芥AtMYB20、AtMYB42、AtMYB43和AtMYB85能够特异性地抑制类黄酮的生物合成(Geng et al., 2020); 棉花(Gossypium hirsutum) GhMYB108-like在干旱和盐胁迫下发挥重要作用(Ullah et al., 2020); 苹果(Malus domestica) MdMYB24-like参与茉莉酸甲酯诱导的花青素苷生物合成(Wang et al., 2019), MdMYB30在苹果角质层蜡质的积累中起重要作用, 并能增强苹果的抗病性(Zhang et al., 2019); 芝麻(Sesamum indicum) SiMYB75在拟南芥中的异位过表达显著促进根系生长, 提高植株对干旱、盐胁迫和渗透胁迫的耐受性(Dossa et al., 2020); 盐、干旱及脱落酸胁迫促进甘薯(Ipomoea batatas) IbMYB3的表达, 暗示IbMYB3在甘薯非生物胁迫应答中起重要作用(李格等, 2020). ...
Arabidopsis transcription factors: genome-wide comparative analysis among eukaryotes 3 2000
... 转录因子在基因转录调节中起重要作用, 通过激活或抑制靶基因的转录, 调控植物的生长发育(Pabo and Sauer, 1992; Kasuga et al., 1999; Riechmann et al., 2000; Valliyodan and Nguyen, 2006).根据转录因子与其靶基因结合位点的相似性, 可将其分成不同的家族, 如AP2/EREBP、bHLH、HB、MYB和WRKY (Riechmann et al., 2000).MYB转录因子具有与DNA结合的特征性MYB结构域, 该结构域在植物中高度保守.MYB转录因子包含1-4个不完全重复的MYB结构域, 分别命名为R1 (Repeat1)、R2、R3和R4, 每个MYB结构域约有52个氨基酸残基, 形成3个α-螺旋, 其中第2和第3螺旋形成螺旋-转角-螺旋(helix-turn-helix)结构并与DNA大沟槽结合(Ogata et al., 1996; Jia et al., 2004; Dubos et al., 2010).根据包含的MYB结构域数目, MYB转录因子可分为4类, 即1R-MYB、2R-MYB (R2R3-MYB)、3R-MYB (R1R2R3- MYB)和4R-MYB (Dubos et al., 2010).1R-MYB转录因子又称MYB-related转录因子, 包含1个完整或部分MYB结构域(Stracke et al., 2001; Chen et al., 2006).3R-MYB转录因子包含3个连续的MYB结构域(R1、R2和R3), 在大多数真核生物基因组中都发现了3R-MYB转录因子, 其在调控细胞周期中发挥作用(Haga et al., 2007).4R-MYB转录因子是最小的一类, 每个基因都含有4个R1/R2 MYB结构域, 它们在植物中发挥的作用尚不明确.包含2个MYB结构域(R2和R3)的R2R3-MYB转录因子是高等植物中MYB转录因子的主要存在形式, 该家族基因可能是因3R-MYB转录因子中R1 MYB结构域缺失进化而来(Rosinski and Atchley, 1998), 也有观点认为3R-MYB转录因子是从R2R3-MYB转录因子进化来的, R1 MYB结构域从R2R3-MYB转录因子获得(Jiang et al., 2004).通常在MYB转录因子的N末端检测到高度保守的MYB结构域, C末端MYB结构域是高度分化的激活结构域, 这也决定了MYB转录因子具有多种调节作用(Dias et al., 2003; Matus et al., 2008). ...
... ).根据转录因子与其靶基因结合位点的相似性, 可将其分成不同的家族, 如AP2/EREBP、bHLH、HB、MYB和WRKY (Riechmann et al., 2000).MYB转录因子具有与DNA结合的特征性MYB结构域, 该结构域在植物中高度保守.MYB转录因子包含1-4个不完全重复的MYB结构域, 分别命名为R1 (Repeat1)、R2、R3和R4, 每个MYB结构域约有52个氨基酸残基, 形成3个α-螺旋, 其中第2和第3螺旋形成螺旋-转角-螺旋(helix-turn-helix)结构并与DNA大沟槽结合(Ogata et al., 1996; Jia et al., 2004; Dubos et al., 2010).根据包含的MYB结构域数目, MYB转录因子可分为4类, 即1R-MYB、2R-MYB (R2R3-MYB)、3R-MYB (R1R2R3- MYB)和4R-MYB (Dubos et al., 2010).1R-MYB转录因子又称MYB-related转录因子, 包含1个完整或部分MYB结构域(Stracke et al., 2001; Chen et al., 2006).3R-MYB转录因子包含3个连续的MYB结构域(R1、R2和R3), 在大多数真核生物基因组中都发现了3R-MYB转录因子, 其在调控细胞周期中发挥作用(Haga et al., 2007).4R-MYB转录因子是最小的一类, 每个基因都含有4个R1/R2 MYB结构域, 它们在植物中发挥的作用尚不明确.包含2个MYB结构域(R2和R3)的R2R3-MYB转录因子是高等植物中MYB转录因子的主要存在形式, 该家族基因可能是因3R-MYB转录因子中R1 MYB结构域缺失进化而来(Rosinski and Atchley, 1998), 也有观点认为3R-MYB转录因子是从R2R3-MYB转录因子进化来的, R1 MYB结构域从R2R3-MYB转录因子获得(Jiang et al., 2004).通常在MYB转录因子的N末端检测到高度保守的MYB结构域, C末端MYB结构域是高度分化的激活结构域, 这也决定了MYB转录因子具有多种调节作用(Dias et al., 2003; Matus et al., 2008). ...
... MYB转录因子广泛分布于植物基因组中, 是植物中最大的转录因子家族之一(Riechmann et al., 2000; Dubos et al., 2010).首个植物MYB转录因子是从玉米(Zea mays)中鉴定出的COLORED1 (Paz-Ares et al., 1987).目前对MYB转录因子研究不断深入, R2R3-MYB转录因子已成为研究重点之一.据报道, R2R3-MYB转录因子参与调控许多重要的生理生化过程, 包括调节植物的初级和次生代谢、控制生长发育以及参与响应各种生物和非生物胁迫(Stracke et al., 2007).例如, 马铃薯(Solanum tuberosum) StMYB44在高温下负调控花青素的生物合成(Liu et al., 2019); 金银花(Lonicera japonica) LjaMYB12在拟南芥(Arabidopsis thaliana)中异位表达可以提高类黄酮含量, 其对类黄酮的生物合成的上下游途径具有不同的调控模式(Qi et al., 2019); 拟南芥AtMYB20、AtMYB42、AtMYB43和AtMYB85能够特异性地抑制类黄酮的生物合成(Geng et al., 2020); 棉花(Gossypium hirsutum) GhMYB108-like在干旱和盐胁迫下发挥重要作用(Ullah et al., 2020); 苹果(Malus domestica) MdMYB24-like参与茉莉酸甲酯诱导的花青素苷生物合成(Wang et al., 2019), MdMYB30在苹果角质层蜡质的积累中起重要作用, 并能增强苹果的抗病性(Zhang et al., 2019); 芝麻(Sesamum indicum) SiMYB75在拟南芥中的异位过表达显著促进根系生长, 提高植株对干旱、盐胁迫和渗透胁迫的耐受性(Dossa et al., 2020); 盐、干旱及脱落酸胁迫促进甘薯(Ipomoea batatas) IbMYB3的表达, 暗示IbMYB3在甘薯非生物胁迫应答中起重要作用(李格等, 2020). ...
Molecular evolution of the MYB family of transcription factors: evidence for polyphyletic origin 1 1998
... 转录因子在基因转录调节中起重要作用, 通过激活或抑制靶基因的转录, 调控植物的生长发育(Pabo and Sauer, 1992; Kasuga et al., 1999; Riechmann et al., 2000; Valliyodan and Nguyen, 2006).根据转录因子与其靶基因结合位点的相似性, 可将其分成不同的家族, 如AP2/EREBP、bHLH、HB、MYB和WRKY (Riechmann et al., 2000).MYB转录因子具有与DNA结合的特征性MYB结构域, 该结构域在植物中高度保守.MYB转录因子包含1-4个不完全重复的MYB结构域, 分别命名为R1 (Repeat1)、R2、R3和R4, 每个MYB结构域约有52个氨基酸残基, 形成3个α-螺旋, 其中第2和第3螺旋形成螺旋-转角-螺旋(helix-turn-helix)结构并与DNA大沟槽结合(Ogata et al., 1996; Jia et al., 2004; Dubos et al., 2010).根据包含的MYB结构域数目, MYB转录因子可分为4类, 即1R-MYB、2R-MYB (R2R3-MYB)、3R-MYB (R1R2R3- MYB)和4R-MYB (Dubos et al., 2010).1R-MYB转录因子又称MYB-related转录因子, 包含1个完整或部分MYB结构域(Stracke et al., 2001; Chen et al., 2006).3R-MYB转录因子包含3个连续的MYB结构域(R1、R2和R3), 在大多数真核生物基因组中都发现了3R-MYB转录因子, 其在调控细胞周期中发挥作用(Haga et al., 2007).4R-MYB转录因子是最小的一类, 每个基因都含有4个R1/R2 MYB结构域, 它们在植物中发挥的作用尚不明确.包含2个MYB结构域(R2和R3)的R2R3-MYB转录因子是高等植物中MYB转录因子的主要存在形式, 该家族基因可能是因3R-MYB转录因子中R1 MYB结构域缺失进化而来(Rosinski and Atchley, 1998), 也有观点认为3R-MYB转录因子是从R2R3-MYB转录因子进化来的, R1 MYB结构域从R2R3-MYB转录因子获得(Jiang et al., 2004).通常在MYB转录因子的N末端检测到高度保守的MYB结构域, C末端MYB结构域是高度分化的激活结构域, 这也决定了MYB转录因子具有多种调节作用(Dias et al., 2003; Matus et al., 2008). ...
Genome sequence of the palaeopolyploid soybean 1 2010
... 随着基因测序技术的不断发展, 已在基因组水平上对许多植物的R2R3-MYB转录因子家族进行了鉴定, 如拟南芥(125个) (Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(249个) (Hajiebrahimi et al., 2017)、大豆(244个) (Du et al., 2012)、番茄(121个) (Zhao et al., 2014)和马铃薯(112个) (Li et al., 2019; Sun et al., 2019), 它们的基因组大小分别为125 Mb (Zapata et al., 2016)、912 Mb (Chalhoub et al., 2014)、1.03 Gb (Schmutz et al., 2010)、828 Mb (The Tomato Genome Consortium, 2012)和663 Mb (The Potato Genome Sequencing Consortium, 2011).这表明物种中R2R3-MYB基因数目与物种基因组的大小并不成正比.一年生辣椒、浆果状辣椒和中国辣椒的基因组大小分别为2.85、2.99和2.80 Gb (Kim et al., 2017).本研究从中分别鉴定出94、92和94个R2R3-MYB基因, 也支持这一观点.与茄科植物番茄(Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019)相比, 3种辣椒中R2R3-MYB基因数目明显较少, 说明R2R3-MYB转录因子家族在辣椒中发生了明显收缩. ...
Differential regulation of closely related R2R3-MYB transcription factors controls flavonol accumulation in different parts of the Arabidopsis thaliana seedling 1 2007
... MYB转录因子广泛分布于植物基因组中, 是植物中最大的转录因子家族之一(Riechmann et al., 2000; Dubos et al., 2010).首个植物MYB转录因子是从玉米(Zea mays)中鉴定出的COLORED1 (Paz-Ares et al., 1987).目前对MYB转录因子研究不断深入, R2R3-MYB转录因子已成为研究重点之一.据报道, R2R3-MYB转录因子参与调控许多重要的生理生化过程, 包括调节植物的初级和次生代谢、控制生长发育以及参与响应各种生物和非生物胁迫(Stracke et al., 2007).例如, 马铃薯(Solanum tuberosum) StMYB44在高温下负调控花青素的生物合成(Liu et al., 2019); 金银花(Lonicera japonica) LjaMYB12在拟南芥(Arabidopsis thaliana)中异位表达可以提高类黄酮含量, 其对类黄酮的生物合成的上下游途径具有不同的调控模式(Qi et al., 2019); 拟南芥AtMYB20、AtMYB42、AtMYB43和AtMYB85能够特异性地抑制类黄酮的生物合成(Geng et al., 2020); 棉花(Gossypium hirsutum) GhMYB108-like在干旱和盐胁迫下发挥重要作用(Ullah et al., 2020); 苹果(Malus domestica) MdMYB24-like参与茉莉酸甲酯诱导的花青素苷生物合成(Wang et al., 2019), MdMYB30在苹果角质层蜡质的积累中起重要作用, 并能增强苹果的抗病性(Zhang et al., 2019); 芝麻(Sesamum indicum) SiMYB75在拟南芥中的异位过表达显著促进根系生长, 提高植株对干旱、盐胁迫和渗透胁迫的耐受性(Dossa et al., 2020); 盐、干旱及脱落酸胁迫促进甘薯(Ipomoea batatas) IbMYB3的表达, 暗示IbMYB3在甘薯非生物胁迫应答中起重要作用(李格等, 2020). ...
The R2R3- MYB gene family in Arabidopsis thaliana 3 2001
... 转录因子在基因转录调节中起重要作用, 通过激活或抑制靶基因的转录, 调控植物的生长发育(Pabo and Sauer, 1992; Kasuga et al., 1999; Riechmann et al., 2000; Valliyodan and Nguyen, 2006).根据转录因子与其靶基因结合位点的相似性, 可将其分成不同的家族, 如AP2/EREBP、bHLH、HB、MYB和WRKY (Riechmann et al., 2000).MYB转录因子具有与DNA结合的特征性MYB结构域, 该结构域在植物中高度保守.MYB转录因子包含1-4个不完全重复的MYB结构域, 分别命名为R1 (Repeat1)、R2、R3和R4, 每个MYB结构域约有52个氨基酸残基, 形成3个α-螺旋, 其中第2和第3螺旋形成螺旋-转角-螺旋(helix-turn-helix)结构并与DNA大沟槽结合(Ogata et al., 1996; Jia et al., 2004; Dubos et al., 2010).根据包含的MYB结构域数目, MYB转录因子可分为4类, 即1R-MYB、2R-MYB (R2R3-MYB)、3R-MYB (R1R2R3- MYB)和4R-MYB (Dubos et al., 2010).1R-MYB转录因子又称MYB-related转录因子, 包含1个完整或部分MYB结构域(Stracke et al., 2001; Chen et al., 2006).3R-MYB转录因子包含3个连续的MYB结构域(R1、R2和R3), 在大多数真核生物基因组中都发现了3R-MYB转录因子, 其在调控细胞周期中发挥作用(Haga et al., 2007).4R-MYB转录因子是最小的一类, 每个基因都含有4个R1/R2 MYB结构域, 它们在植物中发挥的作用尚不明确.包含2个MYB结构域(R2和R3)的R2R3-MYB转录因子是高等植物中MYB转录因子的主要存在形式, 该家族基因可能是因3R-MYB转录因子中R1 MYB结构域缺失进化而来(Rosinski and Atchley, 1998), 也有观点认为3R-MYB转录因子是从R2R3-MYB转录因子进化来的, R1 MYB结构域从R2R3-MYB转录因子获得(Jiang et al., 2004).通常在MYB转录因子的N末端检测到高度保守的MYB结构域, C末端MYB结构域是高度分化的激活结构域, 这也决定了MYB转录因子具有多种调节作用(Dias et al., 2003; Matus et al., 2008). ...
... 随着测序技术的发展, R2R3-MYB转录因子家族先后在许多高等植物的全基因组水平得到系统鉴定和分析, 包括拟南芥(Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(Brassica napus) (Hajiebrahimi et al., 2017)、大豆(Glycine max) (Du et al., 2012)、番茄(Solanum lycopersicum) (Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019).基因组测序技术促进了园艺植物基因组学和遗传学研究(唐嘉瓅等, 2020).辣椒是茄科(Solanaceae)辣椒属(Capsicum)一年生或多年生草本植物, 是具有重要经济价值的蔬菜作物, 在世界范围内广泛种植.辣椒属包含27种, 被人类驯化长期栽培的辣椒有5种, 分别为一年生辣椒(Capsicum annuum)、浆果状辣椒(C. baccatum)、中国辣椒(C. chinense)、灌木状辣椒(C. frutescens)和绒毛辣椒(C. pubescens) (Ibiza et al., 2012).我国主要栽培品种是一年生辣椒, 其它4个栽培种具有许多优良性状, 是对一年生辣椒进行品种改良的重要种质资源(隋益虎和陈劲枫, 2009; 魏家香等, 2018).本研究以一年生辣椒、浆果状辣椒和中国辣椒为材料, 对3种辣椒R2R3-MYB转录因子家族在全基因组范围内进行鉴定, 在此基础上分析了辣椒R2R3-MYB基因的序列、motif分布、外显子-内含子结构、染色体定位、比较进化和不同组织(根、茎、叶和花)中的表达情况.通过共线性分析, 发现3种辣椒都存在特有的R2R3-MYB基因, 一年生辣椒和浆果状辣椒中的R2R3-MYB转录因子家族发生了基因组重复事件.研究结果为揭示辣椒R2R3-MYB转录因子功能及辣椒分子遗传育种奠定了理论基础. ...
... 随着基因测序技术的不断发展, 已在基因组水平上对许多植物的R2R3-MYB转录因子家族进行了鉴定, 如拟南芥(125个) (Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(249个) (Hajiebrahimi et al., 2017)、大豆(244个) (Du et al., 2012)、番茄(121个) (Zhao et al., 2014)和马铃薯(112个) (Li et al., 2019; Sun et al., 2019), 它们的基因组大小分别为125 Mb (Zapata et al., 2016)、912 Mb (Chalhoub et al., 2014)、1.03 Gb (Schmutz et al., 2010)、828 Mb (The Tomato Genome Consortium, 2012)和663 Mb (The Potato Genome Sequencing Consortium, 2011).这表明物种中R2R3-MYB基因数目与物种基因组的大小并不成正比.一年生辣椒、浆果状辣椒和中国辣椒的基因组大小分别为2.85、2.99和2.80 Gb (Kim et al., 2017).本研究从中分别鉴定出94、92和94个R2R3-MYB基因, 也支持这一观点.与茄科植物番茄(Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019)相比, 3种辣椒中R2R3-MYB基因数目明显较少, 说明R2R3-MYB转录因子家族在辣椒中发生了明显收缩. ...
MYB gene family in potato ( Solanum tuberosum L.): genome-wide identification of hormone-responsive reveals their potential functions in growth and development 5 2019
... 随着测序技术的发展, R2R3-MYB转录因子家族先后在许多高等植物的全基因组水平得到系统鉴定和分析, 包括拟南芥(Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(Brassica napus) (Hajiebrahimi et al., 2017)、大豆(Glycine max) (Du et al., 2012)、番茄(Solanum lycopersicum) (Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019).基因组测序技术促进了园艺植物基因组学和遗传学研究(唐嘉瓅等, 2020).辣椒是茄科(Solanaceae)辣椒属(Capsicum)一年生或多年生草本植物, 是具有重要经济价值的蔬菜作物, 在世界范围内广泛种植.辣椒属包含27种, 被人类驯化长期栽培的辣椒有5种, 分别为一年生辣椒(Capsicum annuum)、浆果状辣椒(C. baccatum)、中国辣椒(C. chinense)、灌木状辣椒(C. frutescens)和绒毛辣椒(C. pubescens) (Ibiza et al., 2012).我国主要栽培品种是一年生辣椒, 其它4个栽培种具有许多优良性状, 是对一年生辣椒进行品种改良的重要种质资源(隋益虎和陈劲枫, 2009; 魏家香等, 2018).本研究以一年生辣椒、浆果状辣椒和中国辣椒为材料, 对3种辣椒R2R3-MYB转录因子家族在全基因组范围内进行鉴定, 在此基础上分析了辣椒R2R3-MYB基因的序列、motif分布、外显子-内含子结构、染色体定位、比较进化和不同组织(根、茎、叶和花)中的表达情况.通过共线性分析, 发现3种辣椒都存在特有的R2R3-MYB基因, 一年生辣椒和浆果状辣椒中的R2R3-MYB转录因子家族发生了基因组重复事件.研究结果为揭示辣椒R2R3-MYB转录因子功能及辣椒分子遗传育种奠定了理论基础. ...
... 为了研究辣椒R2R3-MYB基因的进化关系, 利用3种辣椒和拟南芥R2R3-MYB转录因子的蛋白序列构建系统进化树.与拟南芥R2R3-MYB转录因子聚集到同一分支的辣椒R2R3-MYB转录因子被认定为同一亚族, 可分为30个亚家族(S1-S30) (图2).分类中的大多数亚家族与其它植物中的亚家族一致, 但是辣椒R2R3-MYB基因家族未发现有成员属于亚家族S10、S12和S15, 暗示辣椒基因组在长期进化过程中可能丢失了一些R2R3-MYB基因(图2, 图3A).辣椒的近源物种马铃薯R2R3-MYB转录因子家族中不存在亚家族S10和S12也证明了这一点(Li et al., 2019; Sun et al., 2019). ...
... 随着基因测序技术的不断发展, 已在基因组水平上对许多植物的R2R3-MYB转录因子家族进行了鉴定, 如拟南芥(125个) (Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(249个) (Hajiebrahimi et al., 2017)、大豆(244个) (Du et al., 2012)、番茄(121个) (Zhao et al., 2014)和马铃薯(112个) (Li et al., 2019; Sun et al., 2019), 它们的基因组大小分别为125 Mb (Zapata et al., 2016)、912 Mb (Chalhoub et al., 2014)、1.03 Gb (Schmutz et al., 2010)、828 Mb (The Tomato Genome Consortium, 2012)和663 Mb (The Potato Genome Sequencing Consortium, 2011).这表明物种中R2R3-MYB基因数目与物种基因组的大小并不成正比.一年生辣椒、浆果状辣椒和中国辣椒的基因组大小分别为2.85、2.99和2.80 Gb (Kim et al., 2017).本研究从中分别鉴定出94、92和94个R2R3-MYB基因, 也支持这一观点.与茄科植物番茄(Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019)相比, 3种辣椒中R2R3-MYB基因数目明显较少, 说明R2R3-MYB转录因子家族在辣椒中发生了明显收缩. ...
... ; Sun et al., 2019)相比, 3种辣椒中R2R3-MYB基因数目明显较少, 说明R2R3-MYB转录因子家族在辣椒中发生了明显收缩. ...
... 基于3种辣椒和拟南芥R2R3-MYB基因序列的相似性, 可将其划分为30个不同的亚家族(图2).与马铃薯(Li et al., 2019; Sun et al., 2019)类似, 辣椒R2R3-MYB转录因子家族中存在部分亚家族成员丢失, 也获得了一些新的亚家族成员, 表明辣椒R2R3- MYB转录因子家族也有扩张.一年生辣椒、浆果状辣椒和中国辣椒中R2R3-MYB基因的结构分析(图3)显示, 它们主要由2个内含子、3个外显子和1个内含子、2个外显子这两类基因结构组成, 相同亚家族成员也普遍具有相同的基因结构.辣椒R2R3-MYB转录因子中包含2个MYB结构域与所有转录因子共有的motif 相对应, 位于同一亚家族的R2R3-MYB基因motif也都相同, 这说明R2R3-MYB基因的亚家族分类主要由C端序列决定. ...
Genome sequence and analysis of the tuber crop potato 1 2011
... 随着基因测序技术的不断发展, 已在基因组水平上对许多植物的R2R3-MYB转录因子家族进行了鉴定, 如拟南芥(125个) (Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(249个) (Hajiebrahimi et al., 2017)、大豆(244个) (Du et al., 2012)、番茄(121个) (Zhao et al., 2014)和马铃薯(112个) (Li et al., 2019; Sun et al., 2019), 它们的基因组大小分别为125 Mb (Zapata et al., 2016)、912 Mb (Chalhoub et al., 2014)、1.03 Gb (Schmutz et al., 2010)、828 Mb (The Tomato Genome Consortium, 2012)和663 Mb (The Potato Genome Sequencing Consortium, 2011).这表明物种中R2R3-MYB基因数目与物种基因组的大小并不成正比.一年生辣椒、浆果状辣椒和中国辣椒的基因组大小分别为2.85、2.99和2.80 Gb (Kim et al., 2017).本研究从中分别鉴定出94、92和94个R2R3-MYB基因, 也支持这一观点.与茄科植物番茄(Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019)相比, 3种辣椒中R2R3-MYB基因数目明显较少, 说明R2R3-MYB转录因子家族在辣椒中发生了明显收缩. ...
The tomato genome sequence provides insights into fleshy fruit evolution 1 2012
... 随着基因测序技术的不断发展, 已在基因组水平上对许多植物的R2R3-MYB转录因子家族进行了鉴定, 如拟南芥(125个) (Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(249个) (Hajiebrahimi et al., 2017)、大豆(244个) (Du et al., 2012)、番茄(121个) (Zhao et al., 2014)和马铃薯(112个) (Li et al., 2019; Sun et al., 2019), 它们的基因组大小分别为125 Mb (Zapata et al., 2016)、912 Mb (Chalhoub et al., 2014)、1.03 Gb (Schmutz et al., 2010)、828 Mb (The Tomato Genome Consortium, 2012)和663 Mb (The Potato Genome Sequencing Consortium, 2011).这表明物种中R2R3-MYB基因数目与物种基因组的大小并不成正比.一年生辣椒、浆果状辣椒和中国辣椒的基因组大小分别为2.85、2.99和2.80 Gb (Kim et al., 2017).本研究从中分别鉴定出94、92和94个R2R3-MYB基因, 也支持这一观点.与茄科植物番茄(Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019)相比, 3种辣椒中R2R3-MYB基因数目明显较少, 说明R2R3-MYB转录因子家族在辣椒中发生了明显收缩. ...
Characterization of a novel cotton MYB gene, GhMYB108- like responsive to abiotic stresses 1 2020
... MYB转录因子广泛分布于植物基因组中, 是植物中最大的转录因子家族之一(Riechmann et al., 2000; Dubos et al., 2010).首个植物MYB转录因子是从玉米(Zea mays)中鉴定出的COLORED1 (Paz-Ares et al., 1987).目前对MYB转录因子研究不断深入, R2R3-MYB转录因子已成为研究重点之一.据报道, R2R3-MYB转录因子参与调控许多重要的生理生化过程, 包括调节植物的初级和次生代谢、控制生长发育以及参与响应各种生物和非生物胁迫(Stracke et al., 2007).例如, 马铃薯(Solanum tuberosum) StMYB44在高温下负调控花青素的生物合成(Liu et al., 2019); 金银花(Lonicera japonica) LjaMYB12在拟南芥(Arabidopsis thaliana)中异位表达可以提高类黄酮含量, 其对类黄酮的生物合成的上下游途径具有不同的调控模式(Qi et al., 2019); 拟南芥AtMYB20、AtMYB42、AtMYB43和AtMYB85能够特异性地抑制类黄酮的生物合成(Geng et al., 2020); 棉花(Gossypium hirsutum) GhMYB108-like在干旱和盐胁迫下发挥重要作用(Ullah et al., 2020); 苹果(Malus domestica) MdMYB24-like参与茉莉酸甲酯诱导的花青素苷生物合成(Wang et al., 2019), MdMYB30在苹果角质层蜡质的积累中起重要作用, 并能增强苹果的抗病性(Zhang et al., 2019); 芝麻(Sesamum indicum) SiMYB75在拟南芥中的异位过表达显著促进根系生长, 提高植株对干旱、盐胁迫和渗透胁迫的耐受性(Dossa et al., 2020); 盐、干旱及脱落酸胁迫促进甘薯(Ipomoea batatas) IbMYB3的表达, 暗示IbMYB3在甘薯非生物胁迫应答中起重要作用(李格等, 2020). ...
Understanding regulatory networks and engineering for enhanced drought tolerance in plants 1 2006
... 转录因子在基因转录调节中起重要作用, 通过激活或抑制靶基因的转录, 调控植物的生长发育(Pabo and Sauer, 1992; Kasuga et al., 1999; Riechmann et al., 2000; Valliyodan and Nguyen, 2006).根据转录因子与其靶基因结合位点的相似性, 可将其分成不同的家族, 如AP2/EREBP、bHLH、HB、MYB和WRKY (Riechmann et al., 2000).MYB转录因子具有与DNA结合的特征性MYB结构域, 该结构域在植物中高度保守.MYB转录因子包含1-4个不完全重复的MYB结构域, 分别命名为R1 (Repeat1)、R2、R3和R4, 每个MYB结构域约有52个氨基酸残基, 形成3个α-螺旋, 其中第2和第3螺旋形成螺旋-转角-螺旋(helix-turn-helix)结构并与DNA大沟槽结合(Ogata et al., 1996; Jia et al., 2004; Dubos et al., 2010).根据包含的MYB结构域数目, MYB转录因子可分为4类, 即1R-MYB、2R-MYB (R2R3-MYB)、3R-MYB (R1R2R3- MYB)和4R-MYB (Dubos et al., 2010).1R-MYB转录因子又称MYB-related转录因子, 包含1个完整或部分MYB结构域(Stracke et al., 2001; Chen et al., 2006).3R-MYB转录因子包含3个连续的MYB结构域(R1、R2和R3), 在大多数真核生物基因组中都发现了3R-MYB转录因子, 其在调控细胞周期中发挥作用(Haga et al., 2007).4R-MYB转录因子是最小的一类, 每个基因都含有4个R1/R2 MYB结构域, 它们在植物中发挥的作用尚不明确.包含2个MYB结构域(R2和R3)的R2R3-MYB转录因子是高等植物中MYB转录因子的主要存在形式, 该家族基因可能是因3R-MYB转录因子中R1 MYB结构域缺失进化而来(Rosinski and Atchley, 1998), 也有观点认为3R-MYB转录因子是从R2R3-MYB转录因子进化来的, R1 MYB结构域从R2R3-MYB转录因子获得(Jiang et al., 2004).通常在MYB转录因子的N末端检测到高度保守的MYB结构域, C末端MYB结构域是高度分化的激活结构域, 这也决定了MYB转录因子具有多种调节作用(Dias et al., 2003; Matus et al., 2008). ...
γ-MYN: a new algorithm for estimating Ka and Ks with consideration of variable substitution rates 1 2009
... 共线性分析使用MCScanX (Wang et al., 2012)预测物种间和物种内存在的同源基因, 参数设为默认值.将3种辣椒的基因组进行两两比较, 设置E-value≤ 1e-10, 提取R2R3-MYB基因家族中的基因重复对信息, 可筛选出3种辣椒间的直系同源基因; 分别将3种辣椒的基因组与自身基因组进行比较, 设置E-value≤ 1e-10, 根据结果筛选出3种辣椒内的旁系同源基因, 即重复基因; 如果1对重复基因位于染色体上的邻近位置(<100 kb), 就可以认定为串联重复基因.使用InteractiVenn (Heberle et al., 2015)绘制Venn图.使用Circos (Krzywinski et al., 2009)显示一年生辣椒、浆果状辣椒和中国辣椒同源基因的共线性关系.使用ParaAT2.0 (Zhang et al., 2012)对同源基因对的CDS序列和ORF序列进行比对, 参数设为默认值; 使用KaKs_Calculator2.0 (Wang et al., 2010)读取比对结果, 采用γ-MYN法(Wang et al., 2009)计算同源基因对的非同义替换率(Ka)和同义替换率(Ks), 通过Ka/Ks值来评估同源基因对中同源基因在进化过程中受到的选择压力.Ka/Ks>1, 基因受到正选择; Ka/Ks<1, 基因受到纯化选择; Ka/Ks=1, 基因中性进化(Yadav et al., 2015). ...
KaKs_Calculator 2.0: a toolkit incorporating gamma-series methods and sliding window strategies 1 2010
... 共线性分析使用MCScanX (Wang et al., 2012)预测物种间和物种内存在的同源基因, 参数设为默认值.将3种辣椒的基因组进行两两比较, 设置E-value≤ 1e-10, 提取R2R3-MYB基因家族中的基因重复对信息, 可筛选出3种辣椒间的直系同源基因; 分别将3种辣椒的基因组与自身基因组进行比较, 设置E-value≤ 1e-10, 根据结果筛选出3种辣椒内的旁系同源基因, 即重复基因; 如果1对重复基因位于染色体上的邻近位置(<100 kb), 就可以认定为串联重复基因.使用InteractiVenn (Heberle et al., 2015)绘制Venn图.使用Circos (Krzywinski et al., 2009)显示一年生辣椒、浆果状辣椒和中国辣椒同源基因的共线性关系.使用ParaAT2.0 (Zhang et al., 2012)对同源基因对的CDS序列和ORF序列进行比对, 参数设为默认值; 使用KaKs_Calculator2.0 (Wang et al., 2010)读取比对结果, 采用γ-MYN法(Wang et al., 2009)计算同源基因对的非同义替换率(Ka)和同义替换率(Ks), 通过Ka/Ks值来评估同源基因对中同源基因在进化过程中受到的选择压力.Ka/Ks>1, 基因受到正选择; Ka/Ks<1, 基因受到纯化选择; Ka/Ks=1, 基因中性进化(Yadav et al., 2015). ...
The R2R3-MYB transcription factor MdMYB24-like is involved in methyl jasmonate-induced anthocyanin biosynthesis in apple 1 2019
... MYB转录因子广泛分布于植物基因组中, 是植物中最大的转录因子家族之一(Riechmann et al., 2000; Dubos et al., 2010).首个植物MYB转录因子是从玉米(Zea mays)中鉴定出的COLORED1 (Paz-Ares et al., 1987).目前对MYB转录因子研究不断深入, R2R3-MYB转录因子已成为研究重点之一.据报道, R2R3-MYB转录因子参与调控许多重要的生理生化过程, 包括调节植物的初级和次生代谢、控制生长发育以及参与响应各种生物和非生物胁迫(Stracke et al., 2007).例如, 马铃薯(Solanum tuberosum) StMYB44在高温下负调控花青素的生物合成(Liu et al., 2019); 金银花(Lonicera japonica) LjaMYB12在拟南芥(Arabidopsis thaliana)中异位表达可以提高类黄酮含量, 其对类黄酮的生物合成的上下游途径具有不同的调控模式(Qi et al., 2019); 拟南芥AtMYB20、AtMYB42、AtMYB43和AtMYB85能够特异性地抑制类黄酮的生物合成(Geng et al., 2020); 棉花(Gossypium hirsutum) GhMYB108-like在干旱和盐胁迫下发挥重要作用(Ullah et al., 2020); 苹果(Malus domestica) MdMYB24-like参与茉莉酸甲酯诱导的花青素苷生物合成(Wang et al., 2019), MdMYB30在苹果角质层蜡质的积累中起重要作用, 并能增强苹果的抗病性(Zhang et al., 2019); 芝麻(Sesamum indicum) SiMYB75在拟南芥中的异位过表达显著促进根系生长, 提高植株对干旱、盐胁迫和渗透胁迫的耐受性(Dossa et al., 2020); 盐、干旱及脱落酸胁迫促进甘薯(Ipomoea batatas) IbMYB3的表达, 暗示IbMYB3在甘薯非生物胁迫应答中起重要作用(李格等, 2020). ...
MCScanX: a toolkit for detection and evolutionary analysis of gene synteny and collinearity 1 2012
... 共线性分析使用MCScanX (Wang et al., 2012)预测物种间和物种内存在的同源基因, 参数设为默认值.将3种辣椒的基因组进行两两比较, 设置E-value≤ 1e-10, 提取R2R3-MYB基因家族中的基因重复对信息, 可筛选出3种辣椒间的直系同源基因; 分别将3种辣椒的基因组与自身基因组进行比较, 设置E-value≤ 1e-10, 根据结果筛选出3种辣椒内的旁系同源基因, 即重复基因; 如果1对重复基因位于染色体上的邻近位置(<100 kb), 就可以认定为串联重复基因.使用InteractiVenn (Heberle et al., 2015)绘制Venn图.使用Circos (Krzywinski et al., 2009)显示一年生辣椒、浆果状辣椒和中国辣椒同源基因的共线性关系.使用ParaAT2.0 (Zhang et al., 2012)对同源基因对的CDS序列和ORF序列进行比对, 参数设为默认值; 使用KaKs_Calculator2.0 (Wang et al., 2010)读取比对结果, 采用γ-MYN法(Wang et al., 2009)计算同源基因对的非同义替换率(Ka)和同义替换率(Ks), 通过Ka/Ks值来评估同源基因对中同源基因在进化过程中受到的选择压力.Ka/Ks>1, 基因受到正选择; Ka/Ks<1, 基因受到纯化选择; Ka/Ks=1, 基因中性进化(Yadav et al., 2015). ...
Genome-wide development of transposable elements-based markers in foxtail millet and construction of an integrated database 2 2015
... 共线性分析使用MCScanX (Wang et al., 2012)预测物种间和物种内存在的同源基因, 参数设为默认值.将3种辣椒的基因组进行两两比较, 设置E-value≤ 1e-10, 提取R2R3-MYB基因家族中的基因重复对信息, 可筛选出3种辣椒间的直系同源基因; 分别将3种辣椒的基因组与自身基因组进行比较, 设置E-value≤ 1e-10, 根据结果筛选出3种辣椒内的旁系同源基因, 即重复基因; 如果1对重复基因位于染色体上的邻近位置(<100 kb), 就可以认定为串联重复基因.使用InteractiVenn (Heberle et al., 2015)绘制Venn图.使用Circos (Krzywinski et al., 2009)显示一年生辣椒、浆果状辣椒和中国辣椒同源基因的共线性关系.使用ParaAT2.0 (Zhang et al., 2012)对同源基因对的CDS序列和ORF序列进行比对, 参数设为默认值; 使用KaKs_Calculator2.0 (Wang et al., 2010)读取比对结果, 采用γ-MYN法(Wang et al., 2009)计算同源基因对的非同义替换率(Ka)和同义替换率(Ks), 通过Ka/Ks值来评估同源基因对中同源基因在进化过程中受到的选择压力.Ka/Ks>1, 基因受到正选择; Ka/Ks<1, 基因受到纯化选择; Ka/Ks=1, 基因中性进化(Yadav et al., 2015). ...
Chromosome-level assembly of Arabidopsis thaliana Ler reveals the extent of translocation and inversion polymorphisms 1 2016
... 随着基因测序技术的不断发展, 已在基因组水平上对许多植物的R2R3-MYB转录因子家族进行了鉴定, 如拟南芥(125个) (Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(249个) (Hajiebrahimi et al., 2017)、大豆(244个) (Du et al., 2012)、番茄(121个) (Zhao et al., 2014)和马铃薯(112个) (Li et al., 2019; Sun et al., 2019), 它们的基因组大小分别为125 Mb (Zapata et al., 2016)、912 Mb (Chalhoub et al., 2014)、1.03 Gb (Schmutz et al., 2010)、828 Mb (The Tomato Genome Consortium, 2012)和663 Mb (The Potato Genome Sequencing Consortium, 2011).这表明物种中R2R3-MYB基因数目与物种基因组的大小并不成正比.一年生辣椒、浆果状辣椒和中国辣椒的基因组大小分别为2.85、2.99和2.80 Gb (Kim et al., 2017).本研究从中分别鉴定出94、92和94个R2R3-MYB基因, 也支持这一观点.与茄科植物番茄(Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019)相比, 3种辣椒中R2R3-MYB基因数目明显较少, 说明R2R3-MYB转录因子家族在辣椒中发生了明显收缩. ...
The R2R3 MYB transcription factor MdMYB30 modulates plant resistance against pathogens by regulating cuticular wax biosynthesis 1 2019
... MYB转录因子广泛分布于植物基因组中, 是植物中最大的转录因子家族之一(Riechmann et al., 2000; Dubos et al., 2010).首个植物MYB转录因子是从玉米(Zea mays)中鉴定出的COLORED1 (Paz-Ares et al., 1987).目前对MYB转录因子研究不断深入, R2R3-MYB转录因子已成为研究重点之一.据报道, R2R3-MYB转录因子参与调控许多重要的生理生化过程, 包括调节植物的初级和次生代谢、控制生长发育以及参与响应各种生物和非生物胁迫(Stracke et al., 2007).例如, 马铃薯(Solanum tuberosum) StMYB44在高温下负调控花青素的生物合成(Liu et al., 2019); 金银花(Lonicera japonica) LjaMYB12在拟南芥(Arabidopsis thaliana)中异位表达可以提高类黄酮含量, 其对类黄酮的生物合成的上下游途径具有不同的调控模式(Qi et al., 2019); 拟南芥AtMYB20、AtMYB42、AtMYB43和AtMYB85能够特异性地抑制类黄酮的生物合成(Geng et al., 2020); 棉花(Gossypium hirsutum) GhMYB108-like在干旱和盐胁迫下发挥重要作用(Ullah et al., 2020); 苹果(Malus domestica) MdMYB24-like参与茉莉酸甲酯诱导的花青素苷生物合成(Wang et al., 2019), MdMYB30在苹果角质层蜡质的积累中起重要作用, 并能增强苹果的抗病性(Zhang et al., 2019); 芝麻(Sesamum indicum) SiMYB75在拟南芥中的异位过表达显著促进根系生长, 提高植株对干旱、盐胁迫和渗透胁迫的耐受性(Dossa et al., 2020); 盐、干旱及脱落酸胁迫促进甘薯(Ipomoea batatas) IbMYB3的表达, 暗示IbMYB3在甘薯非生物胁迫应答中起重要作用(李格等, 2020). ...
ParaAT: a parallel tool for constructing multiple protein-coding DNA alignments 1 2012
... 共线性分析使用MCScanX (Wang et al., 2012)预测物种间和物种内存在的同源基因, 参数设为默认值.将3种辣椒的基因组进行两两比较, 设置E-value≤ 1e-10, 提取R2R3-MYB基因家族中的基因重复对信息, 可筛选出3种辣椒间的直系同源基因; 分别将3种辣椒的基因组与自身基因组进行比较, 设置E-value≤ 1e-10, 根据结果筛选出3种辣椒内的旁系同源基因, 即重复基因; 如果1对重复基因位于染色体上的邻近位置(<100 kb), 就可以认定为串联重复基因.使用InteractiVenn (Heberle et al., 2015)绘制Venn图.使用Circos (Krzywinski et al., 2009)显示一年生辣椒、浆果状辣椒和中国辣椒同源基因的共线性关系.使用ParaAT2.0 (Zhang et al., 2012)对同源基因对的CDS序列和ORF序列进行比对, 参数设为默认值; 使用KaKs_Calculator2.0 (Wang et al., 2010)读取比对结果, 采用γ-MYN法(Wang et al., 2009)计算同源基因对的非同义替换率(Ka)和同义替换率(Ks), 通过Ka/Ks值来评估同源基因对中同源基因在进化过程中受到的选择压力.Ka/Ks>1, 基因受到正选择; Ka/Ks<1, 基因受到纯化选择; Ka/Ks=1, 基因中性进化(Yadav et al., 2015). ...
Genome- wide identification and characterization of R2R3MYB family in Solanum lycopersicum 3 2014
... 随着测序技术的发展, R2R3-MYB转录因子家族先后在许多高等植物的全基因组水平得到系统鉴定和分析, 包括拟南芥(Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(Brassica napus) (Hajiebrahimi et al., 2017)、大豆(Glycine max) (Du et al., 2012)、番茄(Solanum lycopersicum) (Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019).基因组测序技术促进了园艺植物基因组学和遗传学研究(唐嘉瓅等, 2020).辣椒是茄科(Solanaceae)辣椒属(Capsicum)一年生或多年生草本植物, 是具有重要经济价值的蔬菜作物, 在世界范围内广泛种植.辣椒属包含27种, 被人类驯化长期栽培的辣椒有5种, 分别为一年生辣椒(Capsicum annuum)、浆果状辣椒(C. baccatum)、中国辣椒(C. chinense)、灌木状辣椒(C. frutescens)和绒毛辣椒(C. pubescens) (Ibiza et al., 2012).我国主要栽培品种是一年生辣椒, 其它4个栽培种具有许多优良性状, 是对一年生辣椒进行品种改良的重要种质资源(隋益虎和陈劲枫, 2009; 魏家香等, 2018).本研究以一年生辣椒、浆果状辣椒和中国辣椒为材料, 对3种辣椒R2R3-MYB转录因子家族在全基因组范围内进行鉴定, 在此基础上分析了辣椒R2R3-MYB基因的序列、motif分布、外显子-内含子结构、染色体定位、比较进化和不同组织(根、茎、叶和花)中的表达情况.通过共线性分析, 发现3种辣椒都存在特有的R2R3-MYB基因, 一年生辣椒和浆果状辣椒中的R2R3-MYB转录因子家族发生了基因组重复事件.研究结果为揭示辣椒R2R3-MYB转录因子功能及辣椒分子遗传育种奠定了理论基础. ...
... 随着基因测序技术的不断发展, 已在基因组水平上对许多植物的R2R3-MYB转录因子家族进行了鉴定, 如拟南芥(125个) (Stracke et al., 2001; Chen et al., 2006)、甘蓝型油菜(249个) (Hajiebrahimi et al., 2017)、大豆(244个) (Du et al., 2012)、番茄(121个) (Zhao et al., 2014)和马铃薯(112个) (Li et al., 2019; Sun et al., 2019), 它们的基因组大小分别为125 Mb (Zapata et al., 2016)、912 Mb (Chalhoub et al., 2014)、1.03 Gb (Schmutz et al., 2010)、828 Mb (The Tomato Genome Consortium, 2012)和663 Mb (The Potato Genome Sequencing Consortium, 2011).这表明物种中R2R3-MYB基因数目与物种基因组的大小并不成正比.一年生辣椒、浆果状辣椒和中国辣椒的基因组大小分别为2.85、2.99和2.80 Gb (Kim et al., 2017).本研究从中分别鉴定出94、92和94个R2R3-MYB基因, 也支持这一观点.与茄科植物番茄(Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019)相比, 3种辣椒中R2R3-MYB基因数目明显较少, 说明R2R3-MYB转录因子家族在辣椒中发生了明显收缩. ...
... 基因, 也支持这一观点.与茄科植物番茄(Zhao et al., 2014)和马铃薯(Li et al., 2019; Sun et al., 2019)相比, 3种辣椒中R2R3-MYB基因数目明显较少, 说明R2R3-MYB转录因子家族在辣椒中发生了明显收缩. ...
 ,2,*, 胡能兵2, 隋益虎2, 张开京2, 艾昊2
,2,*, 胡能兵2, 隋益虎2, 张开京2, 艾昊2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT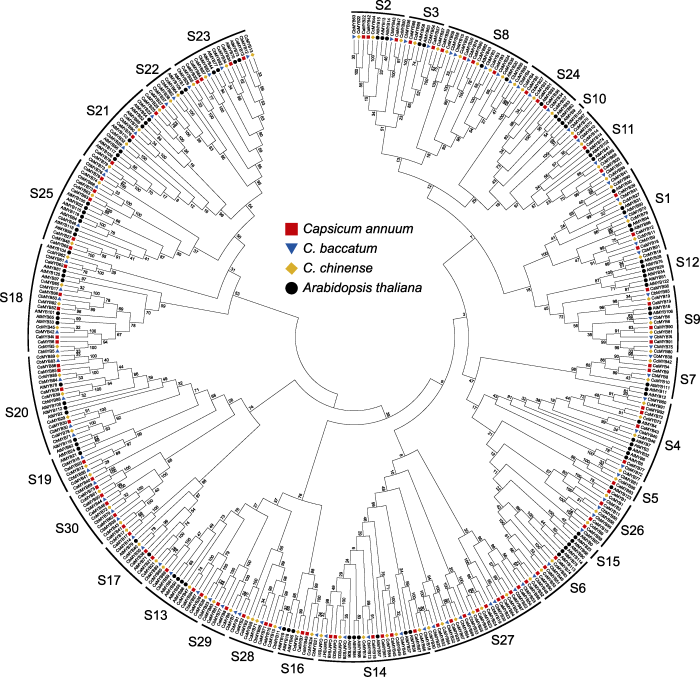 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT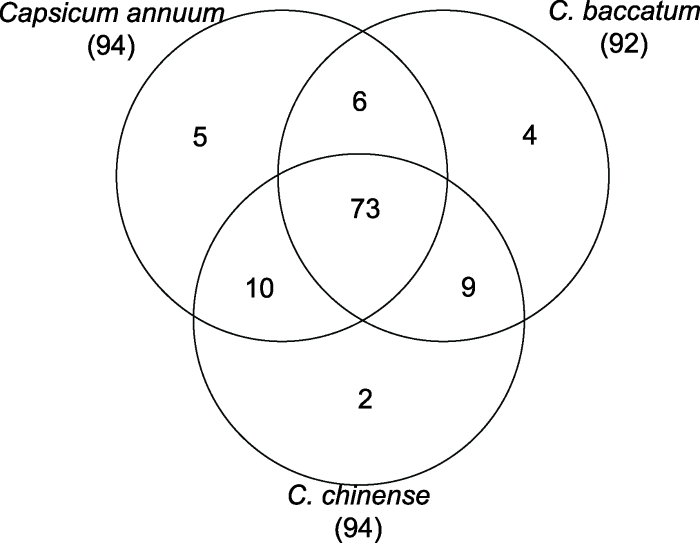 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT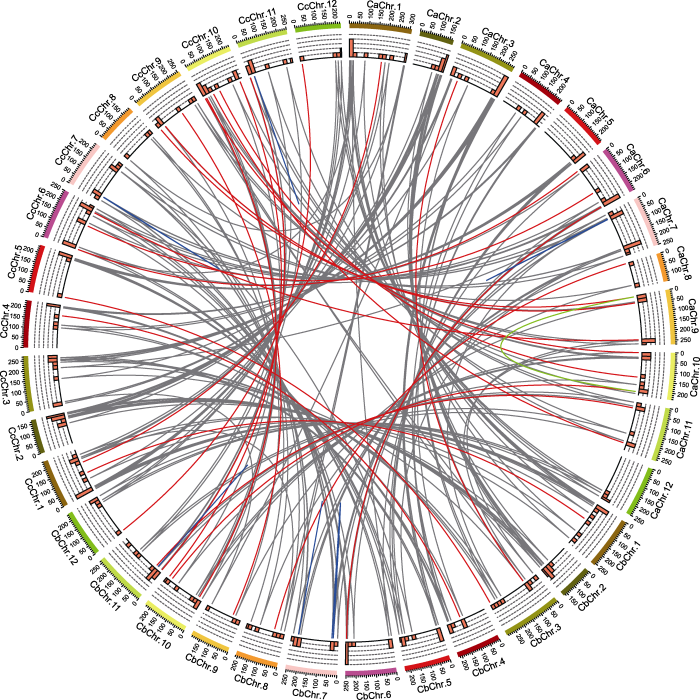 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT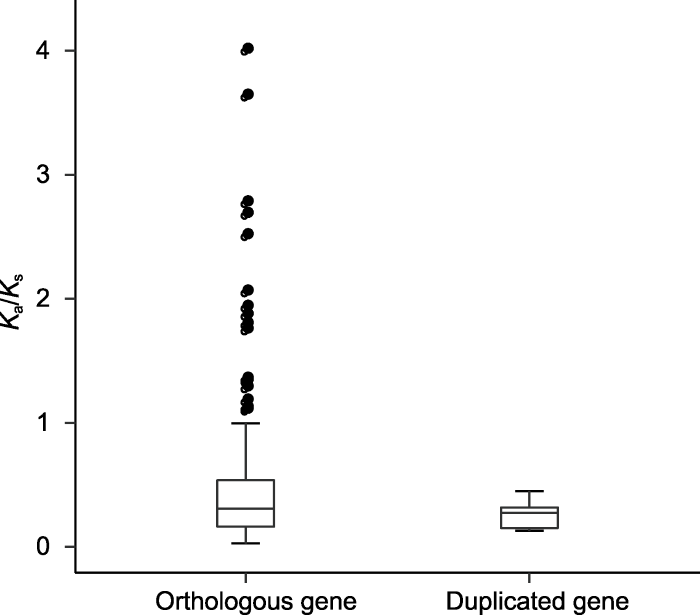 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT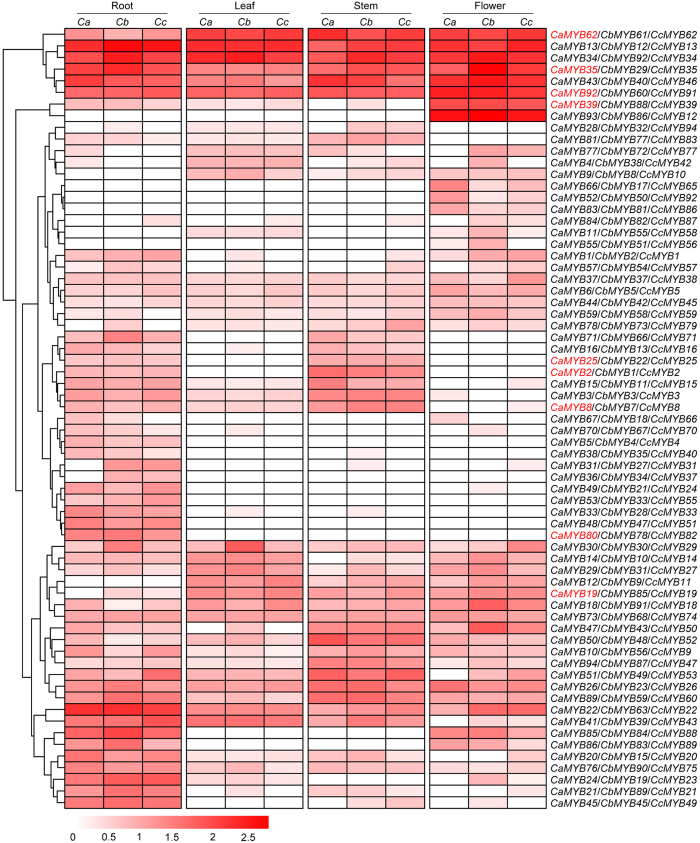 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT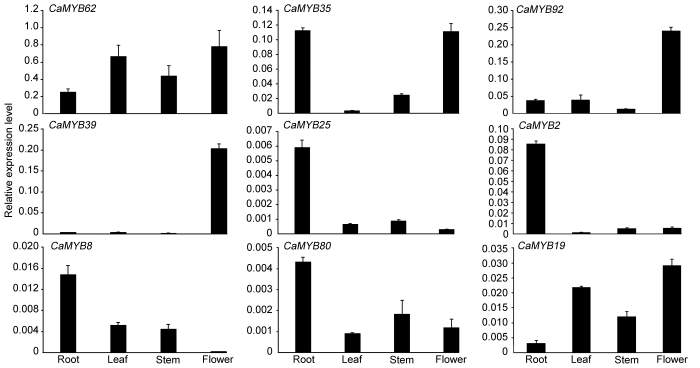 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}