Triploid in Poaceae: Formation, Detection, and Utilization
Xu Yan1,2, Yanchun Zuo1, Honglin Wang1, Yang Li2,3, Yingzheng Li2, Jing Kou1, Qilin Tang1, Xiaokang Zhou,2,*, Zhouhe Du,1,*1Sericulture Research Institute/Animal Husbandry Research Center, Sichuan Academy of Agricultural Sciences, Nanchong 637000, China 2Maize Research Institute, Sichuan Agricultural University, Wenjiang 611130, China 3School of Urban and Rural Planning and Construction, Mianyang Teachers’ College, Mianyang 621000, China
Abstract The formation pathway of triploid in Poaceae includes 2n gamete fusion, interploidy cross, polyspermy, and endosperm culture. The fusion of reduced male (n) and unreduced female gamete (2n) is the main method for forming natural triploids. Interploidy cross is the main method for synthesizing artificial triploids. The application of ploidy identification methods such as morphological observation, chromosome analysis, flow cytometry and molecular markers in gramineous triploids is introduced, and the advantages and disadvantages of different methods are also discussed. At present, triploid has no direct application value in cereal crops, but it can be used as a genetic bridge to synthetize polyploid and aneuploid, as well as to transfer alien genes from wild species to cultivated species. Gramineous triploids (especially allotriploid) are widely cultivated for forage or biofuel production, suggesting that triploidy breeding may be directly performed in this type of grasses. We discuss the future prospect of research on gramineous triploid, e.g., polyploid- and apomixis-triploid breeding. Particularly, endosperm culture can synthesize triploids in one step, and polyspermy can achieve one-step fusion of three genetically different plant genomes, which should be paid attention to in the triploidy research. Due to rare occurrence of 2n gamete fusion and polyspermy, and frequent chromosomal variation in ploidy hybridization and endosperm culture, the development of high-throughput triploid identification technology will become the key for breakthrough in triploidy generation/breeding. Keywords:triploid;2n gamete;interploidy cross;endosperm culture;polyspermy;breeding program
PDF (2274KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 严旭, 左艳春, 王红林, 李杨, 李影正, 寇晶, 唐祈林, 周晓康, 杜周和. 禾本科三倍体: 形成、鉴定与利用. 植物学报, 2021, 56(3): 372-387 doi:10.11983/CBB20166 Yan Xu, Zuo Yanchun, Wang Honglin, Li Yang, Li Yingzheng, Kou Jing, Tang Qilin, Zhou Xiaokang, Du Zhouhe. Triploid in Poaceae: Formation, Detection, and Utilization. Chinese Bulletin of Botany, 2021, 56(3): 372-387 doi:10.11983/CBB20166
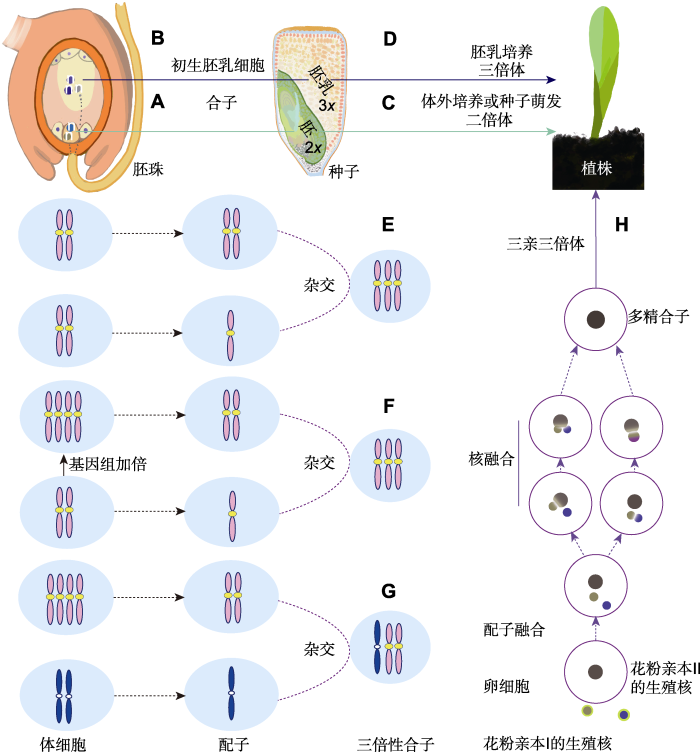
(A) A sperm fertilizes an egg cell to produce diploid zygote which subsequently grows into diploid embryo; (B) A sperm fertilizes a center cell to produce triploid primary endosperm cell which subsequently grows into endosperm; (C) The embryo develops into diploid plant through in vitro culture or seed germination; (D) The endosperm develops into triploid plant via culturing in vitro; (E) Triploid formation by a 2n+n mating; (F) The autotriploid hybrid produced by crossing tetraploid (2n=4x=AAAA) and diploid (2n=2x=AA); (G) The allotriploid hybrid produced by crossing tetraploid (2n=4x=AAAA) and diploid (2n=2x=BB); (H) Formation of triploid by polyspermy
目前, 通过观测小孢子发生在植物中已发现5种2n配子形成机制(Loginova and Silkova, 2017) (图2)。减数分裂II期姐妹染色单体分离形成的FDR (first division restitution)型2n配子(图2C)在很大程度上保留了亲本的杂合性, 而减数分裂II期姐妹染色单体不分离形成的SDR (second division restitution)型2n配子(图2D)高度纯合。因此, 在转移亲本杂合性方面FDR型比SDR型更为有利(Younis et al., 2014)。Harlan和De Wet (1975)认为所有植物都以某种频率产生2n配子, 但物种间发生频率不同。二倍体水稻(Oryza sativa, 2n=2x=24)多胚苗能自发地产生三倍体(张静等, 2002)。在二倍体看麦娘(Alopecurus bulbosus, 2n=2x=14)自然群体中, 二倍体、三倍体与四倍体并存(Sieber and Murray, 1979)。在22个二倍体大麦(Hordeum vulgare, 2n=2x=14)品种中, 三倍体植株的发生率为0.01%-0.29% (Sandfaer, 1975)。在黄花茅(Anthoxanthum alpinum, 2n=2x=10)中2n配子的发生率为6.3% (Bretagnolle, 2001)。对十字花科24个物种60个群体的研究表明, 尽管2n配子先前被认为相对罕见、但普遍发生(通常个体的雄配子2n发生率<2%), 并且在特定个体中较高, 这些极少数产生高频率2n配子的个体可能在多倍体的形成中发挥关键作用(Kreiner et al., 2017)。在同一植物中, 雌雄2n配子的发生率也不一致(Ramsey and Schemske, 1998)。鸭茅(Dactylis glomerata, 2n=2x=14) 2n雌配子发生率为0.49%, 而2n雄配子发生率为0.98% (Maceira et al.,1992)。鉴于2n配子发生率低, 在2x/2x下通过单向有性多倍化(unilateral sexual polyploidization, 2x+x)形成三倍体的频率远高于通过双向有性多倍化(bilateral sexual polyploidization, 2x+2x)形成四倍体的频率。例如, 在黄花茅中2n配子形成三倍体的发生率为0.2%, 但未见四倍体发生(Bretagnolle, 2001)。
(A) 与体细胞相比正常配子的染色体数目减半; (B) 通过胞质融合或多核体形成的四倍体花粉母细胞产生PRD型2n配子, 如鸭茅(Falistocco et al., 1995); (C) 在减数分裂I期同源染色体无配对和分裂形成的FDR型2n配子将保留除重组片段外的所有亲本基因, 如雀稗(Filho et al., 2014)和多年生黑麦草(Chen et al., 1997); (D) 在减数分裂II期因分裂缺失形成的SDR型2n配子将保留除重组片段外比正常配子多1套染色体副本的2n配子, 如多年生黑麦草(Chen et al., 1997); (E) 在减数分裂I期部分染色体以姐妹染色单体分离, 其余以二价体分离, 随后在减数分裂II期分裂缺失形成IMR型2n配子, 如百合(Lim et al., 2001); (F) 减数分裂正常而进入配子体发育阶段时基因组加倍形成PMD型2n配子, 如马铃薯(Bastiaanssen et al., 1998) Figure 2Mechanisms of 2n gamete formation
(A) Chromosome numbers of normal gametes are halved compared with somatic cells; (B) PRD (pre-meiotic doubling) gametes are obtained as a result of the tetraploid pollen mother cells formed by cytomixis or syncytium, e.g., Dactylis glomerata (Falistocco et al., 1995); (C) In the FDR (first division restitution) type, pairing and division of homoeologous chromosomes do not occur during meiosis I, and the FDR 2n gametes maintain all of their parental genes except cross-over fragments, e.g., Paspalum jesuiticum (Filho et al., 2014) and Lolium perenne (Chen et al., 1997); (D) SDR (second division restitution) gametes with two copies of non-recombinant chromosomes are the result of the second division omission or cytokinesis abnormalities after normal anaphase II, e.g., L. perenne (Chen et al., 1997); (E) IMR (indeterminate meiotic restitution) gametes were discovered in Lilium longiflorum × Asiatic hybrid (Lim et al., 2001), in which some chromosomes are separated as univalents during meiosis I whereas the others are separated as bivalents, and subsequently the second division omission occurs during meiosis II; (F) PMD (post-meiotic doubling) gametes occurs due to the genome doubling of haploid spores during microgametogenesis, e.g., Solanum tuberosum (Bastiaanssen et al., 1998)
实践中, 2n配子可通过环境调控、化学诱导、种间杂种和减数分裂突变体获得。2n配子的发生主要受遗传控制, 但暴露于低温或高温、热冲击和高辐射下可诱导或刺激2n配子形成(Younis et al., 2014)。例如, 蔷薇属(Rosa)植物花芽暴露于36°C高温下可提高2n雄配子的发生率(Pécrix et al., 2011)。在禾本科中采用环境调控诱导2n配子发生尚未见报道。N2O在诱导鸭茅(Van Santen et al., 1991)、玉米(Zea mays, 2n=2x=20) (Kato and Birchler, 2006)、小麦(Triticum aestivum, 2n=6x=42)及大麦(Dvorak et al., 1973)的2n配子上取得成功。向玉米雄穗喷洒氟乐灵可诱导2n花粉并获得同源三倍体(Kato, 1999)。种间或属间杂种是产生2n配子的重要途径, 其频率高于祖先物种(Ramsey and Schemske, 1998)。Mason等(2011)分析了3种芸薹属(Brassica)物种及其F1杂种的2n雄配子发生率, 发现5个甘蓝型油菜(B. napus, 2n=4x=38)品系的发生率(0%-0.11%, 平均值为0.03%)较低, 而甘蓝型油菜的11种F1杂种的发生率(0.17%-3.29%, 平均值为1.55%)较高。可见, 种间杂种较亲本基因组2n配子的发生率提高。Zeng等(2020)发现在兰花(Cymbidium ssp.)传统品种中2n雄配子发生率为0.15%-1.0%, 而种间杂种为2.5%-4.03%; 同时作者认为可能由于无性繁殖植物存在可替代的繁殖途径, 2n配子的形成频率相对更高。减数分裂活跃基因的突变可能导致纺锤体形成异常或胞质分裂受阻而形成2n配子, 如拟南芥(Arabidopsis thaliana, 2n= 2x=10)中的cyca1;2/tam (SDR型)、osd1 (SDR型)和Atps1 (FDR型)突变体(Brownfield and K?hler, 2011)。
(A) 三倍体作为品种直接利用; (B) 通过辐射、转基因或组织培养等技术改良三倍体品种; (C) 作为通往更高倍性的“三倍体桥”; (D) 转移异源基因; (E) 通过3x/2x合成非整倍体 Figure 3Application of triploid in Poaceae
(A) The direct use of triploid cultivar; (B) The improvement of triploid cultivar through techniques such as radiation, genetic modification, or tissue culture; (C) Use of “triploids bridge” for production of polyploids; (D) Alien genes transfer; (E) Development of aneuploidy by 3x/2x mating
3.2 桥梁作用
对于有直接利用价值的三倍体, 可通过组培、诱变和转基因等技术进一步改良(图3B)。MS-Supreme源于Tifgreen的自然突变, 与Tifgreen相比其叶片和茎节节间缩小、变短(Krans et al., 1999)。在Tifgreen的自然突变中还选育出Tifdwarf和FloraDwarf等矮生品种; TifwayII、TifSport和TifEagle则由相应三倍体亲本经γ辐射诱变而成(Kamps et al., 2011)。利用体细胞无性系变异育种技术在TifEagle的愈伤组织中长时间连续继代培养能有效诱导体细胞变异(如矮化突变体); 以0.3 mol·L-1 NaCl为选择压筛选出耐盐细胞系(Lu et al., 2007)。以美洲狼尾草(Pennisetum americana)/ 四倍体象草合成的三倍体F1种子为诱变材料, 经γ辐射获得优异变异株闽牧6号(陈钟佃等, 2012)。华南1号杂交狼尾草由三倍体杂交狼尾草芽变而来(陈平等, 2004)。三倍体也可以直接作为亲本利用。在偏序钝叶草(Stenotaphrum secundatum, 2n=2x=18)的3x/ 2x杂交后代中经胚拯救选育出抗灰斑病(由稻瘟病菌(Magnaporthe grisea)引起)和耐旱草坪草新品种(Chandra et al., 2015)。奇岗天然后代群体中16个数量性状出现了丰富的变异(变异系数为14.41%- 151.85%), 在后代中筛选出3份材料, 其生物量优于奇岗(潘志军等, 2017)。通过三倍体狼尾草与摩特矮象草杂交选育出中国南方地区广泛种植的三倍体多年生牧草桂牧1号(罗宗志等, 2016)。宁牧26-2狼尾草也是由三倍体后代衍生而来(顾洪如等, 1992)。三倍体转基因品系可通过2种策略获得。(1) 直接以三倍体为受体, 导入目的基因构建转基因品系, 如三倍体狗牙根Tifeagle (Hu et al., 2005)。(2) 对于愈伤组织形成与再分化率低的三倍体, 首先需要寻找愈伤组织再分化优良的二倍体或四倍体作为转基因桥梁, 然后通过基因枪或农杆菌转化等方法导入目的基因, 最后通过倍性间杂交构建转基因三倍体, 如芒(2n=2x=38) (Wang et al., 2011)。近年来, 基因编辑技术的发展为定向改良三倍体提供了新的育种方案。
三倍体可作为通往其它倍性水平的桥梁, 即三倍体桥(triploid-bridge) (图3C)。在鸭茅三倍体后代中发现了四倍体、五倍体以及混倍体植株(钟声, 2006)。三倍体摩擦禾可产生2n雌配子, 通过3x/2x合成了四倍体(Kindiger and Dewald, 1994)。以三倍体杂交狼尾草(美洲狼尾草/象草, 2n=3x=21)的幼穗离体培养获得的胚性愈伤组织为材料, 通过秋水仙碱诱导获得六倍体(钟小仙等, 2014)。利用秋水仙碱处理象草(2n=4x=28)/珍珠粟(P. glaucum, 2n=2x=14)杂交三倍体幼苗, 体细胞加倍后形成异源六倍体(Campos et al., 2009)。用类似的方法获得M. × miscanthus六倍体(Yu et al., 2009; Ghimire et al., 2016)。水稻(AA)和高秆野生稻(O. alta, CCDD)杂交形成异源三倍体, 加倍后形成三基异源六倍体(AACCDD) (祝剑峰等, 2008)。
BajajYPS, SainiSS, BidaniM (1980). Production of triploid plants from the immature and mature endosperm cultures of rice Theor Appl Genet 58, 17-18. DOI:10.1007/BF00264662PMID:24301212 [本文引用: 1] Triploid plants were induced to regenerate from immature and mature endosperm cultures of three cultivars of rice. The plants were subsequently established in the soil. The significance of the induction of genetic variability in the germplasm of rice is discussed.
BastiaanssenHJM, Van Den BergPMMM, LindhoutP, JacobsenE, RamannaMS (1998). Postmeiotic restitution in 2 n-egg formation of diploid potato Heredity 81, 20-27. DOI:10.1046/j.1365-2540.1998.00370.xURL [本文引用: 2]
BealeKM, LeydonAR, JohnsonMA (2012). Gamete fusion is required to block multiple pollen tubes from entering an Arabidopsis ovule Curr Biol 22, 1090-1094. DOI:10.1016/j.cub.2012.04.041URL [本文引用: 1]
BretagnolleFO (2001). Pollen production and spontaneous polyploidization in diploid populations of Anthoxanthum alpinum Biol J Linn Soc 72, 241-247. DOI:10.1111/j.1095-8312.2001.tb01314.xURL [本文引用: 2]
BrownfieldL, K?hlerC (2011). Unreduced gamete formation in plants: mechanisms and prospects J Exp Bot 62, 1659-1668. DOI:10.1093/jxb/erq371URL [本文引用: 1]
CamposJMS, DavideLC, SalgadoCC, SantosFC, CostaPN, SilvaPS, AlvesCCS, VicciniLF, PereiraAV (2009). In vitro induction of hexaploid plants from triploid hybrids of Pennisetum purpureum and Pennisetum glaucum Plant Breed 128, 101-104. DOI:10.1111/pbr.2009.128.issue-1URL [本文引用: 1]
ChenC, SleperDA, ChaoS, JohalGS, WestCP (1997). RFLP detection of 2 n pollen formation by first and second division restitution in perennial ryegrass Crop Sci 37, 76-80. DOI:10.2135/cropsci1997.0011183X003700010012xURL [本文引用: 5]
De StormeN, ZamariolaL, MauM, SharbelTF, GeelenD (2013). Volume-based pollen size analysis: an advanced method to assess somatic and gametophytic ploidy in flowering plants Plant Reprod 26, 65-81. DOI:10.1007/s00497-012-0209-0PMID:23686220 [本文引用: 1] Pollen size is often used as a biological parameter to estimate the ploidy and viability of mature pollen grains. In general, pollen size quantification is performed one- or two-dimensionally using image-based diameter measurements. As these approaches are elaborate and time consuming, alternative approaches that enable a quick, reliable analysis of pollen size are highly relevant for plant research. In this study, we present the volume-based particle size analysis technique as an alternative method to characterize mature pollen. Based on a comparative assay using different plant species (including tomato, oilseed rape, kiwifruit, clover, among others), we found that volume-based pollen size measurements are not biased by the pollen shape or position and substantially reduce non-biological variation, allowing a more accurate determination of the actual pollen size. As such, volume-based particle size techniques have a strong discriminative power in detecting pollen size differences caused by alterations in the gametophytic ploidy level and therefore allow for a quick and reliable estimation of the somatic ploidy level. Based on observations in Arabidopsis thaliana gametophytic mutants and differentially reproducing Boechera polyantha lines, we additionally found that volume-based pollen size analysis provides quantitative and qualitative data about alterations in male sporogenesis, including aneuploid and diploid gamete formation. Volume-based pollen size analysis therefore not only provides a quick and easy methodology to determine the somatic ploidy level of flowering plants, but can also be used to determine the mode of reproduction and to quantify the level of diplogamete formation.
DelomasTA (2019). Differentiating diploid and triploid individuals using single nucleotide polymorphisms genotyped by amplicon sequencing Mol Ecol Resour 19, 1545-1551. DOI:10.1111/1755-0998.13073PMID:31448561 [本文引用: 1] Triploidy can occur naturally or be induced in fish and shellfish during artificial propagation in order to produce sterile individuals. Fisheries managers often stock these sterile triploids as a means of improving angling opportunities without risking unwanted reproduction of the stocked fish. Additionally, the rearing of all-triploid individuals has been suggested as a means to reduce the possibility of escaped aquaculture fish interbreeding with wild populations. Efficient means of determining if an individual is triploid or diploid are therefore needed both to monitor the efficacy of triploidy-inducing treatments and, when sampling fish from a body of water that has a mixture of diploids and triploids, to determine the ploidy of a fish prior to further analyses. Currently, ploidy is regularly measured through flow cytometry, but this technique typically utilizes a fresh blood sample. This study presents an alternative, cost-effective method of determining ploidy by analysing amplicon-sequencing data for biallelic single-nucleotide polymorphisms (SNPs). For each sample, heterozygous genotypes are identified and the likelihoods of diploidy and triploidy are calculated based on the read counts for each allele. The accuracy of this method is demonstrated using triploid and diploid brook trout (Salvelinus fontinalis) genotyped with a panel of 234 SNPs and Chinook salmon (Oncorhynchus tshawytscha) genotyped with a panel of 298 SNPs following the GT-seq methodology of amplicon sequencing.? 2019 John Wiley & Sons Ltd.
DvorakJ, HarveyBL, CoulmanBE (1973). The use of nitrous oxide for producing eupolyploids and aneuploids in wheat and barley Can J Genet Cytol 15, 205-214. DOI:10.1139/g73-021URL [本文引用: 1]
FalistoccoE, TostiN, FalcinelliM (1995). Cytomixis in pollen mother cells of diploid Dactylis, one of the origins of 2n gametes J Hered 86, 448-453. DOI:10.1093/oxfordjournals.jhered.a111619URL [本文引用: 2]
FilhoRAB, SantosACC, SouzaFHD, VallsJFM, PagliariniMS (2014). Complete asynapsis resulting in 2n pollen formation in Paspalum jesuiticum Parodi (Poaceae) Genet Mol Res 13, 255-261. DOI:10.4238/2014.January.17.9URL [本文引用: 2]
GaoLH, DiarsoM, ZhangA, ZhangHK, DongYZ, LiuLX, LvZL, LiuB (2016). Heritable alteration of DNA methylation induced by whole-chromosome aneuploidy in wheat New Phytol 209, 364-375. DOI:10.1111/nph.2016.209.issue-1URL [本文引用: 1]
GhimireBK, SeongES, NguyenTX, YooJH, YuCY, KimSH, ChungI (2016). Assessment of morphological and phytochemical attributes in triploid and hexaploid plants of the bioenergy crop Miscanthus × giganteus Ind Crop Prod 89, 231-243. DOI:10.1016/j.indcrop.2016.04.051URL [本文引用: 1]
HagerupO (1947). The spontaneous formation of haploid, polyploid, and aneuploid embryos in some orchids Biol Meddel Kongol Danske Vidensk Selsk 20, 1-22. [本文引用: 1]
HannaWW, SchertzKF (1971). Trisome identification in Sorghum bicolor( L.) Moench by observing progeny of triploid × translocation stocks Can J Genet Cytol 13, 105-109. DOI:10.1139/g71-016URL [本文引用: 1]
HarlanJR, DeWetJMJ (1975). On ?. Winge and a prayer: the origins of polyploidy Bot Rev 41, 361-390. DOI:10.1007/BF02860830URL
HenryIM, DilkesBP, MillerES, Burkart-WacoD, ComaiL (2010). Phenotypic consequences of aneuploidy in Arabidopsis thaliana Genetics 186, 1231-1245. DOI:10.1534/genetics.110.121079 [本文引用: 1] Aneuploid cells are characterized by incomplete chromosome sets. The resulting imbalance in gene dosage has phenotypic consequences that are specific to each karyotype. Even in the case of Down syndrome, the most viable and studied form of human aneuploidy, the mechanisms underlying the connected phenotypes remain mostly unclear. Because of their tolerance to aneuploidy, plants provide a powerful system for a genome-wide investigation of aneuploid syndromes, an approach that is not feasible in animal systems. Indeed, in many plant species, populations of aneuploid individuals can be easily obtained from triploid individuals. We phenotyped a population of Arabidopsis thaliana aneuploid individuals containing 25 different karyotypes. Even in this highly heterogeneous population, we demonstrate that certain traits are strongly associated with the dosage of specific chromosome types and that chromosomal effects can be additive. Further, we identified subtle developmental phenotypes expressed in the diploid progeny of aneuploid parent(s) but not in euploid controls from diploid lineages. These results indicate long-term phenotypic consequences of aneuploidy that can persist after chromosomal balance has been restored. We verified the diploid nature of these individuals by whole-genome sequencing and discuss the possibility that trans-generational phenotypic effects stem from epigenetic modifications passed from aneuploid parents to their diploid progeny.
HoshinoY, MiyashitaT, ThomasTD (2011). In vitro culture of endosperm and its application in plant breeding: approaches to polyploidy breeding Sci Hortic 130, 1-8. DOI:10.1016/j.scienta.2011.06.041URL [本文引用: 3]
HuFR, ZhangL, WangXY, DingJ, WuDX (2005). Agrobacterium-mediated transformed transgenic triploid bermudagrass ( Cynodon dactylon × C. transvaalensis) plants are highly resistant to the glufosinate herbicide liberty Plant Cell Tissue Organ Cult 83, 13-19. DOI:10.1007/s11240-005-3705-1URL [本文引用: 1]
IwataN, OmuraT (1975). Studies on the trisomics in rice plants (Oryza sativa L.): III. Relation between trisomics and genetic linkage groups Jpn J Breed 25, 363-368. DOI:10.1270/jsbbs1951.25.363URL [本文引用: 1]
JansenRC, Den NijsAPM (1993). A statistical mixture model for estimating the proportion of unreduced pollen grains in perennial ryegrass (Lolium perenne L.) via the size of pollen grains Euphytica 70, 205-215. DOI:10.1007/BF00023761URL [本文引用: 1]
JohansenB, Von BothmerR (1994). Pollen size in Hordeum L.: correlation between size, ploidy level, and breeding system Sex Plant Reprod 7, 259-263. DOI:10.1007/BF00227707URL [本文引用: 2]
JohriBM, BhojwaniSS (1965). Growth responses of mature endosperm in cultures Nature 208, 1345-1347. DOI:10.1038/2081345a0URL [本文引用: 1]
KampsTL, WilliamsNR, OrtegaVM, ChamuscoKC, Harris-ShultzK, ScullyBT, ChaseCD (2011). DNA polymorphisms at bermudagrass microsatellite loci and their use in genotype fingerprinting Crop Sci 51, 1122-1131. DOI:10.2135/cropsci2010.08.0478URL [本文引用: 2]
KapadiaZJ, GouldFW (1964). Biosystematic studies in the Bouteloua curtipendula complex. III. Pollen size as related to chromosome numbers Am J Bot 51, 166-172. DOI:10.1002/ajb2.1964.51.issue-2URL [本文引用: 1]
KatoA (1999). Induction of bicellular pollen by trifluralin treatment and occurrence of triploids and aneuploids after fertilization in maize Genome 42, 154-157. DOI:10.1139/g98-108URL [本文引用: 1]
KatoA, BirchlerJA (2006). Induction of tetraploid derivatives of maize inbred lines by nitrous oxide gas treatment J Hered 97, 39-44. DOI:10.1093/jhered/esj007URL [本文引用: 1]
KatsiotisA, ForsbergRA (1995). Pollen grain size in four ploidy levels of genus Avena Euphytica 83, 103-108. DOI:10.1007/BF01678036URL [本文引用: 1]
KellerB, FeuilletC (2000). Colinearity and gene density in grass genomes Trends Plant Sci 5, 246-251. PMID:10838615 [本文引用: 1] Grasses are the single most important plant family in agriculture. In the past years, comparative genetic mapping has revealed conserved gene order (colinearity) among many grass species. Recently, the first studies at gene level have demonstrated that microcolinearity of genes is less conserved: small scale rearrangements and deletions complicate the microcolinearity between closely related species, such as sorghum and maize, but also between rice and other crop plants. In spite of these problems, rice remains the model plant for grasses as there is limited useful colinearity between Arabidopsis and grasses. However, studies in rice have to be complemented by more intensive genetic work on grass species with large genomes (maize, Triticeae). Gene-rich chromosomal regions in species with large genomes, such as wheat, have a high gene density and are ideal targets for partial genome sequencing.
KindigerB, DewaldC (1994). Genome accumulation in eastern gamagrass, Tripsacum dactyloides(L.) L. 92, 197-201. [本文引用: 1]
KingIP, MorganWG, HarperJA, ThomasHM (1999). Introgression mapping in the grasses. II. Meiotic analysis of the Lolium perenne/Festuca pratensis triploid hybrid Heredity 82, 107-112. DOI:10.1038/sj.hdy.6884680URL [本文引用: 1]
KirovI, DivashukM, Van LaereK, SolovievA, KhrustalevaL (2014). An easy "SteamDrop" method for high quality plant chromosome preparation Mol Cytogenet 7, 21. DOI:10.1186/1755-8166-7-21PMID:24602284 [本文引用: 1] Background: The chromosome preparation is a crucial step for obtaining satisfactory results in molecular cytogenetic researches. The preparation of plant chromosomes for molecular cytogenetic purposes remains a challenge for some species. In contrast to human chromosome preparation, the processes occurring during plant chromosome preparation and causing chromosome spreading are still poorly understood. Results: We studied the dynamics of plant chromosome spreading after dropping cell suspension on slides. We showed that steam stimulates cytoplasm hydrolysis and rapid chromosome spreading and that chromosomes stretch during this chromosome spreading. Based on these observations, we developed a novel method, named "SteamDrop", for the preparation of well-spread mitotic and pachytene chromosomes and successfully used it for 28 plant species with large and small chromosomes. We applied cell suspensions in ethanol instead of the commonly used ethanol/acetic acid fixative. Mitotic and meiotic chromosomes prepared via "SteamDrop" were used in fluorescent in situ hybridization (FISH) experiments with repetitive and unique DNA probes. Long storage of cell suspensions in ethanol did not impair the quality of chromosome preparations. Conclusion: The SteamDrop procedure provides a robust and routine method for high quality plant chromosome preparations. The method can be applied for metaphase as well as pachytene chromosome preparation in wide range of species. The chromosomes prepared by SteamDrop are well suitable for repetitive and unique DNA visualization.
KreinerJM, KronP, HusbandBC (2017). Frequency and maintenance of unreduced gametes in natural plant populations: associations with reproductive mode, life history and genome size New Phytol 214, 879-889. DOI:10.1111/nph.2017.214.issue-2URL [本文引用: 1]
KronP, HusbandBC (2015). Distinguishing 2N gamete nuclei from doublets in pollen using flow cytometry and pulse analysis Cytometry Part A 87, 943-957. DOI:10.1002/cyto.a.22696URL [本文引用: 2]
La RueCD (1949). Cultures of the endosperm of maize Am J Bot 36, 798. [本文引用: 1]
LamoteV, BaertJ, Roldán-RuizI, De LooseM, Van BockstaeleE (2002). Tracing of 2n egg occurrence in perennial ryegrass (Lolium perenne L.) using interploidy crosses Euphytica 123, 159-164. DOI:10.1023/A:1014980123519URL [本文引用: 1]
LimKB, RamannaMS, De JongJH, JacobsenE, Van TuylJM (2001). Indeterminate meiotic restitution (IMR): a novel type of meiotic nuclear restitution mechanism detected in interspecific lily hybrids by GISH Theor Appl Genet 103, 219-230. DOI:10.1007/s001220100638URL [本文引用: 2]
LoginovaDB, SilkovaOG (2017). Mechanisms of unreduced gamete formation in flowering plants Russ J Genet 53, 741-756. DOI:10.1134/S1022795417070080URL [本文引用: 1]
LuSY, PengXX, GuoZF, ZhangGY, WangZC, WangCY, PangCS, FanZ, WangJH (2007). In vitro selection of salinity tolerant variants from triploid bermudagrass ( Cynodon transvaalensis × C. dactylon) and their physiological responses to salt and drought stress Plant Cell Rep 26, 1413-1420. DOI:10.1007/s00299-007-0339-2URL [本文引用: 1]
MaceiraNO, De HaanAA, LumaretR, BillonM, DelayJ (1992). Production of 2 n gametes in diploid subspecies of Dactylis glomerata L. 1. Occurrence and frequency of 2n pollen Ann Bot 69, 335-343. DOI:10.1093/oxfordjournals.aob.a088350URL [本文引用: 1]
MasonAS, NelsonMN, YanGJ, CowlingWA (2011). Production of viable male unreduced gametes in Brassica interspecific hybrids is genotype specific and stimulated by cold temperatures BMC Plant Biol 11, 103. DOI:10.1186/1471-2229-11-103URL [本文引用: 1]
MultaniDS, JenaKK, BrarDS, De Los ReyesBG, AngelesER, KhushGS (1994). Development of monosomic alien addition lines and introgression of genes from Oryza australiensis Domin. to cultivated rice O sativa L. Theor Appl Genet 88, 102-109. [本文引用: 2]
NaganowskaB, ZwierzykowskiZ, ZwierzykowskaE (2001). Meiosis and fertility of reciprocal triploid hybrids of Lolium multiflorum with Festuca pratensis J Appl Genet 42, 247-255. PMID:14564031 [本文引用: 1] Diploid and tetraploid forms of Lolium multiflorum and Festuca pratensis were crossed under controlled conditions and after embryo rescue all four combinations of autoallotriploid hybrids were obtained. Male and female fertility and chromosome pairing at metaphase I of meiosis were studied in several plants from each hybrid combination. The hybrids with two genomes of L. multiflorum and one of F. pratensis (genomic formulae LmLmFp and FpLmLm) were male and female fertile while the hybrids with two genomes of F. pratensis and one of L. multiflorum had a reduced fertility (FpFpLm) or were completely sterile (LmFpFp). Chromosome pairing at metaphase I varied among hybrid combinations depending on their genomic composition. LmLmFp and FpLmLm hybrids had similar patterns of pairing (1.83I + 5.29II + 2.85III and 2.22I + 5.22II + 2.75III, respectively) but they differed from those of FpFpLm (3.65I + 4.65II + 2.68III) and especially from LmFpFp (4.78I + 5.87II + 1.49III). Conventional analysis of meiosis failed to explain the differences in chromosome behaviour and fertility/sterility levels between the autoallotriploid hybrids with two Lolium or two Festuca genomes.
NakanoH, TashiroT, MaedaE (1975). Plant differentiation in callus tissue induced from immature endosperm of Oryza sauva L Z Pflanzenphysiol 76, 444-449. DOI:10.1016/S0044-328X(75)80006-7URL [本文引用: 2]
NakelT, TekleyohansDG, MaoYB, FuchertG, VoD, Gro?-HardtR (2017). Triparental plants provide direct evidence for polyspermy induced polyploidy Nat Commun 8, 1033. DOI:10.1038/s41467-017-01044-yURL [本文引用: 4]
NorstogK, WallWE, HowlandGP (1969). Cytological characteristics of ten-year-old rye-grass endosperm tissue cultures Bot Gaz 130, 83-86. DOI:10.1086/336473URL [本文引用: 3]
NorstogKJ (1956). Growth of rye-grass endosperm in vitro Bot Gaz 117, 253-259. DOI:10.1086/335914URL [本文引用: 2]
NunesJD, AzevedoALS, PereiraAV, PaulaCMP, CamposJMS, LédoFJS, SantosVB (2013). DNA elimination in embryogenic development of Pennisetum glaucum × Pennisetum purpureum(Poaceae) hybrids Genet Mol Res 12, 4817-4826. DOI:10.4238/2013.October.22.1PMID:24301743 [本文引用: 1] Interspecific hybridization between Napier grass (Pennisetum purpureum), which is widely grown in Brazil for cattle forage, and pearl millet (Pennisetum glaucum) has been used as a breeding strategy for the development of improved cultivars. However, the hybrid between these two species is sterile due to its triploid condition (2n = 3x = 21 chromosomes), which hinders its use in crop breeding programs. It is known that genomic alterations result from the hybridization process. In order to measure the loss of DNA during embryo development, we used flow cytometry to estimate the nuclear DNA content of triploid and tetraploid embryos produced by interspecific hybridization between Napier grass and pearl millet. The triploid and tetraploid hybrids had a mean DNA content of 4.99-4.87 and 5.25-4.84 pg, at 10 and 30 days after pollination, respectively. The mean reduction in DNA content was higher in the tetraploid hybrids. The flow cytometry results revealed progressive genomic instability in these triploid and tetraploid hybrids, with this instability causing significant alterations in the DNA content of the hybrids.
PagliariniMS, ValleCB, SantosEM, MendesDV, BernardoZH, Mendes-BonatoAB, SilvaN, CalistoV (2012). Microsporogenesis in Brachiaria brizantha(Poaceae) as a selection tool for breeding Genet Mol Res 11, 1309-1318. DOI:10.4238/2012.May.15.1PMID:22653577 [本文引用: 1] The genus Brachiaria comprises more than 100 species and is the single most important genus of forage grass in the tropics. Brachiaria brizantha, widely used in Brazilian pastures for beef and dairy production, is native to tropical Africa. As a subsidy to the breeding program underway in Brazil, cytological studies were employed to determine the chromosome number and to evaluate microsporogenesis in 46 accessions of this species available at Embrapa Beef Cattle (Brazil). Thirty-four accessions presented 2n = 36; seven had 2n = 45, and five had 2n = 54 chromosomes. Based on the higher level of chromosome association observed in diakinesis, in tetra-, penta-, and hexavalents, respectively, it was concluded that they are derived from x = 9; consequently, these accessions are tetra- (2n = 4x = 36), penta- (2n = 5x = 45), and hexaploids (2n = 6x = 54). The most common meiotic abnormalities were irregular chromosome segregation due to polyploidy. Chromosome stickiness, abnormal cytokinesis, non-congressed bivalents in metaphase I and chromosomes in metaphase II, and chromosome elimination were recorded at varying frequencies in several accessions. The mean percentage of meiotic abnormalities ranged from 0.36 to 95.76%. All the abnormalities had the potential to affect pollen viability by generating unbalanced gametes. Among the accessions, only the tetraploid ones with less than 40% of abnormalities are suitable as pollen donors in intra- and interspecific crosses. Currently, accessions with a high level of ploidy (5 and 6n) cannot be used as male genitors in crosses because of the lack of sexual female genitors with the same levels of ploidy.
PécrixY, RalloG, FolzerH, CignaM, GudinS, Le BrisM (2011). Polyploidization mechanisms: temperature environment can induce diploid gamete formation in Rosa sp J Exp Bot 62, 3587-3597. DOI:10.1093/jxb/err052URL [本文引用: 1]
PereraD, BarnesDJ, BaldwinBS, ReichertNA (2015). Direct and indirect in vitro regeneration of Miscanthus × giganteus cultivar freedom: effects of explant type and medium on regeneration efficiency In Vitro Cell Dev Biol Plant 51, 294-302. DOI:10.1007/s11627-015-9682-0URL [本文引用: 1]
RamseyJ, SchemskeDW (1998). Pathways, mechanisms, and rates of polyploid formation in flowering plants Annu Rev Ecol Syst 29, 467-501. DOI:10.1146/annurev.ecolsys.29.1.467URL [本文引用: 2]
SchwartzBM, Harris-ShultzKR, ContrerasRN, HansCS, HannaWW, Milla-LewisSR (2013). Creation of artificial triploid and tetraploid centipedegrass using colchicine and breeding Int Turfgrass Soc Res J 12, 327-334. [本文引用: 2]
SehgalCB (1974). Growth of barley and wheat endosperm in cultures Curr Sci 43, 38-40. [本文引用: 1]
SheidaiM, JafariS, TalebanP, KeshavarziM (2009). Cytomixis and unreduced pollen grain formation in Alopecurus L. and Catbrosa Beauv.(Poaceae) Cytologia 74, 31-41. DOI:10.1508/cytologia.74.31URL [本文引用: 1]
SuarezEY, LopezAG, NaranjoCA (1992). Polyspermy versus unreduced male gametes as the origin of nonaploids (9x) common wheat plants Caryologia 45, 21-28. DOI:10.1080/00087114.1992.10797206URL [本文引用: 1]
SunS, WuY, LinXY, WangJ, YuJM, SunY, MiaoYL, LiQP, SanguinetKA, LiuB (2017). Hybrid weakness in a rice interspecific hybrid is nitrogen-dependent, and accompanied by changes in gene expression at both total transcript level and parental allele partitioning PLoS One 12, e0172919. DOI:10.1371/journal.pone.0172919URL [本文引用: 1]
TamaokiT, UllstrupJA (1958). Cultivation in vitro of excised endosperm and meristem tissues of corn Bull Torrey Bot Club 85, 260-272. DOI:10.2307/2482581URL [本文引用: 1]
ThomasH, MorganWG, HumphreysMW (1988). The use of a triploid hybrid for introgression in Lolium species Theor Appl Genet 76, 299-304. DOI:10.1007/BF00257859PMID:24232119 [本文引用: 1] Triploid hybrids of Lolium multiflorum (4x) x L. perenne (2x) behaved cytologically as autotriploids but the segregation of isozyme variants did not always agree with the expected trisomic ratios. The overall effect of these deviations from expectation was a greater proportion than expected of diploid progeny from the cross L. multiflorum (2x) x triploid hybrid which did not include any of the L. perenne alleles at the three marker isozyme loci. The possible mechanisms for these aberrant segregation ratios are discussed together with the advantages of the crossing scheme to accelerate the recovery of the genotype of the recurrent parent in a backcrossing programme to transfer characters from one species to another.
ThomasTD, ChaturvediR (2008). Endosperm culture: a novel method for triploid plant production Plant Cell Tissue Organ Cult 93, 1-14. DOI:10.1007/s11240-008-9336-6URL [本文引用: 4]
TodaE, OhnishiY, OkamotoT (2016). Development of polyspermic rice zygotes Plant Physiol 171, 206-214. DOI:10.1104/pp.15.01953URL [本文引用: 2]
TodaE, OkamotoT (2016). Formation of triploid plants via possible polyspermy Plant Signal Behav 11, e1218107. DOI:10.1080/15592324.2016.1218107URL
Van SantenE, HugessenPM, CaslerMD (1991). Identification and frequency of tetraploid progeny from 2 x-4x and 4x-2x crosses in Dactylis Genome 34, 273-278. DOI:10.1139/g91-043URL [本文引用: 1]
WangC, LiuQ, ShenY, HuaYF, WangJJ, LinJR, WuMG, SunTT, ChengZK, MercierR, WangKJ (2019). Clonal seeds from hybrid rice by simultaneous genome engineering of meiosis and fertilization genes Nat Biotechnol 37, 283-286. DOI:10.1038/s41587-018-0003-0URL [本文引用: 1]
WangX, YamadaT, KongFJ, AbeY, HoshinoY, SatoH, TakamizoT, KanazawaA, YamadaT (2011). Establishment of an efficient in vitro culture and particle bombardment-mediated transformation systems in Miscanthus sinensis Anderss, a potential bioenergy crop GCB Bioenergy 3, 322-332. DOI:10.1111/gcbb.2011.3.issue-4URL [本文引用: 1]
YounisA, HwangYJ, LimKB (2014). Exploitation of induced 2 n-gametes for plant breeding Plant Cell Rep 33, 215-223. DOI:10.1007/s00299-013-1534-yURL [本文引用: 2]
ZengRZ, ZhuJ, XuSY, DuGH, GuoHR, ChenJJ, ZhangSZ, XieL (2020). Unreduced male gamete formation in Cymbidium and its use for developing sexual polyploid cultivars Front Plant Sci 11, 558. DOI:10.3389/fpls.2020.00558URL [本文引用: 1]
... 胚乳发育阶段是胚乳培养的关键因素, 常用授粉后天数(days after pollination, DAP)来表征.通常以未成熟胚乳作外植体, 如多年生黑麦草在9-10 DAP (Norstog, 1956)、玉米在8-11 DAP (Straus and La Rue, 1954; Tamaoki and Ullstrup, 1958)、水稻在4-7 DAP (Nakano et al., 1975)、小麦在10 DAP (赵世绪等, 1984)以及大麦在8 DAP (Sehgal, 1974); 小麦-黑麦杂交胚乳的愈伤组织在14 DAP时诱导频率最佳, 在7 DAP时次之, 而在21 DAP时已不能诱导出愈伤组织(王敬驹等, 1982).以毛竹30-40 DAP的成熟胚乳作为外植体获得成功(袁金玲等, 2015).一般情况下, 成熟胚乳启动细胞分裂在于初始时与胚的联动, 而未成熟胚乳启动细胞分裂与胚是否存在无关(Hoshino et al., 2011).需要特别指出的是, 添加赤霉素等外源激素可以代替胚, 而小麦和黑麦的胚乳在无胚和赤霉素的条件下也可产生愈伤组织并分化成苗(赵世绪等, 1984).培养基、生长调节剂和环境因素对于胚乳三倍体的形成也起着重要作用.例如, 培养基中添加8%的蔗糖有利于小麦-黑麦杂交胚乳的愈伤组织诱导(王敬驹等, 1982).添加氮源对于未成熟胚乳的增殖至关重要(Hoshino et al., 2011).在小麦和黑麦胚乳培养时, 添加激动素和2,4-D可提高愈伤组织诱导频率(赵世绪等, 1984).玉米胚乳在暗处培养效果更佳(Straus and La Rue, 1954), 而在黑麦中光照对胚乳的增殖无影响(Norstog et al.,1969).胚乳培养以组织培养为技术基础, 并未成为主流的三倍体合成方式. ...
... ).培养基、生长调节剂和环境因素对于胚乳三倍体的形成也起着重要作用.例如, 培养基中添加8%的蔗糖有利于小麦-黑麦杂交胚乳的愈伤组织诱导(王敬驹等, 1982).添加氮源对于未成熟胚乳的增殖至关重要(Hoshino et al., 2011).在小麦和黑麦胚乳培养时, 添加激动素和2,4-D可提高愈伤组织诱导频率(赵世绪等, 1984).玉米胚乳在暗处培养效果更佳(Straus and La Rue, 1954), 而在黑麦中光照对胚乳的增殖无影响(Norstog et al.,1969).胚乳培养以组织培养为技术基础, 并未成为主流的三倍体合成方式. ...
... 胚乳发育阶段是胚乳培养的关键因素, 常用授粉后天数(days after pollination, DAP)来表征.通常以未成熟胚乳作外植体, 如多年生黑麦草在9-10 DAP (Norstog, 1956)、玉米在8-11 DAP (Straus and La Rue, 1954; Tamaoki and Ullstrup, 1958)、水稻在4-7 DAP (Nakano et al., 1975)、小麦在10 DAP (赵世绪等, 1984)以及大麦在8 DAP (Sehgal, 1974); 小麦-黑麦杂交胚乳的愈伤组织在14 DAP时诱导频率最佳, 在7 DAP时次之, 而在21 DAP时已不能诱导出愈伤组织(王敬驹等, 1982).以毛竹30-40 DAP的成熟胚乳作为外植体获得成功(袁金玲等, 2015).一般情况下, 成熟胚乳启动细胞分裂在于初始时与胚的联动, 而未成熟胚乳启动细胞分裂与胚是否存在无关(Hoshino et al., 2011).需要特别指出的是, 添加赤霉素等外源激素可以代替胚, 而小麦和黑麦的胚乳在无胚和赤霉素的条件下也可产生愈伤组织并分化成苗(赵世绪等, 1984).培养基、生长调节剂和环境因素对于胚乳三倍体的形成也起着重要作用.例如, 培养基中添加8%的蔗糖有利于小麦-黑麦杂交胚乳的愈伤组织诱导(王敬驹等, 1982).添加氮源对于未成熟胚乳的增殖至关重要(Hoshino et al., 2011).在小麦和黑麦胚乳培养时, 添加激动素和2,4-D可提高愈伤组织诱导频率(赵世绪等, 1984).玉米胚乳在暗处培养效果更佳(Straus and La Rue, 1954), 而在黑麦中光照对胚乳的增殖无影响(Norstog et al.,1969).胚乳培养以组织培养为技术基础, 并未成为主流的三倍体合成方式. ...
... ).需要特别指出的是, 添加赤霉素等外源激素可以代替胚, 而小麦和黑麦的胚乳在无胚和赤霉素的条件下也可产生愈伤组织并分化成苗(赵世绪等, 1984).培养基、生长调节剂和环境因素对于胚乳三倍体的形成也起着重要作用.例如, 培养基中添加8%的蔗糖有利于小麦-黑麦杂交胚乳的愈伤组织诱导(王敬驹等, 1982).添加氮源对于未成熟胚乳的增殖至关重要(Hoshino et al., 2011).在小麦和黑麦胚乳培养时, 添加激动素和2,4-D可提高愈伤组织诱导频率(赵世绪等, 1984).玉米胚乳在暗处培养效果更佳(Straus and La Rue, 1954), 而在黑麦中光照对胚乳的增殖无影响(Norstog et al.,1969).胚乳培养以组织培养为技术基础, 并未成为主流的三倍体合成方式. ...
... ).在小麦和黑麦胚乳培养时, 添加激动素和2,4-D可提高愈伤组织诱导频率(赵世绪等, 1984).玉米胚乳在暗处培养效果更佳(Straus and La Rue, 1954), 而在黑麦中光照对胚乳的增殖无影响(Norstog et al.,1969).胚乳培养以组织培养为技术基础, 并未成为主流的三倍体合成方式. ...
Postmeiotic restitution in 2 n-egg formation of diploid potato 2 1998
... (A) 与体细胞相比正常配子的染色体数目减半; (B) 通过胞质融合或多核体形成的四倍体花粉母细胞产生PRD型2n配子, 如鸭茅(Falistocco et al., 1995); (C) 在减数分裂I期同源染色体无配对和分裂形成的FDR型2n配子将保留除重组片段外的所有亲本基因, 如雀稗(Filho et al., 2014)和多年生黑麦草(Chen et al., 1997); (D) 在减数分裂II期因分裂缺失形成的SDR型2n配子将保留除重组片段外比正常配子多1套染色体副本的2n配子, 如多年生黑麦草(Chen et al., 1997); (E) 在减数分裂I期部分染色体以姐妹染色单体分离, 其余以二价体分离, 随后在减数分裂II期分裂缺失形成IMR型2n配子, 如百合(Lim et al., 2001); (F) 减数分裂正常而进入配子体发育阶段时基因组加倍形成PMD型2n配子, 如马铃薯(Bastiaanssen et al., 1998) ...
... (A) Chromosome numbers of normal gametes are halved compared with somatic cells; (B) PRD (pre-meiotic doubling) gametes are obtained as a result of the tetraploid pollen mother cells formed by cytomixis or syncytium, e.g., Dactylis glomerata (Falistocco et al., 1995); (C) In the FDR (first division restitution) type, pairing and division of homoeologous chromosomes do not occur during meiosis I, and the FDR 2n gametes maintain all of their parental genes except cross-over fragments, e.g., Paspalum jesuiticum (Filho et al., 2014) and Lolium perenne (Chen et al., 1997); (D) SDR (second division restitution) gametes with two copies of non-recombinant chromosomes are the result of the second division omission or cytokinesis abnormalities after normal anaphase II, e.g., L. perenne (Chen et al., 1997); (E) IMR (indeterminate meiotic restitution) gametes were discovered in Lilium longiflorum × Asiatic hybrid (Lim et al., 2001), in which some chromosomes are separated as univalents during meiosis I whereas the others are separated as bivalents, and subsequently the second division omission occurs during meiosis II; (F) PMD (post-meiotic doubling) gametes occurs due to the genome doubling of haploid spores during microgametogenesis, e.g., Solanum tuberosum (Bastiaanssen et al., 1998) ...
Gamete fusion is required to block multiple pollen tubes from entering an Arabidopsis ovule 1 2012
Unreduced gamete formation in plants: mechanisms and prospects 1 2011
... 实践中, 2n配子可通过环境调控、化学诱导、种间杂种和减数分裂突变体获得.2n配子的发生主要受遗传控制, 但暴露于低温或高温、热冲击和高辐射下可诱导或刺激2n配子形成(Younis et al., 2014).例如, 蔷薇属(Rosa)植物花芽暴露于36°C高温下可提高2n雄配子的发生率(Pécrix et al., 2011).在禾本科中采用环境调控诱导2n配子发生尚未见报道.N2O在诱导鸭茅(Van Santen et al., 1991)、玉米(Zea mays, 2n=2x=20) (Kato and Birchler, 2006)、小麦(Triticum aestivum, 2n=6x=42)及大麦(Dvorak et al., 1973)的2n配子上取得成功.向玉米雄穗喷洒氟乐灵可诱导2n花粉并获得同源三倍体(Kato, 1999).种间或属间杂种是产生2n配子的重要途径, 其频率高于祖先物种(Ramsey and Schemske, 1998).Mason等(2011)分析了3种芸薹属(Brassica)物种及其F1杂种的2n雄配子发生率, 发现5个甘蓝型油菜(B. napus, 2n=4x=38)品系的发生率(0%-0.11%, 平均值为0.03%)较低, 而甘蓝型油菜的11种F1杂种的发生率(0.17%-3.29%, 平均值为1.55%)较高.可见, 种间杂种较亲本基因组2n配子的发生率提高.Zeng等(2020)发现在兰花(Cymbidium ssp.)传统品种中2n雄配子发生率为0.15%-1.0%, 而种间杂种为2.5%-4.03%; 同时作者认为可能由于无性繁殖植物存在可替代的繁殖途径, 2n配子的形成频率相对更高.减数分裂活跃基因的突变可能导致纺锤体形成异常或胞质分裂受阻而形成2n配子, 如拟南芥(Arabidopsis thaliana, 2n= 2x=10)中的cyca1;2/tam (SDR型)、osd1 (SDR型)和Atps1 (FDR型)突变体(Brownfield and K?hler, 2011). ...
In vitro induction of hexaploid plants from triploid hybrids of Pennisetum purpureum and Pennisetum glaucum 1 2009
... 三倍体可作为通往其它倍性水平的桥梁, 即三倍体桥(triploid-bridge) (图3C).在鸭茅三倍体后代中发现了四倍体、五倍体以及混倍体植株(钟声, 2006).三倍体摩擦禾可产生2n雌配子, 通过3x/2x合成了四倍体(Kindiger and Dewald, 1994).以三倍体杂交狼尾草(美洲狼尾草/象草, 2n=3x=21)的幼穗离体培养获得的胚性愈伤组织为材料, 通过秋水仙碱诱导获得六倍体(钟小仙等, 2014).利用秋水仙碱处理象草(2n=4x=28)/珍珠粟(P. glaucum, 2n=2x=14)杂交三倍体幼苗, 体细胞加倍后形成异源六倍体(Campos et al., 2009).用类似的方法获得M. × miscanthus六倍体(Yu et al., 2009; Ghimire et al., 2016).水稻(AA)和高秆野生稻(O. alta, CCDD)杂交形成异源三倍体, 加倍后形成三基异源六倍体(AACCDD) (祝剑峰等, 2008). ...
Registration of ‘DALSA 0605’ St. Augustinegrass 1 2015
... 对于有直接利用价值的三倍体, 可通过组培、诱变和转基因等技术进一步改良(图3B).MS-Supreme源于Tifgreen的自然突变, 与Tifgreen相比其叶片和茎节节间缩小、变短(Krans et al., 1999).在Tifgreen的自然突变中还选育出Tifdwarf和FloraDwarf等矮生品种; TifwayII、TifSport和TifEagle则由相应三倍体亲本经γ辐射诱变而成(Kamps et al., 2011).利用体细胞无性系变异育种技术在TifEagle的愈伤组织中长时间连续继代培养能有效诱导体细胞变异(如矮化突变体); 以0.3 mol·L-1 NaCl为选择压筛选出耐盐细胞系(Lu et al., 2007).以美洲狼尾草(Pennisetum americana)/ 四倍体象草合成的三倍体F1种子为诱变材料, 经γ辐射获得优异变异株闽牧6号(陈钟佃等, 2012).华南1号杂交狼尾草由三倍体杂交狼尾草芽变而来(陈平等, 2004).三倍体也可以直接作为亲本利用.在偏序钝叶草(Stenotaphrum secundatum, 2n=2x=18)的3x/ 2x杂交后代中经胚拯救选育出抗灰斑病(由稻瘟病菌(Magnaporthe grisea)引起)和耐旱草坪草新品种(Chandra et al., 2015).奇岗天然后代群体中16个数量性状出现了丰富的变异(变异系数为14.41%- 151.85%), 在后代中筛选出3份材料, 其生物量优于奇岗(潘志军等, 2017).通过三倍体狼尾草与摩特矮象草杂交选育出中国南方地区广泛种植的三倍体多年生牧草桂牧1号(罗宗志等, 2016).宁牧26-2狼尾草也是由三倍体后代衍生而来(顾洪如等, 1992).三倍体转基因品系可通过2种策略获得.(1) 直接以三倍体为受体, 导入目的基因构建转基因品系, 如三倍体狗牙根Tifeagle (Hu et al., 2005).(2) 对于愈伤组织形成与再分化率低的三倍体, 首先需要寻找愈伤组织再分化优良的二倍体或四倍体作为转基因桥梁, 然后通过基因枪或农杆菌转化等方法导入目的基因, 最后通过倍性间杂交构建转基因三倍体, 如芒(2n=2x=38) (Wang et al., 2011).近年来, 基因编辑技术的发展为定向改良三倍体提供了新的育种方案. ...
RFLP detection of 2 n pollen formation by first and second division restitution in perennial ryegrass 5 1997
... (A) 与体细胞相比正常配子的染色体数目减半; (B) 通过胞质融合或多核体形成的四倍体花粉母细胞产生PRD型2n配子, 如鸭茅(Falistocco et al., 1995); (C) 在减数分裂I期同源染色体无配对和分裂形成的FDR型2n配子将保留除重组片段外的所有亲本基因, 如雀稗(Filho et al., 2014)和多年生黑麦草(Chen et al., 1997); (D) 在减数分裂II期因分裂缺失形成的SDR型2n配子将保留除重组片段外比正常配子多1套染色体副本的2n配子, 如多年生黑麦草(Chen et al., 1997); (E) 在减数分裂I期部分染色体以姐妹染色单体分离, 其余以二价体分离, 随后在减数分裂II期分裂缺失形成IMR型2n配子, 如百合(Lim et al., 2001); (F) 减数分裂正常而进入配子体发育阶段时基因组加倍形成PMD型2n配子, 如马铃薯(Bastiaanssen et al., 1998) ...
... 配子, 如多年生黑麦草(Chen et al., 1997); (E) 在减数分裂I期部分染色体以姐妹染色单体分离, 其余以二价体分离, 随后在减数分裂II期分裂缺失形成IMR型2n配子, 如百合(Lim et al., 2001); (F) 减数分裂正常而进入配子体发育阶段时基因组加倍形成PMD型2n配子, 如马铃薯(Bastiaanssen et al., 1998) ...
... (A) Chromosome numbers of normal gametes are halved compared with somatic cells; (B) PRD (pre-meiotic doubling) gametes are obtained as a result of the tetraploid pollen mother cells formed by cytomixis or syncytium, e.g., Dactylis glomerata (Falistocco et al., 1995); (C) In the FDR (first division restitution) type, pairing and division of homoeologous chromosomes do not occur during meiosis I, and the FDR 2n gametes maintain all of their parental genes except cross-over fragments, e.g., Paspalum jesuiticum (Filho et al., 2014) and Lolium perenne (Chen et al., 1997); (D) SDR (second division restitution) gametes with two copies of non-recombinant chromosomes are the result of the second division omission or cytokinesis abnormalities after normal anaphase II, e.g., L. perenne (Chen et al., 1997); (E) IMR (indeterminate meiotic restitution) gametes were discovered in Lilium longiflorum × Asiatic hybrid (Lim et al., 2001), in which some chromosomes are separated as univalents during meiosis I whereas the others are separated as bivalents, and subsequently the second division omission occurs during meiosis II; (F) PMD (post-meiotic doubling) gametes occurs due to the genome doubling of haploid spores during microgametogenesis, e.g., Solanum tuberosum (Bastiaanssen et al., 1998) ...
... (Chen et al., 1997); (E) IMR (indeterminate meiotic restitution) gametes were discovered in Lilium longiflorum × Asiatic hybrid (Lim et al., 2001), in which some chromosomes are separated as univalents during meiosis I whereas the others are separated as bivalents, and subsequently the second division omission occurs during meiosis II; (F) PMD (post-meiotic doubling) gametes occurs due to the genome doubling of haploid spores during microgametogenesis, e.g., Solanum tuberosum (Bastiaanssen et al., 1998) ...
Cytomixis in pollen mother cells of diploid Dactylis, one of the origins of 2n gametes 2 1995
... (A) 与体细胞相比正常配子的染色体数目减半; (B) 通过胞质融合或多核体形成的四倍体花粉母细胞产生PRD型2n配子, 如鸭茅(Falistocco et al., 1995); (C) 在减数分裂I期同源染色体无配对和分裂形成的FDR型2n配子将保留除重组片段外的所有亲本基因, 如雀稗(Filho et al., 2014)和多年生黑麦草(Chen et al., 1997); (D) 在减数分裂II期因分裂缺失形成的SDR型2n配子将保留除重组片段外比正常配子多1套染色体副本的2n配子, 如多年生黑麦草(Chen et al., 1997); (E) 在减数分裂I期部分染色体以姐妹染色单体分离, 其余以二价体分离, 随后在减数分裂II期分裂缺失形成IMR型2n配子, 如百合(Lim et al., 2001); (F) 减数分裂正常而进入配子体发育阶段时基因组加倍形成PMD型2n配子, 如马铃薯(Bastiaanssen et al., 1998) ...
... (A) Chromosome numbers of normal gametes are halved compared with somatic cells; (B) PRD (pre-meiotic doubling) gametes are obtained as a result of the tetraploid pollen mother cells formed by cytomixis or syncytium, e.g., Dactylis glomerata (Falistocco et al., 1995); (C) In the FDR (first division restitution) type, pairing and division of homoeologous chromosomes do not occur during meiosis I, and the FDR 2n gametes maintain all of their parental genes except cross-over fragments, e.g., Paspalum jesuiticum (Filho et al., 2014) and Lolium perenne (Chen et al., 1997); (D) SDR (second division restitution) gametes with two copies of non-recombinant chromosomes are the result of the second division omission or cytokinesis abnormalities after normal anaphase II, e.g., L. perenne (Chen et al., 1997); (E) IMR (indeterminate meiotic restitution) gametes were discovered in Lilium longiflorum × Asiatic hybrid (Lim et al., 2001), in which some chromosomes are separated as univalents during meiosis I whereas the others are separated as bivalents, and subsequently the second division omission occurs during meiosis II; (F) PMD (post-meiotic doubling) gametes occurs due to the genome doubling of haploid spores during microgametogenesis, e.g., Solanum tuberosum (Bastiaanssen et al., 1998) ...
Complete asynapsis resulting in 2n pollen formation in Paspalum jesuiticum Parodi (Poaceae) 2 2014
... (A) 与体细胞相比正常配子的染色体数目减半; (B) 通过胞质融合或多核体形成的四倍体花粉母细胞产生PRD型2n配子, 如鸭茅(Falistocco et al., 1995); (C) 在减数分裂I期同源染色体无配对和分裂形成的FDR型2n配子将保留除重组片段外的所有亲本基因, 如雀稗(Filho et al., 2014)和多年生黑麦草(Chen et al., 1997); (D) 在减数分裂II期因分裂缺失形成的SDR型2n配子将保留除重组片段外比正常配子多1套染色体副本的2n配子, 如多年生黑麦草(Chen et al., 1997); (E) 在减数分裂I期部分染色体以姐妹染色单体分离, 其余以二价体分离, 随后在减数分裂II期分裂缺失形成IMR型2n配子, 如百合(Lim et al., 2001); (F) 减数分裂正常而进入配子体发育阶段时基因组加倍形成PMD型2n配子, 如马铃薯(Bastiaanssen et al., 1998) ...
... (A) Chromosome numbers of normal gametes are halved compared with somatic cells; (B) PRD (pre-meiotic doubling) gametes are obtained as a result of the tetraploid pollen mother cells formed by cytomixis or syncytium, e.g., Dactylis glomerata (Falistocco et al., 1995); (C) In the FDR (first division restitution) type, pairing and division of homoeologous chromosomes do not occur during meiosis I, and the FDR 2n gametes maintain all of their parental genes except cross-over fragments, e.g., Paspalum jesuiticum (Filho et al., 2014) and Lolium perenne (Chen et al., 1997); (D) SDR (second division restitution) gametes with two copies of non-recombinant chromosomes are the result of the second division omission or cytokinesis abnormalities after normal anaphase II, e.g., L. perenne (Chen et al., 1997); (E) IMR (indeterminate meiotic restitution) gametes were discovered in Lilium longiflorum × Asiatic hybrid (Lim et al., 2001), in which some chromosomes are separated as univalents during meiosis I whereas the others are separated as bivalents, and subsequently the second division omission occurs during meiosis II; (F) PMD (post-meiotic doubling) gametes occurs due to the genome doubling of haploid spores during microgametogenesis, e.g., Solanum tuberosum (Bastiaanssen et al., 1998) ...
Heritable alteration of DNA methylation induced by whole-chromosome aneuploidy in wheat 1 2016
... 三倍体也是通往非整倍体的常用桥梁(图3E).对三倍体谷子授以二倍体花粉, 在杂交后代中获得全套三体(王润奇等, 1994).在异源三倍体水稻(AAE)/水稻的杂交后代中, BC1染色体数目变幅为28-31, BC2染色体数目变幅为24-28, 在BC2F2中获得8个附加系(2n=20Os+1Oa) (Multani et al., 1994).在高粱(Hanna and Schertz, 1971)、粳稻(O. sativa subsp. geng) (Iwata and Omura, 1975)和籼稻(O. sativa subsp. xian) (程祝宽等, 1996)等重要禾本科作物中均有通过3x/2x构建的全套初级三体.胚乳组织常表现出有丝分裂不规则, 导致染色体桥和落后染色体进而形成非整倍体(Thomas and Chaturvedi, 2008), 如黑麦草胚乳(Norstog et al., 1969).多花黑麦草4x/2x合成的三倍体与二倍体黑麦草杂交, 后代非整倍体占比为70.89% (56/79); 当以三倍体作父本时, 后代整倍体占比高达87.69% (741/845); 差异源于三倍体作父本时整倍体花粉有选择优势而非整倍体花粉失活(Thomas et al., 1988).随着分子标记和测序技术的发展, 利用非整倍体进行基因定位和构建连锁图谱已无优势.在拟南芥非整倍体自交后代中的整倍体个体, 某些性状的表现与非整倍体母本保持一致(Henry et al., 2010).Gao等(2016)证实了非整倍化在新生多倍体的基因组演化中具有重要作用, 非整倍体小麦的DNA甲基化水平和模式可以稳定地被整倍体后代继承, 这表明非整倍体可以为育种创造表观遗传变异. ...
Assessment of morphological and phytochemical attributes in triploid and hexaploid plants of the bioenergy crop Miscanthus × giganteus 1 2016
... 三倍体可作为通往其它倍性水平的桥梁, 即三倍体桥(triploid-bridge) (图3C).在鸭茅三倍体后代中发现了四倍体、五倍体以及混倍体植株(钟声, 2006).三倍体摩擦禾可产生2n雌配子, 通过3x/2x合成了四倍体(Kindiger and Dewald, 1994).以三倍体杂交狼尾草(美洲狼尾草/象草, 2n=3x=21)的幼穗离体培养获得的胚性愈伤组织为材料, 通过秋水仙碱诱导获得六倍体(钟小仙等, 2014).利用秋水仙碱处理象草(2n=4x=28)/珍珠粟(P. glaucum, 2n=2x=14)杂交三倍体幼苗, 体细胞加倍后形成异源六倍体(Campos et al., 2009).用类似的方法获得M. × miscanthus六倍体(Yu et al., 2009; Ghimire et al., 2016).水稻(AA)和高秆野生稻(O. alta, CCDD)杂交形成异源三倍体, 加倍后形成三基异源六倍体(AACCDD) (祝剑峰等, 2008). ...
The spontaneous formation of haploid, polyploid, and aneuploid embryos in some orchids 1 1947
Trisome identification in Sorghum bicolor( L.) Moench by observing progeny of triploid × translocation stocks 1 1971
... 三倍体也是通往非整倍体的常用桥梁(图3E).对三倍体谷子授以二倍体花粉, 在杂交后代中获得全套三体(王润奇等, 1994).在异源三倍体水稻(AAE)/水稻的杂交后代中, BC1染色体数目变幅为28-31, BC2染色体数目变幅为24-28, 在BC2F2中获得8个附加系(2n=20Os+1Oa) (Multani et al., 1994).在高粱(Hanna and Schertz, 1971)、粳稻(O. sativa subsp. geng) (Iwata and Omura, 1975)和籼稻(O. sativa subsp. xian) (程祝宽等, 1996)等重要禾本科作物中均有通过3x/2x构建的全套初级三体.胚乳组织常表现出有丝分裂不规则, 导致染色体桥和落后染色体进而形成非整倍体(Thomas and Chaturvedi, 2008), 如黑麦草胚乳(Norstog et al., 1969).多花黑麦草4x/2x合成的三倍体与二倍体黑麦草杂交, 后代非整倍体占比为70.89% (56/79); 当以三倍体作父本时, 后代整倍体占比高达87.69% (741/845); 差异源于三倍体作父本时整倍体花粉有选择优势而非整倍体花粉失活(Thomas et al., 1988).随着分子标记和测序技术的发展, 利用非整倍体进行基因定位和构建连锁图谱已无优势.在拟南芥非整倍体自交后代中的整倍体个体, 某些性状的表现与非整倍体母本保持一致(Henry et al., 2010).Gao等(2016)证实了非整倍化在新生多倍体的基因组演化中具有重要作用, 非整倍体小麦的DNA甲基化水平和模式可以稳定地被整倍体后代继承, 这表明非整倍体可以为育种创造表观遗传变异. ...
On ?. Winge and a prayer: the origins of polyploidy 0 1975
Phenotypic consequences of aneuploidy in Arabidopsis thaliana 1 2010
... 三倍体也是通往非整倍体的常用桥梁(图3E).对三倍体谷子授以二倍体花粉, 在杂交后代中获得全套三体(王润奇等, 1994).在异源三倍体水稻(AAE)/水稻的杂交后代中, BC1染色体数目变幅为28-31, BC2染色体数目变幅为24-28, 在BC2F2中获得8个附加系(2n=20Os+1Oa) (Multani et al., 1994).在高粱(Hanna and Schertz, 1971)、粳稻(O. sativa subsp. geng) (Iwata and Omura, 1975)和籼稻(O. sativa subsp. xian) (程祝宽等, 1996)等重要禾本科作物中均有通过3x/2x构建的全套初级三体.胚乳组织常表现出有丝分裂不规则, 导致染色体桥和落后染色体进而形成非整倍体(Thomas and Chaturvedi, 2008), 如黑麦草胚乳(Norstog et al., 1969).多花黑麦草4x/2x合成的三倍体与二倍体黑麦草杂交, 后代非整倍体占比为70.89% (56/79); 当以三倍体作父本时, 后代整倍体占比高达87.69% (741/845); 差异源于三倍体作父本时整倍体花粉有选择优势而非整倍体花粉失活(Thomas et al., 1988).随着分子标记和测序技术的发展, 利用非整倍体进行基因定位和构建连锁图谱已无优势.在拟南芥非整倍体自交后代中的整倍体个体, 某些性状的表现与非整倍体母本保持一致(Henry et al., 2010).Gao等(2016)证实了非整倍化在新生多倍体的基因组演化中具有重要作用, 非整倍体小麦的DNA甲基化水平和模式可以稳定地被整倍体后代继承, 这表明非整倍体可以为育种创造表观遗传变异. ...
In vitro culture of endosperm and its application in plant breeding: approaches to polyploidy breeding 3 2011
... 胚乳发育阶段是胚乳培养的关键因素, 常用授粉后天数(days after pollination, DAP)来表征.通常以未成熟胚乳作外植体, 如多年生黑麦草在9-10 DAP (Norstog, 1956)、玉米在8-11 DAP (Straus and La Rue, 1954; Tamaoki and Ullstrup, 1958)、水稻在4-7 DAP (Nakano et al., 1975)、小麦在10 DAP (赵世绪等, 1984)以及大麦在8 DAP (Sehgal, 1974); 小麦-黑麦杂交胚乳的愈伤组织在14 DAP时诱导频率最佳, 在7 DAP时次之, 而在21 DAP时已不能诱导出愈伤组织(王敬驹等, 1982).以毛竹30-40 DAP的成熟胚乳作为外植体获得成功(袁金玲等, 2015).一般情况下, 成熟胚乳启动细胞分裂在于初始时与胚的联动, 而未成熟胚乳启动细胞分裂与胚是否存在无关(Hoshino et al., 2011).需要特别指出的是, 添加赤霉素等外源激素可以代替胚, 而小麦和黑麦的胚乳在无胚和赤霉素的条件下也可产生愈伤组织并分化成苗(赵世绪等, 1984).培养基、生长调节剂和环境因素对于胚乳三倍体的形成也起着重要作用.例如, 培养基中添加8%的蔗糖有利于小麦-黑麦杂交胚乳的愈伤组织诱导(王敬驹等, 1982).添加氮源对于未成熟胚乳的增殖至关重要(Hoshino et al., 2011).在小麦和黑麦胚乳培养时, 添加激动素和2,4-D可提高愈伤组织诱导频率(赵世绪等, 1984).玉米胚乳在暗处培养效果更佳(Straus and La Rue, 1954), 而在黑麦中光照对胚乳的增殖无影响(Norstog et al.,1969).胚乳培养以组织培养为技术基础, 并未成为主流的三倍体合成方式. ...
... ).添加氮源对于未成熟胚乳的增殖至关重要(Hoshino et al., 2011).在小麦和黑麦胚乳培养时, 添加激动素和2,4-D可提高愈伤组织诱导频率(赵世绪等, 1984).玉米胚乳在暗处培养效果更佳(Straus and La Rue, 1954), 而在黑麦中光照对胚乳的增殖无影响(Norstog et al.,1969).胚乳培养以组织培养为技术基础, 并未成为主流的三倍体合成方式. ...
Agrobacterium-mediated transformed transgenic triploid bermudagrass ( Cynodon dactylon × C. transvaalensis) plants are highly resistant to the glufosinate herbicide liberty 1 2005
... 对于有直接利用价值的三倍体, 可通过组培、诱变和转基因等技术进一步改良(图3B).MS-Supreme源于Tifgreen的自然突变, 与Tifgreen相比其叶片和茎节节间缩小、变短(Krans et al., 1999).在Tifgreen的自然突变中还选育出Tifdwarf和FloraDwarf等矮生品种; TifwayII、TifSport和TifEagle则由相应三倍体亲本经γ辐射诱变而成(Kamps et al., 2011).利用体细胞无性系变异育种技术在TifEagle的愈伤组织中长时间连续继代培养能有效诱导体细胞变异(如矮化突变体); 以0.3 mol·L-1 NaCl为选择压筛选出耐盐细胞系(Lu et al., 2007).以美洲狼尾草(Pennisetum americana)/ 四倍体象草合成的三倍体F1种子为诱变材料, 经γ辐射获得优异变异株闽牧6号(陈钟佃等, 2012).华南1号杂交狼尾草由三倍体杂交狼尾草芽变而来(陈平等, 2004).三倍体也可以直接作为亲本利用.在偏序钝叶草(Stenotaphrum secundatum, 2n=2x=18)的3x/ 2x杂交后代中经胚拯救选育出抗灰斑病(由稻瘟病菌(Magnaporthe grisea)引起)和耐旱草坪草新品种(Chandra et al., 2015).奇岗天然后代群体中16个数量性状出现了丰富的变异(变异系数为14.41%- 151.85%), 在后代中筛选出3份材料, 其生物量优于奇岗(潘志军等, 2017).通过三倍体狼尾草与摩特矮象草杂交选育出中国南方地区广泛种植的三倍体多年生牧草桂牧1号(罗宗志等, 2016).宁牧26-2狼尾草也是由三倍体后代衍生而来(顾洪如等, 1992).三倍体转基因品系可通过2种策略获得.(1) 直接以三倍体为受体, 导入目的基因构建转基因品系, 如三倍体狗牙根Tifeagle (Hu et al., 2005).(2) 对于愈伤组织形成与再分化率低的三倍体, 首先需要寻找愈伤组织再分化优良的二倍体或四倍体作为转基因桥梁, 然后通过基因枪或农杆菌转化等方法导入目的基因, 最后通过倍性间杂交构建转基因三倍体, 如芒(2n=2x=38) (Wang et al., 2011).近年来, 基因编辑技术的发展为定向改良三倍体提供了新的育种方案. ...
Mysterious meiotic behavior of autopolyploid and allopolyploid maize 1 2018
... 在染色体制备的基础上, 基于特异基因重复序列的荧光原位杂交技术(fluorescence in situ hybridization, FISH)和基于基因组中同源性重复序列的基因组原位杂交技术(genomic in situ hybridization, GISH)进一步提高了鉴定的分辨率.例如, 野大麦(H. brevisubutum, 2n=4x=28, H1H1H2H2)与加拿大披碱草(Elymus canadensis, 2n=4x=28, SSHcHc)杂交, 通过GISH鉴定某F1为一组Hc染色体组丢失的异源三倍体(2n=3x=21, SH1H2) (于卓等, 2004).利用GISH技术成功鉴定了多年生黑麦草-羊茅(L. perenne-Festuca pratensis, 2n=3x=14Lp+7Fp) (King et al., 1999)、玉米-四倍体多年生大刍草(Z. mays-Z. perennis, 2n=3x=10Zm+20Zp) (Iqbal et al., 2018)及水稻-斑点野生稻(O. sativa-O. punctata, 2n=3x= 12Os+24Op) (Sun et al., 2017)等异源三倍体的基因组构成.同样地, 该法通常用于特异材料倍性鉴定和亲缘关系分析, 鉴定大批量材料过程更显繁琐. ...
Studies on the trisomics in rice plants (Oryza sativa L.): III. Relation between trisomics and genetic linkage groups 1 1975
... 三倍体也是通往非整倍体的常用桥梁(图3E).对三倍体谷子授以二倍体花粉, 在杂交后代中获得全套三体(王润奇等, 1994).在异源三倍体水稻(AAE)/水稻的杂交后代中, BC1染色体数目变幅为28-31, BC2染色体数目变幅为24-28, 在BC2F2中获得8个附加系(2n=20Os+1Oa) (Multani et al., 1994).在高粱(Hanna and Schertz, 1971)、粳稻(O. sativa subsp. geng) (Iwata and Omura, 1975)和籼稻(O. sativa subsp. xian) (程祝宽等, 1996)等重要禾本科作物中均有通过3x/2x构建的全套初级三体.胚乳组织常表现出有丝分裂不规则, 导致染色体桥和落后染色体进而形成非整倍体(Thomas and Chaturvedi, 2008), 如黑麦草胚乳(Norstog et al., 1969).多花黑麦草4x/2x合成的三倍体与二倍体黑麦草杂交, 后代非整倍体占比为70.89% (56/79); 当以三倍体作父本时, 后代整倍体占比高达87.69% (741/845); 差异源于三倍体作父本时整倍体花粉有选择优势而非整倍体花粉失活(Thomas et al., 1988).随着分子标记和测序技术的发展, 利用非整倍体进行基因定位和构建连锁图谱已无优势.在拟南芥非整倍体自交后代中的整倍体个体, 某些性状的表现与非整倍体母本保持一致(Henry et al., 2010).Gao等(2016)证实了非整倍化在新生多倍体的基因组演化中具有重要作用, 非整倍体小麦的DNA甲基化水平和模式可以稳定地被整倍体后代继承, 这表明非整倍体可以为育种创造表观遗传变异. ...
A statistical mixture model for estimating the proportion of unreduced pollen grains in perennial ryegrass (Lolium perenne L.) via the size of pollen grains 1 1993
... 在通过形态鉴定体细胞或配子体倍性的实践中, 花粉粒大小(花粉直径)是常用参数.例如, 燕麦属(Arena)花粉粒大小与植株倍性呈极显著正相关(r=0.86, P< 0.01) (Katsiotis and Forsberg, 1995).在大麦(Johansen and Vonbothmer, 1994)、看麦娘属(Alepecurus)和沿沟草属(Catabrosa) (Sheidai et al., 2009)中也有相似的结果.然而, 多年生黑麦草花粉粒大小不受体细胞倍性的影响(Jansen and Dennijs, 1993).垂穗草(Bouteloua curtipendula, 2n=2x=20)在六倍体以内花粉粒大小及其变异系数随着倍性的增加而增加, 当超过六倍体后2个参数不再随倍性变化(Kapadia and Gould, 1964).在4个大麦属(Hordeum)物种中, 已露出小花外的花粉粒, 其花粉直径大于未露出小花的花粉粒; 花粉粒大小随着倍性水平的增加而增加(2x、4x和6x) (Johansen and Von Bothmer, 1994).因此, 通过花粉鉴定植株倍性应当注意物种间差异并统一花粉发育时期.无芒雀麦(Bromus inermis, x=7)的花粉粒大小在各倍性(4x、6x和8x)之间存在重叠, 通过气孔长度鉴定倍性优于气孔频率与花粉粒大小(Tan and Dunn, 1973).与通过花粉粒性状鉴定倍性相似, 通过气孔性状鉴定倍性的效果也因物种而异. ...
Pollen size in Hordeum L.: correlation between size, ploidy level, and breeding system 2 1994
... 在通过形态鉴定体细胞或配子体倍性的实践中, 花粉粒大小(花粉直径)是常用参数.例如, 燕麦属(Arena)花粉粒大小与植株倍性呈极显著正相关(r=0.86, P< 0.01) (Katsiotis and Forsberg, 1995).在大麦(Johansen and Vonbothmer, 1994)、看麦娘属(Alepecurus)和沿沟草属(Catabrosa) (Sheidai et al., 2009)中也有相似的结果.然而, 多年生黑麦草花粉粒大小不受体细胞倍性的影响(Jansen and Dennijs, 1993).垂穗草(Bouteloua curtipendula, 2n=2x=20)在六倍体以内花粉粒大小及其变异系数随着倍性的增加而增加, 当超过六倍体后2个参数不再随倍性变化(Kapadia and Gould, 1964).在4个大麦属(Hordeum)物种中, 已露出小花外的花粉粒, 其花粉直径大于未露出小花的花粉粒; 花粉粒大小随着倍性水平的增加而增加(2x、4x和6x) (Johansen and Von Bothmer, 1994).因此, 通过花粉鉴定植株倍性应当注意物种间差异并统一花粉发育时期.无芒雀麦(Bromus inermis, x=7)的花粉粒大小在各倍性(4x、6x和8x)之间存在重叠, 通过气孔长度鉴定倍性优于气孔频率与花粉粒大小(Tan and Dunn, 1973).与通过花粉粒性状鉴定倍性相似, 通过气孔性状鉴定倍性的效果也因物种而异. ...
... ) (Johansen and Von Bothmer, 1994).因此, 通过花粉鉴定植株倍性应当注意物种间差异并统一花粉发育时期.无芒雀麦(Bromus inermis, x=7)的花粉粒大小在各倍性(4x、6x和8x)之间存在重叠, 通过气孔长度鉴定倍性优于气孔频率与花粉粒大小(Tan and Dunn, 1973).与通过花粉粒性状鉴定倍性相似, 通过气孔性状鉴定倍性的效果也因物种而异. ...
Growth responses of mature endosperm in cultures 1 1965
Indeterminate meiotic restitution (IMR): a novel type of meiotic nuclear restitution mechanism detected in interspecific lily hybrids by GISH 2 2001
... (A) 与体细胞相比正常配子的染色体数目减半; (B) 通过胞质融合或多核体形成的四倍体花粉母细胞产生PRD型2n配子, 如鸭茅(Falistocco et al., 1995); (C) 在减数分裂I期同源染色体无配对和分裂形成的FDR型2n配子将保留除重组片段外的所有亲本基因, 如雀稗(Filho et al., 2014)和多年生黑麦草(Chen et al., 1997); (D) 在减数分裂II期因分裂缺失形成的SDR型2n配子将保留除重组片段外比正常配子多1套染色体副本的2n配子, 如多年生黑麦草(Chen et al., 1997); (E) 在减数分裂I期部分染色体以姐妹染色单体分离, 其余以二价体分离, 随后在减数分裂II期分裂缺失形成IMR型2n配子, 如百合(Lim et al., 2001); (F) 减数分裂正常而进入配子体发育阶段时基因组加倍形成PMD型2n配子, 如马铃薯(Bastiaanssen et al., 1998) ...
... (A) Chromosome numbers of normal gametes are halved compared with somatic cells; (B) PRD (pre-meiotic doubling) gametes are obtained as a result of the tetraploid pollen mother cells formed by cytomixis or syncytium, e.g., Dactylis glomerata (Falistocco et al., 1995); (C) In the FDR (first division restitution) type, pairing and division of homoeologous chromosomes do not occur during meiosis I, and the FDR 2n gametes maintain all of their parental genes except cross-over fragments, e.g., Paspalum jesuiticum (Filho et al., 2014) and Lolium perenne (Chen et al., 1997); (D) SDR (second division restitution) gametes with two copies of non-recombinant chromosomes are the result of the second division omission or cytokinesis abnormalities after normal anaphase II, e.g., L. perenne (Chen et al., 1997); (E) IMR (indeterminate meiotic restitution) gametes were discovered in Lilium longiflorum × Asiatic hybrid (Lim et al., 2001), in which some chromosomes are separated as univalents during meiosis I whereas the others are separated as bivalents, and subsequently the second division omission occurs during meiosis II; (F) PMD (post-meiotic doubling) gametes occurs due to the genome doubling of haploid spores during microgametogenesis, e.g., Solanum tuberosum (Bastiaanssen et al., 1998) ...
Mechanisms of unreduced gamete formation in flowering plants 1 2017
... 目前, 通过观测小孢子发生在植物中已发现5种2n配子形成机制(Loginova and Silkova, 2017) (图2).减数分裂II期姐妹染色单体分离形成的FDR (first division restitution)型2n配子(图2C)在很大程度上保留了亲本的杂合性, 而减数分裂II期姐妹染色单体不分离形成的SDR (second division restitution)型2n配子(图2D)高度纯合.因此, 在转移亲本杂合性方面FDR型比SDR型更为有利(Younis et al., 2014).Harlan和De Wet (1975)认为所有植物都以某种频率产生2n配子, 但物种间发生频率不同.二倍体水稻(Oryza sativa, 2n=2x=24)多胚苗能自发地产生三倍体(张静等, 2002).在二倍体看麦娘(Alopecurus bulbosus, 2n=2x=14)自然群体中, 二倍体、三倍体与四倍体并存(Sieber and Murray, 1979).在22个二倍体大麦(Hordeum vulgare, 2n=2x=14)品种中, 三倍体植株的发生率为0.01%-0.29% (Sandfaer, 1975).在黄花茅(Anthoxanthum alpinum, 2n=2x=10)中2n配子的发生率为6.3% (Bretagnolle, 2001).对十字花科24个物种60个群体的研究表明, 尽管2n配子先前被认为相对罕见、但普遍发生(通常个体的雄配子2n发生率<2%), 并且在特定个体中较高, 这些极少数产生高频率2n配子的个体可能在多倍体的形成中发挥关键作用(Kreiner et al., 2017).在同一植物中, 雌雄2n配子的发生率也不一致(Ramsey and Schemske, 1998).鸭茅(Dactylis glomerata, 2n=2x=14) 2n雌配子发生率为0.49%, 而2n雄配子发生率为0.98% (Maceira et al.,1992).鉴于2n配子发生率低, 在2x/2x下通过单向有性多倍化(unilateral sexual polyploidization, 2x+x)形成三倍体的频率远高于通过双向有性多倍化(bilateral sexual polyploidization, 2x+2x)形成四倍体的频率.例如, 在黄花茅中2n配子形成三倍体的发生率为0.2%, 但未见四倍体发生(Bretagnolle, 2001). ...
In vitro selection of salinity tolerant variants from triploid bermudagrass ( Cynodon transvaalensis × C. dactylon) and their physiological responses to salt and drought stress 1 2007
... 对于有直接利用价值的三倍体, 可通过组培、诱变和转基因等技术进一步改良(图3B).MS-Supreme源于Tifgreen的自然突变, 与Tifgreen相比其叶片和茎节节间缩小、变短(Krans et al., 1999).在Tifgreen的自然突变中还选育出Tifdwarf和FloraDwarf等矮生品种; TifwayII、TifSport和TifEagle则由相应三倍体亲本经γ辐射诱变而成(Kamps et al., 2011).利用体细胞无性系变异育种技术在TifEagle的愈伤组织中长时间连续继代培养能有效诱导体细胞变异(如矮化突变体); 以0.3 mol·L-1 NaCl为选择压筛选出耐盐细胞系(Lu et al., 2007).以美洲狼尾草(Pennisetum americana)/ 四倍体象草合成的三倍体F1种子为诱变材料, 经γ辐射获得优异变异株闽牧6号(陈钟佃等, 2012).华南1号杂交狼尾草由三倍体杂交狼尾草芽变而来(陈平等, 2004).三倍体也可以直接作为亲本利用.在偏序钝叶草(Stenotaphrum secundatum, 2n=2x=18)的3x/ 2x杂交后代中经胚拯救选育出抗灰斑病(由稻瘟病菌(Magnaporthe grisea)引起)和耐旱草坪草新品种(Chandra et al., 2015).奇岗天然后代群体中16个数量性状出现了丰富的变异(变异系数为14.41%- 151.85%), 在后代中筛选出3份材料, 其生物量优于奇岗(潘志军等, 2017).通过三倍体狼尾草与摩特矮象草杂交选育出中国南方地区广泛种植的三倍体多年生牧草桂牧1号(罗宗志等, 2016).宁牧26-2狼尾草也是由三倍体后代衍生而来(顾洪如等, 1992).三倍体转基因品系可通过2种策略获得.(1) 直接以三倍体为受体, 导入目的基因构建转基因品系, 如三倍体狗牙根Tifeagle (Hu et al., 2005).(2) 对于愈伤组织形成与再分化率低的三倍体, 首先需要寻找愈伤组织再分化优良的二倍体或四倍体作为转基因桥梁, 然后通过基因枪或农杆菌转化等方法导入目的基因, 最后通过倍性间杂交构建转基因三倍体, 如芒(2n=2x=38) (Wang et al., 2011).近年来, 基因编辑技术的发展为定向改良三倍体提供了新的育种方案. ...
Production of 2 n gametes in diploid subspecies of Dactylis glomerata L. 1. Occurrence and frequency of 2n pollen 1 1992
... 目前, 通过观测小孢子发生在植物中已发现5种2n配子形成机制(Loginova and Silkova, 2017) (图2).减数分裂II期姐妹染色单体分离形成的FDR (first division restitution)型2n配子(图2C)在很大程度上保留了亲本的杂合性, 而减数分裂II期姐妹染色单体不分离形成的SDR (second division restitution)型2n配子(图2D)高度纯合.因此, 在转移亲本杂合性方面FDR型比SDR型更为有利(Younis et al., 2014).Harlan和De Wet (1975)认为所有植物都以某种频率产生2n配子, 但物种间发生频率不同.二倍体水稻(Oryza sativa, 2n=2x=24)多胚苗能自发地产生三倍体(张静等, 2002).在二倍体看麦娘(Alopecurus bulbosus, 2n=2x=14)自然群体中, 二倍体、三倍体与四倍体并存(Sieber and Murray, 1979).在22个二倍体大麦(Hordeum vulgare, 2n=2x=14)品种中, 三倍体植株的发生率为0.01%-0.29% (Sandfaer, 1975).在黄花茅(Anthoxanthum alpinum, 2n=2x=10)中2n配子的发生率为6.3% (Bretagnolle, 2001).对十字花科24个物种60个群体的研究表明, 尽管2n配子先前被认为相对罕见、但普遍发生(通常个体的雄配子2n发生率<2%), 并且在特定个体中较高, 这些极少数产生高频率2n配子的个体可能在多倍体的形成中发挥关键作用(Kreiner et al., 2017).在同一植物中, 雌雄2n配子的发生率也不一致(Ramsey and Schemske, 1998).鸭茅(Dactylis glomerata, 2n=2x=14) 2n雌配子发生率为0.49%, 而2n雄配子发生率为0.98% (Maceira et al.,1992).鉴于2n配子发生率低, 在2x/2x下通过单向有性多倍化(unilateral sexual polyploidization, 2x+x)形成三倍体的频率远高于通过双向有性多倍化(bilateral sexual polyploidization, 2x+2x)形成四倍体的频率.例如, 在黄花茅中2n配子形成三倍体的发生率为0.2%, 但未见四倍体发生(Bretagnolle, 2001). ...
Production of viable male unreduced gametes in Brassica interspecific hybrids is genotype specific and stimulated by cold temperatures 1 2011
... 实践中, 2n配子可通过环境调控、化学诱导、种间杂种和减数分裂突变体获得.2n配子的发生主要受遗传控制, 但暴露于低温或高温、热冲击和高辐射下可诱导或刺激2n配子形成(Younis et al., 2014).例如, 蔷薇属(Rosa)植物花芽暴露于36°C高温下可提高2n雄配子的发生率(Pécrix et al., 2011).在禾本科中采用环境调控诱导2n配子发生尚未见报道.N2O在诱导鸭茅(Van Santen et al., 1991)、玉米(Zea mays, 2n=2x=20) (Kato and Birchler, 2006)、小麦(Triticum aestivum, 2n=6x=42)及大麦(Dvorak et al., 1973)的2n配子上取得成功.向玉米雄穗喷洒氟乐灵可诱导2n花粉并获得同源三倍体(Kato, 1999).种间或属间杂种是产生2n配子的重要途径, 其频率高于祖先物种(Ramsey and Schemske, 1998).Mason等(2011)分析了3种芸薹属(Brassica)物种及其F1杂种的2n雄配子发生率, 发现5个甘蓝型油菜(B. napus, 2n=4x=38)品系的发生率(0%-0.11%, 平均值为0.03%)较低, 而甘蓝型油菜的11种F1杂种的发生率(0.17%-3.29%, 平均值为1.55%)较高.可见, 种间杂种较亲本基因组2n配子的发生率提高.Zeng等(2020)发现在兰花(Cymbidium ssp.)传统品种中2n雄配子发生率为0.15%-1.0%, 而种间杂种为2.5%-4.03%; 同时作者认为可能由于无性繁殖植物存在可替代的繁殖途径, 2n配子的形成频率相对更高.减数分裂活跃基因的突变可能导致纺锤体形成异常或胞质分裂受阻而形成2n配子, 如拟南芥(Arabidopsis thaliana, 2n= 2x=10)中的cyca1;2/tam (SDR型)、osd1 (SDR型)和Atps1 (FDR型)突变体(Brownfield and K?hler, 2011). ...
Development of monosomic alien addition lines and introgression of genes from Oryza australiensis Domin. to cultivated rice O 2 1994
Direct and indirect in vitro regeneration of Miscanthus × giganteus cultivar freedom: effects of explant type and medium on regeneration efficiency 1 2015
Hybrid weakness in a rice interspecific hybrid is nitrogen-dependent, and accompanied by changes in gene expression at both total transcript level and parental allele partitioning 1 2017
... 在染色体制备的基础上, 基于特异基因重复序列的荧光原位杂交技术(fluorescence in situ hybridization, FISH)和基于基因组中同源性重复序列的基因组原位杂交技术(genomic in situ hybridization, GISH)进一步提高了鉴定的分辨率.例如, 野大麦(H. brevisubutum, 2n=4x=28, H1H1H2H2)与加拿大披碱草(Elymus canadensis, 2n=4x=28, SSHcHc)杂交, 通过GISH鉴定某F1为一组Hc染色体组丢失的异源三倍体(2n=3x=21, SH1H2) (于卓等, 2004).利用GISH技术成功鉴定了多年生黑麦草-羊茅(L. perenne-Festuca pratensis, 2n=3x=14Lp+7Fp) (King et al., 1999)、玉米-四倍体多年生大刍草(Z. mays-Z. perennis, 2n=3x=10Zm+20Zp) (Iqbal et al., 2018)及水稻-斑点野生稻(O. sativa-O. punctata, 2n=3x= 12Os+24Op) (Sun et al., 2017)等异源三倍体的基因组构成.同样地, 该法通常用于特异材料倍性鉴定和亲缘关系分析, 鉴定大批量材料过程更显繁琐. ...
Cultivation in vitro of excised endosperm and meristem tissues of corn 1 1958
... 胚乳发育阶段是胚乳培养的关键因素, 常用授粉后天数(days after pollination, DAP)来表征.通常以未成熟胚乳作外植体, 如多年生黑麦草在9-10 DAP (Norstog, 1956)、玉米在8-11 DAP (Straus and La Rue, 1954; Tamaoki and Ullstrup, 1958)、水稻在4-7 DAP (Nakano et al., 1975)、小麦在10 DAP (赵世绪等, 1984)以及大麦在8 DAP (Sehgal, 1974); 小麦-黑麦杂交胚乳的愈伤组织在14 DAP时诱导频率最佳, 在7 DAP时次之, 而在21 DAP时已不能诱导出愈伤组织(王敬驹等, 1982).以毛竹30-40 DAP的成熟胚乳作为外植体获得成功(袁金玲等, 2015).一般情况下, 成熟胚乳启动细胞分裂在于初始时与胚的联动, 而未成熟胚乳启动细胞分裂与胚是否存在无关(Hoshino et al., 2011).需要特别指出的是, 添加赤霉素等外源激素可以代替胚, 而小麦和黑麦的胚乳在无胚和赤霉素的条件下也可产生愈伤组织并分化成苗(赵世绪等, 1984).培养基、生长调节剂和环境因素对于胚乳三倍体的形成也起着重要作用.例如, 培养基中添加8%的蔗糖有利于小麦-黑麦杂交胚乳的愈伤组织诱导(王敬驹等, 1982).添加氮源对于未成熟胚乳的增殖至关重要(Hoshino et al., 2011).在小麦和黑麦胚乳培养时, 添加激动素和2,4-D可提高愈伤组织诱导频率(赵世绪等, 1984).玉米胚乳在暗处培养效果更佳(Straus and La Rue, 1954), 而在黑麦中光照对胚乳的增殖无影响(Norstog et al.,1969).胚乳培养以组织培养为技术基础, 并未成为主流的三倍体合成方式. ...
Relationship of stomatal length and frequency and pollen-grain diameter to ploidy level in Bromus inermis Leyss 1 1973
... 在通过形态鉴定体细胞或配子体倍性的实践中, 花粉粒大小(花粉直径)是常用参数.例如, 燕麦属(Arena)花粉粒大小与植株倍性呈极显著正相关(r=0.86, P< 0.01) (Katsiotis and Forsberg, 1995).在大麦(Johansen and Vonbothmer, 1994)、看麦娘属(Alepecurus)和沿沟草属(Catabrosa) (Sheidai et al., 2009)中也有相似的结果.然而, 多年生黑麦草花粉粒大小不受体细胞倍性的影响(Jansen and Dennijs, 1993).垂穗草(Bouteloua curtipendula, 2n=2x=20)在六倍体以内花粉粒大小及其变异系数随着倍性的增加而增加, 当超过六倍体后2个参数不再随倍性变化(Kapadia and Gould, 1964).在4个大麦属(Hordeum)物种中, 已露出小花外的花粉粒, 其花粉直径大于未露出小花的花粉粒; 花粉粒大小随着倍性水平的增加而增加(2x、4x和6x) (Johansen and Von Bothmer, 1994).因此, 通过花粉鉴定植株倍性应当注意物种间差异并统一花粉发育时期.无芒雀麦(Bromus inermis, x=7)的花粉粒大小在各倍性(4x、6x和8x)之间存在重叠, 通过气孔长度鉴定倍性优于气孔频率与花粉粒大小(Tan and Dunn, 1973).与通过花粉粒性状鉴定倍性相似, 通过气孔性状鉴定倍性的效果也因物种而异. ...
The use of a triploid hybrid for introgression in Lolium species 1 1988
... 三倍体也是通往非整倍体的常用桥梁(图3E).对三倍体谷子授以二倍体花粉, 在杂交后代中获得全套三体(王润奇等, 1994).在异源三倍体水稻(AAE)/水稻的杂交后代中, BC1染色体数目变幅为28-31, BC2染色体数目变幅为24-28, 在BC2F2中获得8个附加系(2n=20Os+1Oa) (Multani et al., 1994).在高粱(Hanna and Schertz, 1971)、粳稻(O. sativa subsp. geng) (Iwata and Omura, 1975)和籼稻(O. sativa subsp. xian) (程祝宽等, 1996)等重要禾本科作物中均有通过3x/2x构建的全套初级三体.胚乳组织常表现出有丝分裂不规则, 导致染色体桥和落后染色体进而形成非整倍体(Thomas and Chaturvedi, 2008), 如黑麦草胚乳(Norstog et al., 1969).多花黑麦草4x/2x合成的三倍体与二倍体黑麦草杂交, 后代非整倍体占比为70.89% (56/79); 当以三倍体作父本时, 后代整倍体占比高达87.69% (741/845); 差异源于三倍体作父本时整倍体花粉有选择优势而非整倍体花粉失活(Thomas et al., 1988).随着分子标记和测序技术的发展, 利用非整倍体进行基因定位和构建连锁图谱已无优势.在拟南芥非整倍体自交后代中的整倍体个体, 某些性状的表现与非整倍体母本保持一致(Henry et al., 2010).Gao等(2016)证实了非整倍化在新生多倍体的基因组演化中具有重要作用, 非整倍体小麦的DNA甲基化水平和模式可以稳定地被整倍体后代继承, 这表明非整倍体可以为育种创造表观遗传变异. ...
Endosperm culture: a novel method for triploid plant production 4 2008
Establishment of an efficient in vitro culture and particle bombardment-mediated transformation systems in Miscanthus sinensis Anderss, a potential bioenergy crop 1 2011
... 对于有直接利用价值的三倍体, 可通过组培、诱变和转基因等技术进一步改良(图3B).MS-Supreme源于Tifgreen的自然突变, 与Tifgreen相比其叶片和茎节节间缩小、变短(Krans et al., 1999).在Tifgreen的自然突变中还选育出Tifdwarf和FloraDwarf等矮生品种; TifwayII、TifSport和TifEagle则由相应三倍体亲本经γ辐射诱变而成(Kamps et al., 2011).利用体细胞无性系变异育种技术在TifEagle的愈伤组织中长时间连续继代培养能有效诱导体细胞变异(如矮化突变体); 以0.3 mol·L-1 NaCl为选择压筛选出耐盐细胞系(Lu et al., 2007).以美洲狼尾草(Pennisetum americana)/ 四倍体象草合成的三倍体F1种子为诱变材料, 经γ辐射获得优异变异株闽牧6号(陈钟佃等, 2012).华南1号杂交狼尾草由三倍体杂交狼尾草芽变而来(陈平等, 2004).三倍体也可以直接作为亲本利用.在偏序钝叶草(Stenotaphrum secundatum, 2n=2x=18)的3x/ 2x杂交后代中经胚拯救选育出抗灰斑病(由稻瘟病菌(Magnaporthe grisea)引起)和耐旱草坪草新品种(Chandra et al., 2015).奇岗天然后代群体中16个数量性状出现了丰富的变异(变异系数为14.41%- 151.85%), 在后代中筛选出3份材料, 其生物量优于奇岗(潘志军等, 2017).通过三倍体狼尾草与摩特矮象草杂交选育出中国南方地区广泛种植的三倍体多年生牧草桂牧1号(罗宗志等, 2016).宁牧26-2狼尾草也是由三倍体后代衍生而来(顾洪如等, 1992).三倍体转基因品系可通过2种策略获得.(1) 直接以三倍体为受体, 导入目的基因构建转基因品系, 如三倍体狗牙根Tifeagle (Hu et al., 2005).(2) 对于愈伤组织形成与再分化率低的三倍体, 首先需要寻找愈伤组织再分化优良的二倍体或四倍体作为转基因桥梁, 然后通过基因枪或农杆菌转化等方法导入目的基因, 最后通过倍性间杂交构建转基因三倍体, 如芒(2n=2x=38) (Wang et al., 2011).近年来, 基因编辑技术的发展为定向改良三倍体提供了新的育种方案. ...
Exploitation of induced 2 n-gametes for plant breeding 2 2014
... 目前, 通过观测小孢子发生在植物中已发现5种2n配子形成机制(Loginova and Silkova, 2017) (图2).减数分裂II期姐妹染色单体分离形成的FDR (first division restitution)型2n配子(图2C)在很大程度上保留了亲本的杂合性, 而减数分裂II期姐妹染色单体不分离形成的SDR (second division restitution)型2n配子(图2D)高度纯合.因此, 在转移亲本杂合性方面FDR型比SDR型更为有利(Younis et al., 2014).Harlan和De Wet (1975)认为所有植物都以某种频率产生2n配子, 但物种间发生频率不同.二倍体水稻(Oryza sativa, 2n=2x=24)多胚苗能自发地产生三倍体(张静等, 2002).在二倍体看麦娘(Alopecurus bulbosus, 2n=2x=14)自然群体中, 二倍体、三倍体与四倍体并存(Sieber and Murray, 1979).在22个二倍体大麦(Hordeum vulgare, 2n=2x=14)品种中, 三倍体植株的发生率为0.01%-0.29% (Sandfaer, 1975).在黄花茅(Anthoxanthum alpinum, 2n=2x=10)中2n配子的发生率为6.3% (Bretagnolle, 2001).对十字花科24个物种60个群体的研究表明, 尽管2n配子先前被认为相对罕见、但普遍发生(通常个体的雄配子2n发生率<2%), 并且在特定个体中较高, 这些极少数产生高频率2n配子的个体可能在多倍体的形成中发挥关键作用(Kreiner et al., 2017).在同一植物中, 雌雄2n配子的发生率也不一致(Ramsey and Schemske, 1998).鸭茅(Dactylis glomerata, 2n=2x=14) 2n雌配子发生率为0.49%, 而2n雄配子发生率为0.98% (Maceira et al.,1992).鉴于2n配子发生率低, 在2x/2x下通过单向有性多倍化(unilateral sexual polyploidization, 2x+x)形成三倍体的频率远高于通过双向有性多倍化(bilateral sexual polyploidization, 2x+2x)形成四倍体的频率.例如, 在黄花茅中2n配子形成三倍体的发生率为0.2%, 但未见四倍体发生(Bretagnolle, 2001). ...
... 实践中, 2n配子可通过环境调控、化学诱导、种间杂种和减数分裂突变体获得.2n配子的发生主要受遗传控制, 但暴露于低温或高温、热冲击和高辐射下可诱导或刺激2n配子形成(Younis et al., 2014).例如, 蔷薇属(Rosa)植物花芽暴露于36°C高温下可提高2n雄配子的发生率(Pécrix et al., 2011).在禾本科中采用环境调控诱导2n配子发生尚未见报道.N2O在诱导鸭茅(Van Santen et al., 1991)、玉米(Zea mays, 2n=2x=20) (Kato and Birchler, 2006)、小麦(Triticum aestivum, 2n=6x=42)及大麦(Dvorak et al., 1973)的2n配子上取得成功.向玉米雄穗喷洒氟乐灵可诱导2n花粉并获得同源三倍体(Kato, 1999).种间或属间杂种是产生2n配子的重要途径, 其频率高于祖先物种(Ramsey and Schemske, 1998).Mason等(2011)分析了3种芸薹属(Brassica)物种及其F1杂种的2n雄配子发生率, 发现5个甘蓝型油菜(B. napus, 2n=4x=38)品系的发生率(0%-0.11%, 平均值为0.03%)较低, 而甘蓝型油菜的11种F1杂种的发生率(0.17%-3.29%, 平均值为1.55%)较高.可见, 种间杂种较亲本基因组2n配子的发生率提高.Zeng等(2020)发现在兰花(Cymbidium ssp.)传统品种中2n雄配子发生率为0.15%-1.0%, 而种间杂种为2.5%-4.03%; 同时作者认为可能由于无性繁殖植物存在可替代的繁殖途径, 2n配子的形成频率相对更高.减数分裂活跃基因的突变可能导致纺锤体形成异常或胞质分裂受阻而形成2n配子, 如拟南芥(Arabidopsis thaliana, 2n= 2x=10)中的cyca1;2/tam (SDR型)、osd1 (SDR型)和Atps1 (FDR型)突变体(Brownfield and K?hler, 2011). ...
Chromosome doubling of the bioenergy crop, Miscanthus × giganteus 1 2009
... 三倍体可作为通往其它倍性水平的桥梁, 即三倍体桥(triploid-bridge) (图3C).在鸭茅三倍体后代中发现了四倍体、五倍体以及混倍体植株(钟声, 2006).三倍体摩擦禾可产生2n雌配子, 通过3x/2x合成了四倍体(Kindiger and Dewald, 1994).以三倍体杂交狼尾草(美洲狼尾草/象草, 2n=3x=21)的幼穗离体培养获得的胚性愈伤组织为材料, 通过秋水仙碱诱导获得六倍体(钟小仙等, 2014).利用秋水仙碱处理象草(2n=4x=28)/珍珠粟(P. glaucum, 2n=2x=14)杂交三倍体幼苗, 体细胞加倍后形成异源六倍体(Campos et al., 2009).用类似的方法获得M. × miscanthus六倍体(Yu et al., 2009; Ghimire et al., 2016).水稻(AA)和高秆野生稻(O. alta, CCDD)杂交形成异源三倍体, 加倍后形成三基异源六倍体(AACCDD) (祝剑峰等, 2008). ...
Unreduced male gamete formation in Cymbidium and its use for developing sexual polyploid cultivars 1 2020
... 实践中, 2n配子可通过环境调控、化学诱导、种间杂种和减数分裂突变体获得.2n配子的发生主要受遗传控制, 但暴露于低温或高温、热冲击和高辐射下可诱导或刺激2n配子形成(Younis et al., 2014).例如, 蔷薇属(Rosa)植物花芽暴露于36°C高温下可提高2n雄配子的发生率(Pécrix et al., 2011).在禾本科中采用环境调控诱导2n配子发生尚未见报道.N2O在诱导鸭茅(Van Santen et al., 1991)、玉米(Zea mays, 2n=2x=20) (Kato and Birchler, 2006)、小麦(Triticum aestivum, 2n=6x=42)及大麦(Dvorak et al., 1973)的2n配子上取得成功.向玉米雄穗喷洒氟乐灵可诱导2n花粉并获得同源三倍体(Kato, 1999).种间或属间杂种是产生2n配子的重要途径, 其频率高于祖先物种(Ramsey and Schemske, 1998).Mason等(2011)分析了3种芸薹属(Brassica)物种及其F1杂种的2n雄配子发生率, 发现5个甘蓝型油菜(B. napus, 2n=4x=38)品系的发生率(0%-0.11%, 平均值为0.03%)较低, 而甘蓝型油菜的11种F1杂种的发生率(0.17%-3.29%, 平均值为1.55%)较高.可见, 种间杂种较亲本基因组2n配子的发生率提高.Zeng等(2020)发现在兰花(Cymbidium ssp.)传统品种中2n雄配子发生率为0.15%-1.0%, 而种间杂种为2.5%-4.03%; 同时作者认为可能由于无性繁殖植物存在可替代的繁殖途径, 2n配子的形成频率相对更高.减数分裂活跃基因的突变可能导致纺锤体形成异常或胞质分裂受阻而形成2n配子, 如拟南芥(Arabidopsis thaliana, 2n= 2x=10)中的cyca1;2/tam (SDR型)、osd1 (SDR型)和Atps1 (FDR型)突变体(Brownfield and K?hler, 2011). ...
 ,2,*, 杜周和
,2,*, 杜周和
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT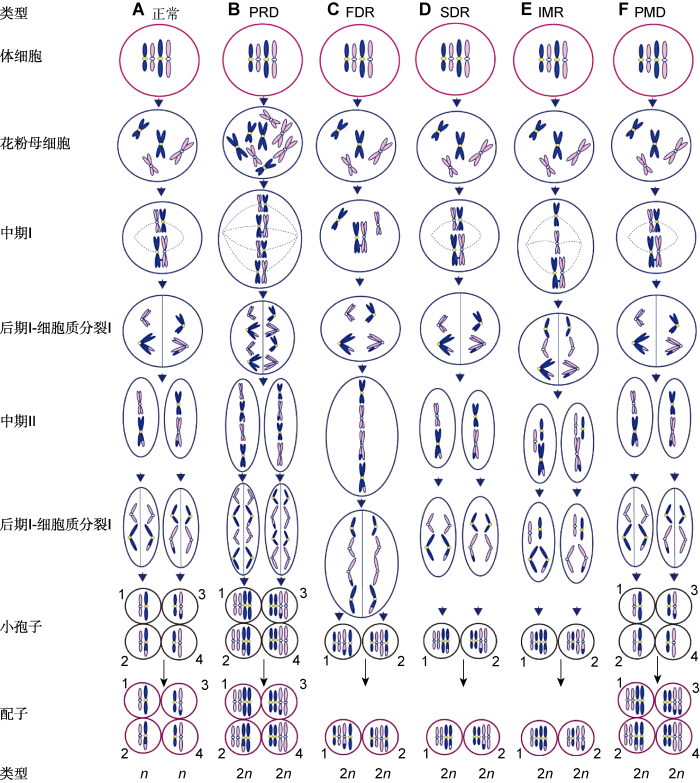 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT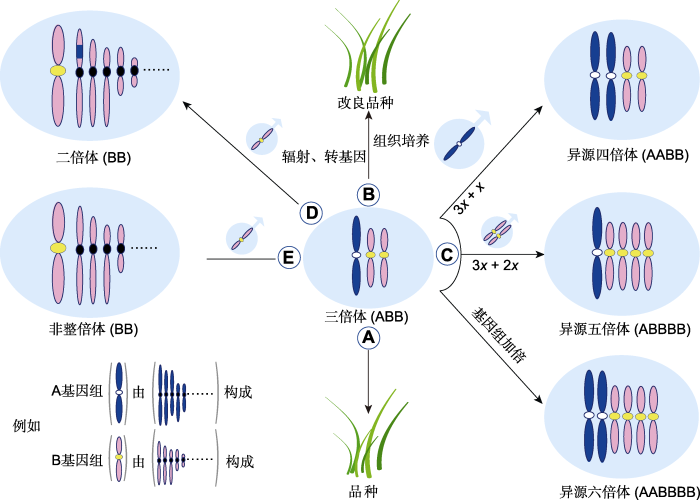 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}