Identification of the Cysteine Protease Family and Corresponding miRNAs in Jatropha curcas and Their Response to Chill-hardening
Dandan Wu1, Yongkun Chen1,2, Yu Yang1, Chunyan Kong1, Ming Gong,1,*1Key Laboratory of Biomass Energy and Environmental Biotechnology of Yunnan Province, Engineering Research Center of Sustainable Development and Utilization of Biomass Energy, Ministry of Education, School of Life Sciences, Yunnan Normal University, Kunming 650500, China 2School of Life Sciences, Xinjiang Normal University, Urumqi 830054, China
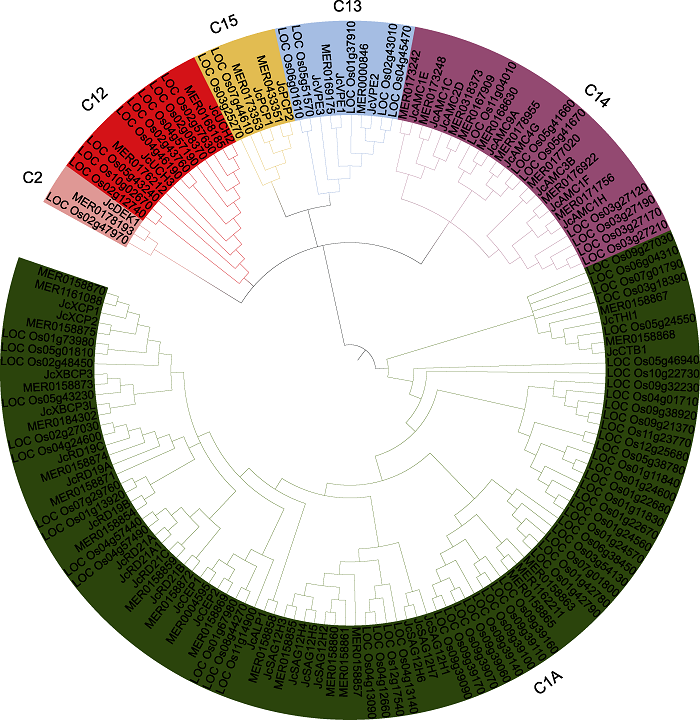
Abstract Jatropha curcas is a kind of promising energy plants, but also a chilling-sensitive plant, which can be chill- hardening at 12°C with significant improvement of its chilling tolerance. In this study, the cysteine protease gene family of J. curcas and their corresponding miRNAs were identified at the genome-wide level. The results showed that a total of 39 cysteine protease genes were identified in J. curcas genome, which were located on 11 chromosomes and could be divided into six subfamilies (C1A, C2, C12, C13, C14 and C15); all encoding 181-2 158 amino acids with Cys and His active sites. Based on the sequencing results of miRNAome and degradome, 283 miRNAs were found to be targeted to 14 members of cysteine protease gene family. In addition, the co-expression analysis of those miRNAs targeting to JcDEK1, JcRD21B and JcXBCP3L during chill-hardening demonstrated significantly negative correlation during the chill-hardening at 12°C, suggesting that these miRNAs are involved in the regulation of the cysteine protease genes, and this regulation should be related to the enhancement of chilling tolerance induced by the chill-hardening. This study will be helpful for better understanding the function of cysteine protease gene family in J. curcas and the interaction of the family genes with their corresponding miRNAs, and how this interaction regulates the response of J. curcas to low temperature. Keywords:Jatropha curcas;cysteine protease;gene family;microRNAs;low temperature response
PDF (2328KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 吴丹丹, 陈永坤, 杨宇, 孔春艳, 龚明. 小桐子半胱氨酸蛋白酶家族和相应miRNAs的鉴定及其对低温锻炼的响应. 植物学报, 2021, 56(5): 544-558 doi:10.11983/CBB21014 Wu Dandan, Chen Yongkun, Yang Yu, Kong Chunyan, Gong Ming. Identification of the Cysteine Protease Family and Corresponding miRNAs in Jatropha curcas and Their Response to Chill-hardening. Chinese Bulletin of Botany, 2021, 56(5): 544-558 doi:10.11983/CBB21014
红色代表半胱氨酸蛋白酶基因, 蓝色代表miRNAs。 Figure 7Networks of targeted regulation among miRNAs and their corresponding cysteine protease family genes in Jatropha curcas
Red represents cysteine protease gene and blue represents miRNAs.
*与**分别表示不同时间的表达量与对照0小时在P<0.05与P<0.01水平差异显著。 Figure 9Co-expression analysis by qRT-PCR of the cysteine protease genes JcDEK1, JcRD21B and JcXBCP3L as well as the miRNAs targeting these genes in Jatropha curcas seedlings during chill-hardening at 12°C
* and ** indicated the significant differences between the expression level at different time and control 0 h at P<0.05 and P<0.01 level, respectively.
AhmadR, Zuily-FodilY, PassaquetC, BethenodO, RocheR, RepellinA (2014). Identification and characterization of MOR-CP, a cysteine protease induced by ozone and developmental senescence in maize ( Zea mays L.) leaves Chemosphere 108, 245-250. DOI:10.1016/j.chemosphere.2014.01.047URL [本文引用: 2]
AoPX, LiZG, FanDM, GongM (2013a). Involvement of antioxidant defense system in chill hardening-induced chilling tolerance in Jatropha curcas seedlings Acta Physiol Plant 35, 153-160. DOI:10.1007/s11738-012-1058-zURL [本文引用: 5]
AoPX, LiZG, GongM (2013b). Involvement of compatible solutes in chill hardening-induced chilling tolerance in Jatropha curcas seedlings Acta Physiol Plant 35, 3457-3464. DOI:10.1007/s11738-013-1381-zURL [本文引用: 4]
ChenC, YuY, DingXD, LiuBD, DuanmuH, ZhuD, SunXL, CaoL, NisaZU, LiQ, ZhuYM (2018). Genome-wide analysis and expression profiling of PP2C clade D under saline and alkali stresses in wild soybean and Arabidopsis Protoplasma 255, 643-654. DOI:10.1007/s00709-017-1172-2URL [本文引用: 1]
ChenCJ, ChenH, ZhangY, ThomasHR, FrankMH, HeYH, XiaR (2020). TBtools: an integrative toolkit developed for interactive analyses of big biological data Mol Plant 13, 1194-1202. DOI:10.1016/j.molp.2020.06.009URL [本文引用: 1]
ClarkK, FrancoJY, SchwizerS, PangZ, HawaraE, LiebrandT (2018). An effector from the huanglongbing-associated pathogen targets citrus proteases Nat Commun 9, 1718. DOI:10.1038/s41467-018-04140-9URL [本文引用: 1]
DandoPM, FortunatoM, StrandGB, SmithTS, BarrettAJ (2003). Pyroglutamyl-peptidase I: cloning, sequencing, and characterisation of the recombinant human enzyme Protein Expr Purif 28, 111-119. DOI:10.1016/S1046-5928(02)00632-0URL [本文引用: 1]
EarnshawWC, MartinsLM, KaufmannSH (1999). Mammalian caspases: structure, activation, substrates, and functions during apoptosis Annu Rev Biochem 68, 383-424. PMID:10872455 [本文引用: 1] Apoptosis is a genetically programmed, morphologically distinct form of cell death that can be triggered by a variety of physiological and pathological stimuli. Studies performed over the past 10 years have demonstrated that proteases play critical roles in initiation and execution of this process. The caspases, a family of cysteine-dependent aspartate-directed proteases, are prominent among the death proteases. Caspases are synthesized as relatively inactive zymogens that become activated by scaffold-mediated transactivation or by cleavage via upstream proteases in an intracellular cascade. Regulation of caspase activation and activity occurs at several different levels: (a) Zymogen gene transcription is regulated; (b) antiapoptotic members of the Bcl-2 family and other cellular polypeptides block proximity-induced activation of certain procaspases; and (c) certain cellular inhibitor of apoptosis proteins (cIAPs) can bind to and inhibit active caspases. Once activated, caspases cleave a variety of intracellular polypeptides, including major structural elements of the cytoplasm and nucleus, components of the DNA repair machinery, and a number of protein kinases. Collectively, these scissions disrupt survival pathways and disassemble important architectural components of the cell, contributing to the stereotypic morphological and biochemical changes that characterize apoptotic cell death.
FinnRD, PenelopeC, EberhardtRY, EddySR, JainaM, MitchellAL, PotterSC, MarcoP, MatloobQ, AmaiaSV (2016). The Pfam protein families database: towards a more sustainable future Nucleic Acids Res 44, D279-D285. DOI:10.1093/nar/gkv1344URL [本文引用: 1]
GrudkowskaM, ZagdanskaB (2004). Multifunctional role of plant cysteine proteinases Acta Biochim Pol 51, 609-624. DOI:10.18388/abp.2004_3547URL [本文引用: 1]
Hara-NishimuraI (1995). Vacuolar processing enzyme responsible for maturation of vacuolar proteins Seikagaku 67, 372-377. PMID:7602211 [本文引用: 1]
HatsugaiN, KuroyanagiM, NishimuraM, Hara-NishimuraI (2006). A cellular suicide strategy of plants: vacuole- mediated cell death Apoptosis 11, 905-911. PMID:16547592 [本文引用: 1] Programmed cell death (PCD) occurs in animals and plants under various stresses and during development. Recently, vacuolar processing enzyme (VPE) was identified as an executioner of plant PCD. VPE is a cysteine protease that cleaves a peptide bond at the C-terminal side of asparagine and aspartic acid. VPE exhibited enzymatic properties similar to that of a caspase, which is a cysteine protease that mediates the PCD pathway in animals, although there is limited sequence identity between the two enzymes. VPE and caspase-1 share several structural properties: the catalytic dyads and three amino acids forming the substrate pockets (Asp pocket) are conserved between VPE and caspase-1. In contrast to such similarities, subcellular localizations of these proteases are completely different from each other. VPE is localized in the vacuoles, while caspases are localized in the cytosol. VPE functions as a key molecule of plant PCD through disrupting the vacuole in pathogenesis and development. Cell death triggered by vacuolar collapse is unique to plants and has not been seen in animals. Plants might have evolved a VPE-mediated vacuolar system as a cellular suicide strategy.
KuroyanagiM, YamadaK, HatsugaiN, KondoM, NishimuraM, Hara-NishimuraI (2005). Vacuolar processing enzyme is essential for mycotoxin-induced cell death in Arabidopsis thaliana J Biol Chem 280, 32914-32920. PMID:16043487 [本文引用: 1] Some compatible pathogens secrete toxins to induce host cell death and promote their growth. The toxin-induced cell death is a pathogen strategy for infection. To clarify the executioner of the toxin-induced cell death, we examined a fungal toxin (fumonisin B1 (FB1))-induced cell death of Arabidopsis plants. FB1-induced cell death was accompanied with disruption of vacuolar membrane followed by lesion formation. The features of FB1-induced cell death were completely abolished in the Arabidopsis vacuolar processing enzyme (VPE)-null mutant, which lacks all four VPE genes of the genome. Interestingly, an inhibitor of caspase-1 abolished FB1-induced lesion formation, as did a VPE inhibitor. The VPE-null mutant had no detectable activities of caspase-1 or VPE in the FB1-treated leaves, although wild-type leaves had the caspase-1 and VPE activities, both of which were inhibited by a caspase-1 inhibitor. gammaVPE is the most essential among the four VPE homologues for FB1-induced cell death in Arabidopsis leaves. Recombinant gammaVPE recognized a VPE substrate with Km = 30.3 microm and a caspase-1 substrate with Km = 44.2 microm, which is comparable with the values for mammalian caspase-1. The gammaVPE precursor was self-catalytically converted into the mature form exhibiting caspase-1 activity. These in vivo and in vitro analyses demonstrate that gammaVPE is the proteinase that exhibits a caspase-1 activity. We show that VPE exhibiting a caspase-1 activity is a key molecule in toxin-induced cell death. Our findings suggest that a susceptible response of toxin-induced cell death is caused by the VPE-mediated vacuolar mechanism similar to a resistance response of hypersensitive cell death (Hatsugai, N., Kuroyanagi, M., Yamada, K., Meshi, T., Tsuda, S., Kondo, M., Nishimura, M., and Hara-Nishimura, I. (2004) Science 305, 855-858).
LiZG, ZengHZ, AoPX, GongM (2014). Lipid response to short-term chilling shock and long-term chill hardening in Jatropha curcas L. seedlings Acta Physiol Plant 36, 2803-2814. DOI:10.1007/s11738-014-1653-2URL [本文引用: 4]
LiuHJ, HuMH, WangQ, ChengL, ZhangZB (2018). Role of papain-like cysteine proteases in plant development Front Plant Sci 9, 1717. DOI:10.3389/fpls.2018.01717URL [本文引用: 1]
MeghaS, BasuU, KavNNV (2018). Regulation of low temperature stress in plants by microRNAs Plant Cell Environ 41, 1-15. DOI:10.1111/pce.v41.1URL [本文引用: 1]
NiñoMC, KimMS, KangKK, ChoYG (2020). Genome- wide identification and molecular characterization of cysteine protease genes in rice Plant Biotechnol Rep 14, 69-87. DOI:10.1007/s11816-019-00583-8URL [本文引用: 2]
RawlingsND, BarrettAJ, BatemanA (2010). MEROPS: the peptidase database Nucl Acids Res 38, D227-D233. DOI:10.1093/nar/gkp971URL [本文引用: 1]
RawlingsND, SalvesenG (2013). Handbook of Proteolytic Enzymes. Salt Lake City: Academic Press. pp. 1253-1257. [本文引用: 1]
RichauKH, KaschaniF, VerdoesM, PansuriyaTC, NiessenS, StüberK, ColbyT, OverkleeftHS, BogyoM, Van der HoornRAL (2012). Subclassification and biochemical analysis of plant papain-like cysteine proteases displays subfamily-specific characteristics Plant Physiol 158, 1583-1599. DOI:10.1104/pp.112.194001URL [本文引用: 1]
ShimadaT, YamadaK, KataokaM, NakauneS, KoumotoY, KuroyanagiM, TabataS, KatoT, ShinozakiK, SekiM, KobayashiM, KondoM, NishimuraM, Hara-NishimuraI (2003). Vacuolar processing enzymes are essential for proper processing of seed storage proteins in Arabidopsis thaliana J Biol Chem 278, 32292-32299. PMID:12799370 [本文引用: 1] The proprotein precursors of storage proteins are post-translationally processed to produce their respective mature forms within the protein storage vacuoles of maturing seeds. To investigate the processing mechanism in vivo, we isolated Arabidopsis mutants that accumulate detectable amounts of the precursors of the storage proteins, 12 S globulins and 2 S albumins, in their seeds. All six mutants isolated have a defect in the beta VPE gene. VPE (vacuolar processing enzyme) is a cysteine proteinase with substrate specificity toward an asparagine residue. We further generated various mutants lacking different VPE isoforms: alpha VPE, beta VPE, and/or gamma VPE. More than 90% of VPE activity is abolished in the beta vpe-3 seeds, and no VPE activity is detected in the alpha vpe-1/beta vpe-3/gamma vpe-1 seeds. The triple mutant seeds accumulate no properly processed mature storage proteins. Instead, large amounts of storage protein precursors are found in the seeds of this mutant. In contrast to beta vpe-3 seeds, which accumulate both precursors and mature storage proteins, the other single (alpha vpe-1 and gamma vpe-1) and double (alpha vpe-1/gamma vpe-1) mutants accumulate no precursors in their seeds at all. Therefore, the vegetative VPEs, alpha VPE and gamma VPE, are not necessary for precursor processing in the presence of beta VPE, but partly compensates for the deficiency in beta VPE in beta vpe-3 seeds. In the absence of functional VPEs, a proportion of pro2S albumin molecules are alternatively cleaved by aspartic proteinase. This cleavage by aspartic proteinase is promoted by the initial processing of pro2S albumins by VPE. Our overall results suggest that seed-type beta VPE is most essential for the processing of storage proteins, and that the vegetative-type VPEs and aspartic proteinase complement beta VPE activity in this processing.
StroeherVL, MaclaganJL, GoodAG (1997). Molecular cloning of a Brassica napus cysteine protease gene inducible by drought and low temperature stress Physiol Plant 101, 389-397. DOI:10.1111/ppl.1997.101.issue-2URL [本文引用: 1]
VorsterBJ, CullisCA, KunertKJ (2019). Plant vacuolar processing enzymes Front Plant Sci 10, 479. DOI:10.3389/fpls.2019.00479PMID:31031794 [本文引用: 1] Plant proteomes contain hundreds of proteases divided into different families based on evolutionary and functional relationship. In particular, plant cysteine proteases of the C1 (papain-like) and C13 (legumain-like) families play key roles in many physiological processes. The legumain-like proteases, also called vacuolar processing enzymes (VPEs), perform a multifunctional role in different plant organs and during different stages of plant development and death. VPEs are similar to animal caspases, and although caspase activity was identified in plants almost 40 years ago, there still remains much research to be done to gain a complete understanding of their various roles and functions in plants. Here we not only summarize the current existing knowledge of plant VPEs, including recent developments in the field, but also highlight the future prospective areas to be investigated to obtain a more detailed understanding of the role of VPEs in plants.
WangHB, GongM, GuoJY, XinH, GaoY, LiuC, DaiDQ, TangLZ (2018a). Genome-wide Identification of Jatropha curcas MAPK, MAPKK, and MAPKKK gene families and their expression profile under cold stress Sci Rep 8, 16163. DOI:10.1038/s41598-018-34614-1URL [本文引用: 1]
WangHB, ZouZR, WangSS, GongM (2013a). Global analysis of transcriptome responses and gene expression profiles to cold stress of Jatropha curcas L PLoS One 8, e82817. DOI:10.1371/journal.pone.0082817URL [本文引用: 2]
WangSS, WangHB, GongM (2013b). Identification of microRNAs involved in chilling response by deep sequencing of Jatropha curcas L. small RNAs at the global genome level. In: Proceedings of the 21st European Biomass Conference and Exhibition. Copenhagen: European Union. pp. 356-363. [本文引用: 2]
WangW, ZhouXM, XiongHX, MaoWY, ZhaoP, SunMX (2018b). Papain-like and legumain-like proteases in rice: genome-wide identification, comprehensive gene feature characterization and expression analysis BMC Plant Biol 18, 87. DOI:10.1186/s12870-018-1298-1PMID:29764367 [本文引用: 2] Background: Papain-like and legumain-like proteases are proteolytic enzymes which play key roles in plant development, senescence and defense. The activities of proteases in both families could be inhibited by a group of small proteins called cystatin. Cystatin family genes have been well characterized both in tobacco and rice, suggesting their potential roles in seed development. However, their potential targets, papain-like and legumain-like proteases, have not been well characterized in plants, especially in rice, a model plant for cereal biology.Results: Here, 33 papain-like and 5 legumain-like proteases have been identified in rice genome, respectively. Gene structure, distribution in rice chromosome, and evolutionary relationship to their counterparts in other plants have been well characterized. Comprehensive expression profile analysis revealed that two family genes display divergent expression pattern, which are regulated temporally and spatially during the process of seed development and germination. Our experiments also revealed that the expression of most genes in these two families is sensitively responsive to plant hormones and different abiotic stresses.Conclusions: Genome-wide identification and comprehensive gene expression pattern analysis of papain-like and legumain-like proteases in rice suggests their multiple and cooperative roles in seed development and response to environmental variations, which provides several useful cues for further in-depth study.
WilkinsonKD, Laleli-SahinE, UrbauerJ, LarsenCN, ShihGH, HaasAL, WalshSTR, WandAJ (1999). The binding site for UCH-L3 on ubiquitin: mutagenesis and NMR studies on the complex between ubiquitin and UCH- L3 J Mol Biol 291, 1067-1077. PMID:10518943 [本文引用: 1] The ubiquitin fold is a versatile and widely used targeting signal that is added post-translationally to a variety of proteins. Covalent attachment of one or more ubiquitin domains results in localization of the target protein to the proteasome, the nucleus, the cytoskeleton or the endocytotic machinery. Recognition of the ubiquitin domain by a variety of enzymes and receptors is vital to the targeting function of ubiquitin. Several parallel pathways exist and these must be able to distinguish among ubiquitin, several different types of polymeric ubiquitin, and the various ubiquitin-like domains. Here we report the first molecular description of the binding site on ubiquitin for ubiquitin C-terminal hydrolase L3 (UCH-L3). The site on ubiquitin was experimentally determined using solution NMR, and site-directed mutagenesis. The site on UCH-L3 was modeled based on X-ray crystallography, multiple sequence alignments, and computer-aided docking. Basic residues located on ubiquitin (K6, K11, R72, and R74) are postulated to contact acidic residues on UCH-L3 (E10, E14, D33, E219). These putative interactions are testable and fully explain the selectivity of ubiquitin domain binding to this enzyme.
ZangQW, WangCX, LiXY, GuoZA, JingRL, ZhaoJ, ChangXP (2010). Isolation and characterization of a gene encoding a polyethylene glycol-induced cysteine protease in common wheat J Biosci 35, 379-388. DOI:10.1007/s12038-010-0043-1URL [本文引用: 1]
ZhangX, PanBZ, ChenMS, ChenW, LiJ, XuZF, LiuCN (2019). JCDB: a comprehensive knowledge base for Jatropha curcas, an emerging model for woody energy plants BMC Genomics 20, 958. DOI:10.1186/s12864-019-6356-zPMID:31874631 [本文引用: 1] Jatropha curcas is an oil-bearing plant, and has seeds with high oil content (~?40%). Several advantages, such as easy genetic transformation and short generation duration, have led to the emergence of J. curcas as a model for woody energy plants. With the development of high-throughput sequencing, the genome of Jatropha curcas has been sequenced by different groups and a mass of transcriptome data was released. How to integrate and analyze these omics data is crucial for functional genomics research on J. curcas.By establishing pipelines for processing novel gene identification, gene function annotation, and gene network construction, we systematically integrated and analyzed a series of J. curcas transcriptome data. Based on these data, we constructed a J. curcas database (JCDB), which not only includes general gene information, gene functional annotation, gene interaction networks, and gene expression matrices but also provides tools for browsing, searching, and downloading data, as well as online BLAST, the JBrowse genome browser, ID conversion, heatmaps, and gene network analysis tools.JCDB is the most comprehensive and well annotated knowledge base for J. curcas. We believe it will make a valuable contribution to the functional genomics study of J. curcas. The database is accessible at http://jcdb.xtbg.ac.cn.
ZhengL, ChenSS, XieLH, LuZC, LiuMY, HanXJ, QiaoGR, JiangJ, ZhuoRY, QiuWM, HeZQ (2018). Overexpression of cysteine protease gene from Salix matsudana enhances salt tolerance in transgenic Arabidopsis Environ Exp Bot 147, 53-62. DOI:10.1016/j.envexpbot.2017.11.008URL [本文引用: 1]
ZouZ, HuangQX, XieGS, YangLF (2018). Genome-wide comparative analysis of papain-like cysteine protease family genes in castor bean and physic nut Sci Rep 8, 331. DOI:10.1038/s41598-017-18760-6URL [本文引用: 5]
miRNA作用机制研究进展 2 2020
... miRNA是一类真核生物中广泛存在的长度为20-24 nt的内源非编码RNA, 可能的作用机制主要包括转录本切割、翻译抑制、次生siRNA形成、翻译激活、转录增强和类激素作用6个方面(郝大海和龚明, 2020).植物miRNA主要通过剪切mRNA或抑制翻译负调控基因表达, 在细胞增殖分化、个体生长发育和抵御逆境胁迫等生理过程中发挥重要作用(张翠桔等, 2020; 王劲东等, 2020).许多研究表明, miRNAs参与植物对低温的响应、信号转导和适应过程(Megha et al., 2018), 但miRNAs是否参与调控半胱氨酸蛋白酶家族基因, 以及这种互作如何调控植物耐冷性的研究尚未见报道.基于本实验室前期的工作(Ao et al., 2013a, 2013b; Wang et al., 2013a, 2013b; Li et al., 2014)和近期完成的小桐子低温锻炼过程中转录组、miRNA组和降解组测序结果, 我们对小桐子半胱氨酸蛋白酶家族进行了全基因组鉴定及系统进化、染色体定位、蛋白结构域及基因表达分析; 并基于miRNA组和降解组测序结果, 鉴定了靶向半胱氨酸蛋白酶基因家族成员的miRNAs, 进行共表达分析, 旨在阐明半胱氨酸蛋白酶基因家族及其与对应miRNAs的互作关系以及这种互作如何参与调控小桐子对低温锻炼的响应过程. ...
 ,1,*
,1,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT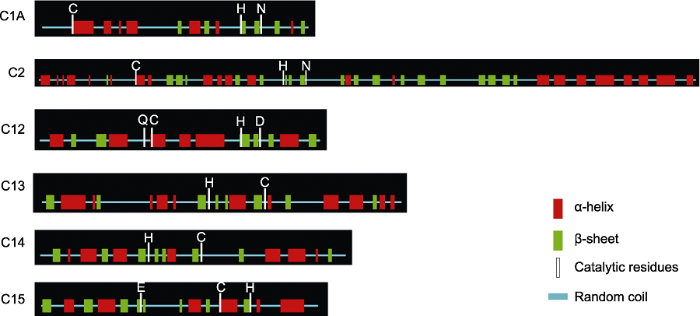 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT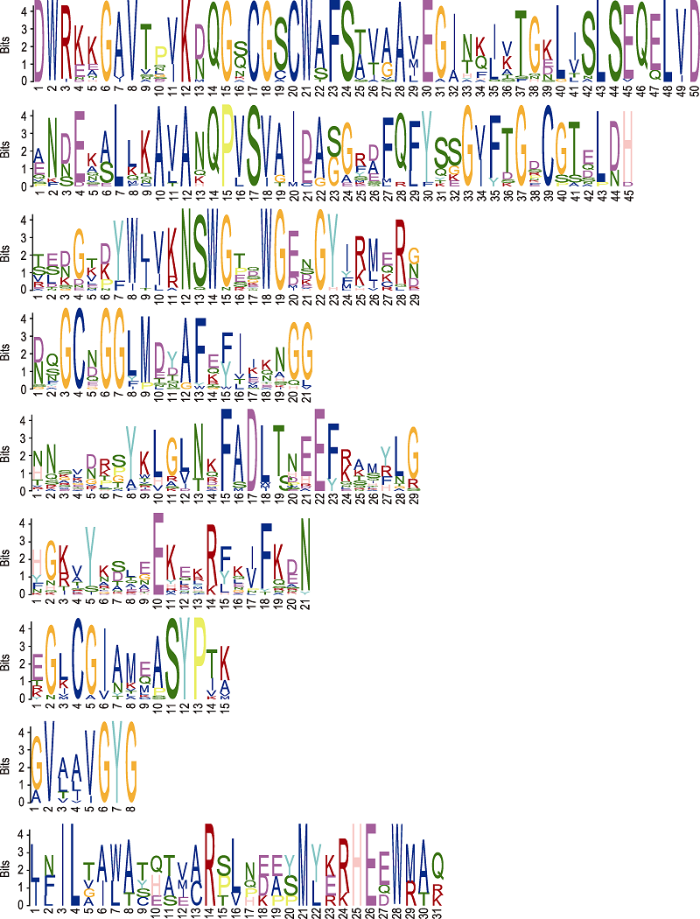 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT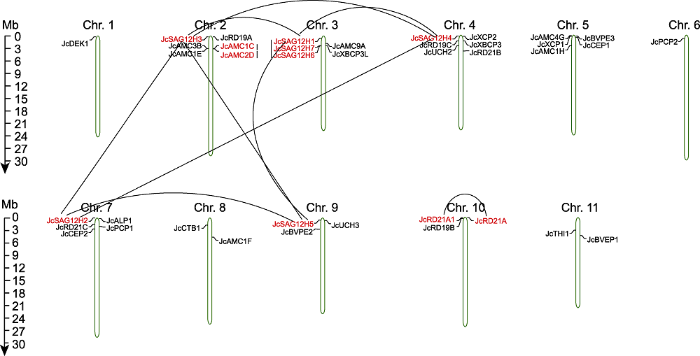 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT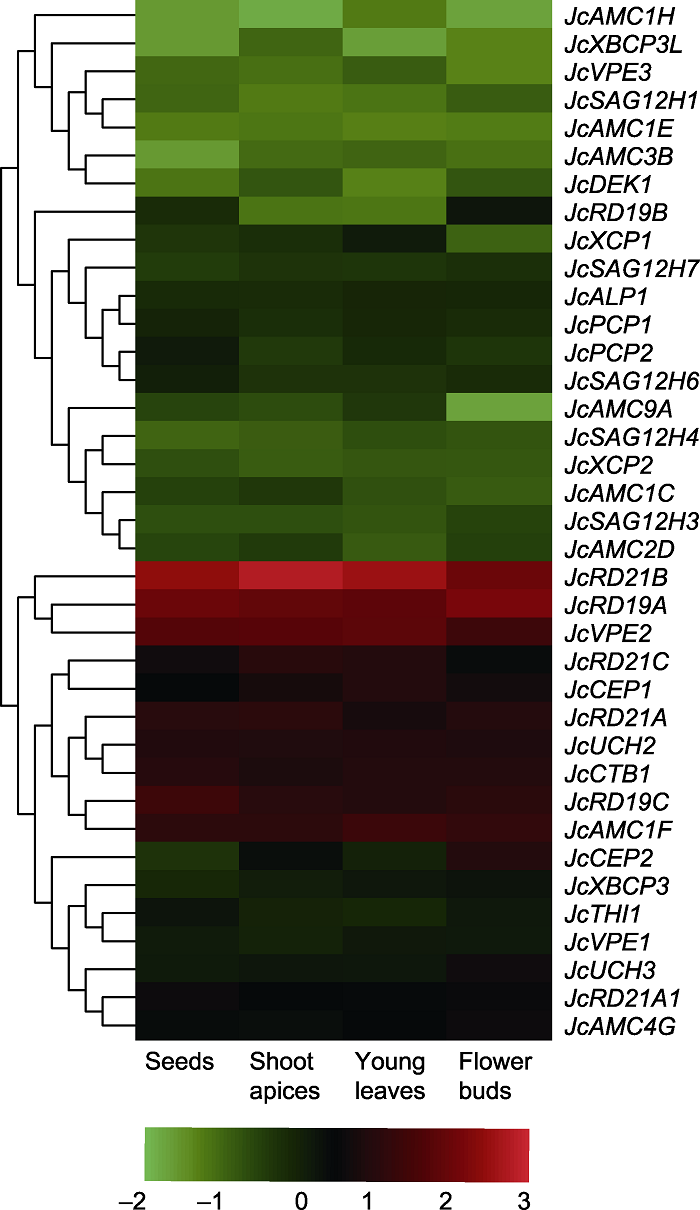 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT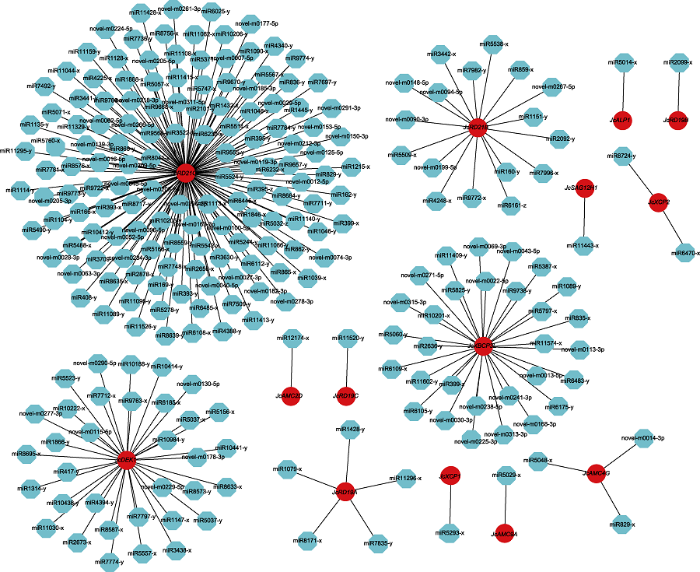 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT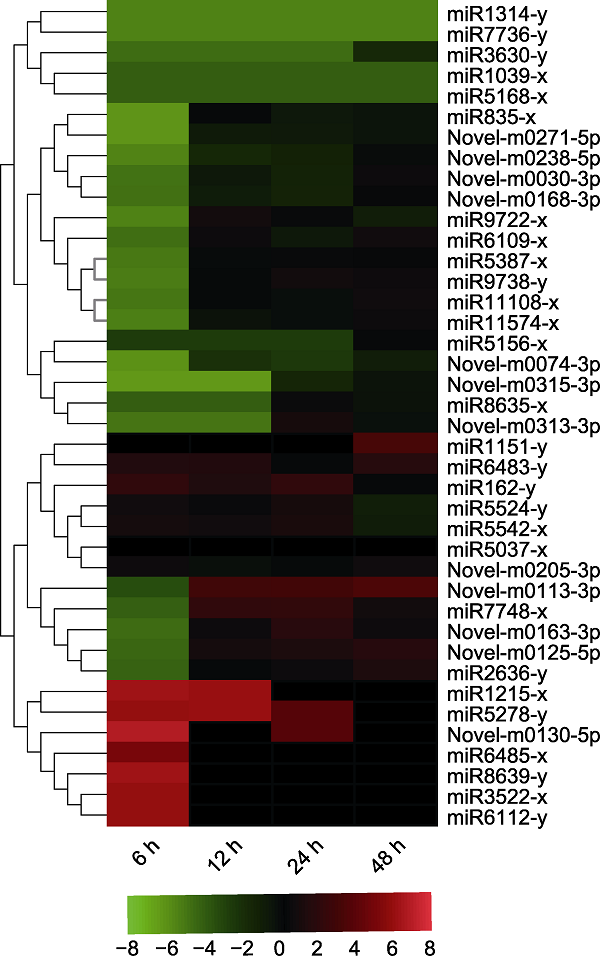 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT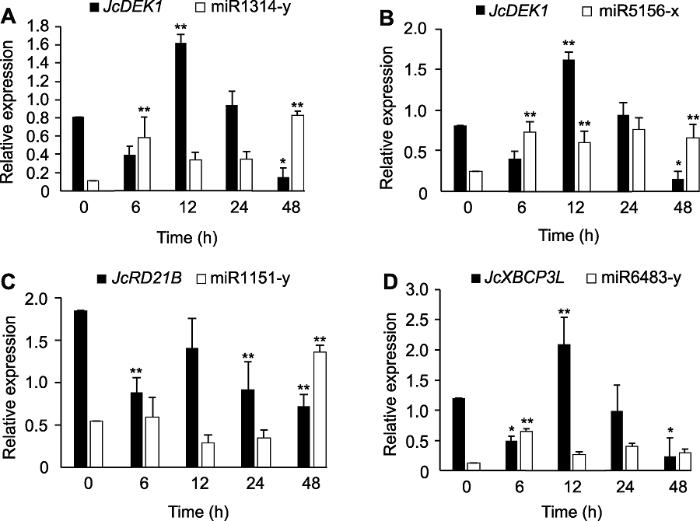 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}