 ,1,*
,1,*Seed Development and Embryo Culture of Endangered Abies beshanzuensis
Ke Liu1, Bin Liu1, Lu Yuan1, Hui Shuai1, Yang Yang1, Tingjin Wang1, Deliang Chen2, Xiaorong Chen2, Kaibin Yang2, Xiaofeng Jin3, Liping Chen,1,*通讯作者: * E-mail:chenliping@zju.edu.cn
责任编辑: 孙冬花
收稿日期:2021-06-14接受日期:2021-09-30
| 基金资助: |
Corresponding authors: * E-mail:chenliping@zju.edu.cn
Received:2021-06-14Accepted:2021-09-30

摘要
百山祖冷杉(Abies beshanzuensis)种子萌发率低, 发育特性尚不明确, 极大地限制了种群在原生境的自然更新。该研究以百山祖冷杉不同发育时期的雌球果为材料, 通过研究球果中种子的胚与胚乳数量、重量和显微结构特性, 及测定种子发育关键时期胚乳的初生代谢物, 探究种子发育特征及影响种子发育的主要因素。结果表明, 7月是百山祖冷杉种子发育的关键时期, 此期间胚乳重量快速增加, 胚乳中碳水化合物代谢活动旺盛。解剖学和形态学分析显示, 7月下旬球果中大量种子发育异常。进一步对发育关键时期胚乳中初生代谢物进行分析表明, 海藻糖-6-磷酸含量显著降低。推测在种子发育的关键时期胚乳中海藻糖-6-磷酸降低可能是种子发育异常的重要原因之一。基于种子发育特征, 该研究建立了百山祖冷杉未成熟胚培养技术体系, 并成功获得试管苗。
关键词:
Abstract
The seed germination rate of Abies beshanzuensis is low, and the characteristics of seed development are elusive, which severely limit the natural regeneration of the population in situ. To illustrate the developmental characteris-tics of seeds and identify the main factors affecting seed development, this article aimed to study the number, weight, and microstructural characteristics of the embryo and endosperm of the seed in the female cones at different development stages, as well as the primary metabolites of the endosperm in the critical period of seed development. The results showed that July was a critical period for seed development of A. beshanzuensis. During this period, the endosperm weight increased rapidly and the carbohydrate metabolism in the endosperm was active. Anatomical and morphological analysis of the female cones showed that a large number of abnormally developed seeds appeared in cones in late July. Analysis of the primary metabolites of endosperm during the critical period showed that the content of trehalose-6-phosp-hate in the endosperm of abnormally developed seeds was significantly decreased. It was speculated that during the critical period of seed development, the decrease of trehalose-6-phosphate in the endosperm may be an important cause of abnormal seed development. Based on the characteristics of A. beshanzuensis seed development, we established the embryo culture technology for A. beshanzuensis, and generated test-tube plantlet.
Keywords:
PDF (4754KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
刘柯, 刘彬, 袁璐, 帅辉, 杨洋, 王挺进, 陈德良, 陈小荣, 杨锴斌, 金孝锋, 陈利萍. 濒危植物百山祖冷杉种子发育特征及其胚培养. 植物学报, 2021, 56(5): 573-583 doi:10.11983/CBB21096
Liu Ke, Liu Bin, Yuan Lu, Shuai Hui, Yang Yang, Wang Tingjin, Chen Deliang, Chen Xiaorong, Yang Kaibin, Jin Xiaofeng, Chen Liping.
百山祖冷杉(Abies beshanzuensis)隶属松科(Pinaceae)冷杉属(Abies), 是我国特有的珍稀濒危野生植物。1963年首次在浙江省庆元县海拔约1 740 m的地区发现百山祖冷杉的自然分布, 1976年正式定名发表, 1987年被国际自然保护联盟物种生存委员会(International Union for Conservation of Nature-Species Survival Commission, IUCN-SSC)列为全球最濒危的12种植物之一, 1999年, 经国务院批准, 百山祖冷杉被列为国家一级重点保护野生植物。百山祖冷杉分布范围狭窄、开花结实周期长且天然更新能力极弱, 自然状态下繁育概率极低。目前, 全世界百山祖冷杉成年野生植株仅存3株, 处于极度濒危状态(李晓笑等, 2012)。
植物发生濒危的原因包括内部和外部两类因素。外部因素包括栖息地被破坏、外来物种引入、过度开发和环境污染等(蒋志刚和马克平, 2014; 徐刚标等, 2015)。内部因素主要表现为植物繁育系统障碍, 包括花粉限制、配子体败育、受精受阻、胚败育和种子不易萌发等(盛茂银等, 2011)。濒危植物繁育能力弱是其濒危的重要原因, 当生境发生剧变时, 濒危植物由于繁育系统存在障碍, 无法快速地恢复种群数量, 进而极大地增加其灭绝的可能性。
种子是植物有性繁殖的重要器官, 其发育直接决定了濒危植物的自然更新能力。裸子植物的种子主要由胚和胚乳组成, 受精后, 胚珠中的受精卵与雌配子体分别发育成胚与胚乳。前期研究发现, 百山祖冷杉成熟种子存在空粒和涩粒, 且萌发率极低等现象(吴友贵等, 2010)。因此, 研究百山祖冷杉种子生长发育特性, 明确种子发育异常的原因是解析百山祖冷杉内部致濒因素的关键。然而, 迄今为止关于百山祖冷杉种子生长发育的研究报道较少。
本研究以百山祖冷杉成年树为研究对象, 分别在2019年和2020年对传粉关键时间和传粉效率, 及不同发育时期球果中种子、胚与胚乳的数量和重量进行观察与统计, 明确种子发育的关键时期和发育特性。利用显微镜对发育关键时期的种子进行解剖和形态结构观察, 进一步明确种子发育关键时期异常种子的显微结构变化特征。利用超高效液相色谱串联质谱(ultra performance liquid chromatography-tandem mass spectrometry, UPLC-MS/MS)系统对胚乳快速生长时期的初生代谢物进行测定与分析, 明确影响胚乳发育的关键物质以及与种子发育异常的相关性。最后, 通过对未成熟胚培养的研究, 建立百山祖冷杉胚培养技术, 获得试管无菌苗。研究结果为揭示百山祖冷杉濒危机制提供了基础资料, 对百山祖冷杉的保护和繁育工作具有一定的理论与实际意义。
1 材料与方法
1.1 材料
试验材料取自生长于浙江省丽水市庆元县凤阳山-百山祖国家级自然保护区(27°42°N, 119°11°E, 海拔1 775 m)的百山祖冷杉(Abies beshanzuensis M.H. Wu)成年树(1978年繁育)。1.2 方法
1.2.1 传粉及授粉情况调查分别于2020年5月15日和5月25日对百山祖冷杉雌球果的传粉情况进行调查。在成年树球果集中分布区间(区间高度1-2 m)进行取样, 每次取样在上午同一时间进行, 选取大小一致的3个雌球果(图1)。随机选取每个球果中部(图1)的种鳞10片, 观察种鳞上的种子, 每个球果20粒种子, 3次生物学重复。将种子从种翅中分离出来, 从珠孔处起始, 将种子纵剖, 在体视显微镜(SZ61, Olympus)下观察并记录每个种子的珠孔开闭情况、花粉位置以及花粉数量。
图1
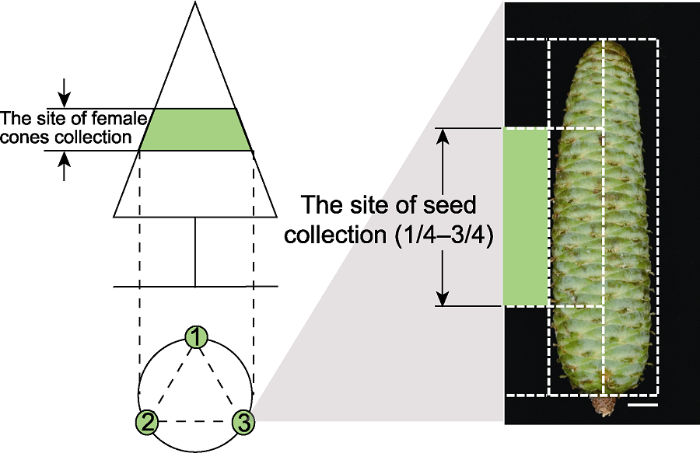 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1百山祖冷杉球果及种子取材示意图
Figure 1Diagram of the sampling of the cones and seeds of Abies beshanzuensis
1.2.2 种子及其胚和胚乳数量、重量统计
分别于2019年和2020年的6-11月的25日采摘百山祖冷杉成年树的雌球果, 统计种子及其胚与胚乳的数量和重量。取样方法见1.2.1节。选取球果中部的种子进行统计, 每个球果60粒种子, 3次生物学重复。将种子与种翅分离后, 立即称重; 去除种皮后, 分离胚和胚乳, 分别称重; 同时利用倒置显微镜(Primovert, Cari Zeisss)和体视显微镜对种子进行解剖观察, 统计种子的含胚率((胚发育正常种子数/种子总数) ×100%)和含胚乳率((胚乳发育正常种子数/种子总数) ×100%)。
1.2.3 胚和胚乳的形态学观察
采集2019年和2020年6-9月的25日球果, 随机选取每个球果中部(1-2层种鳞)的种鳞10片, 取种鳞上的种子, 去除种翅, 利用体视显微镜进行解剖观察, 记录种子及其胚与胚乳的形态结构特征。将种子去除种翅, 置于FAA溶液中固定24小时, 石蜡切片(刘向东和李亚娟, 2012), 番红固绿染色, 中性树胶封片后进行显微结构观察。
1.2.4 胚乳初生代谢物测定与差异代谢物筛选
选取百山祖冷杉6月25日发育正常的胚乳(6_NE)和7月25日发育正常的胚乳(7_NE)与发育异常的胚乳(7_AE)进行初生代谢物测定, 每组样品3次生物学重复。参考Chen等(2013)的方法进行样品制备和提取。获得原始数据后, 进行中心化处理。利用R软件中的Metabo Analyst R包OPLSR.Anal函数进行正交偏最小二乘法判别分析(orthogonal partial least squares discrimination analysis, OPLS-DA)。选择贡献较大的变量, 即VIP (variable importance in projection) ≥1, 差异倍数值FC (fold change) ≥2, 筛选不同类型胚乳的差异代谢物。
1.2.5 胚培养
采集6-11月百山祖冷杉成年树雌球果(每月25日采集1次), 随机选取球果中部21粒发育正常的种子, 3次生物学重复。将种子从球果上分离, 用流水冲洗3小时, 在超净工作台上用70%乙醇处理30秒, 无菌水冲洗3-5次。再用0.1 wt%氯化汞处理10分钟, 无菌水冲洗3-5次, 吸干种子表面水分。剥取胚及其胚乳并接种至培养基(以DCR (Gupta and Durzan medium)为基本培养基), 另外添加蔗糖(20 g·L-1)、水解酪蛋白(500 mg·L-1)和琼脂(8 g·L-1), pH值为5.8。接种后先于(20±2)°C黑暗下培养3天, 再于弱光下培养10天, 统计萌发率(萌发率=(萌发个数/接种个数) ×100%)。
1.2.6 数据统计与分析
利用IBM SPSS 20.0软件对实验数据进行统计, 采用LSD检验的方差分析方法(P<0.05)进行多重比较。
2 结果与讨论
2.1 传粉后不同发育时期球果中含胚率与含胚乳率变化
为明确百山祖冷杉传粉过程, 我们利用体视显微镜对雌球果进行解剖及观察, 结果显示5月15日珠孔张开且其边缘发育形成沟槽(呈漏斗状结构) (图2A), 最长直径为779.59 μm, 此时珠孔周围可见淡黄色花粉(图2B), 表明百山祖冷杉正处于传粉阶段。统计结果表明, 63.33%的胚珠珠孔外可见花粉粒附着。5月25日观察发现珠孔边缘内折(图2C), 花粉进入储粉室且颜色变浅, 体积增大(图2D), 表明传粉阶段结束, 此时百山祖冷杉处于受精前阶段。统计显示, 46.67%的胚珠珠孔内进入了花粉粒, 平均每个珠孔内有4.2个花粉, 表明5月下旬百山祖冷杉完成了传粉过程, 大部分胚珠具备发育成种子的潜力。与大多数被子植物相比, 裸子植物的花粉萌发与花粉管生长速度极其缓慢。冷杉属植物从传粉开始至受精需要2个月(王伏雄, 1990; 林金星等, 2013)。大多数冷杉树种传粉持续时间为18-30天(Politi et al., 2011)。由此确定6月25日为百山祖冷杉种子发育特性观察的初始时间。图2
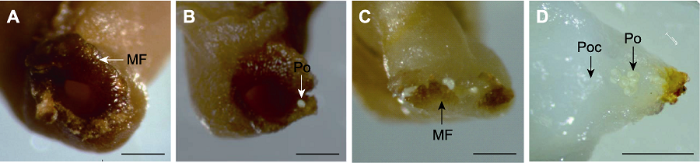 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2百山祖冷杉传粉阶段的珠孔形态变化和花粉转移
(A) 珠被发育形成漏斗状珠孔(箭头); (B) 花粉(箭头)附着于珠孔边缘; (C) 珠被向内折叠, 珠孔闭合(箭头); (D) 花粉(箭头)进入储粉室。MF: 漏斗状珠孔; Po: 花粉; Poc: 储粉室。(A)-(C) Bars=100 μm; (D) Bar=500 μm
Figure 2Morphological changes of the micropylar funnel and pollen transfer during the pollination stage of Abies beshanzuensis
(A) The integument develops a stigmatic micropylar funnel (arrow); (B) The pollen (arrow) attaches to the edge of micropylar funnel; (C) The integument folds inward and the micropylar funnel is closed (arrow); (D) The pollen (arrow) enters the pollen chamber. MF: Micropylar funnel; Po: Pollen; Poc: Pollen chamber. (A)-(C) Bars=100 μm; (D) Bar=500 μm
种子发育过程中, 胚乳的发育动态直接影响胚的发育(Linkies et al., 2010; 刘美善和刘宝, 2012; 李卫星等, 2016; Yuan et al., 2018)。数量变化是胚和胚乳发育动态的直接表现。我们分别对2019年和2020年6-11月的雌球果中胚和胚乳的数量变化进行了统计, 发现百山祖冷杉球果的含胚乳率(图3A)和含胚率(图3B)在发育过程中整体呈递减趋势。2019年的6-7月, 含胚乳率从76.11%迅速下降至15.00%, 而7-11月下降速率明显减缓。2020年胚乳数量变化规律与2019年相似, 但2020年各月的含胚乳率明显高于2019年。观察百山祖冷杉球果中胚的数量变化, 发现2019年6-8月的百山祖冷杉含胚种子数显著减少, 含胚率从33.33%降至3.89%, 8-11月则无显著变化。2020年6-11月各月百山祖冷杉球果中含胚率均高于2019年。以上结果表明, 6-7月, 百山祖冷杉球果的含胚率与含胚乳率短时间迅速降低, 推测在此期间其种子可能存在大量发育异常现象。
图3
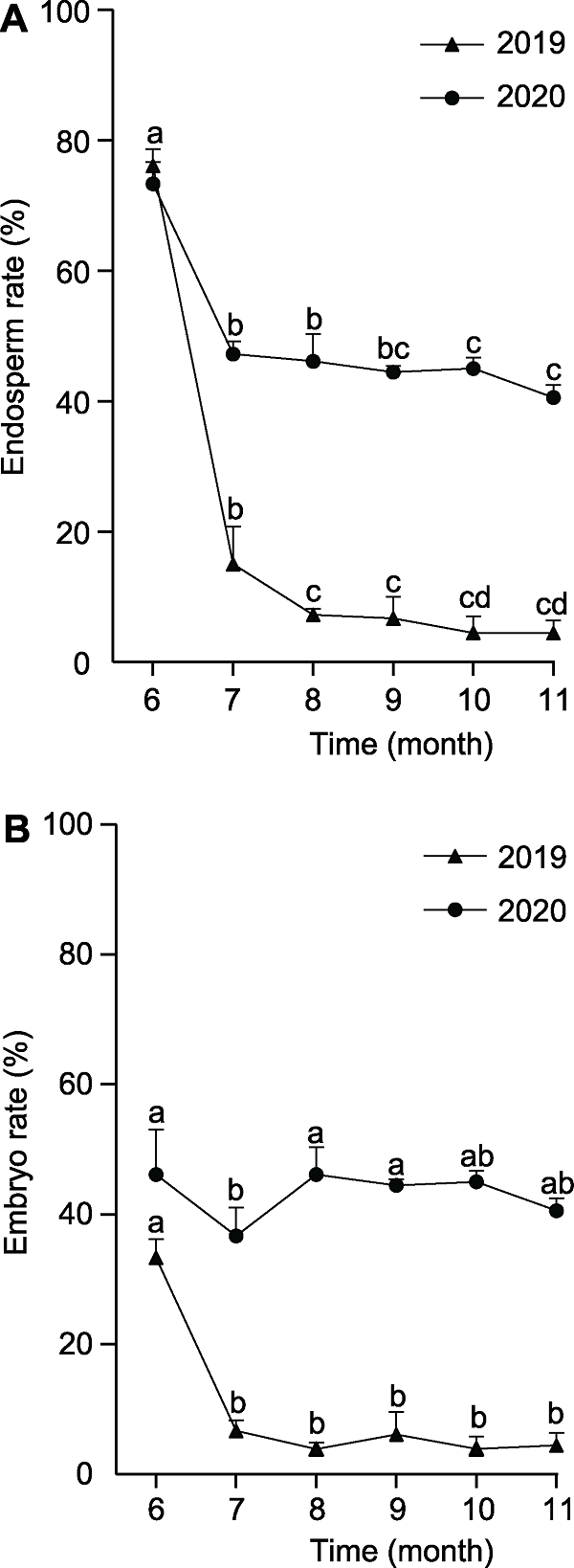 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图32019年和2020年百山祖冷杉不同发育时期球果含胚乳率和含胚率变化
(A) 百山祖冷杉球果含胚乳率随时间的变化; (B) 百山祖冷杉球果含胚率随时间的变化。不同小写字母表示在0.05水平差异显著。
Figure 3The endosperm rate and embryo rate of Abies beshanzuensis cones at different developmental stages in 2019 and 2020
(A) Changes of endosperm rate of A. beshanzuensis seeds with time; (B) Changes of embryo rate of A. beshanzuensis seeds with time. Different lowercase letters indicate significant differences at 0.05 level.
2.2 传粉后不同发育时期种子及其胚与胚乳重量变化
种子发育过程中, 胚和胚乳的细胞数目、体积和物质组成不断变化, 随之重量也发生改变。通过对2019年和2020年百山祖冷杉6-11月的种子、胚和胚乳重量进行测定, 发现6-8月种子的平均重量增加了14.34- 20.14 mg, 8-9月变化不显著, 之后逐月降低(图4A)。2019年6-7月百山祖冷杉的胚乳重量无显著变化, 7- 8月则由8.70 mg迅速上升至24.83 mg, 9月达到最大值(32.07 mg), 之后逐渐下降(图4B); 胚的重量增长主要集中在7-8月(图4B)。2020年百山祖冷杉胚乳和胚重量的总体变化趋势与2019年相似。2020年胚乳重量从6月开始增长(图4C); 7-8月迅速增长, 8-9月增长速率缓慢(图4C)。上述结果表明, 6-7月为胚乳的快速生长阶段; 7-9月, 种子、胚和胚乳重量则同步增长。结合球果的含胚率与含胚乳率变化, 发现胚乳在快速生长阶段, 球果中胚和胚乳的数量骤减, 进一步说明7月是百山祖冷杉种子发育的关键时期。图4
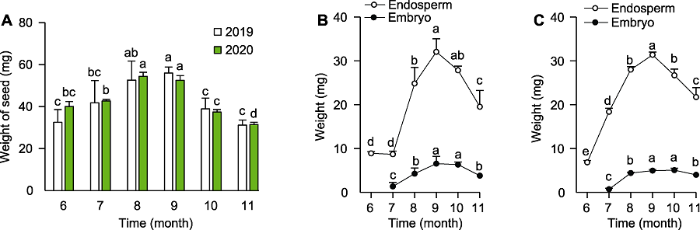 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图42019年与2020年百山祖冷杉不同发育时期种子及其胚和胚乳重量变化
(A) 2019年与2020年种子重量随时间的变化; (B) 2019年胚与胚乳重量随时间的变化; (C) 2020年胚与胚乳重量随时间的变化。不同小写字母表示在0.05水平差异显著。
Figure 4Changes in seed, embryo and endosperm weight of Abies beshanzuensis seeds at different developmental stages in 2019 and 2020
(A) Changes in the weight of seed with time in 2019 and 2020; (B) Changes in the weight of embryo and endosperm with time in 2019; (C) Changes in the weight of embryo and endosperm with time in 2020. Different lowercase letters indicate significant differences at 0.05 level.
2.3 种子重量增长时期胚与胚乳的形态结构变化
为明确百山祖冷杉种子在重量增长时期的内部结构变化, 探究种子发育关键时期是否存在发育异常现象, 我们对6-9月百山祖冷杉胚和胚乳进行了显微观察, 发现5月下旬, 花粉进入储粉室, 珠孔完全闭合, 珠被呈淡黄色(图5A); 雌配子体偏合点区域形成囊状结构, 游离核向中心分裂, 珠被上已形成树脂囊(图5D)。图5
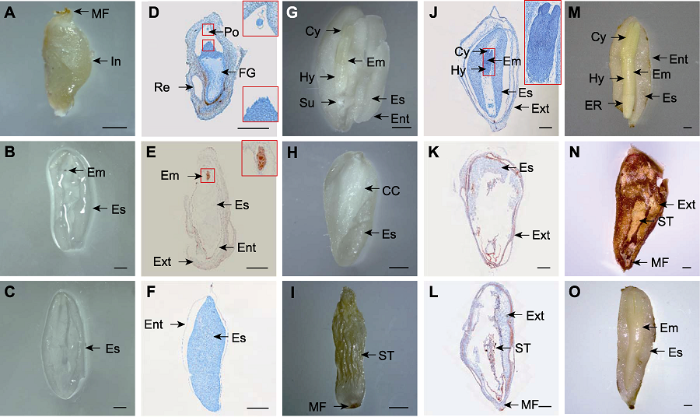 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5百山祖冷杉种子及其胚与胚乳发育过程的形态结构
(A) 5月25日的胚珠; (B) 6月25日类型I种子的胚与胚乳; (C) 6月25日类型II种子的胚乳; (D) 5月25日胚珠的石蜡切片, 红色方框内为花粉(上)与颈卵器(下); (E) 6月25日类型I种子的石蜡切片, 红色方框内为胚; (F) 6月25日类型II种子的石蜡切片; (G) 7月25日类型I种子的胚与胚乳; (H) 7月25日类型II种子的胚乳; (I) 7月25日类型III种子内皱缩的胚乳或/和胚; (J) 7月25日类型I种子的石蜡切片, 红色方框内为胚; (K) 7月25日类型II种子的石蜡切片; (L) 7月25日类型III种子的石蜡切片; (M) 8月25日类型I种子的胚与胚乳; (N) 8月25日类型III种子; (O) 9月25日类型I种子的胚与胚乳。Bars=1 000 μm。MF: 珠孔; In: 珠被; Em: 胚; Es: 胚乳; Po: 花粉; FG: 雌配子体; Re: 树脂囊; Ext: 外种皮; Ent: 内种皮; Cy: 子叶; Hy: 下胚轴; Su: 胚柄系统; ST: 皱缩组织; ER: 胚根; CC: 腐蚀腔
Figure 5Morphology of Abies beshanzuensis seed and its embryo and endosperm during development
(A) The ovule on May 25th; (B) The embryo and endosperm of type I seeds on June 25th; (C) The endosperm of type II seeds on June 25th; (D) The paraffin section of the ovule on May 25th, the pollen (top) and archegonium (bottom) are in the red box; (E) The paraffin section of type I seeds on June 25th, the embryo in the red box; (F) The paraffin section of type II seeds on June 25th; (G) The embryo and endosperm of type I seeds on July 25th; (H) The endosperm of type II seeds on July 25th; (I) The shriveled tissue of endosperm or/and embryo within type III seeds on July 25th; (J) The paraffin section of type I seeds on July 25th, the embryo in the red box; (K) The paraffin section of type II seeds on July 25th; (L) The paraffin section of type III seeds on July 25th; (M) The embryo and endosperm of type I seeds on August 25th; (N) Type III seeds on August 25th; (O) The embryo and endosperm of type I seeds on September 25th. Bars=1 000 μm. MF: Micropylar funnel; In: Integument; Em: Embryo; Es: Endosperm; Po: Pollen; FG: Female gametophyte; Re: Resinocyst; Ext: Exotesta; Ent: Endotesta; Cy: Cotyledon; Hy: Hypocotyl; Su: Suspensor system; ST: Shriveled tissue; ER: Embryo root; CC: Corrosion cavity
6月下旬, 受精作用完成, 在发育正常的种子中, 雌配子体(胚乳)呈白色半透明胶质状(图5B); 珠孔端可见457.60 μm × 160.01 μm (长×宽)的幼胚(图5E); 内、外珠被分别发育成内、外种皮(图5E), 不可分离, 外种皮呈褐色。未受精的胚珠中雌配子体继续发育, 与已受精的种子中的胚乳状态相似(图5C, F)。发育异常的种子中, 雌配子体(胚乳)皱缩成白色片状, 种子外形干瘪, 外观与发育正常的种子有明显差异。
7月下旬, 胚分化形成子叶, 雌球果中存在3种类型的种子。类型I, 发育正常, 主要特征: 胚乳呈乳白色; 胚发育形成子叶, 子叶长度约为胚长度的1/4, 呈淡黄色, 易与胚乳分离; 胚柄系统与优势胚分离, 程序性死亡; 内、外种皮继续发育, 内种皮呈无色透明膜状结构, 易与胚乳分离, 外种皮木质化程度增高, 外种皮上树脂囊区域的厚度较外种皮其它位置更薄, 呈膜状, 其中含有大量的芳香液体(图5G, J)。类型II, 胚乳继续发育、未形成胚或胚消失, 主要特征: 胚乳总体呈乳白色, 珠孔端与合点端呈半透明状, 胚乳中心具有与发育正常种子相似的空腔, 但未发现胚(图5H, K)。类型III, 发育异常, 主要特征: 胚乳和胚皱缩成白色或淡黄色片状, 与发育正常的种子相比, 木质化程度更高(图5I, L)。
8月下旬, 类型I种子进一步发育, 解剖观察发现, 胚乳呈半透明, 胚结构分化明显, 形成子叶、胚轴和胚根等结构, 子叶长度约为胚长度的1/3, 呈黄色; 内种皮颜色变褐, 质地变薄变硬, 可与胚乳分离, 外种皮无明显变化(图5M)。大量的类型II种子转变为类型III种子状态(图5N)。
9月下旬, 类型I种子的解剖结构与8月相似, 主要区别在于胚乳厚度减小, 胚乳中脂类物质增加; 内种皮与胚乳不易分离(图5O)。类型II种子几乎全部消失。
2.4 胚乳快速生长阶段的初生代谢变化
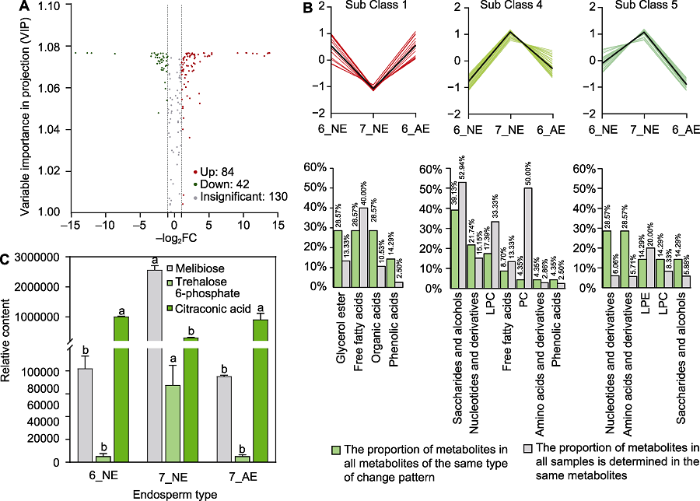
为探究百山祖冷杉种子在胚乳快速生长阶段发育异常种子数迅速增长的原因, 我们利用UPLC-MS/MS检测技术对6月和7月种子中胚乳的初生代谢物类型和含量进行测定。2个时期共检测出256种代谢物, 与6月发育正常的胚乳相比, 7月发育正常的胚乳中有84种代谢物含量显著增加, 42种显著降低(图6A)。图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图66月和7月百山祖冷杉胚乳的初生代谢物变化
(A) 6月和7月发育正常胚乳的差异代谢物; (B) 第1、4、5类模式中关键差异代谢物数量占每类模式全部代谢物总数的比例与占所有被测样品中同类代谢物总数的比例; (C) 6月和7月百山祖冷杉种子胚乳中的蜜二糖、海藻糖-6-磷酸和甲基马来酸相对含量。6_NE: 6月发育正常的胚乳; 7_NE: 7月发育正常的胚乳; 7_AE: 7月发育异常的胚乳。不同小写字母表示在0.05水平差异显著。FC: 差异倍数值; LPC: 溶血磷脂酰胆碱; LPE: 溶血磷脂酰乙醇胺; PC: 磷脂酰胆碱
Figure 6Changes of primary metabolites in endosperm of Abies beshanzuensis in June and July
(A) Differential metabolites of normal endosperm in June and July; (B) The proportion of key difference metabolites in the first, fourth, and fifth models in the same change pattern in all metabolites and the proportion of the same metabolites tested in all samples; (C) Relative content of melibiose, trehalose-6-phosphate and methylmaleic acid in endosperm of A. beshanzuensis seeds in June and July. 6_NE: Normal endosperm in June; 7_NE: Normal endosperm in July; 7_AE: Abnormal endosperm in July. Different lowercase letters indicate significant differences at 0.05 level. FC: Fold change; LPC: Lysophosphatidylcholine; LPE: Lysopnosphatidylethanolamine; PC: Phosphatidylcholine
将6月和7月发育正常的胚乳(6_NE和7_NE)及7月发育异常的胚乳(7_AE) 3种类型样本中的代谢物进行K均值(K means)聚类分析, 发现代谢物在不同样本中存在6类不同的变化模式。其中, 第1类代谢物在发育正常的胚乳中的相对含量降低, 而在发育异常的胚乳中其相对含量维持在6月的水平(图6B), 推测此类代谢物含量与正常胚乳在关键时期的发育呈负相关。第4类代谢物在发育正常的胚乳中相对含量升高, 而在7月发育异常的胚乳中相对含量显著低于7月正常的胚乳(图6B)。第5类代谢物在发育正常的胚乳中的变化模式与第4类代谢物相似, 而在7月发育异常的胚乳中的相对含量显著低于6月正常的胚乳(图6B), 推测第4类与第5类代谢物含量与胚乳在关键时期的发育呈正相关。
为明确7月胚乳发育异常的原因, 我们进一步对上述3类模式中的代谢物进行分析, 发现第1类模式中的关键代谢物(7_NE中的含量分别显著低于6_NE和7_AE的代谢物)为甘油酯类、游离脂肪酸类和有机酸类物质, 其中游离脂肪酸类代谢物数量占所有被测样品中同类代谢物总数的40.00% (图6B), 说明7月异常发育胚乳中游离脂肪酸类物质的分解代谢被抑制。第4类模式中的关键代谢物(7_NE中的含量分别显著高于6_NE以及7_AE的代谢物)为糖及醇类、核苷酸及其衍生物类和溶血磷脂酰胆碱(lysophosphati-dylcholine, LPC)类物质, 其中糖及醇类代谢物数量占所有被测样品中同类代谢物总数的52.94% (图6B); 第5类模式中的关键代谢物(7_NE中的含量显著高于6_NE, 且7_AE中的含量显著低于6_NE)为核苷酸及其衍生物、氨基酸及其衍生物和溶血磷脂酰乙醇胺(lysopnosphatidylethanolamine, LPE)类物质(图6B)。上述结果表明, 胚乳在发育关键时期可能需要第4类模式中的糖及醇类物质提供能量, 且第1类模式中游离脂肪酸类物质的积累可能不利于胚乳发育。
利用KEGG数据库对3类模式中的关键代谢物进行注释和代谢途径分析(Kanehisa and Goto, 2000), 发现大量关键差异代谢物参与碳水化合物代谢途径, 包括半乳糖代谢、蔗糖与淀粉代谢及C5-二元酸代谢。对参与碳水化合物代谢途径中关键差异代谢物含量进行分析, 发现7_NE中的蜜二糖和海藻糖-6-磷酸(trehalose 6-phosphate, T6P)含量比6_NE显著增加(图6C), 甲基马来酸含量显著降低(图6C); 与7_NE相比, 7_AE中的海藻糖-6-磷酸含量显著降低(图6C)。已有研究表明, 海藻糖-6-磷酸可通过维持糖代谢平衡进而调控种子发育(Fichtner and Lunn, 2021)。本研究发现, 百山祖冷杉种子发育关键时期, 胚乳中碳水化合物代谢旺盛, 为其快速生长提供了能量来源; 而胚乳中海藻糖-6-磷酸含量降低, 可能会破坏碳水化合物代谢(糖代谢)平衡, 进而导致种子发育异常。
2.5 未成熟胚培养及其试管苗的获得
百山祖冷杉种子重量在6-8月迅速增长(图4A), 胚乳中碳水化合物代谢旺盛(图6B)。 然而, 7月开始, 大量种子胚乳中的海藻糖-6-磷酸等物质代谢异常, 发育正常的种子数量骤减(图3)。为解决百山祖冷杉种子自然状态下因代谢异常导致的种子发育问题, 本研究尝试利用胚培养技术对百山祖冷杉的胚进行离体培养(图7A, B)。通过比较百山祖冷杉不同发育时期种子中胚培养的萌发率, 发现胚的萌发率从6-11月呈先上升后下降的趋势。其中, 8月和9月的萌发率最高, 分别为86.67%和93.65% (图7C), 高于7月胚的萌发率(54.17%)。6月的胚处于发育早期, 尚未形成完整植株。此外, 10月开始, 胚的萌发率逐渐降低, 根据对种子的显微观察, 此时胚已分化形成子叶, 接近成熟, 胚萌发率低的原因可能是种子将进入休眠状态, 大量抑制胚萌发物质的逐渐积累。图7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7百山祖冷杉不同种子发育时期的胚培养萌发率变化
(A) 百山祖冷杉胚萌发(bar=200 μm); (B) 胚培养获得的试管苗(bar=2 cm); (C) 不同百山祖冷杉种子发育时期的胚培养萌发率变化。不同小写字母表示在0.05水平差异显著。
Figure 7Changes in germination rate of embryo of Abies beshanzuensis seeds at different developmental stages
(A) Embryo germination (bar=200 μm); (B) Seedlings obtained by embryo culture (bar=2 cm); (C) Changes in germination rate of embryo of A. beshanzuensis seeds at different developmental stages. Different lowercase letters indicate significant differences at 0.05 level.
2.6 讨论
百山祖冷杉是松科冷杉属植物, 与大多数松柏纲植物(针叶树)相似, 孢子叶球单性, 雌雄同株(姜在民和贺学礼, 2009), 花粉从雄配子体产生后进入珠孔期间受到开花时间、地理位置和气候条件等诸多因素的影响, 有效传粉是保障种子正常发育的重要前提条件。本研究发现, 百山祖冷杉与大部分冷杉属植物的传粉时间(5月中旬)相似(Singh and Owens, 1982; Owens and Morris, 1998; 樊金拴, 2007; Politi et al., 2011)。传粉结束后(5月下旬), 46.67%的胚珠内进入花粉, 具备了受精条件。6月下旬, 73.00%的雌配子体(胚乳)继续发育, 包括未受精的雌配子体。8月下旬, 仅有46.11%的胚珠中有正常发育的胚乳, 未授粉的胚珠几乎全部发育中止, 导致发育异常种子的数量剧增。冷杉属植物无传粉滴或传粉滴不从珠孔处溢出, 花粉进入珠孔内依赖雨水的作用(Owens et al., 1998), 传粉过程极大地受到气候的制约, 因此, 普遍存在传粉效率低的现象(Politi et al., 2011)。温哥华冷杉(A. amabilis)雌配子体败育发生在2个时期: (1) 游离核中期到后期阶段; (2) 传粉结束1周后, 未授粉的雌配子体发生败育。其中, 第2个时期(6月末至7月)是温哥华冷杉雌配子体败育的主要时期。温哥华冷杉第2个时期雌配子体败育种子的解剖结构与百山祖冷杉类型III部分种子非常相似, 即种子内残留皱缩的组织, 珠被细胞增殖, 种皮变厚(Owens and Morris, 1998)。在松科植物繁育过程中, 利用人工辅助授粉可有效克服因环境条件限制导致的异花传粉困难, 提高植株的传粉和受精机会, 进而增加种子的数量。Iwaizumi和Takahashi (2012)通过对银叶五针松(Pinus monticola)进行不同方式的人工授粉, 发现花粉的数量和质量分别对种子的形成与发育有显著影响。因此, 为确保百山祖冷杉种子的正常发育, 提高球果中胚珠的受精率, 可在5月中旬降雨前进行人工辅助授粉。百山祖冷杉传粉后, 种子重量快速增长, 正常发育的种子数量却急剧下降。代谢组分析表明, 在此期间胚乳的碳水化合物代谢旺盛, 尤其是多个糖类代谢途径。由于每个球果中含有大量的种子(302-384粒), 因此, 种子之间对糖类等能源物质的竞争可能导致部分胚乳发育异常, 最终形成类型II/III种子。与被子植物相比, 目前有关裸子植物种子败育与营养物质供应关系的研究较少。拟南芥(Arabidopsis thaliana)在生殖生长过程中, 同化物匮乏会导致正在发育的种子甚至整个角果发生败育(Lauxmann et al., 2016)。Shen等(2018)研究发现, 玉米(Zea mays)果穗顶部种子由于授粉时间延迟引起的库活性降低, 导致同化物供应不足, 最终发生败育。在裸子和被子植物种子发育过程中, 胚乳是胚发育过程中的主要营养来源, 对大豆(Glycine max)的研究发现, 胚乳的营养物质(糖)供应不足会导致严重的种子败育(Wang et al., 2019)。此外, 2020年是百山祖冷杉成年树结果的大年, 相比2019年(小年), 6-8月胚和胚乳的数量出现波动性下降或下降幅度明显减小。Haim等(2021)研究表明, 营养竞争所导致的营养分配不均衡是多年生木本植物产生大小年现象的重要原因之一, 进一步说明, 营养竞争可能是导致百山祖冷杉种子发育异常的直接原因。在希腊冷杉(A. cephalonic)繁殖生物学研究中也发现相似现象, 大年饱满种子的比例为26.6%, 小年仅有1%的种子饱满。Politi等(2011)推测这可能是希腊冷杉根据植物体内营养的波动改变其繁殖力。在其它木本植物栽培管理中, 通过合理的树体营养管理可有效降低大小年出现的频率(张松文, 2016)。因此, 通过进一步探究百山祖冷杉营养利用与濒危的关系, 有望解析百山祖冷杉濒危的原因。
本研究发现异常胚乳中的海藻糖-6-磷酸(T6P)含量显著低于正常的胚乳, 而T6P是海藻糖的代谢前体, 可通过调控流向糖酵解的糖量, 维持代谢平衡进而调控胚胎发育(Fichtner and Lunn, 2021)。模式植物中, T6P合成酶基因TPS1突变直接导致胚胎在鱼雷期向子叶期早期转变时发生败育(Eastmond et al., 2002)。Meitzel等(2021)研究发现豌豆(Pisum sati-vum)胚发育过程中, T6P通过调控生长素的合成促进种子中的物质积累和发育。因此, 推测T6P可能通过调控百山祖冷杉胚乳中糖代谢平衡进而影响种子的正常发育。利用胚培养技术, 在百山祖冷杉胚乳未出现代谢异常时, 对未成熟胚进行早期离体培养, 为其提供充分营养和适宜的生长环境, 可最大限度地保留百山祖冷杉有性繁殖后代。胚龄和培养基成分是胚培养成功的关键(贺佳玉等, 2008), 本研究比较了不同发育时期种子的胚培养效果, 发现8-9月发育正常种子的胚萌发率最高, 此时, 百山祖冷杉种子及其胚与胚乳的重量达到峰值, 胚发育至早期成熟阶段; 6-7月发育正常种子中的胚处于发育早期, 从生理至形态上远未成熟, 对培养条件要求很高, 尤其是培养基的营养成分需要更加全面, 以满足胚胎发育的要求, 因此胚的萌发率相对较低。然而, 7月球果中发育正常种子的数量处于转折时期, 此时是拯救发育异常种子的最佳时期。未来研究中, 可通过进一步优化培养基成分, 提高百山祖冷杉早期胚培养效果, 克服百山祖冷杉发育不良或中途败育的缺陷, 增加群体遗传多样性。
2.7 结论
7月是百山祖冷杉种子发育的关键时期, 在此期间, 胚乳重量快速增长, 胚乳中碳水化合物代谢活动旺盛。7月下旬球果中出现大量发育异常的种子, 发育异常种子中胚乳的海藻糖-6-磷酸含量显著降低。利用胚培养技术在其发育异常前, 对7月种子中的胚进行培养, 可有效提高有性后代的数量, 对百山祖冷杉保护和人工繁育均具有重要意义。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3724/SP.J.1258.2012.01154 [本文引用: 1]

通过野外实地调查和文献调研, 分析4种极危冷杉属(Abies)植物的地理分布现状和特征, 构建其分布区的气候数据库, 采用国际上常用的研究植被与气候关系的指标和方法, 定量分析冷杉属地理分布与水热条件的关系, 并通过因子分析方法确定影响冷杉属地理分布的限制性气候因子。结果表明: 1)百山祖冷杉(Abies beshanzuensis)、梵净山冷杉(A. fanjingshanensis)、元宝山冷杉(A. yuanbaoshanensis)和资源冷杉(A. beshanzuensis var. ziyuanensis)零星分布于我国亚热带地区, 水平分布范围为25°25′–27°55′ N, 108°42′–119°12′ E, 分布范围狭窄; 垂直分布范围为1 363–2 390 m, 均分布于山体上部。海拔上限高度随着经度的减小而增加。2)整个分布区的气温较低, 年平均气温为7.4–11.2 ℃; 降水较为充沛, 年降水量为1 265.4–1 945.7 mm, 其中生长季降水量占74.78%; 湿度较大, 年相对湿度为77%–86%。气候指标范围狭窄, 表明对水热条件要求很高。分布区寒冷指数较低, 限制其垂直向下扩散, 呈现局限分布于山体上部的现状。3) 4种冷杉各分布区的气候条件基本相近, 梵净山冷杉分布区气温相对偏低, 元宝山冷杉分布区湿度相对偏大。4) 3个气候因子对这4种冷杉的地理分布影响较大, 其作用排序为: 低温因子>极端低温因子>湿度因子。在全球气候变暖的大背景下, 这4种冷杉自然分布区的适宜生境将进一步缩小, 直至物种灭绝, 急需开展迁地保护工作。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3724/SP.J.1259.2012.00101 [本文引用: 1]
被子植物的种子发育从双受精开始, 产生二倍体的胚和三倍体的胚乳。在种子发育和萌发过程中, 胚乳向胚组织提供营养物质, 因此胚乳对胚和种子的正常生长发育至关重要。开花植物发生基因组印迹的主要器官是胚乳。印迹基因的表达受表观遗传学机制的调控, 包括DNA甲基化和组蛋白H3K27甲基化修饰以及依赖于PolIV的siRNAs (p4-siRNAs)调控。基因组印迹的表观遗传学调控对胚乳的正常发育和种子育性具有不可或缺的重要作用。最新研究显示, 胚乳的整个基因组DNA甲基化水平降低, 而且去甲基化作用可能源于雌配子体的中央细胞。该文综述了种子发育的表观遗传学调控机制, 包括基因组印迹机制以及胚乳基因组DNA甲基化变化研究的最新进展。
[本文引用: 1]
DOI:10.1093/mp/sst080PMID:23702596 [本文引用: 1]
Liquid chromatography-mass spectrometry (LC-MS)-based metabolomics has been facilitated by the construction of MS(2) spectral tag (MS2T) library from the total scan ESI MS/MS data, and the development of widely targeted metabolomics method using MS/MS data gathered from authentic standards. In this report, a novel strategy called stepwise multiple ion monitoring-enhanced product ions (stepwise MIM-EPI) was developed to construct the MS2T library, in which stepwise MIM was used as survey scans to trigger the acquisition of EPI. A total number of 698 (almost) non-redundant metabolites with MS(2) spectra were obtained, of which 135 metabolites were identified/annotated. Integrating the data gathered from our MS2T library and other available multiple reaction monitoring (MRM) information, a widely targeted metabolomics method was developed to quantify 277 metabolites, including some phytohormones. Evaluation of the dehydration responses and natural variations of these metabolites in rice leaf not only suggested the coordinated regulation of abscisic acid (ABA) with metabolites such as serotonin derivative(s), polyamine conjugates under drought stress, but also revealed some C-glycosylated flavones as the potential markers for the discrimination of indica and japonica rice subspecies. The new MS2T library construction and widely targeted metabolomics strategy could be used as a tool for rice functional genomics.
PMID:11851922 [本文引用: 1]
Despite the recent discovery that trehalose synthesis is widespread in higher plants very little is known about its physiological significance. Here we report on an Arabidopsis mutant (tps1), disrupted in a gene encoding the first enzyme of trehalose biosynthesis (trehalose-6-phosphate synthase). The tps1 mutant is a recessive embryo lethal. Embryo morphogenesis is normal but development is retarded and stalls early in the phase of cell expansion and storage reserve accumulation. TPS1 is transiently up-regulated at this same developmental stage and is required for the full expression of seed maturation marker genes (2S2 and OLEOSN2). Sucrose levels also increase rapidly in seeds during the onset of cell expansion. In Saccharomyces cerevisiae trehalose-6-phosphate (T-6-P) is required to regulate sugar influx into glycolysis via the inhibition of hexokinase and a deficiency in TPS1 prevents growth on sugars (Thevelein and Hohmann, 1995). The growth of Arabidopsis tps1-1 embryos can be partially rescued in vitro by reducing the sucrose level. However, T-6-P is not an inhibitor of AtHXK1 or AtHXK2. Nor does reducing hexokinase activity rescue tps1-1 embryo growth. Our data establish for the first time that an enzyme of trehalose metabolism is essential in plants and is implicated in the regulation of sugar metabolism/embryo development via a different mechanism to that reported in S. cerevisiae.
DOI:10.1146/arplant.2021.72.issue-1URL [本文引用: 2]
DOI:10.1093/jxb/eraa590PMID:33345278 [本文引用: 1]
In many fruit trees, heavy fruit load in one year reduces flowering in the following year, creating a biennial fluctuation in yield termed alternate bearing (AB). In subtropical trees, where flowering induction is mostly governed by the accumulation of chilling hours, fruit load is thought to generate a signal (AB signal) that blocks the perception of cold induction. Fruit removal during a heavy-fruit-load year is effective at inducing flowering only if performed one to a few months before the onset of the flowering induction period. We previously showed that following fruit removal, the content of the auxin indoleacetic acid (IAA) in citrus buds is reduced, suggesting that the hormone plays a role in the AB signal. Here, we demonstrate that fruit presence generates relatively strong polar auxin transport in citrus and olive stems. Upon fruit removal, polar auxin transport is reduced and allows auxin release from the bud. Furthermore, using immunolocalization, hormone, and gene expression analyses, we show that in citrus, IAA level in the bud and specifically in the apical meristem is reduced upon fruit removal. Overall, our data provide support for the notion that fruit presence generates an auxin signal in the bud, which may affect flowering induction.© The Author(s) 2020. Published by Oxford University Press on behalf of the Society for Experimental Biology. All rights reserved. For permissions, please email: journals.permissions@oup.com.
DOI:10.1007/s10265-011-0463-5URL [本文引用: 1]
PMID:10592173 [本文引用: 1]
KEGG (Kyoto Encyclopedia of Genes and Genomes) is a knowledge base for systematic analysis of gene functions, linking genomic information with higher order functional information. The genomic information is stored in the GENES database, which is a collection of gene catalogs for all the completely sequenced genomes and some partial genomes with up-to-date annotation of gene functions. The higher order functional information is stored in the PATHWAY database, which contains graphical representations of cellular processes, such as metabolism, membrane transport, signal transduction and cell cycle. The PATHWAY database is supplemented by a set of ortholog group tables for the information about conserved subpathways (pathway motifs), which are often encoded by positionally coupled genes on the chromosome and which are especially useful in predicting gene functions. A third database in KEGG is LIGAND for the information about chemical compounds, enzyme molecules and enzymatic reactions. KEGG provides Java graphics tools for browsing genome maps, comparing two genome maps and manipulating expression maps, as well as computational tools for sequence comparison, graph comparison and path computation. The KEGG databases are daily updated and made freely available (http://www. genome.ad.jp/kegg/).
DOI:10.1111/pce.12634URL [本文引用: 1]
DOI:10.1111/j.1469-8137.2010.03249.xPMID:20406407 [本文引用: 1]
The evolution of the seed represents a remarkable life-history transition for photosynthetic organisms. Here, we review the recent literature and historical understanding of how and why seeds evolved. Answering the 'how' question involves a detailed understanding of the developmental morphology and anatomy of seeds, as well as the genetic programs that determine seed size. We complement this with a special emphasis on the evolution of dormancy, the characteristic of seeds that allows for long 'distance' time travel. Answering the 'why' question involves proposed hypotheses of how natural selection has operated to favor the seed life-history phenomenon. The recent flurry of research describing the comparative biology of seeds is discussed. The review will be divided into sections dealing with: (1) the development and anatomy of seeds; (2) the endosperm; (3) dormancy; (4) early seed-like structures and the transition to seeds; and (5) the evolution of seed size (mass). In many cases, a special distinction is made between angiosperm and gymnosperm seeds. Finally, we make some recommendations for future research in seed biology.
[本文引用: 1]
DOI:10.1139/x98-089URL [本文引用: 2]
DOI:10.1016/S1360-1385(98)01337-5URL [本文引用: 1]
DOI:10.1007/s00468-011-0542-1URL [本文引用: 4]
DOI:10.1093/jxb/ery013PMID:29365129 [本文引用: 1]
Selective seed abortion is a survival strategy adopted by many species that sacrifices some seeds to allow the remaining ones to set. While in evolutionary terms this is a successful approach, it causes huge losses to crop yields. A pollination time gap (PTG) has been suggested to be associated with position-related grain abortion. To test this hypothesis, we developed a novel approach to alter the natural pattern of maize (Zea mays L.) pollination and to examine the impact of PTGs on kernel growth and the underlying physiological basis. When apical and basal kernels were synchronously pollinated, the basal kernels set and matured but the apical kernels were aborted at an early stage. Delaying pollination to the basal ovaries suppressed their development and reduced invertase activity and sugar levels, which allowed the apical kernels to set and grow normally. In situ localization revealed normal cell wall invertase activity in apical and basal kernels under synchronous pollination but reduced activity in the delayed-pollinated kernels independent of their position. Starch, which was abundant in basal kernel areas, was absent in the apical kernel regions under synchronous pollination but apparent with delayed pollination. Our analyses identified PTG-related sink strength and a low level of local assimilates as the main causes of grain abortion.
DOI:10.1139/b82-270URL [本文引用: 1]
DOI:10.1104/pp.19.00641URL [本文引用: 1]
DOI:10.1111/nph.15115PMID:29603241 [本文引用: 1]
A defining feature of angiosperms is double fertilization involving the female gametophyte central cell and formation of a nutrient-storing tissue called endosperm. The route for the evolutionary origin of endosperm from a gymnosperm ancestor, particularly the molecular steps involved, has remained elusive. Recently, the histidine kinase gene Cytokinin-Independent 1 (CKI1), an activator of cytokinin signaling, was described as a key to specification of the endosperm precursor central cell in Arabidopsis. Here, we have investigated the function and expression of a putative ortholog of CKI1 in the gymnosperm Ginkgo biloba. We demonstrate that Ginkgo CKI1 can partially rescue an Arabidopsis cki1 mutant and promote weak activation of the cytokinin signaling pathway in the Arabidopsis embryo sac, but does not confer central cell specification. Ginkgo CKI1 is expressed in both male and female gametophytes of Ginkgo. In the latter, it is expressed in the ventral canal cell, which is sister to the egg cell in the archegonium. As in Arabidopsis, Ginkgo CKI1 is not expressed in the egg cell. The similarities in expression patterns of CKI1 in Ginkgo and Arabidopsis female gametophytes suggest that extant gymnosperms possess an essential component of the molecular machinery required for angiosperm endosperm development, and provide new insights into endosperm origin from a gymnospermous ancestor.© 2018 The Authors. New Phytologist © 2018 New Phytologist Trust.
1
2007
... 百山祖冷杉是松科冷杉属植物, 与大多数松柏纲植物(针叶树)相似, 孢子叶球单性, 雌雄同株(
植物胚败育机理及其离体培养挽救技术之研究进展
1
2008
... 本研究发现异常胚乳中的海藻糖-6-磷酸(T6P)含量显著低于正常的胚乳, 而T6P是海藻糖的代谢前体, 可通过调控流向糖酵解的糖量, 维持代谢平衡进而调控胚胎发育(
1
2009
... 百山祖冷杉是松科冷杉属植物, 与大多数松柏纲植物(针叶树)相似, 孢子叶球单性, 雌雄同株(
1
2014
... 植物发生濒危的原因包括内部和外部两类因素.外部因素包括栖息地被破坏、外来物种引入、过度开发和环境污染等(
裸子植物种子发育过程及基因调控研究进展
1
2016
... 种子发育过程中, 胚乳的发育动态直接影响胚的发育(
中国亚热带地区4种极危冷杉属植物的地理分布特征及其与气候的关系
1
2012
... 百山祖冷杉(Abies beshanzuensis)隶属松科(Pinaceae)冷杉属(Abies), 是我国特有的珍稀濒危野生植物.1963年首次在浙江省庆元县海拔约1 740 m的地区发现百山祖冷杉的自然分布, 1976年正式定名发表, 1987年被国际自然保护联盟物种生存委员会(International Union for Conservation of Nature-Species Survival Commission, IUCN-SSC)列为全球最濒危的12种植物之一, 1999年, 经国务院批准, 百山祖冷杉被列为国家一级重点保护野生植物.百山祖冷杉分布范围狭窄、开花结实周期长且天然更新能力极弱, 自然状态下繁育概率极低.目前, 全世界百山祖冷杉成年野生植株仅存3株, 处于极度濒危状态(
1
2013
... 为明确百山祖冷杉传粉过程, 我们利用体视显微镜对雌球果进行解剖及观察, 结果显示5月15日珠孔张开且其边缘发育形成沟槽(呈漏斗状结构) (
1
2012
... 采集2019年和2020年6-9月的25日球果, 随机选取每个球果中部(1-2层种鳞)的种鳞10片, 取种鳞上的种子, 去除种翅, 利用体视显微镜进行解剖观察, 记录种子及其胚与胚乳的形态结构特征.将种子去除种翅, 置于FAA溶液中固定24小时, 石蜡切片(
中国濒危野生植物的资源现状与保护对策
1
2011
... 植物发生濒危的原因包括内部和外部两类因素.外部因素包括栖息地被破坏、外来物种引入、过度开发和环境污染等(
1
1990
... 为明确百山祖冷杉传粉过程, 我们利用体视显微镜对雌球果进行解剖及观察, 结果显示5月15日珠孔张开且其边缘发育形成沟槽(呈漏斗状结构) (
百山祖冷杉种子的人工育苗试验
1
2010
... 种子是植物有性繁殖的重要器官, 其发育直接决定了濒危植物的自然更新能力.裸子植物的种子主要由胚和胚乳组成, 受精后, 胚珠中的受精卵与雌配子体分别发育成胚与胚乳.前期研究发现, 百山祖冷杉成熟种子存在空粒和涩粒, 且萌发率极低等现象(
极度濒危植物水松大孢子发生、雌配子体发育及胚形成
1
2015
... 植物发生濒危的原因包括内部和外部两类因素.外部因素包括栖息地被破坏、外来物种引入、过度开发和环境污染等(
植物胚乳发育的表观遗传学调控
1
2012
... 种子发育过程中, 胚乳的发育动态直接影响胚的发育(
富士响应外源GA3和“大小年结果信号”花芽孕育的生理分子机制
1
2016
... 百山祖冷杉传粉后, 种子重量快速增长, 正常发育的种子数量却急剧下降.代谢组分析表明, 在此期间胚乳的碳水化合物代谢旺盛, 尤其是多个糖类代谢途径.由于每个球果中含有大量的种子(302-384粒), 因此, 种子之间对糖类等能源物质的竞争可能导致部分胚乳发育异常, 最终形成类型II/III种子.与被子植物相比, 目前有关裸子植物种子败育与营养物质供应关系的研究较少.拟南芥(Arabidopsis thaliana)在生殖生长过程中, 同化物匮乏会导致正在发育的种子甚至整个角果发生败育(
A novel integrated method for large-scale detection, identification, and quantification of widely targeted metabolites: application in the study of rice metabolomics
1
2013
... 选取百山祖冷杉6月25日发育正常的胚乳(6_NE)和7月25日发育正常的胚乳(7_NE)与发育异常的胚乳(7_AE)进行初生代谢物测定, 每组样品3次生物学重复.参考
Trehalose-6-phosphate synthase 1, which catalyses the first step in trehalose synthesis, is essential for Arabidopsis embryo maturation
1
2002
... 本研究发现异常胚乳中的海藻糖-6-磷酸(T6P)含量显著低于正常的胚乳, 而T6P是海藻糖的代谢前体, 可通过调控流向糖酵解的糖量, 维持代谢平衡进而调控胚胎发育(
The role of trehalose 6-phosphate (Tre6P) in plant metabolism and development
2
2021
... 利用KEGG数据库对3类模式中的关键代谢物进行注释和代谢途径分析(
... 本研究发现异常胚乳中的海藻糖-6-磷酸(T6P)含量显著低于正常的胚乳, 而T6P是海藻糖的代谢前体, 可通过调控流向糖酵解的糖量, 维持代谢平衡进而调控胚胎发育(
Alternate bearing in fruit trees: fruit presence induces polar auxin transport in citrus and olive stem and represses IAA release from the bud
1
2021
... 百山祖冷杉传粉后, 种子重量快速增长, 正常发育的种子数量却急剧下降.代谢组分析表明, 在此期间胚乳的碳水化合物代谢旺盛, 尤其是多个糖类代谢途径.由于每个球果中含有大量的种子(302-384粒), 因此, 种子之间对糖类等能源物质的竞争可能导致部分胚乳发育异常, 最终形成类型II/III种子.与被子植物相比, 目前有关裸子植物种子败育与营养物质供应关系的研究较少.拟南芥(Arabidopsis thaliana)在生殖生长过程中, 同化物匮乏会导致正在发育的种子甚至整个角果发生败育(
Effects of pollen supply and quality on seed formation and maturation in Pinus densiflora
1
2012
... 百山祖冷杉是松科冷杉属植物, 与大多数松柏纲植物(针叶树)相似, 孢子叶球单性, 雌雄同株(
KEGG: kyoto encyclopedia of genes and genomes
1
2000
... 利用KEGG数据库对3类模式中的关键代谢物进行注释和代谢途径分析(
Reproductive failure in Arabidopsis thaliana under transient carbohydrate limitation: flowers and very young siliques are jettisoned and the meristem is maintained to allow successful resumption of reproductive growth
1
2016
... 百山祖冷杉传粉后, 种子重量快速增长, 正常发育的种子数量却急剧下降.代谢组分析表明, 在此期间胚乳的碳水化合物代谢旺盛, 尤其是多个糖类代谢途径.由于每个球果中含有大量的种子(302-384粒), 因此, 种子之间对糖类等能源物质的竞争可能导致部分胚乳发育异常, 最终形成类型II/III种子.与被子植物相比, 目前有关裸子植物种子败育与营养物质供应关系的研究较少.拟南芥(Arabidopsis thaliana)在生殖生长过程中, 同化物匮乏会导致正在发育的种子甚至整个角果发生败育(
The evolution of seeds
1
2010
... 种子发育过程中, 胚乳的发育动态直接影响胚的发育(
Trehalose 6-phosphate promotes seed filling by activating auxin biosynthesis
1
2021
... 本研究发现异常胚乳中的海藻糖-6-磷酸(T6P)含量显著低于正常的胚乳, 而T6P是海藻糖的代谢前体, 可通过调控流向糖酵解的糖量, 维持代谢平衡进而调控胚胎发育(
Factors affecting seed and cone development in Pacific silver fir ( Abies amabilis)
2
1998
... 百山祖冷杉是松科冷杉属植物, 与大多数松柏纲植物(针叶树)相似, 孢子叶球单性, 雌雄同株(
... )雌配子体败育发生在2个时期: (1) 游离核中期到后期阶段; (2) 传粉结束1周后, 未授粉的雌配子体发生败育.其中, 第2个时期(6月末至7月)是温哥华冷杉雌配子体败育的主要时期.温哥华冷杉第2个时期雌配子体败育种子的解剖结构与百山祖冷杉类型III部分种子非常相似, 即种子内残留皱缩的组织, 珠被细胞增殖, 种皮变厚(
Pollination in conifers
1
1998
... 百山祖冷杉是松科冷杉属植物, 与大多数松柏纲植物(针叶树)相似, 孢子叶球单性, 雌雄同株(
Reproductive biology of Abies cephalonica Loudon in Mount Aenos National Park, Cephalonia, Greece
4
2011
... 为明确百山祖冷杉传粉过程, 我们利用体视显微镜对雌球果进行解剖及观察, 结果显示5月15日珠孔张开且其边缘发育形成沟槽(呈漏斗状结构) (
... 百山祖冷杉是松科冷杉属植物, 与大多数松柏纲植物(针叶树)相似, 孢子叶球单性, 雌雄同株(
... ), 传粉过程极大地受到气候的制约, 因此, 普遍存在传粉效率低的现象(
... 百山祖冷杉传粉后, 种子重量快速增长, 正常发育的种子数量却急剧下降.代谢组分析表明, 在此期间胚乳的碳水化合物代谢旺盛, 尤其是多个糖类代谢途径.由于每个球果中含有大量的种子(302-384粒), 因此, 种子之间对糖类等能源物质的竞争可能导致部分胚乳发育异常, 最终形成类型II/III种子.与被子植物相比, 目前有关裸子植物种子败育与营养物质供应关系的研究较少.拟南芥(Arabidopsis thaliana)在生殖生长过程中, 同化物匮乏会导致正在发育的种子甚至整个角果发生败育(
Delayed pollination and low availability of assimilates are major factors causing maize kernel abortion
1
2018
... 百山祖冷杉传粉后, 种子重量快速增长, 正常发育的种子数量却急剧下降.代谢组分析表明, 在此期间胚乳的碳水化合物代谢旺盛, 尤其是多个糖类代谢途径.由于每个球果中含有大量的种子(302-384粒), 因此, 种子之间对糖类等能源物质的竞争可能导致部分胚乳发育异常, 最终形成类型II/III种子.与被子植物相比, 目前有关裸子植物种子败育与营养物质供应关系的研究较少.拟南芥(Arabidopsis thaliana)在生殖生长过程中, 同化物匮乏会导致正在发育的种子甚至整个角果发生败育(
Sexual reproduction in grand fir ( Abies grandis)
1
1982
... 百山祖冷杉是松科冷杉属植物, 与大多数松柏纲植物(针叶树)相似, 孢子叶球单性, 雌雄同株(
The soybean sugar transporter GmSWEET15 mediates sucrose export from endosperm to early embryo
1
2019
... 百山祖冷杉传粉后, 种子重量快速增长, 正常发育的种子数量却急剧下降.代谢组分析表明, 在此期间胚乳的碳水化合物代谢旺盛, 尤其是多个糖类代谢途径.由于每个球果中含有大量的种子(302-384粒), 因此, 种子之间对糖类等能源物质的竞争可能导致部分胚乳发育异常, 最终形成类型II/III种子.与被子植物相比, 目前有关裸子植物种子败育与营养物质供应关系的研究较少.拟南芥(Arabidopsis thaliana)在生殖生长过程中, 同化物匮乏会导致正在发育的种子甚至整个角果发生败育(
The gymnosperm ortholog of the angiosperm central cell-specification gene CKI1 provides an essential clue to endosperm origin
1
2018
... 种子发育过程中, 胚乳的发育动态直接影响胚的发育(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}