Abstract Calcineurin B-like proteins (CBLs) and their CBL-interacting protein kinases (CIPKs) are important regulatory network in response to abiotic stresses. The CBL-CIPK system senses and decodes Ca2+-signals through phosphorylation to regulate plant response to abiotic stresses. In this review, the basic structures of CBLs and CIPKs, and their phosphorylation on different substrates, as well as regulatory mechanisms of plants in response to abiotic stresses were summarized. We also put forward a perspective on the future research directions of CBLs and CIPKs, as well as their potential applications in genetic improvement of crops for stress tolerance. Keywords:abiotic stresses;calcium signaling;CBL-CIPK;ion channels;phosphorylation
PDF (3726KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 谢玲玲, 王金龙, 伍国强. 植物CBL-CIPK信号系统响应非生物胁迫的调控机制. 植物学报, 2021, 56(5): 614-626 doi:10.11983/CBB21024 Xie Lingling, Wang Jinlong, Wu Guoqiang. Regulatory Mechanisms of the Plant CBL-CIPK Signaling System in Response to Abiotic Stress. Chinese Bulletin of Botany, 2021, 56(5): 614-626 doi:10.11983/CBB21024
蓝色箭头表示离子和化合物的流入/流出; 黑色箭头表示激活; 虚线表示Ca2+信号。RBOHF: 呼吸爆发氧化酶同源因子 Figure 2Responses of CBL-CIPK system to nitrate transport, cellular ammonia toxicity, ROS signaling, and ABA signaling in Arabidopsis thaliana
Blue arrows indicate influx/efflux of ion and compound; black arrows indicate activation; dashed lines indicate Ca2+ signaling. RBOHF: Respiratory burst oxidase homolog factor
目前, CBL-CIPK复合物对植物激素调控的研究主要集中在ABA信号通路。研究发现, CBL9-CIPK3在ABA信号通路中起负向调控作用(Pandey et al., 2008)。拟南芥CBL9-CIPK3复合物通过将转录因子ABI1磷酸化, 形成CBL9-CIPK3-ABR1途径(表2), 从而介导种子萌发和ABA依赖的生理生化反应过程(Sanyal et al., 2017)。在小麦(Triticum aestivum)中, TaCBL1- TaCIPK23介导ABA信号途径, 并在干旱胁迫应答中起重要作用(Cui et al., 2018)。此外, 拟南芥CBL2和CBL3与CIPK9和CIPK27也参与ABA信号通路的负向调控(Song et al., 2018)。OsCBL2可正向调控赤霉素的合成(Hwang et al., 2005), 而AtCIPK6参与调控生长素的运输(Tripathi et al., 2009)。上述结果表明, CBL-CIPK复合物广泛参与多种植物激素的调控。
3.5 CBL-CIPK复合物对信号终止的调控
CIPK的C端存在1个PPI结构域(Guo et al., 2001)。在结构分析中, CBL相互作用域和PPI结构域可能有重叠, CBL和PP2C (protein phosphatase 2C)与CIPK的相互作用可能相反(Sanyal et al., 2020)。此结构特征可以提供一种有利的适应机制, 以防止CIPK被CBL激活和被CIPK磷酸酶失活同时进行, 从而为调控下游底物提供一个激酶-磷酸酶的分子开关(Tang et al., 2020)。
AlemánF, Nieves-CordonesM, MartínezV, RubioF (2011). Root K+ acquisition in plants: the Arabidopsis thaliana model Plant Cell Physiol 52, 1603-1612. DOI:10.1093/pcp/pcr096URL [本文引用: 1]
AlmeidaDM, OliveiraMM, SaiboNJM (2017). Regulation of Na+ and K+ homeostasis in plants: towards improved salt stress tolerance in crop plants Genet Mol Biol 40, 326-345. DOI:S1415-47572017000200326PMID:28350038 [本文引用: 3] Soil salinity is a major abiotic stress that results in considerable crop yield losses worldwide. However, some plant genotypes show a high tolerance to soil salinity, as they manage to maintain a high K+/Na+ ratio in the cytosol, in contrast to salt stress susceptible genotypes. Although, different plant genotypes show different salt tolerance mechanisms, they all rely on the regulation and function of K+ and Na+ transporters and H+ pumps, which generate the driving force for K+ and Na+ transport. In this review we will introduce salt stress responses in plants and summarize the current knowledge about the most important ion transporters that facilitate intra- and intercellular K+ and Na+ homeostasis in these organisms. We will describe and discuss the regulation and function of the H+-ATPases, H+-PPases, SOS1, HKTs, and NHXs, including the specific tissues where they work and their response to salt stress.
AslamM, FakherB, JakadaBH, ZhaoLH, CaoSJ, ChengY, QinY (2019). Genome-wide identification and expression profiling of CBL-CIPK gene family in pineapple ( Ananas comosus) and the role of AcCBL1 in abiotic and biotic stress response Biomolecules 9, 293. DOI:10.3390/biom9070293URL [本文引用: 2]
BatističO, WaadtR, SteinhorstL, HeldK, KudlaJ (2010). CBL-mediated targeting of CIPKs facilitates the decoding of calcium signals emanating from distinct cellular stores Plant J 61, 211-222. DOI:10.1111/j.1365-313X.2009.04045.xURL [本文引用: 4]
BeckmannL, EdelKH, BatističO, KudlaJ (2016). A calcium sensor-protein kinase signaling module diversified in plants and is retained in all lineages of Bikonta species Sci Rep 6, 31645. DOI:10.1038/srep31645PMID:27538881 [本文引用: 1] Calcium (Ca2+) signaling is a universal mechanism of signal transduction and involves Ca2+ signal formation and decoding of information by Ca2+ binding proteins. Calcineurin B-like proteins (CBLs), which upon Ca2+ binding activate CBL-interacting protein kinases (CIPKs) regulate a multitude of physiological processes in plants. Here, we combine phylogenomics and functional analyses to investigate the occurrence and structural conservation of CBL and CIPK proteins in 26 species representing all major clades of eukaryotes. We demonstrate the presence of at least singular CBL-CIPK pairs in representatives of Archaeplastida, Chromalveolates and Excavates and their general absence in Opisthokonta and Amoebozoa. This denotes CBL-CIPK complexes as evolutionary ancient Ca2+ signaling modules that likely evolved in the ancestor of all Bikonta. Furthermore, we functionally characterize the CBLs and CIPK from the parabasalid human pathogen Trichomonas vaginalis. Our results reveal strict evolutionary conservation of functionally important structural features, preservation of biochemical properties and a remarkable cross-kingdom protein-protein interaction potential between CBLs and CIPKs from Arabidopsis thaliana and T. vaginalis. Together our findings suggest an ancient evolutionary origin of a functional CBL-CIPK signaling module close to the root of eukaryotic evolution and provide insights into the initial evolution of signaling networks and Ca2+ signaling specificity.
BeheraS, LongY, Schmitz-ThomI, WangXP, ZhangCX, LiH, SteinhorstL, ManishankarP, RenXL, OffenbornJN, WuWH, KudlaJ, WangY (2017). Two spatially and temporally distinct Ca2+ signals convey Arabidopsis thaliana responses to K+ deficiency New Phytol 213, 739-750. DOI:10.1111/nph.2017.213.issue-2URL [本文引用: 1]
CampbellMT, BandilloN, Al ShiblawiFRA, SharmaS, LiuK, DuQ, SchmitzAJ, ZhangC, VéryA, LorenzAJ, WaliaH (2017). Allelic variants of OsHKT1;1 underlie the divergence between indica and japonica subspecies of rice (Oryza sativa) for root sodium content PLoS Genet 13, e1006823. DOI:10.1371/journal.pgen.1006823URL [本文引用: 1]
Chaves-SanjuanA, Sanchez-BarrenaMJ, Gonzalez-RubioJM, MorenoM, RagelP, JimenezM, PardobJM, Martinez-RipollM, QuinteroFJ, AlbertA (2014). Structural basis of the regulatory mechanism of the plant CIPK family of protein kinases controlling ion homeostasis and abiotic stress Proc Natl Acad Sci USA 111, E4532-E4541. DOI:10.1073/pnas.1407610111URL [本文引用: 1]
ChenXF, GuZM, XinDD, HaoL, LiuCJ, HuangJ, MaBJ, ZhangHS (2011). Identification and characterization of putative CIPK genes in maize J Genet Genomics 38, 77-87. DOI:10.1016/j.jcg.2011.01.005URL [本文引用: 1]
ChuLC, OffenbornJN, SteinhorstL, WuXN, XiL, LiZ, JacquotA, LejayL, KudlaJ, SchulzeWX (2021). Plasma membrane calcineurin B-like calcium-ion sensor proteins function in regulating primary root growth and nitrate uptake by affecting global phosphorylation patterns and microdomain protein distribution New Phytol 229, 2223-2237. DOI:10.1111/nph.v229.4URL [本文引用: 2]
CuiXY, DuYT, FuJD, YuTF, WangTC, ChenM, ChenJ, MaYZ, XuZS (2018). Wheat CBL-interacting protein kinase 23 positively regulates drought stress and ABA responses BMC Plant Biol 18, 93-106. DOI:10.1186/s12870-018-1306-5URL [本文引用: 2]
DongQY, BaiBW, AlmutairiBO, KudlaJ (2021). Emerging roles of the CBL-CIPK calcium signaling network as key regulatory hub in plant nutrition J Plant Physiol 257, 153335. DOI:10.1016/j.jplph.2020.153335URL [本文引用: 3]
GuoY, HalfterU, IshitaniM, ZhuJK (2001). Molecular characterization of functional domains in the protein kinase SOS2 that is required for plant salt tolerance Plant Cell 13, 1383-1400. PMID:11402167 [本文引用: 3] The SOS3 (for SALT OVERLY SENSITIVE3) calcium binding protein and SOS2 protein kinase are required for sodium and potassium ion homeostasis and salt tolerance in Arabidopsis. We have shown previously that SOS3 interacts with and activates the SOS2 protein kinase. We report here the identification of a SOS3 binding motif in SOS2 that also serves as the kinase autoinhibitory domain. Yeast two-hybrid assays as well as in vitro binding assays revealed a 21-amino acid motif in the regulatory domain of SOS2 that is necessary and sufficient for interaction with SOS3. Database searches revealed a large family of SOS2-like protein kinases containing such a SOS3 binding motif. Using a yeast two-hybrid system, we show that these SOS2-like kinases interact with members of the SOS3 family of calcium binding proteins. Two-hybrid assays also revealed interaction between the N-terminal kinase domain and the C-terminal regulatory domain within SOS2, suggesting that the regulatory domain may inhibit kinase activity by blocking substrate access to the catalytic site. Removal of the regulatory domain of SOS2, including the SOS3 binding motif, resulted in constitutive activation of the protein kinase, indicating that the SOS3 binding motif can serve as a kinase autoinhibitory domain. Constitutively active SOS2 that is SOS3 independent also was produced by changing Thr(168) to Asp in the activation loop of the SOS2 kinase domain. Combining the Thr(168)-to-Asp mutation with the autoinhibitory domain deletion created a superactive SOS2 kinase. These results provide insights into regulation of the kinase activities of SOS2 and the SOS2 family of protein kinases.
GuoY, XiongLM, SongCP, GongDM, HalfterU, ZhuJK (2002). A calcium sensor and its interacting protein kinase are global regulators of abscisic acid signaling in Arabidopsis Dev Cell 3, 233-244. DOI:10.1016/S1534-5807(02)00229-0URL [本文引用: 1]
HanJP, KösterP, DrerupMM, ScholzM, LiSZ, EdelKH, HashimotoK, KuchitsuK, HipplerM, KudlaJ (2019). Fine-tuning of RBOHF activity is achieved by differential phosphorylation and Ca2+ binding New Phytol 221, 1935-1949. DOI:10.1111/nph.2019.221.issue-4URL [本文引用: 1]
HashimotoK, EckertC, AnschützU, ScholzM, HeldK, WaadtR, ReyerA, HipplerM, BeckerD, KudlaJ (2012). Phosphorylation of calcineurin B-like (CBL) calcium sensor proteins by their CBL-interacting protein kinases (CIPKs) is required for full activity of CBL-CIPK complexes toward their target proteins J Biol Chem 287, 7956-7968. DOI:10.1074/jbc.M111.279331PMID:22253446 [本文引用: 3] Calcineurin B-like proteins (CBLs) represent a family of calcium sensor proteins that interact with a group of serine/threonine kinases designated as CBL-interacting protein kinases (CIPKs). CBL-CIPK complexes are crucially involved in relaying plant responses to many environmental signals and in regulating ion fluxes. However, the biochemical characterization of CBL-CIPK complexes has so far been hampered by low activities of recombinant CIPKs. Here, we report on an efficient wheat germ extract-based in vitro transcription/translation protocol that yields active full-length wild-type CIPK proteins. We identified a conserved serine residue within the C terminus of CBLs as being phosphorylated by their interacting CIPKs. Remarkably, our studies revealed that CIPK-dependent CBL phosphorylation is strictly dependent on CBL-CIPK interaction via the CIPK NAF domain. The phosphorylation status of CBLs does not appear to influence the stability, localization, or CIPK interaction of these calcium sensor proteins in general. However, proper phosphorylation of CBL1 is absolutely required for the in vivo activation of the AKT1 K(+) channel by CBL1-CIPK23 and CBL9-CIPK23 complexes in oocytes. Moreover, we show that by combining CBL1, CIPK23, and AKT1, we can faithfully reconstitute CBL-dependent enhancement of phosphorylation of target proteins by CIPKs in vitro. In addition, we report that phosphorylation of CBL1 by CIPK23 is also required for the CBL1-dependent enhancement of CIPK23 activity toward its substrate. Together, these data identify a novel general regulatory mechanism of CBL-CIPK complexes in that CBL phosphorylation at their flexible C terminus likely provokes conformational changes that enhance specificity and activity of CBL-CIPK complexes toward their target proteins.
HeplerPK (2005). Calcium: a central regulator of plant growth and development Plant Cell 17, 2142-2155. PMID:16061961 [本文引用: 1]
HuHC, WangYY, TsayYF (2009). AtCIPK8, a CBL-interacting protein kinase, regulates the low-affinity phase of the primary nitrate response Plant J 57, 264-278. DOI:10.1111/tpj.2009.57.issue-2URL [本文引用: 1]
HuW, XiaZQ, YanY, DingZH, TieWW, WangLZ, ZouML, WeiYX, LuC, HouXW, WangXQ, PengM (2015). Genome-wide gene phylogeny of CIPK family in cassava and expression analysis of partial drought-induced genes Front Plant Sci 6, 914. DOI:10.3389/fpls.2015.00914PMID:26579161 [本文引用: 3] Cassava is an important food and potential biofuel crop that is tolerant to multiple abiotic stressors. The mechanisms underlying these tolerances are currently less known. CBL-interacting protein kinases (CIPKs) have been shown to play crucial roles in plant developmental processes, hormone signaling transduction, and in the response to abiotic stress. However, no data is currently available about the CPK family in cassava. In this study, a total of 25 C/PK genes were identified from cassava genome based on our previous genome sequencing data. Phylogenetic analysis suggested that 25 MeCIPKs could be classified into four subfamilies, which was supported by exon-intron organizations and the architectures of conserved protein motifs. Transcriptomic analysis of a wild subspecies and two cultivated varieties showed that most MeCIPKs had different expression patterns between wild subspecies and cultivatars in different tissues or in response to drought stress. Some orthologous genes involved in CIPK interaction networks were identified between Arabidopsis and cassava. The interaction networks and co-expression patterns of these orthologous genes revealed that the crucial pathways controlled by CIPK networks may be involved in the differential response to drought stress in different accessions of cassava. Nine MeC1PK genes were selected to investigate their transcriptional response to various stimuli and the results showed the comprehensive response of the tested MeC1PK genes to osmotic, salt, cold, oxidative stressors, and ABA signaling. The identification and expression analysis of CIPK family suggested that C1PK genes are important components of development and multiple signal transduction pathways in cassava. The findings of this study will help lay a foundation for the functional characterization of the C1PK gene family and provide an improved understanding of abiotic stress responses and signaling transduction in cassava.
HwangYS, BethkePC, CheongYH, ChangHS, ZhuT, JonesRL (2005). A gibberellin-regulated calcineurin B in rice localizes to the tonoplast and is implicated in vacuole function Plant Physiol 138, 1347-1358. DOI:10.1104/pp.105.062703URL [本文引用: 1]
KimBG, WaadtR, CheongYH, PandeyJK, Dominguez- SolisJR, SchültkeS, LeeSC, KudlaJ, LuanS (2007). The calcium sensor CBL10 mediates salt tolerance by regulating ion homeostasis in Arabidopsis Plant J 52, 473-484. DOI:10.1111/j.1365-313X.2007.03249.xURL [本文引用: 1]
KimuraS, KawarazakiT, NiboriH, MichikawaM, ImaiA, KayayzH, KuchitsuK (2013). The CBL-interacting protein kinase CIPK26 is a novel interactor of Arabidopsis NADPH oxidase AtRbohF that negatively modulates its ROS-producing activity in a heterologous expression system J Biochem 153, 191-195. [本文引用: 1]
KleistTJ, SpencleyAL, LuanS (2014). Comparative phylogenomics of the CBL-CIPK calcium-decoding network in the moss Physcomitrella, Arabidopsis, and other green lineages Front Plant Sci 5, 187. [本文引用: 3]
KolukisaogluU, WeinlS, BlazevicD, BatisticO, KudlaJ (2004). Calcium sensors and their interacting protein kinases: genomics of the Arabidopsis and rice CBL-CIPK signaling networks Plant Physiol 134, 43-58. PMID:14730064 [本文引用: 6] Calcium signals mediate a multitude of plant responses to external stimuli and regulate a wide range of physiological processes. Calcium-binding proteins, like calcineurin B-like (CBL) proteins, represent important relays in plant calcium signaling. These proteins form a complex network with their target kinases being the CBL-interacting protein kinases (CIPKs). Here, we present a comparative genomics analysis of the full complement of CBLs and CIPKs in Arabidopsis and rice (Oryza sativa). We confirm the expression and transcript composition of the 10 CBLs and 25 CIPKs encoded in the Arabidopsis genome. Our identification of 10 CBLs and 30 CIPKs from rice indicates a similar complexity of this signaling network in both species. An analysis of the genomic evolution suggests that the extant number of gene family members largely results from segmental duplications. A phylogenetic comparison of protein sequences and intron positions indicates an early diversification of separate branches within both gene families. These branches may represent proteins with different functions. Protein interaction analyses and expression studies of closely related family members suggest that even recently duplicated representatives may fulfill different functions. This work provides a basis for a defined further functional dissection of this important plant-specific signaling system.
KösterP, WallradL, EdelKH, FaisalM, AlatarAA, KudlaJ (2019). The battle of two ions: Ca2+ signaling against Na+ stress Plant Biol 21, 39-48. DOI:10.1111/plb.2019.21.issue-S1URL [本文引用: 2]
LaraA, RódenasR, AndrésZ, MartínezV, QuinteroFJ, Nieves-CordonesM, BotellaMA, RubioF (2020). Arabidopsis K+ transporter HAK5-mediated high-affinity root K+ uptake is regulated by protein kinases CIPK1 and CIPK9 J Exp Bot 71, 5053-5060. DOI:10.1093/jxb/eraa212URL [本文引用: 3]
LéranS, EdelKH, PerventM, HashimotoK, Corratgé- FaillieC, OffenbornJN, TillardP, GojonA, KudlaJ, LacombeB (2015). Nitrate sensing and uptake in Arabidopsis are enhanced by ABI2, a phosphatase inactivated by the stress hormone abscisic acid Sci Signal 8, ra43. [本文引用: 3]
LiJ, JiangMM, LiR, YangL, LiuY, ChenHY (2016). Identification and characterization of CBL and CIPK gene families in eggplant ( Solanum melongena L.) Mol Genet Genomics 291, 1769-1781. DOI:10.1007/s00438-016-1218-8URL [本文引用: 1]
LinHX, DuWM, YangYQ, SchumakerKS, GuoY (2014). A calcium-independent activation of the Arabidopsis SOS2-like protein kinase 24 by its interacting SOS3-like calcium binding protein Plant Physiol 164, 2197-2206. DOI:10.1104/pp.113.232272URL [本文引用: 1]
LiuH, WangYX, LiH, TengRM, WangY, ZhuangJ (2019). Genome-wide identification and expression analysis of calcineurin B-like protein and calcineurin B-like protein-interacting protein kinase family genes in tea plant DNA Cell Biol 38, 824-839. DOI:10.1089/dna.2019.4697URL [本文引用: 1]
LiuLT, ZhengCH, KuangBJ, WeiLQ, YanLF, WangT (2016). Receptor-like kinase RUPO interacts with potassium transporters to regulate pollen tube growth and integrity in rice PLoS Genet 12, e1006085. DOI:10.1371/journal.pgen.1006085URL [本文引用: 1]
LiuP, DuanYH, LiuC, XueQH, GuoJ, QiT, KangZS, GuoJ (2018). Corrigendum to: the calcium sensor TaCBL4 and its interacting protein TaCIPK5 are required for wheat resistance to stripe rust fungus J Exp Bot 69, 5309. DOI:10.1093/jxb/ery307URL [本文引用: 1]
López-ArredondoDL, Leyva-GonzálezMA, Alatorre- CobosF, Herrera-EstrellaL (2013). Biotechnology of nutrient uptake and assimilation in plants Int J Dev Biol 57, 595-610. DOI:10.1387/ijdb.130268lhPMID:24166442 [本文引用: 1] Plants require a complex balance of mineral nutrients to reproduce successfully. Because the availability of many of these nutrients in the soil is compromised by several factors, such as soil pH, cation presence, and microbial activity, crop plants depend directly on nutrients applied as fertilizers to achieve high yields. However, the excessive use of fertilizers is a major environmental concern due to nutrient leaching that causes water eutrophication and promotes toxic algae blooms. This situation generates the urgent need for crop plants with increased nutrient use efficiency and better-designed fertilization schemes. The plant biology revolution triggered by the development of efficient gene transfer systems for plant cells together with the more recent development of next-generation DNA and RNA sequencing and other omics platforms have advanced considerably our understanding on the molecular basis of plant nutrition and how plants respond to nutritional stress. To date, genes encoding sensors, transcription factors, transporters, and metabolic enzymes have been identified as potential candidates to improve nutrient use efficiency. In addition, the study of other genetic resources, such as bacteria and fungi, allows the identification of alternative mechanisms of nutrient assimilation, which are potentially applicable in plants. Although significant progress in this respect has been achieved by conventional breeding, in this review we focus on the biotechnological approaches reported to date aimed at boosting the use of the three most limiting nutrients in the majority of arable lands: nitrogen, phosphorus, and iron.
López-ArredondoDL, Leyva-GonzálezMA, González- MoralesSI, Lopez-BucioJ, Herrera-EstrellaL (2014). Phosphate nutrition: improving low-phosphate tolerance in crops Annu Rev Plant Biol 65, 95-123. DOI:10.1146/annurev-arplant-050213-035949PMID:24579991 [本文引用: 1] Phosphorus is an essential nutrient that is required for all major developmental processes and reproduction in plants. It is also a major constituent of the fertilizers required to sustain high-yield agriculture. Levels of phosphate--the only form of phosphorus that can be assimilated by plants--are suboptimal in most natural and agricultural ecosystems, and when phosphate is applied as fertilizer in soils, it is rapidly immobilized owing to fixation and microbial activity. Thus, cultivated plants use only approximately 20-30% of the applied phosphate, and the rest is lost, eventually causing water eutrophication. Recent advances in the understanding of mechanisms by which wild and cultivated species adapt to low-phosphate stress and the implementation of alternative bacterial pathways for phosphorus metabolism have started to allow the design of more effective breeding and genetic engineering strategies to produce highly phosphate-efficient crops, optimize fertilizer use, and reach agricultural sustainability with a lower environmental cost. In this review, we outline the current advances in research on the complex network of plant responses to low-phosphorus stress and discuss some strategies used to manipulate genes involved in phosphate uptake, remobilization, and metabolism to develop low-phosphate-tolerant crops, which could help in designing more efficient crops.
LuTT, ZhangGF, SunLR, WangJ, HaoFS (2017). Genome-wide identification of CBL family and expression analysis of CBLs in response to potassium deficiency in cotton Peer J 5, e3653. DOI:10.7717/peerj.3653URL [本文引用: 1]
LyzengaWJ, SullivanV, LiuHX, StoneSL (2017). The kinase activity of calcineurin B-like interacting protein kinase 26 (CIPK26) influences its own stability and that of the ABA-regulated ubiquitin ligase, keep on going (KEG) Front Plant Sci 8, 502. DOI:10.3389/fpls.2017.00502PMID:28443108 [本文引用: 3] The Really Interesting New Gene (RING)-type E3 ligase, Keep on Going (KEG) plays a critical role in Arabidopsis growth after germination and the connections between KEG and hormone signaling pathways are expanding. With regards to abscisic acid (ABA) signaling, KEG targets ABA-responsive transcription factors abscisic acid insensitive 5, ABF1 and ABF3 for ubiquitination and subsequent degradation through the 26S proteasome. Regulation of E3 ligases through self-ubiquitination is common to RING-type E3 ligases and ABA promotes KEG self-ubiquitination and degradation. ABA-mediated degradation of KEG is phosphorylation-dependent; however, upstream signaling proteins that may regulate KEG stability have not been characterized. In this report, we show that CBL-Interacting Protein Kinase (CIPK) 26 can phosphorylate KEG in vitro. Using both in vitro and in planta degradation assays we provide evidence which suggests that the kinase activity of CIPK26 promotes the degradation of KEG. Furthermore, we found that the kinase activity of CIPK26 also influences its own stability; a constitutively active version is more stable than a wild type or a kinase dead version. Our results suggest a reciprocal regulation model wherein an activated and stable CIPK26 phosphorylates KEG to promote degradation of the E3.
MaQ, TangRJ, ZhengXJ, WangSM, LuanS (2015). The calcium sensor CBL7 modulates plant responses to low nitrate in Arabidopsis Biochem Biophys Res Commun 468, 59-65. DOI:10.1016/j.bbrc.2015.10.164URL [本文引用: 1]
MaX, GaiWX, QiaoYM, AliM, WeiAM, LuoDX, LiQH, GongZH (2019). Identification of CBL and CIPK gene families and functional characterization of CaCIPK1 under Phytophthora capsici in pepper (Capsicum annuum L.) BMC Genomics 20, 775. DOI:10.1186/s12864-019-6125-zURL [本文引用: 1]
MaX, LiQH, YuYN, QiaoYM, HaqS, GongZH (2020). The CBL-CIPK pathway in plant response to stress signals Int J Mol Sci 21, 5668. DOI:10.3390/ijms21165668URL [本文引用: 1]
MaYC, ChengQK, ChengZM, LiH, ChangYH, LinJ (2017). Identification of important physiological traits and moderators that are associated with improved salt tolerance in CBL and CIPK overexpressors through a meta- analysis Front Plant Sci 8, 856. DOI:10.3389/fpls.2017.00856URL [本文引用: 1]
MahajanS, SoporySK, TutejaN (2006). Cloning and characterization of CBL-CIPK signaling components from a legume ( Pisum sativum) FEBS J 273, 907-925. PMID:16478466 [本文引用: 1] The studies on calcium sensor calcineurin B-like protein (CBL) and CBL interacting protein kinases (CIPK) are limited to Arabidopsis and rice and their functional role is only beginning to emerge. Here, we present cloning and characterization of a protein kinase (PsCIPK) from a legume, pea, with novel properties. The PsCIPK gene is intronless and encodes a protein that showed partial homology to the members of CIPK family. The recombinant PsCIPK protein was autophosphorylated at Thr residue(s). Immunoprecipitation and yeast two-hybrid analysis showed direct interaction of PsCIPK with PsCBL, whose cDNA and genomic DNA were also cloned in this study. PsCBL showed homology to AtCBL3 and contained calcium-binding activity. We demonstrate for the first time that PsCBL is phosphorylated at its Thr residue(s) by PsCIPK. Immunofluorescence/confocal microscopy showed that PsCBL is exclusively localized in the cytosol, whereas PsCIPK is localized in the cytosol and the outer membrane. The exposure of plants to NaCl, cold and wounding co-ordinately upregulated the expression of PsCBL and PsCIPK genes. The transcript levels of both genes were also coordinately stimulated in response to calcium and salicylic acid. However, drought and abscisic acid had no effect on the expression of these genes. These studies show the ubiquitous presence of CBL/CIPK in higher plants and enhance our understanding of their role in abiotic and biotic stress signalling.
MaierhoferT, DiekmannM, OffenbornJN, LindC, BauerH, HashimotoK, Al-RasheidKAS, LuanS, KudlaJ, GeigerD, HedrichR (2014a). Site- and kinase-specific phosphorylation-mediated activation of SLAC1, a guard cell anion channel stimulated by abscisic acid Sci Signal 7, ra86. [本文引用: 2]
ManikSMN, ShiSJ, MaoJJ, DongLH, SuYL, WangQ, LiuHB (2015). The calcium sensor CBL-CIPK is involved in plant’s response to abiotic stresses Int J Genomics2015, 493191. [本文引用: 1]
ManishankarP, WangN, KösterP, AlatarAA, KudlaJ (2018). Calcium signaling during salt stress and in the regulation of ion homeostasis J Exp Bot 69, 4215-4226. DOI:10.1093/jxb/ery201URL [本文引用: 1]
MaoJJ, ManikS, ShiSJ, ChaoJT, JinYR, WangQ, LiuHB (2016). Mechanisms and physiological roles of the CBL-CIPK networking system in Arabidopsis thaliana Genes 7, 62-77. DOI:10.3390/genes7090062URL [本文引用: 1]
MoCY, WanSM, XiaYQ, RenN, ZhouY, JiangXY (2018). Expression patterns and identified protein-protein interactions suggest that cassava CBL-CIPK signal networks function in responses to abiotic stresses Front Plant Sci 9, 269. DOI:10.3389/fpls.2018.00269URL [本文引用: 2]
NiuLL, DongBY, SongZH, MengD, FuYJ (2018). Genome-wide identification and characterization of CIPK family and analysis responses to various stresses in apple (Malus domestica) Int J Mol Sci 19, 2131. DOI:10.3390/ijms19072131URL [本文引用: 1]
OdaY, KobayashiNI, TanoiK, MaJF, ItouY, KatsuharaM, ItouT, HorieT (2018). T-DNA tagging-based gain-of- function of OsHKT1;4 reinforces Na exclusion from leaves and stems but triggers Na toxicity in roots of rice under salt stress Int J Mol Sci 19, 235. DOI:10.3390/ijms19010235URL [本文引用: 1]
PandeyGK, GrantJJ, CheongYH, KimBG, LiLG, LuanS (2008). Calcineurin-B-like protein CBL9 interacts with target kinase CIPK3 in the regulation of ABA response in seed germination Mol Plant 1, 238-248. DOI:10.1093/mp/ssn003URL [本文引用: 1]
ParkerJL, NewsteadS (2014). Molecular basis of nitrate uptake by the plant nitrate transporter NRT1.1 Nature 507, 68-72. DOI:10.1038/nature13116URL [本文引用: 1]
PeckS, MittlerR (2020). Plant signaling in biotic and abiotic stress J Exp Bot 71, 1649-1651. DOI:10.1093/jxb/eraa051URL [本文引用: 1]
PiaoHL, XuanYH, ParkSH, JeBI, ParkSJ, ParkSH, KimCM, HuangJ, WangGK, KimMJ, KangSM, LeeIJ, KwonTR, KimYH, YeoUS, YiG, SonD, HanCD (2010). OsCIPK31, a CBL-interacting protein kinase is involved in germination and seedling growth under abiotic stress conditions in rice plants Mol Cells 30, 19-27. DOI:10.1007/s10059-010-0084-1URL [本文引用: 1]
PlasenciaFA, EstradaY, FloresFB, Ortíz-AtienzaA, LozanoR, EgeaI (2021). The Ca2+ sensor calcineurin B-like protein 10 in plants: emerging new crucial roles for plant abiotic stress tolerance Front Plant Sci 11, 599944. DOI:10.3389/fpls.2020.599944URL [本文引用: 2]
RagelP, RaddatzN, LeidiEO, QuinteroFJ, PardoJM (2019). Regulation of K+ nutrition in plants Front Plant Sci 10, 281. DOI:10.3389/fpls.2019.00281URL [本文引用: 3]
SaitoS, HamamotoS, MoriyaK, MatsuuraA, SatoY, MutoJ, NoguchiH, YamauchiS, TozawaY, UedaM, HashimotoK, KösterP, DongQY, HeldK, KudlaJ, UtsumiT, UozumiN (2018). N-myristoylation and S-acylation are common modifications of Ca2+-regulated Arabidopsis kinases and are required for activation of the SLAC1 anion channel New Phytol 218, 1504-1521. DOI:10.1111/nph.2018.218.issue-4URL [本文引用: 3]
SaitoS, UozumiN (2019). Guard cell membrane anion transport systems and their regulatory components: an elaborate mechanism controlling stress-induced stomatal closure Plants 8, 9. DOI:10.3390/plants8010009URL [本文引用: 2]
SaitoS, UozumiN (2020). Calcium-regulated phosphorylation systems controlling uptake and balance of plant nutrients Front Plant Sci 11, 44. DOI:10.3389/fpls.2020.00044URL [本文引用: 4]
Sánchez-BarrenaMJ, Martínez-RipollM, AlbertA (2013). Structural biology of a major signaling network that regulates plant abiotic stress: the CBL-CIPK mediated pathway Int J Mol Sci 14, 5734-5749. DOI:10.3390/ijms14035734PMID:23481636 [本文引用: 6] The Arabidopsis SOS2 family of twenty-six protein kinases (CIPKs), their interacting activators, the SOS3 family of ten calcium-binding proteins (CBLs) and protein phosphatases type 2C (PP2C), function together in decoding calcium signals elicited by different environmental stimuli. Biochemical data suggest that stable CBL-CIPK or CIPK-PP2C complexes may be regulating the activity of various substrates controlling ion homeostasis. The available structural information provides a general regulatory mechanism in which calcium perception by CBLs and kinase activation is coupled. The structural basis of this molecular mechanism and the specificity of the network is reviewed and discussed in detail.
Sánchez-BarrenaMJ, Martínez-RipollM, ZhuJK, AlbertA (2005). The structure of the Arabidopsis thaliana SOS3: molecular mechanism of sensing calcium for salt stress response J Mol Biol 345, 1253-1264. PMID:15644219 [本文引用: 3] The Arabidopsis thaliana SOS3 gene encodes a calcium sensor that is required for plant salt tolerance. The SOS3 protein binds to and activates the self-inhibited SOS2 protein kinase, which mediates the expression and activities of various transporters important for ion homeostasis under salt stress. SOS3 belongs to a unique family of calcium-binding proteins that contain two pairs of EF hand motifs with four putative metal-binding sites. We report the crystal structure of a dimeric SOS3 protein in complex with calcium, and with calcium and manganese. Analytical ultracentrifugation experiments and circular dichroism measurements show that calcium binding is responsible for the dimerization of SOS3. This leads to a change in the global shape and surface properties of the protein that may be sufficient to transmit the Ca(2+) signal elicited during salt stress.
SanyalSK, KanwarP, YadavAK, SharmaC, KumarA, PandeyGK (2017). Arabidopsis CBL interacting protein kinase 3 interacts with ABR1, an APETALA2 domain transcription factor, to regulate ABA responses Plant Sci 254, 48-59. DOI:10.1016/j.plantsci.2016.11.004URL [本文引用: 3]
SongCP, AgarwalM, OhtaM, GuoY, HalfterU, WangPC, ZhuJK (2005). Role of an Arabidopsis AP2/EREBP- type transcriptional repressor in abscisic acid and drought stress responses Plant Cell 17, 2384-2396. DOI:10.1105/tpc.105.033043URL [本文引用: 2]
SongSJ, FengQN, LiCL, LiE, LiuQ, KangH, ZhangW, ZhangY, LiS (2018). A tonoplast-associated calcium- signaling module dampens ABA signaling during stomatal movement Plant Physiol 177, 1666-1678. DOI:10.1104/pp.18.00377URL [本文引用: 2]
SteinhorstL, KudlaJ (2013). Calcium and reactive oxygen species rule the waves of signaling Plant Physiol 163, 471-485. DOI:10.1104/pp.113.222950PMID:23898042 [本文引用: 1]
StraubT, LudewigU, NeuhäuserB (2017). The kinase CIPK23 inhibits ammonium transport in Arabidopsis thaliana Plant Cell 29, 409-422. DOI:10.1105/tpc.16.00806URL [本文引用: 3]
SuWH, RenYJ, WangDJ, HuangL, FuXQ, LingH, SuYC, HuangN, TangHC, XuLP, QueYX (2020). New insights into the evolution and functional divergence of the CIPK gene family in Saccharum BMC Genomics 21, 868-888. DOI:10.1186/s12864-020-07264-9URL [本文引用: 1]
SunT, WangY, WangM, LiTT, ZhouY, WangXT, WeiSY, HeGY, YangGX (2015). Identification and comprehensive analyses of the CBL and CIPK gene families in wheat (Triticum aestivum L.) BMC Plant Biol 15, 269. DOI:10.1186/s12870-015-0657-4URL [本文引用: 2]
TangRJ, LiuH, YangY, YangL, GaoXS, GarciaVJ, LuanS, ZhangHX (2012). Tonoplast calcium sensors CBL2 and CBL3 control plant growth and ion homeostasis through regulating V-ATPase activity in Arabidopsis Cell Res 22, 1650-1665. DOI:10.1038/cr.2012.161URL [本文引用: 4]
TangRJ, WangC, LiKL, LuanS (2020). The CBL-CIPK calcium signaling network: unified paradigm from 20 years of discoveries Trends Plant Sci 25, 604-617. DOI:10.1016/j.tplants.2020.01.009URL [本文引用: 8]
TripathiV, SyedN, LaxmiA, ChattopadhyayD (2009). Role of CIPK6 in root growth and auxin transport Plant Signal Behav 4, 663-665. DOI:10.1111/j.1365-313-X.2009.03812.xPMID:19820350 [本文引用: 1] In our recent publication, we have shown that a T-DNA insertion in Arabidopsis CIPK6 gene encoding a CBL-interacting protein kinase caused reduction in expression of the gene and emergence of lateral roots. The change in phenotype in the mutant line was likely due to reduction in shoot-to-root acropetal and the root tip basipetal auxin transport. Here we report identification of a homozygous knockout line of AtCIPK6 (atcipk6) with no detectable expression of the gene in normal growth condition. The knockout line exhibited considerable decrease in growth rate of the taproot as well as in emergence of lateral roots. The mutant line also showed reduction in the root tip basipetal and shoot-to-root acropetal auxin transport. Relative rate of auxin transport and the root phenotype of the atcipk6 closely matched with those of pgp4-1, an Arabidopsis line mutated in PGP4. This gene encodes an ABC integral membrane transporter, which functions in polar auxin transport. These observations strengthen our earlier proposal that CIPK6 is probably involved in polar auxin transport and indicate that it may function through the PGP4 auxin transporter.
WangXP, ZhuBP, JiangZH, WangSC (2019). Calcium-mediation of jasmonate biosynthesis and signaling in plants Plant Sci 287, 110192. DOI:10.1016/j.plantsci.2019.110192URL [本文引用: 1]
WangY, LiTT, JohnSJ, ChenMJ, ChangJL, YangGX, HeGY (2018). A CBL-interacting protein kinase TaCIPK27 confers drought tolerance and exogenous ABA sensitivity in transgenic Arabidopsis Plant Physiol Biochem 123, 103-113. DOI:10.1016/j.plaphy.2017.11.019URL [本文引用: 2]
WeinlS, KudlaJ (2009). The CBL-CIPK Ca2+-decoding signaling network: function and perspectives New Phytol 184, 517-528. DOI:10.1111/nph.2009.184.issue-3URL [本文引用: 3]
WengLY, ZhangMX, WangK, ChenGL, DingM, YuanW, ZhuYY, XuWF, XuFY (2020). Potassium alleviates ammonium toxicity in rice by reducing its uptake through activation of plasma membrane H+-ATPase to enhance proton extrusion Plant Physiol Biochem 151, 429-437. DOI:10.1016/j.plaphy.2020.03.040URL [本文引用: 1]
XiY, LiuJY, DongC, ChengZM (2017). The CBL and CIPK gene family in grapevine (Vitis vinifera): genome- wide analysis and expression profiles in response to various abiotic stresses Front Plant Sci 8, 978. DOI:10.3389/fpls.2017.00978URL [本文引用: 2]
YangY, ZhangC, TangRJ, XuHX, LanWZ, ZhaoFG, LuanF (2019). Calcineurin B-like proteins CBL4 and CBL10 mediate two independent salt tolerance pathways in Arabidopsis Int J Mol Sci 20, 2421. DOI:10.3390/ijms20102421URL [本文引用: 1]
YinX, WangQL, ChenQ, XiangN, YangYQ, YangYP (2017). Genome-wide identification and functional analysis of the calcineurin B-like protein and calcineurin B-like protein-interacting protein kinase gene families in turnip ( Brassica rapa var. rapa) Front Plant Sci 8, 1191. DOI:10.3389/fpls.2017.01191URL [本文引用: 2]
YinXC, XiaYQ, XieQ, CaoYX, WangZY, HaoGP, SongJ, ZhouY, JiangXY (2020). The protein kinase complex CBL10-CIPK8-SOS1 functions in Arabidopsis to regulate salt tolerance J Exp Bot 71, 1801-1814. DOI:10.1093/jxb/erz549URL [本文引用: 6]
YuYH, XiaXL, YinWL, ZhangHC (2007). Comparative genomic analysis of CIPK gene family in Arabidopsis and Populus Plant Growth Regul 52, 101-110. DOI:10.1007/s10725-007-9165-3URL [本文引用: 1]
ZhangHF, YangB, LiuWZ, LiHW, WangL, WangYY, DengM, LiangWW, DeyholosMK, JiangYQ (2014a). Identification and characterization of CBL and CIPK gene families in canola (Brassica napus L.) BMC Plant Biol 14, 8. DOI:10.1186/1471-2229-14-8URL [本文引用: 1]
ZhangHW, FengH, ZhangJW, GeRC, ZhangLY, WangYX, LiLG, WeiJH, LiRF (2020). Emerging crosstalk between two signaling pathways coordinates K+ and Na+ homeostasis in the halophyte Hordeum brevisubulatum J Exp Bot 71, 4345-4358. DOI:10.1093/jxb/eraa191URL [本文引用: 1]
ZhangXX, KösterP, SchlückingK, BalcerowiczD, HashimotoK, KuchitsuK, VissenbergK, KudlaJ (2018b). CBL1-CIPK26-mediated phosphorylation enhances activity of the NADPH oxidase RBOHC, but is dispensable for root hair growth FEBS Lett 592, 2582-2593. DOI:10.1002/feb2.2018.592.issue-15URL [本文引用: 1]
ZhangY, LvY, JahanN, ChenG, RenDY, GuoLB (2018a). Sensing of abiotic stress and ionic stress responses in plants Int J Mol Sci 19, 3298. DOI:10.3390/ijms19113298URL [本文引用: 1]
ZhaoJF, YuAL, DuYW, WangGH, LiYF, ZhaoGY, WangXD, ZhangWZ, ChengK, LiuX, WangZH, WangYW (2019). Foxtail millet (Setaria italica (L.) P. Beauv) CIPKs are responsive to ABA and abiotic stresses PLoS One 14, e0225091. DOI:10.1371/journal.pone.0225091URL [本文引用: 1]
ZhengXJ, HeK, KleistT, ChenF, LuanS (2015). Anion channel SLAH3 functions in nitrate-dependent alleviation of ammonium toxicity in Arabidopsis Plant Cell Environ 38, 474-486. DOI:10.1111/pce.2015.38.issue-3URL [本文引用: 2]
ZhouXN, HaoHM, ZhangYG, BaiYL, ZhuWB, QinYX, YuanFF, ZhaoFY, WangMY, HuJJ, XuH, GuoAG, ZhaoHX, ZhaoY, CaoCL, YangYQ, SchumakerKS, GuoY, XieCG (2015). SOS2-like protein kinase 5, an SNF1-related protein kinase 3-type protein kinase, is important for abscisic acid responses in Arabidopsis through phosphorylation of abscisic acid-insensitive 5 Plant Physiol 168, 659-676. DOI:10.1104/pp.114.255455URL [本文引用: 3]
ZhouY, LaiZS, YinXC, YuS, XuYY, WangXX, CongXL, LuoYH, XuHX, JiangXY (2016). Hyperactive mutant of a wheat plasma membrane Na+/H+ antiporter improves the growth and salt tolerance of transgenic tobacco Plant Sci 253, 176-186. DOI:10.1016/j.plantsci.2016.09.016URL [本文引用: 1]
ZhuJK, LiuJP, XiongLM (1998). Genetic analysis of salt tolerance in Arabidopsis: evidence for a critical role of potassium nutrition Plant Cell 10, 1181-1191. PMID:9668136 [本文引用: 4] A large genetic screen for sos (for salt overly sensitive) mutants was performed in an attempt to isolate mutations in any gene with an sos phenotype. Our search yielded 28 new alleles of sos1, nine mutant alleles of a newly identified locus, SOS2, and one allele of a third salt tolerance locus, SOS3. The sos2 mutations, which are recessive, were mapped to the lower arm of chromosome V, approximately 2.3 centimorgans away from the marker PHYC. Growth measurements demonstrated that sos2 mutants are specifically hypersensitive to inhibition by Na+ or Li+ and not hypersensitive to general osmotic stresses. Interestingly, the SOS2 locus is also necessary for K+ nutrition because sos2 mutants were unable to grow on a culture medium with a low level of K+. The expression of several salt-inducible genes was superinduced in sos2 plants. The salt tolerance of sos1, sos2, and sos3 mutants correlated with their K+ tissue content but not their Na+ tissue content. Double mutant analysis indicated that the SOS genes function in the same pathway. Based on these results, a genetic model for salt tolerance mechanisms in Arabidopsis is presented in which SOS1, SOS2, and SOS3 are postulated to encode regulatory components controlling plant K+ nutrition that in turn is essential for salt tolerance. 植物CBL-CIPK信号系统的功能及其作用机理 1 2014
... 在细胞内, Ca2+被认为是无处不在、无时不有的功能性调节者, 对许多生理生化反应和发育过程具有重要调控作用(Yin et al., 2017; Köster et al., 2019).Ca2+信号通过Ca2+感应蛋白解码并传递, 进而引起细胞内各种信号系统对环境变化的响应(张和臣等, 2007; Kudla et al., 2018).在植物中, CBLs (Calcineurin B-like proteins)也称SCaBPs (SOS3-like calcium binding proteins), 是一类独特的Ca2+感应蛋白, 其通过激活CIPKs (CBL-interacting protein kinases)/PKS (protein kinase S)解码Ca2+信号(Sánchez-Barrena et al., 2005; 沈金秋等, 2014; Tang et al., 2020).CBLs和CIPKs互作构成一个复杂的信号网络, 在植物对干旱、低温和盐碱等逆境的应答调控中起核心作用(张和臣等, 2007; Sun et al., 2015; Ma et al., 2017; Xi et al., 2017).CBL家族一个成员可以与CIPK家族一个或多个成员相互作用, 建立不同信号级联, 从而形成精准的CBL-CIPK调控网络(Ma et al., 2020).自CBL-CIPK信号系统被发现以来, 学术界对不同途径的CBL-CIPK复合物的功能、结构、基因表达和调控进行了广泛研究(Weinl and Kudla, 2009; Aslam et al., 2019).本文综述了CBLs和CIPKs的结构、分类及其响应各种非生物胁迫的调控机制, 并展望了未来的研究方向, 以期为作物抗逆性遗传改良提供思路. ...
植物K+通道AKT1的研究进展 1 2017
... K+是植物细胞中最丰富的离子, 在调节细胞渗透压、膜电位和细胞内pH值方面发挥至关重要的作用(伍国强等, 2017; Almeida et al., 2017; Ragel et al., 2019).K+亏缺会导致拟南芥根中Ca2+迅速增加(Behera et al., 2017).维持根系K+吸收的主要系统有K+通道AKT1和高亲和性K+转运蛋白HAK5 (图1; 表2) (Alemán et al., 2011).CBL1/9与CIPK23结合并在质膜上聚集, CIPK23激活后将AKT1和HAK5磷酸化并激活, 以促进K+的吸收和转运(图1; 表2) (Lara et al., 2020).此外, CBL4-CIPK6复合物作用于AKT2, 可应答K+的外排(图1) (Saito and Uozumi, 2020).液泡膜CBL2/3-CIPK3/9/23/26复合物激活TPKs (two- pore K+ channels), 以维持细胞内K+稳态(Tang et al., 2020; Dong et al., 2021). ...
非生物逆境胁迫下植物钙信号转导的分子机制 2 2007
... 在细胞内, Ca2+被认为是无处不在、无时不有的功能性调节者, 对许多生理生化反应和发育过程具有重要调控作用(Yin et al., 2017; Köster et al., 2019).Ca2+信号通过Ca2+感应蛋白解码并传递, 进而引起细胞内各种信号系统对环境变化的响应(张和臣等, 2007; Kudla et al., 2018).在植物中, CBLs (Calcineurin B-like proteins)也称SCaBPs (SOS3-like calcium binding proteins), 是一类独特的Ca2+感应蛋白, 其通过激活CIPKs (CBL-interacting protein kinases)/PKS (protein kinase S)解码Ca2+信号(Sánchez-Barrena et al., 2005; 沈金秋等, 2014; Tang et al., 2020).CBLs和CIPKs互作构成一个复杂的信号网络, 在植物对干旱、低温和盐碱等逆境的应答调控中起核心作用(张和臣等, 2007; Sun et al., 2015; Ma et al., 2017; Xi et al., 2017).CBL家族一个成员可以与CIPK家族一个或多个成员相互作用, 建立不同信号级联, 从而形成精准的CBL-CIPK调控网络(Ma et al., 2020).自CBL-CIPK信号系统被发现以来, 学术界对不同途径的CBL-CIPK复合物的功能、结构、基因表达和调控进行了广泛研究(Weinl and Kudla, 2009; Aslam et al., 2019).本文综述了CBLs和CIPKs的结构、分类及其响应各种非生物胁迫的调控机制, 并展望了未来的研究方向, 以期为作物抗逆性遗传改良提供思路. ...
... ).CBLs和CIPKs互作构成一个复杂的信号网络, 在植物对干旱、低温和盐碱等逆境的应答调控中起核心作用(张和臣等, 2007; Sun et al., 2015; Ma et al., 2017; Xi et al., 2017).CBL家族一个成员可以与CIPK家族一个或多个成员相互作用, 建立不同信号级联, 从而形成精准的CBL-CIPK调控网络(Ma et al., 2020).自CBL-CIPK信号系统被发现以来, 学术界对不同途径的CBL-CIPK复合物的功能、结构、基因表达和调控进行了广泛研究(Weinl and Kudla, 2009; Aslam et al., 2019).本文综述了CBLs和CIPKs的结构、分类及其响应各种非生物胁迫的调控机制, 并展望了未来的研究方向, 以期为作物抗逆性遗传改良提供思路. ...
Root K+ acquisition in plants: the Arabidopsis thaliana model 1 2011
... K+是植物细胞中最丰富的离子, 在调节细胞渗透压、膜电位和细胞内pH值方面发挥至关重要的作用(伍国强等, 2017; Almeida et al., 2017; Ragel et al., 2019).K+亏缺会导致拟南芥根中Ca2+迅速增加(Behera et al., 2017).维持根系K+吸收的主要系统有K+通道AKT1和高亲和性K+转运蛋白HAK5 (图1; 表2) (Alemán et al., 2011).CBL1/9与CIPK23结合并在质膜上聚集, CIPK23激活后将AKT1和HAK5磷酸化并激活, 以促进K+的吸收和转运(图1; 表2) (Lara et al., 2020).此外, CBL4-CIPK6复合物作用于AKT2, 可应答K+的外排(图1) (Saito and Uozumi, 2020).液泡膜CBL2/3-CIPK3/9/23/26复合物激活TPKs (two- pore K+ channels), 以维持细胞内K+稳态(Tang et al., 2020; Dong et al., 2021). ...
Regulation of Na+ and K+ homeostasis in plants: towards improved salt stress tolerance in crop plants 3 2017
... 大量研究表明, CBL与CIPK互作并将其磷酸化是一种普遍的机制, 而且磷酸化能够增强CBL-CIPK复合物的相互作用, 进而提高CIPK的活性(Hashimoto et al., 2012; Sanyal et al., 2020).在植物中, CIPK对CBL的磷酸化最初在豌豆(Pisum sativum)中被报道, PsCIPK可对PsCBL的Thr残基进行磷酸化(Mahajan et al., 2006), 随后在多个物种中相继发现了这种现象(Sanyal et al., 2016).CBL的C端含有1个由23个氨基酸残基组成的基序, 其中有绝对保守的氨基酸残基P、M、L、F、P和F, 因此称作“PFPF基序”(FPSF结构域) (Sanyal et al., 2020).CIPK能够将FPSF结构域中保守的Ser残基磷酸化(Sanyal et al., 2020).海藻(Sargassum sp.)的部分CBL缺少该结构, 以Asp和Glu残基代替(Hashimoto et al., 2012).CIPK介导CBL磷酸化是部分CBL-CIPK信号系统发挥具体功能的必要步骤.例如, AtCIPK24对AtCBL10的磷酸化可激活Na+转运蛋白NHX7 (Almeida et al., 2017), 而CIPK23对CBL1的磷酸化也可激活K+通道AKT1 (Hashimoto et al., 2012; Sánchez-Barrena et al., 2020).因此, CBL的磷酸化一方面可增强其与CIPK互作; 另一方面又促进CIPK对下游靶标蛋白的转磷酸作用(Lin et al., 2014; Sanyal et al., 2016). ...
... 低浓度Na+对植物生长有一定的刺激作用, 但高浓度Na+则会引起细胞渗透和离子毒害(Zhu, 2016).拟南芥SOS信号途径是调节植物根耐盐性的重要机制(Zhu et al., 1998; Yin et al., 2020).在盐胁迫下, AtCBL10与AtCIPK24 (SOS2)互作(图1; 表2), 将Na+区域化到液泡, 以保护植物地上部免受盐胁迫伤害(Hu et al., 2015; Plasencia et al., 2021).这与SOS通路的耐盐机制不同(Yang et al., 2019).被激活的SOS1利用H+-ATPase (AHA2)产生的质子梯度能量将Na+转运出细胞(图1) (Yin et al., 2020).CBL10与CIPK8形成的复合物介导SOS1的激活, 表明CIPK8在调节拟南芥地上部响应盐胁迫中起重要作用(Yin et al., 2020), 但其复合物下游靶标蛋白还有待深入鉴定.细胞内较高的K+/Na+比是植物耐盐性的核心指标(Zhang et al., 2020).研究表明, HKT1 (high affinity potassium transporters 1)能够维持细胞内高浓度的K+和低浓度Na+ (Almeida et al., 2017).水稻(Oryza sativa) OsHKT1;1主要在地上部维管组织中表达, 高盐条件下可增强细胞的Na+外排能力(Campbell et al., 2017).此外, 过表达OsHKT1;4的转基因植株木质部中Na+含量降低, 而根中的Na+含量增加, 从而提高对盐胁迫的敏感性(Oda et al., 2018).CBL2/CBL3-CIPK21也参与植物的耐盐性, 该复合物主要在液泡中发挥作用, 但其下游靶点仍有待确定(Sanyal et al., 2016). ...
... K+是植物细胞中最丰富的离子, 在调节细胞渗透压、膜电位和细胞内pH值方面发挥至关重要的作用(伍国强等, 2017; Almeida et al., 2017; Ragel et al., 2019).K+亏缺会导致拟南芥根中Ca2+迅速增加(Behera et al., 2017).维持根系K+吸收的主要系统有K+通道AKT1和高亲和性K+转运蛋白HAK5 (图1; 表2) (Alemán et al., 2011).CBL1/9与CIPK23结合并在质膜上聚集, CIPK23激活后将AKT1和HAK5磷酸化并激活, 以促进K+的吸收和转运(图1; 表2) (Lara et al., 2020).此外, CBL4-CIPK6复合物作用于AKT2, 可应答K+的外排(图1) (Saito and Uozumi, 2020).液泡膜CBL2/3-CIPK3/9/23/26复合物激活TPKs (two- pore K+ channels), 以维持细胞内K+稳态(Tang et al., 2020; Dong et al., 2021). ...
Genome-wide identification and expression profiling of CBL-CIPK gene family in pineapple ( Ananas comosus) and the role of AcCBL1 in abiotic and biotic stress response 2 2019
... 在细胞内, Ca2+被认为是无处不在、无时不有的功能性调节者, 对许多生理生化反应和发育过程具有重要调控作用(Yin et al., 2017; Köster et al., 2019).Ca2+信号通过Ca2+感应蛋白解码并传递, 进而引起细胞内各种信号系统对环境变化的响应(张和臣等, 2007; Kudla et al., 2018).在植物中, CBLs (Calcineurin B-like proteins)也称SCaBPs (SOS3-like calcium binding proteins), 是一类独特的Ca2+感应蛋白, 其通过激活CIPKs (CBL-interacting protein kinases)/PKS (protein kinase S)解码Ca2+信号(Sánchez-Barrena et al., 2005; 沈金秋等, 2014; Tang et al., 2020).CBLs和CIPKs互作构成一个复杂的信号网络, 在植物对干旱、低温和盐碱等逆境的应答调控中起核心作用(张和臣等, 2007; Sun et al., 2015; Ma et al., 2017; Xi et al., 2017).CBL家族一个成员可以与CIPK家族一个或多个成员相互作用, 建立不同信号级联, 从而形成精准的CBL-CIPK调控网络(Ma et al., 2020).自CBL-CIPK信号系统被发现以来, 学术界对不同途径的CBL-CIPK复合物的功能、结构、基因表达和调控进行了广泛研究(Weinl and Kudla, 2009; Aslam et al., 2019).本文综述了CBLs和CIPKs的结构、分类及其响应各种非生物胁迫的调控机制, 并展望了未来的研究方向, 以期为作物抗逆性遗传改良提供思路. ...
... The CBL and CIPK families in different species Table 1
物种
CBLs数量
CIPKs数量
参考文献
拟南芥(Arabidopsis thaliana)
10
26
Kolukisaoglu et al., 2004
油菜(Brassica napus)
7
23
Zhang et al., 2014a
杨树(Populus trichocarpa)
10
25
Weinl and Kudla, 2009
辣椒(Capsicum annuum)
9
26
Ma et al., 2019
水稻(Oryza sativa)
10
31
Weinl and Kudla, 2009; Piao et al., 2010
茶(Camellia sinensis)
7
18
Liu et al., 2019
小麦(Triticum aestivum)
24
79
Sun et al., 2015; Liu et al., 2018
芜菁(B. rapa var. rapa)
19
51
Yin et al., 2017
木薯(Manihot esculenta)
8
26
Hu et al., 2015; Mo et al., 2018
茄子(Solanum melongena)
5
15
Li et al., 2016
菠萝(Ananas comosus)
8
21
Aslam et al., 2019
葡萄(Vitis vinifera)
8
20
Xi et al., 2017
玉米(Zea mays)
8
43
Chen et al., 2011
甜菜(Beta vulgaris)
7
20
未发表数据
2 植物CBL-CIPK系统与磷酸化 CBL-CIPK复合物对下游靶标蛋白的调节主要通过磷酸化方式进行(Sanyal et al., 2020).靶标蛋白主要有通道蛋白、转运蛋白和转录因子等(Sanyal et al., 2015).已有研究表明, 可被CBL-CIPK复合物磷酸化的阳离子(H+、Na+、K+、Mg2+、Fe2+和NH4+)相关蛋白如AHA2、SOS1、AMT1、ATL31、AKT1、HAK5、TPK1、ARF、ERF、SnRK2D和FIT, 而一些阴离子(OH-和NO3-)转运蛋白如CHL1 (NRT1.1)、RBOH-F和RBOH-C也可被CBL-CIPK磷酸化(Ragel et al.,2019; Lara et al., 2020; Sanyal et al., 2020; Dong et al., 2021). ...
CBL-mediated targeting of CIPKs facilitates the decoding of calcium signals emanating from distinct cellular stores 4 2010
... 系统发育分析表明, 拟南芥AtCBLs可分为3类.AtCBL10是第1类中唯一的成员, 并且在CBL家族中具有最长的N端(Sánchez-Barrena et al., 2013).共聚焦荧光显微分析表明, AtCBL10定位于质膜和液泡膜(Kim et al., 2007; Batistič et al., 2010).第2类有5个成员(AtCBL1、-4、-5、-8和-9), 它们具有N端很短的结构特点(Sánchez-Barrena et al., 2013); 其中大多数包含用于脂双层修饰的保守MGCXXS/T基序, 可帮助CBLs固定在膜上(Tang et al., 2015; Saito et al., 2018).第3类成员具有相对较长的N端(Sánchez- Barrena et al., 2013), 它们均具有液泡膜靶向序列(tonoplast targeting sequence, TTS) MSQCXDGXK HXCXSXXXCF (除AtCBL7外) (Tang et al., 2012; Kleist et al., 2014).TTS则有助于AtCBL2、-3和-6定位于液泡膜(Batistič et al., 2010; Tang et al., 2012).CBL7和CBL3是串联复制, 但它们的N端结构域明显不同(Kleist et al., 2014; Sanyal et al., 2016).CBL7作为Ca2+感应蛋白, 能够解码细胞核和细胞质的Ca2+信号(Batistič et al., 2010). ...
... ).TTS则有助于AtCBL2、-3和-6定位于液泡膜(Batistič et al., 2010; Tang et al., 2012).CBL7和CBL3是串联复制, 但它们的N端结构域明显不同(Kleist et al., 2014; Sanyal et al., 2016).CBL7作为Ca2+感应蛋白, 能够解码细胞核和细胞质的Ca2+信号(Batistič et al., 2010). ...
... 信号(Batistič et al., 2010). ...
... 根据内含子丰富度, 可将拟南芥AtCIPK家族分为富有内含子和稀有内含子2个进化分支(Mao et al., 2016).在稀有内含子的进化分支中有17个成员(AtCIPK2、-4、-5、-6、-7、-10、-11、-12、-13、-14、-15、-16、-18、-19、-20、-22和-25), 其它9个成员(CIPK1、-3、-8、-9、-17、-21、-23、-24和-26)在富有内含子的进化分支中.分段和串联重复的发现促进了这2个进化分支的扩展(Guo et al., 2001).CIPK在细胞中通常定位于细胞质和细胞核, 且可通过与CBLs互作靶向下游蛋白(Batistič et al., 2010).此外, 系统发育分析结果表明, AtCIPKs家族分为5个亚组: A、B、C、D和E.有意思的是, 含有多个内含子的AtCIPK成员都聚集在A亚组中, 而其它4个亚组中的基因均为少有或无内含子的成员(Kolukisaoglu et al., 2004). ...
A calcium sensor-protein kinase signaling module diversified in plants and is retained in all lineages of Bikonta species 1 2016
... 植物CBLs是与酵母(Saccharomyces cerevisiae)和动物中的CNB (calcineurin B subunit)最相似的一个Ca2+传感器蛋白家族(Cui et al., 2018).CBL感知应激信号中的Ca2+信号, 使植物适应环境变化(Bender et al., 2018; Tang et al., 2020).CBL含有1个典型的Ca2+结合基序, 即4个手性延伸因子(elongation factor hands, EF-hands), 每个EF-hands包含1个能与Ca2+结合的保守α-螺旋-环-α-螺旋结构(Lu et al., 2017).EF-hands排列在固定的空间中, 从EF-hand1到EF-hand4依次有22、25和32个氨基酸的距离(Sánchez-Barrena et al., 2005).环区的特征是具有12个残基DKDGDGKIDFEE的保守序列(Kolukisaoglu et al., 2004).位置1(X)、3(Y)、5(Z)、7(-X)、9(-Y)和12(-Z)中的氨基酸高度保守, 并负责与Ca2+配位(Sanyal et al., 2015).值得注意的是, EF-hand1在X位置和Y位置之间包含2个插入氨基酸残基的位置(Sánchez-Barrena et al., 2013).这个位置氨基酸的变化导致与Ca2+结合的亲和力发生变化(Kolukisaoglu et al., 2004).除了具有Ca2+感应的EF-hands, CBL还具有N端定位基序和C端磷酸化基序(Beckmann et al., 2016).研究表明, 拟南芥(Arabidopsis thaliana) CBL家族4个成员(AtCBL1、-4、-5和-9)具有N-豆蔻酰化基序(Wang et al., 2019).在SOS (salt overly sensitive)信号系统中, CBL4/SOS3的N-豆蔻酰化基序是植物耐盐性的必备条件(Kleist et al., 2014; Saito et al., 2018), 这为Ca2+信号的传递提供了结构基础. ...
Two spatially and temporally distinct Ca2+ signals convey Arabidopsis thaliana responses to K+ deficiency 1 2017
... K+是植物细胞中最丰富的离子, 在调节细胞渗透压、膜电位和细胞内pH值方面发挥至关重要的作用(伍国强等, 2017; Almeida et al., 2017; Ragel et al., 2019).K+亏缺会导致拟南芥根中Ca2+迅速增加(Behera et al., 2017).维持根系K+吸收的主要系统有K+通道AKT1和高亲和性K+转运蛋白HAK5 (图1; 表2) (Alemán et al., 2011).CBL1/9与CIPK23结合并在质膜上聚集, CIPK23激活后将AKT1和HAK5磷酸化并激活, 以促进K+的吸收和转运(图1; 表2) (Lara et al., 2020).此外, CBL4-CIPK6复合物作用于AKT2, 可应答K+的外排(图1) (Saito and Uozumi, 2020).液泡膜CBL2/3-CIPK3/9/23/26复合物激活TPKs (two- pore K+ channels), 以维持细胞内K+稳态(Tang et al., 2020; Dong et al., 2021). ...
Revisiting paradigms of Ca2+ signaling protein kinase regulation in plants 1 2018
... 植物CBLs是与酵母(Saccharomyces cerevisiae)和动物中的CNB (calcineurin B subunit)最相似的一个Ca2+传感器蛋白家族(Cui et al., 2018).CBL感知应激信号中的Ca2+信号, 使植物适应环境变化(Bender et al., 2018; Tang et al., 2020).CBL含有1个典型的Ca2+结合基序, 即4个手性延伸因子(elongation factor hands, EF-hands), 每个EF-hands包含1个能与Ca2+结合的保守α-螺旋-环-α-螺旋结构(Lu et al., 2017).EF-hands排列在固定的空间中, 从EF-hand1到EF-hand4依次有22、25和32个氨基酸的距离(Sánchez-Barrena et al., 2005).环区的特征是具有12个残基DKDGDGKIDFEE的保守序列(Kolukisaoglu et al., 2004).位置1(X)、3(Y)、5(Z)、7(-X)、9(-Y)和12(-Z)中的氨基酸高度保守, 并负责与Ca2+配位(Sanyal et al., 2015).值得注意的是, EF-hand1在X位置和Y位置之间包含2个插入氨基酸残基的位置(Sánchez-Barrena et al., 2013).这个位置氨基酸的变化导致与Ca2+结合的亲和力发生变化(Kolukisaoglu et al., 2004).除了具有Ca2+感应的EF-hands, CBL还具有N端定位基序和C端磷酸化基序(Beckmann et al., 2016).研究表明, 拟南芥(Arabidopsis thaliana) CBL家族4个成员(AtCBL1、-4、-5和-9)具有N-豆蔻酰化基序(Wang et al., 2019).在SOS (salt overly sensitive)信号系统中, CBL4/SOS3的N-豆蔻酰化基序是植物耐盐性的必备条件(Kleist et al., 2014; Saito et al., 2018), 这为Ca2+信号的传递提供了结构基础. ...
Allelic variants of OsHKT1;1 underlie the divergence between indica and japonica subspecies of rice (Oryza sativa) for root sodium content 1 2017
... 低浓度Na+对植物生长有一定的刺激作用, 但高浓度Na+则会引起细胞渗透和离子毒害(Zhu, 2016).拟南芥SOS信号途径是调节植物根耐盐性的重要机制(Zhu et al., 1998; Yin et al., 2020).在盐胁迫下, AtCBL10与AtCIPK24 (SOS2)互作(图1; 表2), 将Na+区域化到液泡, 以保护植物地上部免受盐胁迫伤害(Hu et al., 2015; Plasencia et al., 2021).这与SOS通路的耐盐机制不同(Yang et al., 2019).被激活的SOS1利用H+-ATPase (AHA2)产生的质子梯度能量将Na+转运出细胞(图1) (Yin et al., 2020).CBL10与CIPK8形成的复合物介导SOS1的激活, 表明CIPK8在调节拟南芥地上部响应盐胁迫中起重要作用(Yin et al., 2020), 但其复合物下游靶标蛋白还有待深入鉴定.细胞内较高的K+/Na+比是植物耐盐性的核心指标(Zhang et al., 2020).研究表明, HKT1 (high affinity potassium transporters 1)能够维持细胞内高浓度的K+和低浓度Na+ (Almeida et al., 2017).水稻(Oryza sativa) OsHKT1;1主要在地上部维管组织中表达, 高盐条件下可增强细胞的Na+外排能力(Campbell et al., 2017).此外, 过表达OsHKT1;4的转基因植株木质部中Na+含量降低, 而根中的Na+含量增加, 从而提高对盐胁迫的敏感性(Oda et al., 2018).CBL2/CBL3-CIPK21也参与植物的耐盐性, 该复合物主要在液泡中发挥作用, 但其下游靶点仍有待确定(Sanyal et al., 2016). ...
Structural basis of the regulatory mechanism of the plant CIPK family of protein kinases controlling ion homeostasis and abiotic stress 1 2014
... 植物CIPK在功能上类似于酵母SNF1 (sucrose non- fermenting 1)和哺乳动物AMPK (AMP-dependent kinase) (Mo et al., 2018; Niu et al., 2018).CIPK由2个结构域组成, 一个含有磷酸化位点激活环的保守N端激酶催化结构域, 另一个具有NAF/FISL基序和PPI (protein phosphatase interaction)基序高度不同的C端调节域(Sánchez-Barrena et al., 2013; Sanyal et al., 2015).NAF基序以其高度保守的氨基酸Asn (N)、Ala (A)、Phe (F)、Ile (I)、Ser (S)和Leu (L)而得名, 是结合CBL的必需位点, 其对于维持AtCIPK24和AtCBL4互作是必需的, 并且能够连接AtCIPK24的C端调节结构域以覆盖其激活环, 从而使激酶保持在自动抑制状态(Chaves-Sanjuan et al., 2014).CIPK是具有激酶和调节结构域的Ser/Thr蛋白激酶(Tang et al., 2020).显然, 磷酸化是该系统发挥相应功能的主要作用方式.因此, 磷酸化是CIPK调节靶标蛋白以及与CBL互作的优选方式(Sanyal et al., 2020). ...
Identification and characterization of putative CIPK genes in maize 1 2011
... The CBL and CIPK families in different species Table 1
物种
CBLs数量
CIPKs数量
参考文献
拟南芥(Arabidopsis thaliana)
10
26
Kolukisaoglu et al., 2004
油菜(Brassica napus)
7
23
Zhang et al., 2014a
杨树(Populus trichocarpa)
10
25
Weinl and Kudla, 2009
辣椒(Capsicum annuum)
9
26
Ma et al., 2019
水稻(Oryza sativa)
10
31
Weinl and Kudla, 2009; Piao et al., 2010
茶(Camellia sinensis)
7
18
Liu et al., 2019
小麦(Triticum aestivum)
24
79
Sun et al., 2015; Liu et al., 2018
芜菁(B. rapa var. rapa)
19
51
Yin et al., 2017
木薯(Manihot esculenta)
8
26
Hu et al., 2015; Mo et al., 2018
茄子(Solanum melongena)
5
15
Li et al., 2016
菠萝(Ananas comosus)
8
21
Aslam et al., 2019
葡萄(Vitis vinifera)
8
20
Xi et al., 2017
玉米(Zea mays)
8
43
Chen et al., 2011
甜菜(Beta vulgaris)
7
20
未发表数据
2 植物CBL-CIPK系统与磷酸化 CBL-CIPK复合物对下游靶标蛋白的调节主要通过磷酸化方式进行(Sanyal et al., 2020).靶标蛋白主要有通道蛋白、转运蛋白和转录因子等(Sanyal et al., 2015).已有研究表明, 可被CBL-CIPK复合物磷酸化的阳离子(H+、Na+、K+、Mg2+、Fe2+和NH4+)相关蛋白如AHA2、SOS1、AMT1、ATL31、AKT1、HAK5、TPK1、ARF、ERF、SnRK2D和FIT, 而一些阴离子(OH-和NO3-)转运蛋白如CHL1 (NRT1.1)、RBOH-F和RBOH-C也可被CBL-CIPK磷酸化(Ragel et al.,2019; Lara et al., 2020; Sanyal et al., 2020; Dong et al., 2021). ...
Plasma membrane calcineurin B-like calcium-ion sensor proteins function in regulating primary root growth and nitrate uptake by affecting global phosphorylation patterns and microdomain protein distribution 2 2021
... NO3-和NH4+是植物根系从土壤中吸收的2种N素来源(Tang et al., 2020).Ca2+调控多种NRT/NPF和SLAH转运蛋白进行NO3-摄取和转运(Sanyal et al., 2020).NPF6.3通过二聚体作用使NO3-从低亲和性模式转换为高亲和性模式, 此过程通过CBL1/9- CIPK23对Thr101的磷酸化作用进行控制(Ho et al., 2009; Parker and Newstead, 2014; Sun et al., 2014; Chu et al., 2021).NH4+吸收由AMTs (ammonium transporters)介导(Sanyal et al., 2020).高浓度NH4+会替代K+而产生毒害作用, 因此必须控制其在细胞的非毒性水平(Zheng et al., 2015).AMT的2个成员(AMT1;1和AMT1;2)也可被CBL1-CIPK23复合物抑制(表2; 图2) (Straub et al., 2017).因此, CBL1-CIPK23可能在维持根细胞K+稳态和免受NH4+毒害中起关键作用(Zheng et al., 2015).Weng等(2020)研究表明, 高浓度K+降低水稻根系对NH4+的吸收速率, 增强对H+的外排速率, 从而减轻NH4+对水稻的毒害.此外, CBL1/9-CIPK23与SLAH2和SLAH3互作通过吸收或分泌NO3-参与硝酸盐的转运(表2) (Ho et al., 2009; Léran et al., 2015). ...
Wheat CBL-interacting protein kinase 23 positively regulates drought stress and ABA responses 2 2018
... 植物CBLs是与酵母(Saccharomyces cerevisiae)和动物中的CNB (calcineurin B subunit)最相似的一个Ca2+传感器蛋白家族(Cui et al., 2018).CBL感知应激信号中的Ca2+信号, 使植物适应环境变化(Bender et al., 2018; Tang et al., 2020).CBL含有1个典型的Ca2+结合基序, 即4个手性延伸因子(elongation factor hands, EF-hands), 每个EF-hands包含1个能与Ca2+结合的保守α-螺旋-环-α-螺旋结构(Lu et al., 2017).EF-hands排列在固定的空间中, 从EF-hand1到EF-hand4依次有22、25和32个氨基酸的距离(Sánchez-Barrena et al., 2005).环区的特征是具有12个残基DKDGDGKIDFEE的保守序列(Kolukisaoglu et al., 2004).位置1(X)、3(Y)、5(Z)、7(-X)、9(-Y)和12(-Z)中的氨基酸高度保守, 并负责与Ca2+配位(Sanyal et al., 2015).值得注意的是, EF-hand1在X位置和Y位置之间包含2个插入氨基酸残基的位置(Sánchez-Barrena et al., 2013).这个位置氨基酸的变化导致与Ca2+结合的亲和力发生变化(Kolukisaoglu et al., 2004).除了具有Ca2+感应的EF-hands, CBL还具有N端定位基序和C端磷酸化基序(Beckmann et al., 2016).研究表明, 拟南芥(Arabidopsis thaliana) CBL家族4个成员(AtCBL1、-4、-5和-9)具有N-豆蔻酰化基序(Wang et al., 2019).在SOS (salt overly sensitive)信号系统中, CBL4/SOS3的N-豆蔻酰化基序是植物耐盐性的必备条件(Kleist et al., 2014; Saito et al., 2018), 这为Ca2+信号的传递提供了结构基础. ...
... 目前, CBL-CIPK复合物对植物激素调控的研究主要集中在ABA信号通路.研究发现, CBL9-CIPK3在ABA信号通路中起负向调控作用(Pandey et al., 2008).拟南芥CBL9-CIPK3复合物通过将转录因子ABI1磷酸化, 形成CBL9-CIPK3-ABR1途径(表2), 从而介导种子萌发和ABA依赖的生理生化反应过程(Sanyal et al., 2017).在小麦(Triticum aestivum)中, TaCBL1- TaCIPK23介导ABA信号途径, 并在干旱胁迫应答中起重要作用(Cui et al., 2018).此外, 拟南芥CBL2和CBL3与CIPK9和CIPK27也参与ABA信号通路的负向调控(Song et al., 2018).OsCBL2可正向调控赤霉素的合成(Hwang et al., 2005), 而AtCIPK6参与调控生长素的运输(Tripathi et al., 2009).上述结果表明, CBL-CIPK复合物广泛参与多种植物激素的调控. ...
Emerging roles of the CBL-CIPK calcium signaling network as key regulatory hub in plant nutrition 3 2021
... CBL-CIPK复合物对下游靶标蛋白的调节主要通过磷酸化方式进行(Sanyal et al., 2020).靶标蛋白主要有通道蛋白、转运蛋白和转录因子等(Sanyal et al., 2015).已有研究表明, 可被CBL-CIPK复合物磷酸化的阳离子(H+、Na+、K+、Mg2+、Fe2+和NH4+)相关蛋白如AHA2、SOS1、AMT1、ATL31、AKT1、HAK5、TPK1、ARF、ERF、SnRK2D和FIT, 而一些阴离子(OH-和NO3-)转运蛋白如CHL1 (NRT1.1)、RBOH-F和RBOH-C也可被CBL-CIPK磷酸化(Ragel et al.,2019; Lara et al., 2020; Sanyal et al., 2020; Dong et al., 2021). ...
... K+是植物细胞中最丰富的离子, 在调节细胞渗透压、膜电位和细胞内pH值方面发挥至关重要的作用(伍国强等, 2017; Almeida et al., 2017; Ragel et al., 2019).K+亏缺会导致拟南芥根中Ca2+迅速增加(Behera et al., 2017).维持根系K+吸收的主要系统有K+通道AKT1和高亲和性K+转运蛋白HAK5 (图1; 表2) (Alemán et al., 2011).CBL1/9与CIPK23结合并在质膜上聚集, CIPK23激活后将AKT1和HAK5磷酸化并激活, 以促进K+的吸收和转运(图1; 表2) (Lara et al., 2020).此外, CBL4-CIPK6复合物作用于AKT2, 可应答K+的外排(图1) (Saito and Uozumi, 2020).液泡膜CBL2/3-CIPK3/9/23/26复合物激活TPKs (two- pore K+ channels), 以维持细胞内K+稳态(Tang et al., 2020; Dong et al., 2021). ...
Molecular characterization of functional domains in the protein kinase SOS2 that is required for plant salt tolerance 3 2001
... 根据内含子丰富度, 可将拟南芥AtCIPK家族分为富有内含子和稀有内含子2个进化分支(Mao et al., 2016).在稀有内含子的进化分支中有17个成员(AtCIPK2、-4、-5、-6、-7、-10、-11、-12、-13、-14、-15、-16、-18、-19、-20、-22和-25), 其它9个成员(CIPK1、-3、-8、-9、-17、-21、-23、-24和-26)在富有内含子的进化分支中.分段和串联重复的发现促进了这2个进化分支的扩展(Guo et al., 2001).CIPK在细胞中通常定位于细胞质和细胞核, 且可通过与CBLs互作靶向下游蛋白(Batistič et al., 2010).此外, 系统发育分析结果表明, AtCIPKs家族分为5个亚组: A、B、C、D和E.有意思的是, 含有多个内含子的AtCIPK成员都聚集在A亚组中, 而其它4个亚组中的基因均为少有或无内含子的成员(Kolukisaoglu et al., 2004). ...
... 在这些必需的离子中, Ca2+浓度通常相对较低,在细胞质中保持在0.1 mmol·L-1左右(Hepler, 2005).然而, 当植物遭遇干旱、盐碱、病原体侵害或营养缺乏等逆境时, Ca2+通过质膜离子通道流入细胞中或从细胞器中释放, 使胞质Ca2+浓度迅速升高(Zhu, 2016; Manishankar et al., 2018; Toyota et al., 2018).这导致Ca2+结合的感应蛋白构象发生变化, 随之与下游效应蛋白以Ca2+依赖方式相互作用并调控其功能(Tang et al., 2020).胞内Ca2+与CBL在其EF-hands处结合, 而CIPK的C端NAF/FISH结构域与CBL互作(Guo et al., 2001; Kolukisaoglu et al., 2004; Su et al., 2020).CBL与CIPK结合使得CIPK的C端自抑制结构域释放N端激酶结构域, 从而增强CIPK活性(Zhou et al., 2016; Yin et al., 2020). ...
... CIPK的C端存在1个PPI结构域(Guo et al., 2001).在结构分析中, CBL相互作用域和PPI结构域可能有重叠, CBL和PP2C (protein phosphatase 2C)与CIPK的相互作用可能相反(Sanyal et al., 2020).此结构特征可以提供一种有利的适应机制, 以防止CIPK被CBL激活和被CIPK磷酸酶失活同时进行, 从而为调控下游底物提供一个激酶-磷酸酶的分子开关(Tang et al., 2020). ...
A calcium sensor and its interacting protein kinase are global regulators of abscisic acid signaling in Arabidopsis 1 2002
Fine-tuning of RBOHF activity is achieved by differential phosphorylation and Ca2+ binding 1 2019
... 与ROS有关的氧化酶RBOHF (respiratory burst oxidase homolog factor)受到CBL1/9-CIPK26的调控(表2; 图2) (Zhang et al., 2014b).CIPK26可影响定位在质膜上的RBOHF活性(Kimura et al., 2013).然而, 该结果是在异源表达系统中试验得到的, 因此CIPK26对RBOHF活性的调节作用尚缺乏足够的证据.但可以肯定的是, CBL1/CBL9介导CIPK26到质膜的定位导致RBHOF磷酸化(Zhang et al., 2018b; Han et al., 2019). ...
Phosphorylation of calcineurin B-like (CBL) calcium sensor proteins by their CBL-interacting protein kinases (CIPKs) is required for full activity of CBL-CIPK complexes toward their target proteins 3 2012
... 大量研究表明, CBL与CIPK互作并将其磷酸化是一种普遍的机制, 而且磷酸化能够增强CBL-CIPK复合物的相互作用, 进而提高CIPK的活性(Hashimoto et al., 2012; Sanyal et al., 2020).在植物中, CIPK对CBL的磷酸化最初在豌豆(Pisum sativum)中被报道, PsCIPK可对PsCBL的Thr残基进行磷酸化(Mahajan et al., 2006), 随后在多个物种中相继发现了这种现象(Sanyal et al., 2016).CBL的C端含有1个由23个氨基酸残基组成的基序, 其中有绝对保守的氨基酸残基P、M、L、F、P和F, 因此称作“PFPF基序”(FPSF结构域) (Sanyal et al., 2020).CIPK能够将FPSF结构域中保守的Ser残基磷酸化(Sanyal et al., 2020).海藻(Sargassum sp.)的部分CBL缺少该结构, 以Asp和Glu残基代替(Hashimoto et al., 2012).CIPK介导CBL磷酸化是部分CBL-CIPK信号系统发挥具体功能的必要步骤.例如, AtCIPK24对AtCBL10的磷酸化可激活Na+转运蛋白NHX7 (Almeida et al., 2017), 而CIPK23对CBL1的磷酸化也可激活K+通道AKT1 (Hashimoto et al., 2012; Sánchez-Barrena et al., 2020).因此, CBL的磷酸化一方面可增强其与CIPK互作; 另一方面又促进CIPK对下游靶标蛋白的转磷酸作用(Lin et al., 2014; Sanyal et al., 2016). ...
... sp.)的部分CBL缺少该结构, 以Asp和Glu残基代替(Hashimoto et al., 2012).CIPK介导CBL磷酸化是部分CBL-CIPK信号系统发挥具体功能的必要步骤.例如, AtCIPK24对AtCBL10的磷酸化可激活Na+转运蛋白NHX7 (Almeida et al., 2017), 而CIPK23对CBL1的磷酸化也可激活K+通道AKT1 (Hashimoto et al., 2012; Sánchez-Barrena et al., 2020).因此, CBL的磷酸化一方面可增强其与CIPK互作; 另一方面又促进CIPK对下游靶标蛋白的转磷酸作用(Lin et al., 2014; Sanyal et al., 2016). ...
... 通道AKT1 (Hashimoto et al., 2012; Sánchez-Barrena et al., 2020).因此, CBL的磷酸化一方面可增强其与CIPK互作; 另一方面又促进CIPK对下游靶标蛋白的转磷酸作用(Lin et al., 2014; Sanyal et al., 2016). ...
Calcium: a central regulator of plant growth and development 1 2005
... 在这些必需的离子中, Ca2+浓度通常相对较低,在细胞质中保持在0.1 mmol·L-1左右(Hepler, 2005).然而, 当植物遭遇干旱、盐碱、病原体侵害或营养缺乏等逆境时, Ca2+通过质膜离子通道流入细胞中或从细胞器中释放, 使胞质Ca2+浓度迅速升高(Zhu, 2016; Manishankar et al., 2018; Toyota et al., 2018).这导致Ca2+结合的感应蛋白构象发生变化, 随之与下游效应蛋白以Ca2+依赖方式相互作用并调控其功能(Tang et al., 2020).胞内Ca2+与CBL在其EF-hands处结合, 而CIPK的C端NAF/FISH结构域与CBL互作(Guo et al., 2001; Kolukisaoglu et al., 2004; Su et al., 2020).CBL与CIPK结合使得CIPK的C端自抑制结构域释放N端激酶结构域, 从而增强CIPK活性(Zhou et al., 2016; Yin et al., 2020). ...
CHL1 functions as a nitrate sensor in plants 4 2009
... 在拟南芥中, CBL-CIPK信号系统调控许多离子转运蛋白(图1, 图2), 包括介导K+ (KAT1、KAT2、GORK、AKT1、AKT2、HAK5和SPIK)、Na+ (SOS1)、NH4+ (AMT1;1和AMT1;2)、NO3- (NRT1.1、NRT2.4和NRT2.5)、Cl- (SLAC1、SLAH2和SLAH3)和H+ (AHA2和V-ATPase)的通道或转运蛋白(Ho et al., 2009; Léran et al., 2015; Liu et al., 2016; Straub et al., 2017; Ragel et al., 2019; Saito and Uozumi, 2019).CBL-CIPK在响应C/N养分及Mg和Fe的摄取中也发挥重要作用(Sanyal et al., 2020).这种Ca2+依赖性磷酸化调节系统的功能可确保植物的生长, 并使其能耐受各种环境胁迫(Sanyal et al., 2020).由此可见, CBL-CIPK复合物的磷酸化调控对象主要是各种离子通道和转运蛋白. ...
... NO3-和NH4+是植物根系从土壤中吸收的2种N素来源(Tang et al., 2020).Ca2+调控多种NRT/NPF和SLAH转运蛋白进行NO3-摄取和转运(Sanyal et al., 2020).NPF6.3通过二聚体作用使NO3-从低亲和性模式转换为高亲和性模式, 此过程通过CBL1/9- CIPK23对Thr101的磷酸化作用进行控制(Ho et al., 2009; Parker and Newstead, 2014; Sun et al., 2014; Chu et al., 2021).NH4+吸收由AMTs (ammonium transporters)介导(Sanyal et al., 2020).高浓度NH4+会替代K+而产生毒害作用, 因此必须控制其在细胞的非毒性水平(Zheng et al., 2015).AMT的2个成员(AMT1;1和AMT1;2)也可被CBL1-CIPK23复合物抑制(表2; 图2) (Straub et al., 2017).因此, CBL1-CIPK23可能在维持根细胞K+稳态和免受NH4+毒害中起关键作用(Zheng et al., 2015).Weng等(2020)研究表明, 高浓度K+降低水稻根系对NH4+的吸收速率, 增强对H+的外排速率, 从而减轻NH4+对水稻的毒害.此外, CBL1/9-CIPK23与SLAH2和SLAH3互作通过吸收或分泌NO3-参与硝酸盐的转运(表2) (Ho et al., 2009; Léran et al., 2015). ...
... ) (Ho et al., 2009; Léran et al., 2015). ...
AtCIPK8, a CBL-interacting protein kinase, regulates the low-affinity phase of the primary nitrate response 1 2009
Genome-wide gene phylogeny of CIPK family in cassava and expression analysis of partial drought-induced genes 3 2015
... The CBL and CIPK families in different species Table 1
物种
CBLs数量
CIPKs数量
参考文献
拟南芥(Arabidopsis thaliana)
10
26
Kolukisaoglu et al., 2004
油菜(Brassica napus)
7
23
Zhang et al., 2014a
杨树(Populus trichocarpa)
10
25
Weinl and Kudla, 2009
辣椒(Capsicum annuum)
9
26
Ma et al., 2019
水稻(Oryza sativa)
10
31
Weinl and Kudla, 2009; Piao et al., 2010
茶(Camellia sinensis)
7
18
Liu et al., 2019
小麦(Triticum aestivum)
24
79
Sun et al., 2015; Liu et al., 2018
芜菁(B. rapa var. rapa)
19
51
Yin et al., 2017
木薯(Manihot esculenta)
8
26
Hu et al., 2015; Mo et al., 2018
茄子(Solanum melongena)
5
15
Li et al., 2016
菠萝(Ananas comosus)
8
21
Aslam et al., 2019
葡萄(Vitis vinifera)
8
20
Xi et al., 2017
玉米(Zea mays)
8
43
Chen et al., 2011
甜菜(Beta vulgaris)
7
20
未发表数据
2 植物CBL-CIPK系统与磷酸化 CBL-CIPK复合物对下游靶标蛋白的调节主要通过磷酸化方式进行(Sanyal et al., 2020).靶标蛋白主要有通道蛋白、转运蛋白和转录因子等(Sanyal et al., 2015).已有研究表明, 可被CBL-CIPK复合物磷酸化的阳离子(H+、Na+、K+、Mg2+、Fe2+和NH4+)相关蛋白如AHA2、SOS1、AMT1、ATL31、AKT1、HAK5、TPK1、ARF、ERF、SnRK2D和FIT, 而一些阴离子(OH-和NO3-)转运蛋白如CHL1 (NRT1.1)、RBOH-F和RBOH-C也可被CBL-CIPK磷酸化(Ragel et al.,2019; Lara et al., 2020; Sanyal et al., 2020; Dong et al., 2021). ...
... 低浓度Na+对植物生长有一定的刺激作用, 但高浓度Na+则会引起细胞渗透和离子毒害(Zhu, 2016).拟南芥SOS信号途径是调节植物根耐盐性的重要机制(Zhu et al., 1998; Yin et al., 2020).在盐胁迫下, AtCBL10与AtCIPK24 (SOS2)互作(图1; 表2), 将Na+区域化到液泡, 以保护植物地上部免受盐胁迫伤害(Hu et al., 2015; Plasencia et al., 2021).这与SOS通路的耐盐机制不同(Yang et al., 2019).被激活的SOS1利用H+-ATPase (AHA2)产生的质子梯度能量将Na+转运出细胞(图1) (Yin et al., 2020).CBL10与CIPK8形成的复合物介导SOS1的激活, 表明CIPK8在调节拟南芥地上部响应盐胁迫中起重要作用(Yin et al., 2020), 但其复合物下游靶标蛋白还有待深入鉴定.细胞内较高的K+/Na+比是植物耐盐性的核心指标(Zhang et al., 2020).研究表明, HKT1 (high affinity potassium transporters 1)能够维持细胞内高浓度的K+和低浓度Na+ (Almeida et al., 2017).水稻(Oryza sativa) OsHKT1;1主要在地上部维管组织中表达, 高盐条件下可增强细胞的Na+外排能力(Campbell et al., 2017).此外, 过表达OsHKT1;4的转基因植株木质部中Na+含量降低, 而根中的Na+含量增加, 从而提高对盐胁迫的敏感性(Oda et al., 2018).CBL2/CBL3-CIPK21也参与植物的耐盐性, 该复合物主要在液泡中发挥作用, 但其下游靶点仍有待确定(Sanyal et al., 2016). ...
A gibberellin-regulated calcineurin B in rice localizes to the tonoplast and is implicated in vacuole function 1 2005
... 目前, CBL-CIPK复合物对植物激素调控的研究主要集中在ABA信号通路.研究发现, CBL9-CIPK3在ABA信号通路中起负向调控作用(Pandey et al., 2008).拟南芥CBL9-CIPK3复合物通过将转录因子ABI1磷酸化, 形成CBL9-CIPK3-ABR1途径(表2), 从而介导种子萌发和ABA依赖的生理生化反应过程(Sanyal et al., 2017).在小麦(Triticum aestivum)中, TaCBL1- TaCIPK23介导ABA信号途径, 并在干旱胁迫应答中起重要作用(Cui et al., 2018).此外, 拟南芥CBL2和CBL3与CIPK9和CIPK27也参与ABA信号通路的负向调控(Song et al., 2018).OsCBL2可正向调控赤霉素的合成(Hwang et al., 2005), 而AtCIPK6参与调控生长素的运输(Tripathi et al., 2009).上述结果表明, CBL-CIPK复合物广泛参与多种植物激素的调控. ...
The calcium sensor CBL10 mediates salt tolerance by regulating ion homeostasis in Arabidopsis 1 2007
... 系统发育分析表明, 拟南芥AtCBLs可分为3类.AtCBL10是第1类中唯一的成员, 并且在CBL家族中具有最长的N端(Sánchez-Barrena et al., 2013).共聚焦荧光显微分析表明, AtCBL10定位于质膜和液泡膜(Kim et al., 2007; Batistič et al., 2010).第2类有5个成员(AtCBL1、-4、-5、-8和-9), 它们具有N端很短的结构特点(Sánchez-Barrena et al., 2013); 其中大多数包含用于脂双层修饰的保守MGCXXS/T基序, 可帮助CBLs固定在膜上(Tang et al., 2015; Saito et al., 2018).第3类成员具有相对较长的N端(Sánchez- Barrena et al., 2013), 它们均具有液泡膜靶向序列(tonoplast targeting sequence, TTS) MSQCXDGXK HXCXSXXXCF (除AtCBL7外) (Tang et al., 2012; Kleist et al., 2014).TTS则有助于AtCBL2、-3和-6定位于液泡膜(Batistič et al., 2010; Tang et al., 2012).CBL7和CBL3是串联复制, 但它们的N端结构域明显不同(Kleist et al., 2014; Sanyal et al., 2016).CBL7作为Ca2+感应蛋白, 能够解码细胞核和细胞质的Ca2+信号(Batistič et al., 2010). ...
The CBL-interacting protein kinase CIPK26 is a novel interactor of Arabidopsis NADPH oxidase AtRbohF that negatively modulates its ROS-producing activity in a heterologous expression system 1 2013
... 与ROS有关的氧化酶RBOHF (respiratory burst oxidase homolog factor)受到CBL1/9-CIPK26的调控(表2; 图2) (Zhang et al., 2014b).CIPK26可影响定位在质膜上的RBOHF活性(Kimura et al., 2013).然而, 该结果是在异源表达系统中试验得到的, 因此CIPK26对RBOHF活性的调节作用尚缺乏足够的证据.但可以肯定的是, CBL1/CBL9介导CIPK26到质膜的定位导致RBHOF磷酸化(Zhang et al., 2018b; Han et al., 2019). ...
Comparative phylogenomics of the CBL-CIPK calcium-decoding network in the moss Physcomitrella, Arabidopsis, and other green lineages 3 2014
... 植物CBLs是与酵母(Saccharomyces cerevisiae)和动物中的CNB (calcineurin B subunit)最相似的一个Ca2+传感器蛋白家族(Cui et al., 2018).CBL感知应激信号中的Ca2+信号, 使植物适应环境变化(Bender et al., 2018; Tang et al., 2020).CBL含有1个典型的Ca2+结合基序, 即4个手性延伸因子(elongation factor hands, EF-hands), 每个EF-hands包含1个能与Ca2+结合的保守α-螺旋-环-α-螺旋结构(Lu et al., 2017).EF-hands排列在固定的空间中, 从EF-hand1到EF-hand4依次有22、25和32个氨基酸的距离(Sánchez-Barrena et al., 2005).环区的特征是具有12个残基DKDGDGKIDFEE的保守序列(Kolukisaoglu et al., 2004).位置1(X)、3(Y)、5(Z)、7(-X)、9(-Y)和12(-Z)中的氨基酸高度保守, 并负责与Ca2+配位(Sanyal et al., 2015).值得注意的是, EF-hand1在X位置和Y位置之间包含2个插入氨基酸残基的位置(Sánchez-Barrena et al., 2013).这个位置氨基酸的变化导致与Ca2+结合的亲和力发生变化(Kolukisaoglu et al., 2004).除了具有Ca2+感应的EF-hands, CBL还具有N端定位基序和C端磷酸化基序(Beckmann et al., 2016).研究表明, 拟南芥(Arabidopsis thaliana) CBL家族4个成员(AtCBL1、-4、-5和-9)具有N-豆蔻酰化基序(Wang et al., 2019).在SOS (salt overly sensitive)信号系统中, CBL4/SOS3的N-豆蔻酰化基序是植物耐盐性的必备条件(Kleist et al., 2014; Saito et al., 2018), 这为Ca2+信号的传递提供了结构基础. ...
... 系统发育分析表明, 拟南芥AtCBLs可分为3类.AtCBL10是第1类中唯一的成员, 并且在CBL家族中具有最长的N端(Sánchez-Barrena et al., 2013).共聚焦荧光显微分析表明, AtCBL10定位于质膜和液泡膜(Kim et al., 2007; Batistič et al., 2010).第2类有5个成员(AtCBL1、-4、-5、-8和-9), 它们具有N端很短的结构特点(Sánchez-Barrena et al., 2013); 其中大多数包含用于脂双层修饰的保守MGCXXS/T基序, 可帮助CBLs固定在膜上(Tang et al., 2015; Saito et al., 2018).第3类成员具有相对较长的N端(Sánchez- Barrena et al., 2013), 它们均具有液泡膜靶向序列(tonoplast targeting sequence, TTS) MSQCXDGXK HXCXSXXXCF (除AtCBL7外) (Tang et al., 2012; Kleist et al., 2014).TTS则有助于AtCBL2、-3和-6定位于液泡膜(Batistič et al., 2010; Tang et al., 2012).CBL7和CBL3是串联复制, 但它们的N端结构域明显不同(Kleist et al., 2014; Sanyal et al., 2016).CBL7作为Ca2+感应蛋白, 能够解码细胞核和细胞质的Ca2+信号(Batistič et al., 2010). ...
... ).CBL7和CBL3是串联复制, 但它们的N端结构域明显不同(Kleist et al., 2014; Sanyal et al., 2016).CBL7作为Ca2+感应蛋白, 能够解码细胞核和细胞质的Ca2+信号(Batistič et al., 2010). ...
Calcium sensors and their interacting protein kinases: genomics of the Arabidopsis and rice CBL-CIPK signaling networks 6 2004
... 植物CBLs是与酵母(Saccharomyces cerevisiae)和动物中的CNB (calcineurin B subunit)最相似的一个Ca2+传感器蛋白家族(Cui et al., 2018).CBL感知应激信号中的Ca2+信号, 使植物适应环境变化(Bender et al., 2018; Tang et al., 2020).CBL含有1个典型的Ca2+结合基序, 即4个手性延伸因子(elongation factor hands, EF-hands), 每个EF-hands包含1个能与Ca2+结合的保守α-螺旋-环-α-螺旋结构(Lu et al., 2017).EF-hands排列在固定的空间中, 从EF-hand1到EF-hand4依次有22、25和32个氨基酸的距离(Sánchez-Barrena et al., 2005).环区的特征是具有12个残基DKDGDGKIDFEE的保守序列(Kolukisaoglu et al., 2004).位置1(X)、3(Y)、5(Z)、7(-X)、9(-Y)和12(-Z)中的氨基酸高度保守, 并负责与Ca2+配位(Sanyal et al., 2015).值得注意的是, EF-hand1在X位置和Y位置之间包含2个插入氨基酸残基的位置(Sánchez-Barrena et al., 2013).这个位置氨基酸的变化导致与Ca2+结合的亲和力发生变化(Kolukisaoglu et al., 2004).除了具有Ca2+感应的EF-hands, CBL还具有N端定位基序和C端磷酸化基序(Beckmann et al., 2016).研究表明, 拟南芥(Arabidopsis thaliana) CBL家族4个成员(AtCBL1、-4、-5和-9)具有N-豆蔻酰化基序(Wang et al., 2019).在SOS (salt overly sensitive)信号系统中, CBL4/SOS3的N-豆蔻酰化基序是植物耐盐性的必备条件(Kleist et al., 2014; Saito et al., 2018), 这为Ca2+信号的传递提供了结构基础. ...
... 结合的亲和力发生变化(Kolukisaoglu et al., 2004).除了具有Ca2+感应的EF-hands, CBL还具有N端定位基序和C端磷酸化基序(Beckmann et al., 2016).研究表明, 拟南芥(Arabidopsis thaliana) CBL家族4个成员(AtCBL1、-4、-5和-9)具有N-豆蔻酰化基序(Wang et al., 2019).在SOS (salt overly sensitive)信号系统中, CBL4/SOS3的N-豆蔻酰化基序是植物耐盐性的必备条件(Kleist et al., 2014; Saito et al., 2018), 这为Ca2+信号的传递提供了结构基础. ...
... 根据内含子丰富度, 可将拟南芥AtCIPK家族分为富有内含子和稀有内含子2个进化分支(Mao et al., 2016).在稀有内含子的进化分支中有17个成员(AtCIPK2、-4、-5、-6、-7、-10、-11、-12、-13、-14、-15、-16、-18、-19、-20、-22和-25), 其它9个成员(CIPK1、-3、-8、-9、-17、-21、-23、-24和-26)在富有内含子的进化分支中.分段和串联重复的发现促进了这2个进化分支的扩展(Guo et al., 2001).CIPK在细胞中通常定位于细胞质和细胞核, 且可通过与CBLs互作靶向下游蛋白(Batistič et al., 2010).此外, 系统发育分析结果表明, AtCIPKs家族分为5个亚组: A、B、C、D和E.有意思的是, 含有多个内含子的AtCIPK成员都聚集在A亚组中, 而其它4个亚组中的基因均为少有或无内含子的成员(Kolukisaoglu et al., 2004). ...
... CBL和CIPK的生理学功能首次在SOS途径中被发现(Zhu et al., 1998).拟南芥突变体atsos1、atsos2和atsos3在高盐胁迫下产生相同的盐敏感表型.AtSOS3和AtSOS2也分别被称为AtCBL4和AtCIPK24, 而AtSOS3和AtSOS2已被证明协同正向调节质膜Na+/H+转运蛋白AtSOS1的活性, 在高盐胁迫下介导Na+外排(Zhu et al., 1998; Sanyal et al., 2015).随后, 在拟南芥中鉴定出10个CBLs和26个CIPKs (Kolukisaoglu et al., 2004; Yu et al., 2007).随着植物基因组测序的完成, 在不同物种中相继鉴定出数量不等的CBLs和CIPKs家族成员(表1). ...
... The CBL and CIPK families in different species Table 1
物种
CBLs数量
CIPKs数量
参考文献
拟南芥(Arabidopsis thaliana)
10
26
Kolukisaoglu et al., 2004
油菜(Brassica napus)
7
23
Zhang et al., 2014a
杨树(Populus trichocarpa)
10
25
Weinl and Kudla, 2009
辣椒(Capsicum annuum)
9
26
Ma et al., 2019
水稻(Oryza sativa)
10
31
Weinl and Kudla, 2009; Piao et al., 2010
茶(Camellia sinensis)
7
18
Liu et al., 2019
小麦(Triticum aestivum)
24
79
Sun et al., 2015; Liu et al., 2018
芜菁(B. rapa var. rapa)
19
51
Yin et al., 2017
木薯(Manihot esculenta)
8
26
Hu et al., 2015; Mo et al., 2018
茄子(Solanum melongena)
5
15
Li et al., 2016
菠萝(Ananas comosus)
8
21
Aslam et al., 2019
葡萄(Vitis vinifera)
8
20
Xi et al., 2017
玉米(Zea mays)
8
43
Chen et al., 2011
甜菜(Beta vulgaris)
7
20
未发表数据
2 植物CBL-CIPK系统与磷酸化 CBL-CIPK复合物对下游靶标蛋白的调节主要通过磷酸化方式进行(Sanyal et al., 2020).靶标蛋白主要有通道蛋白、转运蛋白和转录因子等(Sanyal et al., 2015).已有研究表明, 可被CBL-CIPK复合物磷酸化的阳离子(H+、Na+、K+、Mg2+、Fe2+和NH4+)相关蛋白如AHA2、SOS1、AMT1、ATL31、AKT1、HAK5、TPK1、ARF、ERF、SnRK2D和FIT, 而一些阴离子(OH-和NO3-)转运蛋白如CHL1 (NRT1.1)、RBOH-F和RBOH-C也可被CBL-CIPK磷酸化(Ragel et al.,2019; Lara et al., 2020; Sanyal et al., 2020; Dong et al., 2021). ...
... 在这些必需的离子中, Ca2+浓度通常相对较低,在细胞质中保持在0.1 mmol·L-1左右(Hepler, 2005).然而, 当植物遭遇干旱、盐碱、病原体侵害或营养缺乏等逆境时, Ca2+通过质膜离子通道流入细胞中或从细胞器中释放, 使胞质Ca2+浓度迅速升高(Zhu, 2016; Manishankar et al., 2018; Toyota et al., 2018).这导致Ca2+结合的感应蛋白构象发生变化, 随之与下游效应蛋白以Ca2+依赖方式相互作用并调控其功能(Tang et al., 2020).胞内Ca2+与CBL在其EF-hands处结合, 而CIPK的C端NAF/FISH结构域与CBL互作(Guo et al., 2001; Kolukisaoglu et al., 2004; Su et al., 2020).CBL与CIPK结合使得CIPK的C端自抑制结构域释放N端激酶结构域, 从而增强CIPK活性(Zhou et al., 2016; Yin et al., 2020). ...
The battle of two ions: Ca2+ signaling against Na+ stress 2 2019
... 在细胞内, Ca2+被认为是无处不在、无时不有的功能性调节者, 对许多生理生化反应和发育过程具有重要调控作用(Yin et al., 2017; Köster et al., 2019).Ca2+信号通过Ca2+感应蛋白解码并传递, 进而引起细胞内各种信号系统对环境变化的响应(张和臣等, 2007; Kudla et al., 2018).在植物中, CBLs (Calcineurin B-like proteins)也称SCaBPs (SOS3-like calcium binding proteins), 是一类独特的Ca2+感应蛋白, 其通过激活CIPKs (CBL-interacting protein kinases)/PKS (protein kinase S)解码Ca2+信号(Sánchez-Barrena et al., 2005; 沈金秋等, 2014; Tang et al., 2020).CBLs和CIPKs互作构成一个复杂的信号网络, 在植物对干旱、低温和盐碱等逆境的应答调控中起核心作用(张和臣等, 2007; Sun et al., 2015; Ma et al., 2017; Xi et al., 2017).CBL家族一个成员可以与CIPK家族一个或多个成员相互作用, 建立不同信号级联, 从而形成精准的CBL-CIPK调控网络(Ma et al., 2020).自CBL-CIPK信号系统被发现以来, 学术界对不同途径的CBL-CIPK复合物的功能、结构、基因表达和调控进行了广泛研究(Weinl and Kudla, 2009; Aslam et al., 2019).本文综述了CBLs和CIPKs的结构、分类及其响应各种非生物胁迫的调控机制, 并展望了未来的研究方向, 以期为作物抗逆性遗传改良提供思路. ...
... 植物通过根系从土壤中吸收必需的离子并分配在整个植物组织中(Köster et al., 2019; Srivastava et al., 2020).大量研究表明, 无机离子在维持植物生长中扮演多种重要角色.K+、NO3-和Cl-可维持细胞渗透压, 从而控制细胞膨压, 这对于细胞扩增、气孔运动和花粉管生长至关重要(Saito and Uozumi, 2019).NO3-、NH4+、SO42-和PO43-等被细胞代谢从而产生各种蛋白质和有机化合物(López-Arredondo et al., 2013, 2014). ...
Advances and current challenges in calcium signaling 2 2018
... 植物在生长过程中经常遇到干旱、盐碱、极端温度、缺K+以及病虫害等多种逆境胁迫(Peck and Mittler, 2020).植物无法逃避逆境, 只能被动应对.因此, 植物在长期进化过程中演化出复杂的信号转导机制, 感知并传递外界变化的信号, 使其对逆境及时做出应激反应(Manik et al., 2015).在植物体内, 许多信号转导途径和调控网络相互作用, 响应特定的生物或非生物胁迫.植物对逆境的响应与激酶、磷酸酶、激素、Ca2+、ROS (reactive oxygen species)以及其它调节蛋白、化合物和小分子相互作用交织在一起, 形成复杂的调控网络(Steinhorst and Kudla, 2013; Sierla et al., 2016; Kudla et al., 2018; Zhang et al., 2018a). ...
... 在细胞内, Ca2+被认为是无处不在、无时不有的功能性调节者, 对许多生理生化反应和发育过程具有重要调控作用(Yin et al., 2017; Köster et al., 2019).Ca2+信号通过Ca2+感应蛋白解码并传递, 进而引起细胞内各种信号系统对环境变化的响应(张和臣等, 2007; Kudla et al., 2018).在植物中, CBLs (Calcineurin B-like proteins)也称SCaBPs (SOS3-like calcium binding proteins), 是一类独特的Ca2+感应蛋白, 其通过激活CIPKs (CBL-interacting protein kinases)/PKS (protein kinase S)解码Ca2+信号(Sánchez-Barrena et al., 2005; 沈金秋等, 2014; Tang et al., 2020).CBLs和CIPKs互作构成一个复杂的信号网络, 在植物对干旱、低温和盐碱等逆境的应答调控中起核心作用(张和臣等, 2007; Sun et al., 2015; Ma et al., 2017; Xi et al., 2017).CBL家族一个成员可以与CIPK家族一个或多个成员相互作用, 建立不同信号级联, 从而形成精准的CBL-CIPK调控网络(Ma et al., 2020).自CBL-CIPK信号系统被发现以来, 学术界对不同途径的CBL-CIPK复合物的功能、结构、基因表达和调控进行了广泛研究(Weinl and Kudla, 2009; Aslam et al., 2019).本文综述了CBLs和CIPKs的结构、分类及其响应各种非生物胁迫的调控机制, 并展望了未来的研究方向, 以期为作物抗逆性遗传改良提供思路. ...
Arabidopsis K+ transporter HAK5-mediated high-affinity root K+ uptake is regulated by protein kinases CIPK1 and CIPK9 3 2020
... CBL-CIPK复合物对下游靶标蛋白的调节主要通过磷酸化方式进行(Sanyal et al., 2020).靶标蛋白主要有通道蛋白、转运蛋白和转录因子等(Sanyal et al., 2015).已有研究表明, 可被CBL-CIPK复合物磷酸化的阳离子(H+、Na+、K+、Mg2+、Fe2+和NH4+)相关蛋白如AHA2、SOS1、AMT1、ATL31、AKT1、HAK5、TPK1、ARF、ERF、SnRK2D和FIT, 而一些阴离子(OH-和NO3-)转运蛋白如CHL1 (NRT1.1)、RBOH-F和RBOH-C也可被CBL-CIPK磷酸化(Ragel et al.,2019; Lara et al., 2020; Sanyal et al., 2020; Dong et al., 2021). ...
... K+是植物细胞中最丰富的离子, 在调节细胞渗透压、膜电位和细胞内pH值方面发挥至关重要的作用(伍国强等, 2017; Almeida et al., 2017; Ragel et al., 2019).K+亏缺会导致拟南芥根中Ca2+迅速增加(Behera et al., 2017).维持根系K+吸收的主要系统有K+通道AKT1和高亲和性K+转运蛋白HAK5 (图1; 表2) (Alemán et al., 2011).CBL1/9与CIPK23结合并在质膜上聚集, CIPK23激活后将AKT1和HAK5磷酸化并激活, 以促进K+的吸收和转运(图1; 表2) (Lara et al., 2020).此外, CBL4-CIPK6复合物作用于AKT2, 可应答K+的外排(图1) (Saito and Uozumi, 2020).液泡膜CBL2/3-CIPK3/9/23/26复合物激活TPKs (two- pore K+ channels), 以维持细胞内K+稳态(Tang et al., 2020; Dong et al., 2021). ...
Nitrate sensing and uptake in Arabidopsis are enhanced by ABI2, a phosphatase inactivated by the stress hormone abscisic acid 3 2015
... 在拟南芥中, CBL-CIPK信号系统调控许多离子转运蛋白(图1, 图2), 包括介导K+ (KAT1、KAT2、GORK、AKT1、AKT2、HAK5和SPIK)、Na+ (SOS1)、NH4+ (AMT1;1和AMT1;2)、NO3- (NRT1.1、NRT2.4和NRT2.5)、Cl- (SLAC1、SLAH2和SLAH3)和H+ (AHA2和V-ATPase)的通道或转运蛋白(Ho et al., 2009; Léran et al., 2015; Liu et al., 2016; Straub et al., 2017; Ragel et al., 2019; Saito and Uozumi, 2019).CBL-CIPK在响应C/N养分及Mg和Fe的摄取中也发挥重要作用(Sanyal et al., 2020).这种Ca2+依赖性磷酸化调节系统的功能可确保植物的生长, 并使其能耐受各种环境胁迫(Sanyal et al., 2020).由此可见, CBL-CIPK复合物的磷酸化调控对象主要是各种离子通道和转运蛋白. ...
... NO3-和NH4+是植物根系从土壤中吸收的2种N素来源(Tang et al., 2020).Ca2+调控多种NRT/NPF和SLAH转运蛋白进行NO3-摄取和转运(Sanyal et al., 2020).NPF6.3通过二聚体作用使NO3-从低亲和性模式转换为高亲和性模式, 此过程通过CBL1/9- CIPK23对Thr101的磷酸化作用进行控制(Ho et al., 2009; Parker and Newstead, 2014; Sun et al., 2014; Chu et al., 2021).NH4+吸收由AMTs (ammonium transporters)介导(Sanyal et al., 2020).高浓度NH4+会替代K+而产生毒害作用, 因此必须控制其在细胞的非毒性水平(Zheng et al., 2015).AMT的2个成员(AMT1;1和AMT1;2)也可被CBL1-CIPK23复合物抑制(表2; 图2) (Straub et al., 2017).因此, CBL1-CIPK23可能在维持根细胞K+稳态和免受NH4+毒害中起关键作用(Zheng et al., 2015).Weng等(2020)研究表明, 高浓度K+降低水稻根系对NH4+的吸收速率, 增强对H+的外排速率, 从而减轻NH4+对水稻的毒害.此外, CBL1/9-CIPK23与SLAH2和SLAH3互作通过吸收或分泌NO3-参与硝酸盐的转运(表2) (Ho et al., 2009; Léran et al., 2015). ...
Identification and characterization of CBL and CIPK gene families in eggplant ( Solanum melongena L.) 1 2016
... The CBL and CIPK families in different species Table 1
物种
CBLs数量
CIPKs数量
参考文献
拟南芥(Arabidopsis thaliana)
10
26
Kolukisaoglu et al., 2004
油菜(Brassica napus)
7
23
Zhang et al., 2014a
杨树(Populus trichocarpa)
10
25
Weinl and Kudla, 2009
辣椒(Capsicum annuum)
9
26
Ma et al., 2019
水稻(Oryza sativa)
10
31
Weinl and Kudla, 2009; Piao et al., 2010
茶(Camellia sinensis)
7
18
Liu et al., 2019
小麦(Triticum aestivum)
24
79
Sun et al., 2015; Liu et al., 2018
芜菁(B. rapa var. rapa)
19
51
Yin et al., 2017
木薯(Manihot esculenta)
8
26
Hu et al., 2015; Mo et al., 2018
茄子(Solanum melongena)
5
15
Li et al., 2016
菠萝(Ananas comosus)
8
21
Aslam et al., 2019
葡萄(Vitis vinifera)
8
20
Xi et al., 2017
玉米(Zea mays)
8
43
Chen et al., 2011
甜菜(Beta vulgaris)
7
20
未发表数据
2 植物CBL-CIPK系统与磷酸化 CBL-CIPK复合物对下游靶标蛋白的调节主要通过磷酸化方式进行(Sanyal et al., 2020).靶标蛋白主要有通道蛋白、转运蛋白和转录因子等(Sanyal et al., 2015).已有研究表明, 可被CBL-CIPK复合物磷酸化的阳离子(H+、Na+、K+、Mg2+、Fe2+和NH4+)相关蛋白如AHA2、SOS1、AMT1、ATL31、AKT1、HAK5、TPK1、ARF、ERF、SnRK2D和FIT, 而一些阴离子(OH-和NO3-)转运蛋白如CHL1 (NRT1.1)、RBOH-F和RBOH-C也可被CBL-CIPK磷酸化(Ragel et al.,2019; Lara et al., 2020; Sanyal et al., 2020; Dong et al., 2021). ...
A calcium-independent activation of the Arabidopsis SOS2-like protein kinase 24 by its interacting SOS3-like calcium binding protein 1 2014
... 大量研究表明, CBL与CIPK互作并将其磷酸化是一种普遍的机制, 而且磷酸化能够增强CBL-CIPK复合物的相互作用, 进而提高CIPK的活性(Hashimoto et al., 2012; Sanyal et al., 2020).在植物中, CIPK对CBL的磷酸化最初在豌豆(Pisum sativum)中被报道, PsCIPK可对PsCBL的Thr残基进行磷酸化(Mahajan et al., 2006), 随后在多个物种中相继发现了这种现象(Sanyal et al., 2016).CBL的C端含有1个由23个氨基酸残基组成的基序, 其中有绝对保守的氨基酸残基P、M、L、F、P和F, 因此称作“PFPF基序”(FPSF结构域) (Sanyal et al., 2020).CIPK能够将FPSF结构域中保守的Ser残基磷酸化(Sanyal et al., 2020).海藻(Sargassum sp.)的部分CBL缺少该结构, 以Asp和Glu残基代替(Hashimoto et al., 2012).CIPK介导CBL磷酸化是部分CBL-CIPK信号系统发挥具体功能的必要步骤.例如, AtCIPK24对AtCBL10的磷酸化可激活Na+转运蛋白NHX7 (Almeida et al., 2017), 而CIPK23对CBL1的磷酸化也可激活K+通道AKT1 (Hashimoto et al., 2012; Sánchez-Barrena et al., 2020).因此, CBL的磷酸化一方面可增强其与CIPK互作; 另一方面又促进CIPK对下游靶标蛋白的转磷酸作用(Lin et al., 2014; Sanyal et al., 2016). ...
Genome-wide identification and expression analysis of calcineurin B-like protein and calcineurin B-like protein-interacting protein kinase family genes in tea plant 1 2019
... The CBL and CIPK families in different species Table 1
物种
CBLs数量
CIPKs数量
参考文献
拟南芥(Arabidopsis thaliana)
10
26
Kolukisaoglu et al., 2004
油菜(Brassica napus)
7
23
Zhang et al., 2014a
杨树(Populus trichocarpa)
10
25
Weinl and Kudla, 2009
辣椒(Capsicum annuum)
9
26
Ma et al., 2019
水稻(Oryza sativa)
10
31
Weinl and Kudla, 2009; Piao et al., 2010
茶(Camellia sinensis)
7
18
Liu et al., 2019
小麦(Triticum aestivum)
24
79
Sun et al., 2015; Liu et al., 2018
芜菁(B. rapa var. rapa)
19
51
Yin et al., 2017
木薯(Manihot esculenta)
8
26
Hu et al., 2015; Mo et al., 2018
茄子(Solanum melongena)
5
15
Li et al., 2016
菠萝(Ananas comosus)
8
21
Aslam et al., 2019
葡萄(Vitis vinifera)
8
20
Xi et al., 2017
玉米(Zea mays)
8
43
Chen et al., 2011
甜菜(Beta vulgaris)
7
20
未发表数据
2 植物CBL-CIPK系统与磷酸化 CBL-CIPK复合物对下游靶标蛋白的调节主要通过磷酸化方式进行(Sanyal et al., 2020).靶标蛋白主要有通道蛋白、转运蛋白和转录因子等(Sanyal et al., 2015).已有研究表明, 可被CBL-CIPK复合物磷酸化的阳离子(H+、Na+、K+、Mg2+、Fe2+和NH4+)相关蛋白如AHA2、SOS1、AMT1、ATL31、AKT1、HAK5、TPK1、ARF、ERF、SnRK2D和FIT, 而一些阴离子(OH-和NO3-)转运蛋白如CHL1 (NRT1.1)、RBOH-F和RBOH-C也可被CBL-CIPK磷酸化(Ragel et al.,2019; Lara et al., 2020; Sanyal et al., 2020; Dong et al., 2021). ...
Receptor-like kinase RUPO interacts with potassium transporters to regulate pollen tube growth and integrity in rice 1 2016
... 在拟南芥中, CBL-CIPK信号系统调控许多离子转运蛋白(图1, 图2), 包括介导K+ (KAT1、KAT2、GORK、AKT1、AKT2、HAK5和SPIK)、Na+ (SOS1)、NH4+ (AMT1;1和AMT1;2)、NO3- (NRT1.1、NRT2.4和NRT2.5)、Cl- (SLAC1、SLAH2和SLAH3)和H+ (AHA2和V-ATPase)的通道或转运蛋白(Ho et al., 2009; Léran et al., 2015; Liu et al., 2016; Straub et al., 2017; Ragel et al., 2019; Saito and Uozumi, 2019).CBL-CIPK在响应C/N养分及Mg和Fe的摄取中也发挥重要作用(Sanyal et al., 2020).这种Ca2+依赖性磷酸化调节系统的功能可确保植物的生长, 并使其能耐受各种环境胁迫(Sanyal et al., 2020).由此可见, CBL-CIPK复合物的磷酸化调控对象主要是各种离子通道和转运蛋白. ...
Corrigendum to: the calcium sensor TaCBL4 and its interacting protein TaCIPK5 are required for wheat resistance to stripe rust fungus 1 2018
... The CBL and CIPK families in different species Table 1
物种
CBLs数量
CIPKs数量
参考文献
拟南芥(Arabidopsis thaliana)
10
26
Kolukisaoglu et al., 2004
油菜(Brassica napus)
7
23
Zhang et al., 2014a
杨树(Populus trichocarpa)
10
25
Weinl and Kudla, 2009
辣椒(Capsicum annuum)
9
26
Ma et al., 2019
水稻(Oryza sativa)
10
31
Weinl and Kudla, 2009; Piao et al., 2010
茶(Camellia sinensis)
7
18
Liu et al., 2019
小麦(Triticum aestivum)
24
79
Sun et al., 2015; Liu et al., 2018
芜菁(B. rapa var. rapa)
19
51
Yin et al., 2017
木薯(Manihot esculenta)
8
26
Hu et al., 2015; Mo et al., 2018
茄子(Solanum melongena)
5
15
Li et al., 2016
菠萝(Ananas comosus)
8
21
Aslam et al., 2019
葡萄(Vitis vinifera)
8
20
Xi et al., 2017
玉米(Zea mays)
8
43
Chen et al., 2011
甜菜(Beta vulgaris)
7
20
未发表数据
2 植物CBL-CIPK系统与磷酸化 CBL-CIPK复合物对下游靶标蛋白的调节主要通过磷酸化方式进行(Sanyal et al., 2020).靶标蛋白主要有通道蛋白、转运蛋白和转录因子等(Sanyal et al., 2015).已有研究表明, 可被CBL-CIPK复合物磷酸化的阳离子(H+、Na+、K+、Mg2+、Fe2+和NH4+)相关蛋白如AHA2、SOS1、AMT1、ATL31、AKT1、HAK5、TPK1、ARF、ERF、SnRK2D和FIT, 而一些阴离子(OH-和NO3-)转运蛋白如CHL1 (NRT1.1)、RBOH-F和RBOH-C也可被CBL-CIPK磷酸化(Ragel et al.,2019; Lara et al., 2020; Sanyal et al., 2020; Dong et al., 2021). ...
Biotechnology of nutrient uptake and assimilation in plants 1 2013
... 植物通过根系从土壤中吸收必需的离子并分配在整个植物组织中(Köster et al., 2019; Srivastava et al., 2020).大量研究表明, 无机离子在维持植物生长中扮演多种重要角色.K+、NO3-和Cl-可维持细胞渗透压, 从而控制细胞膨压, 这对于细胞扩增、气孔运动和花粉管生长至关重要(Saito and Uozumi, 2019).NO3-、NH4+、SO42-和PO43-等被细胞代谢从而产生各种蛋白质和有机化合物(López-Arredondo et al., 2013, 2014). ...
Phosphate nutrition: improving low-phosphate tolerance in crops 1 2014
... 植物通过根系从土壤中吸收必需的离子并分配在整个植物组织中(Köster et al., 2019; Srivastava et al., 2020).大量研究表明, 无机离子在维持植物生长中扮演多种重要角色.K+、NO3-和Cl-可维持细胞渗透压, 从而控制细胞膨压, 这对于细胞扩增、气孔运动和花粉管生长至关重要(Saito and Uozumi, 2019).NO3-、NH4+、SO42-和PO43-等被细胞代谢从而产生各种蛋白质和有机化合物(López-Arredondo et al., 2013, 2014). ...
Genome-wide identification of CBL family and expression analysis of CBLs in response to potassium deficiency in cotton 1 2017
... 植物CBLs是与酵母(Saccharomyces cerevisiae)和动物中的CNB (calcineurin B subunit)最相似的一个Ca2+传感器蛋白家族(Cui et al., 2018).CBL感知应激信号中的Ca2+信号, 使植物适应环境变化(Bender et al., 2018; Tang et al., 2020).CBL含有1个典型的Ca2+结合基序, 即4个手性延伸因子(elongation factor hands, EF-hands), 每个EF-hands包含1个能与Ca2+结合的保守α-螺旋-环-α-螺旋结构(Lu et al., 2017).EF-hands排列在固定的空间中, 从EF-hand1到EF-hand4依次有22、25和32个氨基酸的距离(Sánchez-Barrena et al., 2005).环区的特征是具有12个残基DKDGDGKIDFEE的保守序列(Kolukisaoglu et al., 2004).位置1(X)、3(Y)、5(Z)、7(-X)、9(-Y)和12(-Z)中的氨基酸高度保守, 并负责与Ca2+配位(Sanyal et al., 2015).值得注意的是, EF-hand1在X位置和Y位置之间包含2个插入氨基酸残基的位置(Sánchez-Barrena et al., 2013).这个位置氨基酸的变化导致与Ca2+结合的亲和力发生变化(Kolukisaoglu et al., 2004).除了具有Ca2+感应的EF-hands, CBL还具有N端定位基序和C端磷酸化基序(Beckmann et al., 2016).研究表明, 拟南芥(Arabidopsis thaliana) CBL家族4个成员(AtCBL1、-4、-5和-9)具有N-豆蔻酰化基序(Wang et al., 2019).在SOS (salt overly sensitive)信号系统中, CBL4/SOS3的N-豆蔻酰化基序是植物耐盐性的必备条件(Kleist et al., 2014; Saito et al., 2018), 这为Ca2+信号的传递提供了结构基础. ...
The kinase activity of calcineurin B-like interacting protein kinase 26 (CIPK26) influences its own stability and that of the ABA-regulated ubiquitin ligase, keep on going (KEG) 3 2017
... 已有研究表明, 在ABA信号通路中起作用的转录因子也会被CIPK磷酸化(Sanyal et al., 2016; Zhao et al., 2019).最早的报道是ERF7 (ethylene responsive factor 7)被CIPK15磷酸化(Song et al., 2005).磷酸化后的ERF7可能通过2种机制发挥作用: 一是它们结合含有GCC盒的基因并抑制其转录; 二是ERF7可以将AtSin3和HDA19之间的阻遏复合物靶向移至相关基因的启动子, 以进一步抑制基因转录(Song et al., 2005).与之相反, CIPK26介导的ABI5磷酸化是正向调节因子, 可稳定ABI5并启动ABA的下游反应(Lyzenga et al., 2017).KEG (keep on going)使ABI5和CIPK26受到26S蛋白酶的降解, 从而抵消ABA的作用(Lyzenga et al., 2017).此外, PKS5/CIPK11使ABI5在Ser42处磷酸化并激活, 从而调节基因表达(Zhou et al., 2015).因此, CIPK26和PKS5/CIPK11可能在同一途径中发挥相似作用(Zhou et al., 2015).CIPK3与ABR1 (ABA-repressor 1)相互作用并使其磷酸化, 从而调节种子萌发过程中的ABA应答反应(Sanyal et al., 2017; Wang et al., 2018). ...
... ).KEG (keep on going)使ABI5和CIPK26受到26S蛋白酶的降解, 从而抵消ABA的作用(Lyzenga et al., 2017).此外, PKS5/CIPK11使ABI5在Ser42处磷酸化并激活, 从而调节基因表达(Zhou et al., 2015).因此, CIPK26和PKS5/CIPK11可能在同一途径中发挥相似作用(Zhou et al., 2015).CIPK3与ABR1 (ABA-repressor 1)相互作用并使其磷酸化, 从而调节种子萌发过程中的ABA应答反应(Sanyal et al., 2017; Wang et al., 2018). ...
The calcium sensor CBL7 modulates plant responses to low nitrate in Arabidopsis 1 2015
Identification of CBL and CIPK gene families and functional characterization of CaCIPK1 under Phytophthora capsici in pepper (Capsicum annuum L.) 1 2019
... The CBL and CIPK families in different species Table 1
物种
CBLs数量
CIPKs数量
参考文献
拟南芥(Arabidopsis thaliana)
10
26
Kolukisaoglu et al., 2004
油菜(Brassica napus)
7
23
Zhang et al., 2014a
杨树(Populus trichocarpa)
10
25
Weinl and Kudla, 2009
辣椒(Capsicum annuum)
9
26
Ma et al., 2019
水稻(Oryza sativa)
10
31
Weinl and Kudla, 2009; Piao et al., 2010
茶(Camellia sinensis)
7
18
Liu et al., 2019
小麦(Triticum aestivum)
24
79
Sun et al., 2015; Liu et al., 2018
芜菁(B. rapa var. rapa)
19
51
Yin et al., 2017
木薯(Manihot esculenta)
8
26
Hu et al., 2015; Mo et al., 2018
茄子(Solanum melongena)
5
15
Li et al., 2016
菠萝(Ananas comosus)
8
21
Aslam et al., 2019
葡萄(Vitis vinifera)
8
20
Xi et al., 2017
玉米(Zea mays)
8
43
Chen et al., 2011
甜菜(Beta vulgaris)
7
20
未发表数据
2 植物CBL-CIPK系统与磷酸化 CBL-CIPK复合物对下游靶标蛋白的调节主要通过磷酸化方式进行(Sanyal et al., 2020).靶标蛋白主要有通道蛋白、转运蛋白和转录因子等(Sanyal et al., 2015).已有研究表明, 可被CBL-CIPK复合物磷酸化的阳离子(H+、Na+、K+、Mg2+、Fe2+和NH4+)相关蛋白如AHA2、SOS1、AMT1、ATL31、AKT1、HAK5、TPK1、ARF、ERF、SnRK2D和FIT, 而一些阴离子(OH-和NO3-)转运蛋白如CHL1 (NRT1.1)、RBOH-F和RBOH-C也可被CBL-CIPK磷酸化(Ragel et al.,2019; Lara et al., 2020; Sanyal et al., 2020; Dong et al., 2021). ...
The CBL-CIPK pathway in plant response to stress signals 1 2020
... 在细胞内, Ca2+被认为是无处不在、无时不有的功能性调节者, 对许多生理生化反应和发育过程具有重要调控作用(Yin et al., 2017; Köster et al., 2019).Ca2+信号通过Ca2+感应蛋白解码并传递, 进而引起细胞内各种信号系统对环境变化的响应(张和臣等, 2007; Kudla et al., 2018).在植物中, CBLs (Calcineurin B-like proteins)也称SCaBPs (SOS3-like calcium binding proteins), 是一类独特的Ca2+感应蛋白, 其通过激活CIPKs (CBL-interacting protein kinases)/PKS (protein kinase S)解码Ca2+信号(Sánchez-Barrena et al., 2005; 沈金秋等, 2014; Tang et al., 2020).CBLs和CIPKs互作构成一个复杂的信号网络, 在植物对干旱、低温和盐碱等逆境的应答调控中起核心作用(张和臣等, 2007; Sun et al., 2015; Ma et al., 2017; Xi et al., 2017).CBL家族一个成员可以与CIPK家族一个或多个成员相互作用, 建立不同信号级联, 从而形成精准的CBL-CIPK调控网络(Ma et al., 2020).自CBL-CIPK信号系统被发现以来, 学术界对不同途径的CBL-CIPK复合物的功能、结构、基因表达和调控进行了广泛研究(Weinl and Kudla, 2009; Aslam et al., 2019).本文综述了CBLs和CIPKs的结构、分类及其响应各种非生物胁迫的调控机制, 并展望了未来的研究方向, 以期为作物抗逆性遗传改良提供思路. ...
Identification of important physiological traits and moderators that are associated with improved salt tolerance in CBL and CIPK overexpressors through a meta- analysis 1 2017
... 在细胞内, Ca2+被认为是无处不在、无时不有的功能性调节者, 对许多生理生化反应和发育过程具有重要调控作用(Yin et al., 2017; Köster et al., 2019).Ca2+信号通过Ca2+感应蛋白解码并传递, 进而引起细胞内各种信号系统对环境变化的响应(张和臣等, 2007; Kudla et al., 2018).在植物中, CBLs (Calcineurin B-like proteins)也称SCaBPs (SOS3-like calcium binding proteins), 是一类独特的Ca2+感应蛋白, 其通过激活CIPKs (CBL-interacting protein kinases)/PKS (protein kinase S)解码Ca2+信号(Sánchez-Barrena et al., 2005; 沈金秋等, 2014; Tang et al., 2020).CBLs和CIPKs互作构成一个复杂的信号网络, 在植物对干旱、低温和盐碱等逆境的应答调控中起核心作用(张和臣等, 2007; Sun et al., 2015; Ma et al., 2017; Xi et al., 2017).CBL家族一个成员可以与CIPK家族一个或多个成员相互作用, 建立不同信号级联, 从而形成精准的CBL-CIPK调控网络(Ma et al., 2020).自CBL-CIPK信号系统被发现以来, 学术界对不同途径的CBL-CIPK复合物的功能、结构、基因表达和调控进行了广泛研究(Weinl and Kudla, 2009; Aslam et al., 2019).本文综述了CBLs和CIPKs的结构、分类及其响应各种非生物胁迫的调控机制, 并展望了未来的研究方向, 以期为作物抗逆性遗传改良提供思路. ...
Cloning and characterization of CBL-CIPK signaling components from a legume ( Pisum sativum) 1 2006
... 大量研究表明, CBL与CIPK互作并将其磷酸化是一种普遍的机制, 而且磷酸化能够增强CBL-CIPK复合物的相互作用, 进而提高CIPK的活性(Hashimoto et al., 2012; Sanyal et al., 2020).在植物中, CIPK对CBL的磷酸化最初在豌豆(Pisum sativum)中被报道, PsCIPK可对PsCBL的Thr残基进行磷酸化(Mahajan et al., 2006), 随后在多个物种中相继发现了这种现象(Sanyal et al., 2016).CBL的C端含有1个由23个氨基酸残基组成的基序, 其中有绝对保守的氨基酸残基P、M、L、F、P和F, 因此称作“PFPF基序”(FPSF结构域) (Sanyal et al., 2020).CIPK能够将FPSF结构域中保守的Ser残基磷酸化(Sanyal et al., 2020).海藻(Sargassum sp.)的部分CBL缺少该结构, 以Asp和Glu残基代替(Hashimoto et al., 2012).CIPK介导CBL磷酸化是部分CBL-CIPK信号系统发挥具体功能的必要步骤.例如, AtCIPK24对AtCBL10的磷酸化可激活Na+转运蛋白NHX7 (Almeida et al., 2017), 而CIPK23对CBL1的磷酸化也可激活K+通道AKT1 (Hashimoto et al., 2012; Sánchez-Barrena et al., 2020).因此, CBL的磷酸化一方面可增强其与CIPK互作; 另一方面又促进CIPK对下游靶标蛋白的转磷酸作用(Lin et al., 2014; Sanyal et al., 2016). ...
Site- and kinase-specific phosphorylation-mediated activation of SLAC1, a guard cell anion channel stimulated by abscisic acid 2 2014
... CBL1/9-CIPK23与SLAC1或SLAC3相互作用(表2; 图2), 参与ABA (abscisic acid)对气孔大小的调节, 从而导致气孔关闭(Maierhofer et al., 2014a, 2014b).研究表明, H+-ATPase也与相对应的CBL- CIPK互作而发挥作用(图1; 表2).质膜H+-ATPase受CBL2-CIPK11调控(Saito and Uozumi, 2020), 而液泡膜H+-ATPase则受CBL2/3-CIPK3/9/23/26调控(Tang et al., 2012, 2015). ...
The calcium sensor CBL-CIPK is involved in plant’s response to abiotic stresses 1 2015
... 植物在生长过程中经常遇到干旱、盐碱、极端温度、缺K+以及病虫害等多种逆境胁迫(Peck and Mittler, 2020).植物无法逃避逆境, 只能被动应对.因此, 植物在长期进化过程中演化出复杂的信号转导机制, 感知并传递外界变化的信号, 使其对逆境及时做出应激反应(Manik et al., 2015).在植物体内, 许多信号转导途径和调控网络相互作用, 响应特定的生物或非生物胁迫.植物对逆境的响应与激酶、磷酸酶、激素、Ca2+、ROS (reactive oxygen species)以及其它调节蛋白、化合物和小分子相互作用交织在一起, 形成复杂的调控网络(Steinhorst and Kudla, 2013; Sierla et al., 2016; Kudla et al., 2018; Zhang et al., 2018a). ...
Calcium signaling during salt stress and in the regulation of ion homeostasis 1 2018
... 在这些必需的离子中, Ca2+浓度通常相对较低,在细胞质中保持在0.1 mmol·L-1左右(Hepler, 2005).然而, 当植物遭遇干旱、盐碱、病原体侵害或营养缺乏等逆境时, Ca2+通过质膜离子通道流入细胞中或从细胞器中释放, 使胞质Ca2+浓度迅速升高(Zhu, 2016; Manishankar et al., 2018; Toyota et al., 2018).这导致Ca2+结合的感应蛋白构象发生变化, 随之与下游效应蛋白以Ca2+依赖方式相互作用并调控其功能(Tang et al., 2020).胞内Ca2+与CBL在其EF-hands处结合, 而CIPK的C端NAF/FISH结构域与CBL互作(Guo et al., 2001; Kolukisaoglu et al., 2004; Su et al., 2020).CBL与CIPK结合使得CIPK的C端自抑制结构域释放N端激酶结构域, 从而增强CIPK活性(Zhou et al., 2016; Yin et al., 2020). ...
Mechanisms and physiological roles of the CBL-CIPK networking system in Arabidopsis thaliana 1 2016
... 根据内含子丰富度, 可将拟南芥AtCIPK家族分为富有内含子和稀有内含子2个进化分支(Mao et al., 2016).在稀有内含子的进化分支中有17个成员(AtCIPK2、-4、-5、-6、-7、-10、-11、-12、-13、-14、-15、-16、-18、-19、-20、-22和-25), 其它9个成员(CIPK1、-3、-8、-9、-17、-21、-23、-24和-26)在富有内含子的进化分支中.分段和串联重复的发现促进了这2个进化分支的扩展(Guo et al., 2001).CIPK在细胞中通常定位于细胞质和细胞核, 且可通过与CBLs互作靶向下游蛋白(Batistič et al., 2010).此外, 系统发育分析结果表明, AtCIPKs家族分为5个亚组: A、B、C、D和E.有意思的是, 含有多个内含子的AtCIPK成员都聚集在A亚组中, 而其它4个亚组中的基因均为少有或无内含子的成员(Kolukisaoglu et al., 2004). ...
Expression patterns and identified protein-protein interactions suggest that cassava CBL-CIPK signal networks function in responses to abiotic stresses 2 2018
... 植物CIPK在功能上类似于酵母SNF1 (sucrose non- fermenting 1)和哺乳动物AMPK (AMP-dependent kinase) (Mo et al., 2018; Niu et al., 2018).CIPK由2个结构域组成, 一个含有磷酸化位点激活环的保守N端激酶催化结构域, 另一个具有NAF/FISL基序和PPI (protein phosphatase interaction)基序高度不同的C端调节域(Sánchez-Barrena et al., 2013; Sanyal et al., 2015).NAF基序以其高度保守的氨基酸Asn (N)、Ala (A)、Phe (F)、Ile (I)、Ser (S)和Leu (L)而得名, 是结合CBL的必需位点, 其对于维持AtCIPK24和AtCBL4互作是必需的, 并且能够连接AtCIPK24的C端调节结构域以覆盖其激活环, 从而使激酶保持在自动抑制状态(Chaves-Sanjuan et al., 2014).CIPK是具有激酶和调节结构域的Ser/Thr蛋白激酶(Tang et al., 2020).显然, 磷酸化是该系统发挥相应功能的主要作用方式.因此, 磷酸化是CIPK调节靶标蛋白以及与CBL互作的优选方式(Sanyal et al., 2020). ...
... The CBL and CIPK families in different species Table 1
物种
CBLs数量
CIPKs数量
参考文献
拟南芥(Arabidopsis thaliana)
10
26
Kolukisaoglu et al., 2004
油菜(Brassica napus)
7
23
Zhang et al., 2014a
杨树(Populus trichocarpa)
10
25
Weinl and Kudla, 2009
辣椒(Capsicum annuum)
9
26
Ma et al., 2019
水稻(Oryza sativa)
10
31
Weinl and Kudla, 2009; Piao et al., 2010
茶(Camellia sinensis)
7
18
Liu et al., 2019
小麦(Triticum aestivum)
24
79
Sun et al., 2015; Liu et al., 2018
芜菁(B. rapa var. rapa)
19
51
Yin et al., 2017
木薯(Manihot esculenta)
8
26
Hu et al., 2015; Mo et al., 2018
茄子(Solanum melongena)
5
15
Li et al., 2016
菠萝(Ananas comosus)
8
21
Aslam et al., 2019
葡萄(Vitis vinifera)
8
20
Xi et al., 2017
玉米(Zea mays)
8
43
Chen et al., 2011
甜菜(Beta vulgaris)
7
20
未发表数据
2 植物CBL-CIPK系统与磷酸化 CBL-CIPK复合物对下游靶标蛋白的调节主要通过磷酸化方式进行(Sanyal et al., 2020).靶标蛋白主要有通道蛋白、转运蛋白和转录因子等(Sanyal et al., 2015).已有研究表明, 可被CBL-CIPK复合物磷酸化的阳离子(H+、Na+、K+、Mg2+、Fe2+和NH4+)相关蛋白如AHA2、SOS1、AMT1、ATL31、AKT1、HAK5、TPK1、ARF、ERF、SnRK2D和FIT, 而一些阴离子(OH-和NO3-)转运蛋白如CHL1 (NRT1.1)、RBOH-F和RBOH-C也可被CBL-CIPK磷酸化(Ragel et al.,2019; Lara et al., 2020; Sanyal et al., 2020; Dong et al., 2021). ...
Genome-wide identification and characterization of CIPK family and analysis responses to various stresses in apple (Malus domestica) 1 2018
... 植物CIPK在功能上类似于酵母SNF1 (sucrose non- fermenting 1)和哺乳动物AMPK (AMP-dependent kinase) (Mo et al., 2018; Niu et al., 2018).CIPK由2个结构域组成, 一个含有磷酸化位点激活环的保守N端激酶催化结构域, 另一个具有NAF/FISL基序和PPI (protein phosphatase interaction)基序高度不同的C端调节域(Sánchez-Barrena et al., 2013; Sanyal et al., 2015).NAF基序以其高度保守的氨基酸Asn (N)、Ala (A)、Phe (F)、Ile (I)、Ser (S)和Leu (L)而得名, 是结合CBL的必需位点, 其对于维持AtCIPK24和AtCBL4互作是必需的, 并且能够连接AtCIPK24的C端调节结构域以覆盖其激活环, 从而使激酶保持在自动抑制状态(Chaves-Sanjuan et al., 2014).CIPK是具有激酶和调节结构域的Ser/Thr蛋白激酶(Tang et al., 2020).显然, 磷酸化是该系统发挥相应功能的主要作用方式.因此, 磷酸化是CIPK调节靶标蛋白以及与CBL互作的优选方式(Sanyal et al., 2020). ...
T-DNA tagging-based gain-of- function of OsHKT1;4 reinforces Na exclusion from leaves and stems but triggers Na toxicity in roots of rice under salt stress 1 2018
... 低浓度Na+对植物生长有一定的刺激作用, 但高浓度Na+则会引起细胞渗透和离子毒害(Zhu, 2016).拟南芥SOS信号途径是调节植物根耐盐性的重要机制(Zhu et al., 1998; Yin et al., 2020).在盐胁迫下, AtCBL10与AtCIPK24 (SOS2)互作(图1; 表2), 将Na+区域化到液泡, 以保护植物地上部免受盐胁迫伤害(Hu et al., 2015; Plasencia et al., 2021).这与SOS通路的耐盐机制不同(Yang et al., 2019).被激活的SOS1利用H+-ATPase (AHA2)产生的质子梯度能量将Na+转运出细胞(图1) (Yin et al., 2020).CBL10与CIPK8形成的复合物介导SOS1的激活, 表明CIPK8在调节拟南芥地上部响应盐胁迫中起重要作用(Yin et al., 2020), 但其复合物下游靶标蛋白还有待深入鉴定.细胞内较高的K+/Na+比是植物耐盐性的核心指标(Zhang et al., 2020).研究表明, HKT1 (high affinity potassium transporters 1)能够维持细胞内高浓度的K+和低浓度Na+ (Almeida et al., 2017).水稻(Oryza sativa) OsHKT1;1主要在地上部维管组织中表达, 高盐条件下可增强细胞的Na+外排能力(Campbell et al., 2017).此外, 过表达OsHKT1;4的转基因植株木质部中Na+含量降低, 而根中的Na+含量增加, 从而提高对盐胁迫的敏感性(Oda et al., 2018).CBL2/CBL3-CIPK21也参与植物的耐盐性, 该复合物主要在液泡中发挥作用, 但其下游靶点仍有待确定(Sanyal et al., 2016). ...
Calcineurin-B-like protein CBL9 interacts with target kinase CIPK3 in the regulation of ABA response in seed germination 1 2008
... 目前, CBL-CIPK复合物对植物激素调控的研究主要集中在ABA信号通路.研究发现, CBL9-CIPK3在ABA信号通路中起负向调控作用(Pandey et al., 2008).拟南芥CBL9-CIPK3复合物通过将转录因子ABI1磷酸化, 形成CBL9-CIPK3-ABR1途径(表2), 从而介导种子萌发和ABA依赖的生理生化反应过程(Sanyal et al., 2017).在小麦(Triticum aestivum)中, TaCBL1- TaCIPK23介导ABA信号途径, 并在干旱胁迫应答中起重要作用(Cui et al., 2018).此外, 拟南芥CBL2和CBL3与CIPK9和CIPK27也参与ABA信号通路的负向调控(Song et al., 2018).OsCBL2可正向调控赤霉素的合成(Hwang et al., 2005), 而AtCIPK6参与调控生长素的运输(Tripathi et al., 2009).上述结果表明, CBL-CIPK复合物广泛参与多种植物激素的调控. ...
Molecular basis of nitrate uptake by the plant nitrate transporter NRT1.1 1 2014
... NO3-和NH4+是植物根系从土壤中吸收的2种N素来源(Tang et al., 2020).Ca2+调控多种NRT/NPF和SLAH转运蛋白进行NO3-摄取和转运(Sanyal et al., 2020).NPF6.3通过二聚体作用使NO3-从低亲和性模式转换为高亲和性模式, 此过程通过CBL1/9- CIPK23对Thr101的磷酸化作用进行控制(Ho et al., 2009; Parker and Newstead, 2014; Sun et al., 2014; Chu et al., 2021).NH4+吸收由AMTs (ammonium transporters)介导(Sanyal et al., 2020).高浓度NH4+会替代K+而产生毒害作用, 因此必须控制其在细胞的非毒性水平(Zheng et al., 2015).AMT的2个成员(AMT1;1和AMT1;2)也可被CBL1-CIPK23复合物抑制(表2; 图2) (Straub et al., 2017).因此, CBL1-CIPK23可能在维持根细胞K+稳态和免受NH4+毒害中起关键作用(Zheng et al., 2015).Weng等(2020)研究表明, 高浓度K+降低水稻根系对NH4+的吸收速率, 增强对H+的外排速率, 从而减轻NH4+对水稻的毒害.此外, CBL1/9-CIPK23与SLAH2和SLAH3互作通过吸收或分泌NO3-参与硝酸盐的转运(表2) (Ho et al., 2009; Léran et al., 2015). ...
Plant signaling in biotic and abiotic stress 1 2020
... 植物在生长过程中经常遇到干旱、盐碱、极端温度、缺K+以及病虫害等多种逆境胁迫(Peck and Mittler, 2020).植物无法逃避逆境, 只能被动应对.因此, 植物在长期进化过程中演化出复杂的信号转导机制, 感知并传递外界变化的信号, 使其对逆境及时做出应激反应(Manik et al., 2015).在植物体内, 许多信号转导途径和调控网络相互作用, 响应特定的生物或非生物胁迫.植物对逆境的响应与激酶、磷酸酶、激素、Ca2+、ROS (reactive oxygen species)以及其它调节蛋白、化合物和小分子相互作用交织在一起, 形成复杂的调控网络(Steinhorst and Kudla, 2013; Sierla et al., 2016; Kudla et al., 2018; Zhang et al., 2018a). ...
OsCIPK31, a CBL-interacting protein kinase is involved in germination and seedling growth under abiotic stress conditions in rice plants 1 2010
... The CBL and CIPK families in different species Table 1
物种
CBLs数量
CIPKs数量
参考文献
拟南芥(Arabidopsis thaliana)
10
26
Kolukisaoglu et al., 2004
油菜(Brassica napus)
7
23
Zhang et al., 2014a
杨树(Populus trichocarpa)
10
25
Weinl and Kudla, 2009
辣椒(Capsicum annuum)
9
26
Ma et al., 2019
水稻(Oryza sativa)
10
31
Weinl and Kudla, 2009; Piao et al., 2010
茶(Camellia sinensis)
7
18
Liu et al., 2019
小麦(Triticum aestivum)
24
79
Sun et al., 2015; Liu et al., 2018
芜菁(B. rapa var. rapa)
19
51
Yin et al., 2017
木薯(Manihot esculenta)
8
26
Hu et al., 2015; Mo et al., 2018
茄子(Solanum melongena)
5
15
Li et al., 2016
菠萝(Ananas comosus)
8
21
Aslam et al., 2019
葡萄(Vitis vinifera)
8
20
Xi et al., 2017
玉米(Zea mays)
8
43
Chen et al., 2011
甜菜(Beta vulgaris)
7
20
未发表数据
2 植物CBL-CIPK系统与磷酸化 CBL-CIPK复合物对下游靶标蛋白的调节主要通过磷酸化方式进行(Sanyal et al., 2020).靶标蛋白主要有通道蛋白、转运蛋白和转录因子等(Sanyal et al., 2015).已有研究表明, 可被CBL-CIPK复合物磷酸化的阳离子(H+、Na+、K+、Mg2+、Fe2+和NH4+)相关蛋白如AHA2、SOS1、AMT1、ATL31、AKT1、HAK5、TPK1、ARF、ERF、SnRK2D和FIT, 而一些阴离子(OH-和NO3-)转运蛋白如CHL1 (NRT1.1)、RBOH-F和RBOH-C也可被CBL-CIPK磷酸化(Ragel et al.,2019; Lara et al., 2020; Sanyal et al., 2020; Dong et al., 2021). ...
The Ca2+ sensor calcineurin B-like protein 10 in plants: emerging new crucial roles for plant abiotic stress tolerance 2 2021
... 低浓度Na+对植物生长有一定的刺激作用, 但高浓度Na+则会引起细胞渗透和离子毒害(Zhu, 2016).拟南芥SOS信号途径是调节植物根耐盐性的重要机制(Zhu et al., 1998; Yin et al., 2020).在盐胁迫下, AtCBL10与AtCIPK24 (SOS2)互作(图1; 表2), 将Na+区域化到液泡, 以保护植物地上部免受盐胁迫伤害(Hu et al., 2015; Plasencia et al., 2021).这与SOS通路的耐盐机制不同(Yang et al., 2019).被激活的SOS1利用H+-ATPase (AHA2)产生的质子梯度能量将Na+转运出细胞(图1) (Yin et al., 2020).CBL10与CIPK8形成的复合物介导SOS1的激活, 表明CIPK8在调节拟南芥地上部响应盐胁迫中起重要作用(Yin et al., 2020), 但其复合物下游靶标蛋白还有待深入鉴定.细胞内较高的K+/Na+比是植物耐盐性的核心指标(Zhang et al., 2020).研究表明, HKT1 (high affinity potassium transporters 1)能够维持细胞内高浓度的K+和低浓度Na+ (Almeida et al., 2017).水稻(Oryza sativa) OsHKT1;1主要在地上部维管组织中表达, 高盐条件下可增强细胞的Na+外排能力(Campbell et al., 2017).此外, 过表达OsHKT1;4的转基因植株木质部中Na+含量降低, 而根中的Na+含量增加, 从而提高对盐胁迫的敏感性(Oda et al., 2018).CBL2/CBL3-CIPK21也参与植物的耐盐性, 该复合物主要在液泡中发挥作用, 但其下游靶点仍有待确定(Sanyal et al., 2016). ...
... CBL-CIPK复合物对下游靶标蛋白的调节主要通过磷酸化方式进行(Sanyal et al., 2020).靶标蛋白主要有通道蛋白、转运蛋白和转录因子等(Sanyal et al., 2015).已有研究表明, 可被CBL-CIPK复合物磷酸化的阳离子(H+、Na+、K+、Mg2+、Fe2+和NH4+)相关蛋白如AHA2、SOS1、AMT1、ATL31、AKT1、HAK5、TPK1、ARF、ERF、SnRK2D和FIT, 而一些阴离子(OH-和NO3-)转运蛋白如CHL1 (NRT1.1)、RBOH-F和RBOH-C也可被CBL-CIPK磷酸化(Ragel et al.,2019; Lara et al., 2020; Sanyal et al., 2020; Dong et al., 2021). ...
... 在拟南芥中, CBL-CIPK信号系统调控许多离子转运蛋白(图1, 图2), 包括介导K+ (KAT1、KAT2、GORK、AKT1、AKT2、HAK5和SPIK)、Na+ (SOS1)、NH4+ (AMT1;1和AMT1;2)、NO3- (NRT1.1、NRT2.4和NRT2.5)、Cl- (SLAC1、SLAH2和SLAH3)和H+ (AHA2和V-ATPase)的通道或转运蛋白(Ho et al., 2009; Léran et al., 2015; Liu et al., 2016; Straub et al., 2017; Ragel et al., 2019; Saito and Uozumi, 2019).CBL-CIPK在响应C/N养分及Mg和Fe的摄取中也发挥重要作用(Sanyal et al., 2020).这种Ca2+依赖性磷酸化调节系统的功能可确保植物的生长, 并使其能耐受各种环境胁迫(Sanyal et al., 2020).由此可见, CBL-CIPK复合物的磷酸化调控对象主要是各种离子通道和转运蛋白. ...
... K+是植物细胞中最丰富的离子, 在调节细胞渗透压、膜电位和细胞内pH值方面发挥至关重要的作用(伍国强等, 2017; Almeida et al., 2017; Ragel et al., 2019).K+亏缺会导致拟南芥根中Ca2+迅速增加(Behera et al., 2017).维持根系K+吸收的主要系统有K+通道AKT1和高亲和性K+转运蛋白HAK5 (图1; 表2) (Alemán et al., 2011).CBL1/9与CIPK23结合并在质膜上聚集, CIPK23激活后将AKT1和HAK5磷酸化并激活, 以促进K+的吸收和转运(图1; 表2) (Lara et al., 2020).此外, CBL4-CIPK6复合物作用于AKT2, 可应答K+的外排(图1) (Saito and Uozumi, 2020).液泡膜CBL2/3-CIPK3/9/23/26复合物激活TPKs (two- pore K+ channels), 以维持细胞内K+稳态(Tang et al., 2020; Dong et al., 2021). ...
N-myristoylation and S-acylation are common modifications of Ca2+-regulated Arabidopsis kinases and are required for activation of the SLAC1 anion channel 3 2018
... 植物CBLs是与酵母(Saccharomyces cerevisiae)和动物中的CNB (calcineurin B subunit)最相似的一个Ca2+传感器蛋白家族(Cui et al., 2018).CBL感知应激信号中的Ca2+信号, 使植物适应环境变化(Bender et al., 2018; Tang et al., 2020).CBL含有1个典型的Ca2+结合基序, 即4个手性延伸因子(elongation factor hands, EF-hands), 每个EF-hands包含1个能与Ca2+结合的保守α-螺旋-环-α-螺旋结构(Lu et al., 2017).EF-hands排列在固定的空间中, 从EF-hand1到EF-hand4依次有22、25和32个氨基酸的距离(Sánchez-Barrena et al., 2005).环区的特征是具有12个残基DKDGDGKIDFEE的保守序列(Kolukisaoglu et al., 2004).位置1(X)、3(Y)、5(Z)、7(-X)、9(-Y)和12(-Z)中的氨基酸高度保守, 并负责与Ca2+配位(Sanyal et al., 2015).值得注意的是, EF-hand1在X位置和Y位置之间包含2个插入氨基酸残基的位置(Sánchez-Barrena et al., 2013).这个位置氨基酸的变化导致与Ca2+结合的亲和力发生变化(Kolukisaoglu et al., 2004).除了具有Ca2+感应的EF-hands, CBL还具有N端定位基序和C端磷酸化基序(Beckmann et al., 2016).研究表明, 拟南芥(Arabidopsis thaliana) CBL家族4个成员(AtCBL1、-4、-5和-9)具有N-豆蔻酰化基序(Wang et al., 2019).在SOS (salt overly sensitive)信号系统中, CBL4/SOS3的N-豆蔻酰化基序是植物耐盐性的必备条件(Kleist et al., 2014; Saito et al., 2018), 这为Ca2+信号的传递提供了结构基础. ...
... 系统发育分析表明, 拟南芥AtCBLs可分为3类.AtCBL10是第1类中唯一的成员, 并且在CBL家族中具有最长的N端(Sánchez-Barrena et al., 2013).共聚焦荧光显微分析表明, AtCBL10定位于质膜和液泡膜(Kim et al., 2007; Batistič et al., 2010).第2类有5个成员(AtCBL1、-4、-5、-8和-9), 它们具有N端很短的结构特点(Sánchez-Barrena et al., 2013); 其中大多数包含用于脂双层修饰的保守MGCXXS/T基序, 可帮助CBLs固定在膜上(Tang et al., 2015; Saito et al., 2018).第3类成员具有相对较长的N端(Sánchez- Barrena et al., 2013), 它们均具有液泡膜靶向序列(tonoplast targeting sequence, TTS) MSQCXDGXK HXCXSXXXCF (除AtCBL7外) (Tang et al., 2012; Kleist et al., 2014).TTS则有助于AtCBL2、-3和-6定位于液泡膜(Batistič et al., 2010; Tang et al., 2012).CBL7和CBL3是串联复制, 但它们的N端结构域明显不同(Kleist et al., 2014; Sanyal et al., 2016).CBL7作为Ca2+感应蛋白, 能够解码细胞核和细胞质的Ca2+信号(Batistič et al., 2010). ...
Guard cell membrane anion transport systems and their regulatory components: an elaborate mechanism controlling stress-induced stomatal closure 2 2019
... 植物通过根系从土壤中吸收必需的离子并分配在整个植物组织中(Köster et al., 2019; Srivastava et al., 2020).大量研究表明, 无机离子在维持植物生长中扮演多种重要角色.K+、NO3-和Cl-可维持细胞渗透压, 从而控制细胞膨压, 这对于细胞扩增、气孔运动和花粉管生长至关重要(Saito and Uozumi, 2019).NO3-、NH4+、SO42-和PO43-等被细胞代谢从而产生各种蛋白质和有机化合物(López-Arredondo et al., 2013, 2014). ...
... 在拟南芥中, CBL-CIPK信号系统调控许多离子转运蛋白(图1, 图2), 包括介导K+ (KAT1、KAT2、GORK、AKT1、AKT2、HAK5和SPIK)、Na+ (SOS1)、NH4+ (AMT1;1和AMT1;2)、NO3- (NRT1.1、NRT2.4和NRT2.5)、Cl- (SLAC1、SLAH2和SLAH3)和H+ (AHA2和V-ATPase)的通道或转运蛋白(Ho et al., 2009; Léran et al., 2015; Liu et al., 2016; Straub et al., 2017; Ragel et al., 2019; Saito and Uozumi, 2019).CBL-CIPK在响应C/N养分及Mg和Fe的摄取中也发挥重要作用(Sanyal et al., 2020).这种Ca2+依赖性磷酸化调节系统的功能可确保植物的生长, 并使其能耐受各种环境胁迫(Sanyal et al., 2020).由此可见, CBL-CIPK复合物的磷酸化调控对象主要是各种离子通道和转运蛋白. ...
Calcium-regulated phosphorylation systems controlling uptake and balance of plant nutrients 4 2020
... K+是植物细胞中最丰富的离子, 在调节细胞渗透压、膜电位和细胞内pH值方面发挥至关重要的作用(伍国强等, 2017; Almeida et al., 2017; Ragel et al., 2019).K+亏缺会导致拟南芥根中Ca2+迅速增加(Behera et al., 2017).维持根系K+吸收的主要系统有K+通道AKT1和高亲和性K+转运蛋白HAK5 (图1; 表2) (Alemán et al., 2011).CBL1/9与CIPK23结合并在质膜上聚集, CIPK23激活后将AKT1和HAK5磷酸化并激活, 以促进K+的吸收和转运(图1; 表2) (Lara et al., 2020).此外, CBL4-CIPK6复合物作用于AKT2, 可应答K+的外排(图1) (Saito and Uozumi, 2020).液泡膜CBL2/3-CIPK3/9/23/26复合物激活TPKs (two- pore K+ channels), 以维持细胞内K+稳态(Tang et al., 2020; Dong et al., 2021). ...
... CBL1/9-CIPK23与SLAC1或SLAC3相互作用(表2; 图2), 参与ABA (abscisic acid)对气孔大小的调节, 从而导致气孔关闭(Maierhofer et al., 2014a, 2014b).研究表明, H+-ATPase也与相对应的CBL- CIPK互作而发挥作用(图1; 表2).质膜H+-ATPase受CBL2-CIPK11调控(Saito and Uozumi, 2020), 而液泡膜H+-ATPase则受CBL2/3-CIPK3/9/23/26调控(Tang et al., 2012, 2015). ...
Recognition and activation of the plant AKT1 potassium channel by the kinase CIPK23 1 2020
... 大量研究表明, CBL与CIPK互作并将其磷酸化是一种普遍的机制, 而且磷酸化能够增强CBL-CIPK复合物的相互作用, 进而提高CIPK的活性(Hashimoto et al., 2012; Sanyal et al., 2020).在植物中, CIPK对CBL的磷酸化最初在豌豆(Pisum sativum)中被报道, PsCIPK可对PsCBL的Thr残基进行磷酸化(Mahajan et al., 2006), 随后在多个物种中相继发现了这种现象(Sanyal et al., 2016).CBL的C端含有1个由23个氨基酸残基组成的基序, 其中有绝对保守的氨基酸残基P、M、L、F、P和F, 因此称作“PFPF基序”(FPSF结构域) (Sanyal et al., 2020).CIPK能够将FPSF结构域中保守的Ser残基磷酸化(Sanyal et al., 2020).海藻(Sargassum sp.)的部分CBL缺少该结构, 以Asp和Glu残基代替(Hashimoto et al., 2012).CIPK介导CBL磷酸化是部分CBL-CIPK信号系统发挥具体功能的必要步骤.例如, AtCIPK24对AtCBL10的磷酸化可激活Na+转运蛋白NHX7 (Almeida et al., 2017), 而CIPK23对CBL1的磷酸化也可激活K+通道AKT1 (Hashimoto et al., 2012; Sánchez-Barrena et al., 2020).因此, CBL的磷酸化一方面可增强其与CIPK互作; 另一方面又促进CIPK对下游靶标蛋白的转磷酸作用(Lin et al., 2014; Sanyal et al., 2016). ...
Structural biology of a major signaling network that regulates plant abiotic stress: the CBL-CIPK mediated pathway 6 2013
... CBL和CIPK结构特征为其相互作用提供了基础.CBL4与CIPK24的C端调节域的复合物晶体结构首先被解析出来(Sánchez-Barrena et al., 2005), 进而揭示了CBL4-CIPK24复合物如何解码由细胞外刺激触发的细胞内Ca2+信号转导(Sánchez-Barrena et al., 2013).阐明CBLs和CIPKs结构特征有助于理解信号系统的调控机制. ...
... 植物CBLs是与酵母(Saccharomyces cerevisiae)和动物中的CNB (calcineurin B subunit)最相似的一个Ca2+传感器蛋白家族(Cui et al., 2018).CBL感知应激信号中的Ca2+信号, 使植物适应环境变化(Bender et al., 2018; Tang et al., 2020).CBL含有1个典型的Ca2+结合基序, 即4个手性延伸因子(elongation factor hands, EF-hands), 每个EF-hands包含1个能与Ca2+结合的保守α-螺旋-环-α-螺旋结构(Lu et al., 2017).EF-hands排列在固定的空间中, 从EF-hand1到EF-hand4依次有22、25和32个氨基酸的距离(Sánchez-Barrena et al., 2005).环区的特征是具有12个残基DKDGDGKIDFEE的保守序列(Kolukisaoglu et al., 2004).位置1(X)、3(Y)、5(Z)、7(-X)、9(-Y)和12(-Z)中的氨基酸高度保守, 并负责与Ca2+配位(Sanyal et al., 2015).值得注意的是, EF-hand1在X位置和Y位置之间包含2个插入氨基酸残基的位置(Sánchez-Barrena et al., 2013).这个位置氨基酸的变化导致与Ca2+结合的亲和力发生变化(Kolukisaoglu et al., 2004).除了具有Ca2+感应的EF-hands, CBL还具有N端定位基序和C端磷酸化基序(Beckmann et al., 2016).研究表明, 拟南芥(Arabidopsis thaliana) CBL家族4个成员(AtCBL1、-4、-5和-9)具有N-豆蔻酰化基序(Wang et al., 2019).在SOS (salt overly sensitive)信号系统中, CBL4/SOS3的N-豆蔻酰化基序是植物耐盐性的必备条件(Kleist et al., 2014; Saito et al., 2018), 这为Ca2+信号的传递提供了结构基础. ...
... 系统发育分析表明, 拟南芥AtCBLs可分为3类.AtCBL10是第1类中唯一的成员, 并且在CBL家族中具有最长的N端(Sánchez-Barrena et al., 2013).共聚焦荧光显微分析表明, AtCBL10定位于质膜和液泡膜(Kim et al., 2007; Batistič et al., 2010).第2类有5个成员(AtCBL1、-4、-5、-8和-9), 它们具有N端很短的结构特点(Sánchez-Barrena et al., 2013); 其中大多数包含用于脂双层修饰的保守MGCXXS/T基序, 可帮助CBLs固定在膜上(Tang et al., 2015; Saito et al., 2018).第3类成员具有相对较长的N端(Sánchez- Barrena et al., 2013), 它们均具有液泡膜靶向序列(tonoplast targeting sequence, TTS) MSQCXDGXK HXCXSXXXCF (除AtCBL7外) (Tang et al., 2012; Kleist et al., 2014).TTS则有助于AtCBL2、-3和-6定位于液泡膜(Batistič et al., 2010; Tang et al., 2012).CBL7和CBL3是串联复制, 但它们的N端结构域明显不同(Kleist et al., 2014; Sanyal et al., 2016).CBL7作为Ca2+感应蛋白, 能够解码细胞核和细胞质的Ca2+信号(Batistič et al., 2010). ...
... ).第2类有5个成员(AtCBL1、-4、-5、-8和-9), 它们具有N端很短的结构特点(Sánchez-Barrena et al., 2013); 其中大多数包含用于脂双层修饰的保守MGCXXS/T基序, 可帮助CBLs固定在膜上(Tang et al., 2015; Saito et al., 2018).第3类成员具有相对较长的N端(Sánchez- Barrena et al., 2013), 它们均具有液泡膜靶向序列(tonoplast targeting sequence, TTS) MSQCXDGXK HXCXSXXXCF (除AtCBL7外) (Tang et al., 2012; Kleist et al., 2014).TTS则有助于AtCBL2、-3和-6定位于液泡膜(Batistič et al., 2010; Tang et al., 2012).CBL7和CBL3是串联复制, 但它们的N端结构域明显不同(Kleist et al., 2014; Sanyal et al., 2016).CBL7作为Ca2+感应蛋白, 能够解码细胞核和细胞质的Ca2+信号(Batistič et al., 2010). ...
... ).第3类成员具有相对较长的N端(Sánchez- Barrena et al., 2013), 它们均具有液泡膜靶向序列(tonoplast targeting sequence, TTS) MSQCXDGXK HXCXSXXXCF (除AtCBL7外) (Tang et al., 2012; Kleist et al., 2014).TTS则有助于AtCBL2、-3和-6定位于液泡膜(Batistič et al., 2010; Tang et al., 2012).CBL7和CBL3是串联复制, 但它们的N端结构域明显不同(Kleist et al., 2014; Sanyal et al., 2016).CBL7作为Ca2+感应蛋白, 能够解码细胞核和细胞质的Ca2+信号(Batistič et al., 2010). ...
... 植物CIPK在功能上类似于酵母SNF1 (sucrose non- fermenting 1)和哺乳动物AMPK (AMP-dependent kinase) (Mo et al., 2018; Niu et al., 2018).CIPK由2个结构域组成, 一个含有磷酸化位点激活环的保守N端激酶催化结构域, 另一个具有NAF/FISL基序和PPI (protein phosphatase interaction)基序高度不同的C端调节域(Sánchez-Barrena et al., 2013; Sanyal et al., 2015).NAF基序以其高度保守的氨基酸Asn (N)、Ala (A)、Phe (F)、Ile (I)、Ser (S)和Leu (L)而得名, 是结合CBL的必需位点, 其对于维持AtCIPK24和AtCBL4互作是必需的, 并且能够连接AtCIPK24的C端调节结构域以覆盖其激活环, 从而使激酶保持在自动抑制状态(Chaves-Sanjuan et al., 2014).CIPK是具有激酶和调节结构域的Ser/Thr蛋白激酶(Tang et al., 2020).显然, 磷酸化是该系统发挥相应功能的主要作用方式.因此, 磷酸化是CIPK调节靶标蛋白以及与CBL互作的优选方式(Sanyal et al., 2020). ...
The structure of the Arabidopsis thaliana SOS3: molecular mechanism of sensing calcium for salt stress response 3 2005
... 在细胞内, Ca2+被认为是无处不在、无时不有的功能性调节者, 对许多生理生化反应和发育过程具有重要调控作用(Yin et al., 2017; Köster et al., 2019).Ca2+信号通过Ca2+感应蛋白解码并传递, 进而引起细胞内各种信号系统对环境变化的响应(张和臣等, 2007; Kudla et al., 2018).在植物中, CBLs (Calcineurin B-like proteins)也称SCaBPs (SOS3-like calcium binding proteins), 是一类独特的Ca2+感应蛋白, 其通过激活CIPKs (CBL-interacting protein kinases)/PKS (protein kinase S)解码Ca2+信号(Sánchez-Barrena et al., 2005; 沈金秋等, 2014; Tang et al., 2020).CBLs和CIPKs互作构成一个复杂的信号网络, 在植物对干旱、低温和盐碱等逆境的应答调控中起核心作用(张和臣等, 2007; Sun et al., 2015; Ma et al., 2017; Xi et al., 2017).CBL家族一个成员可以与CIPK家族一个或多个成员相互作用, 建立不同信号级联, 从而形成精准的CBL-CIPK调控网络(Ma et al., 2020).自CBL-CIPK信号系统被发现以来, 学术界对不同途径的CBL-CIPK复合物的功能、结构、基因表达和调控进行了广泛研究(Weinl and Kudla, 2009; Aslam et al., 2019).本文综述了CBLs和CIPKs的结构、分类及其响应各种非生物胁迫的调控机制, 并展望了未来的研究方向, 以期为作物抗逆性遗传改良提供思路. ...
... CBL和CIPK结构特征为其相互作用提供了基础.CBL4与CIPK24的C端调节域的复合物晶体结构首先被解析出来(Sánchez-Barrena et al., 2005), 进而揭示了CBL4-CIPK24复合物如何解码由细胞外刺激触发的细胞内Ca2+信号转导(Sánchez-Barrena et al., 2013).阐明CBLs和CIPKs结构特征有助于理解信号系统的调控机制. ...
... 植物CBLs是与酵母(Saccharomyces cerevisiae)和动物中的CNB (calcineurin B subunit)最相似的一个Ca2+传感器蛋白家族(Cui et al., 2018).CBL感知应激信号中的Ca2+信号, 使植物适应环境变化(Bender et al., 2018; Tang et al., 2020).CBL含有1个典型的Ca2+结合基序, 即4个手性延伸因子(elongation factor hands, EF-hands), 每个EF-hands包含1个能与Ca2+结合的保守α-螺旋-环-α-螺旋结构(Lu et al., 2017).EF-hands排列在固定的空间中, 从EF-hand1到EF-hand4依次有22、25和32个氨基酸的距离(Sánchez-Barrena et al., 2005).环区的特征是具有12个残基DKDGDGKIDFEE的保守序列(Kolukisaoglu et al., 2004).位置1(X)、3(Y)、5(Z)、7(-X)、9(-Y)和12(-Z)中的氨基酸高度保守, 并负责与Ca2+配位(Sanyal et al., 2015).值得注意的是, EF-hand1在X位置和Y位置之间包含2个插入氨基酸残基的位置(Sánchez-Barrena et al., 2013).这个位置氨基酸的变化导致与Ca2+结合的亲和力发生变化(Kolukisaoglu et al., 2004).除了具有Ca2+感应的EF-hands, CBL还具有N端定位基序和C端磷酸化基序(Beckmann et al., 2016).研究表明, 拟南芥(Arabidopsis thaliana) CBL家族4个成员(AtCBL1、-4、-5和-9)具有N-豆蔻酰化基序(Wang et al., 2019).在SOS (salt overly sensitive)信号系统中, CBL4/SOS3的N-豆蔻酰化基序是植物耐盐性的必备条件(Kleist et al., 2014; Saito et al., 2018), 这为Ca2+信号的传递提供了结构基础. ...
Arabidopsis CBL interacting protein kinase 3 interacts with ABR1, an APETALA2 domain transcription factor, to regulate ABA responses 3 2017
... 已有研究表明, 在ABA信号通路中起作用的转录因子也会被CIPK磷酸化(Sanyal et al., 2016; Zhao et al., 2019).最早的报道是ERF7 (ethylene responsive factor 7)被CIPK15磷酸化(Song et al., 2005).磷酸化后的ERF7可能通过2种机制发挥作用: 一是它们结合含有GCC盒的基因并抑制其转录; 二是ERF7可以将AtSin3和HDA19之间的阻遏复合物靶向移至相关基因的启动子, 以进一步抑制基因转录(Song et al., 2005).与之相反, CIPK26介导的ABI5磷酸化是正向调节因子, 可稳定ABI5并启动ABA的下游反应(Lyzenga et al., 2017).KEG (keep on going)使ABI5和CIPK26受到26S蛋白酶的降解, 从而抵消ABA的作用(Lyzenga et al., 2017).此外, PKS5/CIPK11使ABI5在Ser42处磷酸化并激活, 从而调节基因表达(Zhou et al., 2015).因此, CIPK26和PKS5/CIPK11可能在同一途径中发挥相似作用(Zhou et al., 2015).CIPK3与ABR1 (ABA-repressor 1)相互作用并使其磷酸化, 从而调节种子萌发过程中的ABA应答反应(Sanyal et al., 2017; Wang et al., 2018). ...
... 目前, CBL-CIPK复合物对植物激素调控的研究主要集中在ABA信号通路.研究发现, CBL9-CIPK3在ABA信号通路中起负向调控作用(Pandey et al., 2008).拟南芥CBL9-CIPK3复合物通过将转录因子ABI1磷酸化, 形成CBL9-CIPK3-ABR1途径(表2), 从而介导种子萌发和ABA依赖的生理生化反应过程(Sanyal et al., 2017).在小麦(Triticum aestivum)中, TaCBL1- TaCIPK23介导ABA信号途径, 并在干旱胁迫应答中起重要作用(Cui et al., 2018).此外, 拟南芥CBL2和CBL3与CIPK9和CIPK27也参与ABA信号通路的负向调控(Song et al., 2018).OsCBL2可正向调控赤霉素的合成(Hwang et al., 2005), 而AtCIPK6参与调控生长素的运输(Tripathi et al., 2009).上述结果表明, CBL-CIPK复合物广泛参与多种植物激素的调控. ...
CBL-CIPK module-mediated phosphoregulation: facts and hypothesis 11 2020
... 植物CIPK在功能上类似于酵母SNF1 (sucrose non- fermenting 1)和哺乳动物AMPK (AMP-dependent kinase) (Mo et al., 2018; Niu et al., 2018).CIPK由2个结构域组成, 一个含有磷酸化位点激活环的保守N端激酶催化结构域, 另一个具有NAF/FISL基序和PPI (protein phosphatase interaction)基序高度不同的C端调节域(Sánchez-Barrena et al., 2013; Sanyal et al., 2015).NAF基序以其高度保守的氨基酸Asn (N)、Ala (A)、Phe (F)、Ile (I)、Ser (S)和Leu (L)而得名, 是结合CBL的必需位点, 其对于维持AtCIPK24和AtCBL4互作是必需的, 并且能够连接AtCIPK24的C端调节结构域以覆盖其激活环, 从而使激酶保持在自动抑制状态(Chaves-Sanjuan et al., 2014).CIPK是具有激酶和调节结构域的Ser/Thr蛋白激酶(Tang et al., 2020).显然, 磷酸化是该系统发挥相应功能的主要作用方式.因此, 磷酸化是CIPK调节靶标蛋白以及与CBL互作的优选方式(Sanyal et al., 2020). ...
... CBL-CIPK复合物对下游靶标蛋白的调节主要通过磷酸化方式进行(Sanyal et al., 2020).靶标蛋白主要有通道蛋白、转运蛋白和转录因子等(Sanyal et al., 2015).已有研究表明, 可被CBL-CIPK复合物磷酸化的阳离子(H+、Na+、K+、Mg2+、Fe2+和NH4+)相关蛋白如AHA2、SOS1、AMT1、ATL31、AKT1、HAK5、TPK1、ARF、ERF、SnRK2D和FIT, 而一些阴离子(OH-和NO3-)转运蛋白如CHL1 (NRT1.1)、RBOH-F和RBOH-C也可被CBL-CIPK磷酸化(Ragel et al.,2019; Lara et al., 2020; Sanyal et al., 2020; Dong et al., 2021). ...
... ; Sanyal et al., 2020; Dong et al., 2021). ...
... 大量研究表明, CBL与CIPK互作并将其磷酸化是一种普遍的机制, 而且磷酸化能够增强CBL-CIPK复合物的相互作用, 进而提高CIPK的活性(Hashimoto et al., 2012; Sanyal et al., 2020).在植物中, CIPK对CBL的磷酸化最初在豌豆(Pisum sativum)中被报道, PsCIPK可对PsCBL的Thr残基进行磷酸化(Mahajan et al., 2006), 随后在多个物种中相继发现了这种现象(Sanyal et al., 2016).CBL的C端含有1个由23个氨基酸残基组成的基序, 其中有绝对保守的氨基酸残基P、M、L、F、P和F, 因此称作“PFPF基序”(FPSF结构域) (Sanyal et al., 2020).CIPK能够将FPSF结构域中保守的Ser残基磷酸化(Sanyal et al., 2020).海藻(Sargassum sp.)的部分CBL缺少该结构, 以Asp和Glu残基代替(Hashimoto et al., 2012).CIPK介导CBL磷酸化是部分CBL-CIPK信号系统发挥具体功能的必要步骤.例如, AtCIPK24对AtCBL10的磷酸化可激活Na+转运蛋白NHX7 (Almeida et al., 2017), 而CIPK23对CBL1的磷酸化也可激活K+通道AKT1 (Hashimoto et al., 2012; Sánchez-Barrena et al., 2020).因此, CBL的磷酸化一方面可增强其与CIPK互作; 另一方面又促进CIPK对下游靶标蛋白的转磷酸作用(Lin et al., 2014; Sanyal et al., 2016). ...
... ).CBL的C端含有1个由23个氨基酸残基组成的基序, 其中有绝对保守的氨基酸残基P、M、L、F、P和F, 因此称作“PFPF基序”(FPSF结构域) (Sanyal et al., 2020).CIPK能够将FPSF结构域中保守的Ser残基磷酸化(Sanyal et al., 2020).海藻(Sargassum sp.)的部分CBL缺少该结构, 以Asp和Glu残基代替(Hashimoto et al., 2012).CIPK介导CBL磷酸化是部分CBL-CIPK信号系统发挥具体功能的必要步骤.例如, AtCIPK24对AtCBL10的磷酸化可激活Na+转运蛋白NHX7 (Almeida et al., 2017), 而CIPK23对CBL1的磷酸化也可激活K+通道AKT1 (Hashimoto et al., 2012; Sánchez-Barrena et al., 2020).因此, CBL的磷酸化一方面可增强其与CIPK互作; 另一方面又促进CIPK对下游靶标蛋白的转磷酸作用(Lin et al., 2014; Sanyal et al., 2016). ...
... ).CIPK能够将FPSF结构域中保守的Ser残基磷酸化(Sanyal et al., 2020).海藻(Sargassum sp.)的部分CBL缺少该结构, 以Asp和Glu残基代替(Hashimoto et al., 2012).CIPK介导CBL磷酸化是部分CBL-CIPK信号系统发挥具体功能的必要步骤.例如, AtCIPK24对AtCBL10的磷酸化可激活Na+转运蛋白NHX7 (Almeida et al., 2017), 而CIPK23对CBL1的磷酸化也可激活K+通道AKT1 (Hashimoto et al., 2012; Sánchez-Barrena et al., 2020).因此, CBL的磷酸化一方面可增强其与CIPK互作; 另一方面又促进CIPK对下游靶标蛋白的转磷酸作用(Lin et al., 2014; Sanyal et al., 2016). ...
... 在拟南芥中, CBL-CIPK信号系统调控许多离子转运蛋白(图1, 图2), 包括介导K+ (KAT1、KAT2、GORK、AKT1、AKT2、HAK5和SPIK)、Na+ (SOS1)、NH4+ (AMT1;1和AMT1;2)、NO3- (NRT1.1、NRT2.4和NRT2.5)、Cl- (SLAC1、SLAH2和SLAH3)和H+ (AHA2和V-ATPase)的通道或转运蛋白(Ho et al., 2009; Léran et al., 2015; Liu et al., 2016; Straub et al., 2017; Ragel et al., 2019; Saito and Uozumi, 2019).CBL-CIPK在响应C/N养分及Mg和Fe的摄取中也发挥重要作用(Sanyal et al., 2020).这种Ca2+依赖性磷酸化调节系统的功能可确保植物的生长, 并使其能耐受各种环境胁迫(Sanyal et al., 2020).由此可见, CBL-CIPK复合物的磷酸化调控对象主要是各种离子通道和转运蛋白. ...
... 依赖性磷酸化调节系统的功能可确保植物的生长, 并使其能耐受各种环境胁迫(Sanyal et al., 2020).由此可见, CBL-CIPK复合物的磷酸化调控对象主要是各种离子通道和转运蛋白. ...
... NO3-和NH4+是植物根系从土壤中吸收的2种N素来源(Tang et al., 2020).Ca2+调控多种NRT/NPF和SLAH转运蛋白进行NO3-摄取和转运(Sanyal et al., 2020).NPF6.3通过二聚体作用使NO3-从低亲和性模式转换为高亲和性模式, 此过程通过CBL1/9- CIPK23对Thr101的磷酸化作用进行控制(Ho et al., 2009; Parker and Newstead, 2014; Sun et al., 2014; Chu et al., 2021).NH4+吸收由AMTs (ammonium transporters)介导(Sanyal et al., 2020).高浓度NH4+会替代K+而产生毒害作用, 因此必须控制其在细胞的非毒性水平(Zheng et al., 2015).AMT的2个成员(AMT1;1和AMT1;2)也可被CBL1-CIPK23复合物抑制(表2; 图2) (Straub et al., 2017).因此, CBL1-CIPK23可能在维持根细胞K+稳态和免受NH4+毒害中起关键作用(Zheng et al., 2015).Weng等(2020)研究表明, 高浓度K+降低水稻根系对NH4+的吸收速率, 增强对H+的外排速率, 从而减轻NH4+对水稻的毒害.此外, CBL1/9-CIPK23与SLAH2和SLAH3互作通过吸收或分泌NO3-参与硝酸盐的转运(表2) (Ho et al., 2009; Léran et al., 2015). ...
... 吸收由AMTs (ammonium transporters)介导(Sanyal et al., 2020).高浓度NH4+会替代K+而产生毒害作用, 因此必须控制其在细胞的非毒性水平(Zheng et al., 2015).AMT的2个成员(AMT1;1和AMT1;2)也可被CBL1-CIPK23复合物抑制(表2; 图2) (Straub et al., 2017).因此, CBL1-CIPK23可能在维持根细胞K+稳态和免受NH4+毒害中起关键作用(Zheng et al., 2015).Weng等(2020)研究表明, 高浓度K+降低水稻根系对NH4+的吸收速率, 增强对H+的外排速率, 从而减轻NH4+对水稻的毒害.此外, CBL1/9-CIPK23与SLAH2和SLAH3互作通过吸收或分泌NO3-参与硝酸盐的转运(表2) (Ho et al., 2009; Léran et al., 2015). ...
... CIPK的C端存在1个PPI结构域(Guo et al., 2001).在结构分析中, CBL相互作用域和PPI结构域可能有重叠, CBL和PP2C (protein phosphatase 2C)与CIPK的相互作用可能相反(Sanyal et al., 2020).此结构特征可以提供一种有利的适应机制, 以防止CIPK被CBL激活和被CIPK磷酸酶失活同时进行, 从而为调控下游底物提供一个激酶-磷酸酶的分子开关(Tang et al., 2020). ...
The CBL-CIPK signaling module in plants: a mechanistic perspective 4 2015
... 植物CBLs是与酵母(Saccharomyces cerevisiae)和动物中的CNB (calcineurin B subunit)最相似的一个Ca2+传感器蛋白家族(Cui et al., 2018).CBL感知应激信号中的Ca2+信号, 使植物适应环境变化(Bender et al., 2018; Tang et al., 2020).CBL含有1个典型的Ca2+结合基序, 即4个手性延伸因子(elongation factor hands, EF-hands), 每个EF-hands包含1个能与Ca2+结合的保守α-螺旋-环-α-螺旋结构(Lu et al., 2017).EF-hands排列在固定的空间中, 从EF-hand1到EF-hand4依次有22、25和32个氨基酸的距离(Sánchez-Barrena et al., 2005).环区的特征是具有12个残基DKDGDGKIDFEE的保守序列(Kolukisaoglu et al., 2004).位置1(X)、3(Y)、5(Z)、7(-X)、9(-Y)和12(-Z)中的氨基酸高度保守, 并负责与Ca2+配位(Sanyal et al., 2015).值得注意的是, EF-hand1在X位置和Y位置之间包含2个插入氨基酸残基的位置(Sánchez-Barrena et al., 2013).这个位置氨基酸的变化导致与Ca2+结合的亲和力发生变化(Kolukisaoglu et al., 2004).除了具有Ca2+感应的EF-hands, CBL还具有N端定位基序和C端磷酸化基序(Beckmann et al., 2016).研究表明, 拟南芥(Arabidopsis thaliana) CBL家族4个成员(AtCBL1、-4、-5和-9)具有N-豆蔻酰化基序(Wang et al., 2019).在SOS (salt overly sensitive)信号系统中, CBL4/SOS3的N-豆蔻酰化基序是植物耐盐性的必备条件(Kleist et al., 2014; Saito et al., 2018), 这为Ca2+信号的传递提供了结构基础. ...
... 植物CIPK在功能上类似于酵母SNF1 (sucrose non- fermenting 1)和哺乳动物AMPK (AMP-dependent kinase) (Mo et al., 2018; Niu et al., 2018).CIPK由2个结构域组成, 一个含有磷酸化位点激活环的保守N端激酶催化结构域, 另一个具有NAF/FISL基序和PPI (protein phosphatase interaction)基序高度不同的C端调节域(Sánchez-Barrena et al., 2013; Sanyal et al., 2015).NAF基序以其高度保守的氨基酸Asn (N)、Ala (A)、Phe (F)、Ile (I)、Ser (S)和Leu (L)而得名, 是结合CBL的必需位点, 其对于维持AtCIPK24和AtCBL4互作是必需的, 并且能够连接AtCIPK24的C端调节结构域以覆盖其激活环, 从而使激酶保持在自动抑制状态(Chaves-Sanjuan et al., 2014).CIPK是具有激酶和调节结构域的Ser/Thr蛋白激酶(Tang et al., 2020).显然, 磷酸化是该系统发挥相应功能的主要作用方式.因此, 磷酸化是CIPK调节靶标蛋白以及与CBL互作的优选方式(Sanyal et al., 2020). ...
... CBL和CIPK的生理学功能首次在SOS途径中被发现(Zhu et al., 1998).拟南芥突变体atsos1、atsos2和atsos3在高盐胁迫下产生相同的盐敏感表型.AtSOS3和AtSOS2也分别被称为AtCBL4和AtCIPK24, 而AtSOS3和AtSOS2已被证明协同正向调节质膜Na+/H+转运蛋白AtSOS1的活性, 在高盐胁迫下介导Na+外排(Zhu et al., 1998; Sanyal et al., 2015).随后, 在拟南芥中鉴定出10个CBLs和26个CIPKs (Kolukisaoglu et al., 2004; Yu et al., 2007).随着植物基因组测序的完成, 在不同物种中相继鉴定出数量不等的CBLs和CIPKs家族成员(表1). ...
... CBL-CIPK复合物对下游靶标蛋白的调节主要通过磷酸化方式进行(Sanyal et al., 2020).靶标蛋白主要有通道蛋白、转运蛋白和转录因子等(Sanyal et al., 2015).已有研究表明, 可被CBL-CIPK复合物磷酸化的阳离子(H+、Na+、K+、Mg2+、Fe2+和NH4+)相关蛋白如AHA2、SOS1、AMT1、ATL31、AKT1、HAK5、TPK1、ARF、ERF、SnRK2D和FIT, 而一些阴离子(OH-和NO3-)转运蛋白如CHL1 (NRT1.1)、RBOH-F和RBOH-C也可被CBL-CIPK磷酸化(Ragel et al.,2019; Lara et al., 2020; Sanyal et al., 2020; Dong et al., 2021). ...
Plant stress responses mediated by CBL-CIPK phosphorylation network 6 2016
... 系统发育分析表明, 拟南芥AtCBLs可分为3类.AtCBL10是第1类中唯一的成员, 并且在CBL家族中具有最长的N端(Sánchez-Barrena et al., 2013).共聚焦荧光显微分析表明, AtCBL10定位于质膜和液泡膜(Kim et al., 2007; Batistič et al., 2010).第2类有5个成员(AtCBL1、-4、-5、-8和-9), 它们具有N端很短的结构特点(Sánchez-Barrena et al., 2013); 其中大多数包含用于脂双层修饰的保守MGCXXS/T基序, 可帮助CBLs固定在膜上(Tang et al., 2015; Saito et al., 2018).第3类成员具有相对较长的N端(Sánchez- Barrena et al., 2013), 它们均具有液泡膜靶向序列(tonoplast targeting sequence, TTS) MSQCXDGXK HXCXSXXXCF (除AtCBL7外) (Tang et al., 2012; Kleist et al., 2014).TTS则有助于AtCBL2、-3和-6定位于液泡膜(Batistič et al., 2010; Tang et al., 2012).CBL7和CBL3是串联复制, 但它们的N端结构域明显不同(Kleist et al., 2014; Sanyal et al., 2016).CBL7作为Ca2+感应蛋白, 能够解码细胞核和细胞质的Ca2+信号(Batistič et al., 2010). ...
... 大量研究表明, CBL与CIPK互作并将其磷酸化是一种普遍的机制, 而且磷酸化能够增强CBL-CIPK复合物的相互作用, 进而提高CIPK的活性(Hashimoto et al., 2012; Sanyal et al., 2020).在植物中, CIPK对CBL的磷酸化最初在豌豆(Pisum sativum)中被报道, PsCIPK可对PsCBL的Thr残基进行磷酸化(Mahajan et al., 2006), 随后在多个物种中相继发现了这种现象(Sanyal et al., 2016).CBL的C端含有1个由23个氨基酸残基组成的基序, 其中有绝对保守的氨基酸残基P、M、L、F、P和F, 因此称作“PFPF基序”(FPSF结构域) (Sanyal et al., 2020).CIPK能够将FPSF结构域中保守的Ser残基磷酸化(Sanyal et al., 2020).海藻(Sargassum sp.)的部分CBL缺少该结构, 以Asp和Glu残基代替(Hashimoto et al., 2012).CIPK介导CBL磷酸化是部分CBL-CIPK信号系统发挥具体功能的必要步骤.例如, AtCIPK24对AtCBL10的磷酸化可激活Na+转运蛋白NHX7 (Almeida et al., 2017), 而CIPK23对CBL1的磷酸化也可激活K+通道AKT1 (Hashimoto et al., 2012; Sánchez-Barrena et al., 2020).因此, CBL的磷酸化一方面可增强其与CIPK互作; 另一方面又促进CIPK对下游靶标蛋白的转磷酸作用(Lin et al., 2014; Sanyal et al., 2016). ...
... ; Sanyal et al., 2016). ...
... 低浓度Na+对植物生长有一定的刺激作用, 但高浓度Na+则会引起细胞渗透和离子毒害(Zhu, 2016).拟南芥SOS信号途径是调节植物根耐盐性的重要机制(Zhu et al., 1998; Yin et al., 2020).在盐胁迫下, AtCBL10与AtCIPK24 (SOS2)互作(图1; 表2), 将Na+区域化到液泡, 以保护植物地上部免受盐胁迫伤害(Hu et al., 2015; Plasencia et al., 2021).这与SOS通路的耐盐机制不同(Yang et al., 2019).被激活的SOS1利用H+-ATPase (AHA2)产生的质子梯度能量将Na+转运出细胞(图1) (Yin et al., 2020).CBL10与CIPK8形成的复合物介导SOS1的激活, 表明CIPK8在调节拟南芥地上部响应盐胁迫中起重要作用(Yin et al., 2020), 但其复合物下游靶标蛋白还有待深入鉴定.细胞内较高的K+/Na+比是植物耐盐性的核心指标(Zhang et al., 2020).研究表明, HKT1 (high affinity potassium transporters 1)能够维持细胞内高浓度的K+和低浓度Na+ (Almeida et al., 2017).水稻(Oryza sativa) OsHKT1;1主要在地上部维管组织中表达, 高盐条件下可增强细胞的Na+外排能力(Campbell et al., 2017).此外, 过表达OsHKT1;4的转基因植株木质部中Na+含量降低, 而根中的Na+含量增加, 从而提高对盐胁迫的敏感性(Oda et al., 2018).CBL2/CBL3-CIPK21也参与植物的耐盐性, 该复合物主要在液泡中发挥作用, 但其下游靶点仍有待确定(Sanyal et al., 2016). ...
... 已有研究表明, 在ABA信号通路中起作用的转录因子也会被CIPK磷酸化(Sanyal et al., 2016; Zhao et al., 2019).最早的报道是ERF7 (ethylene responsive factor 7)被CIPK15磷酸化(Song et al., 2005).磷酸化后的ERF7可能通过2种机制发挥作用: 一是它们结合含有GCC盒的基因并抑制其转录; 二是ERF7可以将AtSin3和HDA19之间的阻遏复合物靶向移至相关基因的启动子, 以进一步抑制基因转录(Song et al., 2005).与之相反, CIPK26介导的ABI5磷酸化是正向调节因子, 可稳定ABI5并启动ABA的下游反应(Lyzenga et al., 2017).KEG (keep on going)使ABI5和CIPK26受到26S蛋白酶的降解, 从而抵消ABA的作用(Lyzenga et al., 2017).此外, PKS5/CIPK11使ABI5在Ser42处磷酸化并激活, 从而调节基因表达(Zhou et al., 2015).因此, CIPK26和PKS5/CIPK11可能在同一途径中发挥相似作用(Zhou et al., 2015).CIPK3与ABR1 (ABA-repressor 1)相互作用并使其磷酸化, 从而调节种子萌发过程中的ABA应答反应(Sanyal et al., 2017; Wang et al., 2018). ...
Reactive oxygen species in the regulation of stomatal movements 1 2016
... 植物在生长过程中经常遇到干旱、盐碱、极端温度、缺K+以及病虫害等多种逆境胁迫(Peck and Mittler, 2020).植物无法逃避逆境, 只能被动应对.因此, 植物在长期进化过程中演化出复杂的信号转导机制, 感知并传递外界变化的信号, 使其对逆境及时做出应激反应(Manik et al., 2015).在植物体内, 许多信号转导途径和调控网络相互作用, 响应特定的生物或非生物胁迫.植物对逆境的响应与激酶、磷酸酶、激素、Ca2+、ROS (reactive oxygen species)以及其它调节蛋白、化合物和小分子相互作用交织在一起, 形成复杂的调控网络(Steinhorst and Kudla, 2013; Sierla et al., 2016; Kudla et al., 2018; Zhang et al., 2018a). ...
Role of an Arabidopsis AP2/EREBP- type transcriptional repressor in abscisic acid and drought stress responses 2 2005
... 已有研究表明, 在ABA信号通路中起作用的转录因子也会被CIPK磷酸化(Sanyal et al., 2016; Zhao et al., 2019).最早的报道是ERF7 (ethylene responsive factor 7)被CIPK15磷酸化(Song et al., 2005).磷酸化后的ERF7可能通过2种机制发挥作用: 一是它们结合含有GCC盒的基因并抑制其转录; 二是ERF7可以将AtSin3和HDA19之间的阻遏复合物靶向移至相关基因的启动子, 以进一步抑制基因转录(Song et al., 2005).与之相反, CIPK26介导的ABI5磷酸化是正向调节因子, 可稳定ABI5并启动ABA的下游反应(Lyzenga et al., 2017).KEG (keep on going)使ABI5和CIPK26受到26S蛋白酶的降解, 从而抵消ABA的作用(Lyzenga et al., 2017).此外, PKS5/CIPK11使ABI5在Ser42处磷酸化并激活, 从而调节基因表达(Zhou et al., 2015).因此, CIPK26和PKS5/CIPK11可能在同一途径中发挥相似作用(Zhou et al., 2015).CIPK3与ABR1 (ABA-repressor 1)相互作用并使其磷酸化, 从而调节种子萌发过程中的ABA应答反应(Sanyal et al., 2017; Wang et al., 2018). ...
... ).磷酸化后的ERF7可能通过2种机制发挥作用: 一是它们结合含有GCC盒的基因并抑制其转录; 二是ERF7可以将AtSin3和HDA19之间的阻遏复合物靶向移至相关基因的启动子, 以进一步抑制基因转录(Song et al., 2005).与之相反, CIPK26介导的ABI5磷酸化是正向调节因子, 可稳定ABI5并启动ABA的下游反应(Lyzenga et al., 2017).KEG (keep on going)使ABI5和CIPK26受到26S蛋白酶的降解, 从而抵消ABA的作用(Lyzenga et al., 2017).此外, PKS5/CIPK11使ABI5在Ser42处磷酸化并激活, 从而调节基因表达(Zhou et al., 2015).因此, CIPK26和PKS5/CIPK11可能在同一途径中发挥相似作用(Zhou et al., 2015).CIPK3与ABR1 (ABA-repressor 1)相互作用并使其磷酸化, 从而调节种子萌发过程中的ABA应答反应(Sanyal et al., 2017; Wang et al., 2018). ...
A tonoplast-associated calcium- signaling module dampens ABA signaling during stomatal movement 2 2018
... 目前, CBL-CIPK复合物对植物激素调控的研究主要集中在ABA信号通路.研究发现, CBL9-CIPK3在ABA信号通路中起负向调控作用(Pandey et al., 2008).拟南芥CBL9-CIPK3复合物通过将转录因子ABI1磷酸化, 形成CBL9-CIPK3-ABR1途径(表2), 从而介导种子萌发和ABA依赖的生理生化反应过程(Sanyal et al., 2017).在小麦(Triticum aestivum)中, TaCBL1- TaCIPK23介导ABA信号途径, 并在干旱胁迫应答中起重要作用(Cui et al., 2018).此外, 拟南芥CBL2和CBL3与CIPK9和CIPK27也参与ABA信号通路的负向调控(Song et al., 2018).OsCBL2可正向调控赤霉素的合成(Hwang et al., 2005), 而AtCIPK6参与调控生长素的运输(Tripathi et al., 2009).上述结果表明, CBL-CIPK复合物广泛参与多种植物激素的调控. ...
Emerging concepts of potassium homeostasis in plants 1 2020
... 植物通过根系从土壤中吸收必需的离子并分配在整个植物组织中(Köster et al., 2019; Srivastava et al., 2020).大量研究表明, 无机离子在维持植物生长中扮演多种重要角色.K+、NO3-和Cl-可维持细胞渗透压, 从而控制细胞膨压, 这对于细胞扩增、气孔运动和花粉管生长至关重要(Saito and Uozumi, 2019).NO3-、NH4+、SO42-和PO43-等被细胞代谢从而产生各种蛋白质和有机化合物(López-Arredondo et al., 2013, 2014). ...
Calcium and reactive oxygen species rule the waves of signaling 1 2013
... 植物在生长过程中经常遇到干旱、盐碱、极端温度、缺K+以及病虫害等多种逆境胁迫(Peck and Mittler, 2020).植物无法逃避逆境, 只能被动应对.因此, 植物在长期进化过程中演化出复杂的信号转导机制, 感知并传递外界变化的信号, 使其对逆境及时做出应激反应(Manik et al., 2015).在植物体内, 许多信号转导途径和调控网络相互作用, 响应特定的生物或非生物胁迫.植物对逆境的响应与激酶、磷酸酶、激素、Ca2+、ROS (reactive oxygen species)以及其它调节蛋白、化合物和小分子相互作用交织在一起, 形成复杂的调控网络(Steinhorst and Kudla, 2013; Sierla et al., 2016; Kudla et al., 2018; Zhang et al., 2018a). ...
The kinase CIPK23 inhibits ammonium transport in Arabidopsis thaliana 3 2017
... 在拟南芥中, CBL-CIPK信号系统调控许多离子转运蛋白(图1, 图2), 包括介导K+ (KAT1、KAT2、GORK、AKT1、AKT2、HAK5和SPIK)、Na+ (SOS1)、NH4+ (AMT1;1和AMT1;2)、NO3- (NRT1.1、NRT2.4和NRT2.5)、Cl- (SLAC1、SLAH2和SLAH3)和H+ (AHA2和V-ATPase)的通道或转运蛋白(Ho et al., 2009; Léran et al., 2015; Liu et al., 2016; Straub et al., 2017; Ragel et al., 2019; Saito and Uozumi, 2019).CBL-CIPK在响应C/N养分及Mg和Fe的摄取中也发挥重要作用(Sanyal et al., 2020).这种Ca2+依赖性磷酸化调节系统的功能可确保植物的生长, 并使其能耐受各种环境胁迫(Sanyal et al., 2020).由此可见, CBL-CIPK复合物的磷酸化调控对象主要是各种离子通道和转运蛋白. ...
... NO3-和NH4+是植物根系从土壤中吸收的2种N素来源(Tang et al., 2020).Ca2+调控多种NRT/NPF和SLAH转运蛋白进行NO3-摄取和转运(Sanyal et al., 2020).NPF6.3通过二聚体作用使NO3-从低亲和性模式转换为高亲和性模式, 此过程通过CBL1/9- CIPK23对Thr101的磷酸化作用进行控制(Ho et al., 2009; Parker and Newstead, 2014; Sun et al., 2014; Chu et al., 2021).NH4+吸收由AMTs (ammonium transporters)介导(Sanyal et al., 2020).高浓度NH4+会替代K+而产生毒害作用, 因此必须控制其在细胞的非毒性水平(Zheng et al., 2015).AMT的2个成员(AMT1;1和AMT1;2)也可被CBL1-CIPK23复合物抑制(表2; 图2) (Straub et al., 2017).因此, CBL1-CIPK23可能在维持根细胞K+稳态和免受NH4+毒害中起关键作用(Zheng et al., 2015).Weng等(2020)研究表明, 高浓度K+降低水稻根系对NH4+的吸收速率, 增强对H+的外排速率, 从而减轻NH4+对水稻的毒害.此外, CBL1/9-CIPK23与SLAH2和SLAH3互作通过吸收或分泌NO3-参与硝酸盐的转运(表2) (Ho et al., 2009; Léran et al., 2015). ...
New insights into the evolution and functional divergence of the CIPK gene family in Saccharum 1 2020
... 在这些必需的离子中, Ca2+浓度通常相对较低,在细胞质中保持在0.1 mmol·L-1左右(Hepler, 2005).然而, 当植物遭遇干旱、盐碱、病原体侵害或营养缺乏等逆境时, Ca2+通过质膜离子通道流入细胞中或从细胞器中释放, 使胞质Ca2+浓度迅速升高(Zhu, 2016; Manishankar et al., 2018; Toyota et al., 2018).这导致Ca2+结合的感应蛋白构象发生变化, 随之与下游效应蛋白以Ca2+依赖方式相互作用并调控其功能(Tang et al., 2020).胞内Ca2+与CBL在其EF-hands处结合, 而CIPK的C端NAF/FISH结构域与CBL互作(Guo et al., 2001; Kolukisaoglu et al., 2004; Su et al., 2020).CBL与CIPK结合使得CIPK的C端自抑制结构域释放N端激酶结构域, 从而增强CIPK活性(Zhou et al., 2016; Yin et al., 2020). ...
Crystal structure of the plant dual-affinity nitrate transporter NRT1.1 1 2014
... NO3-和NH4+是植物根系从土壤中吸收的2种N素来源(Tang et al., 2020).Ca2+调控多种NRT/NPF和SLAH转运蛋白进行NO3-摄取和转运(Sanyal et al., 2020).NPF6.3通过二聚体作用使NO3-从低亲和性模式转换为高亲和性模式, 此过程通过CBL1/9- CIPK23对Thr101的磷酸化作用进行控制(Ho et al., 2009; Parker and Newstead, 2014; Sun et al., 2014; Chu et al., 2021).NH4+吸收由AMTs (ammonium transporters)介导(Sanyal et al., 2020).高浓度NH4+会替代K+而产生毒害作用, 因此必须控制其在细胞的非毒性水平(Zheng et al., 2015).AMT的2个成员(AMT1;1和AMT1;2)也可被CBL1-CIPK23复合物抑制(表2; 图2) (Straub et al., 2017).因此, CBL1-CIPK23可能在维持根细胞K+稳态和免受NH4+毒害中起关键作用(Zheng et al., 2015).Weng等(2020)研究表明, 高浓度K+降低水稻根系对NH4+的吸收速率, 增强对H+的外排速率, 从而减轻NH4+对水稻的毒害.此外, CBL1/9-CIPK23与SLAH2和SLAH3互作通过吸收或分泌NO3-参与硝酸盐的转运(表2) (Ho et al., 2009; Léran et al., 2015). ...
Identification and comprehensive analyses of the CBL and CIPK gene families in wheat (Triticum aestivum L.) 2 2015
... 在细胞内, Ca2+被认为是无处不在、无时不有的功能性调节者, 对许多生理生化反应和发育过程具有重要调控作用(Yin et al., 2017; Köster et al., 2019).Ca2+信号通过Ca2+感应蛋白解码并传递, 进而引起细胞内各种信号系统对环境变化的响应(张和臣等, 2007; Kudla et al., 2018).在植物中, CBLs (Calcineurin B-like proteins)也称SCaBPs (SOS3-like calcium binding proteins), 是一类独特的Ca2+感应蛋白, 其通过激活CIPKs (CBL-interacting protein kinases)/PKS (protein kinase S)解码Ca2+信号(Sánchez-Barrena et al., 2005; 沈金秋等, 2014; Tang et al., 2020).CBLs和CIPKs互作构成一个复杂的信号网络, 在植物对干旱、低温和盐碱等逆境的应答调控中起核心作用(张和臣等, 2007; Sun et al., 2015; Ma et al., 2017; Xi et al., 2017).CBL家族一个成员可以与CIPK家族一个或多个成员相互作用, 建立不同信号级联, 从而形成精准的CBL-CIPK调控网络(Ma et al., 2020).自CBL-CIPK信号系统被发现以来, 学术界对不同途径的CBL-CIPK复合物的功能、结构、基因表达和调控进行了广泛研究(Weinl and Kudla, 2009; Aslam et al., 2019).本文综述了CBLs和CIPKs的结构、分类及其响应各种非生物胁迫的调控机制, 并展望了未来的研究方向, 以期为作物抗逆性遗传改良提供思路. ...
... The CBL and CIPK families in different species Table 1
物种
CBLs数量
CIPKs数量
参考文献
拟南芥(Arabidopsis thaliana)
10
26
Kolukisaoglu et al., 2004
油菜(Brassica napus)
7
23
Zhang et al., 2014a
杨树(Populus trichocarpa)
10
25
Weinl and Kudla, 2009
辣椒(Capsicum annuum)
9
26
Ma et al., 2019
水稻(Oryza sativa)
10
31
Weinl and Kudla, 2009; Piao et al., 2010
茶(Camellia sinensis)
7
18
Liu et al., 2019
小麦(Triticum aestivum)
24
79
Sun et al., 2015; Liu et al., 2018
芜菁(B. rapa var. rapa)
19
51
Yin et al., 2017
木薯(Manihot esculenta)
8
26
Hu et al., 2015; Mo et al., 2018
茄子(Solanum melongena)
5
15
Li et al., 2016
菠萝(Ananas comosus)
8
21
Aslam et al., 2019
葡萄(Vitis vinifera)
8
20
Xi et al., 2017
玉米(Zea mays)
8
43
Chen et al., 2011
甜菜(Beta vulgaris)
7
20
未发表数据
2 植物CBL-CIPK系统与磷酸化 CBL-CIPK复合物对下游靶标蛋白的调节主要通过磷酸化方式进行(Sanyal et al., 2020).靶标蛋白主要有通道蛋白、转运蛋白和转录因子等(Sanyal et al., 2015).已有研究表明, 可被CBL-CIPK复合物磷酸化的阳离子(H+、Na+、K+、Mg2+、Fe2+和NH4+)相关蛋白如AHA2、SOS1、AMT1、ATL31、AKT1、HAK5、TPK1、ARF、ERF、SnRK2D和FIT, 而一些阴离子(OH-和NO3-)转运蛋白如CHL1 (NRT1.1)、RBOH-F和RBOH-C也可被CBL-CIPK磷酸化(Ragel et al.,2019; Lara et al., 2020; Sanyal et al., 2020; Dong et al., 2021). ...
Tonoplast calcium sensors CBL2 and CBL3 control plant growth and ion homeostasis through regulating V-ATPase activity in Arabidopsis 4 2012
... 系统发育分析表明, 拟南芥AtCBLs可分为3类.AtCBL10是第1类中唯一的成员, 并且在CBL家族中具有最长的N端(Sánchez-Barrena et al., 2013).共聚焦荧光显微分析表明, AtCBL10定位于质膜和液泡膜(Kim et al., 2007; Batistič et al., 2010).第2类有5个成员(AtCBL1、-4、-5、-8和-9), 它们具有N端很短的结构特点(Sánchez-Barrena et al., 2013); 其中大多数包含用于脂双层修饰的保守MGCXXS/T基序, 可帮助CBLs固定在膜上(Tang et al., 2015; Saito et al., 2018).第3类成员具有相对较长的N端(Sánchez- Barrena et al., 2013), 它们均具有液泡膜靶向序列(tonoplast targeting sequence, TTS) MSQCXDGXK HXCXSXXXCF (除AtCBL7外) (Tang et al., 2012; Kleist et al., 2014).TTS则有助于AtCBL2、-3和-6定位于液泡膜(Batistič et al., 2010; Tang et al., 2012).CBL7和CBL3是串联复制, 但它们的N端结构域明显不同(Kleist et al., 2014; Sanyal et al., 2016).CBL7作为Ca2+感应蛋白, 能够解码细胞核和细胞质的Ca2+信号(Batistič et al., 2010). ...
... ; Tang et al., 2012).CBL7和CBL3是串联复制, 但它们的N端结构域明显不同(Kleist et al., 2014; Sanyal et al., 2016).CBL7作为Ca2+感应蛋白, 能够解码细胞核和细胞质的Ca2+信号(Batistič et al., 2010). ...
... CBL1/9-CIPK23与SLAC1或SLAC3相互作用(表2; 图2), 参与ABA (abscisic acid)对气孔大小的调节, 从而导致气孔关闭(Maierhofer et al., 2014a, 2014b).研究表明, H+-ATPase也与相对应的CBL- CIPK互作而发挥作用(图1; 表2).质膜H+-ATPase受CBL2-CIPK11调控(Saito and Uozumi, 2020), 而液泡膜H+-ATPase则受CBL2/3-CIPK3/9/23/26调控(Tang et al., 2012, 2015). ...
The CBL-CIPK calcium signaling network: unified paradigm from 20 years of discoveries 8 2020
... 在细胞内, Ca2+被认为是无处不在、无时不有的功能性调节者, 对许多生理生化反应和发育过程具有重要调控作用(Yin et al., 2017; Köster et al., 2019).Ca2+信号通过Ca2+感应蛋白解码并传递, 进而引起细胞内各种信号系统对环境变化的响应(张和臣等, 2007; Kudla et al., 2018).在植物中, CBLs (Calcineurin B-like proteins)也称SCaBPs (SOS3-like calcium binding proteins), 是一类独特的Ca2+感应蛋白, 其通过激活CIPKs (CBL-interacting protein kinases)/PKS (protein kinase S)解码Ca2+信号(Sánchez-Barrena et al., 2005; 沈金秋等, 2014; Tang et al., 2020).CBLs和CIPKs互作构成一个复杂的信号网络, 在植物对干旱、低温和盐碱等逆境的应答调控中起核心作用(张和臣等, 2007; Sun et al., 2015; Ma et al., 2017; Xi et al., 2017).CBL家族一个成员可以与CIPK家族一个或多个成员相互作用, 建立不同信号级联, 从而形成精准的CBL-CIPK调控网络(Ma et al., 2020).自CBL-CIPK信号系统被发现以来, 学术界对不同途径的CBL-CIPK复合物的功能、结构、基因表达和调控进行了广泛研究(Weinl and Kudla, 2009; Aslam et al., 2019).本文综述了CBLs和CIPKs的结构、分类及其响应各种非生物胁迫的调控机制, 并展望了未来的研究方向, 以期为作物抗逆性遗传改良提供思路. ...
... 植物CBLs是与酵母(Saccharomyces cerevisiae)和动物中的CNB (calcineurin B subunit)最相似的一个Ca2+传感器蛋白家族(Cui et al., 2018).CBL感知应激信号中的Ca2+信号, 使植物适应环境变化(Bender et al., 2018; Tang et al., 2020).CBL含有1个典型的Ca2+结合基序, 即4个手性延伸因子(elongation factor hands, EF-hands), 每个EF-hands包含1个能与Ca2+结合的保守α-螺旋-环-α-螺旋结构(Lu et al., 2017).EF-hands排列在固定的空间中, 从EF-hand1到EF-hand4依次有22、25和32个氨基酸的距离(Sánchez-Barrena et al., 2005).环区的特征是具有12个残基DKDGDGKIDFEE的保守序列(Kolukisaoglu et al., 2004).位置1(X)、3(Y)、5(Z)、7(-X)、9(-Y)和12(-Z)中的氨基酸高度保守, 并负责与Ca2+配位(Sanyal et al., 2015).值得注意的是, EF-hand1在X位置和Y位置之间包含2个插入氨基酸残基的位置(Sánchez-Barrena et al., 2013).这个位置氨基酸的变化导致与Ca2+结合的亲和力发生变化(Kolukisaoglu et al., 2004).除了具有Ca2+感应的EF-hands, CBL还具有N端定位基序和C端磷酸化基序(Beckmann et al., 2016).研究表明, 拟南芥(Arabidopsis thaliana) CBL家族4个成员(AtCBL1、-4、-5和-9)具有N-豆蔻酰化基序(Wang et al., 2019).在SOS (salt overly sensitive)信号系统中, CBL4/SOS3的N-豆蔻酰化基序是植物耐盐性的必备条件(Kleist et al., 2014; Saito et al., 2018), 这为Ca2+信号的传递提供了结构基础. ...
... 植物CIPK在功能上类似于酵母SNF1 (sucrose non- fermenting 1)和哺乳动物AMPK (AMP-dependent kinase) (Mo et al., 2018; Niu et al., 2018).CIPK由2个结构域组成, 一个含有磷酸化位点激活环的保守N端激酶催化结构域, 另一个具有NAF/FISL基序和PPI (protein phosphatase interaction)基序高度不同的C端调节域(Sánchez-Barrena et al., 2013; Sanyal et al., 2015).NAF基序以其高度保守的氨基酸Asn (N)、Ala (A)、Phe (F)、Ile (I)、Ser (S)和Leu (L)而得名, 是结合CBL的必需位点, 其对于维持AtCIPK24和AtCBL4互作是必需的, 并且能够连接AtCIPK24的C端调节结构域以覆盖其激活环, 从而使激酶保持在自动抑制状态(Chaves-Sanjuan et al., 2014).CIPK是具有激酶和调节结构域的Ser/Thr蛋白激酶(Tang et al., 2020).显然, 磷酸化是该系统发挥相应功能的主要作用方式.因此, 磷酸化是CIPK调节靶标蛋白以及与CBL互作的优选方式(Sanyal et al., 2020). ...
... 在这些必需的离子中, Ca2+浓度通常相对较低,在细胞质中保持在0.1 mmol·L-1左右(Hepler, 2005).然而, 当植物遭遇干旱、盐碱、病原体侵害或营养缺乏等逆境时, Ca2+通过质膜离子通道流入细胞中或从细胞器中释放, 使胞质Ca2+浓度迅速升高(Zhu, 2016; Manishankar et al., 2018; Toyota et al., 2018).这导致Ca2+结合的感应蛋白构象发生变化, 随之与下游效应蛋白以Ca2+依赖方式相互作用并调控其功能(Tang et al., 2020).胞内Ca2+与CBL在其EF-hands处结合, 而CIPK的C端NAF/FISH结构域与CBL互作(Guo et al., 2001; Kolukisaoglu et al., 2004; Su et al., 2020).CBL与CIPK结合使得CIPK的C端自抑制结构域释放N端激酶结构域, 从而增强CIPK活性(Zhou et al., 2016; Yin et al., 2020). ...
... K+是植物细胞中最丰富的离子, 在调节细胞渗透压、膜电位和细胞内pH值方面发挥至关重要的作用(伍国强等, 2017; Almeida et al., 2017; Ragel et al., 2019).K+亏缺会导致拟南芥根中Ca2+迅速增加(Behera et al., 2017).维持根系K+吸收的主要系统有K+通道AKT1和高亲和性K+转运蛋白HAK5 (图1; 表2) (Alemán et al., 2011).CBL1/9与CIPK23结合并在质膜上聚集, CIPK23激活后将AKT1和HAK5磷酸化并激活, 以促进K+的吸收和转运(图1; 表2) (Lara et al., 2020).此外, CBL4-CIPK6复合物作用于AKT2, 可应答K+的外排(图1) (Saito and Uozumi, 2020).液泡膜CBL2/3-CIPK3/9/23/26复合物激活TPKs (two- pore K+ channels), 以维持细胞内K+稳态(Tang et al., 2020; Dong et al., 2021). ...
... NO3-和NH4+是植物根系从土壤中吸收的2种N素来源(Tang et al., 2020).Ca2+调控多种NRT/NPF和SLAH转运蛋白进行NO3-摄取和转运(Sanyal et al., 2020).NPF6.3通过二聚体作用使NO3-从低亲和性模式转换为高亲和性模式, 此过程通过CBL1/9- CIPK23对Thr101的磷酸化作用进行控制(Ho et al., 2009; Parker and Newstead, 2014; Sun et al., 2014; Chu et al., 2021).NH4+吸收由AMTs (ammonium transporters)介导(Sanyal et al., 2020).高浓度NH4+会替代K+而产生毒害作用, 因此必须控制其在细胞的非毒性水平(Zheng et al., 2015).AMT的2个成员(AMT1;1和AMT1;2)也可被CBL1-CIPK23复合物抑制(表2; 图2) (Straub et al., 2017).因此, CBL1-CIPK23可能在维持根细胞K+稳态和免受NH4+毒害中起关键作用(Zheng et al., 2015).Weng等(2020)研究表明, 高浓度K+降低水稻根系对NH4+的吸收速率, 增强对H+的外排速率, 从而减轻NH4+对水稻的毒害.此外, CBL1/9-CIPK23与SLAH2和SLAH3互作通过吸收或分泌NO3-参与硝酸盐的转运(表2) (Ho et al., 2009; Léran et al., 2015). ...
... CIPK的C端存在1个PPI结构域(Guo et al., 2001).在结构分析中, CBL相互作用域和PPI结构域可能有重叠, CBL和PP2C (protein phosphatase 2C)与CIPK的相互作用可能相反(Sanyal et al., 2020).此结构特征可以提供一种有利的适应机制, 以防止CIPK被CBL激活和被CIPK磷酸酶失活同时进行, 从而为调控下游底物提供一个激酶-磷酸酶的分子开关(Tang et al., 2020). ...
... 系统发育分析表明, 拟南芥AtCBLs可分为3类.AtCBL10是第1类中唯一的成员, 并且在CBL家族中具有最长的N端(Sánchez-Barrena et al., 2013).共聚焦荧光显微分析表明, AtCBL10定位于质膜和液泡膜(Kim et al., 2007; Batistič et al., 2010).第2类有5个成员(AtCBL1、-4、-5、-8和-9), 它们具有N端很短的结构特点(Sánchez-Barrena et al., 2013); 其中大多数包含用于脂双层修饰的保守MGCXXS/T基序, 可帮助CBLs固定在膜上(Tang et al., 2015; Saito et al., 2018).第3类成员具有相对较长的N端(Sánchez- Barrena et al., 2013), 它们均具有液泡膜靶向序列(tonoplast targeting sequence, TTS) MSQCXDGXK HXCXSXXXCF (除AtCBL7外) (Tang et al., 2012; Kleist et al., 2014).TTS则有助于AtCBL2、-3和-6定位于液泡膜(Batistič et al., 2010; Tang et al., 2012).CBL7和CBL3是串联复制, 但它们的N端结构域明显不同(Kleist et al., 2014; Sanyal et al., 2016).CBL7作为Ca2+感应蛋白, 能够解码细胞核和细胞质的Ca2+信号(Batistič et al., 2010). ...
... 在这些必需的离子中, Ca2+浓度通常相对较低,在细胞质中保持在0.1 mmol·L-1左右(Hepler, 2005).然而, 当植物遭遇干旱、盐碱、病原体侵害或营养缺乏等逆境时, Ca2+通过质膜离子通道流入细胞中或从细胞器中释放, 使胞质Ca2+浓度迅速升高(Zhu, 2016; Manishankar et al., 2018; Toyota et al., 2018).这导致Ca2+结合的感应蛋白构象发生变化, 随之与下游效应蛋白以Ca2+依赖方式相互作用并调控其功能(Tang et al., 2020).胞内Ca2+与CBL在其EF-hands处结合, 而CIPK的C端NAF/FISH结构域与CBL互作(Guo et al., 2001; Kolukisaoglu et al., 2004; Su et al., 2020).CBL与CIPK结合使得CIPK的C端自抑制结构域释放N端激酶结构域, 从而增强CIPK活性(Zhou et al., 2016; Yin et al., 2020). ...
Role of CIPK6 in root growth and auxin transport 1 2009
... 目前, CBL-CIPK复合物对植物激素调控的研究主要集中在ABA信号通路.研究发现, CBL9-CIPK3在ABA信号通路中起负向调控作用(Pandey et al., 2008).拟南芥CBL9-CIPK3复合物通过将转录因子ABI1磷酸化, 形成CBL9-CIPK3-ABR1途径(表2), 从而介导种子萌发和ABA依赖的生理生化反应过程(Sanyal et al., 2017).在小麦(Triticum aestivum)中, TaCBL1- TaCIPK23介导ABA信号途径, 并在干旱胁迫应答中起重要作用(Cui et al., 2018).此外, 拟南芥CBL2和CBL3与CIPK9和CIPK27也参与ABA信号通路的负向调控(Song et al., 2018).OsCBL2可正向调控赤霉素的合成(Hwang et al., 2005), 而AtCIPK6参与调控生长素的运输(Tripathi et al., 2009).上述结果表明, CBL-CIPK复合物广泛参与多种植物激素的调控. ...
Calcium-mediation of jasmonate biosynthesis and signaling in plants 1 2019
... 植物CBLs是与酵母(Saccharomyces cerevisiae)和动物中的CNB (calcineurin B subunit)最相似的一个Ca2+传感器蛋白家族(Cui et al., 2018).CBL感知应激信号中的Ca2+信号, 使植物适应环境变化(Bender et al., 2018; Tang et al., 2020).CBL含有1个典型的Ca2+结合基序, 即4个手性延伸因子(elongation factor hands, EF-hands), 每个EF-hands包含1个能与Ca2+结合的保守α-螺旋-环-α-螺旋结构(Lu et al., 2017).EF-hands排列在固定的空间中, 从EF-hand1到EF-hand4依次有22、25和32个氨基酸的距离(Sánchez-Barrena et al., 2005).环区的特征是具有12个残基DKDGDGKIDFEE的保守序列(Kolukisaoglu et al., 2004).位置1(X)、3(Y)、5(Z)、7(-X)、9(-Y)和12(-Z)中的氨基酸高度保守, 并负责与Ca2+配位(Sanyal et al., 2015).值得注意的是, EF-hand1在X位置和Y位置之间包含2个插入氨基酸残基的位置(Sánchez-Barrena et al., 2013).这个位置氨基酸的变化导致与Ca2+结合的亲和力发生变化(Kolukisaoglu et al., 2004).除了具有Ca2+感应的EF-hands, CBL还具有N端定位基序和C端磷酸化基序(Beckmann et al., 2016).研究表明, 拟南芥(Arabidopsis thaliana) CBL家族4个成员(AtCBL1、-4、-5和-9)具有N-豆蔻酰化基序(Wang et al., 2019).在SOS (salt overly sensitive)信号系统中, CBL4/SOS3的N-豆蔻酰化基序是植物耐盐性的必备条件(Kleist et al., 2014; Saito et al., 2018), 这为Ca2+信号的传递提供了结构基础. ...
A CBL-interacting protein kinase TaCIPK27 confers drought tolerance and exogenous ABA sensitivity in transgenic Arabidopsis 2 2018
... 已有研究表明, 在ABA信号通路中起作用的转录因子也会被CIPK磷酸化(Sanyal et al., 2016; Zhao et al., 2019).最早的报道是ERF7 (ethylene responsive factor 7)被CIPK15磷酸化(Song et al., 2005).磷酸化后的ERF7可能通过2种机制发挥作用: 一是它们结合含有GCC盒的基因并抑制其转录; 二是ERF7可以将AtSin3和HDA19之间的阻遏复合物靶向移至相关基因的启动子, 以进一步抑制基因转录(Song et al., 2005).与之相反, CIPK26介导的ABI5磷酸化是正向调节因子, 可稳定ABI5并启动ABA的下游反应(Lyzenga et al., 2017).KEG (keep on going)使ABI5和CIPK26受到26S蛋白酶的降解, 从而抵消ABA的作用(Lyzenga et al., 2017).此外, PKS5/CIPK11使ABI5在Ser42处磷酸化并激活, 从而调节基因表达(Zhou et al., 2015).因此, CIPK26和PKS5/CIPK11可能在同一途径中发挥相似作用(Zhou et al., 2015).CIPK3与ABR1 (ABA-repressor 1)相互作用并使其磷酸化, 从而调节种子萌发过程中的ABA应答反应(Sanyal et al., 2017; Wang et al., 2018). ...
The CBL-CIPK Ca2+-decoding signaling network: function and perspectives 3 2009
... 在细胞内, Ca2+被认为是无处不在、无时不有的功能性调节者, 对许多生理生化反应和发育过程具有重要调控作用(Yin et al., 2017; Köster et al., 2019).Ca2+信号通过Ca2+感应蛋白解码并传递, 进而引起细胞内各种信号系统对环境变化的响应(张和臣等, 2007; Kudla et al., 2018).在植物中, CBLs (Calcineurin B-like proteins)也称SCaBPs (SOS3-like calcium binding proteins), 是一类独特的Ca2+感应蛋白, 其通过激活CIPKs (CBL-interacting protein kinases)/PKS (protein kinase S)解码Ca2+信号(Sánchez-Barrena et al., 2005; 沈金秋等, 2014; Tang et al., 2020).CBLs和CIPKs互作构成一个复杂的信号网络, 在植物对干旱、低温和盐碱等逆境的应答调控中起核心作用(张和臣等, 2007; Sun et al., 2015; Ma et al., 2017; Xi et al., 2017).CBL家族一个成员可以与CIPK家族一个或多个成员相互作用, 建立不同信号级联, 从而形成精准的CBL-CIPK调控网络(Ma et al., 2020).自CBL-CIPK信号系统被发现以来, 学术界对不同途径的CBL-CIPK复合物的功能、结构、基因表达和调控进行了广泛研究(Weinl and Kudla, 2009; Aslam et al., 2019).本文综述了CBLs和CIPKs的结构、分类及其响应各种非生物胁迫的调控机制, 并展望了未来的研究方向, 以期为作物抗逆性遗传改良提供思路. ...
... The CBL and CIPK families in different species Table 1
物种
CBLs数量
CIPKs数量
参考文献
拟南芥(Arabidopsis thaliana)
10
26
Kolukisaoglu et al., 2004
油菜(Brassica napus)
7
23
Zhang et al., 2014a
杨树(Populus trichocarpa)
10
25
Weinl and Kudla, 2009
辣椒(Capsicum annuum)
9
26
Ma et al., 2019
水稻(Oryza sativa)
10
31
Weinl and Kudla, 2009; Piao et al., 2010
茶(Camellia sinensis)
7
18
Liu et al., 2019
小麦(Triticum aestivum)
24
79
Sun et al., 2015; Liu et al., 2018
芜菁(B. rapa var. rapa)
19
51
Yin et al., 2017
木薯(Manihot esculenta)
8
26
Hu et al., 2015; Mo et al., 2018
茄子(Solanum melongena)
5
15
Li et al., 2016
菠萝(Ananas comosus)
8
21
Aslam et al., 2019
葡萄(Vitis vinifera)
8
20
Xi et al., 2017
玉米(Zea mays)
8
43
Chen et al., 2011
甜菜(Beta vulgaris)
7
20
未发表数据
2 植物CBL-CIPK系统与磷酸化 CBL-CIPK复合物对下游靶标蛋白的调节主要通过磷酸化方式进行(Sanyal et al., 2020).靶标蛋白主要有通道蛋白、转运蛋白和转录因子等(Sanyal et al., 2015).已有研究表明, 可被CBL-CIPK复合物磷酸化的阳离子(H+、Na+、K+、Mg2+、Fe2+和NH4+)相关蛋白如AHA2、SOS1、AMT1、ATL31、AKT1、HAK5、TPK1、ARF、ERF、SnRK2D和FIT, 而一些阴离子(OH-和NO3-)转运蛋白如CHL1 (NRT1.1)、RBOH-F和RBOH-C也可被CBL-CIPK磷酸化(Ragel et al.,2019; Lara et al., 2020; Sanyal et al., 2020; Dong et al., 2021). ...
... Weinl and Kudla, 2009; Piao et al., 2010
茶(Camellia sinensis)
7
18
Liu et al., 2019
小麦(Triticum aestivum)
24
79
Sun et al., 2015; Liu et al., 2018
芜菁(B. rapa var. rapa)
19
51
Yin et al., 2017
木薯(Manihot esculenta)
8
26
Hu et al., 2015; Mo et al., 2018
茄子(Solanum melongena)
5
15
Li et al., 2016
菠萝(Ananas comosus)
8
21
Aslam et al., 2019
葡萄(Vitis vinifera)
8
20
Xi et al., 2017
玉米(Zea mays)
8
43
Chen et al., 2011
甜菜(Beta vulgaris)
7
20
未发表数据
2 植物CBL-CIPK系统与磷酸化 CBL-CIPK复合物对下游靶标蛋白的调节主要通过磷酸化方式进行(Sanyal et al., 2020).靶标蛋白主要有通道蛋白、转运蛋白和转录因子等(Sanyal et al., 2015).已有研究表明, 可被CBL-CIPK复合物磷酸化的阳离子(H+、Na+、K+、Mg2+、Fe2+和NH4+)相关蛋白如AHA2、SOS1、AMT1、ATL31、AKT1、HAK5、TPK1、ARF、ERF、SnRK2D和FIT, 而一些阴离子(OH-和NO3-)转运蛋白如CHL1 (NRT1.1)、RBOH-F和RBOH-C也可被CBL-CIPK磷酸化(Ragel et al.,2019; Lara et al., 2020; Sanyal et al., 2020; Dong et al., 2021). ...
Potassium alleviates ammonium toxicity in rice by reducing its uptake through activation of plasma membrane H+-ATPase to enhance proton extrusion 1 2020
... NO3-和NH4+是植物根系从土壤中吸收的2种N素来源(Tang et al., 2020).Ca2+调控多种NRT/NPF和SLAH转运蛋白进行NO3-摄取和转运(Sanyal et al., 2020).NPF6.3通过二聚体作用使NO3-从低亲和性模式转换为高亲和性模式, 此过程通过CBL1/9- CIPK23对Thr101的磷酸化作用进行控制(Ho et al., 2009; Parker and Newstead, 2014; Sun et al., 2014; Chu et al., 2021).NH4+吸收由AMTs (ammonium transporters)介导(Sanyal et al., 2020).高浓度NH4+会替代K+而产生毒害作用, 因此必须控制其在细胞的非毒性水平(Zheng et al., 2015).AMT的2个成员(AMT1;1和AMT1;2)也可被CBL1-CIPK23复合物抑制(表2; 图2) (Straub et al., 2017).因此, CBL1-CIPK23可能在维持根细胞K+稳态和免受NH4+毒害中起关键作用(Zheng et al., 2015).Weng等(2020)研究表明, 高浓度K+降低水稻根系对NH4+的吸收速率, 增强对H+的外排速率, 从而减轻NH4+对水稻的毒害.此外, CBL1/9-CIPK23与SLAH2和SLAH3互作通过吸收或分泌NO3-参与硝酸盐的转运(表2) (Ho et al., 2009; Léran et al., 2015). ...
The CBL and CIPK gene family in grapevine (Vitis vinifera): genome- wide analysis and expression profiles in response to various abiotic stresses 2 2017
... 在细胞内, Ca2+被认为是无处不在、无时不有的功能性调节者, 对许多生理生化反应和发育过程具有重要调控作用(Yin et al., 2017; Köster et al., 2019).Ca2+信号通过Ca2+感应蛋白解码并传递, 进而引起细胞内各种信号系统对环境变化的响应(张和臣等, 2007; Kudla et al., 2018).在植物中, CBLs (Calcineurin B-like proteins)也称SCaBPs (SOS3-like calcium binding proteins), 是一类独特的Ca2+感应蛋白, 其通过激活CIPKs (CBL-interacting protein kinases)/PKS (protein kinase S)解码Ca2+信号(Sánchez-Barrena et al., 2005; 沈金秋等, 2014; Tang et al., 2020).CBLs和CIPKs互作构成一个复杂的信号网络, 在植物对干旱、低温和盐碱等逆境的应答调控中起核心作用(张和臣等, 2007; Sun et al., 2015; Ma et al., 2017; Xi et al., 2017).CBL家族一个成员可以与CIPK家族一个或多个成员相互作用, 建立不同信号级联, 从而形成精准的CBL-CIPK调控网络(Ma et al., 2020).自CBL-CIPK信号系统被发现以来, 学术界对不同途径的CBL-CIPK复合物的功能、结构、基因表达和调控进行了广泛研究(Weinl and Kudla, 2009; Aslam et al., 2019).本文综述了CBLs和CIPKs的结构、分类及其响应各种非生物胁迫的调控机制, 并展望了未来的研究方向, 以期为作物抗逆性遗传改良提供思路. ...
... The CBL and CIPK families in different species Table 1
物种
CBLs数量
CIPKs数量
参考文献
拟南芥(Arabidopsis thaliana)
10
26
Kolukisaoglu et al., 2004
油菜(Brassica napus)
7
23
Zhang et al., 2014a
杨树(Populus trichocarpa)
10
25
Weinl and Kudla, 2009
辣椒(Capsicum annuum)
9
26
Ma et al., 2019
水稻(Oryza sativa)
10
31
Weinl and Kudla, 2009; Piao et al., 2010
茶(Camellia sinensis)
7
18
Liu et al., 2019
小麦(Triticum aestivum)
24
79
Sun et al., 2015; Liu et al., 2018
芜菁(B. rapa var. rapa)
19
51
Yin et al., 2017
木薯(Manihot esculenta)
8
26
Hu et al., 2015; Mo et al., 2018
茄子(Solanum melongena)
5
15
Li et al., 2016
菠萝(Ananas comosus)
8
21
Aslam et al., 2019
葡萄(Vitis vinifera)
8
20
Xi et al., 2017
玉米(Zea mays)
8
43
Chen et al., 2011
甜菜(Beta vulgaris)
7
20
未发表数据
2 植物CBL-CIPK系统与磷酸化 CBL-CIPK复合物对下游靶标蛋白的调节主要通过磷酸化方式进行(Sanyal et al., 2020).靶标蛋白主要有通道蛋白、转运蛋白和转录因子等(Sanyal et al., 2015).已有研究表明, 可被CBL-CIPK复合物磷酸化的阳离子(H+、Na+、K+、Mg2+、Fe2+和NH4+)相关蛋白如AHA2、SOS1、AMT1、ATL31、AKT1、HAK5、TPK1、ARF、ERF、SnRK2D和FIT, 而一些阴离子(OH-和NO3-)转运蛋白如CHL1 (NRT1.1)、RBOH-F和RBOH-C也可被CBL-CIPK磷酸化(Ragel et al.,2019; Lara et al., 2020; Sanyal et al., 2020; Dong et al., 2021). ...
Calcineurin B-like proteins CBL4 and CBL10 mediate two independent salt tolerance pathways in Arabidopsis 1 2019
... 低浓度Na+对植物生长有一定的刺激作用, 但高浓度Na+则会引起细胞渗透和离子毒害(Zhu, 2016).拟南芥SOS信号途径是调节植物根耐盐性的重要机制(Zhu et al., 1998; Yin et al., 2020).在盐胁迫下, AtCBL10与AtCIPK24 (SOS2)互作(图1; 表2), 将Na+区域化到液泡, 以保护植物地上部免受盐胁迫伤害(Hu et al., 2015; Plasencia et al., 2021).这与SOS通路的耐盐机制不同(Yang et al., 2019).被激活的SOS1利用H+-ATPase (AHA2)产生的质子梯度能量将Na+转运出细胞(图1) (Yin et al., 2020).CBL10与CIPK8形成的复合物介导SOS1的激活, 表明CIPK8在调节拟南芥地上部响应盐胁迫中起重要作用(Yin et al., 2020), 但其复合物下游靶标蛋白还有待深入鉴定.细胞内较高的K+/Na+比是植物耐盐性的核心指标(Zhang et al., 2020).研究表明, HKT1 (high affinity potassium transporters 1)能够维持细胞内高浓度的K+和低浓度Na+ (Almeida et al., 2017).水稻(Oryza sativa) OsHKT1;1主要在地上部维管组织中表达, 高盐条件下可增强细胞的Na+外排能力(Campbell et al., 2017).此外, 过表达OsHKT1;4的转基因植株木质部中Na+含量降低, 而根中的Na+含量增加, 从而提高对盐胁迫的敏感性(Oda et al., 2018).CBL2/CBL3-CIPK21也参与植物的耐盐性, 该复合物主要在液泡中发挥作用, 但其下游靶点仍有待确定(Sanyal et al., 2016). ...
Genome-wide identification and functional analysis of the calcineurin B-like protein and calcineurin B-like protein-interacting protein kinase gene families in turnip ( Brassica rapa var. rapa) 2 2017
... 在细胞内, Ca2+被认为是无处不在、无时不有的功能性调节者, 对许多生理生化反应和发育过程具有重要调控作用(Yin et al., 2017; Köster et al., 2019).Ca2+信号通过Ca2+感应蛋白解码并传递, 进而引起细胞内各种信号系统对环境变化的响应(张和臣等, 2007; Kudla et al., 2018).在植物中, CBLs (Calcineurin B-like proteins)也称SCaBPs (SOS3-like calcium binding proteins), 是一类独特的Ca2+感应蛋白, 其通过激活CIPKs (CBL-interacting protein kinases)/PKS (protein kinase S)解码Ca2+信号(Sánchez-Barrena et al., 2005; 沈金秋等, 2014; Tang et al., 2020).CBLs和CIPKs互作构成一个复杂的信号网络, 在植物对干旱、低温和盐碱等逆境的应答调控中起核心作用(张和臣等, 2007; Sun et al., 2015; Ma et al., 2017; Xi et al., 2017).CBL家族一个成员可以与CIPK家族一个或多个成员相互作用, 建立不同信号级联, 从而形成精准的CBL-CIPK调控网络(Ma et al., 2020).自CBL-CIPK信号系统被发现以来, 学术界对不同途径的CBL-CIPK复合物的功能、结构、基因表达和调控进行了广泛研究(Weinl and Kudla, 2009; Aslam et al., 2019).本文综述了CBLs和CIPKs的结构、分类及其响应各种非生物胁迫的调控机制, 并展望了未来的研究方向, 以期为作物抗逆性遗传改良提供思路. ...
... The CBL and CIPK families in different species Table 1
物种
CBLs数量
CIPKs数量
参考文献
拟南芥(Arabidopsis thaliana)
10
26
Kolukisaoglu et al., 2004
油菜(Brassica napus)
7
23
Zhang et al., 2014a
杨树(Populus trichocarpa)
10
25
Weinl and Kudla, 2009
辣椒(Capsicum annuum)
9
26
Ma et al., 2019
水稻(Oryza sativa)
10
31
Weinl and Kudla, 2009; Piao et al., 2010
茶(Camellia sinensis)
7
18
Liu et al., 2019
小麦(Triticum aestivum)
24
79
Sun et al., 2015; Liu et al., 2018
芜菁(B. rapa var. rapa)
19
51
Yin et al., 2017
木薯(Manihot esculenta)
8
26
Hu et al., 2015; Mo et al., 2018
茄子(Solanum melongena)
5
15
Li et al., 2016
菠萝(Ananas comosus)
8
21
Aslam et al., 2019
葡萄(Vitis vinifera)
8
20
Xi et al., 2017
玉米(Zea mays)
8
43
Chen et al., 2011
甜菜(Beta vulgaris)
7
20
未发表数据
2 植物CBL-CIPK系统与磷酸化 CBL-CIPK复合物对下游靶标蛋白的调节主要通过磷酸化方式进行(Sanyal et al., 2020).靶标蛋白主要有通道蛋白、转运蛋白和转录因子等(Sanyal et al., 2015).已有研究表明, 可被CBL-CIPK复合物磷酸化的阳离子(H+、Na+、K+、Mg2+、Fe2+和NH4+)相关蛋白如AHA2、SOS1、AMT1、ATL31、AKT1、HAK5、TPK1、ARF、ERF、SnRK2D和FIT, 而一些阴离子(OH-和NO3-)转运蛋白如CHL1 (NRT1.1)、RBOH-F和RBOH-C也可被CBL-CIPK磷酸化(Ragel et al.,2019; Lara et al., 2020; Sanyal et al., 2020; Dong et al., 2021). ...
The protein kinase complex CBL10-CIPK8-SOS1 functions in Arabidopsis to regulate salt tolerance 6 2020
... 在这些必需的离子中, Ca2+浓度通常相对较低,在细胞质中保持在0.1 mmol·L-1左右(Hepler, 2005).然而, 当植物遭遇干旱、盐碱、病原体侵害或营养缺乏等逆境时, Ca2+通过质膜离子通道流入细胞中或从细胞器中释放, 使胞质Ca2+浓度迅速升高(Zhu, 2016; Manishankar et al., 2018; Toyota et al., 2018).这导致Ca2+结合的感应蛋白构象发生变化, 随之与下游效应蛋白以Ca2+依赖方式相互作用并调控其功能(Tang et al., 2020).胞内Ca2+与CBL在其EF-hands处结合, 而CIPK的C端NAF/FISH结构域与CBL互作(Guo et al., 2001; Kolukisaoglu et al., 2004; Su et al., 2020).CBL与CIPK结合使得CIPK的C端自抑制结构域释放N端激酶结构域, 从而增强CIPK活性(Zhou et al., 2016; Yin et al., 2020). ...
... 低浓度Na+对植物生长有一定的刺激作用, 但高浓度Na+则会引起细胞渗透和离子毒害(Zhu, 2016).拟南芥SOS信号途径是调节植物根耐盐性的重要机制(Zhu et al., 1998; Yin et al., 2020).在盐胁迫下, AtCBL10与AtCIPK24 (SOS2)互作(图1; 表2), 将Na+区域化到液泡, 以保护植物地上部免受盐胁迫伤害(Hu et al., 2015; Plasencia et al., 2021).这与SOS通路的耐盐机制不同(Yang et al., 2019).被激活的SOS1利用H+-ATPase (AHA2)产生的质子梯度能量将Na+转运出细胞(图1) (Yin et al., 2020).CBL10与CIPK8形成的复合物介导SOS1的激活, 表明CIPK8在调节拟南芥地上部响应盐胁迫中起重要作用(Yin et al., 2020), 但其复合物下游靶标蛋白还有待深入鉴定.细胞内较高的K+/Na+比是植物耐盐性的核心指标(Zhang et al., 2020).研究表明, HKT1 (high affinity potassium transporters 1)能够维持细胞内高浓度的K+和低浓度Na+ (Almeida et al., 2017).水稻(Oryza sativa) OsHKT1;1主要在地上部维管组织中表达, 高盐条件下可增强细胞的Na+外排能力(Campbell et al., 2017).此外, 过表达OsHKT1;4的转基因植株木质部中Na+含量降低, 而根中的Na+含量增加, 从而提高对盐胁迫的敏感性(Oda et al., 2018).CBL2/CBL3-CIPK21也参与植物的耐盐性, 该复合物主要在液泡中发挥作用, 但其下游靶点仍有待确定(Sanyal et al., 2016). ...
... ) (Yin et al., 2020).CBL10与CIPK8形成的复合物介导SOS1的激活, 表明CIPK8在调节拟南芥地上部响应盐胁迫中起重要作用(Yin et al., 2020), 但其复合物下游靶标蛋白还有待深入鉴定.细胞内较高的K+/Na+比是植物耐盐性的核心指标(Zhang et al., 2020).研究表明, HKT1 (high affinity potassium transporters 1)能够维持细胞内高浓度的K+和低浓度Na+ (Almeida et al., 2017).水稻(Oryza sativa) OsHKT1;1主要在地上部维管组织中表达, 高盐条件下可增强细胞的Na+外排能力(Campbell et al., 2017).此外, 过表达OsHKT1;4的转基因植株木质部中Na+含量降低, 而根中的Na+含量增加, 从而提高对盐胁迫的敏感性(Oda et al., 2018).CBL2/CBL3-CIPK21也参与植物的耐盐性, 该复合物主要在液泡中发挥作用, 但其下游靶点仍有待确定(Sanyal et al., 2016). ...
... ).CBL10与CIPK8形成的复合物介导SOS1的激活, 表明CIPK8在调节拟南芥地上部响应盐胁迫中起重要作用(Yin et al., 2020), 但其复合物下游靶标蛋白还有待深入鉴定.细胞内较高的K+/Na+比是植物耐盐性的核心指标(Zhang et al., 2020).研究表明, HKT1 (high affinity potassium transporters 1)能够维持细胞内高浓度的K+和低浓度Na+ (Almeida et al., 2017).水稻(Oryza sativa) OsHKT1;1主要在地上部维管组织中表达, 高盐条件下可增强细胞的Na+外排能力(Campbell et al., 2017).此外, 过表达OsHKT1;4的转基因植株木质部中Na+含量降低, 而根中的Na+含量增加, 从而提高对盐胁迫的敏感性(Oda et al., 2018).CBL2/CBL3-CIPK21也参与植物的耐盐性, 该复合物主要在液泡中发挥作用, 但其下游靶点仍有待确定(Sanyal et al., 2016). ...
Comparative genomic analysis of CIPK gene family in Arabidopsis and Populus 1 2007
... CBL和CIPK的生理学功能首次在SOS途径中被发现(Zhu et al., 1998).拟南芥突变体atsos1、atsos2和atsos3在高盐胁迫下产生相同的盐敏感表型.AtSOS3和AtSOS2也分别被称为AtCBL4和AtCIPK24, 而AtSOS3和AtSOS2已被证明协同正向调节质膜Na+/H+转运蛋白AtSOS1的活性, 在高盐胁迫下介导Na+外排(Zhu et al., 1998; Sanyal et al., 2015).随后, 在拟南芥中鉴定出10个CBLs和26个CIPKs (Kolukisaoglu et al., 2004; Yu et al., 2007).随着植物基因组测序的完成, 在不同物种中相继鉴定出数量不等的CBLs和CIPKs家族成员(表1). ...
Identification and characterization of CBL and CIPK gene families in canola (Brassica napus L.) 1 2014
... The CBL and CIPK families in different species Table 1
物种
CBLs数量
CIPKs数量
参考文献
拟南芥(Arabidopsis thaliana)
10
26
Kolukisaoglu et al., 2004
油菜(Brassica napus)
7
23
Zhang et al., 2014a
杨树(Populus trichocarpa)
10
25
Weinl and Kudla, 2009
辣椒(Capsicum annuum)
9
26
Ma et al., 2019
水稻(Oryza sativa)
10
31
Weinl and Kudla, 2009; Piao et al., 2010
茶(Camellia sinensis)
7
18
Liu et al., 2019
小麦(Triticum aestivum)
24
79
Sun et al., 2015; Liu et al., 2018
芜菁(B. rapa var. rapa)
19
51
Yin et al., 2017
木薯(Manihot esculenta)
8
26
Hu et al., 2015; Mo et al., 2018
茄子(Solanum melongena)
5
15
Li et al., 2016
菠萝(Ananas comosus)
8
21
Aslam et al., 2019
葡萄(Vitis vinifera)
8
20
Xi et al., 2017
玉米(Zea mays)
8
43
Chen et al., 2011
甜菜(Beta vulgaris)
7
20
未发表数据
2 植物CBL-CIPK系统与磷酸化 CBL-CIPK复合物对下游靶标蛋白的调节主要通过磷酸化方式进行(Sanyal et al., 2020).靶标蛋白主要有通道蛋白、转运蛋白和转录因子等(Sanyal et al., 2015).已有研究表明, 可被CBL-CIPK复合物磷酸化的阳离子(H+、Na+、K+、Mg2+、Fe2+和NH4+)相关蛋白如AHA2、SOS1、AMT1、ATL31、AKT1、HAK5、TPK1、ARF、ERF、SnRK2D和FIT, 而一些阴离子(OH-和NO3-)转运蛋白如CHL1 (NRT1.1)、RBOH-F和RBOH-C也可被CBL-CIPK磷酸化(Ragel et al.,2019; Lara et al., 2020; Sanyal et al., 2020; Dong et al., 2021). ...
Emerging crosstalk between two signaling pathways coordinates K+ and Na+ homeostasis in the halophyte Hordeum brevisubulatum 1 2020
... 低浓度Na+对植物生长有一定的刺激作用, 但高浓度Na+则会引起细胞渗透和离子毒害(Zhu, 2016).拟南芥SOS信号途径是调节植物根耐盐性的重要机制(Zhu et al., 1998; Yin et al., 2020).在盐胁迫下, AtCBL10与AtCIPK24 (SOS2)互作(图1; 表2), 将Na+区域化到液泡, 以保护植物地上部免受盐胁迫伤害(Hu et al., 2015; Plasencia et al., 2021).这与SOS通路的耐盐机制不同(Yang et al., 2019).被激活的SOS1利用H+-ATPase (AHA2)产生的质子梯度能量将Na+转运出细胞(图1) (Yin et al., 2020).CBL10与CIPK8形成的复合物介导SOS1的激活, 表明CIPK8在调节拟南芥地上部响应盐胁迫中起重要作用(Yin et al., 2020), 但其复合物下游靶标蛋白还有待深入鉴定.细胞内较高的K+/Na+比是植物耐盐性的核心指标(Zhang et al., 2020).研究表明, HKT1 (high affinity potassium transporters 1)能够维持细胞内高浓度的K+和低浓度Na+ (Almeida et al., 2017).水稻(Oryza sativa) OsHKT1;1主要在地上部维管组织中表达, 高盐条件下可增强细胞的Na+外排能力(Campbell et al., 2017).此外, 过表达OsHKT1;4的转基因植株木质部中Na+含量降低, 而根中的Na+含量增加, 从而提高对盐胁迫的敏感性(Oda et al., 2018).CBL2/CBL3-CIPK21也参与植物的耐盐性, 该复合物主要在液泡中发挥作用, 但其下游靶点仍有待确定(Sanyal et al., 2016). ...
Protein phosphorylation in stomatal movement 2 2014
... 与ROS有关的氧化酶RBOHF (respiratory burst oxidase homolog factor)受到CBL1/9-CIPK26的调控(表2; 图2) (Zhang et al., 2014b).CIPK26可影响定位在质膜上的RBOHF活性(Kimura et al., 2013).然而, 该结果是在异源表达系统中试验得到的, 因此CIPK26对RBOHF活性的调节作用尚缺乏足够的证据.但可以肯定的是, CBL1/CBL9介导CIPK26到质膜的定位导致RBHOF磷酸化(Zhang et al., 2018b; Han et al., 2019). ...
CBL1-CIPK26-mediated phosphorylation enhances activity of the NADPH oxidase RBOHC, but is dispensable for root hair growth 1 2018
... 与ROS有关的氧化酶RBOHF (respiratory burst oxidase homolog factor)受到CBL1/9-CIPK26的调控(表2; 图2) (Zhang et al., 2014b).CIPK26可影响定位在质膜上的RBOHF活性(Kimura et al., 2013).然而, 该结果是在异源表达系统中试验得到的, 因此CIPK26对RBOHF活性的调节作用尚缺乏足够的证据.但可以肯定的是, CBL1/CBL9介导CIPK26到质膜的定位导致RBHOF磷酸化(Zhang et al., 2018b; Han et al., 2019). ...
Sensing of abiotic stress and ionic stress responses in plants 1 2018
... 植物在生长过程中经常遇到干旱、盐碱、极端温度、缺K+以及病虫害等多种逆境胁迫(Peck and Mittler, 2020).植物无法逃避逆境, 只能被动应对.因此, 植物在长期进化过程中演化出复杂的信号转导机制, 感知并传递外界变化的信号, 使其对逆境及时做出应激反应(Manik et al., 2015).在植物体内, 许多信号转导途径和调控网络相互作用, 响应特定的生物或非生物胁迫.植物对逆境的响应与激酶、磷酸酶、激素、Ca2+、ROS (reactive oxygen species)以及其它调节蛋白、化合物和小分子相互作用交织在一起, 形成复杂的调控网络(Steinhorst and Kudla, 2013; Sierla et al., 2016; Kudla et al., 2018; Zhang et al., 2018a). ...
Foxtail millet (Setaria italica (L.) P. Beauv) CIPKs are responsive to ABA and abiotic stresses 1 2019
... 已有研究表明, 在ABA信号通路中起作用的转录因子也会被CIPK磷酸化(Sanyal et al., 2016; Zhao et al., 2019).最早的报道是ERF7 (ethylene responsive factor 7)被CIPK15磷酸化(Song et al., 2005).磷酸化后的ERF7可能通过2种机制发挥作用: 一是它们结合含有GCC盒的基因并抑制其转录; 二是ERF7可以将AtSin3和HDA19之间的阻遏复合物靶向移至相关基因的启动子, 以进一步抑制基因转录(Song et al., 2005).与之相反, CIPK26介导的ABI5磷酸化是正向调节因子, 可稳定ABI5并启动ABA的下游反应(Lyzenga et al., 2017).KEG (keep on going)使ABI5和CIPK26受到26S蛋白酶的降解, 从而抵消ABA的作用(Lyzenga et al., 2017).此外, PKS5/CIPK11使ABI5在Ser42处磷酸化并激活, 从而调节基因表达(Zhou et al., 2015).因此, CIPK26和PKS5/CIPK11可能在同一途径中发挥相似作用(Zhou et al., 2015).CIPK3与ABR1 (ABA-repressor 1)相互作用并使其磷酸化, 从而调节种子萌发过程中的ABA应答反应(Sanyal et al., 2017; Wang et al., 2018). ...
Anion channel SLAH3 functions in nitrate-dependent alleviation of ammonium toxicity in Arabidopsis 2 2015
... NO3-和NH4+是植物根系从土壤中吸收的2种N素来源(Tang et al., 2020).Ca2+调控多种NRT/NPF和SLAH转运蛋白进行NO3-摄取和转运(Sanyal et al., 2020).NPF6.3通过二聚体作用使NO3-从低亲和性模式转换为高亲和性模式, 此过程通过CBL1/9- CIPK23对Thr101的磷酸化作用进行控制(Ho et al., 2009; Parker and Newstead, 2014; Sun et al., 2014; Chu et al., 2021).NH4+吸收由AMTs (ammonium transporters)介导(Sanyal et al., 2020).高浓度NH4+会替代K+而产生毒害作用, 因此必须控制其在细胞的非毒性水平(Zheng et al., 2015).AMT的2个成员(AMT1;1和AMT1;2)也可被CBL1-CIPK23复合物抑制(表2; 图2) (Straub et al., 2017).因此, CBL1-CIPK23可能在维持根细胞K+稳态和免受NH4+毒害中起关键作用(Zheng et al., 2015).Weng等(2020)研究表明, 高浓度K+降低水稻根系对NH4+的吸收速率, 增强对H+的外排速率, 从而减轻NH4+对水稻的毒害.此外, CBL1/9-CIPK23与SLAH2和SLAH3互作通过吸收或分泌NO3-参与硝酸盐的转运(表2) (Ho et al., 2009; Léran et al., 2015). ...
... 毒害中起关键作用(Zheng et al., 2015).Weng等(2020)研究表明, 高浓度K+降低水稻根系对NH4+的吸收速率, 增强对H+的外排速率, 从而减轻NH4+对水稻的毒害.此外, CBL1/9-CIPK23与SLAH2和SLAH3互作通过吸收或分泌NO3-参与硝酸盐的转运(表2) (Ho et al., 2009; Léran et al., 2015). ...
SOS2-like protein kinase 5, an SNF1-related protein kinase 3-type protein kinase, is important for abscisic acid responses in Arabidopsis through phosphorylation of abscisic acid-insensitive 5 3 2015
... 已有研究表明, 在ABA信号通路中起作用的转录因子也会被CIPK磷酸化(Sanyal et al., 2016; Zhao et al., 2019).最早的报道是ERF7 (ethylene responsive factor 7)被CIPK15磷酸化(Song et al., 2005).磷酸化后的ERF7可能通过2种机制发挥作用: 一是它们结合含有GCC盒的基因并抑制其转录; 二是ERF7可以将AtSin3和HDA19之间的阻遏复合物靶向移至相关基因的启动子, 以进一步抑制基因转录(Song et al., 2005).与之相反, CIPK26介导的ABI5磷酸化是正向调节因子, 可稳定ABI5并启动ABA的下游反应(Lyzenga et al., 2017).KEG (keep on going)使ABI5和CIPK26受到26S蛋白酶的降解, 从而抵消ABA的作用(Lyzenga et al., 2017).此外, PKS5/CIPK11使ABI5在Ser42处磷酸化并激活, 从而调节基因表达(Zhou et al., 2015).因此, CIPK26和PKS5/CIPK11可能在同一途径中发挥相似作用(Zhou et al., 2015).CIPK3与ABR1 (ABA-repressor 1)相互作用并使其磷酸化, 从而调节种子萌发过程中的ABA应答反应(Sanyal et al., 2017; Wang et al., 2018). ...
... ).因此, CIPK26和PKS5/CIPK11可能在同一途径中发挥相似作用(Zhou et al., 2015).CIPK3与ABR1 (ABA-repressor 1)相互作用并使其磷酸化, 从而调节种子萌发过程中的ABA应答反应(Sanyal et al., 2017; Wang et al., 2018). ...
Hyperactive mutant of a wheat plasma membrane Na+/H+ antiporter improves the growth and salt tolerance of transgenic tobacco 1 2016
... 在这些必需的离子中, Ca2+浓度通常相对较低,在细胞质中保持在0.1 mmol·L-1左右(Hepler, 2005).然而, 当植物遭遇干旱、盐碱、病原体侵害或营养缺乏等逆境时, Ca2+通过质膜离子通道流入细胞中或从细胞器中释放, 使胞质Ca2+浓度迅速升高(Zhu, 2016; Manishankar et al., 2018; Toyota et al., 2018).这导致Ca2+结合的感应蛋白构象发生变化, 随之与下游效应蛋白以Ca2+依赖方式相互作用并调控其功能(Tang et al., 2020).胞内Ca2+与CBL在其EF-hands处结合, 而CIPK的C端NAF/FISH结构域与CBL互作(Guo et al., 2001; Kolukisaoglu et al., 2004; Su et al., 2020).CBL与CIPK结合使得CIPK的C端自抑制结构域释放N端激酶结构域, 从而增强CIPK活性(Zhou et al., 2016; Yin et al., 2020). ...
Abiotic stress signaling and responses in plants 2 2016
... 在这些必需的离子中, Ca2+浓度通常相对较低,在细胞质中保持在0.1 mmol·L-1左右(Hepler, 2005).然而, 当植物遭遇干旱、盐碱、病原体侵害或营养缺乏等逆境时, Ca2+通过质膜离子通道流入细胞中或从细胞器中释放, 使胞质Ca2+浓度迅速升高(Zhu, 2016; Manishankar et al., 2018; Toyota et al., 2018).这导致Ca2+结合的感应蛋白构象发生变化, 随之与下游效应蛋白以Ca2+依赖方式相互作用并调控其功能(Tang et al., 2020).胞内Ca2+与CBL在其EF-hands处结合, 而CIPK的C端NAF/FISH结构域与CBL互作(Guo et al., 2001; Kolukisaoglu et al., 2004; Su et al., 2020).CBL与CIPK结合使得CIPK的C端自抑制结构域释放N端激酶结构域, 从而增强CIPK活性(Zhou et al., 2016; Yin et al., 2020). ...
... 低浓度Na+对植物生长有一定的刺激作用, 但高浓度Na+则会引起细胞渗透和离子毒害(Zhu, 2016).拟南芥SOS信号途径是调节植物根耐盐性的重要机制(Zhu et al., 1998; Yin et al., 2020).在盐胁迫下, AtCBL10与AtCIPK24 (SOS2)互作(图1; 表2), 将Na+区域化到液泡, 以保护植物地上部免受盐胁迫伤害(Hu et al., 2015; Plasencia et al., 2021).这与SOS通路的耐盐机制不同(Yang et al., 2019).被激活的SOS1利用H+-ATPase (AHA2)产生的质子梯度能量将Na+转运出细胞(图1) (Yin et al., 2020).CBL10与CIPK8形成的复合物介导SOS1的激活, 表明CIPK8在调节拟南芥地上部响应盐胁迫中起重要作用(Yin et al., 2020), 但其复合物下游靶标蛋白还有待深入鉴定.细胞内较高的K+/Na+比是植物耐盐性的核心指标(Zhang et al., 2020).研究表明, HKT1 (high affinity potassium transporters 1)能够维持细胞内高浓度的K+和低浓度Na+ (Almeida et al., 2017).水稻(Oryza sativa) OsHKT1;1主要在地上部维管组织中表达, 高盐条件下可增强细胞的Na+外排能力(Campbell et al., 2017).此外, 过表达OsHKT1;4的转基因植株木质部中Na+含量降低, 而根中的Na+含量增加, 从而提高对盐胁迫的敏感性(Oda et al., 2018).CBL2/CBL3-CIPK21也参与植物的耐盐性, 该复合物主要在液泡中发挥作用, 但其下游靶点仍有待确定(Sanyal et al., 2016). ...
Genetic analysis of salt tolerance in Arabidopsis: evidence for a critical role of potassium nutrition 4 1998
... CBL和CIPK的生理学功能首次在SOS途径中被发现(Zhu et al., 1998).拟南芥突变体atsos1、atsos2和atsos3在高盐胁迫下产生相同的盐敏感表型.AtSOS3和AtSOS2也分别被称为AtCBL4和AtCIPK24, 而AtSOS3和AtSOS2已被证明协同正向调节质膜Na+/H+转运蛋白AtSOS1的活性, 在高盐胁迫下介导Na+外排(Zhu et al., 1998; Sanyal et al., 2015).随后, 在拟南芥中鉴定出10个CBLs和26个CIPKs (Kolukisaoglu et al., 2004; Yu et al., 2007).随着植物基因组测序的完成, 在不同物种中相继鉴定出数量不等的CBLs和CIPKs家族成员(表1). ...
... 外排(Zhu et al., 1998; Sanyal et al., 2015).随后, 在拟南芥中鉴定出10个CBLs和26个CIPKs (Kolukisaoglu et al., 2004; Yu et al., 2007).随着植物基因组测序的完成, 在不同物种中相继鉴定出数量不等的CBLs和CIPKs家族成员(表1). ...
... 低浓度Na+对植物生长有一定的刺激作用, 但高浓度Na+则会引起细胞渗透和离子毒害(Zhu, 2016).拟南芥SOS信号途径是调节植物根耐盐性的重要机制(Zhu et al., 1998; Yin et al., 2020).在盐胁迫下, AtCBL10与AtCIPK24 (SOS2)互作(图1; 表2), 将Na+区域化到液泡, 以保护植物地上部免受盐胁迫伤害(Hu et al., 2015; Plasencia et al., 2021).这与SOS通路的耐盐机制不同(Yang et al., 2019).被激活的SOS1利用H+-ATPase (AHA2)产生的质子梯度能量将Na+转运出细胞(图1) (Yin et al., 2020).CBL10与CIPK8形成的复合物介导SOS1的激活, 表明CIPK8在调节拟南芥地上部响应盐胁迫中起重要作用(Yin et al., 2020), 但其复合物下游靶标蛋白还有待深入鉴定.细胞内较高的K+/Na+比是植物耐盐性的核心指标(Zhang et al., 2020).研究表明, HKT1 (high affinity potassium transporters 1)能够维持细胞内高浓度的K+和低浓度Na+ (Almeida et al., 2017).水稻(Oryza sativa) OsHKT1;1主要在地上部维管组织中表达, 高盐条件下可增强细胞的Na+外排能力(Campbell et al., 2017).此外, 过表达OsHKT1;4的转基因植株木质部中Na+含量降低, 而根中的Na+含量增加, 从而提高对盐胁迫的敏感性(Oda et al., 2018).CBL2/CBL3-CIPK21也参与植物的耐盐性, 该复合物主要在液泡中发挥作用, 但其下游靶点仍有待确定(Sanyal et al., 2016). ...
 ,*兰州理工大学生命科学与工程学院, 兰州 730050
,*兰州理工大学生命科学与工程学院, 兰州 730050
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT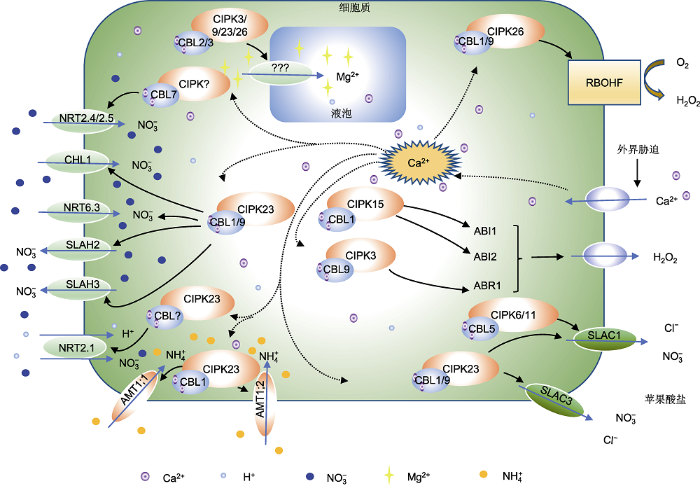 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}