Secondary Metabolites of Syringa and the Linking with Phylogenetic Evolution and Geographical Distributions
Zhaoyu Zhang1,2,3, Qingyun Wang4, Lei Shi3, Wengang Yu4, Yongqing Zhang,1,2,*, Hongxia Cui,3,*1Shandong University of Traditional Chinese Medicine, Jinan 250300, China 2Key Laboratory of Chinese Medicine Resources in Colleges and Universities of Shandong Province, Jinan 250355, China 3Key Laboratory of Plant Resources and Beijing Botanical Garden, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 4Horticulture College, Hainan University, Haikou 570228, China
Abstract Secondary metabolites of plants induced by environmental factors are highly variable, but the given metabolic pathways may have some phylogenetic implications. Due to the difficulty in complete and systematic collections in certain plant groups, the research on the correlation between secondary metabolites and phylogeny is limited. Based on the published papers, 377 secondary metabolites in the roots, stems, leaves and flowers of Syringa were collected, which mainly derived from the mevalonic acid pathway, deoxyxylulose-5-P pathway and shikimic acid pathway. After superimposing phylogenetic background, we found that dominance of a given type of secondary metabolites was high for the firstly diverged series, and the dominance declined for subsequently diverged series with the increase of chemical diversity. Phenylpropanoids and iridoids/secoiridoids were phylogenetically conserved. After superimposing geographical distributions, we found that some local species which were lately diverged had more diverse secondary metabolites compared with widespread species firstly diverged. The high proportion of lignans was highly related to the environmental pressure. This review provided a new clue for the systematic study on the variation pattern of chemical diversity in the taxa within genus in the light of evolution. Keywords:Syringa;secondary metabolites;phylogeny;evolution;environmental stress
PDF (1138KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 张照宇, 王清芸, 石雷, 余文刚, 张永清, 崔洪霞. 丁香属次生代谢产物及其与系统演化和地理环境的关联. 植物学报, 2021, 56(4): 470-479 doi:10.11983/CBB20178 Zhang Zhaoyu, Wang Qingyun, Shi Lei, Yu Wengang, Zhang Yongqing, Cui Hongxia. Secondary Metabolites of Syringa and the Linking with Phylogenetic Evolution and Geographical Distributions. Chinese Bulletin of Botany, 2021, 56(4): 470-479 doi:10.11983/CBB20178
Table 1 表1 表1已报道次生代谢成分的丁香属种质地理分布 Table 1Geographical distributions of Syringa with secondary metabolites reported
组系
物种
自然分布
海拔(m)
经度(E)
纬度(N)
生态幅
欧丁香系 (Ser. Syringa)
欧洲丁香(S. vulgaris)
东南欧(阿尔卑斯山和喀尔巴阡山)
1000-1200
5°36'-66°10'
36°00'-67°46'
广布种
紫丁香(S. oblata)
中国东北、华北、西北、华东和川西北
300-2400
96°23'-135°02'
34°19'-55°33'
广布种
朝阳丁香(S. oblata var. dilatata)
中国华北-东北-朝鲜半岛
300-2400
40°51'-129°40'
33°56'-53°19'
广布种
阿富汗丁香(S. afghanica)
中国青海-阿富汗
60°29'-74°53'
29°21'-38°27'
局域种
羽叶丁香系 (Ser. Pinnatifoliae)
羽叶丁香(S. pinnatifolia)
贺兰山、陕西南部、甘肃、青海东部和四川南部
1700-3100
105°13'-112°32'
26°03'-37°09'
局域种
短花冠管组 (Sect. Ligustrina)
暴马丁香(S. amurensis)
中国东北和西北, 俄罗斯远东地区, 朝鲜
100-1200
103°04'-163°19'
31°09'-72°33'
广布种
日本丁香(S. reticulata)
日本北部(北海道地区)
139°20'-148°53'
40°33'-45°33'
局域种
巧玲花系 (Ser. Pubescentes)
巧玲花(S. pubescens)
河北、陕西东部、山西东部和河南
900-2100
109°29'-119°53'
31°23'-42°37'
局域种
关东丁香(S. velutina)
辽宁和吉林长白山区
300-1200
118°53'-135°05'
38°43'-53°33'
局域种
红丁香系 (Ser. Villosae)
西蜀丁香(S. komarowii)
甘肃南部、陕西南部、 四川和云南北部
105°31'-114°11'
24°20'-35°28'
局域种
辽东丁香(S. wolfii)
中国东北, 朝鲜
500-1600
118°53'-128°28'
37°35'-72°33'
局域种
表中仅列次生代谢成分已报道的9个原种和2个变种, 涉及丁香属下5个组系的所有分类等级。生态幅指丁香属内特定物种的地理分布跨度, 即其分布的经度、纬度或海拔数据中最小值与最大值之间的范围。广布种指经度跨度大于30°或纬度跨度大于20°的物种, 局域种指经度跨度小于30°或纬度跨度小于20°的物种。 The species with secondary metabolites reported in the table include nine native species and two varieties, involving four series and one section of Syringa. Ecological amplitude refers to the geographical span of a species, the range between the highest and lowest points of longitude, latitude and altitude. Widespread species refers to the species whose longitude span is more than 30° or latitude span is more than 20°, and local species refers to the species whose longitude span is less than 30° or latitude span is less than 20°.
Table 2 表2 表2丁香属组系水平的次生代谢产物类别及各类成分计数(单位: 个) Table 2The classification and count of secondary metabolites in Syringa at the level of series (section) (unit: number of components)
代谢产物类别
欧丁香系 (Ser. Syringa)
羽叶丁香系 (Ser. Pinnatifoliae)
短花冠管组 (Sect. Ligustrina)
巧玲花系 (Ser. Pubescentes)
红丁香系 (Ser. Villosae)
环/裂环烯醚萜
96
6
17
9
2
倍半萜
8
46
6
10
-
苯丙素
30
5
11
8
8
木脂素
5
58
8
1
5
黄酮
8
-
-
5
-
三萜
15
8
2
2
12
单萜
3
-
-
-
-
脂肪酸
3
-
-
-
-
醌类
-
2
-
-
-
生物碱
-
-
-
1
3
合计
168
125
44
36
30
- 表示该类产物在特定组系中未见报道。成分来自根、茎、花、叶和茎皮多个器官。各类产物计数指具有相同母核但取代基团、基团数量、基团位置和空间构象不同的化合物个数。 - indicates that the components have not been reported in the series (section). The components came from multiple organs, including root, stem, flower, leaf and stem bark. The component counting refers to the number of the components with the same parent nucleus but different in substituent groups, group numbers, group positions and spatial conformations.
Table 3 表3 表3丁香属物种水平的次生代谢产物类别及成分计数(单位: 个) Table 3The classification and count of secondary metabolites in Syringa at the level of species (unit: number of compound)
代谢产物类别
欧丁香系 (Ser. Syringa)
羽叶丁香系 (Ser. Pinna tifoliae)
短花冠管组 (Sect. Ligustrina)
巧玲花系 (Ser. Pubescentes)
红丁香系 (Ser. Villosae)
欧洲丁香 (S. vulgaris)
紫丁香 (S. ob- lata)
朝阳丁香 (S. oblata var. dilatata)
阿富汗丁香 (S. afghanica)
羽叶丁香 (S. pinnatifolia)
暴马丁香 (S. amu- rensis)
日本丁香 (S. reticulata)
关东丁香 (S. velutina)
巧玲花 (S. pubescens)
辽东丁香 (S.wolfii)
西蜀丁香 (S. komarowii)
环/裂环烯醚萜
60
16
10
20
6
13
5
7
3
-
2
倍半萜
-
6
2
-
46
6
-
-
10
-
-
苯丙素
28
3
-
-
5
6
5
7
1
-
8
木脂素
5
-
-
-
58
1
7
1
-
-
5
黄酮
5
5
-
-
-
-
-
5
-
-
-
三萜
-
14
2
-
8
2
-
2
-
2
10
单萜
-
3
-
-
-
-
-
-
-
-
-
脂肪酸
1
2
-
-
-
-
-
-
-
-
-
醌类
-
-
-
-
2
-
-
-
-
-
-
生物碱
-
-
-
-
-
-
-
1
-
-
3
合计
99
49
14
20
125
28
17
23
14
2
28
- 表示该类产物在特定物种中未见报道。成分来自根、茎、花、叶和茎皮。各类成分计数包括具有相同母核但取代基团、基团数量、基团位置和空间构象不同的化合物。阿富汗丁香、辽东丁香和西蜀丁香数据分别仅来源于1篇文献。 - indicate that the components have not been reported in correspoding species. The components came from root, stem, flower, leaf and stem bark. The component counting includes different compounds with the same parent nucleus but different in substituent groups, group numbers, group positions and spatial conformations. The data of S. afghanica, S. wolfii and S. komarowii were only obtained from one reference, respectively.
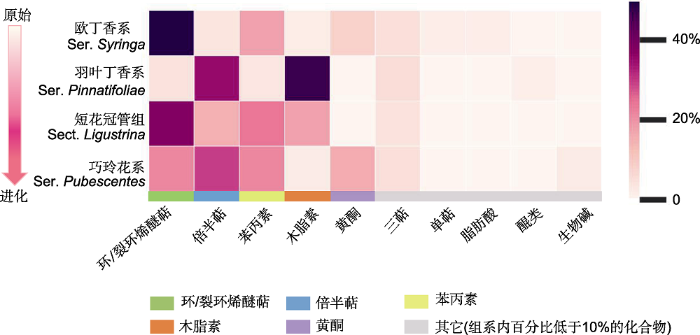
色块内颜色从深到浅表示各类产物的相对数量占比从高到低的连续变化(组系内特定类别产物的相对数量占比=特定类别产物的成分计数/该组系内所有类别产物计数总和), 同一类别产物包含具有相同母核但取代基团、基团数量、基团位置和空间构象不同的化合物。热图下方的不同颜色条表示不同类别产物。为保证成分计数占比分析的客观性, 图中每个物种收入的成分信息需至少来自3篇文献。欧丁香系的阿富汗丁香及红丁香系的辽东丁香和西蜀丁香因各仅有1篇文献而暂未收入。 Figure 1Proportion of secondary metabolites in Syringa at the level of series (section)
The color in the block progressively varying from dark to light represents the continuous change in the percentage of components in the series (section) from high to low (the percentage of specific type of components in a series (section) = the number of specific type of components in a series (section)/the total number of components in this series (section)). The products of the same type include different components with the same parent nucleus but different in substituent groups, group numbers, group positions and conformations. The different color-stipes below the heatmap represent different types of products. In order to ensure the objective counting of the components, at least three reference were required for the metabolite information of each species. Therefore, the S. afghanica in Ser. Syringa as well as the S. wolfii and S. komarowii in Ser. Villosae has not been included because only one related reference was found.
(A) 丁香属组系的系统演化关系(Li et al., 2012)及各类产物在各组系中的占比; (B) 甲戊二羟酸途径(MVA)和脱氧木酮糖磷酸酯途径(DXP) (Mint Evolutionary Genomics Consortium, 2018); (C) 莽草酸途径(刘津等, 2016; 张旭等, 2019)。图(A)组系下方的色条表示各类产物(颜色含义同图1), 色条内的百分数指特定类别产物计数在该组系内成分计数总和中所占的百分比(详见图1图注)。图(B)和图(C)中化合物色框颜色含义与图(A)相同。图(B)和图(C)中虚线表示由多步反应完成。 Figure 2The phylogenetic relationship of Syringa, and the percentage of metabolites and their main metabolic pathways at the level of series (section)
(A) The phylogenetic relationship (Li et al., 2012) and percentage of metabolites of Syringa at the level of series (section); (B) Mevalonic acid pathway (MVA) and Deoxyxylulose-5-P pathway (DXP) (Mint Evolutionary Genomics Consortium, 2018); (C) Shikimic acid pathway (Liu et al., 2016; Zhang et al., 2019, in Chinese). The color-stripes below the series (section) indicate different types of metabolites in (A), and the percentage in the color-stripe represents the percentage of a given type of compounds within the series (section) (please see Figure 1 for details). The meanings of the color frames indicating the compound in (B) and (C) are the same as those of (A). The dotted lines in (B) and (C) represent the process of multi-step reactions.
AllevatoDM, GroppoM, KiyotaE, MazzaferaP, NixonKC (2019). Evolution of phytochemical diversity in Pilocarpus (Rutaceae) Phytochemistry 163, 132-146. DOI:10.1016/j.phytochem.2019.03.027URL [本文引用: 1]
ChadwickM, TrewinH, GawthropF, WagstaffC (2013). Sesquiterpenoids lactones: benefits to plants and people Int J Mol Sci 14, 12780-12805. DOI:10.3390/ijms140612780PMID:23783276 [本文引用: 1] Sesquiterpenoids, and specifically sesquiterpene lactones from Asteraceae, may play a highly significant role in human health, both as part of a balanced diet and as pharmaceutical agents, due to their potential for the treatment of cardiovascular disease and cancer. This review highlights the role of sesquiterpene lactones endogenously in the plants that produce them, and explores mechanisms by which they interact in animal and human consumers of these plants. Several mechanisms are proposed for the reduction of inflammation and tumorigenesis at potentially achievable levels in humans. Plants can be classified by their specific array of produced sesquiterpene lactones, showing high levels of translational control. Studies of folk medicines implicate sesquiterpene lactones as the active ingredient in many treatments for other ailments such as diarrhea, burns, influenza, and neurodegradation. In addition to the anti-inflammatory response, sesquiterpene lactones have been found to sensitize tumor cells to conventional drug treatments. This review explores the varied ecological roles of sesquiterpenes in the plant producer, depending upon the plant and the compound. These include allelopathy with other plants, insects, and microbes, thereby causing behavioural or developmental modification to these secondary organisms to the benefit of the sesquiterpenoid producer. Some sesquiterpenoid lactones are antimicrobial, disrupting the cell wall of fungi and invasive bacteria, whereas others protect the plant from environmental stresses that would otherwise cause oxidative damage. Many of the compounds are effective due to their bitter flavor, which has obvious implications for human consumers. The implications of sesquiterpenoid lactone qualities for future crop production are discussed.
ChenF, ThollD, BohlmannJ, PicherskyE (2011). The family of terpene synthases in plants: a mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom Plant J 66, 212-229. DOI:10.1111/tpj.2011.66.issue-1URL [本文引用: 1]
ChenYG, MulderPPJ, SchaapO, MemelinkJ, KlinkhamerPGL, VrielingK (2020). The evolution of pyrrolizidine alkaloid diversity among and within Jacobaea species J Syst Evol doi: 10.1111/jse.12671. [本文引用: 2]
CuiHX, CongSH, WangXZ, HaoHP, ShiL, ZhangHJ, LiZG, HuTH, QinYS (2016). Mechanistic examination of causes for narrow distribution in an endangered shrub: a comparison of its responses to drought stress with a widespread congeneric species Trees 30, 2227-2236. DOI:10.1007/s00468-016-1448-8URL [本文引用: 2]
ErnstM, Saslis-LagoudakisCH, GraceOH, NilssonN, SimonsenHT, HornJW, RønstedN (2016). Evolutionary prediction of medicinal properties in the genus Euphorbia L Sci Rep 6, 30531. DOI:10.1038/srep30531URL [本文引用: 2]
FilipekA, WyszomierskaJ, MichalakB, KissAK (2019). Syringa vulgaris bark as a source of compounds affecting the release of inflammatory mediators from human neutrophils and monocytes/macrophages Phytochem Lett 30, 309-313. DOI:10.1016/j.phytol.2019.02.008URL [本文引用: 2]
GuittonY, NicolèF, JullienF, CaissardJC, Saint-MarcouxD, LegendreL, PasquierB, MojaS (2018). A comparative study of terpene composition in different clades of the genus Lavandula Bot Lett 165, 494-505. [本文引用: 1]
KimKJ, JansenRK (1998). A chloroplast DNA phylogeny of lilacs (Syringa, Oleaceae): plastome groups show a strong correlation with crossing groups Am J Bot 85, 1338-1351. PMID:21685019 [本文引用: 2] Phylogenetic relationships and genomic compatibility were compared for 60 accessions of Syringa using chloroplast DNA (cpDNA) and nuclear ribosomal DNA (rDNA) markers. A total of 669 cpDNA variants, 653 of which were potentially phylogenetically informative, was detected using 22 restriction enzymes. Phylogenetic analyses reveal four strongly supported plastome groups that correspond to four genetically incompatible crossing groups. Relationships of the four plastome groups (I(II(III,IV))) correlate well with the infrageneric classification except for ser. Syringa and Pinnatifoliae. Group I, which includes subg. Ligustrina, forms a basal lineage within Syringa. Group II includes ser. Syringa and Pinnatifoliae and the two series have high compatibility and low sequence divergence. Group III consists of three well-defined species groups of ser. Pubescentes. Group IV comprises all members of ser. Villosae and has the lowest interspecific cpDNA sequence divergences. Comparison of cpDNA sequence divergence with crossability data indicates that hybrids have not been successfully generated between species with divergence greater than 0.7%. Hybrid barriers are strong among the four major plastome groups, which have sequence divergence estimates ranging from 1.096 to 1.962%. In contrast, fully fertile hybrids occur between species pairs with sequence divergence below 0.4%. Three regions of the plastome have length variants of greater than 100 bp, and these indels identify 12 different plastome types that correlate with phylogenetic trees produced from cpDNA restriction site data. Biparentally inherited nuclear rDNA and maternally inherited cpDNA length variants enable the identification of the specific parentage of several lilac hybrids.
KonnoK, HirayamaC, YasuiH, NakamuraM (1999). Enzymatic activation of oleuropein: a protein crosslinker used as a chemical defense in the privet tree Proc Natl Acad Sci USA 96, 9159-9164. DOI:10.1073/pnas.96.16.9159URL [本文引用: 1]
LendvayB, KadereitJW, WestbergE, CornejoC, PedrycA, HöhnM (2016). Phylogeography of Syringa josikaea (Oleaceae): Early Pleistocene divergence from East Asian relatives and survival in small populations in the Carpathians Biol J Linn Soc 119, 689-703. DOI:10.1111/bij.12499URL [本文引用: 1]
LiJH, Goldman-HuertasB, DeYoungJ, AlexanderJ (2012). Phylogenetics and diversification of Syringa inferred from nuclear and plastid DNA sequences Castanea 77, 82-88. DOI:10.2179/11-016URL [本文引用: 9]
McKelveySD (1928). The Lilac New York, Boston, Chicago, Dallas, Atlanta, San Francisco: The Macmillan Company. pp.1-121. [本文引用: 1]
MintEvolutionary Genomics Consortium (2018). Phylogenomic mining of the mints reveals multiple mechanisms contributing to the evolution of chemical diversity in Lamiaceae Mol Plant 11, 1084-1096. DOI:10.1016/j.molp.2018.06.002URL [本文引用: 4]
RehderA (1940). Manual of Cultivated Trees and Shrubs New York:Macmillan. pp. 777-783. [本文引用: 1]
RønstedN, SavolainenV, MølgaardP, JägerAK (2008). Phylogenetic selection of Narcissus species for drug discovery Biochem Syst Ecol 36, 417-422. DOI:10.1016/j.bse.2007.12.010URL [本文引用: 1]
SedioBE (2017). Recent breakthroughs in metabolomics promise to reveal the cryptic chemical traits that mediate plant community composition, character evolution and lineage diversification New Phytol 214, 952-958. DOI:10.1111/nph.2017.214.issue-3URL [本文引用: 1]
SharmaA, ShahzadB, RehmanA, BhardwajR, LandiM, ZhengBS (2019). Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress Molecules 24, 2452. DOI:10.3390/molecules24132452URL [本文引用: 2]
ThakurM, BhattacharyaS, KhoslaPK, PuriS (2019). Improving production of plant secondary metabolites through biotic and abiotic elicitation J Appl Res Med Aroma Plants 12, 1-12. [本文引用: 1]
VargaE, BarabásC, TóthA, BoldizsárI, NoszálB, TóthG (2019). Phenolic composition, antioxidant and antinociceptive activities of Syringa vulgaris L. bark and leaf extracts. Nat Prod Res 33, 1664-1669. DOI:10.1080/14786419.2018.1425855URL [本文引用: 1]
VleminckxJ, SalazarD, FortunelC, MesonesI, DavilaN, LokvamJ, BeckleyK, BaralotoC, FinePVA (2018). Divergent secondary metabolites and habitat filtering both contribute to tree species coexistence in the Peruvian Amazon Front Plant Sci 9, 836. DOI:10.3389/fpls.2018.00836PMID:29971085 [本文引用: 1] Little is known about the mechanisms promoting or limiting the coexistence of functionally divergent species in hyperdiverse tropical tree genera. Density-dependent enemy attacks have been proposed to be a major driver for the local coexistence of chemically divergent congeneric species. At the same time, we expect local soil conditions to favor the coexistence of species sharing similar functional traits related to resource use strategies, while environmental heterogeneity would promote the diversity of these traits at both local and large spatial scales. To test how these traits mediate species coexistence, we used functional trait data for 29 species from the tree genus Protium (Burseraceae), collected in 19 plots (2 ha each) in the Peruvian Amazon. We characterized the presence-absence of 189 plant secondary metabolites (SM) for 27 of these species, and 14 functional traits associated with resource use strategies (RUT) for 16 species. Based on these data, we found that SM were significantly more dissimilar than null expectations for species co-occurring within plots, whereas RUT were significantly more similar. These results were consistent with the hypothesis that density-dependent enemy attacks contribute to the local coexistence of congeneric species displaying divergent chemical defenses, whereas local habitat conditions filter species with similar RUT. Using measurements of nine soil properties in each plot, we also found a significant turnover of RUT traits with increasing dissimilarity of soil texture and nutrient availabilities, providing support for the hypothesis that soil heterogeneity maintains functional diversity at larger spatial scales (from 500 m up to ca. 200 km) in Protium communities. Our study provides new evidence suggesting that density-dependent enemy attacks and soil heterogeneity both contribute to maintaining high species richness in diverse tropical forests.
WangSC, AlseekhS, FernieAR, LuoJ (2019). The structure and function of major plant metabolite modifications Mol Plant 12, 899-919. DOI:10.1016/j.molp.2019.06.001URL [本文引用: 1]
XuSQ, YaoSC, HuangRS, TanY, HuangD (2020). Transcriptome-wide analysis of the AP2/ERF transcription factor gene family involved in the regulation of gypenoside biosynthesis in Gynostemma pentaphyllum Plant Physiol Biochem 154, 238-247. DOI:10.1016/j.plaphy.2020.05.040URL [本文引用: 1]
ZhangRF, FengX, SuGZ, MuZJ, ZhangHX, ZhaoYN, JiaoSG, CaoL, ChenSYL, TuPF, ChaiXF (2018). Bioactive sesquiterpenoids from the peeled stems of Syringa pinnatifolia J Nat Prod 81, 1711-1720. DOI:10.1021/acs.jnatprod.7b01071URL [本文引用: 1]
... 丰富的次生代谢产物(secondary metabolites)是植物适应复杂环境的策略(Thakur et al., 2019), 也可能在一定条件下带有系统演化的痕迹(Sedio, 2017).植物在生长过程中受到昆虫、微生物和伴生物种等生物因素, 以及温度、光照和水分等非生物因素的影响(杜玮炜和黄宏文, 2008; Wang et al., 2019), 伴随着复杂的时空变化形成了富于变化的次生代谢成分.人们大多认为, 相对于次生代谢产物与环境的紧密联系, 它们与系统学的关联则较为松散(Chen et al., 2020), 与谱系演化的关联具有很大的不确定性(Ernst et al., 2016; Allevato et al., 2019).尽管如此, 仍有****发现在相距2 500 km的秘鲁伊基托斯和巴西马瑙斯保护区中, 经历了海拔、经纬度和土壤环境的巨大变化, 相同物种的种群却共享着超过95%的次生代谢产物, 并且种内化学成分保持极高的一致性(Vleminckx et al., 2018).这意味着在千变万化的环境诱导着丰富的化学多样性背后, 系统保守性仍有迹可循.当今, 人们已经利用快速发展的基因组学和代谢组学工具, 揭示了科以上的分类等级, 以及被子植物门次生代谢关键调控因子家族的演化扩张关系(Chen et al., 2011; 祝志欣和鲁迎青, 2016; 张永增, 2018; Mint Evolutionary Genomics Consortium, 2018; Xu et al., 2020).这表明在较低的分类等级上, 次生代谢或其成分在属内的分布保守性可能同样有章可循(Rønsted et al., 2008; Ernst et al., 2016; Guitton et al., 2018; Chen et al., 2020).尽管由于很难获得具有系统学代表性的属下植物材料而使实证研究较为少见, 但这类研究对于精准而系统地确定属下水平的代谢调控机制具有科学意义, 对功能性成分的系统性挖掘具有现实意义. ...
植物根向水性反应研究进展 2 2018
... 在分别从系统演化和地理环境角度讨论丁香属次生代谢产物的分布趋势之后, 需要面对的问题还有代谢背景下的系统演化与地理分布跨度的关系.研究表明, 紫丁香和欧洲丁香是丁香属系统学原始组系中的2个近缘姊妹系物种(Li et al., 2012), 它们也分别是东亚和欧洲的广布种(张美珍等, 1992; Fiala, 2008), 而后续分化的红丁香系(Li et al., 2012)中的大多数物种都属于生态幅狭窄的局域种.已知温度和水分是限制物种扩散的主要环境因子(高坤等, 2018; 魏华等, 2018), 丁香地理分布范围的差异一定程度上由对温度和水分敏感的光合同化产物形成和水分利用效率等初生代谢机制的差别决定(Cui et al., 2016).紫丁香和欧洲丁香的广布表明其初生代谢机制保证了它们能够适应东亚和东南欧洲的气候变化, 同时强大的系统学保守性使相距万里的2个系统学意义上的近缘姊妹种均具有环/裂环烯醚萜和苯丙素类产物(表1, 表3), 使它们有能力通过这两类优势次生代谢产物的形成, 增强对小环境中生物及非生物扰动的适应能力, 从而改善植株的生长状况并实现高效的繁衍生息(Konno et al., 1999; 刘盟盟等, 2017; Sharma et al., 2019).而对于性喜冷凉的红丁香系中的红丁香和匈牙利丁香2个近缘种, 气孔较大且密度较低的叶片表皮特征很可能使其在初生代谢热量耗散和叶片水分保持能力上存在限制(高艳等, 2008), 因此其分布局限在高海拔的冷凉或邻水生境(张美珍等, 1992; Lendvay et al., 2016)而成为地理分布上的局域种.在这类水热条件适宜的小环境中, 这些较晚分化的局域种不必应对地域扩散中温度和水分的巨大变化, 有条件形成更加多样的次生代谢产物, 在适宜的生境中最大限度地改善生长质量.这种“初生代谢决定生存格局, 次生代谢改善生活质量”(刘晓侠等, 2015)的情况也发生在同样进化却局域分布的巧玲花系中(图1). ...
... 极端环境压力可能会使特定的代谢路径延长.木脂素是丁香属中独具特色的成分, 现有文献显示其在羽叶丁香系和短花冠管组中占据优势.在单种系的羽叶丁香中木脂素占比高达46.40%, 这与其生长的极端干旱环境紧密相关.羽叶丁香生长在贺兰山东、西山麓以及陕西南部、甘肃和青海, 该地区原生境大多降水稀少, 巨大的水分胁迫压力使羽叶丁香的地下生物量投入极大, 以粗壮的根系实现土壤深层水分的向性生长(Cui et al., 2016; 高坤等, 2018).莽草酸途径化合物的积累通常有助于增强植物对水分胁迫的耐受性(刘盟盟等, 2017; Sharma et al., 2019), 而该途径中形成的木脂素可在木质部木质化后成为一种后注入型树脂, 使植物在水分胁迫而韧皮部运输减少的情况下更具干旱耐受性(Gaylord et al., 2013; Zhang et al., 2018).这种被极端干旱强化的木脂素代谢途径是以牺牲莽草酸途径中大量上游产物苯丙素为代价以保证下游产物木脂素的产生, 助力粗壮根系的形成以实现土壤深层的趋向性生长, 从而最大程度地吸收生存所需水分.与此同时, 吸引传粉者也是其需要兼顾的因素, 在1 700-3 100 m的高山生境中, 传粉昆虫比低海拔地区更为稀少, 这使得羽叶丁香在萜类途径中更多地利用底物焦磷酸二甲烯丙酯和焦磷酸烯丙酯向法尼基焦磷酸合成方向延伸, 成为丁香属以倍半萜为优势萜类成分的组系.利用较短途径获得的大量倍半萜可在一定程度上实现对传粉昆虫的诱集(Chadwick et al., 2013). ...
丁香属植物叶片表皮形态特征与环境适应及系统学关联 1 2008
... 在分别从系统演化和地理环境角度讨论丁香属次生代谢产物的分布趋势之后, 需要面对的问题还有代谢背景下的系统演化与地理分布跨度的关系.研究表明, 紫丁香和欧洲丁香是丁香属系统学原始组系中的2个近缘姊妹系物种(Li et al., 2012), 它们也分别是东亚和欧洲的广布种(张美珍等, 1992; Fiala, 2008), 而后续分化的红丁香系(Li et al., 2012)中的大多数物种都属于生态幅狭窄的局域种.已知温度和水分是限制物种扩散的主要环境因子(高坤等, 2018; 魏华等, 2018), 丁香地理分布范围的差异一定程度上由对温度和水分敏感的光合同化产物形成和水分利用效率等初生代谢机制的差别决定(Cui et al., 2016).紫丁香和欧洲丁香的广布表明其初生代谢机制保证了它们能够适应东亚和东南欧洲的气候变化, 同时强大的系统学保守性使相距万里的2个系统学意义上的近缘姊妹种均具有环/裂环烯醚萜和苯丙素类产物(表1, 表3), 使它们有能力通过这两类优势次生代谢产物的形成, 增强对小环境中生物及非生物扰动的适应能力, 从而改善植株的生长状况并实现高效的繁衍生息(Konno et al., 1999; 刘盟盟等, 2017; Sharma et al., 2019).而对于性喜冷凉的红丁香系中的红丁香和匈牙利丁香2个近缘种, 气孔较大且密度较低的叶片表皮特征很可能使其在初生代谢热量耗散和叶片水分保持能力上存在限制(高艳等, 2008), 因此其分布局限在高海拔的冷凉或邻水生境(张美珍等, 1992; Lendvay et al., 2016)而成为地理分布上的局域种.在这类水热条件适宜的小环境中, 这些较晚分化的局域种不必应对地域扩散中温度和水分的巨大变化, 有条件形成更加多样的次生代谢产物, 在适宜的生境中最大限度地改善生长质量.这种“初生代谢决定生存格局, 次生代谢改善生活质量”(刘晓侠等, 2015)的情况也发生在同样进化却局域分布的巧玲花系中(图1). ...
天然木脂素的代谢工程和合成生物学研究进展 2 2016
... (A) 丁香属组系的系统演化关系(Li et al., 2012)及各类产物在各组系中的占比; (B) 甲戊二羟酸途径(MVA)和脱氧木酮糖磷酸酯途径(DXP) (Mint Evolutionary Genomics Consortium, 2018); (C) 莽草酸途径(刘津等, 2016; 张旭等, 2019).图(A)组系下方的色条表示各类产物(颜色含义同图1), 色条内的百分数指特定类别产物计数在该组系内成分计数总和中所占的百分比(详见图1图注).图(B)和图(C)中化合物色框颜色含义与图(A)相同.图(B)和图(C)中虚线表示由多步反应完成. ...
... (A) The phylogenetic relationship (Li et al., 2012) and percentage of metabolites of Syringa at the level of series (section); (B) Mevalonic acid pathway (MVA) and Deoxyxylulose-5-P pathway (DXP) (Mint Evolutionary Genomics Consortium, 2018); (C) Shikimic acid pathway (Liu et al., 2016; Zhang et al., 2019, in Chinese). The color-stripes below the series (section) indicate different types of metabolites in (A), and the percentage in the color-stripe represents the percentage of a given type of compounds within the series (section) (please see Figure 1 for details). The meanings of the color frames indicating the compound in (B) and (C) are the same as those of (A). The dotted lines in (B) and (C) represent the process of multi-step reactions. ...
冷蒿酚酸及其抗氧化防御酶活性对机械损伤的响应 2 2017
... 在分别从系统演化和地理环境角度讨论丁香属次生代谢产物的分布趋势之后, 需要面对的问题还有代谢背景下的系统演化与地理分布跨度的关系.研究表明, 紫丁香和欧洲丁香是丁香属系统学原始组系中的2个近缘姊妹系物种(Li et al., 2012), 它们也分别是东亚和欧洲的广布种(张美珍等, 1992; Fiala, 2008), 而后续分化的红丁香系(Li et al., 2012)中的大多数物种都属于生态幅狭窄的局域种.已知温度和水分是限制物种扩散的主要环境因子(高坤等, 2018; 魏华等, 2018), 丁香地理分布范围的差异一定程度上由对温度和水分敏感的光合同化产物形成和水分利用效率等初生代谢机制的差别决定(Cui et al., 2016).紫丁香和欧洲丁香的广布表明其初生代谢机制保证了它们能够适应东亚和东南欧洲的气候变化, 同时强大的系统学保守性使相距万里的2个系统学意义上的近缘姊妹种均具有环/裂环烯醚萜和苯丙素类产物(表1, 表3), 使它们有能力通过这两类优势次生代谢产物的形成, 增强对小环境中生物及非生物扰动的适应能力, 从而改善植株的生长状况并实现高效的繁衍生息(Konno et al., 1999; 刘盟盟等, 2017; Sharma et al., 2019).而对于性喜冷凉的红丁香系中的红丁香和匈牙利丁香2个近缘种, 气孔较大且密度较低的叶片表皮特征很可能使其在初生代谢热量耗散和叶片水分保持能力上存在限制(高艳等, 2008), 因此其分布局限在高海拔的冷凉或邻水生境(张美珍等, 1992; Lendvay et al., 2016)而成为地理分布上的局域种.在这类水热条件适宜的小环境中, 这些较晚分化的局域种不必应对地域扩散中温度和水分的巨大变化, 有条件形成更加多样的次生代谢产物, 在适宜的生境中最大限度地改善生长质量.这种“初生代谢决定生存格局, 次生代谢改善生活质量”(刘晓侠等, 2015)的情况也发生在同样进化却局域分布的巧玲花系中(图1). ...
... 极端环境压力可能会使特定的代谢路径延长.木脂素是丁香属中独具特色的成分, 现有文献显示其在羽叶丁香系和短花冠管组中占据优势.在单种系的羽叶丁香中木脂素占比高达46.40%, 这与其生长的极端干旱环境紧密相关.羽叶丁香生长在贺兰山东、西山麓以及陕西南部、甘肃和青海, 该地区原生境大多降水稀少, 巨大的水分胁迫压力使羽叶丁香的地下生物量投入极大, 以粗壮的根系实现土壤深层水分的向性生长(Cui et al., 2016; 高坤等, 2018).莽草酸途径化合物的积累通常有助于增强植物对水分胁迫的耐受性(刘盟盟等, 2017; Sharma et al., 2019), 而该途径中形成的木脂素可在木质部木质化后成为一种后注入型树脂, 使植物在水分胁迫而韧皮部运输减少的情况下更具干旱耐受性(Gaylord et al., 2013; Zhang et al., 2018).这种被极端干旱强化的木脂素代谢途径是以牺牲莽草酸途径中大量上游产物苯丙素为代价以保证下游产物木脂素的产生, 助力粗壮根系的形成以实现土壤深层的趋向性生长, 从而最大程度地吸收生存所需水分.与此同时, 吸引传粉者也是其需要兼顾的因素, 在1 700-3 100 m的高山生境中, 传粉昆虫比低海拔地区更为稀少, 这使得羽叶丁香在萜类途径中更多地利用底物焦磷酸二甲烯丙酯和焦磷酸烯丙酯向法尼基焦磷酸合成方向延伸, 成为丁香属以倍半萜为优势萜类成分的组系.利用较短途径获得的大量倍半萜可在一定程度上实现对传粉昆虫的诱集(Chadwick et al., 2013). ...
不同地域枸杞主要次生代谢物含量与初生代谢物含量的关系研究 1 2015
... 在分别从系统演化和地理环境角度讨论丁香属次生代谢产物的分布趋势之后, 需要面对的问题还有代谢背景下的系统演化与地理分布跨度的关系.研究表明, 紫丁香和欧洲丁香是丁香属系统学原始组系中的2个近缘姊妹系物种(Li et al., 2012), 它们也分别是东亚和欧洲的广布种(张美珍等, 1992; Fiala, 2008), 而后续分化的红丁香系(Li et al., 2012)中的大多数物种都属于生态幅狭窄的局域种.已知温度和水分是限制物种扩散的主要环境因子(高坤等, 2018; 魏华等, 2018), 丁香地理分布范围的差异一定程度上由对温度和水分敏感的光合同化产物形成和水分利用效率等初生代谢机制的差别决定(Cui et al., 2016).紫丁香和欧洲丁香的广布表明其初生代谢机制保证了它们能够适应东亚和东南欧洲的气候变化, 同时强大的系统学保守性使相距万里的2个系统学意义上的近缘姊妹种均具有环/裂环烯醚萜和苯丙素类产物(表1, 表3), 使它们有能力通过这两类优势次生代谢产物的形成, 增强对小环境中生物及非生物扰动的适应能力, 从而改善植株的生长状况并实现高效的繁衍生息(Konno et al., 1999; 刘盟盟等, 2017; Sharma et al., 2019).而对于性喜冷凉的红丁香系中的红丁香和匈牙利丁香2个近缘种, 气孔较大且密度较低的叶片表皮特征很可能使其在初生代谢热量耗散和叶片水分保持能力上存在限制(高艳等, 2008), 因此其分布局限在高海拔的冷凉或邻水生境(张美珍等, 1992; Lendvay et al., 2016)而成为地理分布上的局域种.在这类水热条件适宜的小环境中, 这些较晚分化的局域种不必应对地域扩散中温度和水分的巨大变化, 有条件形成更加多样的次生代谢产物, 在适宜的生境中最大限度地改善生长质量.这种“初生代谢决定生存格局, 次生代谢改善生活质量”(刘晓侠等, 2015)的情况也发生在同样进化却局域分布的巧玲花系中(图1). ...
... 对于丁香属内次生代谢产物的讨论离不开物种分类和演化的系统学背景.从经典分类以及既往分子生物学的研究结果看, 丁香属系统演化关系基本明确, 其结果差异主要在于在先分化类群的不同(张美珍等, 1992; Kim and Jansen, 1998; Li et al., 2012).因此,我们依据欧洲丁香与紫丁香存在欧亚间断分布的事实(Kim and Jansen, 1998), 以及核DNA分析在物种演化关系判定上的依据(Li et al., 2012), 支持灌木型欧丁香系是丁香属最先分化的类群.羽叶丁香系在欧丁香系之后独立分化, 乔木型短花冠管组位居灌木型组系之间为中间分化类群(Li et al., 2012), 而对于巧玲花系和红丁香系是丁香属内后续分化类群的观点则不存异议.据此, 我们在进行系统关联讨论时采用Li等(2012)所报道的系统演化模式. ...
... 在分别从系统演化和地理环境角度讨论丁香属次生代谢产物的分布趋势之后, 需要面对的问题还有代谢背景下的系统演化与地理分布跨度的关系.研究表明, 紫丁香和欧洲丁香是丁香属系统学原始组系中的2个近缘姊妹系物种(Li et al., 2012), 它们也分别是东亚和欧洲的广布种(张美珍等, 1992; Fiala, 2008), 而后续分化的红丁香系(Li et al., 2012)中的大多数物种都属于生态幅狭窄的局域种.已知温度和水分是限制物种扩散的主要环境因子(高坤等, 2018; 魏华等, 2018), 丁香地理分布范围的差异一定程度上由对温度和水分敏感的光合同化产物形成和水分利用效率等初生代谢机制的差别决定(Cui et al., 2016).紫丁香和欧洲丁香的广布表明其初生代谢机制保证了它们能够适应东亚和东南欧洲的气候变化, 同时强大的系统学保守性使相距万里的2个系统学意义上的近缘姊妹种均具有环/裂环烯醚萜和苯丙素类产物(表1, 表3), 使它们有能力通过这两类优势次生代谢产物的形成, 增强对小环境中生物及非生物扰动的适应能力, 从而改善植株的生长状况并实现高效的繁衍生息(Konno et al., 1999; 刘盟盟等, 2017; Sharma et al., 2019).而对于性喜冷凉的红丁香系中的红丁香和匈牙利丁香2个近缘种, 气孔较大且密度较低的叶片表皮特征很可能使其在初生代谢热量耗散和叶片水分保持能力上存在限制(高艳等, 2008), 因此其分布局限在高海拔的冷凉或邻水生境(张美珍等, 1992; Lendvay et al., 2016)而成为地理分布上的局域种.在这类水热条件适宜的小环境中, 这些较晚分化的局域种不必应对地域扩散中温度和水分的巨大变化, 有条件形成更加多样的次生代谢产物, 在适宜的生境中最大限度地改善生长质量.这种“初生代谢决定生存格局, 次生代谢改善生活质量”(刘晓侠等, 2015)的情况也发生在同样进化却局域分布的巧玲花系中(图1). ...
... (A) 丁香属组系的系统演化关系(Li et al., 2012)及各类产物在各组系中的占比; (B) 甲戊二羟酸途径(MVA)和脱氧木酮糖磷酸酯途径(DXP) (Mint Evolutionary Genomics Consortium, 2018); (C) 莽草酸途径(刘津等, 2016; 张旭等, 2019).图(A)组系下方的色条表示各类产物(颜色含义同图1), 色条内的百分数指特定类别产物计数在该组系内成分计数总和中所占的百分比(详见图1图注).图(B)和图(C)中化合物色框颜色含义与图(A)相同.图(B)和图(C)中虚线表示由多步反应完成. ...
... (A) The phylogenetic relationship (Li et al., 2012) and percentage of metabolites of Syringa at the level of series (section); (B) Mevalonic acid pathway (MVA) and Deoxyxylulose-5-P pathway (DXP) (Mint Evolutionary Genomics Consortium, 2018); (C) Shikimic acid pathway (Liu et al., 2016; Zhang et al., 2019, in Chinese). The color-stripes below the series (section) indicate different types of metabolites in (A), and the percentage in the color-stripe represents the percentage of a given type of compounds within the series (section) (please see Figure 1 for details). The meanings of the color frames indicating the compound in (B) and (C) are the same as those of (A). The dotted lines in (B) and (C) represent the process of multi-step reactions. ...
次生代谢产物在种子植物生命之树中的分布规律 2 2018
... 丰富的次生代谢产物(secondary metabolites)是植物适应复杂环境的策略(Thakur et al., 2019), 也可能在一定条件下带有系统演化的痕迹(Sedio, 2017).植物在生长过程中受到昆虫、微生物和伴生物种等生物因素, 以及温度、光照和水分等非生物因素的影响(杜玮炜和黄宏文, 2008; Wang et al., 2019), 伴随着复杂的时空变化形成了富于变化的次生代谢成分.人们大多认为, 相对于次生代谢产物与环境的紧密联系, 它们与系统学的关联则较为松散(Chen et al., 2020), 与谱系演化的关联具有很大的不确定性(Ernst et al., 2016; Allevato et al., 2019).尽管如此, 仍有****发现在相距2 500 km的秘鲁伊基托斯和巴西马瑙斯保护区中, 经历了海拔、经纬度和土壤环境的巨大变化, 相同物种的种群却共享着超过95%的次生代谢产物, 并且种内化学成分保持极高的一致性(Vleminckx et al., 2018).这意味着在千变万化的环境诱导着丰富的化学多样性背后, 系统保守性仍有迹可循.当今, 人们已经利用快速发展的基因组学和代谢组学工具, 揭示了科以上的分类等级, 以及被子植物门次生代谢关键调控因子家族的演化扩张关系(Chen et al., 2011; 祝志欣和鲁迎青, 2016; 张永增, 2018; Mint Evolutionary Genomics Consortium, 2018; Xu et al., 2020).这表明在较低的分类等级上, 次生代谢或其成分在属内的分布保守性可能同样有章可循(Rønsted et al., 2008; Ernst et al., 2016; Guitton et al., 2018; Chen et al., 2020).尽管由于很难获得具有系统学代表性的属下植物材料而使实证研究较为少见, 但这类研究对于精准而系统地确定属下水平的代谢调控机制具有科学意义, 对功能性成分的系统性挖掘具有现实意义. ...
... 丰富的次生代谢产物(secondary metabolites)是植物适应复杂环境的策略(Thakur et al., 2019), 也可能在一定条件下带有系统演化的痕迹(Sedio, 2017).植物在生长过程中受到昆虫、微生物和伴生物种等生物因素, 以及温度、光照和水分等非生物因素的影响(杜玮炜和黄宏文, 2008; Wang et al., 2019), 伴随着复杂的时空变化形成了富于变化的次生代谢成分.人们大多认为, 相对于次生代谢产物与环境的紧密联系, 它们与系统学的关联则较为松散(Chen et al., 2020), 与谱系演化的关联具有很大的不确定性(Ernst et al., 2016; Allevato et al., 2019).尽管如此, 仍有****发现在相距2 500 km的秘鲁伊基托斯和巴西马瑙斯保护区中, 经历了海拔、经纬度和土壤环境的巨大变化, 相同物种的种群却共享着超过95%的次生代谢产物, 并且种内化学成分保持极高的一致性(Vleminckx et al., 2018).这意味着在千变万化的环境诱导着丰富的化学多样性背后, 系统保守性仍有迹可循.当今, 人们已经利用快速发展的基因组学和代谢组学工具, 揭示了科以上的分类等级, 以及被子植物门次生代谢关键调控因子家族的演化扩张关系(Chen et al., 2011; 祝志欣和鲁迎青, 2016; 张永增, 2018; Mint Evolutionary Genomics Consortium, 2018; Xu et al., 2020).这表明在较低的分类等级上, 次生代谢或其成分在属内的分布保守性可能同样有章可循(Rønsted et al., 2008; Ernst et al., 2016; Guitton et al., 2018; Chen et al., 2020).尽管由于很难获得具有系统学代表性的属下植物材料而使实证研究较为少见, 但这类研究对于精准而系统地确定属下水平的代谢调控机制具有科学意义, 对功能性成分的系统性挖掘具有现实意义. ...
Evolution of phytochemical diversity in Pilocarpus (Rutaceae) 1 2019
... 丰富的次生代谢产物(secondary metabolites)是植物适应复杂环境的策略(Thakur et al., 2019), 也可能在一定条件下带有系统演化的痕迹(Sedio, 2017).植物在生长过程中受到昆虫、微生物和伴生物种等生物因素, 以及温度、光照和水分等非生物因素的影响(杜玮炜和黄宏文, 2008; Wang et al., 2019), 伴随着复杂的时空变化形成了富于变化的次生代谢成分.人们大多认为, 相对于次生代谢产物与环境的紧密联系, 它们与系统学的关联则较为松散(Chen et al., 2020), 与谱系演化的关联具有很大的不确定性(Ernst et al., 2016; Allevato et al., 2019).尽管如此, 仍有****发现在相距2 500 km的秘鲁伊基托斯和巴西马瑙斯保护区中, 经历了海拔、经纬度和土壤环境的巨大变化, 相同物种的种群却共享着超过95%的次生代谢产物, 并且种内化学成分保持极高的一致性(Vleminckx et al., 2018).这意味着在千变万化的环境诱导着丰富的化学多样性背后, 系统保守性仍有迹可循.当今, 人们已经利用快速发展的基因组学和代谢组学工具, 揭示了科以上的分类等级, 以及被子植物门次生代谢关键调控因子家族的演化扩张关系(Chen et al., 2011; 祝志欣和鲁迎青, 2016; 张永增, 2018; Mint Evolutionary Genomics Consortium, 2018; Xu et al., 2020).这表明在较低的分类等级上, 次生代谢或其成分在属内的分布保守性可能同样有章可循(Rønsted et al., 2008; Ernst et al., 2016; Guitton et al., 2018; Chen et al., 2020).尽管由于很难获得具有系统学代表性的属下植物材料而使实证研究较为少见, 但这类研究对于精准而系统地确定属下水平的代谢调控机制具有科学意义, 对功能性成分的系统性挖掘具有现实意义. ...
Sesquiterpenoids lactones: benefits to plants and people 1 2013
... 极端环境压力可能会使特定的代谢路径延长.木脂素是丁香属中独具特色的成分, 现有文献显示其在羽叶丁香系和短花冠管组中占据优势.在单种系的羽叶丁香中木脂素占比高达46.40%, 这与其生长的极端干旱环境紧密相关.羽叶丁香生长在贺兰山东、西山麓以及陕西南部、甘肃和青海, 该地区原生境大多降水稀少, 巨大的水分胁迫压力使羽叶丁香的地下生物量投入极大, 以粗壮的根系实现土壤深层水分的向性生长(Cui et al., 2016; 高坤等, 2018).莽草酸途径化合物的积累通常有助于增强植物对水分胁迫的耐受性(刘盟盟等, 2017; Sharma et al., 2019), 而该途径中形成的木脂素可在木质部木质化后成为一种后注入型树脂, 使植物在水分胁迫而韧皮部运输减少的情况下更具干旱耐受性(Gaylord et al., 2013; Zhang et al., 2018).这种被极端干旱强化的木脂素代谢途径是以牺牲莽草酸途径中大量上游产物苯丙素为代价以保证下游产物木脂素的产生, 助力粗壮根系的形成以实现土壤深层的趋向性生长, 从而最大程度地吸收生存所需水分.与此同时, 吸引传粉者也是其需要兼顾的因素, 在1 700-3 100 m的高山生境中, 传粉昆虫比低海拔地区更为稀少, 这使得羽叶丁香在萜类途径中更多地利用底物焦磷酸二甲烯丙酯和焦磷酸烯丙酯向法尼基焦磷酸合成方向延伸, 成为丁香属以倍半萜为优势萜类成分的组系.利用较短途径获得的大量倍半萜可在一定程度上实现对传粉昆虫的诱集(Chadwick et al., 2013). ...
The family of terpene synthases in plants: a mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom 1 2011
... 丰富的次生代谢产物(secondary metabolites)是植物适应复杂环境的策略(Thakur et al., 2019), 也可能在一定条件下带有系统演化的痕迹(Sedio, 2017).植物在生长过程中受到昆虫、微生物和伴生物种等生物因素, 以及温度、光照和水分等非生物因素的影响(杜玮炜和黄宏文, 2008; Wang et al., 2019), 伴随着复杂的时空变化形成了富于变化的次生代谢成分.人们大多认为, 相对于次生代谢产物与环境的紧密联系, 它们与系统学的关联则较为松散(Chen et al., 2020), 与谱系演化的关联具有很大的不确定性(Ernst et al., 2016; Allevato et al., 2019).尽管如此, 仍有****发现在相距2 500 km的秘鲁伊基托斯和巴西马瑙斯保护区中, 经历了海拔、经纬度和土壤环境的巨大变化, 相同物种的种群却共享着超过95%的次生代谢产物, 并且种内化学成分保持极高的一致性(Vleminckx et al., 2018).这意味着在千变万化的环境诱导着丰富的化学多样性背后, 系统保守性仍有迹可循.当今, 人们已经利用快速发展的基因组学和代谢组学工具, 揭示了科以上的分类等级, 以及被子植物门次生代谢关键调控因子家族的演化扩张关系(Chen et al., 2011; 祝志欣和鲁迎青, 2016; 张永增, 2018; Mint Evolutionary Genomics Consortium, 2018; Xu et al., 2020).这表明在较低的分类等级上, 次生代谢或其成分在属内的分布保守性可能同样有章可循(Rønsted et al., 2008; Ernst et al., 2016; Guitton et al., 2018; Chen et al., 2020).尽管由于很难获得具有系统学代表性的属下植物材料而使实证研究较为少见, 但这类研究对于精准而系统地确定属下水平的代谢调控机制具有科学意义, 对功能性成分的系统性挖掘具有现实意义. ...
The evolution of pyrrolizidine alkaloid diversity among and within Jacobaea species 2 2020
... 丰富的次生代谢产物(secondary metabolites)是植物适应复杂环境的策略(Thakur et al., 2019), 也可能在一定条件下带有系统演化的痕迹(Sedio, 2017).植物在生长过程中受到昆虫、微生物和伴生物种等生物因素, 以及温度、光照和水分等非生物因素的影响(杜玮炜和黄宏文, 2008; Wang et al., 2019), 伴随着复杂的时空变化形成了富于变化的次生代谢成分.人们大多认为, 相对于次生代谢产物与环境的紧密联系, 它们与系统学的关联则较为松散(Chen et al., 2020), 与谱系演化的关联具有很大的不确定性(Ernst et al., 2016; Allevato et al., 2019).尽管如此, 仍有****发现在相距2 500 km的秘鲁伊基托斯和巴西马瑙斯保护区中, 经历了海拔、经纬度和土壤环境的巨大变化, 相同物种的种群却共享着超过95%的次生代谢产物, 并且种内化学成分保持极高的一致性(Vleminckx et al., 2018).这意味着在千变万化的环境诱导着丰富的化学多样性背后, 系统保守性仍有迹可循.当今, 人们已经利用快速发展的基因组学和代谢组学工具, 揭示了科以上的分类等级, 以及被子植物门次生代谢关键调控因子家族的演化扩张关系(Chen et al., 2011; 祝志欣和鲁迎青, 2016; 张永增, 2018; Mint Evolutionary Genomics Consortium, 2018; Xu et al., 2020).这表明在较低的分类等级上, 次生代谢或其成分在属内的分布保守性可能同样有章可循(Rønsted et al., 2008; Ernst et al., 2016; Guitton et al., 2018; Chen et al., 2020).尽管由于很难获得具有系统学代表性的属下植物材料而使实证研究较为少见, 但这类研究对于精准而系统地确定属下水平的代谢调控机制具有科学意义, 对功能性成分的系统性挖掘具有现实意义. ...
... ; Chen et al., 2020).尽管由于很难获得具有系统学代表性的属下植物材料而使实证研究较为少见, 但这类研究对于精准而系统地确定属下水平的代谢调控机制具有科学意义, 对功能性成分的系统性挖掘具有现实意义. ...
Mechanistic examination of causes for narrow distribution in an endangered shrub: a comparison of its responses to drought stress with a widespread congeneric species 2 2016
... 在分别从系统演化和地理环境角度讨论丁香属次生代谢产物的分布趋势之后, 需要面对的问题还有代谢背景下的系统演化与地理分布跨度的关系.研究表明, 紫丁香和欧洲丁香是丁香属系统学原始组系中的2个近缘姊妹系物种(Li et al., 2012), 它们也分别是东亚和欧洲的广布种(张美珍等, 1992; Fiala, 2008), 而后续分化的红丁香系(Li et al., 2012)中的大多数物种都属于生态幅狭窄的局域种.已知温度和水分是限制物种扩散的主要环境因子(高坤等, 2018; 魏华等, 2018), 丁香地理分布范围的差异一定程度上由对温度和水分敏感的光合同化产物形成和水分利用效率等初生代谢机制的差别决定(Cui et al., 2016).紫丁香和欧洲丁香的广布表明其初生代谢机制保证了它们能够适应东亚和东南欧洲的气候变化, 同时强大的系统学保守性使相距万里的2个系统学意义上的近缘姊妹种均具有环/裂环烯醚萜和苯丙素类产物(表1, 表3), 使它们有能力通过这两类优势次生代谢产物的形成, 增强对小环境中生物及非生物扰动的适应能力, 从而改善植株的生长状况并实现高效的繁衍生息(Konno et al., 1999; 刘盟盟等, 2017; Sharma et al., 2019).而对于性喜冷凉的红丁香系中的红丁香和匈牙利丁香2个近缘种, 气孔较大且密度较低的叶片表皮特征很可能使其在初生代谢热量耗散和叶片水分保持能力上存在限制(高艳等, 2008), 因此其分布局限在高海拔的冷凉或邻水生境(张美珍等, 1992; Lendvay et al., 2016)而成为地理分布上的局域种.在这类水热条件适宜的小环境中, 这些较晚分化的局域种不必应对地域扩散中温度和水分的巨大变化, 有条件形成更加多样的次生代谢产物, 在适宜的生境中最大限度地改善生长质量.这种“初生代谢决定生存格局, 次生代谢改善生活质量”(刘晓侠等, 2015)的情况也发生在同样进化却局域分布的巧玲花系中(图1). ...
... 极端环境压力可能会使特定的代谢路径延长.木脂素是丁香属中独具特色的成分, 现有文献显示其在羽叶丁香系和短花冠管组中占据优势.在单种系的羽叶丁香中木脂素占比高达46.40%, 这与其生长的极端干旱环境紧密相关.羽叶丁香生长在贺兰山东、西山麓以及陕西南部、甘肃和青海, 该地区原生境大多降水稀少, 巨大的水分胁迫压力使羽叶丁香的地下生物量投入极大, 以粗壮的根系实现土壤深层水分的向性生长(Cui et al., 2016; 高坤等, 2018).莽草酸途径化合物的积累通常有助于增强植物对水分胁迫的耐受性(刘盟盟等, 2017; Sharma et al., 2019), 而该途径中形成的木脂素可在木质部木质化后成为一种后注入型树脂, 使植物在水分胁迫而韧皮部运输减少的情况下更具干旱耐受性(Gaylord et al., 2013; Zhang et al., 2018).这种被极端干旱强化的木脂素代谢途径是以牺牲莽草酸途径中大量上游产物苯丙素为代价以保证下游产物木脂素的产生, 助力粗壮根系的形成以实现土壤深层的趋向性生长, 从而最大程度地吸收生存所需水分.与此同时, 吸引传粉者也是其需要兼顾的因素, 在1 700-3 100 m的高山生境中, 传粉昆虫比低海拔地区更为稀少, 这使得羽叶丁香在萜类途径中更多地利用底物焦磷酸二甲烯丙酯和焦磷酸烯丙酯向法尼基焦磷酸合成方向延伸, 成为丁香属以倍半萜为优势萜类成分的组系.利用较短途径获得的大量倍半萜可在一定程度上实现对传粉昆虫的诱集(Chadwick et al., 2013). ...
Evolutionary prediction of medicinal properties in the genus Euphorbia L 2 2016
... 丰富的次生代谢产物(secondary metabolites)是植物适应复杂环境的策略(Thakur et al., 2019), 也可能在一定条件下带有系统演化的痕迹(Sedio, 2017).植物在生长过程中受到昆虫、微生物和伴生物种等生物因素, 以及温度、光照和水分等非生物因素的影响(杜玮炜和黄宏文, 2008; Wang et al., 2019), 伴随着复杂的时空变化形成了富于变化的次生代谢成分.人们大多认为, 相对于次生代谢产物与环境的紧密联系, 它们与系统学的关联则较为松散(Chen et al., 2020), 与谱系演化的关联具有很大的不确定性(Ernst et al., 2016; Allevato et al., 2019).尽管如此, 仍有****发现在相距2 500 km的秘鲁伊基托斯和巴西马瑙斯保护区中, 经历了海拔、经纬度和土壤环境的巨大变化, 相同物种的种群却共享着超过95%的次生代谢产物, 并且种内化学成分保持极高的一致性(Vleminckx et al., 2018).这意味着在千变万化的环境诱导着丰富的化学多样性背后, 系统保守性仍有迹可循.当今, 人们已经利用快速发展的基因组学和代谢组学工具, 揭示了科以上的分类等级, 以及被子植物门次生代谢关键调控因子家族的演化扩张关系(Chen et al., 2011; 祝志欣和鲁迎青, 2016; 张永增, 2018; Mint Evolutionary Genomics Consortium, 2018; Xu et al., 2020).这表明在较低的分类等级上, 次生代谢或其成分在属内的分布保守性可能同样有章可循(Rønsted et al., 2008; Ernst et al., 2016; Guitton et al., 2018; Chen et al., 2020).尽管由于很难获得具有系统学代表性的属下植物材料而使实证研究较为少见, 但这类研究对于精准而系统地确定属下水平的代谢调控机制具有科学意义, 对功能性成分的系统性挖掘具有现实意义. ...
... ; Ernst et al., 2016; Guitton et al., 2018; Chen et al., 2020).尽管由于很难获得具有系统学代表性的属下植物材料而使实证研究较为少见, 但这类研究对于精准而系统地确定属下水平的代谢调控机制具有科学意义, 对功能性成分的系统性挖掘具有现实意义. ...
1 2008
... 在分别从系统演化和地理环境角度讨论丁香属次生代谢产物的分布趋势之后, 需要面对的问题还有代谢背景下的系统演化与地理分布跨度的关系.研究表明, 紫丁香和欧洲丁香是丁香属系统学原始组系中的2个近缘姊妹系物种(Li et al., 2012), 它们也分别是东亚和欧洲的广布种(张美珍等, 1992; Fiala, 2008), 而后续分化的红丁香系(Li et al., 2012)中的大多数物种都属于生态幅狭窄的局域种.已知温度和水分是限制物种扩散的主要环境因子(高坤等, 2018; 魏华等, 2018), 丁香地理分布范围的差异一定程度上由对温度和水分敏感的光合同化产物形成和水分利用效率等初生代谢机制的差别决定(Cui et al., 2016).紫丁香和欧洲丁香的广布表明其初生代谢机制保证了它们能够适应东亚和东南欧洲的气候变化, 同时强大的系统学保守性使相距万里的2个系统学意义上的近缘姊妹种均具有环/裂环烯醚萜和苯丙素类产物(表1, 表3), 使它们有能力通过这两类优势次生代谢产物的形成, 增强对小环境中生物及非生物扰动的适应能力, 从而改善植株的生长状况并实现高效的繁衍生息(Konno et al., 1999; 刘盟盟等, 2017; Sharma et al., 2019).而对于性喜冷凉的红丁香系中的红丁香和匈牙利丁香2个近缘种, 气孔较大且密度较低的叶片表皮特征很可能使其在初生代谢热量耗散和叶片水分保持能力上存在限制(高艳等, 2008), 因此其分布局限在高海拔的冷凉或邻水生境(张美珍等, 1992; Lendvay et al., 2016)而成为地理分布上的局域种.在这类水热条件适宜的小环境中, 这些较晚分化的局域种不必应对地域扩散中温度和水分的巨大变化, 有条件形成更加多样的次生代谢产物, 在适宜的生境中最大限度地改善生长质量.这种“初生代谢决定生存格局, 次生代谢改善生活质量”(刘晓侠等, 2015)的情况也发生在同样进化却局域分布的巧玲花系中(图1). ...
Syringa vulgaris bark as a source of compounds affecting the release of inflammatory mediators from human neutrophils and monocytes/macrophages 2 2019
... 丁香属中的环/裂环烯醚萜类和苯丙素类产物呈现较强的系统遗传保守性, 甚至遥远的地理隔离也未造成近缘种优势代谢产物的明显差异.紫丁香和欧洲丁香是系统学意义上的姊妹系(Li et al., 2012), 虽然二者在自然分布上存在着东亚与巴尔干半岛的遥远亚欧间断分布(表1), 却均具有丰富的环/裂环烯醚萜类和苯丙素类产物(表3).苯丙素类产物形成的保守性还体现在短花冠管组的2个近缘种暴马丁香和日本丁香中.暴马丁香广泛分布于中国北方及远东地区, 日本丁香仅在日本呈局域分布, 但2个系统学的近缘种同样具有占比相近的苯丙素类成分(表1, 表3).由此表明, 环/裂环烯醚萜类和苯丙素类成分的形成具有较强的遗传保守性, 强烈的保守性没有因遥远的地理距离而消弱.与此同时, 在相邻的近缘组系中出现了较高占比的特定类别化合物的连续分布(图1).图2A显示, 木脂素以较高的占比出现在相邻的羽叶丁香系(46.40%)和短花冠管组(18.18%)中; 倍半萜分别以36.80%、13.64%和27.78%的较高占比出现在相邻的羽叶丁香系、短花冠管组和巧玲花系中; 苯丙素以25.00%和22.22%的占比出现在相邻的短花冠管组和巧玲花系中; 环/裂环烯醚萜类分别以38.64%和25.00%的较高占比出现在相邻的短花冠管组和巧玲花系中.值得一提的是, 在针对同一个物种的诸多文献进行化合物汇总时, 我们发现欧洲丁香的样品取自罗马尼亚(Varga et al., 2019)和波兰(Filipek et al., 2019)多地, 但这些来自不同环境的同一物种却具有高度相似的次生代谢产物.这表明在丁香属特定的次生代谢途径中确实存在无法被环境异质性改变的系统学保守性. ...
Drought predisposes piñon-juniper woodlands to insect attacks and mortality 1 2013
... 极端环境压力可能会使特定的代谢路径延长.木脂素是丁香属中独具特色的成分, 现有文献显示其在羽叶丁香系和短花冠管组中占据优势.在单种系的羽叶丁香中木脂素占比高达46.40%, 这与其生长的极端干旱环境紧密相关.羽叶丁香生长在贺兰山东、西山麓以及陕西南部、甘肃和青海, 该地区原生境大多降水稀少, 巨大的水分胁迫压力使羽叶丁香的地下生物量投入极大, 以粗壮的根系实现土壤深层水分的向性生长(Cui et al., 2016; 高坤等, 2018).莽草酸途径化合物的积累通常有助于增强植物对水分胁迫的耐受性(刘盟盟等, 2017; Sharma et al., 2019), 而该途径中形成的木脂素可在木质部木质化后成为一种后注入型树脂, 使植物在水分胁迫而韧皮部运输减少的情况下更具干旱耐受性(Gaylord et al., 2013; Zhang et al., 2018).这种被极端干旱强化的木脂素代谢途径是以牺牲莽草酸途径中大量上游产物苯丙素为代价以保证下游产物木脂素的产生, 助力粗壮根系的形成以实现土壤深层的趋向性生长, 从而最大程度地吸收生存所需水分.与此同时, 吸引传粉者也是其需要兼顾的因素, 在1 700-3 100 m的高山生境中, 传粉昆虫比低海拔地区更为稀少, 这使得羽叶丁香在萜类途径中更多地利用底物焦磷酸二甲烯丙酯和焦磷酸烯丙酯向法尼基焦磷酸合成方向延伸, 成为丁香属以倍半萜为优势萜类成分的组系.利用较短途径获得的大量倍半萜可在一定程度上实现对传粉昆虫的诱集(Chadwick et al., 2013). ...
A comparative study of terpene composition in different clades of the genus Lavandula 1 2018
... 丰富的次生代谢产物(secondary metabolites)是植物适应复杂环境的策略(Thakur et al., 2019), 也可能在一定条件下带有系统演化的痕迹(Sedio, 2017).植物在生长过程中受到昆虫、微生物和伴生物种等生物因素, 以及温度、光照和水分等非生物因素的影响(杜玮炜和黄宏文, 2008; Wang et al., 2019), 伴随着复杂的时空变化形成了富于变化的次生代谢成分.人们大多认为, 相对于次生代谢产物与环境的紧密联系, 它们与系统学的关联则较为松散(Chen et al., 2020), 与谱系演化的关联具有很大的不确定性(Ernst et al., 2016; Allevato et al., 2019).尽管如此, 仍有****发现在相距2 500 km的秘鲁伊基托斯和巴西马瑙斯保护区中, 经历了海拔、经纬度和土壤环境的巨大变化, 相同物种的种群却共享着超过95%的次生代谢产物, 并且种内化学成分保持极高的一致性(Vleminckx et al., 2018).这意味着在千变万化的环境诱导着丰富的化学多样性背后, 系统保守性仍有迹可循.当今, 人们已经利用快速发展的基因组学和代谢组学工具, 揭示了科以上的分类等级, 以及被子植物门次生代谢关键调控因子家族的演化扩张关系(Chen et al., 2011; 祝志欣和鲁迎青, 2016; 张永增, 2018; Mint Evolutionary Genomics Consortium, 2018; Xu et al., 2020).这表明在较低的分类等级上, 次生代谢或其成分在属内的分布保守性可能同样有章可循(Rønsted et al., 2008; Ernst et al., 2016; Guitton et al., 2018; Chen et al., 2020).尽管由于很难获得具有系统学代表性的属下植物材料而使实证研究较为少见, 但这类研究对于精准而系统地确定属下水平的代谢调控机制具有科学意义, 对功能性成分的系统性挖掘具有现实意义. ...
A chloroplast DNA phylogeny of lilacs (Syringa, Oleaceae): plastome groups show a strong correlation with crossing groups 2 1998
... 对于丁香属内次生代谢产物的讨论离不开物种分类和演化的系统学背景.从经典分类以及既往分子生物学的研究结果看, 丁香属系统演化关系基本明确, 其结果差异主要在于在先分化类群的不同(张美珍等, 1992; Kim and Jansen, 1998; Li et al., 2012).因此,我们依据欧洲丁香与紫丁香存在欧亚间断分布的事实(Kim and Jansen, 1998), 以及核DNA分析在物种演化关系判定上的依据(Li et al., 2012), 支持灌木型欧丁香系是丁香属最先分化的类群.羽叶丁香系在欧丁香系之后独立分化, 乔木型短花冠管组位居灌木型组系之间为中间分化类群(Li et al., 2012), 而对于巧玲花系和红丁香系是丁香属内后续分化类群的观点则不存异议.据此, 我们在进行系统关联讨论时采用Li等(2012)所报道的系统演化模式. ...
... ).因此,我们依据欧洲丁香与紫丁香存在欧亚间断分布的事实(Kim and Jansen, 1998), 以及核DNA分析在物种演化关系判定上的依据(Li et al., 2012), 支持灌木型欧丁香系是丁香属最先分化的类群.羽叶丁香系在欧丁香系之后独立分化, 乔木型短花冠管组位居灌木型组系之间为中间分化类群(Li et al., 2012), 而对于巧玲花系和红丁香系是丁香属内后续分化类群的观点则不存异议.据此, 我们在进行系统关联讨论时采用Li等(2012)所报道的系统演化模式. ...
Enzymatic activation of oleuropein: a protein crosslinker used as a chemical defense in the privet tree 1 1999
... 在分别从系统演化和地理环境角度讨论丁香属次生代谢产物的分布趋势之后, 需要面对的问题还有代谢背景下的系统演化与地理分布跨度的关系.研究表明, 紫丁香和欧洲丁香是丁香属系统学原始组系中的2个近缘姊妹系物种(Li et al., 2012), 它们也分别是东亚和欧洲的广布种(张美珍等, 1992; Fiala, 2008), 而后续分化的红丁香系(Li et al., 2012)中的大多数物种都属于生态幅狭窄的局域种.已知温度和水分是限制物种扩散的主要环境因子(高坤等, 2018; 魏华等, 2018), 丁香地理分布范围的差异一定程度上由对温度和水分敏感的光合同化产物形成和水分利用效率等初生代谢机制的差别决定(Cui et al., 2016).紫丁香和欧洲丁香的广布表明其初生代谢机制保证了它们能够适应东亚和东南欧洲的气候变化, 同时强大的系统学保守性使相距万里的2个系统学意义上的近缘姊妹种均具有环/裂环烯醚萜和苯丙素类产物(表1, 表3), 使它们有能力通过这两类优势次生代谢产物的形成, 增强对小环境中生物及非生物扰动的适应能力, 从而改善植株的生长状况并实现高效的繁衍生息(Konno et al., 1999; 刘盟盟等, 2017; Sharma et al., 2019).而对于性喜冷凉的红丁香系中的红丁香和匈牙利丁香2个近缘种, 气孔较大且密度较低的叶片表皮特征很可能使其在初生代谢热量耗散和叶片水分保持能力上存在限制(高艳等, 2008), 因此其分布局限在高海拔的冷凉或邻水生境(张美珍等, 1992; Lendvay et al., 2016)而成为地理分布上的局域种.在这类水热条件适宜的小环境中, 这些较晚分化的局域种不必应对地域扩散中温度和水分的巨大变化, 有条件形成更加多样的次生代谢产物, 在适宜的生境中最大限度地改善生长质量.这种“初生代谢决定生存格局, 次生代谢改善生活质量”(刘晓侠等, 2015)的情况也发生在同样进化却局域分布的巧玲花系中(图1). ...
Phylogeography of Syringa josikaea (Oleaceae): Early Pleistocene divergence from East Asian relatives and survival in small populations in the Carpathians 1 2016
... 在分别从系统演化和地理环境角度讨论丁香属次生代谢产物的分布趋势之后, 需要面对的问题还有代谢背景下的系统演化与地理分布跨度的关系.研究表明, 紫丁香和欧洲丁香是丁香属系统学原始组系中的2个近缘姊妹系物种(Li et al., 2012), 它们也分别是东亚和欧洲的广布种(张美珍等, 1992; Fiala, 2008), 而后续分化的红丁香系(Li et al., 2012)中的大多数物种都属于生态幅狭窄的局域种.已知温度和水分是限制物种扩散的主要环境因子(高坤等, 2018; 魏华等, 2018), 丁香地理分布范围的差异一定程度上由对温度和水分敏感的光合同化产物形成和水分利用效率等初生代谢机制的差别决定(Cui et al., 2016).紫丁香和欧洲丁香的广布表明其初生代谢机制保证了它们能够适应东亚和东南欧洲的气候变化, 同时强大的系统学保守性使相距万里的2个系统学意义上的近缘姊妹种均具有环/裂环烯醚萜和苯丙素类产物(表1, 表3), 使它们有能力通过这两类优势次生代谢产物的形成, 增强对小环境中生物及非生物扰动的适应能力, 从而改善植株的生长状况并实现高效的繁衍生息(Konno et al., 1999; 刘盟盟等, 2017; Sharma et al., 2019).而对于性喜冷凉的红丁香系中的红丁香和匈牙利丁香2个近缘种, 气孔较大且密度较低的叶片表皮特征很可能使其在初生代谢热量耗散和叶片水分保持能力上存在限制(高艳等, 2008), 因此其分布局限在高海拔的冷凉或邻水生境(张美珍等, 1992; Lendvay et al., 2016)而成为地理分布上的局域种.在这类水热条件适宜的小环境中, 这些较晚分化的局域种不必应对地域扩散中温度和水分的巨大变化, 有条件形成更加多样的次生代谢产物, 在适宜的生境中最大限度地改善生长质量.这种“初生代谢决定生存格局, 次生代谢改善生活质量”(刘晓侠等, 2015)的情况也发生在同样进化却局域分布的巧玲花系中(图1). ...
Phylogenetics and diversification of Syringa inferred from nuclear and plastid DNA sequences 9 2012
... (A) 丁香属组系的系统演化关系(Li et al., 2012)及各类产物在各组系中的占比; (B) 甲戊二羟酸途径(MVA)和脱氧木酮糖磷酸酯途径(DXP) (Mint Evolutionary Genomics Consortium, 2018); (C) 莽草酸途径(刘津等, 2016; 张旭等, 2019).图(A)组系下方的色条表示各类产物(颜色含义同图1), 色条内的百分数指特定类别产物计数在该组系内成分计数总和中所占的百分比(详见图1图注).图(B)和图(C)中化合物色框颜色含义与图(A)相同.图(B)和图(C)中虚线表示由多步反应完成. ...
... (A) The phylogenetic relationship (Li et al., 2012) and percentage of metabolites of Syringa at the level of series (section); (B) Mevalonic acid pathway (MVA) and Deoxyxylulose-5-P pathway (DXP) (Mint Evolutionary Genomics Consortium, 2018); (C) Shikimic acid pathway (Liu et al., 2016; Zhang et al., 2019, in Chinese). The color-stripes below the series (section) indicate different types of metabolites in (A), and the percentage in the color-stripe represents the percentage of a given type of compounds within the series (section) (please see Figure 1 for details). The meanings of the color frames indicating the compound in (B) and (C) are the same as those of (A). The dotted lines in (B) and (C) represent the process of multi-step reactions. ...
... 对于丁香属内次生代谢产物的讨论离不开物种分类和演化的系统学背景.从经典分类以及既往分子生物学的研究结果看, 丁香属系统演化关系基本明确, 其结果差异主要在于在先分化类群的不同(张美珍等, 1992; Kim and Jansen, 1998; Li et al., 2012).因此,我们依据欧洲丁香与紫丁香存在欧亚间断分布的事实(Kim and Jansen, 1998), 以及核DNA分析在物种演化关系判定上的依据(Li et al., 2012), 支持灌木型欧丁香系是丁香属最先分化的类群.羽叶丁香系在欧丁香系之后独立分化, 乔木型短花冠管组位居灌木型组系之间为中间分化类群(Li et al., 2012), 而对于巧玲花系和红丁香系是丁香属内后续分化类群的观点则不存异议.据此, 我们在进行系统关联讨论时采用Li等(2012)所报道的系统演化模式. ...
... ), 以及核DNA分析在物种演化关系判定上的依据(Li et al., 2012), 支持灌木型欧丁香系是丁香属最先分化的类群.羽叶丁香系在欧丁香系之后独立分化, 乔木型短花冠管组位居灌木型组系之间为中间分化类群(Li et al., 2012), 而对于巧玲花系和红丁香系是丁香属内后续分化类群的观点则不存异议.据此, 我们在进行系统关联讨论时采用Li等(2012)所报道的系统演化模式. ...
... ), 支持灌木型欧丁香系是丁香属最先分化的类群.羽叶丁香系在欧丁香系之后独立分化, 乔木型短花冠管组位居灌木型组系之间为中间分化类群(Li et al., 2012), 而对于巧玲花系和红丁香系是丁香属内后续分化类群的观点则不存异议.据此, 我们在进行系统关联讨论时采用Li等(2012)所报道的系统演化模式. ...
Phylogenomic mining of the mints reveals multiple mechanisms contributing to the evolution of chemical diversity in Lamiaceae 4 2018
... 丰富的次生代谢产物(secondary metabolites)是植物适应复杂环境的策略(Thakur et al., 2019), 也可能在一定条件下带有系统演化的痕迹(Sedio, 2017).植物在生长过程中受到昆虫、微生物和伴生物种等生物因素, 以及温度、光照和水分等非生物因素的影响(杜玮炜和黄宏文, 2008; Wang et al., 2019), 伴随着复杂的时空变化形成了富于变化的次生代谢成分.人们大多认为, 相对于次生代谢产物与环境的紧密联系, 它们与系统学的关联则较为松散(Chen et al., 2020), 与谱系演化的关联具有很大的不确定性(Ernst et al., 2016; Allevato et al., 2019).尽管如此, 仍有****发现在相距2 500 km的秘鲁伊基托斯和巴西马瑙斯保护区中, 经历了海拔、经纬度和土壤环境的巨大变化, 相同物种的种群却共享着超过95%的次生代谢产物, 并且种内化学成分保持极高的一致性(Vleminckx et al., 2018).这意味着在千变万化的环境诱导着丰富的化学多样性背后, 系统保守性仍有迹可循.当今, 人们已经利用快速发展的基因组学和代谢组学工具, 揭示了科以上的分类等级, 以及被子植物门次生代谢关键调控因子家族的演化扩张关系(Chen et al., 2011; 祝志欣和鲁迎青, 2016; 张永增, 2018; Mint Evolutionary Genomics Consortium, 2018; Xu et al., 2020).这表明在较低的分类等级上, 次生代谢或其成分在属内的分布保守性可能同样有章可循(Rønsted et al., 2008; Ernst et al., 2016; Guitton et al., 2018; Chen et al., 2020).尽管由于很难获得具有系统学代表性的属下植物材料而使实证研究较为少见, 但这类研究对于精准而系统地确定属下水平的代谢调控机制具有科学意义, 对功能性成分的系统性挖掘具有现实意义. ...
... (A) 丁香属组系的系统演化关系(Li et al., 2012)及各类产物在各组系中的占比; (B) 甲戊二羟酸途径(MVA)和脱氧木酮糖磷酸酯途径(DXP) (Mint Evolutionary Genomics Consortium, 2018); (C) 莽草酸途径(刘津等, 2016; 张旭等, 2019).图(A)组系下方的色条表示各类产物(颜色含义同图1), 色条内的百分数指特定类别产物计数在该组系内成分计数总和中所占的百分比(详见图1图注).图(B)和图(C)中化合物色框颜色含义与图(A)相同.图(B)和图(C)中虚线表示由多步反应完成. ...
... (A) The phylogenetic relationship (Li et al., 2012) and percentage of metabolites of Syringa at the level of series (section); (B) Mevalonic acid pathway (MVA) and Deoxyxylulose-5-P pathway (DXP) (Mint Evolutionary Genomics Consortium, 2018); (C) Shikimic acid pathway (Liu et al., 2016; Zhang et al., 2019, in Chinese). The color-stripes below the series (section) indicate different types of metabolites in (A), and the percentage in the color-stripe represents the percentage of a given type of compounds within the series (section) (please see Figure 1 for details). The meanings of the color frames indicating the compound in (B) and (C) are the same as those of (A). The dotted lines in (B) and (C) represent the process of multi-step reactions. ...
Phylogenetic selection of Narcissus species for drug discovery 1 2008
... 丰富的次生代谢产物(secondary metabolites)是植物适应复杂环境的策略(Thakur et al., 2019), 也可能在一定条件下带有系统演化的痕迹(Sedio, 2017).植物在生长过程中受到昆虫、微生物和伴生物种等生物因素, 以及温度、光照和水分等非生物因素的影响(杜玮炜和黄宏文, 2008; Wang et al., 2019), 伴随着复杂的时空变化形成了富于变化的次生代谢成分.人们大多认为, 相对于次生代谢产物与环境的紧密联系, 它们与系统学的关联则较为松散(Chen et al., 2020), 与谱系演化的关联具有很大的不确定性(Ernst et al., 2016; Allevato et al., 2019).尽管如此, 仍有****发现在相距2 500 km的秘鲁伊基托斯和巴西马瑙斯保护区中, 经历了海拔、经纬度和土壤环境的巨大变化, 相同物种的种群却共享着超过95%的次生代谢产物, 并且种内化学成分保持极高的一致性(Vleminckx et al., 2018).这意味着在千变万化的环境诱导着丰富的化学多样性背后, 系统保守性仍有迹可循.当今, 人们已经利用快速发展的基因组学和代谢组学工具, 揭示了科以上的分类等级, 以及被子植物门次生代谢关键调控因子家族的演化扩张关系(Chen et al., 2011; 祝志欣和鲁迎青, 2016; 张永增, 2018; Mint Evolutionary Genomics Consortium, 2018; Xu et al., 2020).这表明在较低的分类等级上, 次生代谢或其成分在属内的分布保守性可能同样有章可循(Rønsted et al., 2008; Ernst et al., 2016; Guitton et al., 2018; Chen et al., 2020).尽管由于很难获得具有系统学代表性的属下植物材料而使实证研究较为少见, 但这类研究对于精准而系统地确定属下水平的代谢调控机制具有科学意义, 对功能性成分的系统性挖掘具有现实意义. ...
Recent breakthroughs in metabolomics promise to reveal the cryptic chemical traits that mediate plant community composition, character evolution and lineage diversification 1 2017
... 丰富的次生代谢产物(secondary metabolites)是植物适应复杂环境的策略(Thakur et al., 2019), 也可能在一定条件下带有系统演化的痕迹(Sedio, 2017).植物在生长过程中受到昆虫、微生物和伴生物种等生物因素, 以及温度、光照和水分等非生物因素的影响(杜玮炜和黄宏文, 2008; Wang et al., 2019), 伴随着复杂的时空变化形成了富于变化的次生代谢成分.人们大多认为, 相对于次生代谢产物与环境的紧密联系, 它们与系统学的关联则较为松散(Chen et al., 2020), 与谱系演化的关联具有很大的不确定性(Ernst et al., 2016; Allevato et al., 2019).尽管如此, 仍有****发现在相距2 500 km的秘鲁伊基托斯和巴西马瑙斯保护区中, 经历了海拔、经纬度和土壤环境的巨大变化, 相同物种的种群却共享着超过95%的次生代谢产物, 并且种内化学成分保持极高的一致性(Vleminckx et al., 2018).这意味着在千变万化的环境诱导着丰富的化学多样性背后, 系统保守性仍有迹可循.当今, 人们已经利用快速发展的基因组学和代谢组学工具, 揭示了科以上的分类等级, 以及被子植物门次生代谢关键调控因子家族的演化扩张关系(Chen et al., 2011; 祝志欣和鲁迎青, 2016; 张永增, 2018; Mint Evolutionary Genomics Consortium, 2018; Xu et al., 2020).这表明在较低的分类等级上, 次生代谢或其成分在属内的分布保守性可能同样有章可循(Rønsted et al., 2008; Ernst et al., 2016; Guitton et al., 2018; Chen et al., 2020).尽管由于很难获得具有系统学代表性的属下植物材料而使实证研究较为少见, 但这类研究对于精准而系统地确定属下水平的代谢调控机制具有科学意义, 对功能性成分的系统性挖掘具有现实意义. ...
Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress 2 2019
... 在分别从系统演化和地理环境角度讨论丁香属次生代谢产物的分布趋势之后, 需要面对的问题还有代谢背景下的系统演化与地理分布跨度的关系.研究表明, 紫丁香和欧洲丁香是丁香属系统学原始组系中的2个近缘姊妹系物种(Li et al., 2012), 它们也分别是东亚和欧洲的广布种(张美珍等, 1992; Fiala, 2008), 而后续分化的红丁香系(Li et al., 2012)中的大多数物种都属于生态幅狭窄的局域种.已知温度和水分是限制物种扩散的主要环境因子(高坤等, 2018; 魏华等, 2018), 丁香地理分布范围的差异一定程度上由对温度和水分敏感的光合同化产物形成和水分利用效率等初生代谢机制的差别决定(Cui et al., 2016).紫丁香和欧洲丁香的广布表明其初生代谢机制保证了它们能够适应东亚和东南欧洲的气候变化, 同时强大的系统学保守性使相距万里的2个系统学意义上的近缘姊妹种均具有环/裂环烯醚萜和苯丙素类产物(表1, 表3), 使它们有能力通过这两类优势次生代谢产物的形成, 增强对小环境中生物及非生物扰动的适应能力, 从而改善植株的生长状况并实现高效的繁衍生息(Konno et al., 1999; 刘盟盟等, 2017; Sharma et al., 2019).而对于性喜冷凉的红丁香系中的红丁香和匈牙利丁香2个近缘种, 气孔较大且密度较低的叶片表皮特征很可能使其在初生代谢热量耗散和叶片水分保持能力上存在限制(高艳等, 2008), 因此其分布局限在高海拔的冷凉或邻水生境(张美珍等, 1992; Lendvay et al., 2016)而成为地理分布上的局域种.在这类水热条件适宜的小环境中, 这些较晚分化的局域种不必应对地域扩散中温度和水分的巨大变化, 有条件形成更加多样的次生代谢产物, 在适宜的生境中最大限度地改善生长质量.这种“初生代谢决定生存格局, 次生代谢改善生活质量”(刘晓侠等, 2015)的情况也发生在同样进化却局域分布的巧玲花系中(图1). ...
... 极端环境压力可能会使特定的代谢路径延长.木脂素是丁香属中独具特色的成分, 现有文献显示其在羽叶丁香系和短花冠管组中占据优势.在单种系的羽叶丁香中木脂素占比高达46.40%, 这与其生长的极端干旱环境紧密相关.羽叶丁香生长在贺兰山东、西山麓以及陕西南部、甘肃和青海, 该地区原生境大多降水稀少, 巨大的水分胁迫压力使羽叶丁香的地下生物量投入极大, 以粗壮的根系实现土壤深层水分的向性生长(Cui et al., 2016; 高坤等, 2018).莽草酸途径化合物的积累通常有助于增强植物对水分胁迫的耐受性(刘盟盟等, 2017; Sharma et al., 2019), 而该途径中形成的木脂素可在木质部木质化后成为一种后注入型树脂, 使植物在水分胁迫而韧皮部运输减少的情况下更具干旱耐受性(Gaylord et al., 2013; Zhang et al., 2018).这种被极端干旱强化的木脂素代谢途径是以牺牲莽草酸途径中大量上游产物苯丙素为代价以保证下游产物木脂素的产生, 助力粗壮根系的形成以实现土壤深层的趋向性生长, 从而最大程度地吸收生存所需水分.与此同时, 吸引传粉者也是其需要兼顾的因素, 在1 700-3 100 m的高山生境中, 传粉昆虫比低海拔地区更为稀少, 这使得羽叶丁香在萜类途径中更多地利用底物焦磷酸二甲烯丙酯和焦磷酸烯丙酯向法尼基焦磷酸合成方向延伸, 成为丁香属以倍半萜为优势萜类成分的组系.利用较短途径获得的大量倍半萜可在一定程度上实现对传粉昆虫的诱集(Chadwick et al., 2013). ...
Improving production of plant secondary metabolites through biotic and abiotic elicitation 1 2019
... 丰富的次生代谢产物(secondary metabolites)是植物适应复杂环境的策略(Thakur et al., 2019), 也可能在一定条件下带有系统演化的痕迹(Sedio, 2017).植物在生长过程中受到昆虫、微生物和伴生物种等生物因素, 以及温度、光照和水分等非生物因素的影响(杜玮炜和黄宏文, 2008; Wang et al., 2019), 伴随着复杂的时空变化形成了富于变化的次生代谢成分.人们大多认为, 相对于次生代谢产物与环境的紧密联系, 它们与系统学的关联则较为松散(Chen et al., 2020), 与谱系演化的关联具有很大的不确定性(Ernst et al., 2016; Allevato et al., 2019).尽管如此, 仍有****发现在相距2 500 km的秘鲁伊基托斯和巴西马瑙斯保护区中, 经历了海拔、经纬度和土壤环境的巨大变化, 相同物种的种群却共享着超过95%的次生代谢产物, 并且种内化学成分保持极高的一致性(Vleminckx et al., 2018).这意味着在千变万化的环境诱导着丰富的化学多样性背后, 系统保守性仍有迹可循.当今, 人们已经利用快速发展的基因组学和代谢组学工具, 揭示了科以上的分类等级, 以及被子植物门次生代谢关键调控因子家族的演化扩张关系(Chen et al., 2011; 祝志欣和鲁迎青, 2016; 张永增, 2018; Mint Evolutionary Genomics Consortium, 2018; Xu et al., 2020).这表明在较低的分类等级上, 次生代谢或其成分在属内的分布保守性可能同样有章可循(Rønsted et al., 2008; Ernst et al., 2016; Guitton et al., 2018; Chen et al., 2020).尽管由于很难获得具有系统学代表性的属下植物材料而使实证研究较为少见, 但这类研究对于精准而系统地确定属下水平的代谢调控机制具有科学意义, 对功能性成分的系统性挖掘具有现实意义. ...
Phenolic composition, antioxidant and antinociceptive activities of Syringa vulgaris L. bark and leaf extracts. 1 2019
... 丁香属中的环/裂环烯醚萜类和苯丙素类产物呈现较强的系统遗传保守性, 甚至遥远的地理隔离也未造成近缘种优势代谢产物的明显差异.紫丁香和欧洲丁香是系统学意义上的姊妹系(Li et al., 2012), 虽然二者在自然分布上存在着东亚与巴尔干半岛的遥远亚欧间断分布(表1), 却均具有丰富的环/裂环烯醚萜类和苯丙素类产物(表3).苯丙素类产物形成的保守性还体现在短花冠管组的2个近缘种暴马丁香和日本丁香中.暴马丁香广泛分布于中国北方及远东地区, 日本丁香仅在日本呈局域分布, 但2个系统学的近缘种同样具有占比相近的苯丙素类成分(表1, 表3).由此表明, 环/裂环烯醚萜类和苯丙素类成分的形成具有较强的遗传保守性, 强烈的保守性没有因遥远的地理距离而消弱.与此同时, 在相邻的近缘组系中出现了较高占比的特定类别化合物的连续分布(图1).图2A显示, 木脂素以较高的占比出现在相邻的羽叶丁香系(46.40%)和短花冠管组(18.18%)中; 倍半萜分别以36.80%、13.64%和27.78%的较高占比出现在相邻的羽叶丁香系、短花冠管组和巧玲花系中; 苯丙素以25.00%和22.22%的占比出现在相邻的短花冠管组和巧玲花系中; 环/裂环烯醚萜类分别以38.64%和25.00%的较高占比出现在相邻的短花冠管组和巧玲花系中.值得一提的是, 在针对同一个物种的诸多文献进行化合物汇总时, 我们发现欧洲丁香的样品取自罗马尼亚(Varga et al., 2019)和波兰(Filipek et al., 2019)多地, 但这些来自不同环境的同一物种却具有高度相似的次生代谢产物.这表明在丁香属特定的次生代谢途径中确实存在无法被环境异质性改变的系统学保守性. ...
Divergent secondary metabolites and habitat filtering both contribute to tree species coexistence in the Peruvian Amazon 1 2018
... 丰富的次生代谢产物(secondary metabolites)是植物适应复杂环境的策略(Thakur et al., 2019), 也可能在一定条件下带有系统演化的痕迹(Sedio, 2017).植物在生长过程中受到昆虫、微生物和伴生物种等生物因素, 以及温度、光照和水分等非生物因素的影响(杜玮炜和黄宏文, 2008; Wang et al., 2019), 伴随着复杂的时空变化形成了富于变化的次生代谢成分.人们大多认为, 相对于次生代谢产物与环境的紧密联系, 它们与系统学的关联则较为松散(Chen et al., 2020), 与谱系演化的关联具有很大的不确定性(Ernst et al., 2016; Allevato et al., 2019).尽管如此, 仍有****发现在相距2 500 km的秘鲁伊基托斯和巴西马瑙斯保护区中, 经历了海拔、经纬度和土壤环境的巨大变化, 相同物种的种群却共享着超过95%的次生代谢产物, 并且种内化学成分保持极高的一致性(Vleminckx et al., 2018).这意味着在千变万化的环境诱导着丰富的化学多样性背后, 系统保守性仍有迹可循.当今, 人们已经利用快速发展的基因组学和代谢组学工具, 揭示了科以上的分类等级, 以及被子植物门次生代谢关键调控因子家族的演化扩张关系(Chen et al., 2011; 祝志欣和鲁迎青, 2016; 张永增, 2018; Mint Evolutionary Genomics Consortium, 2018; Xu et al., 2020).这表明在较低的分类等级上, 次生代谢或其成分在属内的分布保守性可能同样有章可循(Rønsted et al., 2008; Ernst et al., 2016; Guitton et al., 2018; Chen et al., 2020).尽管由于很难获得具有系统学代表性的属下植物材料而使实证研究较为少见, 但这类研究对于精准而系统地确定属下水平的代谢调控机制具有科学意义, 对功能性成分的系统性挖掘具有现实意义. ...
The structure and function of major plant metabolite modifications 1 2019
... 丰富的次生代谢产物(secondary metabolites)是植物适应复杂环境的策略(Thakur et al., 2019), 也可能在一定条件下带有系统演化的痕迹(Sedio, 2017).植物在生长过程中受到昆虫、微生物和伴生物种等生物因素, 以及温度、光照和水分等非生物因素的影响(杜玮炜和黄宏文, 2008; Wang et al., 2019), 伴随着复杂的时空变化形成了富于变化的次生代谢成分.人们大多认为, 相对于次生代谢产物与环境的紧密联系, 它们与系统学的关联则较为松散(Chen et al., 2020), 与谱系演化的关联具有很大的不确定性(Ernst et al., 2016; Allevato et al., 2019).尽管如此, 仍有****发现在相距2 500 km的秘鲁伊基托斯和巴西马瑙斯保护区中, 经历了海拔、经纬度和土壤环境的巨大变化, 相同物种的种群却共享着超过95%的次生代谢产物, 并且种内化学成分保持极高的一致性(Vleminckx et al., 2018).这意味着在千变万化的环境诱导着丰富的化学多样性背后, 系统保守性仍有迹可循.当今, 人们已经利用快速发展的基因组学和代谢组学工具, 揭示了科以上的分类等级, 以及被子植物门次生代谢关键调控因子家族的演化扩张关系(Chen et al., 2011; 祝志欣和鲁迎青, 2016; 张永增, 2018; Mint Evolutionary Genomics Consortium, 2018; Xu et al., 2020).这表明在较低的分类等级上, 次生代谢或其成分在属内的分布保守性可能同样有章可循(Rønsted et al., 2008; Ernst et al., 2016; Guitton et al., 2018; Chen et al., 2020).尽管由于很难获得具有系统学代表性的属下植物材料而使实证研究较为少见, 但这类研究对于精准而系统地确定属下水平的代谢调控机制具有科学意义, 对功能性成分的系统性挖掘具有现实意义. ...
Transcriptome-wide analysis of the AP2/ERF transcription factor gene family involved in the regulation of gypenoside biosynthesis in Gynostemma pentaphyllum 1 2020
... 丰富的次生代谢产物(secondary metabolites)是植物适应复杂环境的策略(Thakur et al., 2019), 也可能在一定条件下带有系统演化的痕迹(Sedio, 2017).植物在生长过程中受到昆虫、微生物和伴生物种等生物因素, 以及温度、光照和水分等非生物因素的影响(杜玮炜和黄宏文, 2008; Wang et al., 2019), 伴随着复杂的时空变化形成了富于变化的次生代谢成分.人们大多认为, 相对于次生代谢产物与环境的紧密联系, 它们与系统学的关联则较为松散(Chen et al., 2020), 与谱系演化的关联具有很大的不确定性(Ernst et al., 2016; Allevato et al., 2019).尽管如此, 仍有****发现在相距2 500 km的秘鲁伊基托斯和巴西马瑙斯保护区中, 经历了海拔、经纬度和土壤环境的巨大变化, 相同物种的种群却共享着超过95%的次生代谢产物, 并且种内化学成分保持极高的一致性(Vleminckx et al., 2018).这意味着在千变万化的环境诱导着丰富的化学多样性背后, 系统保守性仍有迹可循.当今, 人们已经利用快速发展的基因组学和代谢组学工具, 揭示了科以上的分类等级, 以及被子植物门次生代谢关键调控因子家族的演化扩张关系(Chen et al., 2011; 祝志欣和鲁迎青, 2016; 张永增, 2018; Mint Evolutionary Genomics Consortium, 2018; Xu et al., 2020).这表明在较低的分类等级上, 次生代谢或其成分在属内的分布保守性可能同样有章可循(Rønsted et al., 2008; Ernst et al., 2016; Guitton et al., 2018; Chen et al., 2020).尽管由于很难获得具有系统学代表性的属下植物材料而使实证研究较为少见, 但这类研究对于精准而系统地确定属下水平的代谢调控机制具有科学意义, 对功能性成分的系统性挖掘具有现实意义. ...
Bioactive sesquiterpenoids from the peeled stems of Syringa pinnatifolia 1 2018
... 极端环境压力可能会使特定的代谢路径延长.木脂素是丁香属中独具特色的成分, 现有文献显示其在羽叶丁香系和短花冠管组中占据优势.在单种系的羽叶丁香中木脂素占比高达46.40%, 这与其生长的极端干旱环境紧密相关.羽叶丁香生长在贺兰山东、西山麓以及陕西南部、甘肃和青海, 该地区原生境大多降水稀少, 巨大的水分胁迫压力使羽叶丁香的地下生物量投入极大, 以粗壮的根系实现土壤深层水分的向性生长(Cui et al., 2016; 高坤等, 2018).莽草酸途径化合物的积累通常有助于增强植物对水分胁迫的耐受性(刘盟盟等, 2017; Sharma et al., 2019), 而该途径中形成的木脂素可在木质部木质化后成为一种后注入型树脂, 使植物在水分胁迫而韧皮部运输减少的情况下更具干旱耐受性(Gaylord et al., 2013; Zhang et al., 2018).这种被极端干旱强化的木脂素代谢途径是以牺牲莽草酸途径中大量上游产物苯丙素为代价以保证下游产物木脂素的产生, 助力粗壮根系的形成以实现土壤深层的趋向性生长, 从而最大程度地吸收生存所需水分.与此同时, 吸引传粉者也是其需要兼顾的因素, 在1 700-3 100 m的高山生境中, 传粉昆虫比低海拔地区更为稀少, 这使得羽叶丁香在萜类途径中更多地利用底物焦磷酸二甲烯丙酯和焦磷酸烯丙酯向法尼基焦磷酸合成方向延伸, 成为丁香属以倍半萜为优势萜类成分的组系.利用较短途径获得的大量倍半萜可在一定程度上实现对传粉昆虫的诱集(Chadwick et al., 2013). ...
 ,1,2,*, 崔洪霞
,1,2,*, 崔洪霞
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT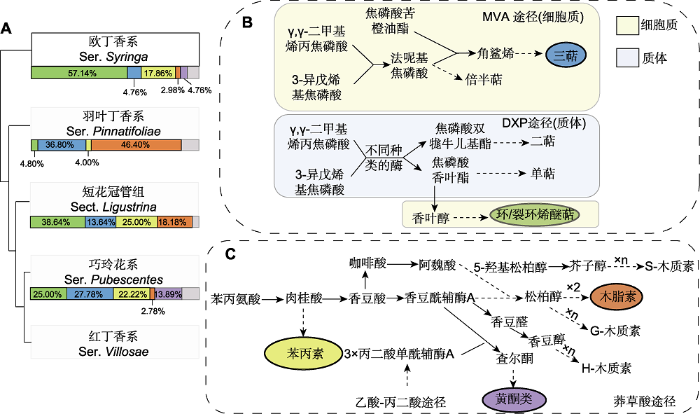 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}