罗钱, 张燕莎, 欧静
, * 贵州大学林学院, 贵阳 550025
Callus Induction and Plant Regeneration of Cerasus serrulata var. lannesiana cv. ‘Grandiflora’ Qian Luo, Yansha Zhang, Jing Ou
, * College of Forestry, Guizhou University, Guiyang 550025, China
通讯作者: *E-mail:
coloroj@126.com 责任编辑: 朱亚娜
收稿日期: 2020-12-19
接受日期: 2021-05-7
基金资助: 贵州省农业科技支撑项目 (20182287 )
Corresponding authors: *E-mail:
coloroj@126.com Received: 2020-12-19
Accepted: 2021-05-7
摘要 为建立郁金樱(
Cerasus serrulata var
. lannesiana cv. ‘Grandiflora’)再生体系, 以多年生母株小叶、一年生嫁接苗小叶、腋芽诱导小叶和增殖一代小叶为外植体, 探讨不同外植体和植物激素组合对郁金樱愈伤组织诱导、不定芽分化、增殖和生根的影响。结果表明, 4种外植体均可诱导出愈伤组织, 除多年生母株小叶外皆分化出不定芽, 外植体幼化程度越高, 后期培养潜力越大。以增殖一代小叶为外植体效果最佳, 其最适愈伤组织诱导培养基为MS+0.5 mg·L
-1 6-BA+1.0 mg·L
-1 2,4-D, 诱导率达96.22%; 最佳分化培养基为MS+1.0 mg·L
-1 6-BA+0.1 mg·L
-1 2,4-D+0.1 mg·L
-1 TDZ, 分化率达78.14%; 最佳增殖培养基为MS+1.0 mg·L
-1 6-BA, 增殖系数可达7.85; 最佳生根培养基为不含任何激素的1/2MS培养基, 生根率达100%。不同外植体获得的再生植株移栽成活后生长差异显著, 以增殖一代小叶诱导的再生植株长势最佳。
关键词: 郁金樱 ;
愈伤组织 ;
外植体 ;
植株再生 ;
组织培养 Abstract In order to establish the regeneration system of
Cerasus serrulata var.
lannesiana cv. ‘Grandiflora’, the effects of different explants and plant hormone combinations on callus induction, adventitious bud differentiation, proliferation and rooting were studied using perennial mother plant leaflets, annual grafted seedling leaflets, axillary bud induction leaflets and proliferative first-generation leaflets as explants. The results showed that callus could be induced from all four explants, and adventitious buds could be derived from all explants except the leaflets of perennial mother plants. The higher the degree of explants’ juvenility, the greater the success of subsequent culture, with the best explants were the proliferative first generation leaflets. The best medium for callus induction was MS+0.5 mg·L
-1 6-BA+1.0 mg·L
-1 2,4-D, and the induction rate was 96.22%. The optimal medium for differentiation was MS+1.0 mg·L
-1 6-BA+0.1 mg·L
-1 2,4-D+0.1 mg·L
-1 TDZ, and the differentiation rate was 78.14%. The optimal medium for proliferation was MS+1.0 mg·L
-1 6-BA, and the proliferation coefficient reached 7.85. The optimal medium for rooting was 1/2MS without any hormone, and the regenerated plants with 100% rooting rate were obtained. The regenerated plants from different explants grew very differently, and those induced from first generation of proliferative leaflets had the best growth.
Keywords: Cerasus serrulata var. lannesiana cv. ‘Grandiflora’;
callus ;
explants ;
plant regeneration ;
tissue culture PDF (4746KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 罗钱, 张燕莎, 欧静 . 郁金樱愈伤组织诱导及植株再生. 植物学报, 2021,
56 (4): 451-461 doi:10.11983/CBB20205
Luo Qian, Zhang Yansha, Ou Jing .
Callus Induction and Plant Regeneration of Cerasus serrulata var. lannesiana cv. ‘Grandiflora’ .
Chinese Bulletin of Botany , 2021,
56 (4): 451-461 doi:10.11983/CBB20205
郁金樱(
Cerasus serrulata var
. lannesiana cv. ‘Grandiflora’)是蔷薇科(Rosaceae)樱属(
Cerasus )落叶乔木, 花叶同开, 花色淡黄至绿色, 花期在3月下旬至4月上旬, 在我国长三角地区有栽培, 现已推广到全国。樱花品种花色大多为白色、粉色或红色, 郁金樱因其绿色花而独特, 在盛花期和落花期均有极高的观赏价值, 园林开发潜力巨大(
刘晓莉, 2012 )。郁金樱具有良好的适应性和观赏性(
朱继军等, 2015 ), 其在校园绿地、居住区绿地、道路绿地、综合公园和城市广场上皆有分布, 苗木市场需求较大(
蒋冬月等, 2019 ;
杨小燕等, 2019 )。生产上樱花主要通过嫁接繁殖, 但该繁殖方法存在占用空间资源大、成苗周期长及种苗价格高等问题, 且易受时间、气候和环境等因素影响, 经多代繁殖后, 易造成病毒积累, 影响樱花的产量和品质。随着现代生物技术的发展, 利用基因工程技术进行植物性状改良已成为可行的育种方式(
张旭红等, 2018 ), 而建立高效且快速的再生体系是进行遗传转化研究的前提。因此, 应用植物组织培养技术建立无性快繁体系, 对郁金樱的遗传改良具有重要意义。
目前, 关于樱花的组培研究主要集中在福建山樱花(
Prunus campanulata ) (
黄守印等, 2003 ;
吕月良等, 2006 ;
Chen et al., 2016 )、日本晚樱杨贵妃(
C. serrulata var.
lannesiana cv. ‘Mollis’)、日本晚樱醉红(
C. serrulata var
. lannesiana cv. ‘Zuihong’) (
张灵灵和蒋细旺, 2015 )、华中樱(
C. conradinae )、迎春樱(
C. discoidea ) (
宋斯妤, 2018 )、染井吉野樱(
C. ×
yedoensis cv. ‘Somei-yoshino’) (
徐晨捷和欧静, 2020 )和喜马拉雅樱花(
C. cerasoides ) (
李水根等, 2020 )等少数品种(
李艳敏等, 2008 ,
2012 )。关于樱花植株再生和遗传转化方面的研究较少, 仅见福建山樱花以腋芽诱导小叶为外植体, 成功建立了植株再生体系和体细胞胚胎发生途径(
邹娜等, 2013 ); 以雪落樱(
C. xueluoensis ) (
史港影等, 2014 )和御殿场樱(
C. ×
incisa cv. ‘Gotenba-zakura’) (
雷巾茗, 2020 )叶片为外植体建立了愈伤组织再生体系, 目前尚未见有关郁金樱再生体系及组织培养的报道。通过对北玄参(
Scrophularia buergeriana ) (
任如意等, 2017 )、黄花乌头(
Aconitum coreanum ) (
刘莉莉等, 2015 )、绒毛白蜡(
Fraxinus velutina ) (
燕丽萍等, 2016 )和月季(
Rosa chinensis ) (
陈雪等, 2011 )等植株再生的研究表明, 不同外植体对植株再生影响极显著, 直接关系试验成功与否, 故选择正确外植体是试验成功的前提。本研究以郁金樱不同幼化程度的小叶为材料, 以期通过愈伤组织诱导不定芽分化、不定芽增殖和生根, 建立高效、稳定的郁金樱再生体系, 为其生物技术育种及后续的遗传转化研究提供参考。
1 植物材料 试验材料郁金樱(
Cerasus serrulata G. Don var
. lannesiana cv.
‘ Grandiflora’)采自贵阳市白云区牛场乡石龙基地。在天气晴朗、无下雨时, 分别从多年生郁金樱母株(7年生, 以下称多年生母株)、1年生郁金樱嫁接苗(砧木为1年生山樱花, 接穗为7年生郁金樱母株)剪取当年生颜色嫩绿、半木质化且无病虫害的健壮枝条, 置于保鲜袋中带回实验室备用。其中, 腋芽诱导小叶以7年生母株半木质化茎段诱导而来, 增殖一代小叶外植体采自7年生母株, 经诱导、增殖培养一代后取幼嫩小叶进行试验。
2 培养基成分与培养条件 2.1 郁金樱无菌体的获得 将郁金樱枝条去掉叶片, 保留部分叶柄, 剪成4 cm左右带腋芽茎段。挑选颜色嫩绿的叶片, 保留叶柄, 分别于流水下用软毛刷蘸洗洁精液轻刷茎段和叶片表面, 用流水冲洗1小时刷洗干净。在超净工作台内进行外植体消毒处理: 用75%乙醇摇晃消毒30秒, 用无菌水冲洗3次, 然后转至0.1% (m/v)升汞(HgCl
2 )溶液摇晃消毒6分钟, 最后用无菌水冲洗5次, 滤纸吸干水分备用。将带腋芽茎段切掉两端, 切成2 cm左右接种于MS+2 mg·L
-1 6-BA+0.2 mg·L
-1 IBA培养基进行腋芽诱导。
2.2 愈伤组织诱导 分别将多年生母株幼嫩小叶、一年生嫁接苗幼嫩小叶、腋芽诱导小叶(当腋芽生长至3-4 cm时完全展开的幼嫩小叶)和增殖培养一代的幼嫩小叶(带叶柄)用解剖刀于叶主脉垂直处横切3-4刀但不切断, 叶面朝上接种于含不同浓度6-BA和2,4-D的MS愈伤组织诱导培养基。每个处理接种15个外植体, 每瓶接种1-2片小叶, 3次重复。20天后统计出愈叶片数, 并计算愈伤组织诱导率。愈伤诱导率(%)=(出愈叶片数/接种叶片数)×100。
2.3 愈伤组织分化 将获得的愈伤组织增殖后, 选择质地紧密的胚性愈伤组织, 转入含不同浓度TDZ、6-BA和2,4-D配比组合的MS分化培养基(
表1 )。每个处理接种20瓶, 每瓶接种3块胚性愈伤组织, 重复3次。30天后统计愈伤组织分化率。分化率(%)=(分化出不定芽愈伤组织块数/接种总块数)×100。
Table 1 表1 表1 郁金樱愈伤组织分化不同植物激素正交设计
Table 1
Orthogonal design of different plant hormones in callus differentiation of Cerasus serrulata Concentrations of plant -1 ) Treatment code 1 2 3 4 5 6 7 8 9 6-BA 0.5 0.5 0.5 1 1 1 2 2 2 2,4-D 0.1 0.5 1 0.1 0.5 1 0.1 0.5 1 TDZ 0.01 0.1 0.5 0.1 0.5 0.01 0.5 0.01 0.1
TDZ: 噻苯隆; TDZ: Thidiazuron 新窗口打开 |
下载CSV 2.4 增殖及生根培养 将分化成功的愈伤组织带不定芽切成小块, 连同基部愈伤组织一起转入含不同6-BA和NAA组合的MS增殖培养基。在增殖培养基上不定芽迅速生长和增殖, 当不定芽长至2-5 cm高时, 从愈伤基部切掉后将不定芽转入含不同浓度IBA和NAA的1/2MS生根培养基。35天后统计生根率及生根数。增殖系数=有效芽数/接种芽数; 生根率(%)=(生根组培苗数/接种组培苗数)×100。
2.5 炼苗移栽 选择生长健壮、根系发育良好的郁金樱组培苗, 半打开瓶盖后于室内炼苗2天, 然后全打开瓶盖炼苗5天。用流水冲洗掉根部附着的培养基, 然后将组培苗移栽于泥炭土:蛭石:河沙=2:1:1 (v/v/v)的混合基质中, 注意保温保湿。30天后统计成活率。成活率(%)=(移栽成活植株数/移栽总株数)×100。
2.6 培养条件 上述培养基中均添加30 g·L
-1 蔗糖、6 g·L
-1 琼脂, pH值为5.8, 121°C高压灭菌20分钟。将组培材料置于温度为(25±1)°C、光照强度为160 μmol·m
-2 ·s
-1 、光周期为12小时光照/12小时黑暗的光照培养箱中培养。
2.7 数据处理及分析 用单反相机拍摄郁金樱再生过程各阶段形态及发生情况。采用SPSS 24.0软件分析数据, 采用Photoshop 2017处理图片, 用Microsoft Excel 2014制表, 采用Greship 8.0.2制图。
3 结果与讨论 3.1 不同激素组合对不同外植体愈伤组织诱导的影响 为筛选郁金樱愈伤组织诱导最佳外植体及激素组合, 以郁金樱多年生母株小叶、一年生嫁接苗小叶、腋芽诱导小叶和增殖一代小叶为外植体进行愈伤组织诱导(
图1 )。将不同外植体小叶接种于诱导培养基1周后, 叶片边缘朝叶脉中心卷曲拱起; 2周后, 小叶主叶脉切口及叶柄端出现细小球形愈伤突起, 愈伤突起由主叶脉向切口两端衍生逐渐变小。球形突起多发生于各支叶脉切口处, 可能是叶脉作为物质运输交换的组织, 最先感受到外界环境的激素变化从而脱分化产生愈伤组织(
房洪舟等, 2019 ), 后期叶片质地逐渐变硬变脆。不同外植体诱导愈伤组织从颜色、质地到形态皆有差异(
图2A -E7)。以多年生母株小叶为外植体诱导的愈伤组织颜色多呈黄褐色, 部分为红色, 质地较紧实坚硬, 表面有粗糙颗粒突起(
图2B 1)。其它3种外植体诱导的愈伤组织质地较松软, 表面分布有大量细小颗粒, 其中一年生嫁接苗小叶诱导的愈伤组织呈浅黄色(
图2C 1), 腋芽小叶愈伤组织和增殖一代小叶愈伤组织为透明白色, 表面有光泽(
图2D 1, E1)。多年生母株诱导的愈伤组织无分化能力, 为非胚性愈伤组织, 后期培养逐渐褐化(
图2B 2, B3)。嫁接苗诱导愈伤组织(
图2C 2)随着后期培养激素的调整可转化为胚性愈伤组织, 腋芽及增殖一代小叶诱导的愈伤组织多为胚性愈伤组织, 以增殖一代小叶为外植体诱导效果最佳(
图2D 2, D3, E2)。
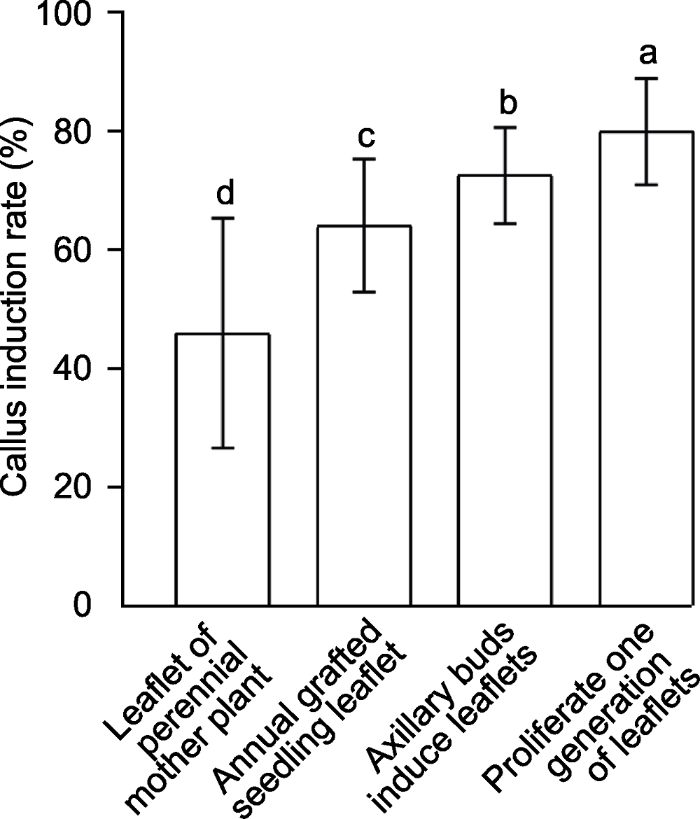
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1郁金樱不同外植体愈伤组织的平均诱导率 不同小写字母表示不同处理间差异显著(
P <0.05)。
Figure 1Average callus induction rates in different explants of Cerasus serrulata Different lowercase letters indicate significant differences among different treatments (
P <0.05).
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2郁金樱愈伤组织诱导、分化及增殖 (A) 腋芽诱导小叶; (B1)-(B3) 多年生母株小叶愈伤组织诱导及分化; (C1)-(C7) 一年生嫁接苗小叶愈伤组织诱导、不定芽分化及增殖过程; (D1)-(D7) 腋芽诱导小叶的愈伤组织诱导、不定芽分化及增殖过程; (E1)-(E7) 增殖一代小叶的愈伤组织诱导、不定芽分化及增殖过程。Bars=1 cm
Figure 2Callus induction, differentiation and proliferation of Cerasus serrulata (A) Axillary buds induce leaflets; (B1)-(B3) Leaflet callus induction and differentiation of perennial mother plant; (C1)-(C7) Leaflet callus induction, adventitious bud differentiation and proliferation of one-year grafted seedlings; (D1)-(D7) The process of callus induction, adventitious bud differentiation and proliferation induced from axillary buds; (E1)-(E7) The process of callus induction, adventitious bud differentiation and proliferation of the proliferative generation of leaflets. Bars=1 cm
本研究表明, 不同浓度6-BA和2,4-D组合激素处理后, 各郁金樱外植体愈伤发生表现出明显差异(
表2 )。多年生母树小叶、一年生嫁接苗小叶、腋芽诱导小叶和增殖一代小叶4种不同外植体的愈伤组织诱导率排序为: 46.31%<64.41%<72.86%<80.27% (
图1 )。分析多年生母株愈伤组织诱导率(
表2 )发现, 当2,4-D浓度在2-3 mg·L
-1 时愈伤诱导效果显著, 诱导率均在57%以上, 配合0.5 mg·L
-1 6-BA能使诱导率达到最大(68.55%), 比单独使用2,4-D时提高11.20%, 过高浓度2,4-D和6-BA对其皆有抑制效应。就一年生嫁接苗愈伤组织诱导而言, 当6-BA浓度为0.5 mg·L
-1 时, 配合2.0 mg·L
-1 2,4-D能使愈伤组织诱导率达到最大(80.71%); 腋芽愈伤组织诱导率仅次于增殖一代小叶, 两者对激素响应敏感, 当6-BA和2,4-D浓度分别为0.5和2.0 mg·L
-1 时腋芽愈伤组织诱导率最高(88.74%)。而增殖一代小叶愈伤组织诱导率在2,4-D浓度为0.5 mg·L
-1 、6-BA浓度为1.0 mg·L
-1 时达到最大(96.22%), 低浓度2,4-D及6-BA配合使用对增殖一代小叶愈伤组织的诱导效果较好, 形成的愈伤组织多为胚性愈伤组织且生长良好。
Table 2 表2 表2 不同激素组合及外植体对郁金樱愈伤组织诱导的影响(平均值±标准误)
Table 2
Effects of different hormone combinations and explants on Cerasus serrulata callus induction (means±SE) No. 6-BA -1 ) 2,4-D -1 ) Callus induction rate (%) Leaflet of perennial mother plant Annual grafted Axillary buds induce leaflets Proliferate one generation of leaflets 1 0 0.5 0 l 42.87±1.51 i 66.09±0.72 g 66.51±0.33 i 2 0 1.0 23.91±0.51 j 49.32±1.01 h 68.46±1.00 ef 72.86±0.59 g 3 0 1.5 46.28±0.58 h 58.92±0.72 f 67.64±0.80 f 76.23±0.96 ef 4 0 2.0 57.35±0.68 e 68.63±0.60 d 72.52±1.16 d 80.77±0.59 d 5 0 3.0 57.15±0.32 e 61.31±1.34 e 69.61±0.67 e 77.63±0.71 e 6 0.5 0.5 16.68±1.45 k 55.08±0.82 g 63.67±0.54 h 71.12±0.20 h 7 0.5 1.0 60.09±0.98 d 61.43±1.40 e 69.04±0.70 ef 96.22±0.94 a 8 0.5 1.5 49.31±0.70 g 74.11±0.78 c 83.07±0.73 c 94.63±0.42 b 9 0.5 2.0 68.55±1.04 a 80.71±0.49 a 88.74±0.45 a 91.40±0.80 c 10 0.5 3.0 62.52±1.09 c 77.62±1.11 b 86.17±0.72 b 77.05±0.77 ef 11 1 0.5 27.85±0.69 i 62.06±0.73 e 68.09±0.23 f 76.63±0.82 ef 12 1 1.0 45.64±0.99 h 69.25±0.42 d 73.28±0.64 d 81.36±1.09 d 13 1 1.5 51.03±1.17 f 73.81±0.92 c 83.64±1.06 c 92.18±2.00 c 14 1 2.0 65.46±0.61 b 60.95±0.30 e 67.87±0.98 f 75.57±0.90 f 15 1 3.0 59.89±1.22 d 54.64±1.00 g 65.07±0.82 f 73.85±0.63 g
同列不同小写字母表示差异显著(P <0.05)。 Different lowercase letters in the same column indicate significant differences (P <0.05). 新窗口打开 |
下载CSV 3.2 不同浓度植物激素对外植体不定芽分化的影响 将获得的愈伤组织切割成小块后接种到分化培养基上。随着培养时间的推移, 愈伤组织逐渐紧实, 生长变缓, 表面出现颗粒状绿色愈伤组织, 部分绿色愈伤组织分化出芽点(
图2B 4, D3, E2)。以增殖一代小叶为外植体分化出的不定芽最多且长势最好(
图2E 3), 其次为腋芽诱导小叶外植体(
图2D 4), 以嫁接小叶为外植体诱导的不定芽效果较差, 部分出现褐化且不定芽较少(
图2C 5), 多年生母株愈伤组织在不同激素组合的培养基上均未分化出不定芽, 且随着培养时间的延长逐渐褐化死亡(
图2B 3)。
不同外植体愈伤组织平均分化率排序为: 增殖一代小叶(51.29%)>腋芽诱导小叶(32.01%)>一年生嫁接苗小叶(19.72%) (
图3 )。增殖一代小叶最佳愈伤组织分化培养基为: MS+1.0 mg·L
-1 6-BA+0.1 mg·L
-1 2,4-D+0.1 mg·L
-1 TDZ, 分化率最大为78.14%; 腋芽诱导小叶最佳愈伤分化培养基为: MS+2.0 mg·L
-1 6-BA+0.5 mg·L
-1 2,4-D, 分化率最大为45.42%; 一年生嫁接苗小叶最佳愈伤分化培养基为: MS+2.0 mg·L
-1 6-BA+0.1 mg·L
-1 2,4-D+0.5 mg·L
-1 TDZ, 分化率最大为35.23%。除不同外植体对愈伤组织的不定芽分化起决定作用外, 不同植物激素组合对不定芽分化也有重要影响, 不同外植体对激素的响应差异显著(
表3 )。结果表明, 6-BA对一年生嫁接小叶、腋芽诱导小叶及增殖一代小叶的愈伤分化均产生极显著影响(
P < 0.01); 2,4-D对腋芽诱导小叶及增殖一代小叶愈伤分化的影响均达显著性水平, 对一年生嫁接苗小叶愈伤分化的影响不显著; TDZ对一年生嫁接苗愈伤分化的影响显著, 对腋芽诱导小叶及增殖一代小叶愈伤分化的影响均不显著。综合各因素最优水平, 以增殖一代小叶为外植体, 在MS+1.0 mg·L
-1 6-BA+0.1 mg·L
-1 2,4-D+0.1 mg·L
-1 TDZ培养基上愈伤组织分化效果最佳(分化率为78.14%)。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3不同植物激素对郁金樱不同外植体愈伤组织分化的影响 分化培养基编号1-9同
表1。不同小写字母表示不同处理间差异显著(P <0.05)。Figure 3Effects of different plant hormones on Cerasus serrulata callus differentiation in different explants Table 1. Different lowercase letters indicate significant differences at P <0.05 among different treatments.Table 3 表3 表3 不同植物激素对郁金樱不同外植体愈伤组织分化的方差分析Variance analysis of Cerasus serrulata callus differentiation of different explants with different plant hormones Different explants Source df Sum of squares Mean square F P Annual grafted 6-BA 2 2622.375 1311.187 77.591 0.000** 2,4-D 2 57.819 28.910 1.711 0.206 TDZ 2 128.789 64.394 3.811 0.040* Error 20 337.975 16.899 - - Sun 27 13723.723 - - - Axillary buds induce leaflets 6-BA 2 1476.646 738.323 42.619 0.000** 2,4-D 2 144.656 72.328 4.175 0.031* TDZ 2 19.924 9.962 0.575 0.572 Error 20 346.474 17.324 - - Sun 27 29649.142 - - - Proliferate one generation of leaflets 6-BA 2 3360.867 1680.433 29.026 0.000** 2,4-D 2 600.965 300.483 5.19 0.015* TDZ 2 182.009 91.005 1.572 0.232 Error 20 1157.866 57.893 - - Sun 27 79113.749 - - -
TDZ: 噻苯隆; TDZ: Thidiazuron * P <0.05; ** P <0.01 新窗口打开 |下载CSV 3.3 不同浓度植物激素对外植体不定芽增殖的影响 将不同外植体分化的不定芽团带愈伤组织切成大小相等的小块, 接种于增殖培养基。结果表明, 外植体类型及外源激素浓度对不定芽增殖均有显著影响。一年生嫁接苗小叶、腋芽诱导小叶和增殖一代小叶3种不同外植体平均增殖系数分别为: 2.22<4.19<4.79 (表4 )。其中, 一年生嫁接苗小叶芽团在MS+2.0 mg·L-1 6-BA+0.5 mg·L-1 NAA增殖培养基上增殖系数最大(3.53) (图2C 7); 腋芽诱导小叶芽团在MS+2.0 mg·L-1 6-BA无NAA的增殖培养基上增殖系数最大(5.39) (图2D 7); 增殖一代小叶芽团在MS+1.0 mg·L-1 6-BA增殖培养基上增殖系数最大(7.85) (图2E 7)。6-BA和NAA对郁金樱一年生嫁接苗小叶芽团的不定芽增殖具有显著促进作用。随着6-BA和NAA浓度的增大, 不定芽增殖系数增大; NAA对腋芽诱导小叶芽团不定芽增殖则呈现出先上升后下降的趋势, 6-BA浓度与不定芽增殖系数呈正相关, 说明高浓度6-BA及低浓度NAA有利于腋芽诱导小叶芽团的不定芽增殖。对增殖一代小叶芽团而言, 6-BA和NAA则呈现出高浓度抑制低浓度促进的共同现象, 表明过高浓度6-BA和NAA不利于增殖一代小叶芽团增殖。综合不定芽增殖系数及芽团生长状况, 以增殖一代小叶为外植体分化的芽团接种于MS+1.0 mg·L-1 6-BA培养基上不定芽增殖效果最佳。Table 4 表4 表4 不同植物激素对郁金樱外植体不定芽增殖的影响(平均值±标准误)Effects of different plant hormones on Cerasus serrulata adventitious bud proliferation of different explants (means±SE) No. 6-BA -1 ) NAA -1 ) Proliferation coefficient Annual grafted seedling leaflets Axillary buds induce leaflets Proliferate one generation of leaflets 1 0.5 0 0 e 2.42±0.04 g 3.32±0.46 e 2 0.5 0.1 0.88±0.77 d 3.52±0.07 f 2.70±0.24 f 3 0.5 0.5 2.17±0.27 c 2.39±0.05 g 2.91±0.29 ef 4 1 0 2.38±0.04 c 4.46±0.12 e 7.85±0.33 a 5 1 0.1 2.17±0.62 c 4.99±0.11 c 5.57±0.20 cd 6 1 0.5 3.13±0.05 ab 4.64±0.08 d 6.30±0.14 b 7 2 0 2.61±0.07 bc 5.39±0.05 a 5.26±0.27 d 8 2 0.1 3.07±0.06 b 5.18±0.19 b 5.82±0.19 c 9 2 0.5 3.53±0.04 a 4.73±0.08 d 3.38±0.15 e Average 2.22±1.12 C 4.19±1.10 B 4.79±1.73 A
同列不同小写字母表示差异显著(P <0.05), 同行数值后不同大写字母表示差异显著(P <0.05)。 Different lowercase letters in the same column indicate significant differences (P <0.05), different capital letters after peer values indicate significant differences (P <0.05). 新窗口打开 |下载CSV 3.4 生根培养及炼苗移栽 切取不定芽增殖培养基上生长健壮且高度大于1.0 cm的组培苗, 接种于生根培养基(表5 )。生长7天左右, 不添加任何激素的1/2MS培养基中组培苗底部出现不定根原始体突起, 20天后出现大量须根(图4D ); 在1/2MS+0.5 mg·L-1 NAA培养基中, 组培苗培养7天后出现少量透明愈伤, 12天左右底部出现不定根原始体突起(图4A ), 约20天后出现5-6条粗壮的肉质主根(图4B ), 约25天后从主根上长出大量侧根(图4C )。在上述2种培养基生根完成后瓶盖半敞开于室内炼苗2天, 然后完全打开瓶盖炼苗5天, 洗净根部培养基, 移植到泥炭土:蛭石:河沙=2:1:1 (v/v/v)的混合基质营养钵中, 用塑料袋套住营养钵保湿保温, 30天后移栽成活率达96%以上。与上述2种生根培养基相比, 其它生根培养基上组培苗生根数少且多为肉质主根, 移栽成活率仅为65%。Table 5 表5 表5 不同植物激素对郁金樱生根的影响(平均值±标准误)Effects of plant hormones on rooting of Cerasus serrulata (means±SE) IBA -1 ) NAA -1 ) Rooting rate Number of roots 0 0 100 a 12.82±0.54 a 0 0.5 86.72±1.62 b 9.24±1.24 b 0 1 0 e 0 e 0.5 0 68.32±2.95 c 5.64±0.26 c 0.5 0.5 44.09±1.74 d 3.24±0.48 d
同列不同小写字母表示差异显著(P <0.05)。 Different lowercase letters in the same column indicate significant differences (P <0.05). 新窗口打开 |下载CSV 图4 新窗口打开 |下载原图ZIP |生成PPT 图4郁金樱不同外植体诱导再生植株生根及移载 Figure 4Rooting and transplantation of regenerated plants induced by different explants of Cerasus serrulata 图5 )表明, 增殖一代小叶诱导的再生组培苗的根、茎、叶增长量均为最优, 幼苗高度达23- 25 cm, 移栽成活后生长迅速且长势最好(图4K -M), 表明增殖一代小叶诱导的再生组培苗比一年生嫁接苗和腋芽诱导的再生组培苗更具生长优势。图5 新窗口打开 |下载原图ZIP |生成PPT 图5郁金樱不同外植体诱导的再生植株的生长状态 Figure 5The growth status of regenerated plants induced by different explants of Cerasus serrulata 3.5 讨论 研究发现, 郁金樱具有复杂的季节性周期, 在生长动力学上表现出强烈的阵发性生长(episodic growth)特性, 其生长势在上一个生长季节已预先确定(闫国华等, 2002 ), 而这些由植物基因决定的生长生理规律通过后期植物激素和营养手段较难调控(郭希梅等, 2011 ), 从而增加了组织培养的难度。以冬樱花(Prunus majestica )叶片、花托、花梗和子房为外植体未分化出不定芽(和凤美等, 2010 )。现阶段成功分化出不定芽的樱花愈伤组织再生研究皆以叶片为外植体。福建山樱花以腋芽诱导小叶和茎段为外植体诱导愈伤组织, 仅腋芽诱导小叶分化出不定芽(邹娜等, 2013 )。雪落樱和御殿场樱以叶片为外植体虽分化出不定芽, 但仍存在褐化率高和分化困难等问题(史港影等, 2014 ; 雷巾茗, 2020 ), 这表明在樱花的愈伤组织再生研究中外植体的选择尤为重要, 可能与樱花基因型和外植体生理状态有关。研究表明, 在树番茄(Cyphomandra betacea ) (Correia et al., 2011 )、小粒咖啡(Coffea arabica ) (Bartos et al., 2018 )和冬青栎(Quercus ilex ) (Martínez et al., 2017 )等物种上, 胚性愈伤的诱导也很大程度依赖于外植体种类, 这种由基因驱动的外植体“抗拒”是很多品种无法建立稳定培养体系的主要原因(McCown, 2000 )。采用郁金樱不同分化程度的小叶诱导再生植株, 我们发现郁金樱在离体培养中表现出强烈的顽拗现象(recalcitrance), 这与樱桃砧木酸樱桃(P. cerasus )相似(Ben Mahmoud et al., 2017 )。本研究以多年生郁金樱小叶为外植体未分化出不定芽, 愈伤组织诱导率低且胚性愈伤较少, 同时随着培养时间的延长褐化现象严重, 顽拗现象出现于多年生小叶组织培养的诱导和分化阶段, 这与前人的研究结果一致(Hu et al., 2017 ; Wu et al., 2020 )。为克服遗传性基因导致的顽拗现象, 本研究采用郁金樱一年生嫁接苗小叶、腋芽诱导小叶及增殖一代小叶为外植体, 相当于间接对多年生郁金樱母株进行不同程度的幼化(rejuvennation)或返童, 幼年外植体相比发育成熟或分化组织具有更大的潜能(Wu et al., 2019 )。通过采集多年生郁金樱母株嫁接于一年生山樱花砧木上, 相当于直接操作原植物来克服郁金樱阵发性生长特性。利用一年生砧木生长旺盛、生物活性强的特点获得幼化接穗; 以腋芽诱导小叶为外植体, 相当于采用器官发生来克服顽拗现象(Singh et al., 2015 ; Díaz-Sala, 2019 ), 因为分生组织的诱导就是一种幼化过程; 而通过诱导、分化和增殖等器官发生刺激途径, 使增殖一代小叶外植体达到高度幼化, 是其在愈伤诱导、分化及不定芽增殖中表现出较强生长势及分裂活性的原因。本研究中, 不同幼化处理的外植体在愈伤诱导、分化、增殖及生长上均表现出显著差异, 即采用的外植体幼化程度越高, 后期生长潜力越大(于波等, 2020 ), 各项生长生理指标越优, 以增殖一代小叶为外植体效果最佳。研究发现, 随着郁金樱幼化程度的加大, 各项生理指标渐优, 而连续继代后随着幼化的深入, 能否逆转成年态? 这些问题尚无相关报道, 有待深入研究。我们也曾连续对郁金樱进行多次增殖继代, 发现随着继代次数的增加, 出现叶发黄、毒害作用加强、对激素敏感性增强及大量死亡等问题, 尚未尝试以增殖继代多次的小叶为外植体进行愈伤组织再生, 这些都有待后期深入研究。表3 ), 其具有较强的生长素和细胞分裂素双重活性, 对克服顽拗现象有显著效果(Bernula et al., 2020 )。随着外植体幼化程度的提高, 不同外植体诱导的愈伤组织对激素敏感性增强(Ming et al., 2019 ), 表现为具有高活性的TDZ对愈伤分化影响不显著, 低浓度6-BA及2,4-D对分化的影响逐渐加强, 到不定芽增殖时, 腋芽诱导小叶和增殖一代小叶不定芽能在不含NAA的培养基中大量增殖。生根培养更出现组培苗在不含任何激素的1/2MS培养基上生根率达到100%的现象, 在含有NAA或IBA的培养基中, 根系诱导受到不同程度的抑制。综上表明, 在植物组织培养过程中, 植物激素可能随着培养的深入出现富集, 导致后期植物对激素敏感性增强, 对激素依赖度降低。还有可能是郁金樱组培再生过程中出现了激素自养性的适应性现象, 即获得了不依赖外援生长素和细胞分裂素的能力。通过对郁金樱植株再生各阶段激素浓度的横向比较, 发现郁金樱再生过程中不同外植体对激素的敏感性均发生改变, 幼化程度最高的增殖一代小叶对激素敏感性最强。[1] 陈雪 , 张金柱 , 潘兵兵 , 桑成瑾 , 马雪 , 杨涛 , 车代弟 (2011 ). 月季愈伤组织的诱导及植株再生植物学报 46, 569 -574 . [本文引用: 1] [2] 房洪舟 , 鲁敏 , 安华明 (2019 ). 刺梨叶片愈伤组织培养体系建立及其主要活性物质分析植物生理学报 55, 1147 -1155 . [本文引用: 1] [3] 郭希梅 , 丛日晨 , 张常青 , 古润泽 , 高俊平 (2011 ). 古油松衰弱衰老诊断的生理指标林业科学 47(4 ), 43 -48 . [本文引用: 1] [4] 和凤美 , 李璇 , 邵琬珊 , 朱永平 , 杨晓红 (2010 ). 冬樱花愈伤组织诱导和抑制褐化初探中国农学通报 26(12 ), 130 -134 . [本文引用: 1] [5] 黄守印 , 池井存 , 苏淑欣 , 尚文艳 , 任艳平 (2003 ). 雾灵山地区野生樱花的组织培养与快速繁殖植物生理学通讯 (3 ), 228 . [本文引用: 1] [6] 蒋冬月 , 邹宜含 , 柳新红 , 程亚平 , 王平 , 沈鑫 (2019 ). 樱花粉红及黄绿色系品种苗期生长特性及适应性江西农业大学学报 41, 673 -682 . [本文引用: 1] [7] 雷巾茗 (2020 ). 樱花组培快繁与扦插繁殖研究硕士论文. 北京: 北京林业大学 . pp. 1 -92 . [本文引用: 2] [8] 李水根 , 李秀芬 , 殷丽青 , 高晨 , 朱建军 (2020 ). 喜马拉雅樱花嫩茎离体快繁体系优化分子植物育种 18, 8217 -8222 . [本文引用: 1] [9] 李艳敏 , 孟月娥 , 张玉 , 赵秀山 , 王利民 , 王慧娟 (2012 ). 新优彩叶植物红叶樱花外植体采集及离体培养技术研究河南农业科学 41(9 ), 127 -130, 142 . [本文引用: 1] [10] 李艳敏 , 孟月娥 , 赵秀山 , 王慧娟 , 张强 , 王利民 (2008 ). ‘红叶樱花’的组织培养和快速繁殖植物生理学通讯 44, 1163 -1164 . [本文引用: 1] [11] 刘莉莉 , 卢淑波 , 徐佳萍 , 张庆田 , 李昌禹 (2015 ). 以黄花乌头发根为外植体的再生培养体系建立植物学报 50, 623 -627 . [本文引用: 1] [12] 刘晓莉 (2012 ). 14个樱花品种观赏性状综合评价和樱花园林应用研究硕士论文. 杭州: 浙江农林大学 . pp. 1 -89 . [本文引用: 1] [13] 吕月良 , 陈璋 , 施季森 , 黄宇翔 , 刘金燕 , 谢建丽 (2006 ). 福建山樱花不定芽诱导和植株再生规模化繁殖试验南京林业大学学报(自然科学版) (3 ), 105 -108 . [本文引用: 1] [14] 任如意 , 薛巨坤 , 国会艳 , 魏继承 (2017 ). 北玄参毛状根诱导及其植株再生植物学报 52, 783 -787 . [本文引用: 1] [15] 史港影 , 南程慧 , 伊贤贵 , 张开文 , 王贤荣 (2014 ). 雪落樱再生体系的建立南京林业大学学报(自然科学版) 38, 20 -24 . [本文引用: 2] [16] 宋斯妤 (2018 ). 迎春樱和华中樱优良品系组培快繁技术的研究硕士论文. 杭州: 浙江农林大学 . pp. 1 -62 . [本文引用: 1] [17] 徐晨捷 , 欧静 (2020 ). 染井吉野樱的茎段培养与胚培养比较北方园艺 (23 ), 65 -71 . [本文引用: 1] [18] 闫国华 , 周宇 , 张晓明 , 张开春 (2002 ). 植物离体培养中的顽拗现象及其生理和遗传基础植物生理学通讯 38, 481 -486 . [本文引用: 1] [19] 燕丽萍 , 李丽 , 刘翠兰 , 吴德军 , 王因花 , 任飞 , 赵梁军 (2016 ). 绒毛白蜡体胚诱导和植株再生植物学报 51, 807 -816 . 10.11983/CBB15222 [本文引用: 1] [20] 杨小燕 , 欧静 , 张凤泉 , 翁钰舟 , 于瀚 , 曹时波 (2019 ). 贵阳市樱花资源及其园林应用研究山地农业生物学报 38(6 ), 14 -20 . [本文引用: 1] [21] 于波 , 黄丽丽 , 朱玉 , 朱根发 , 孙映波 (2020 ). 朱顶红幼嫩花梗胚性愈伤组织诱导和高效植株再生园艺学报 47, 907 -915 . [本文引用: 1] [22] 张灵灵 , 蒋细旺 (2015 ). 2个日本晚樱品种组织培养和快繁技术研究西南林业大学学报 35(4 ), 27 -32 . [本文引用: 1] [23] 张旭红 , 王頔 , 梁振旭 , 孙美玉 , 张金政 , 石雷 (2018 ). 欧洲百合愈伤组织诱导及植株再生体系的建立植物学报 53, 840 -847 . [本文引用: 1] [24] 朱继军 , 奉树成 , 陈必胜 (2015 ). 晚樱花品种的引种与筛选中国园艺文摘 31(5 ), 1 -3, 24 . [本文引用: 1] [25] 邹娜 , 陈璋 , 林思祖 , 林庆良 (2013 ). 福建山樱花愈伤组织的诱导及植株再生核农学报 27, 1417 -1423 . [本文引用: 2] [26] Bartos PMC Gomes HT do Amaral LIV Teixeira JB Scherwinski-Pereira JE 2018 ). Biochemical events during somatic embryogenesis in Coffea arabica L3 Biotech 209 . 10.1007/s13205-018-1238-7 URL [本文引用: 1] [27] Ben Mahmoud K Jedidi E Delporte F Muhovski Y Jemmali A Druart P 2017 ). Molecular investigations of the somatic embryogenesis recalcitrance in the cherry ( Prunus cerasus L.) rootstock CAB 6PTurk J Biol 158 -165 . 10.3906/biy-1604-105 URL [本文引用: 1] [28] Bernula D Benkő P Kaszler N Domonkos I Szőllősi R Ferenc G Ayaydin F Fehér A Gémes K 2020 ). Timely removal of exogenous cytokinin and the prevention of auxin transport from the shoot to the root affect the regeneration potential of Arabidopsis rootsPlant Cell Tissue Organ Cult 327 -339 . 10.1007/s11240-019-01730-3 URL [本文引用: 1] [29] Chen BH Li JM Zhang J Wu ZX Fan HH Li QZ 2016 ). Optimizing the rapid technique for propagation of Cerasus campanulata by tissue culturePak J Bot 305 -309 . [本文引用: 1] [30] Correia S Lopes ML Canhoto JM 2011 ). Somatic embryogenesis induction system for cloning an adult Cyphomandra betacea (Cav.) Sendt. (tamarillo)Trees 1009 -1020 . 10.1007/s00468-011-0575-5 URL [本文引用: 1] [31] Díaz-Sala C 2019 ). Molecular dissection of the regenerative capacity of forest tree species: special focus on conifersFront Plant Sci 1943 . 10.3389/fpls.2018.01943 URL [本文引用: 1] [32] Hu RY Sun YH Wu B Duan HJ Zheng HQ Hu DL Lin HZ Tong ZK Xu JL Li Y 2017 ). Somatic embryogenesis of immature Cunninghamia lanceolata (Lamb.) hook zygotic embryosSci Rep 56 . 10.1038/s41598-017-00156-1 URL [本文引用: 1] [33] Martínez MT San José MC Vieitez AM Cernadas MJ Ballester A Corredoira E 2017 ). Propagation of mature Quercus ilex L. (holm oak) trees by somatic embryogenesisPlant Cell Tissue Organ Cult 321 -333 . 10.1007/s11240-017-1286-4 URL [本文引用: 1] [34] McCown BH 2000 ). Special symposium: in vitro plant recalcitrance recalcitrance of woody and herbaceous perennial plants: dealing with genetic predeterminismIn Vitro Cell Dev Biol Plant 149 -154 . 10.1007/s11627-000-0030-6 URL [本文引用: 1] [35] Ming NJ Mostafiz SB Johon NS Zulkifli NSA Wagiran A 2019 ). Combination of plant growth regulators, maltose, and partial desiccation treatment enhance somatic embryogenesis in selected Malaysian rice cultivarPlants 144 . 10.3390/plants8060144 URL [本文引用: 1] [36] Singh R Rai MK Kumari N 2015 ). Somatic embryogenesis and plant regeneration in Sapindus mukorossi gaertn. from leaf-derived callus induced with 6-benzylaminopurineAppl Biochem Biotechnol 498 -510 . 10.1007/s12010-015-1758-0 URL [本文引用: 1] [37] Wu GY Wei XL Wang X Wei Y 2020 ). Induction of somatic embryogenesis in different explants from Ormosia henryi PrainPlant Cell Tissue Organ Cult 229 -240 . 10.1007/s11240-020-01822-5 URL [本文引用: 1] [38] Wu H Chen BJ Fiers M Wróbel-Marek J Kodde J Groot SPC Angenent G Feng H Bentsink L Boutilier K 2019 ). Seed maturation and post-harvest ripening negatively affect Arabidopsis somatic embryogenesisPlant Cell Tissue Organ Cult 17 -27 . 10.1007/s11240-019-01658-8 URL [本文引用: 1] Prunus campanulata ) (黄守印等, 2003 ; 吕月良等, 2006 ; Chen et al., 2016 )、日本晚樱杨贵妃(C. serrulata var. lannesiana cv. ‘Mollis’)、日本晚樱醉红(C. serrulata var. lannesiana cv. ‘Zuihong’) (张灵灵和蒋细旺, 2015 )、华中樱(C. conradinae )、迎春樱(C. discoidea ) (宋斯妤, 2018 )、染井吉野樱(C. × yedoensis cv. ‘Somei-yoshino’) (徐晨捷和欧静, 2020 )和喜马拉雅樱花(C. cerasoides ) (李水根等, 2020 )等少数品种(李艳敏等, 2008 , 2012 ).关于樱花植株再生和遗传转化方面的研究较少, 仅见福建山樱花以腋芽诱导小叶为外植体, 成功建立了植株再生体系和体细胞胚胎发生途径(邹娜等, 2013 ); 以雪落樱(C. xueluoensis ) (史港影等, 2014 )和御殿场樱(C. × incisa cv. ‘Gotenba-zakura’) (雷巾茗, 2020 )叶片为外植体建立了愈伤组织再生体系, 目前尚未见有关郁金樱再生体系及组织培养的报道.通过对北玄参(Scrophularia buergeriana ) (任如意等, 2017 )、黄花乌头(Aconitum coreanum ) (刘莉莉等, 2015 )、绒毛白蜡(Fraxinus velutina ) (燕丽萍等, 2016 )和月季(Rosa chinensis ) (陈雪等, 2011 )等植株再生的研究表明, 不同外植体对植株再生影响极显著, 直接关系试验成功与否, 故选择正确外植体是试验成功的前提.本研究以郁金樱不同幼化程度的小叶为材料, 以期通过愈伤组织诱导不定芽分化、不定芽增殖和生根, 建立高效、稳定的郁金樱再生体系, 为其生物技术育种及后续的遗传转化研究提供参考. ...图1 ).将不同外植体小叶接种于诱导培养基1周后, 叶片边缘朝叶脉中心卷曲拱起; 2周后, 小叶主叶脉切口及叶柄端出现细小球形愈伤突起, 愈伤突起由主叶脉向切口两端衍生逐渐变小.球形突起多发生于各支叶脉切口处, 可能是叶脉作为物质运输交换的组织, 最先感受到外界环境的激素变化从而脱分化产生愈伤组织(房洪舟等, 2019 ), 后期叶片质地逐渐变硬变脆.不同外植体诱导愈伤组织从颜色、质地到形态皆有差异(图2A -E7).以多年生母株小叶为外植体诱导的愈伤组织颜色多呈黄褐色, 部分为红色, 质地较紧实坚硬, 表面有粗糙颗粒突起(图2B 1).其它3种外植体诱导的愈伤组织质地较松软, 表面分布有大量细小颗粒, 其中一年生嫁接苗小叶诱导的愈伤组织呈浅黄色(图2C 1), 腋芽小叶愈伤组织和增殖一代小叶愈伤组织为透明白色, 表面有光泽(图2D 1, E1).多年生母株诱导的愈伤组织无分化能力, 为非胚性愈伤组织, 后期培养逐渐褐化(图2B 2, B3).嫁接苗诱导愈伤组织(图2C 2)随着后期培养激素的调整可转化为胚性愈伤组织, 腋芽及增殖一代小叶诱导的愈伤组织多为胚性愈伤组织, 以增殖一代小叶为外植体诱导效果最佳(图2D 2, D3, E2). ...闫国华等, 2002 ), 而这些由植物基因决定的生长生理规律通过后期植物激素和营养手段较难调控(郭希梅等, 2011 ), 从而增加了组织培养的难度.以冬樱花(Prunus majestica )叶片、花托、花梗和子房为外植体未分化出不定芽(和凤美等, 2010 ).现阶段成功分化出不定芽的樱花愈伤组织再生研究皆以叶片为外植体.福建山樱花以腋芽诱导小叶和茎段为外植体诱导愈伤组织, 仅腋芽诱导小叶分化出不定芽(邹娜等, 2013 ).雪落樱和御殿场樱以叶片为外植体虽分化出不定芽, 但仍存在褐化率高和分化困难等问题(史港影等, 2014 ; 雷巾茗, 2020 ), 这表明在樱花的愈伤组织再生研究中外植体的选择尤为重要, 可能与樱花基因型和外植体生理状态有关.研究表明, 在树番茄(Cyphomandra betacea ) (Correia et al., 2011 )、小粒咖啡(Coffea arabica ) (Bartos et al., 2018 )和冬青栎(Quercus ilex ) (Martínez et al., 2017 )等物种上, 胚性愈伤的诱导也很大程度依赖于外植体种类, 这种由基因驱动的外植体“抗拒”是很多品种无法建立稳定培养体系的主要原因(McCown, 2000 ).采用郁金樱不同分化程度的小叶诱导再生植株, 我们发现郁金樱在离体培养中表现出强烈的顽拗现象(recalcitrance), 这与樱桃砧木酸樱桃(P. cerasus )相似(Ben Mahmoud et al., 2017 ).本研究以多年生郁金樱小叶为外植体未分化出不定芽, 愈伤组织诱导率低且胚性愈伤较少, 同时随着培养时间的延长褐化现象严重, 顽拗现象出现于多年生小叶组织培养的诱导和分化阶段, 这与前人的研究结果一致(Hu et al., 2017 ; Wu et al., 2020 ).为克服遗传性基因导致的顽拗现象, 本研究采用郁金樱一年生嫁接苗小叶、腋芽诱导小叶及增殖一代小叶为外植体, 相当于间接对多年生郁金樱母株进行不同程度的幼化(rejuvennation)或返童, 幼年外植体相比发育成熟或分化组织具有更大的潜能(Wu et al., 2019 ).通过采集多年生郁金樱母株嫁接于一年生山樱花砧木上, 相当于直接操作原植物来克服郁金樱阵发性生长特性.利用一年生砧木生长旺盛、生物活性强的特点获得幼化接穗; 以腋芽诱导小叶为外植体, 相当于采用器官发生来克服顽拗现象(Singh et al., 2015 ; Díaz-Sala, 2019 ), 因为分生组织的诱导就是一种幼化过程; 而通过诱导、分化和增殖等器官发生刺激途径, 使增殖一代小叶外植体达到高度幼化, 是其在愈伤诱导、分化及不定芽增殖中表现出较强生长势及分裂活性的原因.本研究中, 不同幼化处理的外植体在愈伤诱导、分化、增殖及生长上均表现出显著差异, 即采用的外植体幼化程度越高, 后期生长潜力越大(于波等, 2020 ), 各项生长生理指标越优, 以增殖一代小叶为外植体效果最佳.研究发现, 随着郁金樱幼化程度的加大, 各项生理指标渐优, 而连续继代后随着幼化的深入, 能否逆转成年态? 这些问题尚无相关报道, 有待深入研究.我们也曾连续对郁金樱进行多次增殖继代, 发现随着继代次数的增加, 出现叶发黄、毒害作用加强、对激素敏感性增强及大量死亡等问题, 尚未尝试以增殖继代多次的小叶为外植体进行愈伤组织再生, 这些都有待后期深入研究. ...闫国华等, 2002 ), 而这些由植物基因决定的生长生理规律通过后期植物激素和营养手段较难调控(郭希梅等, 2011 ), 从而增加了组织培养的难度.以冬樱花(Prunus majestica )叶片、花托、花梗和子房为外植体未分化出不定芽(和凤美等, 2010 ).现阶段成功分化出不定芽的樱花愈伤组织再生研究皆以叶片为外植体.福建山樱花以腋芽诱导小叶和茎段为外植体诱导愈伤组织, 仅腋芽诱导小叶分化出不定芽(邹娜等, 2013 ).雪落樱和御殿场樱以叶片为外植体虽分化出不定芽, 但仍存在褐化率高和分化困难等问题(史港影等, 2014 ; 雷巾茗, 2020 ), 这表明在樱花的愈伤组织再生研究中外植体的选择尤为重要, 可能与樱花基因型和外植体生理状态有关.研究表明, 在树番茄(Cyphomandra betacea ) (Correia et al., 2011 )、小粒咖啡(Coffea arabica ) (Bartos et al., 2018 )和冬青栎(Quercus ilex ) (Martínez et al., 2017 )等物种上, 胚性愈伤的诱导也很大程度依赖于外植体种类, 这种由基因驱动的外植体“抗拒”是很多品种无法建立稳定培养体系的主要原因(McCown, 2000 ).采用郁金樱不同分化程度的小叶诱导再生植株, 我们发现郁金樱在离体培养中表现出强烈的顽拗现象(recalcitrance), 这与樱桃砧木酸樱桃(P. cerasus )相似(Ben Mahmoud et al., 2017 ).本研究以多年生郁金樱小叶为外植体未分化出不定芽, 愈伤组织诱导率低且胚性愈伤较少, 同时随着培养时间的延长褐化现象严重, 顽拗现象出现于多年生小叶组织培养的诱导和分化阶段, 这与前人的研究结果一致(Hu et al., 2017 ; Wu et al., 2020 ).为克服遗传性基因导致的顽拗现象, 本研究采用郁金樱一年生嫁接苗小叶、腋芽诱导小叶及增殖一代小叶为外植体, 相当于间接对多年生郁金樱母株进行不同程度的幼化(rejuvennation)或返童, 幼年外植体相比发育成熟或分化组织具有更大的潜能(Wu et al., 2019 ).通过采集多年生郁金樱母株嫁接于一年生山樱花砧木上, 相当于直接操作原植物来克服郁金樱阵发性生长特性.利用一年生砧木生长旺盛、生物活性强的特点获得幼化接穗; 以腋芽诱导小叶为外植体, 相当于采用器官发生来克服顽拗现象(Singh et al., 2015 ; Díaz-Sala, 2019 ), 因为分生组织的诱导就是一种幼化过程; 而通过诱导、分化和增殖等器官发生刺激途径, 使增殖一代小叶外植体达到高度幼化, 是其在愈伤诱导、分化及不定芽增殖中表现出较强生长势及分裂活性的原因.本研究中, 不同幼化处理的外植体在愈伤诱导、分化、增殖及生长上均表现出显著差异, 即采用的外植体幼化程度越高, 后期生长潜力越大(于波等, 2020 ), 各项生长生理指标越优, 以增殖一代小叶为外植体效果最佳.研究发现, 随着郁金樱幼化程度的加大, 各项生理指标渐优, 而连续继代后随着幼化的深入, 能否逆转成年态? 这些问题尚无相关报道, 有待深入研究.我们也曾连续对郁金樱进行多次增殖继代, 发现随着继代次数的增加, 出现叶发黄、毒害作用加强、对激素敏感性增强及大量死亡等问题, 尚未尝试以增殖继代多次的小叶为外植体进行愈伤组织再生, 这些都有待后期深入研究. ...Prunus campanulata ) (黄守印等, 2003 ; 吕月良等, 2006 ; Chen et al., 2016 )、日本晚樱杨贵妃(C. serrulata var. lannesiana cv. ‘Mollis’)、日本晚樱醉红(C. serrulata var. lannesiana cv. ‘Zuihong’) (张灵灵和蒋细旺, 2015 )、华中樱(C. conradinae )、迎春樱(C. discoidea ) (宋斯妤, 2018 )、染井吉野樱(C. × yedoensis cv. ‘Somei-yoshino’) (徐晨捷和欧静, 2020 )和喜马拉雅樱花(C. cerasoides ) (李水根等, 2020 )等少数品种(李艳敏等, 2008 , 2012 ).关于樱花植株再生和遗传转化方面的研究较少, 仅见福建山樱花以腋芽诱导小叶为外植体, 成功建立了植株再生体系和体细胞胚胎发生途径(邹娜等, 2013 ); 以雪落樱(C. xueluoensis ) (史港影等, 2014 )和御殿场樱(C. × incisa cv. ‘Gotenba-zakura’) (雷巾茗, 2020 )叶片为外植体建立了愈伤组织再生体系, 目前尚未见有关郁金樱再生体系及组织培养的报道.通过对北玄参(Scrophularia buergeriana ) (任如意等, 2017 )、黄花乌头(Aconitum coreanum ) (刘莉莉等, 2015 )、绒毛白蜡(Fraxinus velutina ) (燕丽萍等, 2016 )和月季(Rosa chinensis ) (陈雪等, 2011 )等植株再生的研究表明, 不同外植体对植株再生影响极显著, 直接关系试验成功与否, 故选择正确外植体是试验成功的前提.本研究以郁金樱不同幼化程度的小叶为材料, 以期通过愈伤组织诱导不定芽分化、不定芽增殖和生根, 建立高效、稳定的郁金樱再生体系, 为其生物技术育种及后续的遗传转化研究提供参考. ...Cerasus serrulata var. lannesiana cv. ‘Grandiflora’)是蔷薇科(Rosaceae)樱属(Cerasus )落叶乔木, 花叶同开, 花色淡黄至绿色, 花期在3月下旬至4月上旬, 在我国长三角地区有栽培, 现已推广到全国.樱花品种花色大多为白色、粉色或红色, 郁金樱因其绿色花而独特, 在盛花期和落花期均有极高的观赏价值, 园林开发潜力巨大(刘晓莉, 2012 ).郁金樱具有良好的适应性和观赏性(朱继军等, 2015 ), 其在校园绿地、居住区绿地、道路绿地、综合公园和城市广场上皆有分布, 苗木市场需求较大(蒋冬月等, 2019 ; 杨小燕等, 2019 ).生产上樱花主要通过嫁接繁殖, 但该繁殖方法存在占用空间资源大、成苗周期长及种苗价格高等问题, 且易受时间、气候和环境等因素影响, 经多代繁殖后, 易造成病毒积累, 影响樱花的产量和品质.随着现代生物技术的发展, 利用基因工程技术进行植物性状改良已成为可行的育种方式(张旭红等, 2018 ), 而建立高效且快速的再生体系是进行遗传转化研究的前提.因此, 应用植物组织培养技术建立无性快繁体系, 对郁金樱的遗传改良具有重要意义. ...Prunus campanulata ) (黄守印等, 2003 ; 吕月良等, 2006 ; Chen et al., 2016 )、日本晚樱杨贵妃(C. serrulata var. lannesiana cv. ‘Mollis’)、日本晚樱醉红(C. serrulata var. lannesiana cv. ‘Zuihong’) (张灵灵和蒋细旺, 2015 )、华中樱(C. conradinae )、迎春樱(C. discoidea ) (宋斯妤, 2018 )、染井吉野樱(C. × yedoensis cv. ‘Somei-yoshino’) (徐晨捷和欧静, 2020 )和喜马拉雅樱花(C. cerasoides ) (李水根等, 2020 )等少数品种(李艳敏等, 2008 , 2012 ).关于樱花植株再生和遗传转化方面的研究较少, 仅见福建山樱花以腋芽诱导小叶为外植体, 成功建立了植株再生体系和体细胞胚胎发生途径(邹娜等, 2013 ); 以雪落樱(C. xueluoensis ) (史港影等, 2014 )和御殿场樱(C. × incisa cv. ‘Gotenba-zakura’) (雷巾茗, 2020 )叶片为外植体建立了愈伤组织再生体系, 目前尚未见有关郁金樱再生体系及组织培养的报道.通过对北玄参(Scrophularia buergeriana ) (任如意等, 2017 )、黄花乌头(Aconitum coreanum ) (刘莉莉等, 2015 )、绒毛白蜡(Fraxinus velutina ) (燕丽萍等, 2016 )和月季(Rosa chinensis ) (陈雪等, 2011 )等植株再生的研究表明, 不同外植体对植株再生影响极显著, 直接关系试验成功与否, 故选择正确外植体是试验成功的前提.本研究以郁金樱不同幼化程度的小叶为材料, 以期通过愈伤组织诱导不定芽分化、不定芽增殖和生根, 建立高效、稳定的郁金樱再生体系, 为其生物技术育种及后续的遗传转化研究提供参考. ...闫国华等, 2002 ), 而这些由植物基因决定的生长生理规律通过后期植物激素和营养手段较难调控(郭希梅等, 2011 ), 从而增加了组织培养的难度.以冬樱花(Prunus majestica )叶片、花托、花梗和子房为外植体未分化出不定芽(和凤美等, 2010 ).现阶段成功分化出不定芽的樱花愈伤组织再生研究皆以叶片为外植体.福建山樱花以腋芽诱导小叶和茎段为外植体诱导愈伤组织, 仅腋芽诱导小叶分化出不定芽(邹娜等, 2013 ).雪落樱和御殿场樱以叶片为外植体虽分化出不定芽, 但仍存在褐化率高和分化困难等问题(史港影等, 2014 ; 雷巾茗, 2020 ), 这表明在樱花的愈伤组织再生研究中外植体的选择尤为重要, 可能与樱花基因型和外植体生理状态有关.研究表明, 在树番茄(Cyphomandra betacea ) (Correia et al., 2011 )、小粒咖啡(Coffea arabica ) (Bartos et al., 2018 )和冬青栎(Quercus ilex ) (Martínez et al., 2017 )等物种上, 胚性愈伤的诱导也很大程度依赖于外植体种类, 这种由基因驱动的外植体“抗拒”是很多品种无法建立稳定培养体系的主要原因(McCown, 2000 ).采用郁金樱不同分化程度的小叶诱导再生植株, 我们发现郁金樱在离体培养中表现出强烈的顽拗现象(recalcitrance), 这与樱桃砧木酸樱桃(P. cerasus )相似(Ben Mahmoud et al., 2017 ).本研究以多年生郁金樱小叶为外植体未分化出不定芽, 愈伤组织诱导率低且胚性愈伤较少, 同时随着培养时间的延长褐化现象严重, 顽拗现象出现于多年生小叶组织培养的诱导和分化阶段, 这与前人的研究结果一致(Hu et al., 2017 ; Wu et al., 2020 ).为克服遗传性基因导致的顽拗现象, 本研究采用郁金樱一年生嫁接苗小叶、腋芽诱导小叶及增殖一代小叶为外植体, 相当于间接对多年生郁金樱母株进行不同程度的幼化(rejuvennation)或返童, 幼年外植体相比发育成熟或分化组织具有更大的潜能(Wu et al., 2019 ).通过采集多年生郁金樱母株嫁接于一年生山樱花砧木上, 相当于直接操作原植物来克服郁金樱阵发性生长特性.利用一年生砧木生长旺盛、生物活性强的特点获得幼化接穗; 以腋芽诱导小叶为外植体, 相当于采用器官发生来克服顽拗现象(Singh et al., 2015 ; Díaz-Sala, 2019 ), 因为分生组织的诱导就是一种幼化过程; 而通过诱导、分化和增殖等器官发生刺激途径, 使增殖一代小叶外植体达到高度幼化, 是其在愈伤诱导、分化及不定芽增殖中表现出较强生长势及分裂活性的原因.本研究中, 不同幼化处理的外植体在愈伤诱导、分化、增殖及生长上均表现出显著差异, 即采用的外植体幼化程度越高, 后期生长潜力越大(于波等, 2020 ), 各项生长生理指标越优, 以增殖一代小叶为外植体效果最佳.研究发现, 随着郁金樱幼化程度的加大, 各项生理指标渐优, 而连续继代后随着幼化的深入, 能否逆转成年态? 这些问题尚无相关报道, 有待深入研究.我们也曾连续对郁金樱进行多次增殖继代, 发现随着继代次数的增加, 出现叶发黄、毒害作用加强、对激素敏感性增强及大量死亡等问题, 尚未尝试以增殖继代多次的小叶为外植体进行愈伤组织再生, 这些都有待后期深入研究. ...Prunus campanulata ) (黄守印等, 2003 ; 吕月良等, 2006 ; Chen et al., 2016 )、日本晚樱杨贵妃(C. serrulata var. lannesiana cv. ‘Mollis’)、日本晚樱醉红(C. serrulata var. lannesiana cv. ‘Zuihong’) (张灵灵和蒋细旺, 2015 )、华中樱(C. conradinae )、迎春樱(C. discoidea ) (宋斯妤, 2018 )、染井吉野樱(C. × yedoensis cv. ‘Somei-yoshino’) (徐晨捷和欧静, 2020 )和喜马拉雅樱花(C. cerasoides ) (李水根等, 2020 )等少数品种(李艳敏等, 2008 , 2012 ).关于樱花植株再生和遗传转化方面的研究较少, 仅见福建山樱花以腋芽诱导小叶为外植体, 成功建立了植株再生体系和体细胞胚胎发生途径(邹娜等, 2013 ); 以雪落樱(C. xueluoensis ) (史港影等, 2014 )和御殿场樱(C. × incisa cv. ‘Gotenba-zakura’) (雷巾茗, 2020 )叶片为外植体建立了愈伤组织再生体系, 目前尚未见有关郁金樱再生体系及组织培养的报道.通过对北玄参(Scrophularia buergeriana ) (任如意等, 2017 )、黄花乌头(Aconitum coreanum ) (刘莉莉等, 2015 )、绒毛白蜡(Fraxinus velutina ) (燕丽萍等, 2016 )和月季(Rosa chinensis ) (陈雪等, 2011 )等植株再生的研究表明, 不同外植体对植株再生影响极显著, 直接关系试验成功与否, 故选择正确外植体是试验成功的前提.本研究以郁金樱不同幼化程度的小叶为材料, 以期通过愈伤组织诱导不定芽分化、不定芽增殖和生根, 建立高效、稳定的郁金樱再生体系, 为其生物技术育种及后续的遗传转化研究提供参考. ...Prunus campanulata ) (黄守印等, 2003 ; 吕月良等, 2006 ; Chen et al., 2016 )、日本晚樱杨贵妃(C. serrulata var. lannesiana cv. ‘Mollis’)、日本晚樱醉红(C. serrulata var. lannesiana cv. ‘Zuihong’) (张灵灵和蒋细旺, 2015 )、华中樱(C. conradinae )、迎春樱(C. discoidea ) (宋斯妤, 2018 )、染井吉野樱(C. × yedoensis cv. ‘Somei-yoshino’) (徐晨捷和欧静, 2020 )和喜马拉雅樱花(C. cerasoides ) (李水根等, 2020 )等少数品种(李艳敏等, 2008 , 2012 ).关于樱花植株再生和遗传转化方面的研究较少, 仅见福建山樱花以腋芽诱导小叶为外植体, 成功建立了植株再生体系和体细胞胚胎发生途径(邹娜等, 2013 ); 以雪落樱(C. xueluoensis ) (史港影等, 2014 )和御殿场樱(C. × incisa cv. ‘Gotenba-zakura’) (雷巾茗, 2020 )叶片为外植体建立了愈伤组织再生体系, 目前尚未见有关郁金樱再生体系及组织培养的报道.通过对北玄参(Scrophularia buergeriana ) (任如意等, 2017 )、黄花乌头(Aconitum coreanum ) (刘莉莉等, 2015 )、绒毛白蜡(Fraxinus velutina ) (燕丽萍等, 2016 )和月季(Rosa chinensis ) (陈雪等, 2011 )等植株再生的研究表明, 不同外植体对植株再生影响极显著, 直接关系试验成功与否, 故选择正确外植体是试验成功的前提.本研究以郁金樱不同幼化程度的小叶为材料, 以期通过愈伤组织诱导不定芽分化、不定芽增殖和生根, 建立高效、稳定的郁金樱再生体系, 为其生物技术育种及后续的遗传转化研究提供参考. ...Prunus campanulata ) (黄守印等, 2003 ; 吕月良等, 2006 ; Chen et al., 2016 )、日本晚樱杨贵妃(C. serrulata var. lannesiana cv. ‘Mollis’)、日本晚樱醉红(C. serrulata var. lannesiana cv. ‘Zuihong’) (张灵灵和蒋细旺, 2015 )、华中樱(C. conradinae )、迎春樱(C. discoidea ) (宋斯妤, 2018 )、染井吉野樱(C. × yedoensis cv. ‘Somei-yoshino’) (徐晨捷和欧静, 2020 )和喜马拉雅樱花(C. cerasoides ) (李水根等, 2020 )等少数品种(李艳敏等, 2008 , 2012 ).关于樱花植株再生和遗传转化方面的研究较少, 仅见福建山樱花以腋芽诱导小叶为外植体, 成功建立了植株再生体系和体细胞胚胎发生途径(邹娜等, 2013 ); 以雪落樱(C. xueluoensis ) (史港影等, 2014 )和御殿场樱(C. × incisa cv. ‘Gotenba-zakura’) (雷巾茗, 2020 )叶片为外植体建立了愈伤组织再生体系, 目前尚未见有关郁金樱再生体系及组织培养的报道.通过对北玄参(Scrophularia buergeriana ) (任如意等, 2017 )、黄花乌头(Aconitum coreanum ) (刘莉莉等, 2015 )、绒毛白蜡(Fraxinus velutina ) (燕丽萍等, 2016 )和月季(Rosa chinensis ) (陈雪等, 2011 )等植株再生的研究表明, 不同外植体对植株再生影响极显著, 直接关系试验成功与否, 故选择正确外植体是试验成功的前提.本研究以郁金樱不同幼化程度的小叶为材料, 以期通过愈伤组织诱导不定芽分化、不定芽增殖和生根, 建立高效、稳定的郁金樱再生体系, 为其生物技术育种及后续的遗传转化研究提供参考. ...Prunus campanulata ) (黄守印等, 2003 ; 吕月良等, 2006 ; Chen et al., 2016 )、日本晚樱杨贵妃(C. serrulata var. lannesiana cv. ‘Mollis’)、日本晚樱醉红(C. serrulata var. lannesiana cv. ‘Zuihong’) (张灵灵和蒋细旺, 2015 )、华中樱(C. conradinae )、迎春樱(C. discoidea ) (宋斯妤, 2018 )、染井吉野樱(C. × yedoensis cv. ‘Somei-yoshino’) (徐晨捷和欧静, 2020 )和喜马拉雅樱花(C. cerasoides ) (李水根等, 2020 )等少数品种(李艳敏等, 2008 , 2012 ).关于樱花植株再生和遗传转化方面的研究较少, 仅见福建山樱花以腋芽诱导小叶为外植体, 成功建立了植株再生体系和体细胞胚胎发生途径(邹娜等, 2013 ); 以雪落樱(C. xueluoensis ) (史港影等, 2014 )和御殿场樱(C. × incisa cv. ‘Gotenba-zakura’) (雷巾茗, 2020 )叶片为外植体建立了愈伤组织再生体系, 目前尚未见有关郁金樱再生体系及组织培养的报道.通过对北玄参(Scrophularia buergeriana ) (任如意等, 2017 )、黄花乌头(Aconitum coreanum ) (刘莉莉等, 2015 )、绒毛白蜡(Fraxinus velutina ) (燕丽萍等, 2016 )和月季(Rosa chinensis ) (陈雪等, 2011 )等植株再生的研究表明, 不同外植体对植株再生影响极显著, 直接关系试验成功与否, 故选择正确外植体是试验成功的前提.本研究以郁金樱不同幼化程度的小叶为材料, 以期通过愈伤组织诱导不定芽分化、不定芽增殖和生根, 建立高效、稳定的郁金樱再生体系, 为其生物技术育种及后续的遗传转化研究提供参考. ...Cerasus serrulata var. lannesiana cv. ‘Grandiflora’)是蔷薇科(Rosaceae)樱属(Cerasus )落叶乔木, 花叶同开, 花色淡黄至绿色, 花期在3月下旬至4月上旬, 在我国长三角地区有栽培, 现已推广到全国.樱花品种花色大多为白色、粉色或红色, 郁金樱因其绿色花而独特, 在盛花期和落花期均有极高的观赏价值, 园林开发潜力巨大(刘晓莉, 2012 ).郁金樱具有良好的适应性和观赏性(朱继军等, 2015 ), 其在校园绿地、居住区绿地、道路绿地、综合公园和城市广场上皆有分布, 苗木市场需求较大(蒋冬月等, 2019 ; 杨小燕等, 2019 ).生产上樱花主要通过嫁接繁殖, 但该繁殖方法存在占用空间资源大、成苗周期长及种苗价格高等问题, 且易受时间、气候和环境等因素影响, 经多代繁殖后, 易造成病毒积累, 影响樱花的产量和品质.随着现代生物技术的发展, 利用基因工程技术进行植物性状改良已成为可行的育种方式(张旭红等, 2018 ), 而建立高效且快速的再生体系是进行遗传转化研究的前提.因此, 应用植物组织培养技术建立无性快繁体系, 对郁金樱的遗传改良具有重要意义. ...Prunus campanulata ) (黄守印等, 2003 ; 吕月良等, 2006 ; Chen et al., 2016 )、日本晚樱杨贵妃(C. serrulata var. lannesiana cv. ‘Mollis’)、日本晚樱醉红(C. serrulata var. lannesiana cv. ‘Zuihong’) (张灵灵和蒋细旺, 2015 )、华中樱(C. conradinae )、迎春樱(C. discoidea ) (宋斯妤, 2018 )、染井吉野樱(C. × yedoensis cv. ‘Somei-yoshino’) (徐晨捷和欧静, 2020 )和喜马拉雅樱花(C. cerasoides ) (李水根等, 2020 )等少数品种(李艳敏等, 2008 , 2012 ).关于樱花植株再生和遗传转化方面的研究较少, 仅见福建山樱花以腋芽诱导小叶为外植体, 成功建立了植株再生体系和体细胞胚胎发生途径(邹娜等, 2013 ); 以雪落樱(C. xueluoensis ) (史港影等, 2014 )和御殿场樱(C. × incisa cv. ‘Gotenba-zakura’) (雷巾茗, 2020 )叶片为外植体建立了愈伤组织再生体系, 目前尚未见有关郁金樱再生体系及组织培养的报道.通过对北玄参(Scrophularia buergeriana ) (任如意等, 2017 )、黄花乌头(Aconitum coreanum ) (刘莉莉等, 2015 )、绒毛白蜡(Fraxinus velutina ) (燕丽萍等, 2016 )和月季(Rosa chinensis ) (陈雪等, 2011 )等植株再生的研究表明, 不同外植体对植株再生影响极显著, 直接关系试验成功与否, 故选择正确外植体是试验成功的前提.本研究以郁金樱不同幼化程度的小叶为材料, 以期通过愈伤组织诱导不定芽分化、不定芽增殖和生根, 建立高效、稳定的郁金樱再生体系, 为其生物技术育种及后续的遗传转化研究提供参考. ...Prunus campanulata ) (黄守印等, 2003 ; 吕月良等, 2006 ; Chen et al., 2016 )、日本晚樱杨贵妃(C. serrulata var. lannesiana cv. ‘Mollis’)、日本晚樱醉红(C. serrulata var. lannesiana cv. ‘Zuihong’) (张灵灵和蒋细旺, 2015 )、华中樱(C. conradinae )、迎春樱(C. discoidea ) (宋斯妤, 2018 )、染井吉野樱(C. × yedoensis cv. ‘Somei-yoshino’) (徐晨捷和欧静, 2020 )和喜马拉雅樱花(C. cerasoides ) (李水根等, 2020 )等少数品种(李艳敏等, 2008 , 2012 ).关于樱花植株再生和遗传转化方面的研究较少, 仅见福建山樱花以腋芽诱导小叶为外植体, 成功建立了植株再生体系和体细胞胚胎发生途径(邹娜等, 2013 ); 以雪落樱(C. xueluoensis ) (史港影等, 2014 )和御殿场樱(C. × incisa cv. ‘Gotenba-zakura’) (雷巾茗, 2020 )叶片为外植体建立了愈伤组织再生体系, 目前尚未见有关郁金樱再生体系及组织培养的报道.通过对北玄参(Scrophularia buergeriana ) (任如意等, 2017 )、黄花乌头(Aconitum coreanum ) (刘莉莉等, 2015 )、绒毛白蜡(Fraxinus velutina ) (燕丽萍等, 2016 )和月季(Rosa chinensis ) (陈雪等, 2011 )等植株再生的研究表明, 不同外植体对植株再生影响极显著, 直接关系试验成功与否, 故选择正确外植体是试验成功的前提.本研究以郁金樱不同幼化程度的小叶为材料, 以期通过愈伤组织诱导不定芽分化、不定芽增殖和生根, 建立高效、稳定的郁金樱再生体系, 为其生物技术育种及后续的遗传转化研究提供参考. ...Prunus campanulata ) (黄守印等, 2003 ; 吕月良等, 2006 ; Chen et al., 2016 )、日本晚樱杨贵妃(C. serrulata var. lannesiana cv. ‘Mollis’)、日本晚樱醉红(C. serrulata var. lannesiana cv. ‘Zuihong’) (张灵灵和蒋细旺, 2015 )、华中樱(C. conradinae )、迎春樱(C. discoidea ) (宋斯妤, 2018 )、染井吉野樱(C. × yedoensis cv. ‘Somei-yoshino’) (徐晨捷和欧静, 2020 )和喜马拉雅樱花(C. cerasoides ) (李水根等, 2020 )等少数品种(李艳敏等, 2008 , 2012 ).关于樱花植株再生和遗传转化方面的研究较少, 仅见福建山樱花以腋芽诱导小叶为外植体, 成功建立了植株再生体系和体细胞胚胎发生途径(邹娜等, 2013 ); 以雪落樱(C. xueluoensis ) (史港影等, 2014 )和御殿场樱(C. × incisa cv. ‘Gotenba-zakura’) (雷巾茗, 2020 )叶片为外植体建立了愈伤组织再生体系, 目前尚未见有关郁金樱再生体系及组织培养的报道.通过对北玄参(Scrophularia buergeriana ) (任如意等, 2017 )、黄花乌头(Aconitum coreanum ) (刘莉莉等, 2015 )、绒毛白蜡(Fraxinus velutina ) (燕丽萍等, 2016 )和月季(Rosa chinensis ) (陈雪等, 2011 )等植株再生的研究表明, 不同外植体对植株再生影响极显著, 直接关系试验成功与否, 故选择正确外植体是试验成功的前提.本研究以郁金樱不同幼化程度的小叶为材料, 以期通过愈伤组织诱导不定芽分化、不定芽增殖和生根, 建立高效、稳定的郁金樱再生体系, 为其生物技术育种及后续的遗传转化研究提供参考. ...闫国华等, 2002 ), 而这些由植物基因决定的生长生理规律通过后期植物激素和营养手段较难调控(郭希梅等, 2011 ), 从而增加了组织培养的难度.以冬樱花(Prunus majestica )叶片、花托、花梗和子房为外植体未分化出不定芽(和凤美等, 2010 ).现阶段成功分化出不定芽的樱花愈伤组织再生研究皆以叶片为外植体.福建山樱花以腋芽诱导小叶和茎段为外植体诱导愈伤组织, 仅腋芽诱导小叶分化出不定芽(邹娜等, 2013 ).雪落樱和御殿场樱以叶片为外植体虽分化出不定芽, 但仍存在褐化率高和分化困难等问题(史港影等, 2014 ; 雷巾茗, 2020 ), 这表明在樱花的愈伤组织再生研究中外植体的选择尤为重要, 可能与樱花基因型和外植体生理状态有关.研究表明, 在树番茄(Cyphomandra betacea ) (Correia et al., 2011 )、小粒咖啡(Coffea arabica ) (Bartos et al., 2018 )和冬青栎(Quercus ilex ) (Martínez et al., 2017 )等物种上, 胚性愈伤的诱导也很大程度依赖于外植体种类, 这种由基因驱动的外植体“抗拒”是很多品种无法建立稳定培养体系的主要原因(McCown, 2000 ).采用郁金樱不同分化程度的小叶诱导再生植株, 我们发现郁金樱在离体培养中表现出强烈的顽拗现象(recalcitrance), 这与樱桃砧木酸樱桃(P. cerasus )相似(Ben Mahmoud et al., 2017 ).本研究以多年生郁金樱小叶为外植体未分化出不定芽, 愈伤组织诱导率低且胚性愈伤较少, 同时随着培养时间的延长褐化现象严重, 顽拗现象出现于多年生小叶组织培养的诱导和分化阶段, 这与前人的研究结果一致(Hu et al., 2017 ; Wu et al., 2020 ).为克服遗传性基因导致的顽拗现象, 本研究采用郁金樱一年生嫁接苗小叶、腋芽诱导小叶及增殖一代小叶为外植体, 相当于间接对多年生郁金樱母株进行不同程度的幼化(rejuvennation)或返童, 幼年外植体相比发育成熟或分化组织具有更大的潜能(Wu et al., 2019 ).通过采集多年生郁金樱母株嫁接于一年生山樱花砧木上, 相当于直接操作原植物来克服郁金樱阵发性生长特性.利用一年生砧木生长旺盛、生物活性强的特点获得幼化接穗; 以腋芽诱导小叶为外植体, 相当于采用器官发生来克服顽拗现象(Singh et al., 2015 ; Díaz-Sala, 2019 ), 因为分生组织的诱导就是一种幼化过程; 而通过诱导、分化和增殖等器官发生刺激途径, 使增殖一代小叶外植体达到高度幼化, 是其在愈伤诱导、分化及不定芽增殖中表现出较强生长势及分裂活性的原因.本研究中, 不同幼化处理的外植体在愈伤诱导、分化、增殖及生长上均表现出显著差异, 即采用的外植体幼化程度越高, 后期生长潜力越大(于波等, 2020 ), 各项生长生理指标越优, 以增殖一代小叶为外植体效果最佳.研究发现, 随着郁金樱幼化程度的加大, 各项生理指标渐优, 而连续继代后随着幼化的深入, 能否逆转成年态? 这些问题尚无相关报道, 有待深入研究.我们也曾连续对郁金樱进行多次增殖继代, 发现随着继代次数的增加, 出现叶发黄、毒害作用加强、对激素敏感性增强及大量死亡等问题, 尚未尝试以增殖继代多次的小叶为外植体进行愈伤组织再生, 这些都有待后期深入研究. ...Prunus campanulata ) (黄守印等, 2003 ; 吕月良等, 2006 ; Chen et al., 2016 )、日本晚樱杨贵妃(C. serrulata var. lannesiana cv. ‘Mollis’)、日本晚樱醉红(C. serrulata var. lannesiana cv. ‘Zuihong’) (张灵灵和蒋细旺, 2015 )、华中樱(C. conradinae )、迎春樱(C. discoidea ) (宋斯妤, 2018 )、染井吉野樱(C. × yedoensis cv. ‘Somei-yoshino’) (徐晨捷和欧静, 2020 )和喜马拉雅樱花(C. cerasoides ) (李水根等, 2020 )等少数品种(李艳敏等, 2008 , 2012 ).关于樱花植株再生和遗传转化方面的研究较少, 仅见福建山樱花以腋芽诱导小叶为外植体, 成功建立了植株再生体系和体细胞胚胎发生途径(邹娜等, 2013 ); 以雪落樱(C. xueluoensis ) (史港影等, 2014 )和御殿场樱(C. × incisa cv. ‘Gotenba-zakura’) (雷巾茗, 2020 )叶片为外植体建立了愈伤组织再生体系, 目前尚未见有关郁金樱再生体系及组织培养的报道.通过对北玄参(Scrophularia buergeriana ) (任如意等, 2017 )、黄花乌头(Aconitum coreanum ) (刘莉莉等, 2015 )、绒毛白蜡(Fraxinus velutina ) (燕丽萍等, 2016 )和月季(Rosa chinensis ) (陈雪等, 2011 )等植株再生的研究表明, 不同外植体对植株再生影响极显著, 直接关系试验成功与否, 故选择正确外植体是试验成功的前提.本研究以郁金樱不同幼化程度的小叶为材料, 以期通过愈伤组织诱导不定芽分化、不定芽增殖和生根, 建立高效、稳定的郁金樱再生体系, 为其生物技术育种及后续的遗传转化研究提供参考. ...Prunus campanulata ) (黄守印等, 2003 ; 吕月良等, 2006 ; Chen et al., 2016 )、日本晚樱杨贵妃(C. serrulata var. lannesiana cv. ‘Mollis’)、日本晚樱醉红(C. serrulata var. lannesiana cv. ‘Zuihong’) (张灵灵和蒋细旺, 2015 )、华中樱(C. conradinae )、迎春樱(C. discoidea ) (宋斯妤, 2018 )、染井吉野樱(C. × yedoensis cv. ‘Somei-yoshino’) (徐晨捷和欧静, 2020 )和喜马拉雅樱花(C. cerasoides ) (李水根等, 2020 )等少数品种(李艳敏等, 2008 , 2012 ).关于樱花植株再生和遗传转化方面的研究较少, 仅见福建山樱花以腋芽诱导小叶为外植体, 成功建立了植株再生体系和体细胞胚胎发生途径(邹娜等, 2013 ); 以雪落樱(C. xueluoensis ) (史港影等, 2014 )和御殿场樱(C. × incisa cv. ‘Gotenba-zakura’) (雷巾茗, 2020 )叶片为外植体建立了愈伤组织再生体系, 目前尚未见有关郁金樱再生体系及组织培养的报道.通过对北玄参(Scrophularia buergeriana ) (任如意等, 2017 )、黄花乌头(Aconitum coreanum ) (刘莉莉等, 2015 )、绒毛白蜡(Fraxinus velutina ) (燕丽萍等, 2016 )和月季(Rosa chinensis ) (陈雪等, 2011 )等植株再生的研究表明, 不同外植体对植株再生影响极显著, 直接关系试验成功与否, 故选择正确外植体是试验成功的前提.本研究以郁金樱不同幼化程度的小叶为材料, 以期通过愈伤组织诱导不定芽分化、不定芽增殖和生根, 建立高效、稳定的郁金樱再生体系, 为其生物技术育种及后续的遗传转化研究提供参考. ...闫国华等, 2002 ), 而这些由植物基因决定的生长生理规律通过后期植物激素和营养手段较难调控(郭希梅等, 2011 ), 从而增加了组织培养的难度.以冬樱花(Prunus majestica )叶片、花托、花梗和子房为外植体未分化出不定芽(和凤美等, 2010 ).现阶段成功分化出不定芽的樱花愈伤组织再生研究皆以叶片为外植体.福建山樱花以腋芽诱导小叶和茎段为外植体诱导愈伤组织, 仅腋芽诱导小叶分化出不定芽(邹娜等, 2013 ).雪落樱和御殿场樱以叶片为外植体虽分化出不定芽, 但仍存在褐化率高和分化困难等问题(史港影等, 2014 ; 雷巾茗, 2020 ), 这表明在樱花的愈伤组织再生研究中外植体的选择尤为重要, 可能与樱花基因型和外植体生理状态有关.研究表明, 在树番茄(Cyphomandra betacea ) (Correia et al., 2011 )、小粒咖啡(Coffea arabica ) (Bartos et al., 2018 )和冬青栎(Quercus ilex ) (Martínez et al., 2017 )等物种上, 胚性愈伤的诱导也很大程度依赖于外植体种类, 这种由基因驱动的外植体“抗拒”是很多品种无法建立稳定培养体系的主要原因(McCown, 2000 ).采用郁金樱不同分化程度的小叶诱导再生植株, 我们发现郁金樱在离体培养中表现出强烈的顽拗现象(recalcitrance), 这与樱桃砧木酸樱桃(P. cerasus )相似(Ben Mahmoud et al., 2017 ).本研究以多年生郁金樱小叶为外植体未分化出不定芽, 愈伤组织诱导率低且胚性愈伤较少, 同时随着培养时间的延长褐化现象严重, 顽拗现象出现于多年生小叶组织培养的诱导和分化阶段, 这与前人的研究结果一致(Hu et al., 2017 ; Wu et al., 2020 ).为克服遗传性基因导致的顽拗现象, 本研究采用郁金樱一年生嫁接苗小叶、腋芽诱导小叶及增殖一代小叶为外植体, 相当于间接对多年生郁金樱母株进行不同程度的幼化(rejuvennation)或返童, 幼年外植体相比发育成熟或分化组织具有更大的潜能(Wu et al., 2019 ).通过采集多年生郁金樱母株嫁接于一年生山樱花砧木上, 相当于直接操作原植物来克服郁金樱阵发性生长特性.利用一年生砧木生长旺盛、生物活性强的特点获得幼化接穗; 以腋芽诱导小叶为外植体, 相当于采用器官发生来克服顽拗现象(Singh et al., 2015 ; Díaz-Sala, 2019 ), 因为分生组织的诱导就是一种幼化过程; 而通过诱导、分化和增殖等器官发生刺激途径, 使增殖一代小叶外植体达到高度幼化, 是其在愈伤诱导、分化及不定芽增殖中表现出较强生长势及分裂活性的原因.本研究中, 不同幼化处理的外植体在愈伤诱导、分化、增殖及生长上均表现出显著差异, 即采用的外植体幼化程度越高, 后期生长潜力越大(于波等, 2020 ), 各项生长生理指标越优, 以增殖一代小叶为外植体效果最佳.研究发现, 随着郁金樱幼化程度的加大, 各项生理指标渐优, 而连续继代后随着幼化的深入, 能否逆转成年态? 这些问题尚无相关报道, 有待深入研究.我们也曾连续对郁金樱进行多次增殖继代, 发现随着继代次数的增加, 出现叶发黄、毒害作用加强、对激素敏感性增强及大量死亡等问题, 尚未尝试以增殖继代多次的小叶为外植体进行愈伤组织再生, 这些都有待后期深入研究. ...Prunus campanulata ) (黄守印等, 2003 ; 吕月良等, 2006 ; Chen et al., 2016 )、日本晚樱杨贵妃(C. serrulata var. lannesiana cv. ‘Mollis’)、日本晚樱醉红(C. serrulata var. lannesiana cv. ‘Zuihong’) (张灵灵和蒋细旺, 2015 )、华中樱(C. conradinae )、迎春樱(C. discoidea ) (宋斯妤, 2018 )、染井吉野樱(C. × yedoensis cv. ‘Somei-yoshino’) (徐晨捷和欧静, 2020 )和喜马拉雅樱花(C. cerasoides ) (李水根等, 2020 )等少数品种(李艳敏等, 2008 , 2012 ).关于樱花植株再生和遗传转化方面的研究较少, 仅见福建山樱花以腋芽诱导小叶为外植体, 成功建立了植株再生体系和体细胞胚胎发生途径(邹娜等, 2013 ); 以雪落樱(C. xueluoensis ) (史港影等, 2014 )和御殿场樱(C. × incisa cv. ‘Gotenba-zakura’) (雷巾茗, 2020 )叶片为外植体建立了愈伤组织再生体系, 目前尚未见有关郁金樱再生体系及组织培养的报道.通过对北玄参(Scrophularia buergeriana ) (任如意等, 2017 )、黄花乌头(Aconitum coreanum ) (刘莉莉等, 2015 )、绒毛白蜡(Fraxinus velutina ) (燕丽萍等, 2016 )和月季(Rosa chinensis ) (陈雪等, 2011 )等植株再生的研究表明, 不同外植体对植株再生影响极显著, 直接关系试验成功与否, 故选择正确外植体是试验成功的前提.本研究以郁金樱不同幼化程度的小叶为材料, 以期通过愈伤组织诱导不定芽分化、不定芽增殖和生根, 建立高效、稳定的郁金樱再生体系, 为其生物技术育种及后续的遗传转化研究提供参考. ...Cerasus serrulata var. lannesiana cv. ‘Grandiflora’)是蔷薇科(Rosaceae)樱属(Cerasus )落叶乔木, 花叶同开, 花色淡黄至绿色, 花期在3月下旬至4月上旬, 在我国长三角地区有栽培, 现已推广到全国.樱花品种花色大多为白色、粉色或红色, 郁金樱因其绿色花而独特, 在盛花期和落花期均有极高的观赏价值, 园林开发潜力巨大(刘晓莉, 2012 ).郁金樱具有良好的适应性和观赏性(朱继军等, 2015 ), 其在校园绿地、居住区绿地、道路绿地、综合公园和城市广场上皆有分布, 苗木市场需求较大(蒋冬月等, 2019 ; 杨小燕等, 2019 ).生产上樱花主要通过嫁接繁殖, 但该繁殖方法存在占用空间资源大、成苗周期长及种苗价格高等问题, 且易受时间、气候和环境等因素影响, 经多代繁殖后, 易造成病毒积累, 影响樱花的产量和品质.随着现代生物技术的发展, 利用基因工程技术进行植物性状改良已成为可行的育种方式(张旭红等, 2018 ), 而建立高效且快速的再生体系是进行遗传转化研究的前提.因此, 应用植物组织培养技术建立无性快繁体系, 对郁金樱的遗传改良具有重要意义. ...闫国华等, 2002 ), 而这些由植物基因决定的生长生理规律通过后期植物激素和营养手段较难调控(郭希梅等, 2011 ), 从而增加了组织培养的难度.以冬樱花(Prunus majestica )叶片、花托、花梗和子房为外植体未分化出不定芽(和凤美等, 2010 ).现阶段成功分化出不定芽的樱花愈伤组织再生研究皆以叶片为外植体.福建山樱花以腋芽诱导小叶和茎段为外植体诱导愈伤组织, 仅腋芽诱导小叶分化出不定芽(邹娜等, 2013 ).雪落樱和御殿场樱以叶片为外植体虽分化出不定芽, 但仍存在褐化率高和分化困难等问题(史港影等, 2014 ; 雷巾茗, 2020 ), 这表明在樱花的愈伤组织再生研究中外植体的选择尤为重要, 可能与樱花基因型和外植体生理状态有关.研究表明, 在树番茄(Cyphomandra betacea ) (Correia et al., 2011 )、小粒咖啡(Coffea arabica ) (Bartos et al., 2018 )和冬青栎(Quercus ilex ) (Martínez et al., 2017 )等物种上, 胚性愈伤的诱导也很大程度依赖于外植体种类, 这种由基因驱动的外植体“抗拒”是很多品种无法建立稳定培养体系的主要原因(McCown, 2000 ).采用郁金樱不同分化程度的小叶诱导再生植株, 我们发现郁金樱在离体培养中表现出强烈的顽拗现象(recalcitrance), 这与樱桃砧木酸樱桃(P. cerasus )相似(Ben Mahmoud et al., 2017 ).本研究以多年生郁金樱小叶为外植体未分化出不定芽, 愈伤组织诱导率低且胚性愈伤较少, 同时随着培养时间的延长褐化现象严重, 顽拗现象出现于多年生小叶组织培养的诱导和分化阶段, 这与前人的研究结果一致(Hu et al., 2017 ; Wu et al., 2020 ).为克服遗传性基因导致的顽拗现象, 本研究采用郁金樱一年生嫁接苗小叶、腋芽诱导小叶及增殖一代小叶为外植体, 相当于间接对多年生郁金樱母株进行不同程度的幼化(rejuvennation)或返童, 幼年外植体相比发育成熟或分化组织具有更大的潜能(Wu et al., 2019 ).通过采集多年生郁金樱母株嫁接于一年生山樱花砧木上, 相当于直接操作原植物来克服郁金樱阵发性生长特性.利用一年生砧木生长旺盛、生物活性强的特点获得幼化接穗; 以腋芽诱导小叶为外植体, 相当于采用器官发生来克服顽拗现象(Singh et al., 2015 ; Díaz-Sala, 2019 ), 因为分生组织的诱导就是一种幼化过程; 而通过诱导、分化和增殖等器官发生刺激途径, 使增殖一代小叶外植体达到高度幼化, 是其在愈伤诱导、分化及不定芽增殖中表现出较强生长势及分裂活性的原因.本研究中, 不同幼化处理的外植体在愈伤诱导、分化、增殖及生长上均表现出显著差异, 即采用的外植体幼化程度越高, 后期生长潜力越大(于波等, 2020 ), 各项生长生理指标越优, 以增殖一代小叶为外植体效果最佳.研究发现, 随着郁金樱幼化程度的加大, 各项生理指标渐优, 而连续继代后随着幼化的深入, 能否逆转成年态? 这些问题尚无相关报道, 有待深入研究.我们也曾连续对郁金樱进行多次增殖继代, 发现随着继代次数的增加, 出现叶发黄、毒害作用加强、对激素敏感性增强及大量死亡等问题, 尚未尝试以增殖继代多次的小叶为外植体进行愈伤组织再生, 这些都有待后期深入研究. ...Prunus campanulata ) (黄守印等, 2003 ; 吕月良等, 2006 ; Chen et al., 2016 )、日本晚樱杨贵妃(C. serrulata var. lannesiana cv. ‘Mollis’)、日本晚樱醉红(C. serrulata var. lannesiana cv. ‘Zuihong’) (张灵灵和蒋细旺, 2015 )、华中樱(C. conradinae )、迎春樱(C. discoidea ) (宋斯妤, 2018 )、染井吉野樱(C. × yedoensis cv. ‘Somei-yoshino’) (徐晨捷和欧静, 2020 )和喜马拉雅樱花(C. cerasoides ) (李水根等, 2020 )等少数品种(李艳敏等, 2008 , 2012 ).关于樱花植株再生和遗传转化方面的研究较少, 仅见福建山樱花以腋芽诱导小叶为外植体, 成功建立了植株再生体系和体细胞胚胎发生途径(邹娜等, 2013 ); 以雪落樱(C. xueluoensis ) (史港影等, 2014 )和御殿场樱(C. × incisa cv. ‘Gotenba-zakura’) (雷巾茗, 2020 )叶片为外植体建立了愈伤组织再生体系, 目前尚未见有关郁金樱再生体系及组织培养的报道.通过对北玄参(Scrophularia buergeriana ) (任如意等, 2017 )、黄花乌头(Aconitum coreanum ) (刘莉莉等, 2015 )、绒毛白蜡(Fraxinus velutina ) (燕丽萍等, 2016 )和月季(Rosa chinensis ) (陈雪等, 2011 )等植株再生的研究表明, 不同外植体对植株再生影响极显著, 直接关系试验成功与否, 故选择正确外植体是试验成功的前提.本研究以郁金樱不同幼化程度的小叶为材料, 以期通过愈伤组织诱导不定芽分化、不定芽增殖和生根, 建立高效、稳定的郁金樱再生体系, 为其生物技术育种及后续的遗传转化研究提供参考. ...Cerasus serrulata var. lannesiana cv. ‘Grandiflora’)是蔷薇科(Rosaceae)樱属(Cerasus )落叶乔木, 花叶同开, 花色淡黄至绿色, 花期在3月下旬至4月上旬, 在我国长三角地区有栽培, 现已推广到全国.樱花品种花色大多为白色、粉色或红色, 郁金樱因其绿色花而独特, 在盛花期和落花期均有极高的观赏价值, 园林开发潜力巨大(刘晓莉, 2012 ).郁金樱具有良好的适应性和观赏性(朱继军等, 2015 ), 其在校园绿地、居住区绿地、道路绿地、综合公园和城市广场上皆有分布, 苗木市场需求较大(蒋冬月等, 2019 ; 杨小燕等, 2019 ).生产上樱花主要通过嫁接繁殖, 但该繁殖方法存在占用空间资源大、成苗周期长及种苗价格高等问题, 且易受时间、气候和环境等因素影响, 经多代繁殖后, 易造成病毒积累, 影响樱花的产量和品质.随着现代生物技术的发展, 利用基因工程技术进行植物性状改良已成为可行的育种方式(张旭红等, 2018 ), 而建立高效且快速的再生体系是进行遗传转化研究的前提.因此, 应用植物组织培养技术建立无性快繁体系, 对郁金樱的遗传改良具有重要意义. ...Cerasus serrulata var. lannesiana cv. ‘Grandiflora’)是蔷薇科(Rosaceae)樱属(Cerasus )落叶乔木, 花叶同开, 花色淡黄至绿色, 花期在3月下旬至4月上旬, 在我国长三角地区有栽培, 现已推广到全国.樱花品种花色大多为白色、粉色或红色, 郁金樱因其绿色花而独特, 在盛花期和落花期均有极高的观赏价值, 园林开发潜力巨大(刘晓莉, 2012 ).郁金樱具有良好的适应性和观赏性(朱继军等, 2015 ), 其在校园绿地、居住区绿地、道路绿地、综合公园和城市广场上皆有分布, 苗木市场需求较大(蒋冬月等, 2019 ; 杨小燕等, 2019 ).生产上樱花主要通过嫁接繁殖, 但该繁殖方法存在占用空间资源大、成苗周期长及种苗价格高等问题, 且易受时间、气候和环境等因素影响, 经多代繁殖后, 易造成病毒积累, 影响樱花的产量和品质.随着现代生物技术的发展, 利用基因工程技术进行植物性状改良已成为可行的育种方式(张旭红等, 2018 ), 而建立高效且快速的再生体系是进行遗传转化研究的前提.因此, 应用植物组织培养技术建立无性快繁体系, 对郁金樱的遗传改良具有重要意义. ...Prunus campanulata ) (黄守印等, 2003 ; 吕月良等, 2006 ; Chen et al., 2016 )、日本晚樱杨贵妃(C. serrulata var. lannesiana cv. ‘Mollis’)、日本晚樱醉红(C. serrulata var. lannesiana cv. ‘Zuihong’) (张灵灵和蒋细旺, 2015 )、华中樱(C. conradinae )、迎春樱(C. discoidea ) (宋斯妤, 2018 )、染井吉野樱(C. × yedoensis cv. ‘Somei-yoshino’) (徐晨捷和欧静, 2020 )和喜马拉雅樱花(C. cerasoides ) (李水根等, 2020 )等少数品种(李艳敏等, 2008 , 2012 ).关于樱花植株再生和遗传转化方面的研究较少, 仅见福建山樱花以腋芽诱导小叶为外植体, 成功建立了植株再生体系和体细胞胚胎发生途径(邹娜等, 2013 ); 以雪落樱(C. xueluoensis ) (史港影等, 2014 )和御殿场樱(C. × incisa cv. ‘Gotenba-zakura’) (雷巾茗, 2020 )叶片为外植体建立了愈伤组织再生体系, 目前尚未见有关郁金樱再生体系及组织培养的报道.通过对北玄参(Scrophularia buergeriana ) (任如意等, 2017 )、黄花乌头(Aconitum coreanum ) (刘莉莉等, 2015 )、绒毛白蜡(Fraxinus velutina ) (燕丽萍等, 2016 )和月季(Rosa chinensis ) (陈雪等, 2011 )等植株再生的研究表明, 不同外植体对植株再生影响极显著, 直接关系试验成功与否, 故选择正确外植体是试验成功的前提.本研究以郁金樱不同幼化程度的小叶为材料, 以期通过愈伤组织诱导不定芽分化、不定芽增殖和生根, 建立高效、稳定的郁金樱再生体系, 为其生物技术育种及后续的遗传转化研究提供参考. ...闫国华等, 2002 ), 而这些由植物基因决定的生长生理规律通过后期植物激素和营养手段较难调控(郭希梅等, 2011 ), 从而增加了组织培养的难度.以冬樱花(Prunus majestica )叶片、花托、花梗和子房为外植体未分化出不定芽(和凤美等, 2010 ).现阶段成功分化出不定芽的樱花愈伤组织再生研究皆以叶片为外植体.福建山樱花以腋芽诱导小叶和茎段为外植体诱导愈伤组织, 仅腋芽诱导小叶分化出不定芽(邹娜等, 2013 ).雪落樱和御殿场樱以叶片为外植体虽分化出不定芽, 但仍存在褐化率高和分化困难等问题(史港影等, 2014 ; 雷巾茗, 2020 ), 这表明在樱花的愈伤组织再生研究中外植体的选择尤为重要, 可能与樱花基因型和外植体生理状态有关.研究表明, 在树番茄(Cyphomandra betacea ) (Correia et al., 2011 )、小粒咖啡(Coffea arabica ) (Bartos et al., 2018 )和冬青栎(Quercus ilex ) (Martínez et al., 2017 )等物种上, 胚性愈伤的诱导也很大程度依赖于外植体种类, 这种由基因驱动的外植体“抗拒”是很多品种无法建立稳定培养体系的主要原因(McCown, 2000 ).采用郁金樱不同分化程度的小叶诱导再生植株, 我们发现郁金樱在离体培养中表现出强烈的顽拗现象(recalcitrance), 这与樱桃砧木酸樱桃(P. cerasus )相似(Ben Mahmoud et al., 2017 ).本研究以多年生郁金樱小叶为外植体未分化出不定芽, 愈伤组织诱导率低且胚性愈伤较少, 同时随着培养时间的延长褐化现象严重, 顽拗现象出现于多年生小叶组织培养的诱导和分化阶段, 这与前人的研究结果一致(Hu et al., 2017 ; Wu et al., 2020 ).为克服遗传性基因导致的顽拗现象, 本研究采用郁金樱一年生嫁接苗小叶、腋芽诱导小叶及增殖一代小叶为外植体, 相当于间接对多年生郁金樱母株进行不同程度的幼化(rejuvennation)或返童, 幼年外植体相比发育成熟或分化组织具有更大的潜能(Wu et al., 2019 ).通过采集多年生郁金樱母株嫁接于一年生山樱花砧木上, 相当于直接操作原植物来克服郁金樱阵发性生长特性.利用一年生砧木生长旺盛、生物活性强的特点获得幼化接穗; 以腋芽诱导小叶为外植体, 相当于采用器官发生来克服顽拗现象(Singh et al., 2015 ; Díaz-Sala, 2019 ), 因为分生组织的诱导就是一种幼化过程; 而通过诱导、分化和增殖等器官发生刺激途径, 使增殖一代小叶外植体达到高度幼化, 是其在愈伤诱导、分化及不定芽增殖中表现出较强生长势及分裂活性的原因.本研究中, 不同幼化处理的外植体在愈伤诱导、分化、增殖及生长上均表现出显著差异, 即采用的外植体幼化程度越高, 后期生长潜力越大(于波等, 2020 ), 各项生长生理指标越优, 以增殖一代小叶为外植体效果最佳.研究发现, 随着郁金樱幼化程度的加大, 各项生理指标渐优, 而连续继代后随着幼化的深入, 能否逆转成年态? 这些问题尚无相关报道, 有待深入研究.我们也曾连续对郁金樱进行多次增殖继代, 发现随着继代次数的增加, 出现叶发黄、毒害作用加强、对激素敏感性增强及大量死亡等问题, 尚未尝试以增殖继代多次的小叶为外植体进行愈伤组织再生, 这些都有待后期深入研究. ...Coffea arabica L闫国华等, 2002 ), 而这些由植物基因决定的生长生理规律通过后期植物激素和营养手段较难调控(郭希梅等, 2011 ), 从而增加了组织培养的难度.以冬樱花(Prunus majestica )叶片、花托、花梗和子房为外植体未分化出不定芽(和凤美等, 2010 ).现阶段成功分化出不定芽的樱花愈伤组织再生研究皆以叶片为外植体.福建山樱花以腋芽诱导小叶和茎段为外植体诱导愈伤组织, 仅腋芽诱导小叶分化出不定芽(邹娜等, 2013 ).雪落樱和御殿场樱以叶片为外植体虽分化出不定芽, 但仍存在褐化率高和分化困难等问题(史港影等, 2014 ; 雷巾茗, 2020 ), 这表明在樱花的愈伤组织再生研究中外植体的选择尤为重要, 可能与樱花基因型和外植体生理状态有关.研究表明, 在树番茄(Cyphomandra betacea ) (Correia et al., 2011 )、小粒咖啡(Coffea arabica ) (Bartos et al., 2018 )和冬青栎(Quercus ilex ) (Martínez et al., 2017 )等物种上, 胚性愈伤的诱导也很大程度依赖于外植体种类, 这种由基因驱动的外植体“抗拒”是很多品种无法建立稳定培养体系的主要原因(McCown, 2000 ).采用郁金樱不同分化程度的小叶诱导再生植株, 我们发现郁金樱在离体培养中表现出强烈的顽拗现象(recalcitrance), 这与樱桃砧木酸樱桃(P. cerasus )相似(Ben Mahmoud et al., 2017 ).本研究以多年生郁金樱小叶为外植体未分化出不定芽, 愈伤组织诱导率低且胚性愈伤较少, 同时随着培养时间的延长褐化现象严重, 顽拗现象出现于多年生小叶组织培养的诱导和分化阶段, 这与前人的研究结果一致(Hu et al., 2017 ; Wu et al., 2020 ).为克服遗传性基因导致的顽拗现象, 本研究采用郁金樱一年生嫁接苗小叶、腋芽诱导小叶及增殖一代小叶为外植体, 相当于间接对多年生郁金樱母株进行不同程度的幼化(rejuvennation)或返童, 幼年外植体相比发育成熟或分化组织具有更大的潜能(Wu et al., 2019 ).通过采集多年生郁金樱母株嫁接于一年生山樱花砧木上, 相当于直接操作原植物来克服郁金樱阵发性生长特性.利用一年生砧木生长旺盛、生物活性强的特点获得幼化接穗; 以腋芽诱导小叶为外植体, 相当于采用器官发生来克服顽拗现象(Singh et al., 2015 ; Díaz-Sala, 2019 ), 因为分生组织的诱导就是一种幼化过程; 而通过诱导、分化和增殖等器官发生刺激途径, 使增殖一代小叶外植体达到高度幼化, 是其在愈伤诱导、分化及不定芽增殖中表现出较强生长势及分裂活性的原因.本研究中, 不同幼化处理的外植体在愈伤诱导、分化、增殖及生长上均表现出显著差异, 即采用的外植体幼化程度越高, 后期生长潜力越大(于波等, 2020 ), 各项生长生理指标越优, 以增殖一代小叶为外植体效果最佳.研究发现, 随着郁金樱幼化程度的加大, 各项生理指标渐优, 而连续继代后随着幼化的深入, 能否逆转成年态? 这些问题尚无相关报道, 有待深入研究.我们也曾连续对郁金樱进行多次增殖继代, 发现随着继代次数的增加, 出现叶发黄、毒害作用加强、对激素敏感性增强及大量死亡等问题, 尚未尝试以增殖继代多次的小叶为外植体进行愈伤组织再生, 这些都有待后期深入研究. ...Prunus cerasus L.) rootstock CAB 6P闫国华等, 2002 ), 而这些由植物基因决定的生长生理规律通过后期植物激素和营养手段较难调控(郭希梅等, 2011 ), 从而增加了组织培养的难度.以冬樱花(Prunus majestica )叶片、花托、花梗和子房为外植体未分化出不定芽(和凤美等, 2010 ).现阶段成功分化出不定芽的樱花愈伤组织再生研究皆以叶片为外植体.福建山樱花以腋芽诱导小叶和茎段为外植体诱导愈伤组织, 仅腋芽诱导小叶分化出不定芽(邹娜等, 2013 ).雪落樱和御殿场樱以叶片为外植体虽分化出不定芽, 但仍存在褐化率高和分化困难等问题(史港影等, 2014 ; 雷巾茗, 2020 ), 这表明在樱花的愈伤组织再生研究中外植体的选择尤为重要, 可能与樱花基因型和外植体生理状态有关.研究表明, 在树番茄(Cyphomandra betacea ) (Correia et al., 2011 )、小粒咖啡(Coffea arabica ) (Bartos et al., 2018 )和冬青栎(Quercus ilex ) (Martínez et al., 2017 )等物种上, 胚性愈伤的诱导也很大程度依赖于外植体种类, 这种由基因驱动的外植体“抗拒”是很多品种无法建立稳定培养体系的主要原因(McCown, 2000 ).采用郁金樱不同分化程度的小叶诱导再生植株, 我们发现郁金樱在离体培养中表现出强烈的顽拗现象(recalcitrance), 这与樱桃砧木酸樱桃(P. cerasus )相似(Ben Mahmoud et al., 2017 ).本研究以多年生郁金樱小叶为外植体未分化出不定芽, 愈伤组织诱导率低且胚性愈伤较少, 同时随着培养时间的延长褐化现象严重, 顽拗现象出现于多年生小叶组织培养的诱导和分化阶段, 这与前人的研究结果一致(Hu et al., 2017 ; Wu et al., 2020 ).为克服遗传性基因导致的顽拗现象, 本研究采用郁金樱一年生嫁接苗小叶、腋芽诱导小叶及增殖一代小叶为外植体, 相当于间接对多年生郁金樱母株进行不同程度的幼化(rejuvennation)或返童, 幼年外植体相比发育成熟或分化组织具有更大的潜能(Wu et al., 2019 ).通过采集多年生郁金樱母株嫁接于一年生山樱花砧木上, 相当于直接操作原植物来克服郁金樱阵发性生长特性.利用一年生砧木生长旺盛、生物活性强的特点获得幼化接穗; 以腋芽诱导小叶为外植体, 相当于采用器官发生来克服顽拗现象(Singh et al., 2015 ; Díaz-Sala, 2019 ), 因为分生组织的诱导就是一种幼化过程; 而通过诱导、分化和增殖等器官发生刺激途径, 使增殖一代小叶外植体达到高度幼化, 是其在愈伤诱导、分化及不定芽增殖中表现出较强生长势及分裂活性的原因.本研究中, 不同幼化处理的外植体在愈伤诱导、分化、增殖及生长上均表现出显著差异, 即采用的外植体幼化程度越高, 后期生长潜力越大(于波等, 2020 ), 各项生长生理指标越优, 以增殖一代小叶为外植体效果最佳.研究发现, 随着郁金樱幼化程度的加大, 各项生理指标渐优, 而连续继代后随着幼化的深入, 能否逆转成年态? 这些问题尚无相关报道, 有待深入研究.我们也曾连续对郁金樱进行多次增殖继代, 发现随着继代次数的增加, 出现叶发黄、毒害作用加强、对激素敏感性增强及大量死亡等问题, 尚未尝试以增殖继代多次的小叶为外植体进行愈伤组织再生, 这些都有待后期深入研究. ...Arabidopsis roots表3 ), 其具有较强的生长素和细胞分裂素双重活性, 对克服顽拗现象有显著效果(Bernula et al., 2020 ).随着外植体幼化程度的提高, 不同外植体诱导的愈伤组织对激素敏感性增强(Ming et al., 2019 ), 表现为具有高活性的TDZ对愈伤分化影响不显著, 低浓度6-BA及2,4-D对分化的影响逐渐加强, 到不定芽增殖时, 腋芽诱导小叶和增殖一代小叶不定芽能在不含NAA的培养基中大量增殖.生根培养更出现组培苗在不含任何激素的1/2MS培养基上生根率达到100%的现象, 在含有NAA或IBA的培养基中, 根系诱导受到不同程度的抑制.综上表明, 在植物组织培养过程中, 植物激素可能随着培养的深入出现富集, 导致后期植物对激素敏感性增强, 对激素依赖度降低.还有可能是郁金樱组培再生过程中出现了激素自养性的适应性现象, 即获得了不依赖外援生长素和细胞分裂素的能力.通过对郁金樱植株再生各阶段激素浓度的横向比较, 发现郁金樱再生过程中不同外植体对激素的敏感性均发生改变, 幼化程度最高的增殖一代小叶对激素敏感性最强. ...Cerasus campanulata by tissue culturePrunus campanulata ) (黄守印等, 2003 ; 吕月良等, 2006 ; Chen et al., 2016 )、日本晚樱杨贵妃(C. serrulata var. lannesiana cv. ‘Mollis’)、日本晚樱醉红(C. serrulata var. lannesiana cv. ‘Zuihong’) (张灵灵和蒋细旺, 2015 )、华中樱(C. conradinae )、迎春樱(C. discoidea ) (宋斯妤, 2018 )、染井吉野樱(C. × yedoensis cv. ‘Somei-yoshino’) (徐晨捷和欧静, 2020 )和喜马拉雅樱花(C. cerasoides ) (李水根等, 2020 )等少数品种(李艳敏等, 2008 , 2012 ).关于樱花植株再生和遗传转化方面的研究较少, 仅见福建山樱花以腋芽诱导小叶为外植体, 成功建立了植株再生体系和体细胞胚胎发生途径(邹娜等, 2013 ); 以雪落樱(C. xueluoensis ) (史港影等, 2014 )和御殿场樱(C. × incisa cv. ‘Gotenba-zakura’) (雷巾茗, 2020 )叶片为外植体建立了愈伤组织再生体系, 目前尚未见有关郁金樱再生体系及组织培养的报道.通过对北玄参(Scrophularia buergeriana ) (任如意等, 2017 )、黄花乌头(Aconitum coreanum ) (刘莉莉等, 2015 )、绒毛白蜡(Fraxinus velutina ) (燕丽萍等, 2016 )和月季(Rosa chinensis ) (陈雪等, 2011 )等植株再生的研究表明, 不同外植体对植株再生影响极显著, 直接关系试验成功与否, 故选择正确外植体是试验成功的前提.本研究以郁金樱不同幼化程度的小叶为材料, 以期通过愈伤组织诱导不定芽分化、不定芽增殖和生根, 建立高效、稳定的郁金樱再生体系, 为其生物技术育种及后续的遗传转化研究提供参考. ...Cyphomandra betacea (Cav.) Sendt. (tamarillo)闫国华等, 2002 ), 而这些由植物基因决定的生长生理规律通过后期植物激素和营养手段较难调控(郭希梅等, 2011 ), 从而增加了组织培养的难度.以冬樱花(Prunus majestica )叶片、花托、花梗和子房为外植体未分化出不定芽(和凤美等, 2010 ).现阶段成功分化出不定芽的樱花愈伤组织再生研究皆以叶片为外植体.福建山樱花以腋芽诱导小叶和茎段为外植体诱导愈伤组织, 仅腋芽诱导小叶分化出不定芽(邹娜等, 2013 ).雪落樱和御殿场樱以叶片为外植体虽分化出不定芽, 但仍存在褐化率高和分化困难等问题(史港影等, 2014 ; 雷巾茗, 2020 ), 这表明在樱花的愈伤组织再生研究中外植体的选择尤为重要, 可能与樱花基因型和外植体生理状态有关.研究表明, 在树番茄(Cyphomandra betacea ) (Correia et al., 2011 )、小粒咖啡(Coffea arabica ) (Bartos et al., 2018 )和冬青栎(Quercus ilex ) (Martínez et al., 2017 )等物种上, 胚性愈伤的诱导也很大程度依赖于外植体种类, 这种由基因驱动的外植体“抗拒”是很多品种无法建立稳定培养体系的主要原因(McCown, 2000 ).采用郁金樱不同分化程度的小叶诱导再生植株, 我们发现郁金樱在离体培养中表现出强烈的顽拗现象(recalcitrance), 这与樱桃砧木酸樱桃(P. cerasus )相似(Ben Mahmoud et al., 2017 ).本研究以多年生郁金樱小叶为外植体未分化出不定芽, 愈伤组织诱导率低且胚性愈伤较少, 同时随着培养时间的延长褐化现象严重, 顽拗现象出现于多年生小叶组织培养的诱导和分化阶段, 这与前人的研究结果一致(Hu et al., 2017 ; Wu et al., 2020 ).为克服遗传性基因导致的顽拗现象, 本研究采用郁金樱一年生嫁接苗小叶、腋芽诱导小叶及增殖一代小叶为外植体, 相当于间接对多年生郁金樱母株进行不同程度的幼化(rejuvennation)或返童, 幼年外植体相比发育成熟或分化组织具有更大的潜能(Wu et al., 2019 ).通过采集多年生郁金樱母株嫁接于一年生山樱花砧木上, 相当于直接操作原植物来克服郁金樱阵发性生长特性.利用一年生砧木生长旺盛、生物活性强的特点获得幼化接穗; 以腋芽诱导小叶为外植体, 相当于采用器官发生来克服顽拗现象(Singh et al., 2015 ; Díaz-Sala, 2019 ), 因为分生组织的诱导就是一种幼化过程; 而通过诱导、分化和增殖等器官发生刺激途径, 使增殖一代小叶外植体达到高度幼化, 是其在愈伤诱导、分化及不定芽增殖中表现出较强生长势及分裂活性的原因.本研究中, 不同幼化处理的外植体在愈伤诱导、分化、增殖及生长上均表现出显著差异, 即采用的外植体幼化程度越高, 后期生长潜力越大(于波等, 2020 ), 各项生长生理指标越优, 以增殖一代小叶为外植体效果最佳.研究发现, 随着郁金樱幼化程度的加大, 各项生理指标渐优, 而连续继代后随着幼化的深入, 能否逆转成年态? 这些问题尚无相关报道, 有待深入研究.我们也曾连续对郁金樱进行多次增殖继代, 发现随着继代次数的增加, 出现叶发黄、毒害作用加强、对激素敏感性增强及大量死亡等问题, 尚未尝试以增殖继代多次的小叶为外植体进行愈伤组织再生, 这些都有待后期深入研究. ...闫国华等, 2002 ), 而这些由植物基因决定的生长生理规律通过后期植物激素和营养手段较难调控(郭希梅等, 2011 ), 从而增加了组织培养的难度.以冬樱花(Prunus majestica )叶片、花托、花梗和子房为外植体未分化出不定芽(和凤美等, 2010 ).现阶段成功分化出不定芽的樱花愈伤组织再生研究皆以叶片为外植体.福建山樱花以腋芽诱导小叶和茎段为外植体诱导愈伤组织, 仅腋芽诱导小叶分化出不定芽(邹娜等, 2013 ).雪落樱和御殿场樱以叶片为外植体虽分化出不定芽, 但仍存在褐化率高和分化困难等问题(史港影等, 2014 ; 雷巾茗, 2020 ), 这表明在樱花的愈伤组织再生研究中外植体的选择尤为重要, 可能与樱花基因型和外植体生理状态有关.研究表明, 在树番茄(Cyphomandra betacea ) (Correia et al., 2011 )、小粒咖啡(Coffea arabica ) (Bartos et al., 2018 )和冬青栎(Quercus ilex ) (Martínez et al., 2017 )等物种上, 胚性愈伤的诱导也很大程度依赖于外植体种类, 这种由基因驱动的外植体“抗拒”是很多品种无法建立稳定培养体系的主要原因(McCown, 2000 ).采用郁金樱不同分化程度的小叶诱导再生植株, 我们发现郁金樱在离体培养中表现出强烈的顽拗现象(recalcitrance), 这与樱桃砧木酸樱桃(P. cerasus )相似(Ben Mahmoud et al., 2017 ).本研究以多年生郁金樱小叶为外植体未分化出不定芽, 愈伤组织诱导率低且胚性愈伤较少, 同时随着培养时间的延长褐化现象严重, 顽拗现象出现于多年生小叶组织培养的诱导和分化阶段, 这与前人的研究结果一致(Hu et al., 2017 ; Wu et al., 2020 ).为克服遗传性基因导致的顽拗现象, 本研究采用郁金樱一年生嫁接苗小叶、腋芽诱导小叶及增殖一代小叶为外植体, 相当于间接对多年生郁金樱母株进行不同程度的幼化(rejuvennation)或返童, 幼年外植体相比发育成熟或分化组织具有更大的潜能(Wu et al., 2019 ).通过采集多年生郁金樱母株嫁接于一年生山樱花砧木上, 相当于直接操作原植物来克服郁金樱阵发性生长特性.利用一年生砧木生长旺盛、生物活性强的特点获得幼化接穗; 以腋芽诱导小叶为外植体, 相当于采用器官发生来克服顽拗现象(Singh et al., 2015 ; Díaz-Sala, 2019 ), 因为分生组织的诱导就是一种幼化过程; 而通过诱导、分化和增殖等器官发生刺激途径, 使增殖一代小叶外植体达到高度幼化, 是其在愈伤诱导、分化及不定芽增殖中表现出较强生长势及分裂活性的原因.本研究中, 不同幼化处理的外植体在愈伤诱导、分化、增殖及生长上均表现出显著差异, 即采用的外植体幼化程度越高, 后期生长潜力越大(于波等, 2020 ), 各项生长生理指标越优, 以增殖一代小叶为外植体效果最佳.研究发现, 随着郁金樱幼化程度的加大, 各项生理指标渐优, 而连续继代后随着幼化的深入, 能否逆转成年态? 这些问题尚无相关报道, 有待深入研究.我们也曾连续对郁金樱进行多次增殖继代, 发现随着继代次数的增加, 出现叶发黄、毒害作用加强、对激素敏感性增强及大量死亡等问题, 尚未尝试以增殖继代多次的小叶为外植体进行愈伤组织再生, 这些都有待后期深入研究. ... Cunninghamia lanceolata (Lamb.) hook zygotic embryos闫国华等, 2002 ), 而这些由植物基因决定的生长生理规律通过后期植物激素和营养手段较难调控(郭希梅等, 2011 ), 从而增加了组织培养的难度.以冬樱花(Prunus majestica )叶片、花托、花梗和子房为外植体未分化出不定芽(和凤美等, 2010 ).现阶段成功分化出不定芽的樱花愈伤组织再生研究皆以叶片为外植体.福建山樱花以腋芽诱导小叶和茎段为外植体诱导愈伤组织, 仅腋芽诱导小叶分化出不定芽(邹娜等, 2013 ).雪落樱和御殿场樱以叶片为外植体虽分化出不定芽, 但仍存在褐化率高和分化困难等问题(史港影等, 2014 ; 雷巾茗, 2020 ), 这表明在樱花的愈伤组织再生研究中外植体的选择尤为重要, 可能与樱花基因型和外植体生理状态有关.研究表明, 在树番茄(Cyphomandra betacea ) (Correia et al., 2011 )、小粒咖啡(Coffea arabica ) (Bartos et al., 2018 )和冬青栎(Quercus ilex ) (Martínez et al., 2017 )等物种上, 胚性愈伤的诱导也很大程度依赖于外植体种类, 这种由基因驱动的外植体“抗拒”是很多品种无法建立稳定培养体系的主要原因(McCown, 2000 ).采用郁金樱不同分化程度的小叶诱导再生植株, 我们发现郁金樱在离体培养中表现出强烈的顽拗现象(recalcitrance), 这与樱桃砧木酸樱桃(P. cerasus )相似(Ben Mahmoud et al., 2017 ).本研究以多年生郁金樱小叶为外植体未分化出不定芽, 愈伤组织诱导率低且胚性愈伤较少, 同时随着培养时间的延长褐化现象严重, 顽拗现象出现于多年生小叶组织培养的诱导和分化阶段, 这与前人的研究结果一致(Hu et al., 2017 ; Wu et al., 2020 ).为克服遗传性基因导致的顽拗现象, 本研究采用郁金樱一年生嫁接苗小叶、腋芽诱导小叶及增殖一代小叶为外植体, 相当于间接对多年生郁金樱母株进行不同程度的幼化(rejuvennation)或返童, 幼年外植体相比发育成熟或分化组织具有更大的潜能(Wu et al., 2019 ).通过采集多年生郁金樱母株嫁接于一年生山樱花砧木上, 相当于直接操作原植物来克服郁金樱阵发性生长特性.利用一年生砧木生长旺盛、生物活性强的特点获得幼化接穗; 以腋芽诱导小叶为外植体, 相当于采用器官发生来克服顽拗现象(Singh et al., 2015 ; Díaz-Sala, 2019 ), 因为分生组织的诱导就是一种幼化过程; 而通过诱导、分化和增殖等器官发生刺激途径, 使增殖一代小叶外植体达到高度幼化, 是其在愈伤诱导、分化及不定芽增殖中表现出较强生长势及分裂活性的原因.本研究中, 不同幼化处理的外植体在愈伤诱导、分化、增殖及生长上均表现出显著差异, 即采用的外植体幼化程度越高, 后期生长潜力越大(于波等, 2020 ), 各项生长生理指标越优, 以增殖一代小叶为外植体效果最佳.研究发现, 随着郁金樱幼化程度的加大, 各项生理指标渐优, 而连续继代后随着幼化的深入, 能否逆转成年态? 这些问题尚无相关报道, 有待深入研究.我们也曾连续对郁金樱进行多次增殖继代, 发现随着继代次数的增加, 出现叶发黄、毒害作用加强、对激素敏感性增强及大量死亡等问题, 尚未尝试以增殖继代多次的小叶为外植体进行愈伤组织再生, 这些都有待后期深入研究. ...Quercus ilex L. (holm oak) trees by somatic embryogenesis闫国华等, 2002 ), 而这些由植物基因决定的生长生理规律通过后期植物激素和营养手段较难调控(郭希梅等, 2011 ), 从而增加了组织培养的难度.以冬樱花(Prunus majestica )叶片、花托、花梗和子房为外植体未分化出不定芽(和凤美等, 2010 ).现阶段成功分化出不定芽的樱花愈伤组织再生研究皆以叶片为外植体.福建山樱花以腋芽诱导小叶和茎段为外植体诱导愈伤组织, 仅腋芽诱导小叶分化出不定芽(邹娜等, 2013 ).雪落樱和御殿场樱以叶片为外植体虽分化出不定芽, 但仍存在褐化率高和分化困难等问题(史港影等, 2014 ; 雷巾茗, 2020 ), 这表明在樱花的愈伤组织再生研究中外植体的选择尤为重要, 可能与樱花基因型和外植体生理状态有关.研究表明, 在树番茄(Cyphomandra betacea ) (Correia et al., 2011 )、小粒咖啡(Coffea arabica ) (Bartos et al., 2018 )和冬青栎(Quercus ilex ) (Martínez et al., 2017 )等物种上, 胚性愈伤的诱导也很大程度依赖于外植体种类, 这种由基因驱动的外植体“抗拒”是很多品种无法建立稳定培养体系的主要原因(McCown, 2000 ).采用郁金樱不同分化程度的小叶诱导再生植株, 我们发现郁金樱在离体培养中表现出强烈的顽拗现象(recalcitrance), 这与樱桃砧木酸樱桃(P. cerasus )相似(Ben Mahmoud et al., 2017 ).本研究以多年生郁金樱小叶为外植体未分化出不定芽, 愈伤组织诱导率低且胚性愈伤较少, 同时随着培养时间的延长褐化现象严重, 顽拗现象出现于多年生小叶组织培养的诱导和分化阶段, 这与前人的研究结果一致(Hu et al., 2017 ; Wu et al., 2020 ).为克服遗传性基因导致的顽拗现象, 本研究采用郁金樱一年生嫁接苗小叶、腋芽诱导小叶及增殖一代小叶为外植体, 相当于间接对多年生郁金樱母株进行不同程度的幼化(rejuvennation)或返童, 幼年外植体相比发育成熟或分化组织具有更大的潜能(Wu et al., 2019 ).通过采集多年生郁金樱母株嫁接于一年生山樱花砧木上, 相当于直接操作原植物来克服郁金樱阵发性生长特性.利用一年生砧木生长旺盛、生物活性强的特点获得幼化接穗; 以腋芽诱导小叶为外植体, 相当于采用器官发生来克服顽拗现象(Singh et al., 2015 ; Díaz-Sala, 2019 ), 因为分生组织的诱导就是一种幼化过程; 而通过诱导、分化和增殖等器官发生刺激途径, 使增殖一代小叶外植体达到高度幼化, 是其在愈伤诱导、分化及不定芽增殖中表现出较强生长势及分裂活性的原因.本研究中, 不同幼化处理的外植体在愈伤诱导、分化、增殖及生长上均表现出显著差异, 即采用的外植体幼化程度越高, 后期生长潜力越大(于波等, 2020 ), 各项生长生理指标越优, 以增殖一代小叶为外植体效果最佳.研究发现, 随着郁金樱幼化程度的加大, 各项生理指标渐优, 而连续继代后随着幼化的深入, 能否逆转成年态? 这些问题尚无相关报道, 有待深入研究.我们也曾连续对郁金樱进行多次增殖继代, 发现随着继代次数的增加, 出现叶发黄、毒害作用加强、对激素敏感性增强及大量死亡等问题, 尚未尝试以增殖继代多次的小叶为外植体进行愈伤组织再生, 这些都有待后期深入研究. ...in vitro plant recalcitrance recalcitrance of woody and herbaceous perennial plants: dealing with genetic predeterminism闫国华等, 2002 ), 而这些由植物基因决定的生长生理规律通过后期植物激素和营养手段较难调控(郭希梅等, 2011 ), 从而增加了组织培养的难度.以冬樱花(Prunus majestica )叶片、花托、花梗和子房为外植体未分化出不定芽(和凤美等, 2010 ).现阶段成功分化出不定芽的樱花愈伤组织再生研究皆以叶片为外植体.福建山樱花以腋芽诱导小叶和茎段为外植体诱导愈伤组织, 仅腋芽诱导小叶分化出不定芽(邹娜等, 2013 ).雪落樱和御殿场樱以叶片为外植体虽分化出不定芽, 但仍存在褐化率高和分化困难等问题(史港影等, 2014 ; 雷巾茗, 2020 ), 这表明在樱花的愈伤组织再生研究中外植体的选择尤为重要, 可能与樱花基因型和外植体生理状态有关.研究表明, 在树番茄(Cyphomandra betacea ) (Correia et al., 2011 )、小粒咖啡(Coffea arabica ) (Bartos et al., 2018 )和冬青栎(Quercus ilex ) (Martínez et al., 2017 )等物种上, 胚性愈伤的诱导也很大程度依赖于外植体种类, 这种由基因驱动的外植体“抗拒”是很多品种无法建立稳定培养体系的主要原因(McCown, 2000 ).采用郁金樱不同分化程度的小叶诱导再生植株, 我们发现郁金樱在离体培养中表现出强烈的顽拗现象(recalcitrance), 这与樱桃砧木酸樱桃(P. cerasus )相似(Ben Mahmoud et al., 2017 ).本研究以多年生郁金樱小叶为外植体未分化出不定芽, 愈伤组织诱导率低且胚性愈伤较少, 同时随着培养时间的延长褐化现象严重, 顽拗现象出现于多年生小叶组织培养的诱导和分化阶段, 这与前人的研究结果一致(Hu et al., 2017 ; Wu et al., 2020 ).为克服遗传性基因导致的顽拗现象, 本研究采用郁金樱一年生嫁接苗小叶、腋芽诱导小叶及增殖一代小叶为外植体, 相当于间接对多年生郁金樱母株进行不同程度的幼化(rejuvennation)或返童, 幼年外植体相比发育成熟或分化组织具有更大的潜能(Wu et al., 2019 ).通过采集多年生郁金樱母株嫁接于一年生山樱花砧木上, 相当于直接操作原植物来克服郁金樱阵发性生长特性.利用一年生砧木生长旺盛、生物活性强的特点获得幼化接穗; 以腋芽诱导小叶为外植体, 相当于采用器官发生来克服顽拗现象(Singh et al., 2015 ; Díaz-Sala, 2019 ), 因为分生组织的诱导就是一种幼化过程; 而通过诱导、分化和增殖等器官发生刺激途径, 使增殖一代小叶外植体达到高度幼化, 是其在愈伤诱导、分化及不定芽增殖中表现出较强生长势及分裂活性的原因.本研究中, 不同幼化处理的外植体在愈伤诱导、分化、增殖及生长上均表现出显著差异, 即采用的外植体幼化程度越高, 后期生长潜力越大(于波等, 2020 ), 各项生长生理指标越优, 以增殖一代小叶为外植体效果最佳.研究发现, 随着郁金樱幼化程度的加大, 各项生理指标渐优, 而连续继代后随着幼化的深入, 能否逆转成年态? 这些问题尚无相关报道, 有待深入研究.我们也曾连续对郁金樱进行多次增殖继代, 发现随着继代次数的增加, 出现叶发黄、毒害作用加强、对激素敏感性增强及大量死亡等问题, 尚未尝试以增殖继代多次的小叶为外植体进行愈伤组织再生, 这些都有待后期深入研究. ...表3 ), 其具有较强的生长素和细胞分裂素双重活性, 对克服顽拗现象有显著效果(Bernula et al., 2020 ).随着外植体幼化程度的提高, 不同外植体诱导的愈伤组织对激素敏感性增强(Ming et al., 2019 ), 表现为具有高活性的TDZ对愈伤分化影响不显著, 低浓度6-BA及2,4-D对分化的影响逐渐加强, 到不定芽增殖时, 腋芽诱导小叶和增殖一代小叶不定芽能在不含NAA的培养基中大量增殖.生根培养更出现组培苗在不含任何激素的1/2MS培养基上生根率达到100%的现象, 在含有NAA或IBA的培养基中, 根系诱导受到不同程度的抑制.综上表明, 在植物组织培养过程中, 植物激素可能随着培养的深入出现富集, 导致后期植物对激素敏感性增强, 对激素依赖度降低.还有可能是郁金樱组培再生过程中出现了激素自养性的适应性现象, 即获得了不依赖外援生长素和细胞分裂素的能力.通过对郁金樱植株再生各阶段激素浓度的横向比较, 发现郁金樱再生过程中不同外植体对激素的敏感性均发生改变, 幼化程度最高的增殖一代小叶对激素敏感性最强. ...Sapindus mukorossi gaertn. from leaf-derived callus induced with 6-benzylaminopurine闫国华等, 2002 ), 而这些由植物基因决定的生长生理规律通过后期植物激素和营养手段较难调控(郭希梅等, 2011 ), 从而增加了组织培养的难度.以冬樱花(Prunus majestica )叶片、花托、花梗和子房为外植体未分化出不定芽(和凤美等, 2010 ).现阶段成功分化出不定芽的樱花愈伤组织再生研究皆以叶片为外植体.福建山樱花以腋芽诱导小叶和茎段为外植体诱导愈伤组织, 仅腋芽诱导小叶分化出不定芽(邹娜等, 2013 ).雪落樱和御殿场樱以叶片为外植体虽分化出不定芽, 但仍存在褐化率高和分化困难等问题(史港影等, 2014 ; 雷巾茗, 2020 ), 这表明在樱花的愈伤组织再生研究中外植体的选择尤为重要, 可能与樱花基因型和外植体生理状态有关.研究表明, 在树番茄(Cyphomandra betacea ) (Correia et al., 2011 )、小粒咖啡(Coffea arabica ) (Bartos et al., 2018 )和冬青栎(Quercus ilex ) (Martínez et al., 2017 )等物种上, 胚性愈伤的诱导也很大程度依赖于外植体种类, 这种由基因驱动的外植体“抗拒”是很多品种无法建立稳定培养体系的主要原因(McCown, 2000 ).采用郁金樱不同分化程度的小叶诱导再生植株, 我们发现郁金樱在离体培养中表现出强烈的顽拗现象(recalcitrance), 这与樱桃砧木酸樱桃(P. cerasus )相似(Ben Mahmoud et al., 2017 ).本研究以多年生郁金樱小叶为外植体未分化出不定芽, 愈伤组织诱导率低且胚性愈伤较少, 同时随着培养时间的延长褐化现象严重, 顽拗现象出现于多年生小叶组织培养的诱导和分化阶段, 这与前人的研究结果一致(Hu et al., 2017 ; Wu et al., 2020 ).为克服遗传性基因导致的顽拗现象, 本研究采用郁金樱一年生嫁接苗小叶、腋芽诱导小叶及增殖一代小叶为外植体, 相当于间接对多年生郁金樱母株进行不同程度的幼化(rejuvennation)或返童, 幼年外植体相比发育成熟或分化组织具有更大的潜能(Wu et al., 2019 ).通过采集多年生郁金樱母株嫁接于一年生山樱花砧木上, 相当于直接操作原植物来克服郁金樱阵发性生长特性.利用一年生砧木生长旺盛、生物活性强的特点获得幼化接穗; 以腋芽诱导小叶为外植体, 相当于采用器官发生来克服顽拗现象(Singh et al., 2015 ; Díaz-Sala, 2019 ), 因为分生组织的诱导就是一种幼化过程; 而通过诱导、分化和增殖等器官发生刺激途径, 使增殖一代小叶外植体达到高度幼化, 是其在愈伤诱导、分化及不定芽增殖中表现出较强生长势及分裂活性的原因.本研究中, 不同幼化处理的外植体在愈伤诱导、分化、增殖及生长上均表现出显著差异, 即采用的外植体幼化程度越高, 后期生长潜力越大(于波等, 2020 ), 各项生长生理指标越优, 以增殖一代小叶为外植体效果最佳.研究发现, 随着郁金樱幼化程度的加大, 各项生理指标渐优, 而连续继代后随着幼化的深入, 能否逆转成年态? 这些问题尚无相关报道, 有待深入研究.我们也曾连续对郁金樱进行多次增殖继代, 发现随着继代次数的增加, 出现叶发黄、毒害作用加强、对激素敏感性增强及大量死亡等问题, 尚未尝试以增殖继代多次的小叶为外植体进行愈伤组织再生, 这些都有待后期深入研究. ...Ormosia henryi Prain闫国华等, 2002 ), 而这些由植物基因决定的生长生理规律通过后期植物激素和营养手段较难调控(郭希梅等, 2011 ), 从而增加了组织培养的难度.以冬樱花(Prunus majestica )叶片、花托、花梗和子房为外植体未分化出不定芽(和凤美等, 2010 ).现阶段成功分化出不定芽的樱花愈伤组织再生研究皆以叶片为外植体.福建山樱花以腋芽诱导小叶和茎段为外植体诱导愈伤组织, 仅腋芽诱导小叶分化出不定芽(邹娜等, 2013 ).雪落樱和御殿场樱以叶片为外植体虽分化出不定芽, 但仍存在褐化率高和分化困难等问题(史港影等, 2014 ; 雷巾茗, 2020 ), 这表明在樱花的愈伤组织再生研究中外植体的选择尤为重要, 可能与樱花基因型和外植体生理状态有关.研究表明, 在树番茄(Cyphomandra betacea ) (Correia et al., 2011 )、小粒咖啡(Coffea arabica ) (Bartos et al., 2018 )和冬青栎(Quercus ilex ) (Martínez et al., 2017 )等物种上, 胚性愈伤的诱导也很大程度依赖于外植体种类, 这种由基因驱动的外植体“抗拒”是很多品种无法建立稳定培养体系的主要原因(McCown, 2000 ).采用郁金樱不同分化程度的小叶诱导再生植株, 我们发现郁金樱在离体培养中表现出强烈的顽拗现象(recalcitrance), 这与樱桃砧木酸樱桃(P. cerasus )相似(Ben Mahmoud et al., 2017 ).本研究以多年生郁金樱小叶为外植体未分化出不定芽, 愈伤组织诱导率低且胚性愈伤较少, 同时随着培养时间的延长褐化现象严重, 顽拗现象出现于多年生小叶组织培养的诱导和分化阶段, 这与前人的研究结果一致(Hu et al., 2017 ; Wu et al., 2020 ).为克服遗传性基因导致的顽拗现象, 本研究采用郁金樱一年生嫁接苗小叶、腋芽诱导小叶及增殖一代小叶为外植体, 相当于间接对多年生郁金樱母株进行不同程度的幼化(rejuvennation)或返童, 幼年外植体相比发育成熟或分化组织具有更大的潜能(Wu et al., 2019 ).通过采集多年生郁金樱母株嫁接于一年生山樱花砧木上, 相当于直接操作原植物来克服郁金樱阵发性生长特性.利用一年生砧木生长旺盛、生物活性强的特点获得幼化接穗; 以腋芽诱导小叶为外植体, 相当于采用器官发生来克服顽拗现象(Singh et al., 2015 ; Díaz-Sala, 2019 ), 因为分生组织的诱导就是一种幼化过程; 而通过诱导、分化和增殖等器官发生刺激途径, 使增殖一代小叶外植体达到高度幼化, 是其在愈伤诱导、分化及不定芽增殖中表现出较强生长势及分裂活性的原因.本研究中, 不同幼化处理的外植体在愈伤诱导、分化、增殖及生长上均表现出显著差异, 即采用的外植体幼化程度越高, 后期生长潜力越大(于波等, 2020 ), 各项生长生理指标越优, 以增殖一代小叶为外植体效果最佳.研究发现, 随着郁金樱幼化程度的加大, 各项生理指标渐优, 而连续继代后随着幼化的深入, 能否逆转成年态? 这些问题尚无相关报道, 有待深入研究.我们也曾连续对郁金樱进行多次增殖继代, 发现随着继代次数的增加, 出现叶发黄、毒害作用加强、对激素敏感性增强及大量死亡等问题, 尚未尝试以增殖继代多次的小叶为外植体进行愈伤组织再生, 这些都有待后期深入研究. ...Arabidopsis somatic embryogenesis闫国华等, 2002 ), 而这些由植物基因决定的生长生理规律通过后期植物激素和营养手段较难调控(郭希梅等, 2011 ), 从而增加了组织培养的难度.以冬樱花(Prunus majestica )叶片、花托、花梗和子房为外植体未分化出不定芽(和凤美等, 2010 ).现阶段成功分化出不定芽的樱花愈伤组织再生研究皆以叶片为外植体.福建山樱花以腋芽诱导小叶和茎段为外植体诱导愈伤组织, 仅腋芽诱导小叶分化出不定芽(邹娜等, 2013 ).雪落樱和御殿场樱以叶片为外植体虽分化出不定芽, 但仍存在褐化率高和分化困难等问题(史港影等, 2014 ; 雷巾茗, 2020 ), 这表明在樱花的愈伤组织再生研究中外植体的选择尤为重要, 可能与樱花基因型和外植体生理状态有关.研究表明, 在树番茄(Cyphomandra betacea ) (Correia et al., 2011 )、小粒咖啡(Coffea arabica ) (Bartos et al., 2018 )和冬青栎(Quercus ilex ) (Martínez et al., 2017 )等物种上, 胚性愈伤的诱导也很大程度依赖于外植体种类, 这种由基因驱动的外植体“抗拒”是很多品种无法建立稳定培养体系的主要原因(McCown, 2000 ).采用郁金樱不同分化程度的小叶诱导再生植株, 我们发现郁金樱在离体培养中表现出强烈的顽拗现象(recalcitrance), 这与樱桃砧木酸樱桃(P. cerasus )相似(Ben Mahmoud et al., 2017 ).本研究以多年生郁金樱小叶为外植体未分化出不定芽, 愈伤组织诱导率低且胚性愈伤较少, 同时随着培养时间的延长褐化现象严重, 顽拗现象出现于多年生小叶组织培养的诱导和分化阶段, 这与前人的研究结果一致(Hu et al., 2017 ; Wu et al., 2020 ).为克服遗传性基因导致的顽拗现象, 本研究采用郁金樱一年生嫁接苗小叶、腋芽诱导小叶及增殖一代小叶为外植体, 相当于间接对多年生郁金樱母株进行不同程度的幼化(rejuvennation)或返童, 幼年外植体相比发育成熟或分化组织具有更大的潜能(Wu et al., 2019 ).通过采集多年生郁金樱母株嫁接于一年生山樱花砧木上, 相当于直接操作原植物来克服郁金樱阵发性生长特性.利用一年生砧木生长旺盛、生物活性强的特点获得幼化接穗; 以腋芽诱导小叶为外植体, 相当于采用器官发生来克服顽拗现象(Singh et al., 2015 ; Díaz-Sala, 2019 ), 因为分生组织的诱导就是一种幼化过程; 而通过诱导、分化和增殖等器官发生刺激途径, 使增殖一代小叶外植体达到高度幼化, 是其在愈伤诱导、分化及不定芽增殖中表现出较强生长势及分裂活性的原因.本研究中, 不同幼化处理的外植体在愈伤诱导、分化、增殖及生长上均表现出显著差异, 即采用的外植体幼化程度越高, 后期生长潜力越大(于波等, 2020 ), 各项生长生理指标越优, 以增殖一代小叶为外植体效果最佳.研究发现, 随着郁金樱幼化程度的加大, 各项生理指标渐优, 而连续继代后随着幼化的深入, 能否逆转成年态? 这些问题尚无相关报道, 有待深入研究.我们也曾连续对郁金樱进行多次增殖继代, 发现随着继代次数的增加, 出现叶发黄、毒害作用加强、对激素敏感性增强及大量死亡等问题, 尚未尝试以增殖继代多次的小叶为外植体进行愈伤组织再生, 这些都有待后期深入研究. ...  ,*贵州大学林学院, 贵阳 550025
,*贵州大学林学院, 贵阳 550025

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT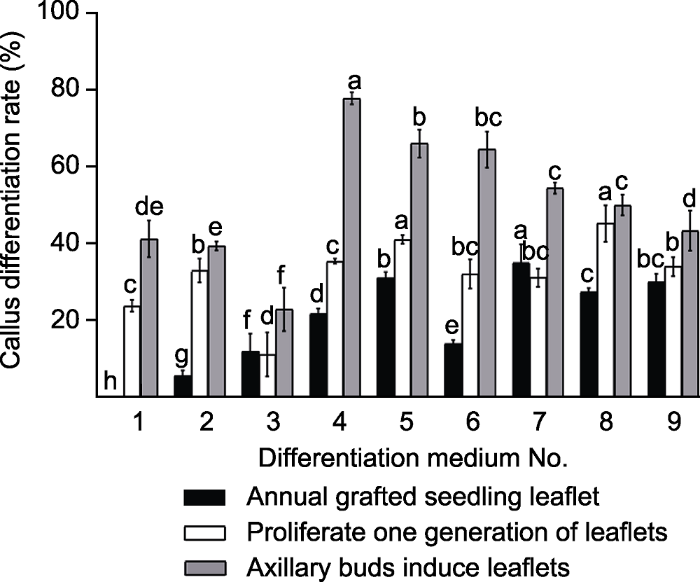 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT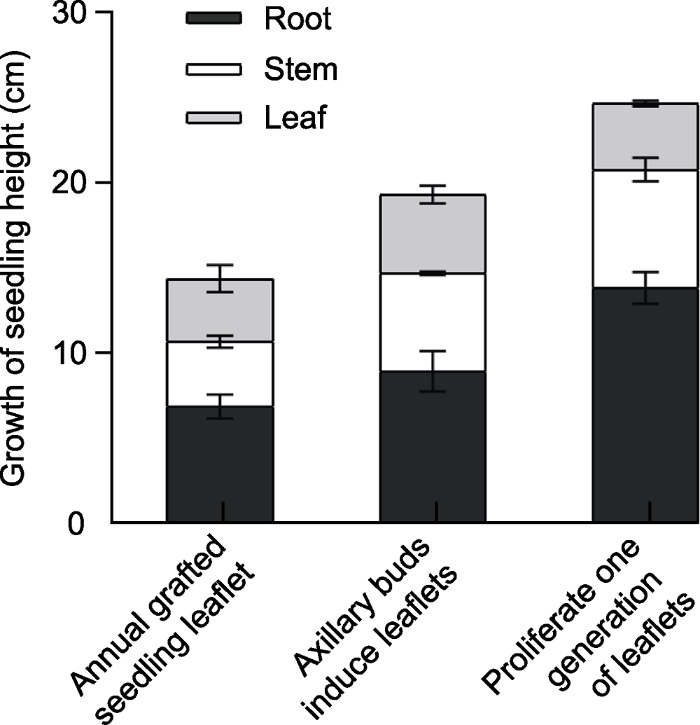 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}