Research Progress of CURT1 on Regulating Thylakoid Membrane Curvature
Qiuxin Li1,2, Wei Chi1,2, Daili Ji,1,*1Photosynthesis Research Center, Key Laboratory of Photobiology, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 2University of Chinese Academy of Sciences, Beijing 100049, China
Abstract The grana in chloroplast of higher plants is a structure composed of many thylakoid discs stacked together. The formation of the grana facilitates the distribution of the photosynthetic protein complex in different positions in thylakoids, that is, it has lateral heterogeneity and can effectively carry out photosynthesis. The key step to promote the formation of grana is to bend the thylakoid membrane. CURVATURE THYLAKOID 1 (CURT1) protein has been found to be the key factor leading to membrane bending. In this review, the recent research progresses of CURT1 protein in Arabidopsis thaliana and Cyanobacteria were summarized, and the prospect of CURT1 protein research in the future was put forward. Keywords:CURT1;membrane curvature;Arabidopsis; Cyanobacteria;CurT
PDF (1816KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 李秋信, 迟伟, 季代丽. CURT1调控类囊体膜弯曲的研究进展. 植物学报, 2021, 56(4): 462-469 doi:10.11983/CBB20194 Li Qiuxin, Chi Wei, Ji Daili. Research Progress of CURT1 on Regulating Thylakoid Membrane Curvature. Chinese Bulletin of Botany, 2021, 56(4): 462-469 doi:10.11983/CBB20194
高等植物的叶绿体是进行光合作用的重要细胞器, 由叶绿体被膜、类囊体和基质3部分组成。根据类囊体膜之间是否垛叠可分为基粒类囊体与基质类囊体(Pribil et al., 2014), 这两种类囊体膜并不是独立存在, 而是彼此相互连接, 构成一个连续且封闭的整体(Shimoni et al., 2005)。关于基粒类囊体的结构曾提出2种模型, 即螺旋模型和分叉模型(Daum and Kühlbrandt, 2011; Kirchhoff, 2013)。随着研究技术与仪器的发展, 目前更倾向于螺旋模型(Austin II and Staehelin, 2011)。基粒类囊体结构具有可变性, 当周围环境发生变化时, 基粒类囊体的超微结构会根据功能的需要发生改变, 但是叶绿体感知外部条件变化而促使类囊体膜结构发生改变的分子机制目前仍不十分清楚。直到2013年, Armbruster等(2013)发现弯曲类囊体膜蛋白1 (CURVATURE THYLAKOID1, CURT1)家族蛋白具有使类囊体膜发生弯曲的能力, 该蛋白家族以一种剂量依赖方式对类囊体超微结构的改变起重要调控作用。目前, 对CURT1蛋白的研究主要集中于拟南芥(Arabidopsis thaliana)和蓝细菌(Cyanobacteria)中。本文将分别对这两种光合生物中有关CURT1蛋白调控类囊体膜结构变化的最新研究进展进行阐述。
1 CURT1在拟南芥中的研究进展
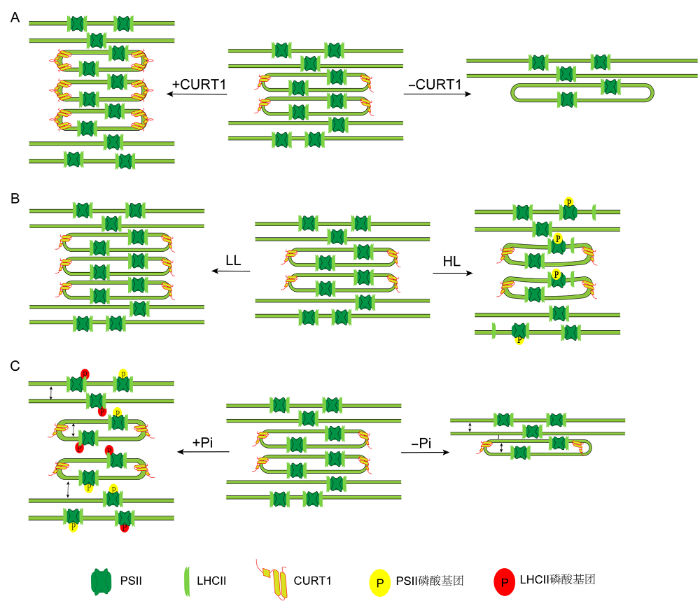
高等植物叶绿体形成基粒这一特殊功能结构的前提条件是使类囊体膜发生弯曲。Armbruster等(2013)在拟南芥中发现一种小分子蛋白CURT1具有使类囊体膜发生弯曲的能力。该类蛋白是由核基因编码、定位于叶绿体类囊体膜且由100多个氨基酸组成的小蛋白, 主要分布于基粒边缘(Armbruster et al., 2013)。CURT1蛋白家族有4种CURT1蛋白, 分别为CURT1A、CURT1B、CURT1C和CURT1D。其中CURT1A含量最丰富, CURT1D含量最少(Armbruster et al., 2013)。与其它光合蛋白复合体的含量相比, CURT1蛋白含量并不多, 却对类囊体膜结构起至关重要的作用。CURT1蛋白含量的减少和增多均会影响类囊体结构, 进而影响光合作用。其中, 对缺失突变体光合作用的影响远大于对过表达植株(Armbruster et al., 2013; Pribil et al., 2018)。在curt1a/b/c/d四突变植株中基粒直径变宽, 堆积层数减少, 形成没有基粒边缘的结构; 而过表达CURT1A植株中, 基粒直径变小, 堆积层数增加, 呈现管状的基粒片层(图1A)。值得注意的是, CURT1蛋白的改变并不影响光合蛋白复合体的含量, 光系统II (photosystem II, PSII)的功能也未受到直接损伤。因此认为在CURT1突变体中光合作用受到的影响是间接原因所致, 最可能的原因是CURT1蛋白含量变化导致类囊体的超微结构改变, 从而影响膜上蛋白的分布以及相关的生物学过程, 最终影响光合作用(Armbruster et al., 2013; Pribil et al., 2018)。
(A) Effects of CURT1 levels on granum dimensions: With the level of CURT1 proteins increased, the grana displays more layers of membrane and decreased diameter; Grana without CURT1 proteins significantly increased in diameter but contain far fewer layers of membrane; (B) Effects of changes in light conditions on granum dimensions: under low light levels, the numbers of layers in grana stacks are increased, and high light intensities lead to significant reduction in the diameter, and to partial transversal unstacking of grana discs; (C) Effects of PSII phosphorylation on granum dimensions: increased levels of PSII phosphorylation lead to significant reduction in the diameter, the enlargement of the vertical gaps between thylakoid layers, and swelling of the thylakoid lumen; Decreased levels of PSII phosphorylation lead to increased diameter, decreased gaps of adjacent layers within the granum, and fewer layers of membrane. LL: Low light; HL: High light; PSII: Photosystem II; LHC II: Light-harvesting complex II; CURT1: CURVATURE THYLAKOID 1
过表达以及缺失CURT1植株在高光下对PSII修复的影响以及弱光下对状态转变的影响。PSI: 光系统I。PSII、LHCII和CURT1同图1。 Figure 3CURT1 modulate grana structure to regulate photosynthesis in response to different light conditions
Overexpression or knock out of CURT1 protein influence the PSII repair process under high light condition and state transition during low light condition. PSI: Photosystem I. PSII, LHCII and CURT1 see Figure 1.
AustinII JR, StaehelinLA (2011). Three-dimensional architecture of grana and stroma thylakoids of higher plants as determined by electron tomography Plant Physiol 155, 1601-1611. DOI:10.1104/pp.110.170647URL [本文引用: 1]
ChuartzmanSG, NevoR, ShimoniE, CharuviD, KissV, OhadI, BrumfeldV, ReichZ (2008). Thylakoid membrane remodeling during state transitions in Arabidopsis Plant Cell 20, 1029-1039. DOI:10.1105/tpc.107.055830PMID:18398051 [本文引用: 1] Adaptability of oxygenic photosynthetic organisms to fluctuations in light spectral composition and intensity is conferred by state transitions, short-term regulatory processes that enable the photosynthetic apparatus to rapidly adjust to variations in light quality. In green algae and higher plants, these processes are accompanied by reversible structural rearrangements in the thylakoid membranes. We studied these structural changes in the thylakoid membranes of Arabidopsis thaliana chloroplasts using atomic force microscopy, scanning and transmission electron microscopy, and confocal imaging. Based on our results and on the recently determined three-dimensional structure of higher-plant thylakoids trapped in one of the two major light-adapted states, we propose a model for the transitions in membrane architecture. The model suggests that reorganization of the membranes involves fission and fusion events that occur at the interface between the appressed (granal) and nonappressed (stroma lamellar) domains of the thylakoid membranes. Vertical and lateral displacements of the grana layers presumably follow these localized events, eventually leading to macroscopic rearrangements of the entire membrane network.
DaumB, KühlbrandtW (2011). Electron tomography of plant thylakoid membranes J Exp Bot 62, 2393-2402. DOI:10.1093/jxb/err034URL [本文引用: 1]
GkekaP, SarkisovL (2010). Interactions of phospholipid bilayers with several classes of amphiphilic α-helical peptides: insights from coarse-grained molecular dynamics simulations J Phys Chem B 114, 826-839. DOI:10.1021/jp908320bURL [本文引用: 1]
HanssonM, VenerAV (2003). Identification of three previously unknown in vivo protein phosphorylation sites in thylakoid membranes of Arabidopsis thaliana Mol Cell Proteomics 2, 550-559. PMID:12883043 [本文引用: 1] The proteins in plant photosynthetic thylakoid membranes undergo light-induced phosphorylation, but only a few phosphoproteins have been characterized. To access the unknown sites of in vivo protein phosphorylation the thylakoid membranes were isolated from Arabidopsis thaliana grown in normal light, and the surface-exposed peptides were cleaved from the membranes by trypsin. The peptides were methylated and subjected to immobilized metal affinity chromatography, and the enriched phosphopeptides were sequenced using tandem nanospray quadrupole time-of-flight mass spectrometry. Three new phosphopeptides were revealed in addition to the five known phosphorylation sites in photosystem II proteins. All phosphopeptides are found phosphorylated at threonine residues implementing a strict threonine specificity of the thylakoid kinases. For the first time protein phosphorylation is found in photosystem I. The phosphorylation site is localized to the first threonine in the N terminus of PsaD protein that assists in the electron transfer from photosystem I to ferredoxin. A new phosphorylation site is also revealed in the acetylated N terminus of the minor chlorophyll a-binding protein CP29. The third novel phosphopeptide, composed of 25 amino acids, belongs to a nuclear encoded protein annotated as "expressed protein" in the Arabidopsis database. The protein precursor has a chloroplast-targeting peptide followed by the mature protein with two transmembrane helices and a molecular mass of 14 kDa. This previously uncharacterized protein is named thylakoid membrane phosphoprotein of 14 kDa (TMP14). The finding of the novel phosphoproteins extends involvement of the redox-regulated protein phosphorylation in photosynthetic membranes beyond the photosystem II and its light-harvesting antennae.
HeinzS, RastA, ShaoL, GutuA, GügelIL, HeynoE, LabsM, RengstlB, ViolaS, NowaczykMM, LeisterD, NickelsenJ (2016). Thylakoid membrane architecture in Synechocystis depends on CurT, a homolog of the granal CURVATURE THYLAKOID1 proteins Plant Cell 28, 2238-2260. DOI:10.1105/tpc.16.00491URL [本文引用: 6]
KirchhoffH (2014). Structural changes of the thylakoid membrane network induced by high light stress in plant chloroplasts Philos Trans R Soc Lond B Biol Sci 369, 20130225. [本文引用: 5]
KönnelA, BugaevaW, GügelIL, PhilipparK (2019). BANFF: bending of bilayer membranes by amphiphilic α-helices is necessary for form and function of organelles Biochem Cell Biol 97, 243-256. DOI:10.1139/bcb-2018-0150URL [本文引用: 1]
MarešJ, StruneckýO, BučinskáL, WiedermannováJ (2019). Evolutionary patterns of thylakoid architecture in cyanobacteria Front Microbiol 10, 277. DOI:10.3389/fmicb.2019.00277PMID:30853950 [本文引用: 1] While photosynthetic processes have become increasingly understood in cyanobacterial model strains, differences in the spatial distribution of thylakoid membranes among various lineages have been largely unexplored. Cyanobacterial cells exhibit an intriguing diversity in thylakoid arrangements, ranging from simple parietal to radial, coiled, parallel, and special types. Although metabolic background of their variability remains unknown, it has been suggested that thylakoid patterns are stable in certain phylogenetic clades. For decades, thylakoid arrangements have been used in cyanobacterial classification as one of the crucial characters for definition of taxa. The last comprehensive study addressing their evolutionary history in cyanobacteria was published 15 years ago. Since then both DNA sequence and electron microscopy data have grown rapidly. In the current study, we map ultrastructural data of >200 strains onto the SSU rRNA gene tree, and the resulting phylogeny is compared to a phylogenomic tree. Changes in thylakoid architecture in general follow the phylogeny of housekeeping loci. Parietal arrangement is resolved as the original thylakoid organization, evolving into complex arrangement in the most derived group of heterocytous cyanobacteria. Cyanobacteria occupying intermediate phylogenetic positions (greater filamentous, coccoid, and baeocytous types) exhibit fascicular, radial, and parallel arrangements, partly tracing the reconstructed course of phylogenetic branching. Contrary to previous studies, taxonomic value of thylakoid morphology seems very limited. Only special cases such as thylakoid absence or the parallel arrangement could be used as taxonomically informative apomorphies. The phylogenetic trees provide evidence of both paraphyly and reversion from more derived architectures in the simple parietal thylakoid pattern. Repeated convergent evolution is suggested for the radial and fascicular architectures. Moreover, thylakoid arrangement is constrained by cell size, excluding the occurrence of complex architectures in cyanobacteria smaller than 2 μm in width. It may further be dependent on unknown (eco)physiological factors as suggested by recurrence of the radial type in unrelated but morphologically similar cyanobacteria, and occurrence of special features throughout the phylogeny. No straightforward phylogenetic congruences have been found between proteins involved in photosynthesis and thylakoid formation, and the thylakoid patterns. Remarkably, several postulated thylakoid biogenesis factors are partly or completely missing in cyanobacteria, challenging their proposed essential roles.
McMahonHT, GallopJL (2005). Membrane curvature and mechanisms of dynamic cell membrane remodelling Nature 438, 590-596. DOI:10.1038/nature04396URL [本文引用: 1]
NixonPJ, MichouxF, YuJF, BoehmM, KomendaJ (2010). Recent advances in understanding the assembly and repair of photosystem II Ann Bot 106, 1-16. DOI:10.1093/aob/mcq059URL [本文引用: 1]
PeterBJ, KentHM, MillsIG, VallisY, ButlerPJG, EvansPR, McMahonHT (2004). BAR domains as sensors of membrane curvature: the amphiphysin BAR structure Science 303, 495-499. DOI:10.1126/science.1092586URL [本文引用: 1]
PribilM, LabsM, LeisterD (2014). Structure and dynamics of thylakoids in land plants J Exp Bot 65, 1955-1972. DOI:10.1093/jxb/eru090URL [本文引用: 3]
ShimoniE, Rav-HonO, OhadI, BrumfeldV, ReichZ (2005). Three-dimensional organization of higher-plant chloroplast thylakoid membranes revealed by electron tomography Plant Cell 17, 2580-2586. PMID:16055630 [本文引用: 1] The light-harvesting and energy-transducing functions of the chloroplast are performed within an intricate lamellar system of membranes, called thylakoid membranes, which are differentiated into granum and stroma lamellar domains. Using dual-axis electron microscope tomography, we determined the three-dimensional organization of the chloroplast thylakoid membranes within cryo-immobilized, freeze-substituted lettuce (Lactuca sativa) leaves. We found that the grana are built of repeating units that consist of paired layers formed by bifurcations of stroma lamellar sheets, which fuse within the granum body. These units are rotated relative to each other around the axis of the granum cylinder. One of the layers that makes up the pair bends upwards at its edge and fuses with the layer above it, whereas the other layer bends in the opposite direction and merges with the layer below. As a result, each unit in the granum is directly connected to its neighbors as well as to the surrounding stroma lamellae. This highly connected morphology has important consequences for the formation and function of the thylakoid membranes as well as for their stacking/unstacking response to variations in light conditions.
StengelA, GügelIL, HilgerD, RengstlB, JungH, NickelsenJ (2012). Initial steps of photosystem II de novo assembly and preloading with manganese take place in biogenesis centers in Synechocystis Plant Cell 24, 660-675. DOI:10.1105/tpc.111.093914URL [本文引用: 2]
TrottaA, BajwaAA, ManciniI, PaakkarinenV, PribilM, AroEM (2019). The role of phosphorylation dynamics of CURVATURE THYLAKOID 1B in plant thylakoid membranes Plant Physiol 181, 1615-1631. DOI:10.1104/pp.19.00942URL [本文引用: 5]
WoodWHJ, BarnettSFH, FlanneryS, HunterCN, JohnsonMP (2019). Dynamic thylakoid stacking is regulated by LHCII phosphorylation but not its interaction with PSI Plant Physiol 180, 2152-2166. DOI:10.1104/pp.19.00503URL [本文引用: 1]
Three-dimensional architecture of grana and stroma thylakoids of higher plants as determined by electron tomography 1 2011
... 高等植物的叶绿体是进行光合作用的重要细胞器, 由叶绿体被膜、类囊体和基质3部分组成.根据类囊体膜之间是否垛叠可分为基粒类囊体与基质类囊体(Pribil et al., 2014), 这两种类囊体膜并不是独立存在, 而是彼此相互连接, 构成一个连续且封闭的整体(Shimoni et al., 2005).关于基粒类囊体的结构曾提出2种模型, 即螺旋模型和分叉模型(Daum and Kühlbrandt, 2011; Kirchhoff, 2013).随着研究技术与仪器的发展, 目前更倾向于螺旋模型(Austin II and Staehelin, 2011).基粒类囊体结构具有可变性, 当周围环境发生变化时, 基粒类囊体的超微结构会根据功能的需要发生改变, 但是叶绿体感知外部条件变化而促使类囊体膜结构发生改变的分子机制目前仍不十分清楚.直到2013年, Armbruster等(2013)发现弯曲类囊体膜蛋白1 (CURVATURE THYLAKOID1, CURT1)家族蛋白具有使类囊体膜发生弯曲的能力, 该蛋白家族以一种剂量依赖方式对类囊体超微结构的改变起重要调控作用.目前, 对CURT1蛋白的研究主要集中于拟南芥(Arabidopsis thaliana)和蓝细菌(Cyanobacteria)中.本文将分别对这两种光合生物中有关CURT1蛋白调控类囊体膜结构变化的最新研究进展进行阐述. ...
Thylakoid membrane remodeling during state transitions in Arabidopsis 1 2008
... 典型的拟南芥基粒结构为: 类囊体膜厚度约为4 nm, 囊腔厚度为4.7 nm, 基粒直径为300-600 nm, 基粒内部类囊体膜间隙约为3.6 nm (Jensen and Leister, 2014).导致类囊体膜结构发生变化的因素有很多, 不同条件会使基粒的直径、囊腔厚度、类囊体膜间隙以及堆叠层数发生相应变化.首先是光照强度引起的变化: 弱光下, 基粒类囊体堆叠层数增多, 囊腔增厚(Anderson, 1986); 高光下, 基粒类囊体膜间隙变大, 基粒直径减小, 囊腔增厚, 基粒边缘面积增大, 且向外突出(Kirchhoff, 2014; Pribil et al., 2018) (图1B).其次是光质引起的变化: 红光或者蓝光有助于PSII和捕光蛋白复合物II (light-harvesting complex II, LHCII)磷酸化, 基粒直径变小, 膜间隙加厚; 远红光下, 优先激发光系统I (photosystem I, PSI), 使PSII与LHCII去磷酸化, 基粒直径变大(Chuartzman et al., 2008; Trotta et al., 2019).此外, 光系统蛋白磷酸化水平也会引起类囊体膜结构改变, 在stn7/stn8中, PSII和LHCII磷酸化水平下降, 基粒直径增大, 层数减少, 基粒中类囊体膜间隙变小, 囊腔厚度变小, 基粒变得宽且矮; tap38中, LHCII维持高水平磷酸化, 基粒直径变小, 层数增加, 基粒变窄且高(Wood et al., 2019) (图1C). ...
Electron tomography of plant thylakoid membranes 1 2011
... 高等植物的叶绿体是进行光合作用的重要细胞器, 由叶绿体被膜、类囊体和基质3部分组成.根据类囊体膜之间是否垛叠可分为基粒类囊体与基质类囊体(Pribil et al., 2014), 这两种类囊体膜并不是独立存在, 而是彼此相互连接, 构成一个连续且封闭的整体(Shimoni et al., 2005).关于基粒类囊体的结构曾提出2种模型, 即螺旋模型和分叉模型(Daum and Kühlbrandt, 2011; Kirchhoff, 2013).随着研究技术与仪器的发展, 目前更倾向于螺旋模型(Austin II and Staehelin, 2011).基粒类囊体结构具有可变性, 当周围环境发生变化时, 基粒类囊体的超微结构会根据功能的需要发生改变, 但是叶绿体感知外部条件变化而促使类囊体膜结构发生改变的分子机制目前仍不十分清楚.直到2013年, Armbruster等(2013)发现弯曲类囊体膜蛋白1 (CURVATURE THYLAKOID1, CURT1)家族蛋白具有使类囊体膜发生弯曲的能力, 该蛋白家族以一种剂量依赖方式对类囊体超微结构的改变起重要调控作用.目前, 对CURT1蛋白的研究主要集中于拟南芥(Arabidopsis thaliana)和蓝细菌(Cyanobacteria)中.本文将分别对这两种光合生物中有关CURT1蛋白调控类囊体膜结构变化的最新研究进展进行阐述. ...
Interactions of phospholipid bilayers with several classes of amphiphilic α-helical peptides: insights from coarse-grained molecular dynamics simulations 1 2010
... CURT1蛋白N端有1个两亲性螺旋, 该螺旋通常可像楔子一样插入到膜的一个小叶中, 其在膜中位置固定后, 就会使该位置的膜结构发生变化(Mcmahon and Gallop, 2005; Jarsch et al., 2016) (图2).两亲性α螺旋具有疏水性氨基酸的一面位于疏水尾部中, 带有正电荷的极性残基与带有负电荷的磷脂极性头部基团相互作用, 产生膜弯曲(Könnel, 2019).已知的两亲性α螺旋的作用方式有4类, CURT1蛋白的作用方式属于第2类.两亲性α螺旋平行于膜表面插入, 这种不对称插入导致严重膜弯曲, 膜弯曲程度取决于插入程度, 可以确定在CURT1中对膜弯曲产生关键作用的是两亲性螺旋(Gkeka and Sarkisov, 2010).然而, 目前CURT1两亲性螺旋调控类囊体膜结构变化的作用机理尚不明确, 还未发现与CURT1两亲性螺旋相互作用的蛋白, 因此无法判断CURT1是单独起作用还是需要其它蛋白把它固定在膜周围.通过分析CURT1过表达植株以及缺失突变体中类囊体膜结构的差异, 可以确定CURT1蛋白以剂量依赖方式对类囊体膜弯曲产生影响(Armbruster et al., 2013). ...
Identification of three previously unknown in vivo protein phosphorylation sites in thylakoid membranes of Arabidopsis thaliana 1 2003
... 比较CurT与CURT1的蛋白结构, 发现CurT蛋白的C端与CURT1的同源性较高, N端的相似性较低, 但仍具有预测的两亲性α螺旋, 进一步说明两亲性螺旋的存在是使CURT1蛋白具有膜弯曲能力的关键(Heinz et al., 2016).结构上的不同导致CurT与CURT1蛋白功能分化.首先, 在拟南芥中CURT1蛋白可发生磷酸化, 这种蛋白磷酸化的变化对类囊体超微结构的改变具有重要意义, 但是在蓝藻中CurT缺少磷酸化位点(Hansson and Vener, 2003).蓝藻中的CurT大部分位于类囊体上, 分子量较大, 主要有a、b和c型, 并且类囊体表面的CurT蛋白分布不对称, 呈现凸面多凹面少, 也表明CurT蛋白对类囊体的膜弯曲有作用.此外, 还有小部分CurT蛋白集中在生物发生中心PDMs (PratA-defined membranes) (PratA蛋白决定了PDM是否存在, 因此称为PratA定义的膜)上, PDM位于类囊体膜与质膜之间, 此区域的叶绿素荧光低, CurT以低分子量的形式存在, 主要是b和d型(Heinz et al., 2016).其次, 在蓝藻中CurT蛋白缺失导致生物发生中心PDM消失(Heinz et al., 2016), 在PDM上可能会进行类囊体膜的生物发生以及PSII组装的初始步骤(Stengel et al., 2012).PratA存在情况下, 可在周质中与Mn2+结合, 也可在PDM上与pD1的C端直接互作, 并由蛋白酶CtpA对pD1进行加工, 通过PratA可将Mn2+转运到PDM的pD1上, 随后该中间体再进行CP43和CP47的组装(Stengel et al., 2012).不存在CurT蛋白时, PratA含量下降, 生物发生中心消失, 不仅影响PSII组装的初始步骤(尤其是PSII二聚体的形成), 而且影响D1蛋白的降解.但即使缺少生物发生中心, 蓝细菌中也会有PSII积累, 表明生物发生中心并不是蓝细菌所必需的, 其存在对于PSII来说可能仅起到促进作用(Heinz et al., 2016). ...
Thylakoid membrane architecture in Synechocystis depends on CurT, a homolog of the granal CURVATURE THYLAKOID1 proteins 6 2016
 ,1,*
,1,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT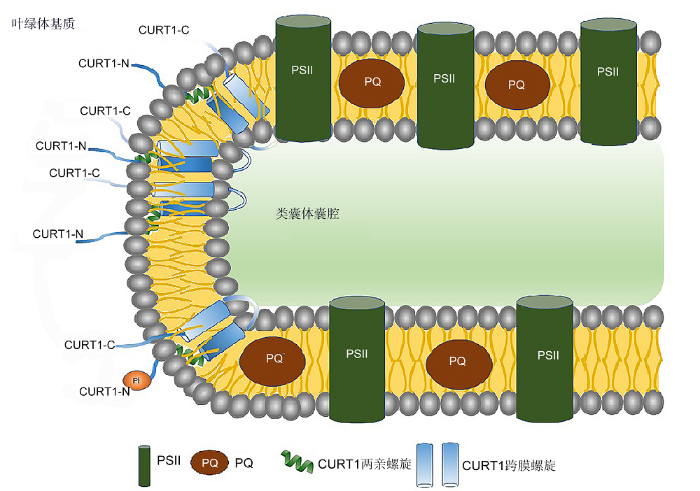 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT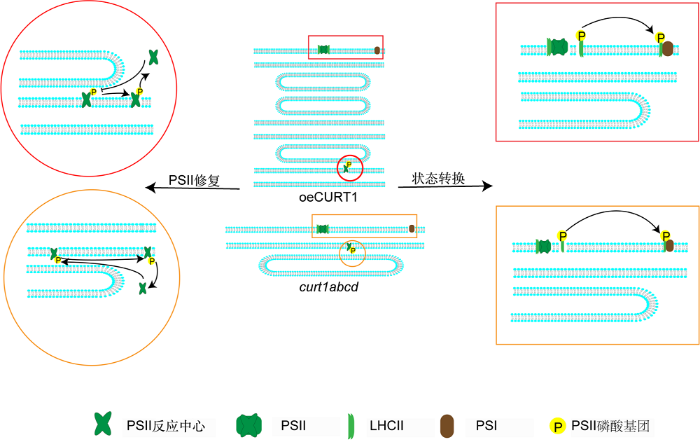 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}