Abstract During sexual plant reproduction, pollen-stigma recognition is a critical step for the germination of compatible pollen to ensure successful fertilization and genetic stability of offspring. It is also the first barrier for interspecific hybridization in crop breeding. Thus, great efforts have been made in relevant investigations during past decades. However, how the compatible pollen is recognized by the stigma remains a mystery. Recently, Chao Li’s group from East China Normal University published their work inScience, which reveals that POLLEN COAT PROTEIN B-class peptides (PCP-Bs) could compete with stigma RALF23/33 for binding to the ANJ-FER complex on stigma surface, resulting in a decline of stigmatic reactive oxygen species (ROS) that facilitates compatible pollen hydration. This finding represents a breakthrough in the field. Keywords:pollen-stigma recognition;pollen coat protein;ANJ-FER receptor kinase complex;peptide;reactive oxygen species (ROS)
PDF (658KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 王伟, 孙蒙祥. 花粉外被蛋白B类小肽, 花粉打开柱头大门的一把钥匙. 植物学报, 2021, 56(2): 147-150 doi:10.11983/CBB21048 Wang Wei, Sun Mengxiang. POLLENCOAT PROTEIN B-class Peptides (PCP-Bs), a Key of Compatible Pollen to Open the Gate of Stigma. Bulletin of Botany, 2021, 56(2): 147-150 doi:10.11983/CBB21048
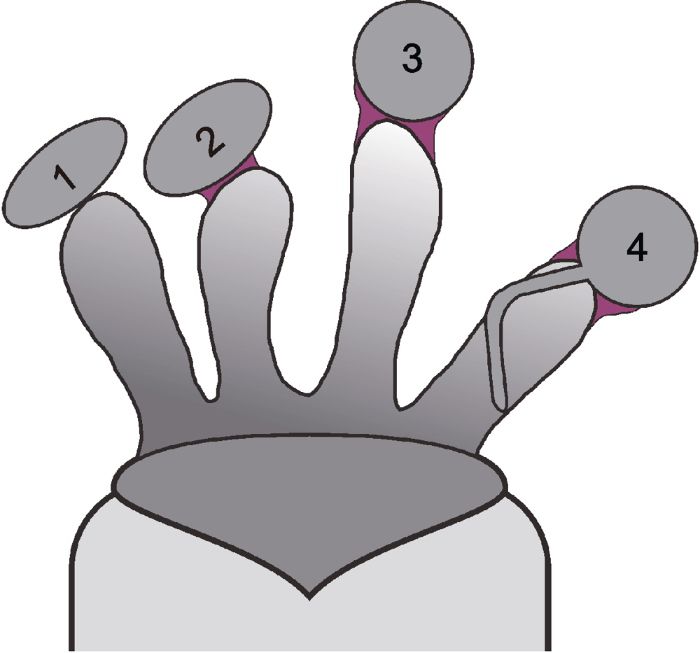
1: Pollen landing on the stigma; 2, 3: Pollen adhesion and hydration; 4: Pollen germination. After pollen adhesion a foot structure (red) established, which bridges pollen and papillar cells for the signaling between them. Currently, the known pollen factors involved in its hydration include pollen exine lipid and relevant lipidase, lipid binding proteins, and PCP-Bs, etc. The stigma factors involved in the hydration include cal-cium ion and Exo70A1 (vesicle trafficking), etc.
令人兴奋的是, 由于一项突破性研究进展, 这种尴尬的局面即将成为过去。近期, 国际权威杂志Science在线发表了华东师范大学生命科学学院李超团队撰写的研究论文“PCP-B peptides unlock a stigma 1 peptide-receptor kinase gated mechanism for pollination”(Liuet al.2021), 该文解析了第一句花粉-柱头间清晰的对答。
CabrillacD, CockJM, DumasC, GaudeT (2001). The S-locus receptor kinase is inhibited by thioredoxins and activated by pollen coat proteins Nature410,220-223. [本文引用: 1]
ChapmanLA, GoringDR (2010). Pollen-pistil interactions regulating successful fertilization in the Brassicaceae J Exp Bot61, 1987-1999. [本文引用: 1]
DoucetJ, LeeHK, GoringDR (2016). Pollen acceptance or rejection: a tale of two pathways Trends Plant Sci21,1058-1067. [本文引用: 1]
Heslop-HarrisonJ (1975). Incompatibility and the pollen- stigma interaction Annu Rev Plant Physiol26,403-425. [本文引用: 1]
Heslop-HarrisonY (1977). The pollen-stigma interaction: pollen-tube penetration in Crocus Ann Bot41,913-922. [本文引用: 1]
LinZ, EavesDJ, Sanchez-MoranE, FranklinFCH, Frank-alin-TongVE (2015). The Papaver rhoeas S determinants confer self-incompatibility to Arabidopsis thaliana in plan- ta Science350,684-687. [本文引用: 1]
MattssonO, KnoxRB, Heslop-HarrisonJ, Heslop-Harri-sonY (1974). Protein pellicle of stigmatic papillae as a probable recognition site in incompatibility reactions Nature247,298-300. [本文引用: 1]
NasrallahME, LiuP, NasrallahJB (2002). Generation of self-incompatible Arabidopsis thaliana by transfer of two S locus genes from A. lyrata Science297,247-249. [本文引用: 1]
SchopferCR, NasrallahME, NasrallahJB (1999). The male determinant of self-incompatibility in Brassica Science286, 1697-1700. [本文引用: 1]
TakayamaS, ShibaH, IwanoM, ShimosatoH, CheFS, KaiN, WatanabeM, SuzukiG, HinataK, IsogaiA (2000). The pollen determinant of self-incompatibility in Brassica campestris Proc Natl Acad Sci USA97, 1920- 1925. [本文引用: 1]
WangLD, ClarkeLA, EasonRJ, ParkerCC, QiBX, ScottRJ, DoughtyJ (2017). PCP-B class pollen coat proteins are key regulators of the hydration checkpoint in Arabidopsis thaliana pollen-stigma interactions New Phytol213,764-777. [本文引用: 1]
WheelerMJ, deGraaf BHJ, HadjiosifN, PerryRM, PoulterNS, OsmanK, VatovecS, HarperA, FranklinFCH, Franklin-TongVE (2009). Identification of the pollen self-incompatibility determinant in Papaver rhoeas Nature459,992-995. [本文引用: 1]
The S-locus receptor kinase is inhibited by thioredoxins and activated by pollen coat proteins 1 2001
... 20世纪70年代前后, 以Heslop-Harrison教授为代表的植物学家对各种植物花粉-柱头相互作用进行了长期细致的观察.经过研究, 形成了对花粉落到柱头上后, 通过黏附、水合和萌发等具体过程的基本认识; 形成了对不同类型柱头和柱头表面“接受”与“拒绝”花粉时形态结构特征的基本认识(图1); 也形成了花粉和柱头表面的某些蛋白可能是两者间识别位点的认识(Mattsson et al.1974; Heslop-Harrison, 1975, 1977).这些成果奠定了此后该领域研究的必要基础和基本格局.随着电镜技术和生化分析技术的广泛应用, 科学家对花粉外壁结构、柱头表面结构、花粉外壁蛋白和外被(pollen coat)蛋白有了更细致且准确的了解; 对柱头在亲和与不亲和反应中的动态变化, 包括同工酶、各种水解酶和表皮细胞角质层的动态变化有了全新的认识.20世纪90年代, 分子生物学技术快速发展, 给该领域研究带来了生机.在80年代研究的基础上, 自交不亲和机制的研究取得了重要进展.到21世纪初, 该研究已渐成系统, 明确了不同类型的自交不亲和系统的作用方式和主要作用机制(Schopfer et al.1999; Takayama et al.2000; Cabrillac et al.2001; Nasrallah et al.2002; Wheeler et al.2009; Lin et al.2015).相比之下, 关于花粉-柱头亲和作用机制的研究则缺乏实质性突破(Chapman and Goring, 2010; Doucet et al.2016).甚至进入21世纪, 尽管借助转录组和蛋白质组学分析手段, 获得了大量授粉前后差异表达基因与蛋白的信息, 研究者仍然感到困惑.一方面因为缺乏有效的分析手段, 很难快速锁定密切相关的基因或蛋白, 用以探讨其在花粉-柱头识别中的作用; 另一方面, 由于截至目前对花粉-柱头相互作用的了解仍很肤浅, 对其大致过程和途径都不甚了解, 因此难以判断这些数据对花粉-柱头相互作用研究的价值和有效性, 加之单基因突变常常没有明显表型, 需要经过长时间测试多基因的组合效应后才有可能做出明确判断.以致研究者即使坐拥丰富的数据库仍步履艰难.可以形象地说, 几十年来, 我们虽然明确地知道花粉-柱头在热烈的交谈, 也采用了各种技术手段企图聆听它们的谈话, 然而, 除了满耳似是而非的单词, 迄今为止仍未能清晰地听懂一句符合逻辑的对答. ...
Pollen-pistil interactions regulating successful fertilization in the Brassicaceae 1 2010
... 20世纪70年代前后, 以Heslop-Harrison教授为代表的植物学家对各种植物花粉-柱头相互作用进行了长期细致的观察.经过研究, 形成了对花粉落到柱头上后, 通过黏附、水合和萌发等具体过程的基本认识; 形成了对不同类型柱头和柱头表面“接受”与“拒绝”花粉时形态结构特征的基本认识(图1); 也形成了花粉和柱头表面的某些蛋白可能是两者间识别位点的认识(Mattsson et al.1974; Heslop-Harrison, 1975, 1977).这些成果奠定了此后该领域研究的必要基础和基本格局.随着电镜技术和生化分析技术的广泛应用, 科学家对花粉外壁结构、柱头表面结构、花粉外壁蛋白和外被(pollen coat)蛋白有了更细致且准确的了解; 对柱头在亲和与不亲和反应中的动态变化, 包括同工酶、各种水解酶和表皮细胞角质层的动态变化有了全新的认识.20世纪90年代, 分子生物学技术快速发展, 给该领域研究带来了生机.在80年代研究的基础上, 自交不亲和机制的研究取得了重要进展.到21世纪初, 该研究已渐成系统, 明确了不同类型的自交不亲和系统的作用方式和主要作用机制(Schopfer et al.1999; Takayama et al.2000; Cabrillac et al.2001; Nasrallah et al.2002; Wheeler et al.2009; Lin et al.2015).相比之下, 关于花粉-柱头亲和作用机制的研究则缺乏实质性突破(Chapman and Goring, 2010; Doucet et al.2016).甚至进入21世纪, 尽管借助转录组和蛋白质组学分析手段, 获得了大量授粉前后差异表达基因与蛋白的信息, 研究者仍然感到困惑.一方面因为缺乏有效的分析手段, 很难快速锁定密切相关的基因或蛋白, 用以探讨其在花粉-柱头识别中的作用; 另一方面, 由于截至目前对花粉-柱头相互作用的了解仍很肤浅, 对其大致过程和途径都不甚了解, 因此难以判断这些数据对花粉-柱头相互作用研究的价值和有效性, 加之单基因突变常常没有明显表型, 需要经过长时间测试多基因的组合效应后才有可能做出明确判断.以致研究者即使坐拥丰富的数据库仍步履艰难.可以形象地说, 几十年来, 我们虽然明确地知道花粉-柱头在热烈的交谈, 也采用了各种技术手段企图聆听它们的谈话, 然而, 除了满耳似是而非的单词, 迄今为止仍未能清晰地听懂一句符合逻辑的对答. ...
Pollen acceptance or rejection: a tale of two pathways 1 2016
... 20世纪70年代前后, 以Heslop-Harrison教授为代表的植物学家对各种植物花粉-柱头相互作用进行了长期细致的观察.经过研究, 形成了对花粉落到柱头上后, 通过黏附、水合和萌发等具体过程的基本认识; 形成了对不同类型柱头和柱头表面“接受”与“拒绝”花粉时形态结构特征的基本认识(图1); 也形成了花粉和柱头表面的某些蛋白可能是两者间识别位点的认识(Mattsson et al.1974; Heslop-Harrison, 1975, 1977).这些成果奠定了此后该领域研究的必要基础和基本格局.随着电镜技术和生化分析技术的广泛应用, 科学家对花粉外壁结构、柱头表面结构、花粉外壁蛋白和外被(pollen coat)蛋白有了更细致且准确的了解; 对柱头在亲和与不亲和反应中的动态变化, 包括同工酶、各种水解酶和表皮细胞角质层的动态变化有了全新的认识.20世纪90年代, 分子生物学技术快速发展, 给该领域研究带来了生机.在80年代研究的基础上, 自交不亲和机制的研究取得了重要进展.到21世纪初, 该研究已渐成系统, 明确了不同类型的自交不亲和系统的作用方式和主要作用机制(Schopfer et al.1999; Takayama et al.2000; Cabrillac et al.2001; Nasrallah et al.2002; Wheeler et al.2009; Lin et al.2015).相比之下, 关于花粉-柱头亲和作用机制的研究则缺乏实质性突破(Chapman and Goring, 2010; Doucet et al.2016).甚至进入21世纪, 尽管借助转录组和蛋白质组学分析手段, 获得了大量授粉前后差异表达基因与蛋白的信息, 研究者仍然感到困惑.一方面因为缺乏有效的分析手段, 很难快速锁定密切相关的基因或蛋白, 用以探讨其在花粉-柱头识别中的作用; 另一方面, 由于截至目前对花粉-柱头相互作用的了解仍很肤浅, 对其大致过程和途径都不甚了解, 因此难以判断这些数据对花粉-柱头相互作用研究的价值和有效性, 加之单基因突变常常没有明显表型, 需要经过长时间测试多基因的组合效应后才有可能做出明确判断.以致研究者即使坐拥丰富的数据库仍步履艰难.可以形象地说, 几十年来, 我们虽然明确地知道花粉-柱头在热烈的交谈, 也采用了各种技术手段企图聆听它们的谈话, 然而, 除了满耳似是而非的单词, 迄今为止仍未能清晰地听懂一句符合逻辑的对答. ...
Incompatibility and the pollen- stigma interaction 1 1975
... 20世纪70年代前后, 以Heslop-Harrison教授为代表的植物学家对各种植物花粉-柱头相互作用进行了长期细致的观察.经过研究, 形成了对花粉落到柱头上后, 通过黏附、水合和萌发等具体过程的基本认识; 形成了对不同类型柱头和柱头表面“接受”与“拒绝”花粉时形态结构特征的基本认识(图1); 也形成了花粉和柱头表面的某些蛋白可能是两者间识别位点的认识(Mattsson et al.1974; Heslop-Harrison, 1975, 1977).这些成果奠定了此后该领域研究的必要基础和基本格局.随着电镜技术和生化分析技术的广泛应用, 科学家对花粉外壁结构、柱头表面结构、花粉外壁蛋白和外被(pollen coat)蛋白有了更细致且准确的了解; 对柱头在亲和与不亲和反应中的动态变化, 包括同工酶、各种水解酶和表皮细胞角质层的动态变化有了全新的认识.20世纪90年代, 分子生物学技术快速发展, 给该领域研究带来了生机.在80年代研究的基础上, 自交不亲和机制的研究取得了重要进展.到21世纪初, 该研究已渐成系统, 明确了不同类型的自交不亲和系统的作用方式和主要作用机制(Schopfer et al.1999; Takayama et al.2000; Cabrillac et al.2001; Nasrallah et al.2002; Wheeler et al.2009; Lin et al.2015).相比之下, 关于花粉-柱头亲和作用机制的研究则缺乏实质性突破(Chapman and Goring, 2010; Doucet et al.2016).甚至进入21世纪, 尽管借助转录组和蛋白质组学分析手段, 获得了大量授粉前后差异表达基因与蛋白的信息, 研究者仍然感到困惑.一方面因为缺乏有效的分析手段, 很难快速锁定密切相关的基因或蛋白, 用以探讨其在花粉-柱头识别中的作用; 另一方面, 由于截至目前对花粉-柱头相互作用的了解仍很肤浅, 对其大致过程和途径都不甚了解, 因此难以判断这些数据对花粉-柱头相互作用研究的价值和有效性, 加之单基因突变常常没有明显表型, 需要经过长时间测试多基因的组合效应后才有可能做出明确判断.以致研究者即使坐拥丰富的数据库仍步履艰难.可以形象地说, 几十年来, 我们虽然明确地知道花粉-柱头在热烈的交谈, 也采用了各种技术手段企图聆听它们的谈话, 然而, 除了满耳似是而非的单词, 迄今为止仍未能清晰地听懂一句符合逻辑的对答. ...
The pollen-stigma interaction: pollen-tube penetration in Crocus 1 1977
... 20世纪70年代前后, 以Heslop-Harrison教授为代表的植物学家对各种植物花粉-柱头相互作用进行了长期细致的观察.经过研究, 形成了对花粉落到柱头上后, 通过黏附、水合和萌发等具体过程的基本认识; 形成了对不同类型柱头和柱头表面“接受”与“拒绝”花粉时形态结构特征的基本认识(图1); 也形成了花粉和柱头表面的某些蛋白可能是两者间识别位点的认识(Mattsson et al.1974; Heslop-Harrison, 1975, 1977).这些成果奠定了此后该领域研究的必要基础和基本格局.随着电镜技术和生化分析技术的广泛应用, 科学家对花粉外壁结构、柱头表面结构、花粉外壁蛋白和外被(pollen coat)蛋白有了更细致且准确的了解; 对柱头在亲和与不亲和反应中的动态变化, 包括同工酶、各种水解酶和表皮细胞角质层的动态变化有了全新的认识.20世纪90年代, 分子生物学技术快速发展, 给该领域研究带来了生机.在80年代研究的基础上, 自交不亲和机制的研究取得了重要进展.到21世纪初, 该研究已渐成系统, 明确了不同类型的自交不亲和系统的作用方式和主要作用机制(Schopfer et al.1999; Takayama et al.2000; Cabrillac et al.2001; Nasrallah et al.2002; Wheeler et al.2009; Lin et al.2015).相比之下, 关于花粉-柱头亲和作用机制的研究则缺乏实质性突破(Chapman and Goring, 2010; Doucet et al.2016).甚至进入21世纪, 尽管借助转录组和蛋白质组学分析手段, 获得了大量授粉前后差异表达基因与蛋白的信息, 研究者仍然感到困惑.一方面因为缺乏有效的分析手段, 很难快速锁定密切相关的基因或蛋白, 用以探讨其在花粉-柱头识别中的作用; 另一方面, 由于截至目前对花粉-柱头相互作用的了解仍很肤浅, 对其大致过程和途径都不甚了解, 因此难以判断这些数据对花粉-柱头相互作用研究的价值和有效性, 加之单基因突变常常没有明显表型, 需要经过长时间测试多基因的组合效应后才有可能做出明确判断.以致研究者即使坐拥丰富的数据库仍步履艰难.可以形象地说, 几十年来, 我们虽然明确地知道花粉-柱头在热烈的交谈, 也采用了各种技术手段企图聆听它们的谈话, 然而, 除了满耳似是而非的单词, 迄今为止仍未能清晰地听懂一句符合逻辑的对答. ...
The Papaver rhoeas S determinants confer self-incompatibility to Arabidopsis thaliana in plan- ta 1 2015
... 20世纪70年代前后, 以Heslop-Harrison教授为代表的植物学家对各种植物花粉-柱头相互作用进行了长期细致的观察.经过研究, 形成了对花粉落到柱头上后, 通过黏附、水合和萌发等具体过程的基本认识; 形成了对不同类型柱头和柱头表面“接受”与“拒绝”花粉时形态结构特征的基本认识(图1); 也形成了花粉和柱头表面的某些蛋白可能是两者间识别位点的认识(Mattsson et al.1974; Heslop-Harrison, 1975, 1977).这些成果奠定了此后该领域研究的必要基础和基本格局.随着电镜技术和生化分析技术的广泛应用, 科学家对花粉外壁结构、柱头表面结构、花粉外壁蛋白和外被(pollen coat)蛋白有了更细致且准确的了解; 对柱头在亲和与不亲和反应中的动态变化, 包括同工酶、各种水解酶和表皮细胞角质层的动态变化有了全新的认识.20世纪90年代, 分子生物学技术快速发展, 给该领域研究带来了生机.在80年代研究的基础上, 自交不亲和机制的研究取得了重要进展.到21世纪初, 该研究已渐成系统, 明确了不同类型的自交不亲和系统的作用方式和主要作用机制(Schopfer et al.1999; Takayama et al.2000; Cabrillac et al.2001; Nasrallah et al.2002; Wheeler et al.2009; Lin et al.2015).相比之下, 关于花粉-柱头亲和作用机制的研究则缺乏实质性突破(Chapman and Goring, 2010; Doucet et al.2016).甚至进入21世纪, 尽管借助转录组和蛋白质组学分析手段, 获得了大量授粉前后差异表达基因与蛋白的信息, 研究者仍然感到困惑.一方面因为缺乏有效的分析手段, 很难快速锁定密切相关的基因或蛋白, 用以探讨其在花粉-柱头识别中的作用; 另一方面, 由于截至目前对花粉-柱头相互作用的了解仍很肤浅, 对其大致过程和途径都不甚了解, 因此难以判断这些数据对花粉-柱头相互作用研究的价值和有效性, 加之单基因突变常常没有明显表型, 需要经过长时间测试多基因的组合效应后才有可能做出明确判断.以致研究者即使坐拥丰富的数据库仍步履艰难.可以形象地说, 几十年来, 我们虽然明确地知道花粉-柱头在热烈的交谈, 也采用了各种技术手段企图聆听它们的谈话, 然而, 除了满耳似是而非的单词, 迄今为止仍未能清晰地听懂一句符合逻辑的对答. ...
PCP-B peptides unlock a stigma 1 pep-tide-receptor kinase gated mechanism for pollination 1 2021
... 令人兴奋的是, 由于一项突破性研究进展, 这种尴尬的局面即将成为过去.近期, 国际权威杂志Science在线发表了华东师范大学生命科学学院李超团队撰写的研究论文“PCP-B peptides unlock a stigma 1 peptide-receptor kinase gated mechanism for pollination”(Liuet al.2021), 该文解析了第一句花粉-柱头间清晰的对答. ...
Protein pellicle of stigmatic papillae as a probable recognition site in incompatibility reactions 1 1974
... 20世纪70年代前后, 以Heslop-Harrison教授为代表的植物学家对各种植物花粉-柱头相互作用进行了长期细致的观察.经过研究, 形成了对花粉落到柱头上后, 通过黏附、水合和萌发等具体过程的基本认识; 形成了对不同类型柱头和柱头表面“接受”与“拒绝”花粉时形态结构特征的基本认识(图1); 也形成了花粉和柱头表面的某些蛋白可能是两者间识别位点的认识(Mattsson et al.1974; Heslop-Harrison, 1975, 1977).这些成果奠定了此后该领域研究的必要基础和基本格局.随着电镜技术和生化分析技术的广泛应用, 科学家对花粉外壁结构、柱头表面结构、花粉外壁蛋白和外被(pollen coat)蛋白有了更细致且准确的了解; 对柱头在亲和与不亲和反应中的动态变化, 包括同工酶、各种水解酶和表皮细胞角质层的动态变化有了全新的认识.20世纪90年代, 分子生物学技术快速发展, 给该领域研究带来了生机.在80年代研究的基础上, 自交不亲和机制的研究取得了重要进展.到21世纪初, 该研究已渐成系统, 明确了不同类型的自交不亲和系统的作用方式和主要作用机制(Schopfer et al.1999; Takayama et al.2000; Cabrillac et al.2001; Nasrallah et al.2002; Wheeler et al.2009; Lin et al.2015).相比之下, 关于花粉-柱头亲和作用机制的研究则缺乏实质性突破(Chapman and Goring, 2010; Doucet et al.2016).甚至进入21世纪, 尽管借助转录组和蛋白质组学分析手段, 获得了大量授粉前后差异表达基因与蛋白的信息, 研究者仍然感到困惑.一方面因为缺乏有效的分析手段, 很难快速锁定密切相关的基因或蛋白, 用以探讨其在花粉-柱头识别中的作用; 另一方面, 由于截至目前对花粉-柱头相互作用的了解仍很肤浅, 对其大致过程和途径都不甚了解, 因此难以判断这些数据对花粉-柱头相互作用研究的价值和有效性, 加之单基因突变常常没有明显表型, 需要经过长时间测试多基因的组合效应后才有可能做出明确判断.以致研究者即使坐拥丰富的数据库仍步履艰难.可以形象地说, 几十年来, 我们虽然明确地知道花粉-柱头在热烈的交谈, 也采用了各种技术手段企图聆听它们的谈话, 然而, 除了满耳似是而非的单词, 迄今为止仍未能清晰地听懂一句符合逻辑的对答. ...
Generation of self-incompatible Arabidopsis thaliana by transfer of two S locus genes from A. lyrata 1 2002
... 20世纪70年代前后, 以Heslop-Harrison教授为代表的植物学家对各种植物花粉-柱头相互作用进行了长期细致的观察.经过研究, 形成了对花粉落到柱头上后, 通过黏附、水合和萌发等具体过程的基本认识; 形成了对不同类型柱头和柱头表面“接受”与“拒绝”花粉时形态结构特征的基本认识(图1); 也形成了花粉和柱头表面的某些蛋白可能是两者间识别位点的认识(Mattsson et al.1974; Heslop-Harrison, 1975, 1977).这些成果奠定了此后该领域研究的必要基础和基本格局.随着电镜技术和生化分析技术的广泛应用, 科学家对花粉外壁结构、柱头表面结构、花粉外壁蛋白和外被(pollen coat)蛋白有了更细致且准确的了解; 对柱头在亲和与不亲和反应中的动态变化, 包括同工酶、各种水解酶和表皮细胞角质层的动态变化有了全新的认识.20世纪90年代, 分子生物学技术快速发展, 给该领域研究带来了生机.在80年代研究的基础上, 自交不亲和机制的研究取得了重要进展.到21世纪初, 该研究已渐成系统, 明确了不同类型的自交不亲和系统的作用方式和主要作用机制(Schopfer et al.1999; Takayama et al.2000; Cabrillac et al.2001; Nasrallah et al.2002; Wheeler et al.2009; Lin et al.2015).相比之下, 关于花粉-柱头亲和作用机制的研究则缺乏实质性突破(Chapman and Goring, 2010; Doucet et al.2016).甚至进入21世纪, 尽管借助转录组和蛋白质组学分析手段, 获得了大量授粉前后差异表达基因与蛋白的信息, 研究者仍然感到困惑.一方面因为缺乏有效的分析手段, 很难快速锁定密切相关的基因或蛋白, 用以探讨其在花粉-柱头识别中的作用; 另一方面, 由于截至目前对花粉-柱头相互作用的了解仍很肤浅, 对其大致过程和途径都不甚了解, 因此难以判断这些数据对花粉-柱头相互作用研究的价值和有效性, 加之单基因突变常常没有明显表型, 需要经过长时间测试多基因的组合效应后才有可能做出明确判断.以致研究者即使坐拥丰富的数据库仍步履艰难.可以形象地说, 几十年来, 我们虽然明确地知道花粉-柱头在热烈的交谈, 也采用了各种技术手段企图聆听它们的谈话, 然而, 除了满耳似是而非的单词, 迄今为止仍未能清晰地听懂一句符合逻辑的对答. ...
The male determinant of self-incompatibility in Brassica 1 1999
... 20世纪70年代前后, 以Heslop-Harrison教授为代表的植物学家对各种植物花粉-柱头相互作用进行了长期细致的观察.经过研究, 形成了对花粉落到柱头上后, 通过黏附、水合和萌发等具体过程的基本认识; 形成了对不同类型柱头和柱头表面“接受”与“拒绝”花粉时形态结构特征的基本认识(图1); 也形成了花粉和柱头表面的某些蛋白可能是两者间识别位点的认识(Mattsson et al.1974; Heslop-Harrison, 1975, 1977).这些成果奠定了此后该领域研究的必要基础和基本格局.随着电镜技术和生化分析技术的广泛应用, 科学家对花粉外壁结构、柱头表面结构、花粉外壁蛋白和外被(pollen coat)蛋白有了更细致且准确的了解; 对柱头在亲和与不亲和反应中的动态变化, 包括同工酶、各种水解酶和表皮细胞角质层的动态变化有了全新的认识.20世纪90年代, 分子生物学技术快速发展, 给该领域研究带来了生机.在80年代研究的基础上, 自交不亲和机制的研究取得了重要进展.到21世纪初, 该研究已渐成系统, 明确了不同类型的自交不亲和系统的作用方式和主要作用机制(Schopfer et al.1999; Takayama et al.2000; Cabrillac et al.2001; Nasrallah et al.2002; Wheeler et al.2009; Lin et al.2015).相比之下, 关于花粉-柱头亲和作用机制的研究则缺乏实质性突破(Chapman and Goring, 2010; Doucet et al.2016).甚至进入21世纪, 尽管借助转录组和蛋白质组学分析手段, 获得了大量授粉前后差异表达基因与蛋白的信息, 研究者仍然感到困惑.一方面因为缺乏有效的分析手段, 很难快速锁定密切相关的基因或蛋白, 用以探讨其在花粉-柱头识别中的作用; 另一方面, 由于截至目前对花粉-柱头相互作用的了解仍很肤浅, 对其大致过程和途径都不甚了解, 因此难以判断这些数据对花粉-柱头相互作用研究的价值和有效性, 加之单基因突变常常没有明显表型, 需要经过长时间测试多基因的组合效应后才有可能做出明确判断.以致研究者即使坐拥丰富的数据库仍步履艰难.可以形象地说, 几十年来, 我们虽然明确地知道花粉-柱头在热烈的交谈, 也采用了各种技术手段企图聆听它们的谈话, 然而, 除了满耳似是而非的单词, 迄今为止仍未能清晰地听懂一句符合逻辑的对答. ...
The pollen determinant of self-incompatibility in Brassica campestris 1 2000
... 20世纪70年代前后, 以Heslop-Harrison教授为代表的植物学家对各种植物花粉-柱头相互作用进行了长期细致的观察.经过研究, 形成了对花粉落到柱头上后, 通过黏附、水合和萌发等具体过程的基本认识; 形成了对不同类型柱头和柱头表面“接受”与“拒绝”花粉时形态结构特征的基本认识(图1); 也形成了花粉和柱头表面的某些蛋白可能是两者间识别位点的认识(Mattsson et al.1974; Heslop-Harrison, 1975, 1977).这些成果奠定了此后该领域研究的必要基础和基本格局.随着电镜技术和生化分析技术的广泛应用, 科学家对花粉外壁结构、柱头表面结构、花粉外壁蛋白和外被(pollen coat)蛋白有了更细致且准确的了解; 对柱头在亲和与不亲和反应中的动态变化, 包括同工酶、各种水解酶和表皮细胞角质层的动态变化有了全新的认识.20世纪90年代, 分子生物学技术快速发展, 给该领域研究带来了生机.在80年代研究的基础上, 自交不亲和机制的研究取得了重要进展.到21世纪初, 该研究已渐成系统, 明确了不同类型的自交不亲和系统的作用方式和主要作用机制(Schopfer et al.1999; Takayama et al.2000; Cabrillac et al.2001; Nasrallah et al.2002; Wheeler et al.2009; Lin et al.2015).相比之下, 关于花粉-柱头亲和作用机制的研究则缺乏实质性突破(Chapman and Goring, 2010; Doucet et al.2016).甚至进入21世纪, 尽管借助转录组和蛋白质组学分析手段, 获得了大量授粉前后差异表达基因与蛋白的信息, 研究者仍然感到困惑.一方面因为缺乏有效的分析手段, 很难快速锁定密切相关的基因或蛋白, 用以探讨其在花粉-柱头识别中的作用; 另一方面, 由于截至目前对花粉-柱头相互作用的了解仍很肤浅, 对其大致过程和途径都不甚了解, 因此难以判断这些数据对花粉-柱头相互作用研究的价值和有效性, 加之单基因突变常常没有明显表型, 需要经过长时间测试多基因的组合效应后才有可能做出明确判断.以致研究者即使坐拥丰富的数据库仍步履艰难.可以形象地说, 几十年来, 我们虽然明确地知道花粉-柱头在热烈的交谈, 也采用了各种技术手段企图聆听它们的谈话, 然而, 除了满耳似是而非的单词, 迄今为止仍未能清晰地听懂一句符合逻辑的对答. ...
PCP-B class pollen coat proteins are key regulators of the hydration checkpoint in Arabidopsis thaliana pollen-stigma interactions 1 2017
 ,*
,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}