High-throughput Identification of Meiotic Anti-CO Mutants by Fluorescent Reporters
Fan Li, Jiwei Ruan,*Yunnan Key Laboratory of Flower Breeding, National Engineering Research Center for Ornamental Horticulture, Flower Research Institute, Yunnan Academy of Agricultural Sciences, Kunming 650200, China
Abstract The forward genetic approach has become a widespread methodology to reveal genetic factors involved in meiosis, such as the crossover negative regulators, which limit the class II crossover formation. Here we developed a forward genetics mutant screen to identify anti-CO mutants under the Col wild-type background of Arabidopsis thaliana. We isolated 18 mutant lines showing more than three-fold increase in male meiotic recombination frequency as compared with the wild type, including dominant and recessive mutants. Thus, the EMS screen based on fluorescent reporters allows for high-throughput identification of meiotic anti-CO mutants and provides a novel approach and genetic materials to study the molecular mechanism of meiotic recombination regulation. Keywords:meiosis;anti-CO mutants;fluorescent marker;high-throughput visual assay
PDF (2253KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 李帆, 阮继伟. 利用荧光标记高通量鉴定减数分裂重组抑制突变体. 植物学报, 2019, 54(4): 522-530 doi:10.11983/CBB18229 Li Fan, Ruan Jiwei. High-throughput Identification of Meiotic Anti-CO Mutants by Fluorescent Reporters. Chinese Bulletin of Botany, 2019, 54(4): 522-530 doi:10.11983/CBB18229
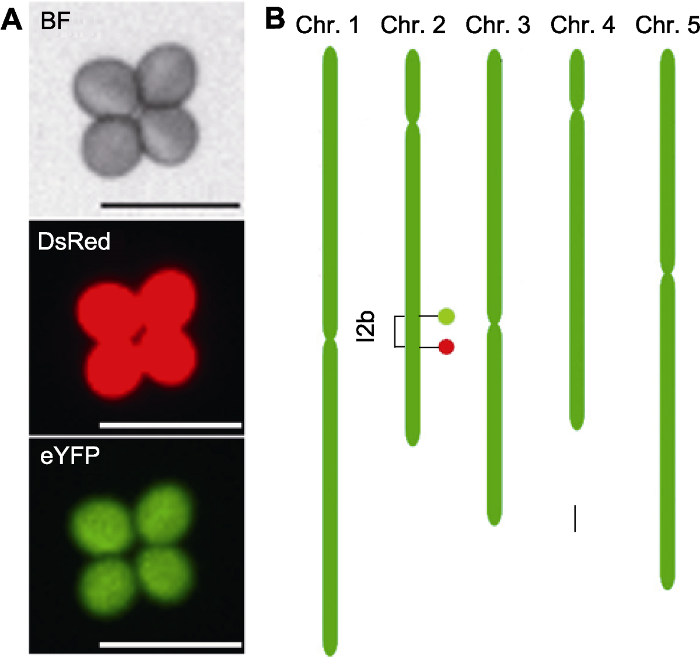
(A) Fluorescence micrograph of a tetrad pollen of FTL-I2b, which contains a DsRed and an eYFP fluorescent marker and expresses red fluorescence and green fluorescence under different fluorescence excitations respectively (Bars=50 μm); (B) The genomic location of FTL-I2b fluorescent markers on the chromosome (Bar=1 Mb), the DsRed and eYFP fluorescent markers are indicated by filled circles colored by red (down) and green (up), respectively, which constructing a I2b interval (1.45 Mb). BF: Bright field
(A) A schematic illustration of FTL-I2b cross with Col, the hybrid F1 contains a hemizygous fluorescent markers after hybridization, the recombined and unrecombined gametes are formed through meiosis; (B) Fluorescence micrograph of pollen in hybrid F1 generation, in which yellow pollen (GR) is unrecombined gamete, and monochromatic red (R) and green (G) pollen is recombined gametes (Bars=50 μm); (C) The recombination frequency (RF) of I2b interval can be calculated by counting the fluorescent marker pattern in the gametes via flow cytometry, the formula calculating recombination frequency=(G+R)/(2GR+G+R)×100.
为了确定EMS诱变最佳处理时间, 我们用75 mmol∙L-1 EMS分别对F1代种子进行不同的时间梯度处理(0、6、7、8和9小时, 每个处理400粒种子)。根据理想EMS诱变后50%的种子萌发率及在幼苗中出现白化和毛状等隐性表型的标准(Kim et al., 2006), 采用75 mmol∙L-1 EMS处理8小时为最佳诱变处理(图3)。由于EMS诱变在基因组中会产生多个点突变, 5 000株M1代突变群体足以满足在任意基因中发现突变的可能(Lukowitz et al., 2000; Qu and Qin, 2014)。同时, 受限于F1代杂交种子的制备效率, 我们对10 000粒F1代种子进行诱变, 获得了5 000多株M1代突变植株。
The forward genetics mutants screen was used the FTL-I2b containing linked hemizygous fluorescent reporters (I2b/++) on the genetic background of Arabidopsis wild type (WT). High-throughput detecting the recombination frequency by flow cytometry could identify the mutants with increased recombination rate. The DsRed and eYFP fluorescent reporters of FTL-I2b are indicated by R and G, respectively. The m refers to a dominant or recessive mutation. RF: Recombination frequency
(A) 拟南芥野生型(WT)花粉大小为(21.82±0.98) µm (n=10); (B) 大花粉突变体1 (big pollen 1)的大花粉率为100%, 花粉大小为(38.36±1.37) µm (n=10); (C) 大花粉突变体2 (big pollen 2)的大花粉率为50%, 花粉大小为(36.00±2.70) µm (n=10)。Bars=50 µm Figure 5The pollen micrograph of Arabidopsis wild type and big pollen mutants
(A) The pollen size of Arabidopsis wild type (WT) is (21.82±0.98) µm (n=10); (B) The large pollen rate of big pollen 1 mutant is 100%, pollen size is (38.36±1.37) µm (n=10); (C) The large pollen rate of big pollen 2 mutant is 50%, pollen size is (36.00±2.70) µm (n=10). Bars=50 µm
BerchowitzLE, CopenhaverGP ( 2008). Fluorescent Arabidopsis tetrads: a visual assay for quickly developing large crossover and crossover interference data sets Nat Protoc 3, 41-50. [本文引用: 2]
BerchowitzLE, CopenhaverGP ( 2010). Genetic interference: don't stand so close to me Curr Genomics 11, 91-102. [本文引用: 1]
BerchowitzLE, FrancisKE, BeyAL, CopenhaverGP ( 2007). The role of AtMUS81 in interference-insensitive crossovers in A. thaliana PLoS Genet 3, e132. [本文引用: 1]
HigginsJD, BucklingEF, FranklinFC, JonesGH ( 2008). Expression and functional analysis of AtMUS81 in Arabidopsis meiosis reveals a role in the second pathway of crossing-over Plant J 54, 152-162. [本文引用: 2]
HuQ, LiYF, WangHJ, ShenY, ZhangC, DuGJ, TangD, ChengZK ( 2017). Meiotic chromosome association 1 interacts with TOP3α and regulates meiotic recombination in rice Plant Cell 29, 1697-1708. [本文引用: 1]
HuangJY, ChengZH, WangC, HongY, SuH, WangJ, CopenhaverGP, MaH, WangYX ( 2015). Formation of interference-sensitive meiotic cross-overs requires sufficient DNA leading-strand elongation Proc Natl Acad Sci USA 112, 12534-12539. [本文引用: 1]
LiF, De StormeN, GeelenD ( 2017). Dynamics of male meiotic recombination frequency during plant development using fluorescent tagged lines in Arabidopsis thaliana Sci Rep 7, 42535. [本文引用: 1]
LuPL, HanXW, QiJ, YangJG, WijeratneAJ, LiT, MaH ( 2012). Analysis of Arabidopsis genome-wide variations before and after meiosis and meiotic recombination by resequencing Landsberg erecta and all four products of a single meiosis Genome Res 22, 508-518. [本文引用: 1]
LuPL, WijeratneAJ, WangZJ, CopenhaverGP, MaH ( 2014). Arabidopsis PTD is required for type I crossover formation and affects recombination frequency in two different chromosomal regions J Genet Genomics 41, 165-175. [本文引用: 1]
LukowitzW, GillmorCS, ScheibleWR ( 2000). Positional cloning in Arabidopsis. Why it feels good to have a genome initiative working for you Plant Physiol 123, 795-806. [本文引用: 1]
MacaisneN, NovatchkovaM, PeireraL, VezonD, JolivetS, FrogerN, ChelyshevaL, GrelonM, MercierR ( 2008). SHOC1, an XPF endonuclease-related protein, is essential for the formation of class I meiotic crossovers Curr Biol 18, 1432-1437. [本文引用: 1]
MacaisneN, VignardJ, MercierR ( 2011). SHOC1 and PTD form an XPF-ERCC1-like complex that is required for formation of class I crossovers J Cell Sci 124, 2687-2691. [本文引用: 1]
MercierR, MézardC, JenczewskiE, MacaisneN, GrelonM ( 2015). The molecular biology of meiosis in plants Annu Rev Plant Biol 66, 297-327. [本文引用: 1]
QiJ, ChenYM, CopenhaverGP, MaH ( 2014). Detection of genomic variations and DNA polymorphisms and impact on analysis of meiotic recombination and genetic mapping Proc Natl Acad Sci USA 111, 10007-10012. [本文引用: 1]
QuLJ, QinGJ ( 2014). Generation and identification of Arabidopsis EMS mutants In: Sanchez-Serrano JJ, Salinas J, eds. Arabidopsis Protocols. Totowa: Humana Press. pp. 225-239. [本文引用: 1]
Séguéla-ArnaudM, ChoinardS, LarchevêqueC, GirardC, FrogerN, CrismaniW, MercierR ( 2017). RMI1 and TOP3α limit meiotic CO formation through their C-terminal domains Nucleic Acids Res 4, 1860-1871. [本文引用: 1]
Séguéla-ArnaudM, CrismaniW, LarchevêqueC, MazelJ, FrogerN, ChoinardS, LemhemdiA, MacaisneN, Van LeeneJ, GevaertK, De JaegerG, ChelyshevaL, MercierR ( 2015). Multiple mechanisms limit meiotic crossovers: TOP3α and two BLM homologs antagonize crossovers in parallel to FANCM Proc Natl Acad Sci USA 112, 4713-4718. [本文引用: 1]
WangYX, ChengZH, HuangJY, ShiQ, HongY, CopenhaverGP, GongZZ, MaH ( 2012). The DNA replication factor RFC1 is required for interference-sensitive meiotic crossovers in Arabidopsis thaliana PLoS Genet 8, e1003039. [本文引用: 1]
YelinaNE, LambingC, HardcastleTJ, ZhaoXH, SantosB, HendersonIR ( 2015). DNA methylation epigenetically silences crossover hot spots and controls chromosomal domains of meiotic recombination in Arabidopsis Gene Dev 29, 2183-2202. [本文引用: 1]
YelinaNE, ZiolkowskiPA, MillerN, ZhaoX, KellyKA, Mu?ozDF, MannDJ, CopenhaverGP, HendersonIR ( 2013). High-throughput analysis of meiotic crossover frequency and interference via flow cytometry of fluorescent pollen in Arabidopsis thaliana Nat Protoc 8, 2119-2134. [本文引用: 2]
Fluorescent Arabidopsis tetrads: a visual assay for quickly developing large crossover and crossover interference data sets 2 2008
... 拟南芥花粉荧光标记系统(fluorescent tagged lines, FTLs)能高通量快速检测染色体特定标记区间的重组率(Yelina et al., 2013), 为高通量筛选减数分裂重组抑制突变体提供了可能.拟南芥花粉荧光标记系统是由一系列T-DNA插入片段编码的荧光标记构成, 由花粉特异性LAT52启动子激活, 在花粉中表达红荧光(DsRed)和绿荧光(eYFP), 通过统计花粉荧光标记的分离模式可以测量标记区间的重组率(Francis et al., 2007; Berchowitz and Copenhaver, 2008).本研究采用正向遗传学方法, 创新性地利用拟南芥花粉荧光标记系统和流式细胞技术对拟南芥野生型Col遗传背景下的EMS突变体进行大规模重组率检测和筛选, 进而获得重组率提高的突变体, 旨在为植物减数分裂重组抑制基因及其调控途径研究奠定基础. ...
... 我们的研究表明, 利用拟南芥花粉荧光标记系进行高通量重组抑制突变体筛选具有以下几个优势.首先, 该方法克服了基于zmm重组恢复系只能筛选获得II型交叉重组抑制基因的缺陷, 为发现I型交叉重组抑制基因提供了新途径.其次, 基于可视化的荧光标记突变体筛选, 不仅能鉴定出重组率提高的突变体, 还能筛选获得重组率降低和大花粉突变体等副产物, 为植物减数分裂相关分子调控机理研究提供了研究材料.更重要的是, 荧光标记系分别在拟南芥5条染色体上分布着大量荧光标记(包括35个DsRed、41个eYFP和37个eCFP), 这些荧光标记的不同组合为研究拟南芥不同染色体区间的重组率提供了强大而高效的工具(Francis et al., 2007; Berchowitz and Copenhaver, 2008; Li et al., 2017).例如, FTL-CEN3和FTL-CEN5的荧光标记分别横跨3号和5号染色体着丝粒区域, 而该区域为抑制减数分裂重组的“冷点”, 这为研究调控该区域的重组抑制基因提供了可视化的分析方法.因此, 利用不同荧光标记系进行高通量减数分裂重组抑制突变体的筛选, 鉴定减数分裂重组抑制突变体有待深入研究, 如染色体着丝粒异染色质区域的重组抑制突变体, 这对充分利用植物减数分裂重组的自然潜力进行育种具有重要意义. ...
Genetic interference: don't stand so close to me 1 2010
... 最近, 44个不同物种的基因组研究表明, 尽管不同物种基因组大小存在极大差异, 但重组率波动幅度并不大, 这表明减数分裂重组率受到严格的约束和限制(Fernandes et al., 2018).在大多数物种中, 78%的染色体重组率低于1.5 cM, 而其中绝大多数染色体的重组率仅为0.5 cM, 这意味着每个配对同源染色体只形成了1个交叉, 这与为保障同源染色体后期的精准分离至少需要形成1个交叉相一致(Jones and Franklin, 2006; Mercier et al., 2015).在大部分生物中, 交叉的形成和分布受到交叉干涉的影响, 即一个交叉的产生会抑制其沿染色体两侧一定范围内另一个交叉的形成, 导致交叉在染色体上呈非随机分布(Berchowitz and Copenhaver, 2010).根据对交叉干扰是否敏感, 可分为干涉敏感型(I型)和干涉不敏感型(II型)交叉(Hatkevich et al., 2017).近年来, 利用正向遗传学和全基因组测序已在拟南芥(Arabidopsis thaliana)中鉴定出3种减数分裂重组抑制途径: (1) FANCM与其DNA结合辅助因子MHF1/2的解旋酶途径(Crismani et al., 2012; Girard et al., 2014); (2) RECQ4A/B、TOP3α和RMI1形成的BTR重组抑制途径(Séguéla-Arnaud et al., 2015, 2017); (3) FIGL1- FLIP复合体调节单链入侵抑制II型交叉形成途径(Girard et al., 2015; Fernandes et al., 2017; Hu et al., 2017).这3种减数分裂重组抑制途径平行共存, 且通过不同机制限制II型交叉形成(Mieulet et al., 2018).然而, 在大多数真核生物减数分裂过程中, II型交叉仅占交叉总数的10%左右, 且依赖于MUS81和FANCD2调控途径(Berchowitz et al., 2007; Higgins et al., 2008; Kurzbauer et al., 2018), 绝大多数交叉主要通过I型交叉形成途径产生(Macaisne et al., 2008, 2011; Wang et al., 2012; Lu et al., 2014; Huang et al., 2015).更重要的是, 同时关闭I型和II型交叉形成途径后, 在拟南芥msh4/mus81/ fancd2突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
The role of AtMUS81 in interference-insensitive crossovers in A. thaliana 1 2007
... 最近, 44个不同物种的基因组研究表明, 尽管不同物种基因组大小存在极大差异, 但重组率波动幅度并不大, 这表明减数分裂重组率受到严格的约束和限制(Fernandes et al., 2018).在大多数物种中, 78%的染色体重组率低于1.5 cM, 而其中绝大多数染色体的重组率仅为0.5 cM, 这意味着每个配对同源染色体只形成了1个交叉, 这与为保障同源染色体后期的精准分离至少需要形成1个交叉相一致(Jones and Franklin, 2006; Mercier et al., 2015).在大部分生物中, 交叉的形成和分布受到交叉干涉的影响, 即一个交叉的产生会抑制其沿染色体两侧一定范围内另一个交叉的形成, 导致交叉在染色体上呈非随机分布(Berchowitz and Copenhaver, 2010).根据对交叉干扰是否敏感, 可分为干涉敏感型(I型)和干涉不敏感型(II型)交叉(Hatkevich et al., 2017).近年来, 利用正向遗传学和全基因组测序已在拟南芥(Arabidopsis thaliana)中鉴定出3种减数分裂重组抑制途径: (1) FANCM与其DNA结合辅助因子MHF1/2的解旋酶途径(Crismani et al., 2012; Girard et al., 2014); (2) RECQ4A/B、TOP3α和RMI1形成的BTR重组抑制途径(Séguéla-Arnaud et al., 2015, 2017); (3) FIGL1- FLIP复合体调节单链入侵抑制II型交叉形成途径(Girard et al., 2015; Fernandes et al., 2017; Hu et al., 2017).这3种减数分裂重组抑制途径平行共存, 且通过不同机制限制II型交叉形成(Mieulet et al., 2018).然而, 在大多数真核生物减数分裂过程中, II型交叉仅占交叉总数的10%左右, 且依赖于MUS81和FANCD2调控途径(Berchowitz et al., 2007; Higgins et al., 2008; Kurzbauer et al., 2018), 绝大多数交叉主要通过I型交叉形成途径产生(Macaisne et al., 2008, 2011; Wang et al., 2012; Lu et al., 2014; Huang et al., 2015).更重要的是, 同时关闭I型和II型交叉形成途径后, 在拟南芥msh4/mus81/ fancd2突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
FANCM limits meiotic crossovers 1 2012
... 最近, 44个不同物种的基因组研究表明, 尽管不同物种基因组大小存在极大差异, 但重组率波动幅度并不大, 这表明减数分裂重组率受到严格的约束和限制(Fernandes et al., 2018).在大多数物种中, 78%的染色体重组率低于1.5 cM, 而其中绝大多数染色体的重组率仅为0.5 cM, 这意味着每个配对同源染色体只形成了1个交叉, 这与为保障同源染色体后期的精准分离至少需要形成1个交叉相一致(Jones and Franklin, 2006; Mercier et al., 2015).在大部分生物中, 交叉的形成和分布受到交叉干涉的影响, 即一个交叉的产生会抑制其沿染色体两侧一定范围内另一个交叉的形成, 导致交叉在染色体上呈非随机分布(Berchowitz and Copenhaver, 2010).根据对交叉干扰是否敏感, 可分为干涉敏感型(I型)和干涉不敏感型(II型)交叉(Hatkevich et al., 2017).近年来, 利用正向遗传学和全基因组测序已在拟南芥(Arabidopsis thaliana)中鉴定出3种减数分裂重组抑制途径: (1) FANCM与其DNA结合辅助因子MHF1/2的解旋酶途径(Crismani et al., 2012; Girard et al., 2014); (2) RECQ4A/B、TOP3α和RMI1形成的BTR重组抑制途径(Séguéla-Arnaud et al., 2015, 2017); (3) FIGL1- FLIP复合体调节单链入侵抑制II型交叉形成途径(Girard et al., 2015; Fernandes et al., 2017; Hu et al., 2017).这3种减数分裂重组抑制途径平行共存, 且通过不同机制限制II型交叉形成(Mieulet et al., 2018).然而, 在大多数真核生物减数分裂过程中, II型交叉仅占交叉总数的10%左右, 且依赖于MUS81和FANCD2调控途径(Berchowitz et al., 2007; Higgins et al., 2008; Kurzbauer et al., 2018), 绝大多数交叉主要通过I型交叉形成途径产生(Macaisne et al., 2008, 2011; Wang et al., 2012; Lu et al., 2014; Huang et al., 2015).更重要的是, 同时关闭I型和II型交叉形成途径后, 在拟南芥msh4/mus81/ fancd2突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
FIGL1 and its novel partner FLIP form a conserved complex that regulates homologous recombination 1 2017
... 最近, 44个不同物种的基因组研究表明, 尽管不同物种基因组大小存在极大差异, 但重组率波动幅度并不大, 这表明减数分裂重组率受到严格的约束和限制(Fernandes et al., 2018).在大多数物种中, 78%的染色体重组率低于1.5 cM, 而其中绝大多数染色体的重组率仅为0.5 cM, 这意味着每个配对同源染色体只形成了1个交叉, 这与为保障同源染色体后期的精准分离至少需要形成1个交叉相一致(Jones and Franklin, 2006; Mercier et al., 2015).在大部分生物中, 交叉的形成和分布受到交叉干涉的影响, 即一个交叉的产生会抑制其沿染色体两侧一定范围内另一个交叉的形成, 导致交叉在染色体上呈非随机分布(Berchowitz and Copenhaver, 2010).根据对交叉干扰是否敏感, 可分为干涉敏感型(I型)和干涉不敏感型(II型)交叉(Hatkevich et al., 2017).近年来, 利用正向遗传学和全基因组测序已在拟南芥(Arabidopsis thaliana)中鉴定出3种减数分裂重组抑制途径: (1) FANCM与其DNA结合辅助因子MHF1/2的解旋酶途径(Crismani et al., 2012; Girard et al., 2014); (2) RECQ4A/B、TOP3α和RMI1形成的BTR重组抑制途径(Séguéla-Arnaud et al., 2015, 2017); (3) FIGL1- FLIP复合体调节单链入侵抑制II型交叉形成途径(Girard et al., 2015; Fernandes et al., 2017; Hu et al., 2017).这3种减数分裂重组抑制途径平行共存, 且通过不同机制限制II型交叉形成(Mieulet et al., 2018).然而, 在大多数真核生物减数分裂过程中, II型交叉仅占交叉总数的10%左右, 且依赖于MUS81和FANCD2调控途径(Berchowitz et al., 2007; Higgins et al., 2008; Kurzbauer et al., 2018), 绝大多数交叉主要通过I型交叉形成途径产生(Macaisne et al., 2008, 2011; Wang et al., 2012; Lu et al., 2014; Huang et al., 2015).更重要的是, 同时关闭I型和II型交叉形成途径后, 在拟南芥msh4/mus81/ fancd2突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
Unleashing meiotic crossovers in hybrid plants 1 2018
... 最近, 44个不同物种的基因组研究表明, 尽管不同物种基因组大小存在极大差异, 但重组率波动幅度并不大, 这表明减数分裂重组率受到严格的约束和限制(Fernandes et al., 2018).在大多数物种中, 78%的染色体重组率低于1.5 cM, 而其中绝大多数染色体的重组率仅为0.5 cM, 这意味着每个配对同源染色体只形成了1个交叉, 这与为保障同源染色体后期的精准分离至少需要形成1个交叉相一致(Jones and Franklin, 2006; Mercier et al., 2015).在大部分生物中, 交叉的形成和分布受到交叉干涉的影响, 即一个交叉的产生会抑制其沿染色体两侧一定范围内另一个交叉的形成, 导致交叉在染色体上呈非随机分布(Berchowitz and Copenhaver, 2010).根据对交叉干扰是否敏感, 可分为干涉敏感型(I型)和干涉不敏感型(II型)交叉(Hatkevich et al., 2017).近年来, 利用正向遗传学和全基因组测序已在拟南芥(Arabidopsis thaliana)中鉴定出3种减数分裂重组抑制途径: (1) FANCM与其DNA结合辅助因子MHF1/2的解旋酶途径(Crismani et al., 2012; Girard et al., 2014); (2) RECQ4A/B、TOP3α和RMI1形成的BTR重组抑制途径(Séguéla-Arnaud et al., 2015, 2017); (3) FIGL1- FLIP复合体调节单链入侵抑制II型交叉形成途径(Girard et al., 2015; Fernandes et al., 2017; Hu et al., 2017).这3种减数分裂重组抑制途径平行共存, 且通过不同机制限制II型交叉形成(Mieulet et al., 2018).然而, 在大多数真核生物减数分裂过程中, II型交叉仅占交叉总数的10%左右, 且依赖于MUS81和FANCD2调控途径(Berchowitz et al., 2007; Higgins et al., 2008; Kurzbauer et al., 2018), 绝大多数交叉主要通过I型交叉形成途径产生(Macaisne et al., 2008, 2011; Wang et al., 2012; Lu et al., 2014; Huang et al., 2015).更重要的是, 同时关闭I型和II型交叉形成途径后, 在拟南芥msh4/mus81/ fancd2突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
Pollen tetrad-based visual assay for meiotic recombination in Arabidopsis 2 2007
... 拟南芥花粉荧光标记系统(fluorescent tagged lines, FTLs)能高通量快速检测染色体特定标记区间的重组率(Yelina et al., 2013), 为高通量筛选减数分裂重组抑制突变体提供了可能.拟南芥花粉荧光标记系统是由一系列T-DNA插入片段编码的荧光标记构成, 由花粉特异性LAT52启动子激活, 在花粉中表达红荧光(DsRed)和绿荧光(eYFP), 通过统计花粉荧光标记的分离模式可以测量标记区间的重组率(Francis et al., 2007; Berchowitz and Copenhaver, 2008).本研究采用正向遗传学方法, 创新性地利用拟南芥花粉荧光标记系统和流式细胞技术对拟南芥野生型Col遗传背景下的EMS突变体进行大规模重组率检测和筛选, 进而获得重组率提高的突变体, 旨在为植物减数分裂重组抑制基因及其调控途径研究奠定基础. ...
... 我们的研究表明, 利用拟南芥花粉荧光标记系进行高通量重组抑制突变体筛选具有以下几个优势.首先, 该方法克服了基于zmm重组恢复系只能筛选获得II型交叉重组抑制基因的缺陷, 为发现I型交叉重组抑制基因提供了新途径.其次, 基于可视化的荧光标记突变体筛选, 不仅能鉴定出重组率提高的突变体, 还能筛选获得重组率降低和大花粉突变体等副产物, 为植物减数分裂相关分子调控机理研究提供了研究材料.更重要的是, 荧光标记系分别在拟南芥5条染色体上分布着大量荧光标记(包括35个DsRed、41个eYFP和37个eCFP), 这些荧光标记的不同组合为研究拟南芥不同染色体区间的重组率提供了强大而高效的工具(Francis et al., 2007; Berchowitz and Copenhaver, 2008; Li et al., 2017).例如, FTL-CEN3和FTL-CEN5的荧光标记分别横跨3号和5号染色体着丝粒区域, 而该区域为抑制减数分裂重组的“冷点”, 这为研究调控该区域的重组抑制基因提供了可视化的分析方法.因此, 利用不同荧光标记系进行高通量减数分裂重组抑制突变体的筛选, 鉴定减数分裂重组抑制突变体有待深入研究, 如染色体着丝粒异染色质区域的重组抑制突变体, 这对充分利用植物减数分裂重组的自然潜力进行育种具有重要意义. ...
AAA-ATPase FIDGETIN-LIKE 1 and helicase FANCM antagonize meiotic crossovers by distinct mechanisms 1 2015
... 最近, 44个不同物种的基因组研究表明, 尽管不同物种基因组大小存在极大差异, 但重组率波动幅度并不大, 这表明减数分裂重组率受到严格的约束和限制(Fernandes et al., 2018).在大多数物种中, 78%的染色体重组率低于1.5 cM, 而其中绝大多数染色体的重组率仅为0.5 cM, 这意味着每个配对同源染色体只形成了1个交叉, 这与为保障同源染色体后期的精准分离至少需要形成1个交叉相一致(Jones and Franklin, 2006; Mercier et al., 2015).在大部分生物中, 交叉的形成和分布受到交叉干涉的影响, 即一个交叉的产生会抑制其沿染色体两侧一定范围内另一个交叉的形成, 导致交叉在染色体上呈非随机分布(Berchowitz and Copenhaver, 2010).根据对交叉干扰是否敏感, 可分为干涉敏感型(I型)和干涉不敏感型(II型)交叉(Hatkevich et al., 2017).近年来, 利用正向遗传学和全基因组测序已在拟南芥(Arabidopsis thaliana)中鉴定出3种减数分裂重组抑制途径: (1) FANCM与其DNA结合辅助因子MHF1/2的解旋酶途径(Crismani et al., 2012; Girard et al., 2014); (2) RECQ4A/B、TOP3α和RMI1形成的BTR重组抑制途径(Séguéla-Arnaud et al., 2015, 2017); (3) FIGL1- FLIP复合体调节单链入侵抑制II型交叉形成途径(Girard et al., 2015; Fernandes et al., 2017; Hu et al., 2017).这3种减数分裂重组抑制途径平行共存, 且通过不同机制限制II型交叉形成(Mieulet et al., 2018).然而, 在大多数真核生物减数分裂过程中, II型交叉仅占交叉总数的10%左右, 且依赖于MUS81和FANCD2调控途径(Berchowitz et al., 2007; Higgins et al., 2008; Kurzbauer et al., 2018), 绝大多数交叉主要通过I型交叉形成途径产生(Macaisne et al., 2008, 2011; Wang et al., 2012; Lu et al., 2014; Huang et al., 2015).更重要的是, 同时关闭I型和II型交叉形成途径后, 在拟南芥msh4/mus81/ fancd2突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
FANCM-associated proteins MHF1 and MHF2, but not the other Fanconi anemia factors, limit meiotic crossovers 1 2014
... 最近, 44个不同物种的基因组研究表明, 尽管不同物种基因组大小存在极大差异, 但重组率波动幅度并不大, 这表明减数分裂重组率受到严格的约束和限制(Fernandes et al., 2018).在大多数物种中, 78%的染色体重组率低于1.5 cM, 而其中绝大多数染色体的重组率仅为0.5 cM, 这意味着每个配对同源染色体只形成了1个交叉, 这与为保障同源染色体后期的精准分离至少需要形成1个交叉相一致(Jones and Franklin, 2006; Mercier et al., 2015).在大部分生物中, 交叉的形成和分布受到交叉干涉的影响, 即一个交叉的产生会抑制其沿染色体两侧一定范围内另一个交叉的形成, 导致交叉在染色体上呈非随机分布(Berchowitz and Copenhaver, 2010).根据对交叉干扰是否敏感, 可分为干涉敏感型(I型)和干涉不敏感型(II型)交叉(Hatkevich et al., 2017).近年来, 利用正向遗传学和全基因组测序已在拟南芥(Arabidopsis thaliana)中鉴定出3种减数分裂重组抑制途径: (1) FANCM与其DNA结合辅助因子MHF1/2的解旋酶途径(Crismani et al., 2012; Girard et al., 2014); (2) RECQ4A/B、TOP3α和RMI1形成的BTR重组抑制途径(Séguéla-Arnaud et al., 2015, 2017); (3) FIGL1- FLIP复合体调节单链入侵抑制II型交叉形成途径(Girard et al., 2015; Fernandes et al., 2017; Hu et al., 2017).这3种减数分裂重组抑制途径平行共存, 且通过不同机制限制II型交叉形成(Mieulet et al., 2018).然而, 在大多数真核生物减数分裂过程中, II型交叉仅占交叉总数的10%左右, 且依赖于MUS81和FANCD2调控途径(Berchowitz et al., 2007; Higgins et al., 2008; Kurzbauer et al., 2018), 绝大多数交叉主要通过I型交叉形成途径产生(Macaisne et al., 2008, 2011; Wang et al., 2012; Lu et al., 2014; Huang et al., 2015).更重要的是, 同时关闭I型和II型交叉形成途径后, 在拟南芥msh4/mus81/ fancd2突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
Genome-wide crossover distribution in Arabidopsis thaliana meiosis reveals sex-specific patterns along chromosomes 1 2011
... 减数分裂重组起始于减数分裂初期产生的大量DNA双链断裂, 而只有极少数的断裂双链被修复形成交叉, 其余的大量断裂双链通过不同的途径和机制修复形成非交叉(Giraut et al., 2011; Lu et al., 2012; Qi et al., 2014).在模式植物拟南芥中, 细胞学分析表明每个减数分裂的细胞大约形成200个双链断裂, 但只有约10个断裂双链被修复形成交叉, 其余的断裂双链则被修复产生非交叉(Yelina et al., 2015).近年来, 利用拟南芥zmm突变体短角果表型(因缺乏I型交叉形成的ZMM基因而育性降低)进行果荚表型恢复系的正向遗传学筛选, 已经获得了多个重组率提高的突变体, 通过全基因组测序鉴定出多个II型交叉重组抑制基因, 如FANCM、RECQ4和FIGL1.然而, 由于该突变体筛选是建立在zmm突变体的基础之上, 不能揭示调控绝大多数交叉(I型交叉)形成的重组抑制基因.因此, 我们利用拟南芥花粉荧光标记系作为重组率增加的报告因子, 在拟南芥野生型(WT)遗传背景下进行EMS突变体的正向遗传学筛选, 共获得了18个重组率显著提高(3倍以上)的重组抑制突变体, 其中包括显性和隐性遗传突变.通常EMS诱变产生较多基因功能缺失的隐性突变体和少量功能获得型的显性突变体(Kim et al., 2006), 这与我们的筛选结果一致, 表明利用拟南芥花粉荧光标记系进行重组抑制突变体筛选是成功的.通过全基因组测序, 我们在隐性突变体中鉴定出1个包含FANCM-G540D突变位点的重组抑制突变体rrs1, 该突变体与fancm-1突变体具有相似的重组提高能力, 表明rrs1突变体可能是由FANCM-G540D突变造成, 这也进一步证明利用拟南芥花粉荧光标记系进行重组抑制突变体筛选可行. ...
... 最近, 44个不同物种的基因组研究表明, 尽管不同物种基因组大小存在极大差异, 但重组率波动幅度并不大, 这表明减数分裂重组率受到严格的约束和限制(Fernandes et al., 2018).在大多数物种中, 78%的染色体重组率低于1.5 cM, 而其中绝大多数染色体的重组率仅为0.5 cM, 这意味着每个配对同源染色体只形成了1个交叉, 这与为保障同源染色体后期的精准分离至少需要形成1个交叉相一致(Jones and Franklin, 2006; Mercier et al., 2015).在大部分生物中, 交叉的形成和分布受到交叉干涉的影响, 即一个交叉的产生会抑制其沿染色体两侧一定范围内另一个交叉的形成, 导致交叉在染色体上呈非随机分布(Berchowitz and Copenhaver, 2010).根据对交叉干扰是否敏感, 可分为干涉敏感型(I型)和干涉不敏感型(II型)交叉(Hatkevich et al., 2017).近年来, 利用正向遗传学和全基因组测序已在拟南芥(Arabidopsis thaliana)中鉴定出3种减数分裂重组抑制途径: (1) FANCM与其DNA结合辅助因子MHF1/2的解旋酶途径(Crismani et al., 2012; Girard et al., 2014); (2) RECQ4A/B、TOP3α和RMI1形成的BTR重组抑制途径(Séguéla-Arnaud et al., 2015, 2017); (3) FIGL1- FLIP复合体调节单链入侵抑制II型交叉形成途径(Girard et al., 2015; Fernandes et al., 2017; Hu et al., 2017).这3种减数分裂重组抑制途径平行共存, 且通过不同机制限制II型交叉形成(Mieulet et al., 2018).然而, 在大多数真核生物减数分裂过程中, II型交叉仅占交叉总数的10%左右, 且依赖于MUS81和FANCD2调控途径(Berchowitz et al., 2007; Higgins et al., 2008; Kurzbauer et al., 2018), 绝大多数交叉主要通过I型交叉形成途径产生(Macaisne et al., 2008, 2011; Wang et al., 2012; Lu et al., 2014; Huang et al., 2015).更重要的是, 同时关闭I型和II型交叉形成途径后, 在拟南芥msh4/mus81/ fancd2突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
Expression and functional analysis of AtMUS81 in Arabidopsis meiosis reveals a role in the second pathway of crossing-over 2 2008
... 最近, 44个不同物种的基因组研究表明, 尽管不同物种基因组大小存在极大差异, 但重组率波动幅度并不大, 这表明减数分裂重组率受到严格的约束和限制(Fernandes et al., 2018).在大多数物种中, 78%的染色体重组率低于1.5 cM, 而其中绝大多数染色体的重组率仅为0.5 cM, 这意味着每个配对同源染色体只形成了1个交叉, 这与为保障同源染色体后期的精准分离至少需要形成1个交叉相一致(Jones and Franklin, 2006; Mercier et al., 2015).在大部分生物中, 交叉的形成和分布受到交叉干涉的影响, 即一个交叉的产生会抑制其沿染色体两侧一定范围内另一个交叉的形成, 导致交叉在染色体上呈非随机分布(Berchowitz and Copenhaver, 2010).根据对交叉干扰是否敏感, 可分为干涉敏感型(I型)和干涉不敏感型(II型)交叉(Hatkevich et al., 2017).近年来, 利用正向遗传学和全基因组测序已在拟南芥(Arabidopsis thaliana)中鉴定出3种减数分裂重组抑制途径: (1) FANCM与其DNA结合辅助因子MHF1/2的解旋酶途径(Crismani et al., 2012; Girard et al., 2014); (2) RECQ4A/B、TOP3α和RMI1形成的BTR重组抑制途径(Séguéla-Arnaud et al., 2015, 2017); (3) FIGL1- FLIP复合体调节单链入侵抑制II型交叉形成途径(Girard et al., 2015; Fernandes et al., 2017; Hu et al., 2017).这3种减数分裂重组抑制途径平行共存, 且通过不同机制限制II型交叉形成(Mieulet et al., 2018).然而, 在大多数真核生物减数分裂过程中, II型交叉仅占交叉总数的10%左右, 且依赖于MUS81和FANCD2调控途径(Berchowitz et al., 2007; Higgins et al., 2008; Kurzbauer et al., 2018), 绝大多数交叉主要通过I型交叉形成途径产生(Macaisne et al., 2008, 2011; Wang et al., 2012; Lu et al., 2014; Huang et al., 2015).更重要的是, 同时关闭I型和II型交叉形成途径后, 在拟南芥msh4/mus81/ fancd2突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
... 突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
Meiotic chromosome association 1 interacts with TOP3α and regulates meiotic recombination in rice 1 2017
... 最近, 44个不同物种的基因组研究表明, 尽管不同物种基因组大小存在极大差异, 但重组率波动幅度并不大, 这表明减数分裂重组率受到严格的约束和限制(Fernandes et al., 2018).在大多数物种中, 78%的染色体重组率低于1.5 cM, 而其中绝大多数染色体的重组率仅为0.5 cM, 这意味着每个配对同源染色体只形成了1个交叉, 这与为保障同源染色体后期的精准分离至少需要形成1个交叉相一致(Jones and Franklin, 2006; Mercier et al., 2015).在大部分生物中, 交叉的形成和分布受到交叉干涉的影响, 即一个交叉的产生会抑制其沿染色体两侧一定范围内另一个交叉的形成, 导致交叉在染色体上呈非随机分布(Berchowitz and Copenhaver, 2010).根据对交叉干扰是否敏感, 可分为干涉敏感型(I型)和干涉不敏感型(II型)交叉(Hatkevich et al., 2017).近年来, 利用正向遗传学和全基因组测序已在拟南芥(Arabidopsis thaliana)中鉴定出3种减数分裂重组抑制途径: (1) FANCM与其DNA结合辅助因子MHF1/2的解旋酶途径(Crismani et al., 2012; Girard et al., 2014); (2) RECQ4A/B、TOP3α和RMI1形成的BTR重组抑制途径(Séguéla-Arnaud et al., 2015, 2017); (3) FIGL1- FLIP复合体调节单链入侵抑制II型交叉形成途径(Girard et al., 2015; Fernandes et al., 2017; Hu et al., 2017).这3种减数分裂重组抑制途径平行共存, 且通过不同机制限制II型交叉形成(Mieulet et al., 2018).然而, 在大多数真核生物减数分裂过程中, II型交叉仅占交叉总数的10%左右, 且依赖于MUS81和FANCD2调控途径(Berchowitz et al., 2007; Higgins et al., 2008; Kurzbauer et al., 2018), 绝大多数交叉主要通过I型交叉形成途径产生(Macaisne et al., 2008, 2011; Wang et al., 2012; Lu et al., 2014; Huang et al., 2015).更重要的是, 同时关闭I型和II型交叉形成途径后, 在拟南芥msh4/mus81/ fancd2突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
Formation of interference-sensitive meiotic cross-overs requires sufficient DNA leading-strand elongation 1 2015
... 最近, 44个不同物种的基因组研究表明, 尽管不同物种基因组大小存在极大差异, 但重组率波动幅度并不大, 这表明减数分裂重组率受到严格的约束和限制(Fernandes et al., 2018).在大多数物种中, 78%的染色体重组率低于1.5 cM, 而其中绝大多数染色体的重组率仅为0.5 cM, 这意味着每个配对同源染色体只形成了1个交叉, 这与为保障同源染色体后期的精准分离至少需要形成1个交叉相一致(Jones and Franklin, 2006; Mercier et al., 2015).在大部分生物中, 交叉的形成和分布受到交叉干涉的影响, 即一个交叉的产生会抑制其沿染色体两侧一定范围内另一个交叉的形成, 导致交叉在染色体上呈非随机分布(Berchowitz and Copenhaver, 2010).根据对交叉干扰是否敏感, 可分为干涉敏感型(I型)和干涉不敏感型(II型)交叉(Hatkevich et al., 2017).近年来, 利用正向遗传学和全基因组测序已在拟南芥(Arabidopsis thaliana)中鉴定出3种减数分裂重组抑制途径: (1) FANCM与其DNA结合辅助因子MHF1/2的解旋酶途径(Crismani et al., 2012; Girard et al., 2014); (2) RECQ4A/B、TOP3α和RMI1形成的BTR重组抑制途径(Séguéla-Arnaud et al., 2015, 2017); (3) FIGL1- FLIP复合体调节单链入侵抑制II型交叉形成途径(Girard et al., 2015; Fernandes et al., 2017; Hu et al., 2017).这3种减数分裂重组抑制途径平行共存, 且通过不同机制限制II型交叉形成(Mieulet et al., 2018).然而, 在大多数真核生物减数分裂过程中, II型交叉仅占交叉总数的10%左右, 且依赖于MUS81和FANCD2调控途径(Berchowitz et al., 2007; Higgins et al., 2008; Kurzbauer et al., 2018), 绝大多数交叉主要通过I型交叉形成途径产生(Macaisne et al., 2008, 2011; Wang et al., 2012; Lu et al., 2014; Huang et al., 2015).更重要的是, 同时关闭I型和II型交叉形成途径后, 在拟南芥msh4/mus81/ fancd2突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
Meiotic crossing-over: obligation and interference 1 2006
... 最近, 44个不同物种的基因组研究表明, 尽管不同物种基因组大小存在极大差异, 但重组率波动幅度并不大, 这表明减数分裂重组率受到严格的约束和限制(Fernandes et al., 2018).在大多数物种中, 78%的染色体重组率低于1.5 cM, 而其中绝大多数染色体的重组率仅为0.5 cM, 这意味着每个配对同源染色体只形成了1个交叉, 这与为保障同源染色体后期的精准分离至少需要形成1个交叉相一致(Jones and Franklin, 2006; Mercier et al., 2015).在大部分生物中, 交叉的形成和分布受到交叉干涉的影响, 即一个交叉的产生会抑制其沿染色体两侧一定范围内另一个交叉的形成, 导致交叉在染色体上呈非随机分布(Berchowitz and Copenhaver, 2010).根据对交叉干扰是否敏感, 可分为干涉敏感型(I型)和干涉不敏感型(II型)交叉(Hatkevich et al., 2017).近年来, 利用正向遗传学和全基因组测序已在拟南芥(Arabidopsis thaliana)中鉴定出3种减数分裂重组抑制途径: (1) FANCM与其DNA结合辅助因子MHF1/2的解旋酶途径(Crismani et al., 2012; Girard et al., 2014); (2) RECQ4A/B、TOP3α和RMI1形成的BTR重组抑制途径(Séguéla-Arnaud et al., 2015, 2017); (3) FIGL1- FLIP复合体调节单链入侵抑制II型交叉形成途径(Girard et al., 2015; Fernandes et al., 2017; Hu et al., 2017).这3种减数分裂重组抑制途径平行共存, 且通过不同机制限制II型交叉形成(Mieulet et al., 2018).然而, 在大多数真核生物减数分裂过程中, II型交叉仅占交叉总数的10%左右, 且依赖于MUS81和FANCD2调控途径(Berchowitz et al., 2007; Higgins et al., 2008; Kurzbauer et al., 2018), 绝大多数交叉主要通过I型交叉形成途径产生(Macaisne et al., 2008, 2011; Wang et al., 2012; Lu et al., 2014; Huang et al., 2015).更重要的是, 同时关闭I型和II型交叉形成途径后, 在拟南芥msh4/mus81/ fancd2突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
EMS mutagenesis of Arabidopsis 2 2006
... 为了确定EMS诱变最佳处理时间, 我们用75 mmol∙L-1 EMS分别对F1代种子进行不同的时间梯度处理(0、6、7、8和9小时, 每个处理400粒种子).根据理想EMS诱变后50%的种子萌发率及在幼苗中出现白化和毛状等隐性表型的标准(Kim et al., 2006), 采用75 mmol∙L-1 EMS处理8小时为最佳诱变处理(图3).由于EMS诱变在基因组中会产生多个点突变, 5 000株M1代突变群体足以满足在任意基因中发现突变的可能(Lukowitz et al., 2000; Qu and Qin, 2014).同时, 受限于F1代杂交种子的制备效率, 我们对10 000粒F1代种子进行诱变, 获得了5 000多株M1代突变植株. ...
... 减数分裂重组起始于减数分裂初期产生的大量DNA双链断裂, 而只有极少数的断裂双链被修复形成交叉, 其余的大量断裂双链通过不同的途径和机制修复形成非交叉(Giraut et al., 2011; Lu et al., 2012; Qi et al., 2014).在模式植物拟南芥中, 细胞学分析表明每个减数分裂的细胞大约形成200个双链断裂, 但只有约10个断裂双链被修复形成交叉, 其余的断裂双链则被修复产生非交叉(Yelina et al., 2015).近年来, 利用拟南芥zmm突变体短角果表型(因缺乏I型交叉形成的ZMM基因而育性降低)进行果荚表型恢复系的正向遗传学筛选, 已经获得了多个重组率提高的突变体, 通过全基因组测序鉴定出多个II型交叉重组抑制基因, 如FANCM、RECQ4和FIGL1.然而, 由于该突变体筛选是建立在zmm突变体的基础之上, 不能揭示调控绝大多数交叉(I型交叉)形成的重组抑制基因.因此, 我们利用拟南芥花粉荧光标记系作为重组率增加的报告因子, 在拟南芥野生型(WT)遗传背景下进行EMS突变体的正向遗传学筛选, 共获得了18个重组率显著提高(3倍以上)的重组抑制突变体, 其中包括显性和隐性遗传突变.通常EMS诱变产生较多基因功能缺失的隐性突变体和少量功能获得型的显性突变体(Kim et al., 2006), 这与我们的筛选结果一致, 表明利用拟南芥花粉荧光标记系进行重组抑制突变体筛选是成功的.通过全基因组测序, 我们在隐性突变体中鉴定出1个包含FANCM-G540D突变位点的重组抑制突变体rrs1, 该突变体与fancm-1突变体具有相似的重组提高能力, 表明rrs1突变体可能是由FANCM-G540D突变造成, 这也进一步证明利用拟南芥花粉荧光标记系进行重组抑制突变体筛选可行. ...
... 最近, 44个不同物种的基因组研究表明, 尽管不同物种基因组大小存在极大差异, 但重组率波动幅度并不大, 这表明减数分裂重组率受到严格的约束和限制(Fernandes et al., 2018).在大多数物种中, 78%的染色体重组率低于1.5 cM, 而其中绝大多数染色体的重组率仅为0.5 cM, 这意味着每个配对同源染色体只形成了1个交叉, 这与为保障同源染色体后期的精准分离至少需要形成1个交叉相一致(Jones and Franklin, 2006; Mercier et al., 2015).在大部分生物中, 交叉的形成和分布受到交叉干涉的影响, 即一个交叉的产生会抑制其沿染色体两侧一定范围内另一个交叉的形成, 导致交叉在染色体上呈非随机分布(Berchowitz and Copenhaver, 2010).根据对交叉干扰是否敏感, 可分为干涉敏感型(I型)和干涉不敏感型(II型)交叉(Hatkevich et al., 2017).近年来, 利用正向遗传学和全基因组测序已在拟南芥(Arabidopsis thaliana)中鉴定出3种减数分裂重组抑制途径: (1) FANCM与其DNA结合辅助因子MHF1/2的解旋酶途径(Crismani et al., 2012; Girard et al., 2014); (2) RECQ4A/B、TOP3α和RMI1形成的BTR重组抑制途径(Séguéla-Arnaud et al., 2015, 2017); (3) FIGL1- FLIP复合体调节单链入侵抑制II型交叉形成途径(Girard et al., 2015; Fernandes et al., 2017; Hu et al., 2017).这3种减数分裂重组抑制途径平行共存, 且通过不同机制限制II型交叉形成(Mieulet et al., 2018).然而, 在大多数真核生物减数分裂过程中, II型交叉仅占交叉总数的10%左右, 且依赖于MUS81和FANCD2调控途径(Berchowitz et al., 2007; Higgins et al., 2008; Kurzbauer et al., 2018), 绝大多数交叉主要通过I型交叉形成途径产生(Macaisne et al., 2008, 2011; Wang et al., 2012; Lu et al., 2014; Huang et al., 2015).更重要的是, 同时关闭I型和II型交叉形成途径后, 在拟南芥msh4/mus81/ fancd2突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
... ; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
Dynamics of male meiotic recombination frequency during plant development using fluorescent tagged lines in Arabidopsis thaliana 1 2017
... 我们的研究表明, 利用拟南芥花粉荧光标记系进行高通量重组抑制突变体筛选具有以下几个优势.首先, 该方法克服了基于zmm重组恢复系只能筛选获得II型交叉重组抑制基因的缺陷, 为发现I型交叉重组抑制基因提供了新途径.其次, 基于可视化的荧光标记突变体筛选, 不仅能鉴定出重组率提高的突变体, 还能筛选获得重组率降低和大花粉突变体等副产物, 为植物减数分裂相关分子调控机理研究提供了研究材料.更重要的是, 荧光标记系分别在拟南芥5条染色体上分布着大量荧光标记(包括35个DsRed、41个eYFP和37个eCFP), 这些荧光标记的不同组合为研究拟南芥不同染色体区间的重组率提供了强大而高效的工具(Francis et al., 2007; Berchowitz and Copenhaver, 2008; Li et al., 2017).例如, FTL-CEN3和FTL-CEN5的荧光标记分别横跨3号和5号染色体着丝粒区域, 而该区域为抑制减数分裂重组的“冷点”, 这为研究调控该区域的重组抑制基因提供了可视化的分析方法.因此, 利用不同荧光标记系进行高通量减数分裂重组抑制突变体的筛选, 鉴定减数分裂重组抑制突变体有待深入研究, 如染色体着丝粒异染色质区域的重组抑制突变体, 这对充分利用植物减数分裂重组的自然潜力进行育种具有重要意义. ...
Analysis of Arabidopsis genome-wide variations before and after meiosis and meiotic recombination by resequencing Landsberg erecta and all four products of a single meiosis 1 2012
... 减数分裂重组起始于减数分裂初期产生的大量DNA双链断裂, 而只有极少数的断裂双链被修复形成交叉, 其余的大量断裂双链通过不同的途径和机制修复形成非交叉(Giraut et al., 2011; Lu et al., 2012; Qi et al., 2014).在模式植物拟南芥中, 细胞学分析表明每个减数分裂的细胞大约形成200个双链断裂, 但只有约10个断裂双链被修复形成交叉, 其余的断裂双链则被修复产生非交叉(Yelina et al., 2015).近年来, 利用拟南芥zmm突变体短角果表型(因缺乏I型交叉形成的ZMM基因而育性降低)进行果荚表型恢复系的正向遗传学筛选, 已经获得了多个重组率提高的突变体, 通过全基因组测序鉴定出多个II型交叉重组抑制基因, 如FANCM、RECQ4和FIGL1.然而, 由于该突变体筛选是建立在zmm突变体的基础之上, 不能揭示调控绝大多数交叉(I型交叉)形成的重组抑制基因.因此, 我们利用拟南芥花粉荧光标记系作为重组率增加的报告因子, 在拟南芥野生型(WT)遗传背景下进行EMS突变体的正向遗传学筛选, 共获得了18个重组率显著提高(3倍以上)的重组抑制突变体, 其中包括显性和隐性遗传突变.通常EMS诱变产生较多基因功能缺失的隐性突变体和少量功能获得型的显性突变体(Kim et al., 2006), 这与我们的筛选结果一致, 表明利用拟南芥花粉荧光标记系进行重组抑制突变体筛选是成功的.通过全基因组测序, 我们在隐性突变体中鉴定出1个包含FANCM-G540D突变位点的重组抑制突变体rrs1, 该突变体与fancm-1突变体具有相似的重组提高能力, 表明rrs1突变体可能是由FANCM-G540D突变造成, 这也进一步证明利用拟南芥花粉荧光标记系进行重组抑制突变体筛选可行. ...
Arabidopsis PTD is required for type I crossover formation and affects recombination frequency in two different chromosomal regions 1 2014
... 最近, 44个不同物种的基因组研究表明, 尽管不同物种基因组大小存在极大差异, 但重组率波动幅度并不大, 这表明减数分裂重组率受到严格的约束和限制(Fernandes et al., 2018).在大多数物种中, 78%的染色体重组率低于1.5 cM, 而其中绝大多数染色体的重组率仅为0.5 cM, 这意味着每个配对同源染色体只形成了1个交叉, 这与为保障同源染色体后期的精准分离至少需要形成1个交叉相一致(Jones and Franklin, 2006; Mercier et al., 2015).在大部分生物中, 交叉的形成和分布受到交叉干涉的影响, 即一个交叉的产生会抑制其沿染色体两侧一定范围内另一个交叉的形成, 导致交叉在染色体上呈非随机分布(Berchowitz and Copenhaver, 2010).根据对交叉干扰是否敏感, 可分为干涉敏感型(I型)和干涉不敏感型(II型)交叉(Hatkevich et al., 2017).近年来, 利用正向遗传学和全基因组测序已在拟南芥(Arabidopsis thaliana)中鉴定出3种减数分裂重组抑制途径: (1) FANCM与其DNA结合辅助因子MHF1/2的解旋酶途径(Crismani et al., 2012; Girard et al., 2014); (2) RECQ4A/B、TOP3α和RMI1形成的BTR重组抑制途径(Séguéla-Arnaud et al., 2015, 2017); (3) FIGL1- FLIP复合体调节单链入侵抑制II型交叉形成途径(Girard et al., 2015; Fernandes et al., 2017; Hu et al., 2017).这3种减数分裂重组抑制途径平行共存, 且通过不同机制限制II型交叉形成(Mieulet et al., 2018).然而, 在大多数真核生物减数分裂过程中, II型交叉仅占交叉总数的10%左右, 且依赖于MUS81和FANCD2调控途径(Berchowitz et al., 2007; Higgins et al., 2008; Kurzbauer et al., 2018), 绝大多数交叉主要通过I型交叉形成途径产生(Macaisne et al., 2008, 2011; Wang et al., 2012; Lu et al., 2014; Huang et al., 2015).更重要的是, 同时关闭I型和II型交叉形成途径后, 在拟南芥msh4/mus81/ fancd2突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
Positional cloning in Arabidopsis. Why it feels good to have a genome initiative working for you 1 2000
... 为了确定EMS诱变最佳处理时间, 我们用75 mmol∙L-1 EMS分别对F1代种子进行不同的时间梯度处理(0、6、7、8和9小时, 每个处理400粒种子).根据理想EMS诱变后50%的种子萌发率及在幼苗中出现白化和毛状等隐性表型的标准(Kim et al., 2006), 采用75 mmol∙L-1 EMS处理8小时为最佳诱变处理(图3).由于EMS诱变在基因组中会产生多个点突变, 5 000株M1代突变群体足以满足在任意基因中发现突变的可能(Lukowitz et al., 2000; Qu and Qin, 2014).同时, 受限于F1代杂交种子的制备效率, 我们对10 000粒F1代种子进行诱变, 获得了5 000多株M1代突变植株. ...
SHOC1, an XPF endonuclease-related protein, is essential for the formation of class I meiotic crossovers 1 2008
... 最近, 44个不同物种的基因组研究表明, 尽管不同物种基因组大小存在极大差异, 但重组率波动幅度并不大, 这表明减数分裂重组率受到严格的约束和限制(Fernandes et al., 2018).在大多数物种中, 78%的染色体重组率低于1.5 cM, 而其中绝大多数染色体的重组率仅为0.5 cM, 这意味着每个配对同源染色体只形成了1个交叉, 这与为保障同源染色体后期的精准分离至少需要形成1个交叉相一致(Jones and Franklin, 2006; Mercier et al., 2015).在大部分生物中, 交叉的形成和分布受到交叉干涉的影响, 即一个交叉的产生会抑制其沿染色体两侧一定范围内另一个交叉的形成, 导致交叉在染色体上呈非随机分布(Berchowitz and Copenhaver, 2010).根据对交叉干扰是否敏感, 可分为干涉敏感型(I型)和干涉不敏感型(II型)交叉(Hatkevich et al., 2017).近年来, 利用正向遗传学和全基因组测序已在拟南芥(Arabidopsis thaliana)中鉴定出3种减数分裂重组抑制途径: (1) FANCM与其DNA结合辅助因子MHF1/2的解旋酶途径(Crismani et al., 2012; Girard et al., 2014); (2) RECQ4A/B、TOP3α和RMI1形成的BTR重组抑制途径(Séguéla-Arnaud et al., 2015, 2017); (3) FIGL1- FLIP复合体调节单链入侵抑制II型交叉形成途径(Girard et al., 2015; Fernandes et al., 2017; Hu et al., 2017).这3种减数分裂重组抑制途径平行共存, 且通过不同机制限制II型交叉形成(Mieulet et al., 2018).然而, 在大多数真核生物减数分裂过程中, II型交叉仅占交叉总数的10%左右, 且依赖于MUS81和FANCD2调控途径(Berchowitz et al., 2007; Higgins et al., 2008; Kurzbauer et al., 2018), 绝大多数交叉主要通过I型交叉形成途径产生(Macaisne et al., 2008, 2011; Wang et al., 2012; Lu et al., 2014; Huang et al., 2015).更重要的是, 同时关闭I型和II型交叉形成途径后, 在拟南芥msh4/mus81/ fancd2突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
SHOC1 and PTD form an XPF-ERCC1-like complex that is required for formation of class I crossovers 1 2011
... 最近, 44个不同物种的基因组研究表明, 尽管不同物种基因组大小存在极大差异, 但重组率波动幅度并不大, 这表明减数分裂重组率受到严格的约束和限制(Fernandes et al., 2018).在大多数物种中, 78%的染色体重组率低于1.5 cM, 而其中绝大多数染色体的重组率仅为0.5 cM, 这意味着每个配对同源染色体只形成了1个交叉, 这与为保障同源染色体后期的精准分离至少需要形成1个交叉相一致(Jones and Franklin, 2006; Mercier et al., 2015).在大部分生物中, 交叉的形成和分布受到交叉干涉的影响, 即一个交叉的产生会抑制其沿染色体两侧一定范围内另一个交叉的形成, 导致交叉在染色体上呈非随机分布(Berchowitz and Copenhaver, 2010).根据对交叉干扰是否敏感, 可分为干涉敏感型(I型)和干涉不敏感型(II型)交叉(Hatkevich et al., 2017).近年来, 利用正向遗传学和全基因组测序已在拟南芥(Arabidopsis thaliana)中鉴定出3种减数分裂重组抑制途径: (1) FANCM与其DNA结合辅助因子MHF1/2的解旋酶途径(Crismani et al., 2012; Girard et al., 2014); (2) RECQ4A/B、TOP3α和RMI1形成的BTR重组抑制途径(Séguéla-Arnaud et al., 2015, 2017); (3) FIGL1- FLIP复合体调节单链入侵抑制II型交叉形成途径(Girard et al., 2015; Fernandes et al., 2017; Hu et al., 2017).这3种减数分裂重组抑制途径平行共存, 且通过不同机制限制II型交叉形成(Mieulet et al., 2018).然而, 在大多数真核生物减数分裂过程中, II型交叉仅占交叉总数的10%左右, 且依赖于MUS81和FANCD2调控途径(Berchowitz et al., 2007; Higgins et al., 2008; Kurzbauer et al., 2018), 绝大多数交叉主要通过I型交叉形成途径产生(Macaisne et al., 2008, 2011; Wang et al., 2012; Lu et al., 2014; Huang et al., 2015).更重要的是, 同时关闭I型和II型交叉形成途径后, 在拟南芥msh4/mus81/ fancd2突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
The molecular biology of meiosis in plants 1 2015
... 最近, 44个不同物种的基因组研究表明, 尽管不同物种基因组大小存在极大差异, 但重组率波动幅度并不大, 这表明减数分裂重组率受到严格的约束和限制(Fernandes et al., 2018).在大多数物种中, 78%的染色体重组率低于1.5 cM, 而其中绝大多数染色体的重组率仅为0.5 cM, 这意味着每个配对同源染色体只形成了1个交叉, 这与为保障同源染色体后期的精准分离至少需要形成1个交叉相一致(Jones and Franklin, 2006; Mercier et al., 2015).在大部分生物中, 交叉的形成和分布受到交叉干涉的影响, 即一个交叉的产生会抑制其沿染色体两侧一定范围内另一个交叉的形成, 导致交叉在染色体上呈非随机分布(Berchowitz and Copenhaver, 2010).根据对交叉干扰是否敏感, 可分为干涉敏感型(I型)和干涉不敏感型(II型)交叉(Hatkevich et al., 2017).近年来, 利用正向遗传学和全基因组测序已在拟南芥(Arabidopsis thaliana)中鉴定出3种减数分裂重组抑制途径: (1) FANCM与其DNA结合辅助因子MHF1/2的解旋酶途径(Crismani et al., 2012; Girard et al., 2014); (2) RECQ4A/B、TOP3α和RMI1形成的BTR重组抑制途径(Séguéla-Arnaud et al., 2015, 2017); (3) FIGL1- FLIP复合体调节单链入侵抑制II型交叉形成途径(Girard et al., 2015; Fernandes et al., 2017; Hu et al., 2017).这3种减数分裂重组抑制途径平行共存, 且通过不同机制限制II型交叉形成(Mieulet et al., 2018).然而, 在大多数真核生物减数分裂过程中, II型交叉仅占交叉总数的10%左右, 且依赖于MUS81和FANCD2调控途径(Berchowitz et al., 2007; Higgins et al., 2008; Kurzbauer et al., 2018), 绝大多数交叉主要通过I型交叉形成途径产生(Macaisne et al., 2008, 2011; Wang et al., 2012; Lu et al., 2014; Huang et al., 2015).更重要的是, 同时关闭I型和II型交叉形成途径后, 在拟南芥msh4/mus81/ fancd2突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
Unleashing meiotic crossovers in crops 1 2018
... 最近, 44个不同物种的基因组研究表明, 尽管不同物种基因组大小存在极大差异, 但重组率波动幅度并不大, 这表明减数分裂重组率受到严格的约束和限制(Fernandes et al., 2018).在大多数物种中, 78%的染色体重组率低于1.5 cM, 而其中绝大多数染色体的重组率仅为0.5 cM, 这意味着每个配对同源染色体只形成了1个交叉, 这与为保障同源染色体后期的精准分离至少需要形成1个交叉相一致(Jones and Franklin, 2006; Mercier et al., 2015).在大部分生物中, 交叉的形成和分布受到交叉干涉的影响, 即一个交叉的产生会抑制其沿染色体两侧一定范围内另一个交叉的形成, 导致交叉在染色体上呈非随机分布(Berchowitz and Copenhaver, 2010).根据对交叉干扰是否敏感, 可分为干涉敏感型(I型)和干涉不敏感型(II型)交叉(Hatkevich et al., 2017).近年来, 利用正向遗传学和全基因组测序已在拟南芥(Arabidopsis thaliana)中鉴定出3种减数分裂重组抑制途径: (1) FANCM与其DNA结合辅助因子MHF1/2的解旋酶途径(Crismani et al., 2012; Girard et al., 2014); (2) RECQ4A/B、TOP3α和RMI1形成的BTR重组抑制途径(Séguéla-Arnaud et al., 2015, 2017); (3) FIGL1- FLIP复合体调节单链入侵抑制II型交叉形成途径(Girard et al., 2015; Fernandes et al., 2017; Hu et al., 2017).这3种减数分裂重组抑制途径平行共存, 且通过不同机制限制II型交叉形成(Mieulet et al., 2018).然而, 在大多数真核生物减数分裂过程中, II型交叉仅占交叉总数的10%左右, 且依赖于MUS81和FANCD2调控途径(Berchowitz et al., 2007; Higgins et al., 2008; Kurzbauer et al., 2018), 绝大多数交叉主要通过I型交叉形成途径产生(Macaisne et al., 2008, 2011; Wang et al., 2012; Lu et al., 2014; Huang et al., 2015).更重要的是, 同时关闭I型和II型交叉形成途径后, 在拟南芥msh4/mus81/ fancd2突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
Detection of genomic variations and DNA polymorphisms and impact on analysis of meiotic recombination and genetic mapping 1 2014
... 减数分裂重组起始于减数分裂初期产生的大量DNA双链断裂, 而只有极少数的断裂双链被修复形成交叉, 其余的大量断裂双链通过不同的途径和机制修复形成非交叉(Giraut et al., 2011; Lu et al., 2012; Qi et al., 2014).在模式植物拟南芥中, 细胞学分析表明每个减数分裂的细胞大约形成200个双链断裂, 但只有约10个断裂双链被修复形成交叉, 其余的断裂双链则被修复产生非交叉(Yelina et al., 2015).近年来, 利用拟南芥zmm突变体短角果表型(因缺乏I型交叉形成的ZMM基因而育性降低)进行果荚表型恢复系的正向遗传学筛选, 已经获得了多个重组率提高的突变体, 通过全基因组测序鉴定出多个II型交叉重组抑制基因, 如FANCM、RECQ4和FIGL1.然而, 由于该突变体筛选是建立在zmm突变体的基础之上, 不能揭示调控绝大多数交叉(I型交叉)形成的重组抑制基因.因此, 我们利用拟南芥花粉荧光标记系作为重组率增加的报告因子, 在拟南芥野生型(WT)遗传背景下进行EMS突变体的正向遗传学筛选, 共获得了18个重组率显著提高(3倍以上)的重组抑制突变体, 其中包括显性和隐性遗传突变.通常EMS诱变产生较多基因功能缺失的隐性突变体和少量功能获得型的显性突变体(Kim et al., 2006), 这与我们的筛选结果一致, 表明利用拟南芥花粉荧光标记系进行重组抑制突变体筛选是成功的.通过全基因组测序, 我们在隐性突变体中鉴定出1个包含FANCM-G540D突变位点的重组抑制突变体rrs1, 该突变体与fancm-1突变体具有相似的重组提高能力, 表明rrs1突变体可能是由FANCM-G540D突变造成, 这也进一步证明利用拟南芥花粉荧光标记系进行重组抑制突变体筛选可行. ...
Generation and identification of Arabidopsis EMS mutants 1 2014
... 为了确定EMS诱变最佳处理时间, 我们用75 mmol∙L-1 EMS分别对F1代种子进行不同的时间梯度处理(0、6、7、8和9小时, 每个处理400粒种子).根据理想EMS诱变后50%的种子萌发率及在幼苗中出现白化和毛状等隐性表型的标准(Kim et al., 2006), 采用75 mmol∙L-1 EMS处理8小时为最佳诱变处理(图3).由于EMS诱变在基因组中会产生多个点突变, 5 000株M1代突变群体足以满足在任意基因中发现突变的可能(Lukowitz et al., 2000; Qu and Qin, 2014).同时, 受限于F1代杂交种子的制备效率, 我们对10 000粒F1代种子进行诱变, 获得了5 000多株M1代突变植株. ...
RMI1 and TOP3α limit meiotic CO formation through their C-terminal domains 1 2017
... 最近, 44个不同物种的基因组研究表明, 尽管不同物种基因组大小存在极大差异, 但重组率波动幅度并不大, 这表明减数分裂重组率受到严格的约束和限制(Fernandes et al., 2018).在大多数物种中, 78%的染色体重组率低于1.5 cM, 而其中绝大多数染色体的重组率仅为0.5 cM, 这意味着每个配对同源染色体只形成了1个交叉, 这与为保障同源染色体后期的精准分离至少需要形成1个交叉相一致(Jones and Franklin, 2006; Mercier et al., 2015).在大部分生物中, 交叉的形成和分布受到交叉干涉的影响, 即一个交叉的产生会抑制其沿染色体两侧一定范围内另一个交叉的形成, 导致交叉在染色体上呈非随机分布(Berchowitz and Copenhaver, 2010).根据对交叉干扰是否敏感, 可分为干涉敏感型(I型)和干涉不敏感型(II型)交叉(Hatkevich et al., 2017).近年来, 利用正向遗传学和全基因组测序已在拟南芥(Arabidopsis thaliana)中鉴定出3种减数分裂重组抑制途径: (1) FANCM与其DNA结合辅助因子MHF1/2的解旋酶途径(Crismani et al., 2012; Girard et al., 2014); (2) RECQ4A/B、TOP3α和RMI1形成的BTR重组抑制途径(Séguéla-Arnaud et al., 2015, 2017); (3) FIGL1- FLIP复合体调节单链入侵抑制II型交叉形成途径(Girard et al., 2015; Fernandes et al., 2017; Hu et al., 2017).这3种减数分裂重组抑制途径平行共存, 且通过不同机制限制II型交叉形成(Mieulet et al., 2018).然而, 在大多数真核生物减数分裂过程中, II型交叉仅占交叉总数的10%左右, 且依赖于MUS81和FANCD2调控途径(Berchowitz et al., 2007; Higgins et al., 2008; Kurzbauer et al., 2018), 绝大多数交叉主要通过I型交叉形成途径产生(Macaisne et al., 2008, 2011; Wang et al., 2012; Lu et al., 2014; Huang et al., 2015).更重要的是, 同时关闭I型和II型交叉形成途径后, 在拟南芥msh4/mus81/ fancd2突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
Multiple mechanisms limit meiotic crossovers: TOP3α and two BLM homologs antagonize crossovers in parallel to FANCM 1 2015
... 最近, 44个不同物种的基因组研究表明, 尽管不同物种基因组大小存在极大差异, 但重组率波动幅度并不大, 这表明减数分裂重组率受到严格的约束和限制(Fernandes et al., 2018).在大多数物种中, 78%的染色体重组率低于1.5 cM, 而其中绝大多数染色体的重组率仅为0.5 cM, 这意味着每个配对同源染色体只形成了1个交叉, 这与为保障同源染色体后期的精准分离至少需要形成1个交叉相一致(Jones and Franklin, 2006; Mercier et al., 2015).在大部分生物中, 交叉的形成和分布受到交叉干涉的影响, 即一个交叉的产生会抑制其沿染色体两侧一定范围内另一个交叉的形成, 导致交叉在染色体上呈非随机分布(Berchowitz and Copenhaver, 2010).根据对交叉干扰是否敏感, 可分为干涉敏感型(I型)和干涉不敏感型(II型)交叉(Hatkevich et al., 2017).近年来, 利用正向遗传学和全基因组测序已在拟南芥(Arabidopsis thaliana)中鉴定出3种减数分裂重组抑制途径: (1) FANCM与其DNA结合辅助因子MHF1/2的解旋酶途径(Crismani et al., 2012; Girard et al., 2014); (2) RECQ4A/B、TOP3α和RMI1形成的BTR重组抑制途径(Séguéla-Arnaud et al., 2015, 2017); (3) FIGL1- FLIP复合体调节单链入侵抑制II型交叉形成途径(Girard et al., 2015; Fernandes et al., 2017; Hu et al., 2017).这3种减数分裂重组抑制途径平行共存, 且通过不同机制限制II型交叉形成(Mieulet et al., 2018).然而, 在大多数真核生物减数分裂过程中, II型交叉仅占交叉总数的10%左右, 且依赖于MUS81和FANCD2调控途径(Berchowitz et al., 2007; Higgins et al., 2008; Kurzbauer et al., 2018), 绝大多数交叉主要通过I型交叉形成途径产生(Macaisne et al., 2008, 2011; Wang et al., 2012; Lu et al., 2014; Huang et al., 2015).更重要的是, 同时关闭I型和II型交叉形成途径后, 在拟南芥msh4/mus81/ fancd2突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
The DNA replication factor RFC1 is required for interference-sensitive meiotic crossovers in Arabidopsis thaliana 1 2012
... 最近, 44个不同物种的基因组研究表明, 尽管不同物种基因组大小存在极大差异, 但重组率波动幅度并不大, 这表明减数分裂重组率受到严格的约束和限制(Fernandes et al., 2018).在大多数物种中, 78%的染色体重组率低于1.5 cM, 而其中绝大多数染色体的重组率仅为0.5 cM, 这意味着每个配对同源染色体只形成了1个交叉, 这与为保障同源染色体后期的精准分离至少需要形成1个交叉相一致(Jones and Franklin, 2006; Mercier et al., 2015).在大部分生物中, 交叉的形成和分布受到交叉干涉的影响, 即一个交叉的产生会抑制其沿染色体两侧一定范围内另一个交叉的形成, 导致交叉在染色体上呈非随机分布(Berchowitz and Copenhaver, 2010).根据对交叉干扰是否敏感, 可分为干涉敏感型(I型)和干涉不敏感型(II型)交叉(Hatkevich et al., 2017).近年来, 利用正向遗传学和全基因组测序已在拟南芥(Arabidopsis thaliana)中鉴定出3种减数分裂重组抑制途径: (1) FANCM与其DNA结合辅助因子MHF1/2的解旋酶途径(Crismani et al., 2012; Girard et al., 2014); (2) RECQ4A/B、TOP3α和RMI1形成的BTR重组抑制途径(Séguéla-Arnaud et al., 2015, 2017); (3) FIGL1- FLIP复合体调节单链入侵抑制II型交叉形成途径(Girard et al., 2015; Fernandes et al., 2017; Hu et al., 2017).这3种减数分裂重组抑制途径平行共存, 且通过不同机制限制II型交叉形成(Mieulet et al., 2018).然而, 在大多数真核生物减数分裂过程中, II型交叉仅占交叉总数的10%左右, 且依赖于MUS81和FANCD2调控途径(Berchowitz et al., 2007; Higgins et al., 2008; Kurzbauer et al., 2018), 绝大多数交叉主要通过I型交叉形成途径产生(Macaisne et al., 2008, 2011; Wang et al., 2012; Lu et al., 2014; Huang et al., 2015).更重要的是, 同时关闭I型和II型交叉形成途径后, 在拟南芥msh4/mus81/ fancd2突变体中仍然可观察到交叉形成, 这表明还存在其它交叉形成途径(III型) (Higgins et al., 2008; Kurzbauer et al., 2018).但是, 目前抑制I型和III型交叉形成的基因及其调控途径尚不清楚, 而这些重组调控途径的分子机制对植物育种至关重要. ...
DNA methylation epigenetically silences crossover hot spots and controls chromosomal domains of meiotic recombination in Arabidopsis 1 2015
... 减数分裂重组起始于减数分裂初期产生的大量DNA双链断裂, 而只有极少数的断裂双链被修复形成交叉, 其余的大量断裂双链通过不同的途径和机制修复形成非交叉(Giraut et al., 2011; Lu et al., 2012; Qi et al., 2014).在模式植物拟南芥中, 细胞学分析表明每个减数分裂的细胞大约形成200个双链断裂, 但只有约10个断裂双链被修复形成交叉, 其余的断裂双链则被修复产生非交叉(Yelina et al., 2015).近年来, 利用拟南芥zmm突变体短角果表型(因缺乏I型交叉形成的ZMM基因而育性降低)进行果荚表型恢复系的正向遗传学筛选, 已经获得了多个重组率提高的突变体, 通过全基因组测序鉴定出多个II型交叉重组抑制基因, 如FANCM、RECQ4和FIGL1.然而, 由于该突变体筛选是建立在zmm突变体的基础之上, 不能揭示调控绝大多数交叉(I型交叉)形成的重组抑制基因.因此, 我们利用拟南芥花粉荧光标记系作为重组率增加的报告因子, 在拟南芥野生型(WT)遗传背景下进行EMS突变体的正向遗传学筛选, 共获得了18个重组率显著提高(3倍以上)的重组抑制突变体, 其中包括显性和隐性遗传突变.通常EMS诱变产生较多基因功能缺失的隐性突变体和少量功能获得型的显性突变体(Kim et al., 2006), 这与我们的筛选结果一致, 表明利用拟南芥花粉荧光标记系进行重组抑制突变体筛选是成功的.通过全基因组测序, 我们在隐性突变体中鉴定出1个包含FANCM-G540D突变位点的重组抑制突变体rrs1, 该突变体与fancm-1突变体具有相似的重组提高能力, 表明rrs1突变体可能是由FANCM-G540D突变造成, 这也进一步证明利用拟南芥花粉荧光标记系进行重组抑制突变体筛选可行. ...
High-throughput analysis of meiotic crossover frequency and interference via flow cytometry of fluorescent pollen in Arabidopsis thaliana 2 2013
... 拟南芥花粉荧光标记系统(fluorescent tagged lines, FTLs)能高通量快速检测染色体特定标记区间的重组率(Yelina et al., 2013), 为高通量筛选减数分裂重组抑制突变体提供了可能.拟南芥花粉荧光标记系统是由一系列T-DNA插入片段编码的荧光标记构成, 由花粉特异性LAT52启动子激活, 在花粉中表达红荧光(DsRed)和绿荧光(eYFP), 通过统计花粉荧光标记的分离模式可以测量标记区间的重组率(Francis et al., 2007; Berchowitz and Copenhaver, 2008).本研究采用正向遗传学方法, 创新性地利用拟南芥花粉荧光标记系统和流式细胞技术对拟南芥野生型Col遗传背景下的EMS突变体进行大规模重组率检测和筛选, 进而获得重组率提高的突变体, 旨在为植物减数分裂重组抑制基因及其调控途径研究奠定基础. ...
 ,*云南省农业科学院花卉研究所, 国家观赏园艺工程技术研究中心, 云南省花卉育种重点实验室, 昆明 650200
,*云南省农业科学院花卉研究所, 国家观赏园艺工程技术研究中心, 云南省花卉育种重点实验室, 昆明 650200
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT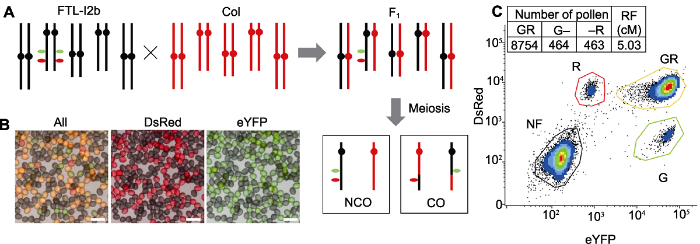 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT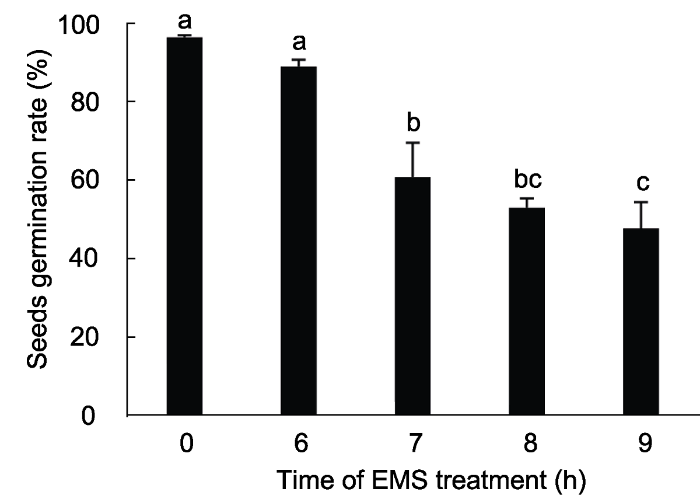 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT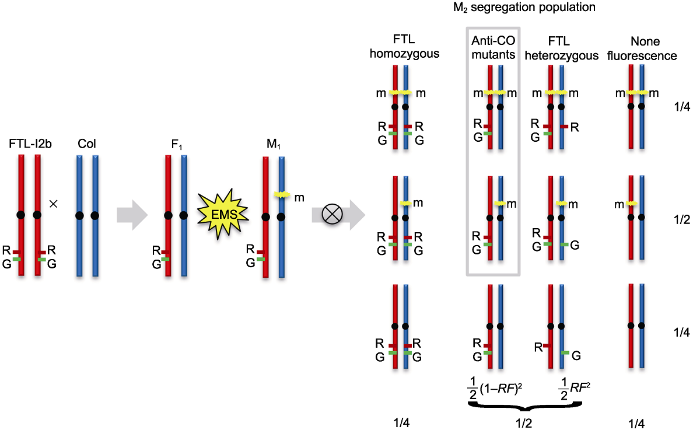 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}