Use of DNA Barcoding in Identifying Tropical Trees from Dipterocarpaceae
Jianlin Hu1,2, Zhifang Liu1,2, Xiuqin Ci1, Jie Li1,*1. Laboratory of Plant Phylogenetics and Conservation, Center for Integrative Conservation, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Kunming 650223, China 2. University of Chinese Academy of Sciences, Beijing 100049, China
摘要 龙脑香科植物是东南亚地区重要的热带木材来源树种, 对其开展DNA条形码评估在林业监管及森林资源保护等方面具有非常重要的实际应用价值。通过对龙脑香科植物样品进行rbcL、matK、trnL-trnF和ITS2四个片段的扩增和测序, 结合GenBank下载的数据, 共获得龙脑香科树种14属244种共计899条序列。通过比较4个片段的通用性、序列特征、种内和种间的遗传变异, 基于Best Match (BM)、Best Close Match (BCM)、相似性搜索算法(BLAST)和邻接树(NJ) 4种方法评估DNA条形码对于龙脑香科树种的鉴定能力。结果表明, ITS2在龙脑香科树种中鉴定效率最高, 通过优化的扩增体系能够从该科植物叶片中获得较高质量的ITS2片段; 叶绿体matK片段扩增和测序效率为100%, 且种内及种间遗传变异明显, 鉴定成功率高于其它叶绿体片段, 并据此提出ITS2和matK适合作为龙脑香科树种的DNA条形码。 关键词:龙脑香科;DNA条形码;ITS2; matK;热带地区
Abstract Plants from Dipterocarpaceae are vital tropical timber trees in Southeast Asia. They have high potential in forest supervision, forest resource protection and other aspects to evaluate the efficiency of DNA barcodes. We obtained 899 sequences from 244 species for 14 genera combined with sequences from the GenBank. We amplified and sequenced four DNA regions (rbcL, matK, trnL-trnF and ITS2) in samples from the family Dipterocarpaceae. The discrimination ability of DNA barcodes was evaluated by using effective sequence ratios, characteristics of sequences, divergence of intra- and inter-varieties, and identification success rates (Best Match, Best Close Match, BLAST and neighbor-joining methods). The ITS2 fragment showed the highest identification efficiency. High-quality ITS2 sequence can be obtained from the leaves of Dipterocarpaceae plants by using an optimized amplification system. The matK fragment showed obvious genetic variation and 100% amplification and sequencing efficiency, and its identification ability was higher than with other chloroplast markers. Therefore, ITS2 and matK are suitable markers for tropical trees from Dipterocarpaceae. Keywords:Dipterocarpaceae;DNA barcoding;ITS2; matK;tropical region
PDF (1185KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 胡建霖, 刘志芳, 慈秀芹, 李捷. DNA条形码在热带龙脑香科树种鉴定中的应用. 植物学报, 2019, 54(3): 350-359 doi:10.11983/CBB18205 Hu Jianlin, Liu Zhifang, Ci Xiuqin, Li Jie. Use of DNA Barcoding in Identifying Tropical Trees from Dipterocarpaceae. Chinese Bulletin of Botany, 2019, 54(3): 350-359 doi:10.11983/CBB18205
DNA条形码技术首先由Paul Hebert提出, 是利用1个或少数几个DNA片段对地球上所有物种进行识别和鉴定(Hebert et al., 2003; Kress et al., 2005)。自DNA条形码技术的概念被提出以来, 研究者制订了DNA条形码的选择标准: (1) 尽量短的片段, 片段两端连接相对保守的区域, 易于开发通用引物; (2) 要有足够的变异可将物种区分开来(CBOL Plant Working Group et al., 2009; 任保青和陈之端, 2010)。利用以上标准, 研究者陆续提出将matK、rbcL、trnH-psbA和ITS/ITS2作为陆地植物的候选DNA条形码(Kress et al., 2005; China Plant BOL Group et al., 2011)。近些年, DNA条形码技术的发展十分迅速, 已经应用于一些热带珍贵植物树种的鉴定。例如, Hassold等(2016)对马达加斯加的红木类树种(red wood)进行DNA条形码研究, 评估3个DNA条形码片段(matK、rbcL和trnL)对于不同产地及不同种类红木的鉴定效率; Jiao等(2018)利用6种紫檀属(Pterocarpus Jacq.)木材标本和干燥叶片样品材料, 评估ITS2、matK、ndhF-rpl32和rbcL四个DNA条形码鉴定效果, 并构建6种紫檀属DNA条形码数据库。在龙脑香科树种中也开展了不少相关研究。例如, Tsumura等(2011)通过比较trnL、trnL-trnF、trnH-trnK和psbC-trnS片段构建了84种娑罗双属(Shorea Roxb.)植物分子数据库; Trang等(2015)评估rbcL、matK和trnH-psbA片段在东南亚地区4种坡垒属(Hopea Roxb.)近缘树种中的鉴定效果, 发现matK鉴定效果优于rbcL和trnH-psbA。目前, 龙脑香科植物的DNA条形码研究仅见于对娑罗双属和坡垒属等少数类群, 而整个龙脑香科的条形码尚未形成统一的标准(Kamiya et al., 2005; Tsumura et al., 2011; Trang et al., 2015)。
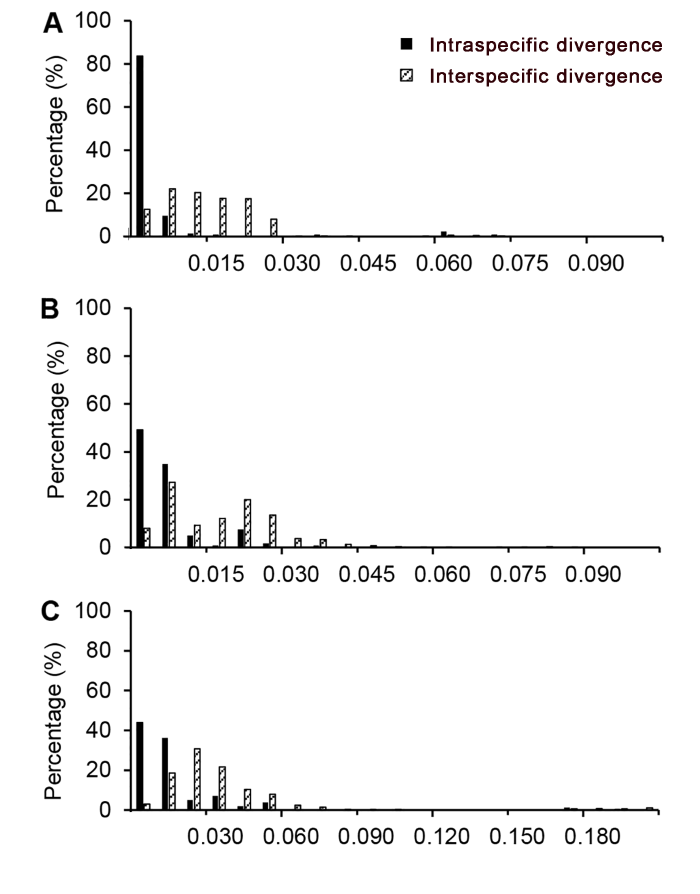
计算结果见表3。叶绿体片段rbcL、matK和trnL-trnF均获得100%的扩增成功率和测序成功率。通过采用引物ITS2F/ITS3R (Chen et al., 2010)进行扩增, ITS2的扩增成功率为100%, 而有效序列获得率仅为47.4%; 通过调整引物ITS3/26SE (White et al., 1990; Sun et al., 1994)并优化扩增体系, ITS2的有效序列获得率提高到89.5%。19个龙脑香科植物样品材料的扩增和测序结果表明, 叶绿体片段的有效序列获得率明显高于ITS2, 但采用2对ITS2引物进行扩增能够获得较高质量的ITS2序列。
Table 3 表3 表3条形码扩增和测序成功率 Table 3The success rate of PCR amplification and sequencing
Table 5 表5 表5叶绿体条形码片段在龙脑香科物种种内变异Wilcoxon检验 Table 5Wilcoxon signed rank tests of intraspecific divergence among chloroplast markers in the species of Dipterocarpaceae
Table 6 表6 表6叶绿体条形码片段在龙脑香科物种种间变异Wilcoxon检验 Table 6Wilcoxon signed rank tests of interspecific divergence among chloroplast markers in the species of Dipterocarpaceae
Figure 2Comparison of the identification efficiency of barcodes base on four methods
2.4 讨论
2.4.1 龙脑香科树种DNA条形码综合评价
理想的DNA条形码需具备片段较短、种间变异较大且鉴定效率高等特点(Kress et al., 2005; Taberlet et al., 2007; 任保青和陈之端, 2010)。本研究中, 我们发现ITS2片段在龙脑香科植物树种中呈现较高的扩增率和测序成功率(分别为100%和89.5%), 且基于BM、BCM、BLAST和NJ树4种条形码评估方法均获得最高的鉴定成功率, 因此我们认为ITS2是龙脑香科树种首选的DNA条形码。能否成功获得高质量条形码序列决定着条形码技术可否应用到实际物种鉴定工作中, 因此条形码扩增率和测序成功率是条形码筛选时的重要指标。ITS2是ITS序列的1个片段, 位于5.8S和26S rRNA之间, 其序列长度较短, 易于扩增和测序, 且其二级结构能够提供更多的信息(Schultz and Wolf, 2009; China Plant BOL Group et al., 2011)。Chen等(2010)通过大规模的取样分析, 认为可将ITS2作为药用植物鉴定的核心条形码; 同时, ITS2在山葡萄(Vitis amurensis)、兰科及红景天属等植物中也显示较好的鉴定效果(宋慧芳等, 2017; De Boer et al., 2017; Zhu et al., 2017)。本研究采用引物ITS3/26SE并优化扩增体系, 通过在扩增体系中添加DMSO及BSA提高PCR的产物特异性和产量。此外, 根据ITS2具有较高的GC/AT含量, 使用高保真DNA聚合酶和GC Enhancer优化PCR过程, 提高了ITS2在龙脑香科树种的获得率, 我们建议将20 μL体系作为龙脑香科植物树种ITS2片段扩增的标准体系。
以往的研究表明, 叶绿体基因组进化速率相对较慢, 能够提供鉴定分析的变异位点较少(Newmaster et al., 2006; Chase et al., 2007; Kress and Erickson, 2007)。根据Wilcoxon Signed Rank检验结果, matK和trnL-trnF片段的种间遗传变异较高, 且差异显著, 显示出较高的应用潜力。matK基因序列是植物叶绿体基因中进化较快的编码基因, 基于样品序列matK片段在龙脑香科树种中进行物种鉴定的成功率仅次于ITS2片段, 表明matK用于龙脑香科树种鉴定效果较好。叶绿体基因间隔区trnL-trnF常用于龙脑香科树种的分子系统学研究, 在GenBank数据库中有较多的数据, 但在本研究中基于4种评估方法获得的鉴定成功率较低。rbcL片段是DNA条形码的核心之一, 由于其引物通用性较高而被广泛使用, 但因变异较小不适合用于种级水平的物种鉴定(Kress and Erickson, 2007; Lahaye et al., 2008)。本研究结果显示, 叶绿体rbcL片段种内种间遗传变异不明显, 且鉴定成功率低于其它片段, 因此不适合作为热带龙脑香科树种的DNA条形码。综上, 我们建议将ITS2和matK片段作为热带龙脑香科树种的DNA条形码。
AshtonPS ( 1982). Flora Malesiana. Series I—Spermatophyta. Flowering Plants, Vol. 9, Part 2. Dipterocarpaceae. The Netherlands: Martinun Nijhoff Pub. [本文引用: 1]
CBOL Plant WorkingGroup, HollingsworthPM, ForrestLL, SpougeJL, HajibabaeiM, RatnasinghamS, van der BankM, ChaseMW, CowanRS, EricksonDL, FazekasAJ, GrahamSW, JamesKE, KimKJ, KressWJ, SchneiderH, van AlphenStahlJ, BarrettSCH, van den BergC, BogarinD, BurgessKS, CameronKM, CarineM, ChacónJ, ClarkA, ClarksonJJ, ConradF, DeveyDS, FordCS, HeddersonTAJ, HollingsworthML, HusbandBC, KellyLJ, KesanakurtiPR, KimJS, KimYD, LahayeR, LeeHL, LongDG, Madri?ánS, MaurinO, MeusnierI, NewmasterSG, ParkCW, PercyDM, PetersenG, RichardsonJE, SalazarGA, SavolainenV, SebergO, WilkinsonMJ, YiDK, LittleDP ( 2009). A DNA barcode for land plants Proc Natl Acad Sci USA 106, 12794-12797. [本文引用: 1]
ChaseMW, CowanRS, HollingsworthPM, van den BergC, Madri?ánS, PetersenG, SebergO, J?rgsensenT, CameronKM, CarineM, PedersenN, HeddersonTAJ, ConradF, SalazarGA, RichardsonJE, HollingsworthML, BarracloughTG, KellyL, WilkinsonM ( 2007). A proposal for a standardised protocol to barcode all land plants Taxon 56, 295-299. [本文引用: 1]
ChenSL, YaoH, HanJP, LiuC, SongJY, ShiLC, ZhuYJ, MaXY, GaoT, PangXH, LuoK, LiY, LiXW, JiaXC, LinYL, LeonC ( 2010). Validation of the ITS2 region as a novel DNA barcode for identifying medicinal plant species. PLoS One 5, e8613. [本文引用: 2]
China Plant BOLGroup, LiDZ, GaoLM, LiHT, WangH, GeXJ, LiuJQ, ChenZD, ZhouSL, ChenSL, YangJB, FuCX, ZengCX, YanHF, ZhuYJ, SunYS, ChenSY, ZhaoL, WangK, YangT, DuanGW ( 2011). Comparative analysis of a large dataset indicates that internal transcribed spacer (ITS) should be incorporated into the core barcode for seed plants Proc Natl Acad Sci USA 108, 19641-19646. [本文引用: 2]
CuénoudP, SavolainenV, ChatrouLW, PowellM, GrayerRJ, ChaseMW ( 2002). Molecular phylogenetics of Caryo- phyllales based on nuclear 18S rDNA and plastid rbcL, atpB, and matK DNA sequences. Am J Bot 89, 132-144.
de BoerHJ, GhorbaniA, ManzanillaV, RaclariuAC, KreziouA, OunjaiS, OsathanunkulM, GravendeelB ( 2017). DNA metabarcoding of orchid-derived products reveals widespread illegal orchid trade Proc Roy Soc B 284, 20171182.
GassonP, MillerR, StekelDJ, WhinderF, ZiemińskaK ( 2010). Wood identification of Dalbergia nigra( CITES Appendix I) using quantitative wood anatomy, principal components analysis and na?ve Bayes classification. Ann Bot 105, 45-56. [本文引用: 1]
GonzalezMA, BaralotoC, EngelJ, MoriSA, PétronelliP, RiéraB, RogerA, ThébaudC, ChaveJ ( 2009). Iden- tification of Amazonian trees with DNA barcodes PLoS One 4, e7483.
HassoldS, Lowry IIPP, BauertMR, RazafintsalamaA, RamamonjisoaL, WidmerA ( 2016). DNA barcoding of Malagasy rosewoods: towards a molecular identification of CITES-listed Dalbergia species. PLoS One 11, e0157881.
HebertPDN, CywinskaA, BallSL, deWaardJR ( 2003). Biological identifications through DNA barcodes Proc Roy Sci B 270, 313-321. [本文引用: 2]
HeckenhauerJ, SamuelR, AshtonPS, TurnerB, BarfussMHJ, JangTS, TemschEM, MccannJ, SalimKA, AttanayakeAMAS, ChaseMW ( 2017). Phylogenetic analyses of plastid DNA suggest a different interpretation of morphological evolution than those used as the basis for previous classifications of Dipterocarpaceae (Malvales) Bot J Linn Soc 185, 1-26. [本文引用: 1]
HollingsworthPM, LiDZ, van der BankM, TwyfordAD ( 2016). Telling plant species apart with DNA: from barcodes to genomes Philos Trans Roy Soc B 371, 2015-0338. [本文引用: 1]
HuangXC, CiXQ, ConranJG, LiJ ( 2015). Application of DNA barcodes in Asian tropical trees—a case study from Xishuangbanna Nature Reserve, Southwest China PLoS One 10, e0129295. [本文引用: 1]
JiaoLC, YuM, WiedenhoeftAC, HeT, LiJN, LiuB, JiangXM, YinYF ( 2018). DNA barcode authentication and library development for the wood of six commercial Pterocarpus species: the critical role of xylarium specimens. Sci Rep 8, 1945. [本文引用: 1]
KamiyaK, HaradaK, TachidaH, AshtonPS ( 2005). Phylogeny of PgiC gene in Shorea and its closely related genera (Dipterocarpaceae), the dominant trees in southeast Asian tropical rain forests. Am J Bot 92, 775-788. [本文引用: 1]
KressWJ, EricksonDL ( 2007). A two-locus global DNA barcode for land plants: the coding rbcL gene complements the non-coding trnH-psbA spacer region. PLoS One 2, e508. [本文引用: 2]
KressWJ, WurdackKJ, ZimmerEA, WeigtLA, JanzenDH ( 2005). Use of DNA barcodes to identify flowering plants Proc Natl Acad Sci USA 102, 8369-8374. [本文引用: 3]
LahayeR, van der BankM, BogarinD, WarnerJ, PupulinF, GigotG, MaurinO, DuthoitS, BarracloughTG, SavolainenV ( 2008). DNA barcoding the floras of biodiversity hotspots Proc Natl Acad Sci USA 105, 2923-2928. [本文引用: 2]
MeierR, ShiyangK, VaidyaG, NgPKL ( 2006). DNA barcoding and taxonomy in Diptera: a tale of high intraspecific variability and low identification success Syst Biol 55, 715-728. [本文引用: 1]
NewmasterSG, FazekasAJ, RagupathyS ( 2006). DNA barcoding in land plants: evaluation of rbcL in a multigene tiered approach. Can J Bot 84, 335-341. [本文引用: 1]
NgCH, LeeSL, TnahLH, NgKKS, LeeCT, MariaM ( 2016). Genome size variation and evolution in Dipterocarpaceae Plant Ecol Divers 9, 437-446.
OlmsteadRG, MichaelsHJ, ScottKM, PalmerJD ( 1992). Monophyly of the Asteridae and identification of their major lineages inferred from DNA sequences of rbcL. Ann Mo Bot Gard 79, 249-265.
SchultzJ, WolfM ( 2009). ITS2 sequence-structure analysis in phylogenetics: a how-to manual for molecular syste- matics Mol Phylogenet Evol 52, 520-523. [本文引用: 1]
SunY, SkinnerDZ, LiangGH, HulbertSH ( 1994). Phylogenetic analysis of Sorghum and related taxa using internal transcribed spacers of nuclear ribosomal DNA. Theor Appl Genet 89, 26-32. [本文引用: 1]
TaberletP, CoissacE, PompanonF, GiellyL, MiquelC, ValentiniA, VermatT, CorthierG, BrochmannC, WillerslevE ( 2007). Power and limitations of the chloroplast trnL( UAA) intron for plant DNA barcoding. Nucleic Acids Res 35, e14. [本文引用: 1]
TaberletP, GiellyL, PautouG, BouvetJ ( 1991). Universal primers for amplification of three non-coding regions of chloroplast DNA Plant Mol Biol 17, 1105-1109.
TrangNTP, DucNM, SinhNV, TriestL ( 2015). Application of DNA barcoding markers to the identification of Hopea species. Genet Mol Res 14, 9181-9190. [本文引用: 1]
TsumuraY, KadoT, YoshidaK, AbeH, OhtaniM, TaguchiY, FukueY, TaniN, UenoS, YoshimuraK, KamiyaK, HaradaK, TakeuchiY, DiwayB, FinkeldeyR, Na’iemM, IndriokoS, NgKKS, MuhammadN, LeeSL ( 2011). Molecular database for classifying Shorea species (Dipterocarpaceae) and techniques for checking the legitimacy of timber and wood products. J Plant Res 124, 35-48. [本文引用: 1]
WhiteTJ, BrunsTD, LeeS, TaylorJ ( 1990). Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ, eds. PCR Protocols: a Guide to Methods and Applications. San Diego: Academic Press. pp. 315-322.
WilsonEO ( 2016). Half-Earth: Our Planet’s Fight for Life. New York: Liveright Publishing Corporation.
YuM, JiaoLC, GuoJ, WiedenhoeftAC, HeT, JiangXM, YinYF ( 2017). DNA barcoding of vouchered xylarium wood specimens of nine endangered Dalbergia species. Planta 246, 1165-1176. [本文引用: 1]
ZengCX, HollingsworthPM, YangJ, HeZS, ZhangZR, LiDZ, YangJB ( 2018). Genome skimming herbarium spe- cimens for DNA barcoding and phylogenomics Plant Methods 14, 43. [本文引用: 1]
ZhuRW, LiYC, ZhongDL, ZhangJQ ( 2017). Establishment of the most comprehensive ITS2 barcode database to date of the traditional medicinal plant Rhodiola ( Crassulaceae). Sci Rep 7, 10051. [本文引用: 1]
... 随着世界各国对森林资源保护的重视, 使用合法认证的木材和木制品已成为国际共识, 因此对木材种类进行准确鉴定的重要性日趋凸显(Dormontt et al., 2015).基于宏观和微观特征的木材鉴定方法往往依赖于经验丰富的技术专家, 且大多数解剖学特征也只能将木材产品鉴定到属级水平(Gasson et al., 2010; Dormontt et al., 2015).近年来, 植物幼苗和木材等非标准材料DNA提取方法的改进大大提高了条形码技术在木材树种及其产品鉴定中的可行性, 研究者在树木形成层(Huang et al., 2015)、植物幼苗(Gon- zalez et al., 2009)和木材标本(Yu et al., 2017; Jiao et al., 2018; 乔梦吉等, 2018)等样品材料中均成功获得条形码片段, 尤其是Tsumura等(2011)在龙脑香科木材样品中成功扩增到trnL、trnL-trnF、trnH- trnK和psbC-trnS片段.这些研究表明以植物叶片为材料扩增到的植物条形码片段也能用于龙脑香科木材产品的鉴定.本研究以龙脑香科树种的干燥叶片为样品材料, 结合GenBank数据分析评估4种条形码片段在龙脑香科植物鉴定方面的应用潜力, 能够为龙脑香科树种及木材产品的条形码鉴定提供借鉴. ...
植物DNA条形码技术 2 2010
... DNA条形码技术首先由Paul Hebert提出, 是利用1个或少数几个DNA片段对地球上所有物种进行识别和鉴定(Hebert et al., 2003; Kress et al., 2005).自DNA条形码技术的概念被提出以来, 研究者制订了DNA条形码的选择标准: (1) 尽量短的片段, 片段两端连接相对保守的区域, 易于开发通用引物; (2) 要有足够的变异可将物种区分开来(CBOL Plant Working Group et al., 2009; 任保青和陈之端, 2010).利用以上标准, 研究者陆续提出将matK、rbcL、trnH-psbA和ITS/ITS2作为陆地植物的候选DNA条形码(Kress et al., 2005; China Plant BOL Group et al., 2011).近些年, DNA条形码技术的发展十分迅速, 已经应用于一些热带珍贵植物树种的鉴定.例如, Hassold等(2016)对马达加斯加的红木类树种(red wood)进行DNA条形码研究, 评估3个DNA条形码片段(matK、rbcL和trnL)对于不同产地及不同种类红木的鉴定效率; Jiao等(2018)利用6种紫檀属(Pterocarpus Jacq.)木材标本和干燥叶片样品材料, 评估ITS2、matK、ndhF-rpl32和rbcL四个DNA条形码鉴定效果, 并构建6种紫檀属DNA条形码数据库.在龙脑香科树种中也开展了不少相关研究.例如, Tsumura等(2011)通过比较trnL、trnL-trnF、trnH-trnK和psbC-trnS片段构建了84种娑罗双属(Shorea Roxb.)植物分子数据库; Trang等(2015)评估rbcL、matK和trnH-psbA片段在东南亚地区4种坡垒属(Hopea Roxb.)近缘树种中的鉴定效果, 发现matK鉴定效果优于rbcL和trnH-psbA.目前, 龙脑香科植物的DNA条形码研究仅见于对娑罗双属和坡垒属等少数类群, 而整个龙脑香科的条形码尚未形成统一的标准(Kamiya et al., 2005; Tsumura et al., 2011; Trang et al., 2015). ...
... 理想的DNA条形码需具备片段较短、种间变异较大且鉴定效率高等特点(Kress et al., 2005; Taberlet et al., 2007; 任保青和陈之端, 2010).本研究中, 我们发现ITS2片段在龙脑香科植物树种中呈现较高的扩增率和测序成功率(分别为100%和89.5%), 且基于BM、BCM、BLAST和NJ树4种条形码评估方法均获得最高的鉴定成功率, 因此我们认为ITS2是龙脑香科树种首选的DNA条形码.能否成功获得高质量条形码序列决定着条形码技术可否应用到实际物种鉴定工作中, 因此条形码扩增率和测序成功率是条形码筛选时的重要指标.ITS2是ITS序列的1个片段, 位于5.8S和26S rRNA之间, 其序列长度较短, 易于扩增和测序, 且其二级结构能够提供更多的信息(Schultz and Wolf, 2009; China Plant BOL Group et al., 2011).Chen等(2010)通过大规模的取样分析, 认为可将ITS2作为药用植物鉴定的核心条形码; 同时, ITS2在山葡萄(Vitis amurensis)、兰科及红景天属等植物中也显示较好的鉴定效果(宋慧芳等, 2017; De Boer et al., 2017; Zhu et al., 2017).本研究采用引物ITS3/26SE并优化扩增体系, 通过在扩增体系中添加DMSO及BSA提高PCR的产物特异性和产量.此外, 根据ITS2具有较高的GC/AT含量, 使用高保真DNA聚合酶和GC Enhancer优化PCR过程, 提高了ITS2在龙脑香科树种的获得率, 我们建议将20 μL体系作为龙脑香科植物树种ITS2片段扩增的标准体系. ...
山葡萄种质资源DNA条形码通用序列的筛选 1 2017
... 理想的DNA条形码需具备片段较短、种间变异较大且鉴定效率高等特点(Kress et al., 2005; Taberlet et al., 2007; 任保青和陈之端, 2010).本研究中, 我们发现ITS2片段在龙脑香科植物树种中呈现较高的扩增率和测序成功率(分别为100%和89.5%), 且基于BM、BCM、BLAST和NJ树4种条形码评估方法均获得最高的鉴定成功率, 因此我们认为ITS2是龙脑香科树种首选的DNA条形码.能否成功获得高质量条形码序列决定着条形码技术可否应用到实际物种鉴定工作中, 因此条形码扩增率和测序成功率是条形码筛选时的重要指标.ITS2是ITS序列的1个片段, 位于5.8S和26S rRNA之间, 其序列长度较短, 易于扩增和测序, 且其二级结构能够提供更多的信息(Schultz and Wolf, 2009; China Plant BOL Group et al., 2011).Chen等(2010)通过大规模的取样分析, 认为可将ITS2作为药用植物鉴定的核心条形码; 同时, ITS2在山葡萄(Vitis amurensis)、兰科及红景天属等植物中也显示较好的鉴定效果(宋慧芳等, 2017; De Boer et al., 2017; Zhu et al., 2017).本研究采用引物ITS3/26SE并优化扩增体系, 通过在扩增体系中添加DMSO及BSA提高PCR的产物特异性和产量.此外, 根据ITS2具有较高的GC/AT含量, 使用高保真DNA聚合酶和GC Enhancer优化PCR过程, 提高了ITS2在龙脑香科树种的获得率, 我们建议将20 μL体系作为龙脑香科植物树种ITS2片段扩增的标准体系. ...
... DNA条形码技术首先由Paul Hebert提出, 是利用1个或少数几个DNA片段对地球上所有物种进行识别和鉴定(Hebert et al., 2003; Kress et al., 2005).自DNA条形码技术的概念被提出以来, 研究者制订了DNA条形码的选择标准: (1) 尽量短的片段, 片段两端连接相对保守的区域, 易于开发通用引物; (2) 要有足够的变异可将物种区分开来(CBOL Plant Working Group et al., 2009; 任保青和陈之端, 2010).利用以上标准, 研究者陆续提出将matK、rbcL、trnH-psbA和ITS/ITS2作为陆地植物的候选DNA条形码(Kress et al., 2005; China Plant BOL Group et al., 2011).近些年, DNA条形码技术的发展十分迅速, 已经应用于一些热带珍贵植物树种的鉴定.例如, Hassold等(2016)对马达加斯加的红木类树种(red wood)进行DNA条形码研究, 评估3个DNA条形码片段(matK、rbcL和trnL)对于不同产地及不同种类红木的鉴定效率; Jiao等(2018)利用6种紫檀属(Pterocarpus Jacq.)木材标本和干燥叶片样品材料, 评估ITS2、matK、ndhF-rpl32和rbcL四个DNA条形码鉴定效果, 并构建6种紫檀属DNA条形码数据库.在龙脑香科树种中也开展了不少相关研究.例如, Tsumura等(2011)通过比较trnL、trnL-trnF、trnH-trnK和psbC-trnS片段构建了84种娑罗双属(Shorea Roxb.)植物分子数据库; Trang等(2015)评估rbcL、matK和trnH-psbA片段在东南亚地区4种坡垒属(Hopea Roxb.)近缘树种中的鉴定效果, 发现matK鉴定效果优于rbcL和trnH-psbA.目前, 龙脑香科植物的DNA条形码研究仅见于对娑罗双属和坡垒属等少数类群, 而整个龙脑香科的条形码尚未形成统一的标准(Kamiya et al., 2005; Tsumura et al., 2011; Trang et al., 2015). ...
A proposal for a standardised protocol to barcode all land plants 1 2007
... 以往的研究表明, 叶绿体基因组进化速率相对较慢, 能够提供鉴定分析的变异位点较少(Newmaster et al., 2006; Chase et al., 2007; Kress and Erickson, 2007).根据Wilcoxon Signed Rank检验结果, matK和trnL-trnF片段的种间遗传变异较高, 且差异显著, 显示出较高的应用潜力.matK基因序列是植物叶绿体基因中进化较快的编码基因, 基于样品序列matK片段在龙脑香科树种中进行物种鉴定的成功率仅次于ITS2片段, 表明matK用于龙脑香科树种鉴定效果较好.叶绿体基因间隔区trnL-trnF常用于龙脑香科树种的分子系统学研究, 在GenBank数据库中有较多的数据, 但在本研究中基于4种评估方法获得的鉴定成功率较低.rbcL片段是DNA条形码的核心之一, 由于其引物通用性较高而被广泛使用, 但因变异较小不适合用于种级水平的物种鉴定(Kress and Erickson, 2007; Lahaye et al., 2008).本研究结果显示, 叶绿体rbcL片段种内种间遗传变异不明显, 且鉴定成功率低于其它片段, 因此不适合作为热带龙脑香科树种的DNA条形码.综上, 我们建议将ITS2和matK片段作为热带龙脑香科树种的DNA条形码. ...
... 计算结果见表3.叶绿体片段rbcL、matK和trnL-trnF均获得100%的扩增成功率和测序成功率.通过采用引物ITS2F/ITS3R (Chen et al., 2010)进行扩增, ITS2的扩增成功率为100%, 而有效序列获得率仅为47.4%; 通过调整引物ITS3/26SE (White et al., 1990; Sun et al., 1994)并优化扩增体系, ITS2的有效序列获得率提高到89.5%.19个龙脑香科植物样品材料的扩增和测序结果表明, 叶绿体片段的有效序列获得率明显高于ITS2, 但采用2对ITS2引物进行扩增能够获得较高质量的ITS2序列. ...
Comparative analysis of a large dataset indicates that internal transcribed spacer (ITS) should be incorporated into the core barcode for seed plants 2 2011
... DNA条形码技术首先由Paul Hebert提出, 是利用1个或少数几个DNA片段对地球上所有物种进行识别和鉴定(Hebert et al., 2003; Kress et al., 2005).自DNA条形码技术的概念被提出以来, 研究者制订了DNA条形码的选择标准: (1) 尽量短的片段, 片段两端连接相对保守的区域, 易于开发通用引物; (2) 要有足够的变异可将物种区分开来(CBOL Plant Working Group et al., 2009; 任保青和陈之端, 2010).利用以上标准, 研究者陆续提出将matK、rbcL、trnH-psbA和ITS/ITS2作为陆地植物的候选DNA条形码(Kress et al., 2005; China Plant BOL Group et al., 2011).近些年, DNA条形码技术的发展十分迅速, 已经应用于一些热带珍贵植物树种的鉴定.例如, Hassold等(2016)对马达加斯加的红木类树种(red wood)进行DNA条形码研究, 评估3个DNA条形码片段(matK、rbcL和trnL)对于不同产地及不同种类红木的鉴定效率; Jiao等(2018)利用6种紫檀属(Pterocarpus Jacq.)木材标本和干燥叶片样品材料, 评估ITS2、matK、ndhF-rpl32和rbcL四个DNA条形码鉴定效果, 并构建6种紫檀属DNA条形码数据库.在龙脑香科树种中也开展了不少相关研究.例如, Tsumura等(2011)通过比较trnL、trnL-trnF、trnH-trnK和psbC-trnS片段构建了84种娑罗双属(Shorea Roxb.)植物分子数据库; Trang等(2015)评估rbcL、matK和trnH-psbA片段在东南亚地区4种坡垒属(Hopea Roxb.)近缘树种中的鉴定效果, 发现matK鉴定效果优于rbcL和trnH-psbA.目前, 龙脑香科植物的DNA条形码研究仅见于对娑罗双属和坡垒属等少数类群, 而整个龙脑香科的条形码尚未形成统一的标准(Kamiya et al., 2005; Tsumura et al., 2011; Trang et al., 2015). ...
... 理想的DNA条形码需具备片段较短、种间变异较大且鉴定效率高等特点(Kress et al., 2005; Taberlet et al., 2007; 任保青和陈之端, 2010).本研究中, 我们发现ITS2片段在龙脑香科植物树种中呈现较高的扩增率和测序成功率(分别为100%和89.5%), 且基于BM、BCM、BLAST和NJ树4种条形码评估方法均获得最高的鉴定成功率, 因此我们认为ITS2是龙脑香科树种首选的DNA条形码.能否成功获得高质量条形码序列决定着条形码技术可否应用到实际物种鉴定工作中, 因此条形码扩增率和测序成功率是条形码筛选时的重要指标.ITS2是ITS序列的1个片段, 位于5.8S和26S rRNA之间, 其序列长度较短, 易于扩增和测序, 且其二级结构能够提供更多的信息(Schultz and Wolf, 2009; China Plant BOL Group et al., 2011).Chen等(2010)通过大规模的取样分析, 认为可将ITS2作为药用植物鉴定的核心条形码; 同时, ITS2在山葡萄(Vitis amurensis)、兰科及红景天属等植物中也显示较好的鉴定效果(宋慧芳等, 2017; De Boer et al., 2017; Zhu et al., 2017).本研究采用引物ITS3/26SE并优化扩增体系, 通过在扩增体系中添加DMSO及BSA提高PCR的产物特异性和产量.此外, 根据ITS2具有较高的GC/AT含量, 使用高保真DNA聚合酶和GC Enhancer优化PCR过程, 提高了ITS2在龙脑香科树种的获得率, 我们建议将20 μL体系作为龙脑香科植物树种ITS2片段扩增的标准体系. ...
Molecular phylogenetics of Caryo- phyllales based on nuclear 18S rDNA and plastid 0 2002
DNA metabarcoding of orchid-derived products reveals widespread illegal orchid trade 0 2017
Forensic timber identification: it's time to integrate disciplines to combat illegal logging 2 2015
... 随着世界各国对森林资源保护的重视, 使用合法认证的木材和木制品已成为国际共识, 因此对木材种类进行准确鉴定的重要性日趋凸显(Dormontt et al., 2015).基于宏观和微观特征的木材鉴定方法往往依赖于经验丰富的技术专家, 且大多数解剖学特征也只能将木材产品鉴定到属级水平(Gasson et al., 2010; Dormontt et al., 2015).近年来, 植物幼苗和木材等非标准材料DNA提取方法的改进大大提高了条形码技术在木材树种及其产品鉴定中的可行性, 研究者在树木形成层(Huang et al., 2015)、植物幼苗(Gon- zalez et al., 2009)和木材标本(Yu et al., 2017; Jiao et al., 2018; 乔梦吉等, 2018)等样品材料中均成功获得条形码片段, 尤其是Tsumura等(2011)在龙脑香科木材样品中成功扩增到trnL、trnL-trnF、trnH- trnK和psbC-trnS片段.这些研究表明以植物叶片为材料扩增到的植物条形码片段也能用于龙脑香科木材产品的鉴定.本研究以龙脑香科树种的干燥叶片为样品材料, 结合GenBank数据分析评估4种条形码片段在龙脑香科植物鉴定方面的应用潜力, 能够为龙脑香科树种及木材产品的条形码鉴定提供借鉴. ...
... ; Dormontt et al., 2015).近年来, 植物幼苗和木材等非标准材料DNA提取方法的改进大大提高了条形码技术在木材树种及其产品鉴定中的可行性, 研究者在树木形成层(Huang et al., 2015)、植物幼苗(Gon- zalez et al., 2009)和木材标本(Yu et al., 2017; Jiao et al., 2018; 乔梦吉等, 2018)等样品材料中均成功获得条形码片段, 尤其是Tsumura等(2011)在龙脑香科木材样品中成功扩增到trnL、trnL-trnF、trnH- trnK和psbC-trnS片段.这些研究表明以植物叶片为材料扩增到的植物条形码片段也能用于龙脑香科木材产品的鉴定.本研究以龙脑香科树种的干燥叶片为样品材料, 结合GenBank数据分析评估4种条形码片段在龙脑香科植物鉴定方面的应用潜力, 能够为龙脑香科树种及木材产品的条形码鉴定提供借鉴. ...
A rapid DNA isolation procedure for small quantities of fresh leaf tissue 1 1987
... DNA条形码技术首先由Paul Hebert提出, 是利用1个或少数几个DNA片段对地球上所有物种进行识别和鉴定(Hebert et al., 2003; Kress et al., 2005).自DNA条形码技术的概念被提出以来, 研究者制订了DNA条形码的选择标准: (1) 尽量短的片段, 片段两端连接相对保守的区域, 易于开发通用引物; (2) 要有足够的变异可将物种区分开来(CBOL Plant Working Group et al., 2009; 任保青和陈之端, 2010).利用以上标准, 研究者陆续提出将matK、rbcL、trnH-psbA和ITS/ITS2作为陆地植物的候选DNA条形码(Kress et al., 2005; China Plant BOL Group et al., 2011).近些年, DNA条形码技术的发展十分迅速, 已经应用于一些热带珍贵植物树种的鉴定.例如, Hassold等(2016)对马达加斯加的红木类树种(red wood)进行DNA条形码研究, 评估3个DNA条形码片段(matK、rbcL和trnL)对于不同产地及不同种类红木的鉴定效率; Jiao等(2018)利用6种紫檀属(Pterocarpus Jacq.)木材标本和干燥叶片样品材料, 评估ITS2、matK、ndhF-rpl32和rbcL四个DNA条形码鉴定效果, 并构建6种紫檀属DNA条形码数据库.在龙脑香科树种中也开展了不少相关研究.例如, Tsumura等(2011)通过比较trnL、trnL-trnF、trnH-trnK和psbC-trnS片段构建了84种娑罗双属(Shorea Roxb.)植物分子数据库; Trang等(2015)评估rbcL、matK和trnH-psbA片段在东南亚地区4种坡垒属(Hopea Roxb.)近缘树种中的鉴定效果, 发现matK鉴定效果优于rbcL和trnH-psbA.目前, 龙脑香科植物的DNA条形码研究仅见于对娑罗双属和坡垒属等少数类群, 而整个龙脑香科的条形码尚未形成统一的标准(Kamiya et al., 2005; Tsumura et al., 2011; Trang et al., 2015). ...
Phylogenetic analyses of plastid DNA suggest a different interpretation of morphological evolution than those used as the basis for previous classifications of Dipterocarpaceae (Malvales) 1 2017
... 龙脑香科植物是热带雨林中具有重要生态价值和经济意义的树种, 其木材产品在东南亚地区热带木材市场中有重要地位.本研究涉及的条形码序列来自龙脑香科14属244种, 共计899条, 涵盖了龙脑香科树种82.4%的属和46.1%的种, 序列数据大多来源于东南亚地区龙脑香科树种的相关研究, 在较大范围内代表了龙脑香科树种的遗传变异.目前, 龙脑香科植物的物种亲缘关系还未得到很好的解决与澄清, 这可能在一定程度上影响了DNA条形码在此类群中的应用.分子系统学研究结果不支持将龙脑香亚科(Dipterocarpoideae)分为Dipterocarpeae和Shoreeae 2个族, 理由是龙脑香属(Dipterocarpus Gaertn. f.)被认为是冰片香属(Dryobalanops C.F. Gaertn.)和Shoreeae的姐妹类群, 且娑罗双属(Shorea Roxb.)为非单系分支(Heckenhauer et al., 2017; Ng et al., 2017).利用DNA条形码进行物种鉴定需要较为完善的物种系统发育框架, 因此龙脑香科条形码的研究有赖于其系统发育关系树的构建.随着高通量测序技术(next generation sequencing)的发展, DNA条形码技术衍生出新的发展趋势.例如, 基因组浅层测序(genome skimming)通过获得全基因组较低测序深度基因组数据, 借助生物信息学手段可获得叶绿体全基因组以及核基因组中高度重复区域(nrDNA)等序列(Hollingsworth et al., 2016; Zeng et al., 2018).利用高通量测序技术能够获得物种的基因组水平信息, 为快速分化的近缘物种鉴定提供更加有效的解决办法(李德铢和曾春霞, 2015).随着条形码技术的不断成熟, 以及龙脑香科木材树种DNA条形码数据库的不断完善和龙脑香科系统发育关系的日趋明确, 相信DNA条形码技术将成为热带地区龙脑香科木材树种鉴定的有力工具. ...
Telling plant species apart with DNA: from barcodes to genomes 1 2016
... 龙脑香科植物是热带雨林中具有重要生态价值和经济意义的树种, 其木材产品在东南亚地区热带木材市场中有重要地位.本研究涉及的条形码序列来自龙脑香科14属244种, 共计899条, 涵盖了龙脑香科树种82.4%的属和46.1%的种, 序列数据大多来源于东南亚地区龙脑香科树种的相关研究, 在较大范围内代表了龙脑香科树种的遗传变异.目前, 龙脑香科植物的物种亲缘关系还未得到很好的解决与澄清, 这可能在一定程度上影响了DNA条形码在此类群中的应用.分子系统学研究结果不支持将龙脑香亚科(Dipterocarpoideae)分为Dipterocarpeae和Shoreeae 2个族, 理由是龙脑香属(Dipterocarpus Gaertn. f.)被认为是冰片香属(Dryobalanops C.F. Gaertn.)和Shoreeae的姐妹类群, 且娑罗双属(Shorea Roxb.)为非单系分支(Heckenhauer et al., 2017; Ng et al., 2017).利用DNA条形码进行物种鉴定需要较为完善的物种系统发育框架, 因此龙脑香科条形码的研究有赖于其系统发育关系树的构建.随着高通量测序技术(next generation sequencing)的发展, DNA条形码技术衍生出新的发展趋势.例如, 基因组浅层测序(genome skimming)通过获得全基因组较低测序深度基因组数据, 借助生物信息学手段可获得叶绿体全基因组以及核基因组中高度重复区域(nrDNA)等序列(Hollingsworth et al., 2016; Zeng et al., 2018).利用高通量测序技术能够获得物种的基因组水平信息, 为快速分化的近缘物种鉴定提供更加有效的解决办法(李德铢和曾春霞, 2015).随着条形码技术的不断成熟, 以及龙脑香科木材树种DNA条形码数据库的不断完善和龙脑香科系统发育关系的日趋明确, 相信DNA条形码技术将成为热带地区龙脑香科木材树种鉴定的有力工具. ...
Application of DNA barcodes in Asian tropical trees—a case study from Xishuangbanna Nature Reserve, Southwest China 1 2015
... 随着世界各国对森林资源保护的重视, 使用合法认证的木材和木制品已成为国际共识, 因此对木材种类进行准确鉴定的重要性日趋凸显(Dormontt et al., 2015).基于宏观和微观特征的木材鉴定方法往往依赖于经验丰富的技术专家, 且大多数解剖学特征也只能将木材产品鉴定到属级水平(Gasson et al., 2010; Dormontt et al., 2015).近年来, 植物幼苗和木材等非标准材料DNA提取方法的改进大大提高了条形码技术在木材树种及其产品鉴定中的可行性, 研究者在树木形成层(Huang et al., 2015)、植物幼苗(Gon- zalez et al., 2009)和木材标本(Yu et al., 2017; Jiao et al., 2018; 乔梦吉等, 2018)等样品材料中均成功获得条形码片段, 尤其是Tsumura等(2011)在龙脑香科木材样品中成功扩增到trnL、trnL-trnF、trnH- trnK和psbC-trnS片段.这些研究表明以植物叶片为材料扩增到的植物条形码片段也能用于龙脑香科木材产品的鉴定.本研究以龙脑香科树种的干燥叶片为样品材料, 结合GenBank数据分析评估4种条形码片段在龙脑香科植物鉴定方面的应用潜力, 能够为龙脑香科树种及木材产品的条形码鉴定提供借鉴. ...
DNA barcode authentication and library development for the wood of six commercial 1 2018
... 随着世界各国对森林资源保护的重视, 使用合法认证的木材和木制品已成为国际共识, 因此对木材种类进行准确鉴定的重要性日趋凸显(Dormontt et al., 2015).基于宏观和微观特征的木材鉴定方法往往依赖于经验丰富的技术专家, 且大多数解剖学特征也只能将木材产品鉴定到属级水平(Gasson et al., 2010; Dormontt et al., 2015).近年来, 植物幼苗和木材等非标准材料DNA提取方法的改进大大提高了条形码技术在木材树种及其产品鉴定中的可行性, 研究者在树木形成层(Huang et al., 2015)、植物幼苗(Gon- zalez et al., 2009)和木材标本(Yu et al., 2017; Jiao et al., 2018; 乔梦吉等, 2018)等样品材料中均成功获得条形码片段, 尤其是Tsumura等(2011)在龙脑香科木材样品中成功扩增到trnL、trnL-trnF、trnH- trnK和psbC-trnS片段.这些研究表明以植物叶片为材料扩增到的植物条形码片段也能用于龙脑香科木材产品的鉴定.本研究以龙脑香科树种的干燥叶片为样品材料, 结合GenBank数据分析评估4种条形码片段在龙脑香科植物鉴定方面的应用潜力, 能够为龙脑香科树种及木材产品的条形码鉴定提供借鉴. ...
Phylogeny of 1 2005
... DNA条形码技术首先由Paul Hebert提出, 是利用1个或少数几个DNA片段对地球上所有物种进行识别和鉴定(Hebert et al., 2003; Kress et al., 2005).自DNA条形码技术的概念被提出以来, 研究者制订了DNA条形码的选择标准: (1) 尽量短的片段, 片段两端连接相对保守的区域, 易于开发通用引物; (2) 要有足够的变异可将物种区分开来(CBOL Plant Working Group et al., 2009; 任保青和陈之端, 2010).利用以上标准, 研究者陆续提出将matK、rbcL、trnH-psbA和ITS/ITS2作为陆地植物的候选DNA条形码(Kress et al., 2005; China Plant BOL Group et al., 2011).近些年, DNA条形码技术的发展十分迅速, 已经应用于一些热带珍贵植物树种的鉴定.例如, Hassold等(2016)对马达加斯加的红木类树种(red wood)进行DNA条形码研究, 评估3个DNA条形码片段(matK、rbcL和trnL)对于不同产地及不同种类红木的鉴定效率; Jiao等(2018)利用6种紫檀属(Pterocarpus Jacq.)木材标本和干燥叶片样品材料, 评估ITS2、matK、ndhF-rpl32和rbcL四个DNA条形码鉴定效果, 并构建6种紫檀属DNA条形码数据库.在龙脑香科树种中也开展了不少相关研究.例如, Tsumura等(2011)通过比较trnL、trnL-trnF、trnH-trnK和psbC-trnS片段构建了84种娑罗双属(Shorea Roxb.)植物分子数据库; Trang等(2015)评估rbcL、matK和trnH-psbA片段在东南亚地区4种坡垒属(Hopea Roxb.)近缘树种中的鉴定效果, 发现matK鉴定效果优于rbcL和trnH-psbA.目前, 龙脑香科植物的DNA条形码研究仅见于对娑罗双属和坡垒属等少数类群, 而整个龙脑香科的条形码尚未形成统一的标准(Kamiya et al., 2005; Tsumura et al., 2011; Trang et al., 2015). ...
A two-locus global DNA barcode for land plants: the coding 2 2007
... 以往的研究表明, 叶绿体基因组进化速率相对较慢, 能够提供鉴定分析的变异位点较少(Newmaster et al., 2006; Chase et al., 2007; Kress and Erickson, 2007).根据Wilcoxon Signed Rank检验结果, matK和trnL-trnF片段的种间遗传变异较高, 且差异显著, 显示出较高的应用潜力.matK基因序列是植物叶绿体基因中进化较快的编码基因, 基于样品序列matK片段在龙脑香科树种中进行物种鉴定的成功率仅次于ITS2片段, 表明matK用于龙脑香科树种鉴定效果较好.叶绿体基因间隔区trnL-trnF常用于龙脑香科树种的分子系统学研究, 在GenBank数据库中有较多的数据, 但在本研究中基于4种评估方法获得的鉴定成功率较低.rbcL片段是DNA条形码的核心之一, 由于其引物通用性较高而被广泛使用, 但因变异较小不适合用于种级水平的物种鉴定(Kress and Erickson, 2007; Lahaye et al., 2008).本研究结果显示, 叶绿体rbcL片段种内种间遗传变异不明显, 且鉴定成功率低于其它片段, 因此不适合作为热带龙脑香科树种的DNA条形码.综上, 我们建议将ITS2和matK片段作为热带龙脑香科树种的DNA条形码. ...
... 片段是DNA条形码的核心之一, 由于其引物通用性较高而被广泛使用, 但因变异较小不适合用于种级水平的物种鉴定(Kress and Erickson, 2007; Lahaye et al., 2008).本研究结果显示, 叶绿体rbcL片段种内种间遗传变异不明显, 且鉴定成功率低于其它片段, 因此不适合作为热带龙脑香科树种的DNA条形码.综上, 我们建议将ITS2和matK片段作为热带龙脑香科树种的DNA条形码. ...
Use of DNA barcodes to identify flowering plants 3 2005
... DNA条形码技术首先由Paul Hebert提出, 是利用1个或少数几个DNA片段对地球上所有物种进行识别和鉴定(Hebert et al., 2003; Kress et al., 2005).自DNA条形码技术的概念被提出以来, 研究者制订了DNA条形码的选择标准: (1) 尽量短的片段, 片段两端连接相对保守的区域, 易于开发通用引物; (2) 要有足够的变异可将物种区分开来(CBOL Plant Working Group et al., 2009; 任保青和陈之端, 2010).利用以上标准, 研究者陆续提出将matK、rbcL、trnH-psbA和ITS/ITS2作为陆地植物的候选DNA条形码(Kress et al., 2005; China Plant BOL Group et al., 2011).近些年, DNA条形码技术的发展十分迅速, 已经应用于一些热带珍贵植物树种的鉴定.例如, Hassold等(2016)对马达加斯加的红木类树种(red wood)进行DNA条形码研究, 评估3个DNA条形码片段(matK、rbcL和trnL)对于不同产地及不同种类红木的鉴定效率; Jiao等(2018)利用6种紫檀属(Pterocarpus Jacq.)木材标本和干燥叶片样品材料, 评估ITS2、matK、ndhF-rpl32和rbcL四个DNA条形码鉴定效果, 并构建6种紫檀属DNA条形码数据库.在龙脑香科树种中也开展了不少相关研究.例如, Tsumura等(2011)通过比较trnL、trnL-trnF、trnH-trnK和psbC-trnS片段构建了84种娑罗双属(Shorea Roxb.)植物分子数据库; Trang等(2015)评估rbcL、matK和trnH-psbA片段在东南亚地区4种坡垒属(Hopea Roxb.)近缘树种中的鉴定效果, 发现matK鉴定效果优于rbcL和trnH-psbA.目前, 龙脑香科植物的DNA条形码研究仅见于对娑罗双属和坡垒属等少数类群, 而整个龙脑香科的条形码尚未形成统一的标准(Kamiya et al., 2005; Tsumura et al., 2011; Trang et al., 2015). ...
... 作为陆地植物的候选DNA条形码(Kress et al., 2005; China Plant BOL Group et al., 2011).近些年, DNA条形码技术的发展十分迅速, 已经应用于一些热带珍贵植物树种的鉴定.例如, Hassold等(2016)对马达加斯加的红木类树种(red wood)进行DNA条形码研究, 评估3个DNA条形码片段(matK、rbcL和trnL)对于不同产地及不同种类红木的鉴定效率; Jiao等(2018)利用6种紫檀属(Pterocarpus Jacq.)木材标本和干燥叶片样品材料, 评估ITS2、matK、ndhF-rpl32和rbcL四个DNA条形码鉴定效果, 并构建6种紫檀属DNA条形码数据库.在龙脑香科树种中也开展了不少相关研究.例如, Tsumura等(2011)通过比较trnL、trnL-trnF、trnH-trnK和psbC-trnS片段构建了84种娑罗双属(Shorea Roxb.)植物分子数据库; Trang等(2015)评估rbcL、matK和trnH-psbA片段在东南亚地区4种坡垒属(Hopea Roxb.)近缘树种中的鉴定效果, 发现matK鉴定效果优于rbcL和trnH-psbA.目前, 龙脑香科植物的DNA条形码研究仅见于对娑罗双属和坡垒属等少数类群, 而整个龙脑香科的条形码尚未形成统一的标准(Kamiya et al., 2005; Tsumura et al., 2011; Trang et al., 2015). ...
... 理想的DNA条形码需具备片段较短、种间变异较大且鉴定效率高等特点(Kress et al., 2005; Taberlet et al., 2007; 任保青和陈之端, 2010).本研究中, 我们发现ITS2片段在龙脑香科植物树种中呈现较高的扩增率和测序成功率(分别为100%和89.5%), 且基于BM、BCM、BLAST和NJ树4种条形码评估方法均获得最高的鉴定成功率, 因此我们认为ITS2是龙脑香科树种首选的DNA条形码.能否成功获得高质量条形码序列决定着条形码技术可否应用到实际物种鉴定工作中, 因此条形码扩增率和测序成功率是条形码筛选时的重要指标.ITS2是ITS序列的1个片段, 位于5.8S和26S rRNA之间, 其序列长度较短, 易于扩增和测序, 且其二级结构能够提供更多的信息(Schultz and Wolf, 2009; China Plant BOL Group et al., 2011).Chen等(2010)通过大规模的取样分析, 认为可将ITS2作为药用植物鉴定的核心条形码; 同时, ITS2在山葡萄(Vitis amurensis)、兰科及红景天属等植物中也显示较好的鉴定效果(宋慧芳等, 2017; De Boer et al., 2017; Zhu et al., 2017).本研究采用引物ITS3/26SE并优化扩增体系, 通过在扩增体系中添加DMSO及BSA提高PCR的产物特异性和产量.此外, 根据ITS2具有较高的GC/AT含量, 使用高保真DNA聚合酶和GC Enhancer优化PCR过程, 提高了ITS2在龙脑香科树种的获得率, 我们建议将20 μL体系作为龙脑香科植物树种ITS2片段扩增的标准体系. ...
DNA barcoding the floras of biodiversity hotspots 2 2008
... 使用Sequencher 4.14软件(GeneCodes Corp., Ann Arbor, Michigan, USA)对双向测序结果进行序列拼接和人工校对, 去除低质量序列和两端引物区.使用MEGA 6.0软件(Tamura et al., 2013)对获得的序列进行比对, 采用K2P (Kimura-2-parameter)模型计算种内及种间平均遗传距离.种内、种间遗传变异通过Taxton DNA软件(Meier et al., 2006)进行计算, 随后使用SPSS V21.0软件(IBM Corp., Armonk, New York, USA)对各片段的种内、种间遗传距离差异进行Wilcoxon Signed Rank检验(Lahaye et al., 2008).采用Taxon DNA软件中的BM (Best Match)和BCM (Best Close Match)两种方法分析鉴定成功率.利用Geneious 6.15软件(Biomatters Ltd.)中Custom Blast功能实现本地BLAST, 以相同位点的百分比(identical sites)作为量化标准, 如果相同物种的identical sites值都大于与其它所有物种个体间的identical sites值, 即认为这个物种被准确鉴定(卢孟孟等, 2013).使用Geneious 6.15软件构建邻接(NJ)树, 设置bootstrap值为1 000, 检验各支点支持率.如果同一个物种的所有个体形成1个单系, 且节点支持率大于50%, 则认为该物种被准确鉴定. ...
... 以往的研究表明, 叶绿体基因组进化速率相对较慢, 能够提供鉴定分析的变异位点较少(Newmaster et al., 2006; Chase et al., 2007; Kress and Erickson, 2007).根据Wilcoxon Signed Rank检验结果, matK和trnL-trnF片段的种间遗传变异较高, 且差异显著, 显示出较高的应用潜力.matK基因序列是植物叶绿体基因中进化较快的编码基因, 基于样品序列matK片段在龙脑香科树种中进行物种鉴定的成功率仅次于ITS2片段, 表明matK用于龙脑香科树种鉴定效果较好.叶绿体基因间隔区trnL-trnF常用于龙脑香科树种的分子系统学研究, 在GenBank数据库中有较多的数据, 但在本研究中基于4种评估方法获得的鉴定成功率较低.rbcL片段是DNA条形码的核心之一, 由于其引物通用性较高而被广泛使用, 但因变异较小不适合用于种级水平的物种鉴定(Kress and Erickson, 2007; Lahaye et al., 2008).本研究结果显示, 叶绿体rbcL片段种内种间遗传变异不明显, 且鉴定成功率低于其它片段, 因此不适合作为热带龙脑香科树种的DNA条形码.综上, 我们建议将ITS2和matK片段作为热带龙脑香科树种的DNA条形码. ...
DNA barcoding and taxonomy in Diptera: a tale of high intraspecific variability and low identification success 1 2006
... 使用Sequencher 4.14软件(GeneCodes Corp., Ann Arbor, Michigan, USA)对双向测序结果进行序列拼接和人工校对, 去除低质量序列和两端引物区.使用MEGA 6.0软件(Tamura et al., 2013)对获得的序列进行比对, 采用K2P (Kimura-2-parameter)模型计算种内及种间平均遗传距离.种内、种间遗传变异通过Taxton DNA软件(Meier et al., 2006)进行计算, 随后使用SPSS V21.0软件(IBM Corp., Armonk, New York, USA)对各片段的种内、种间遗传距离差异进行Wilcoxon Signed Rank检验(Lahaye et al., 2008).采用Taxon DNA软件中的BM (Best Match)和BCM (Best Close Match)两种方法分析鉴定成功率.利用Geneious 6.15软件(Biomatters Ltd.)中Custom Blast功能实现本地BLAST, 以相同位点的百分比(identical sites)作为量化标准, 如果相同物种的identical sites值都大于与其它所有物种个体间的identical sites值, 即认为这个物种被准确鉴定(卢孟孟等, 2013).使用Geneious 6.15软件构建邻接(NJ)树, 设置bootstrap值为1 000, 检验各支点支持率.如果同一个物种的所有个体形成1个单系, 且节点支持率大于50%, 则认为该物种被准确鉴定. ...
DNA barcoding in land plants: evaluation of 1 2006
... 以往的研究表明, 叶绿体基因组进化速率相对较慢, 能够提供鉴定分析的变异位点较少(Newmaster et al., 2006; Chase et al., 2007; Kress and Erickson, 2007).根据Wilcoxon Signed Rank检验结果, matK和trnL-trnF片段的种间遗传变异较高, 且差异显著, 显示出较高的应用潜力.matK基因序列是植物叶绿体基因中进化较快的编码基因, 基于样品序列matK片段在龙脑香科树种中进行物种鉴定的成功率仅次于ITS2片段, 表明matK用于龙脑香科树种鉴定效果较好.叶绿体基因间隔区trnL-trnF常用于龙脑香科树种的分子系统学研究, 在GenBank数据库中有较多的数据, 但在本研究中基于4种评估方法获得的鉴定成功率较低.rbcL片段是DNA条形码的核心之一, 由于其引物通用性较高而被广泛使用, 但因变异较小不适合用于种级水平的物种鉴定(Kress and Erickson, 2007; Lahaye et al., 2008).本研究结果显示, 叶绿体rbcL片段种内种间遗传变异不明显, 且鉴定成功率低于其它片段, 因此不适合作为热带龙脑香科树种的DNA条形码.综上, 我们建议将ITS2和matK片段作为热带龙脑香科树种的DNA条形码. ...
Genome size variation and evolution in Dipterocarpaceae 0 2016
Monophyly of the Asteridae and identification of their major lineages inferred from DNA sequences of 0 1992
ITS2 sequence-structure analysis in phylogenetics: a how-to manual for molecular syste- matics 1 2009
... 理想的DNA条形码需具备片段较短、种间变异较大且鉴定效率高等特点(Kress et al., 2005; Taberlet et al., 2007; 任保青和陈之端, 2010).本研究中, 我们发现ITS2片段在龙脑香科植物树种中呈现较高的扩增率和测序成功率(分别为100%和89.5%), 且基于BM、BCM、BLAST和NJ树4种条形码评估方法均获得最高的鉴定成功率, 因此我们认为ITS2是龙脑香科树种首选的DNA条形码.能否成功获得高质量条形码序列决定着条形码技术可否应用到实际物种鉴定工作中, 因此条形码扩增率和测序成功率是条形码筛选时的重要指标.ITS2是ITS序列的1个片段, 位于5.8S和26S rRNA之间, 其序列长度较短, 易于扩增和测序, 且其二级结构能够提供更多的信息(Schultz and Wolf, 2009; China Plant BOL Group et al., 2011).Chen等(2010)通过大规模的取样分析, 认为可将ITS2作为药用植物鉴定的核心条形码; 同时, ITS2在山葡萄(Vitis amurensis)、兰科及红景天属等植物中也显示较好的鉴定效果(宋慧芳等, 2017; De Boer et al., 2017; Zhu et al., 2017).本研究采用引物ITS3/26SE并优化扩增体系, 通过在扩增体系中添加DMSO及BSA提高PCR的产物特异性和产量.此外, 根据ITS2具有较高的GC/AT含量, 使用高保真DNA聚合酶和GC Enhancer优化PCR过程, 提高了ITS2在龙脑香科树种的获得率, 我们建议将20 μL体系作为龙脑香科植物树种ITS2片段扩增的标准体系. ...
Phylogenetic analysis of 1 1994
... 计算结果见表3.叶绿体片段rbcL、matK和trnL-trnF均获得100%的扩增成功率和测序成功率.通过采用引物ITS2F/ITS3R (Chen et al., 2010)进行扩增, ITS2的扩增成功率为100%, 而有效序列获得率仅为47.4%; 通过调整引物ITS3/26SE (White et al., 1990; Sun et al., 1994)并优化扩增体系, ITS2的有效序列获得率提高到89.5%.19个龙脑香科植物样品材料的扩增和测序结果表明, 叶绿体片段的有效序列获得率明显高于ITS2, 但采用2对ITS2引物进行扩增能够获得较高质量的ITS2序列. ...
Power and limitations of the chloroplast 1 2007
... 理想的DNA条形码需具备片段较短、种间变异较大且鉴定效率高等特点(Kress et al., 2005; Taberlet et al., 2007; 任保青和陈之端, 2010).本研究中, 我们发现ITS2片段在龙脑香科植物树种中呈现较高的扩增率和测序成功率(分别为100%和89.5%), 且基于BM、BCM、BLAST和NJ树4种条形码评估方法均获得最高的鉴定成功率, 因此我们认为ITS2是龙脑香科树种首选的DNA条形码.能否成功获得高质量条形码序列决定着条形码技术可否应用到实际物种鉴定工作中, 因此条形码扩增率和测序成功率是条形码筛选时的重要指标.ITS2是ITS序列的1个片段, 位于5.8S和26S rRNA之间, 其序列长度较短, 易于扩增和测序, 且其二级结构能够提供更多的信息(Schultz and Wolf, 2009; China Plant BOL Group et al., 2011).Chen等(2010)通过大规模的取样分析, 认为可将ITS2作为药用植物鉴定的核心条形码; 同时, ITS2在山葡萄(Vitis amurensis)、兰科及红景天属等植物中也显示较好的鉴定效果(宋慧芳等, 2017; De Boer et al., 2017; Zhu et al., 2017).本研究采用引物ITS3/26SE并优化扩增体系, 通过在扩增体系中添加DMSO及BSA提高PCR的产物特异性和产量.此外, 根据ITS2具有较高的GC/AT含量, 使用高保真DNA聚合酶和GC Enhancer优化PCR过程, 提高了ITS2在龙脑香科树种的获得率, 我们建议将20 μL体系作为龙脑香科植物树种ITS2片段扩增的标准体系. ...
Universal primers for amplification of three non-coding regions of chloroplast DNA 0 1991
MEGA6: molecular evolutionary genetics analysis version 6.0 1 2013
... 使用Sequencher 4.14软件(GeneCodes Corp., Ann Arbor, Michigan, USA)对双向测序结果进行序列拼接和人工校对, 去除低质量序列和两端引物区.使用MEGA 6.0软件(Tamura et al., 2013)对获得的序列进行比对, 采用K2P (Kimura-2-parameter)模型计算种内及种间平均遗传距离.种内、种间遗传变异通过Taxton DNA软件(Meier et al., 2006)进行计算, 随后使用SPSS V21.0软件(IBM Corp., Armonk, New York, USA)对各片段的种内、种间遗传距离差异进行Wilcoxon Signed Rank检验(Lahaye et al., 2008).采用Taxon DNA软件中的BM (Best Match)和BCM (Best Close Match)两种方法分析鉴定成功率.利用Geneious 6.15软件(Biomatters Ltd.)中Custom Blast功能实现本地BLAST, 以相同位点的百分比(identical sites)作为量化标准, 如果相同物种的identical sites值都大于与其它所有物种个体间的identical sites值, 即认为这个物种被准确鉴定(卢孟孟等, 2013).使用Geneious 6.15软件构建邻接(NJ)树, 设置bootstrap值为1 000, 检验各支点支持率.如果同一个物种的所有个体形成1个单系, 且节点支持率大于50%, 则认为该物种被准确鉴定. ...
Application of DNA barcoding markers to the identification of 1 2015
... DNA条形码技术首先由Paul Hebert提出, 是利用1个或少数几个DNA片段对地球上所有物种进行识别和鉴定(Hebert et al., 2003; Kress et al., 2005).自DNA条形码技术的概念被提出以来, 研究者制订了DNA条形码的选择标准: (1) 尽量短的片段, 片段两端连接相对保守的区域, 易于开发通用引物; (2) 要有足够的变异可将物种区分开来(CBOL Plant Working Group et al., 2009; 任保青和陈之端, 2010).利用以上标准, 研究者陆续提出将matK、rbcL、trnH-psbA和ITS/ITS2作为陆地植物的候选DNA条形码(Kress et al., 2005; China Plant BOL Group et al., 2011).近些年, DNA条形码技术的发展十分迅速, 已经应用于一些热带珍贵植物树种的鉴定.例如, Hassold等(2016)对马达加斯加的红木类树种(red wood)进行DNA条形码研究, 评估3个DNA条形码片段(matK、rbcL和trnL)对于不同产地及不同种类红木的鉴定效率; Jiao等(2018)利用6种紫檀属(Pterocarpus Jacq.)木材标本和干燥叶片样品材料, 评估ITS2、matK、ndhF-rpl32和rbcL四个DNA条形码鉴定效果, 并构建6种紫檀属DNA条形码数据库.在龙脑香科树种中也开展了不少相关研究.例如, Tsumura等(2011)通过比较trnL、trnL-trnF、trnH-trnK和psbC-trnS片段构建了84种娑罗双属(Shorea Roxb.)植物分子数据库; Trang等(2015)评估rbcL、matK和trnH-psbA片段在东南亚地区4种坡垒属(Hopea Roxb.)近缘树种中的鉴定效果, 发现matK鉴定效果优于rbcL和trnH-psbA.目前, 龙脑香科植物的DNA条形码研究仅见于对娑罗双属和坡垒属等少数类群, 而整个龙脑香科的条形码尚未形成统一的标准(Kamiya et al., 2005; Tsumura et al., 2011; Trang et al., 2015). ...
Molecular database for classifying 1 2011
... DNA条形码技术首先由Paul Hebert提出, 是利用1个或少数几个DNA片段对地球上所有物种进行识别和鉴定(Hebert et al., 2003; Kress et al., 2005).自DNA条形码技术的概念被提出以来, 研究者制订了DNA条形码的选择标准: (1) 尽量短的片段, 片段两端连接相对保守的区域, 易于开发通用引物; (2) 要有足够的变异可将物种区分开来(CBOL Plant Working Group et al., 2009; 任保青和陈之端, 2010).利用以上标准, 研究者陆续提出将matK、rbcL、trnH-psbA和ITS/ITS2作为陆地植物的候选DNA条形码(Kress et al., 2005; China Plant BOL Group et al., 2011).近些年, DNA条形码技术的发展十分迅速, 已经应用于一些热带珍贵植物树种的鉴定.例如, Hassold等(2016)对马达加斯加的红木类树种(red wood)进行DNA条形码研究, 评估3个DNA条形码片段(matK、rbcL和trnL)对于不同产地及不同种类红木的鉴定效率; Jiao等(2018)利用6种紫檀属(Pterocarpus Jacq.)木材标本和干燥叶片样品材料, 评估ITS2、matK、ndhF-rpl32和rbcL四个DNA条形码鉴定效果, 并构建6种紫檀属DNA条形码数据库.在龙脑香科树种中也开展了不少相关研究.例如, Tsumura等(2011)通过比较trnL、trnL-trnF、trnH-trnK和psbC-trnS片段构建了84种娑罗双属(Shorea Roxb.)植物分子数据库; Trang等(2015)评估rbcL、matK和trnH-psbA片段在东南亚地区4种坡垒属(Hopea Roxb.)近缘树种中的鉴定效果, 发现matK鉴定效果优于rbcL和trnH-psbA.目前, 龙脑香科植物的DNA条形码研究仅见于对娑罗双属和坡垒属等少数类群, 而整个龙脑香科的条形码尚未形成统一的标准(Kamiya et al., 2005; Tsumura et al., 2011; Trang et al., 2015). ...
0
0
DNA barcoding of vouchered xylarium wood specimens of nine endangered 1 2017
... 随着世界各国对森林资源保护的重视, 使用合法认证的木材和木制品已成为国际共识, 因此对木材种类进行准确鉴定的重要性日趋凸显(Dormontt et al., 2015).基于宏观和微观特征的木材鉴定方法往往依赖于经验丰富的技术专家, 且大多数解剖学特征也只能将木材产品鉴定到属级水平(Gasson et al., 2010; Dormontt et al., 2015).近年来, 植物幼苗和木材等非标准材料DNA提取方法的改进大大提高了条形码技术在木材树种及其产品鉴定中的可行性, 研究者在树木形成层(Huang et al., 2015)、植物幼苗(Gon- zalez et al., 2009)和木材标本(Yu et al., 2017; Jiao et al., 2018; 乔梦吉等, 2018)等样品材料中均成功获得条形码片段, 尤其是Tsumura等(2011)在龙脑香科木材样品中成功扩增到trnL、trnL-trnF、trnH- trnK和psbC-trnS片段.这些研究表明以植物叶片为材料扩增到的植物条形码片段也能用于龙脑香科木材产品的鉴定.本研究以龙脑香科树种的干燥叶片为样品材料, 结合GenBank数据分析评估4种条形码片段在龙脑香科植物鉴定方面的应用潜力, 能够为龙脑香科树种及木材产品的条形码鉴定提供借鉴. ...
Genome skimming herbarium spe- cimens for DNA barcoding and phylogenomics 1 2018
... 龙脑香科植物是热带雨林中具有重要生态价值和经济意义的树种, 其木材产品在东南亚地区热带木材市场中有重要地位.本研究涉及的条形码序列来自龙脑香科14属244种, 共计899条, 涵盖了龙脑香科树种82.4%的属和46.1%的种, 序列数据大多来源于东南亚地区龙脑香科树种的相关研究, 在较大范围内代表了龙脑香科树种的遗传变异.目前, 龙脑香科植物的物种亲缘关系还未得到很好的解决与澄清, 这可能在一定程度上影响了DNA条形码在此类群中的应用.分子系统学研究结果不支持将龙脑香亚科(Dipterocarpoideae)分为Dipterocarpeae和Shoreeae 2个族, 理由是龙脑香属(Dipterocarpus Gaertn. f.)被认为是冰片香属(Dryobalanops C.F. Gaertn.)和Shoreeae的姐妹类群, 且娑罗双属(Shorea Roxb.)为非单系分支(Heckenhauer et al., 2017; Ng et al., 2017).利用DNA条形码进行物种鉴定需要较为完善的物种系统发育框架, 因此龙脑香科条形码的研究有赖于其系统发育关系树的构建.随着高通量测序技术(next generation sequencing)的发展, DNA条形码技术衍生出新的发展趋势.例如, 基因组浅层测序(genome skimming)通过获得全基因组较低测序深度基因组数据, 借助生物信息学手段可获得叶绿体全基因组以及核基因组中高度重复区域(nrDNA)等序列(Hollingsworth et al., 2016; Zeng et al., 2018).利用高通量测序技术能够获得物种的基因组水平信息, 为快速分化的近缘物种鉴定提供更加有效的解决办法(李德铢和曾春霞, 2015).随着条形码技术的不断成熟, 以及龙脑香科木材树种DNA条形码数据库的不断完善和龙脑香科系统发育关系的日趋明确, 相信DNA条形码技术将成为热带地区龙脑香科木材树种鉴定的有力工具. ...
Establishment of the most comprehensive 1 2017
... 理想的DNA条形码需具备片段较短、种间变异较大且鉴定效率高等特点(Kress et al., 2005; Taberlet et al., 2007; 任保青和陈之端, 2010).本研究中, 我们发现ITS2片段在龙脑香科植物树种中呈现较高的扩增率和测序成功率(分别为100%和89.5%), 且基于BM、BCM、BLAST和NJ树4种条形码评估方法均获得最高的鉴定成功率, 因此我们认为ITS2是龙脑香科树种首选的DNA条形码.能否成功获得高质量条形码序列决定着条形码技术可否应用到实际物种鉴定工作中, 因此条形码扩增率和测序成功率是条形码筛选时的重要指标.ITS2是ITS序列的1个片段, 位于5.8S和26S rRNA之间, 其序列长度较短, 易于扩增和测序, 且其二级结构能够提供更多的信息(Schultz and Wolf, 2009; China Plant BOL Group et al., 2011).Chen等(2010)通过大规模的取样分析, 认为可将ITS2作为药用植物鉴定的核心条形码; 同时, ITS2在山葡萄(Vitis amurensis)、兰科及红景天属等植物中也显示较好的鉴定效果(宋慧芳等, 2017; De Boer et al., 2017; Zhu et al., 2017).本研究采用引物ITS3/26SE并优化扩增体系, 通过在扩增体系中添加DMSO及BSA提高PCR的产物特异性和产量.此外, 根据ITS2具有较高的GC/AT含量, 使用高保真DNA聚合酶和GC Enhancer优化PCR过程, 提高了ITS2在龙脑香科树种的获得率, 我们建议将20 μL体系作为龙脑香科植物树种ITS2片段扩增的标准体系. ...

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT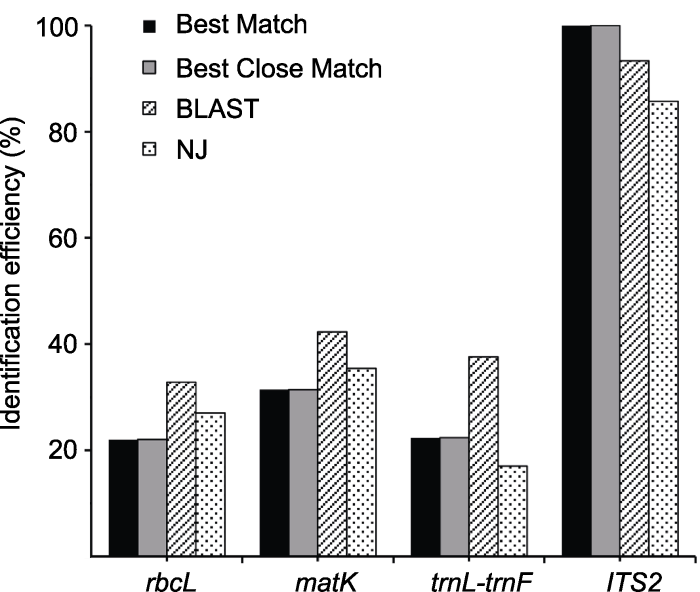 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}