Abstract: Grazing represents the main way for utilization of grassland, and different seasonal grazing affects compensatory growth and regrowth by affecting the trade-off among plant functional traits. To understand the trade-off and variation among the ‘soft’ and ‘hard’ traits under different seasonal grazing, we measured soft traits such as plant height, internode length, and tillers and hard traits such as photosynthetic capacity, antioxidant system and substance contents of osmotic adjustment in root, stem and leaf of Leymus chinensis. Compared with spring grazing, under continuous grazing, L. chinensis adopts stronger avoidance and tolerance strategies. The soft traits and some hard traits, such as photosynthetic capacity, chlorophyII fluorescence characteristics and energy partition show the same trend: net photosynthetic rate decreasing in June and increasing in August. However, the plants have higher photosynthetic product in leaf under spring grazing, which is harmful for regrowth, with vast organic matter contents eaten after grazing. However, under continuous grazing, L. chinensis allocates more to stem and root, which is conducive to uptake water from soil and regrow by using the remaining organic matter. Thus, under three-year grazing, continuous grazing is better for regrowth of L. chinensis and sustainable for grassland.
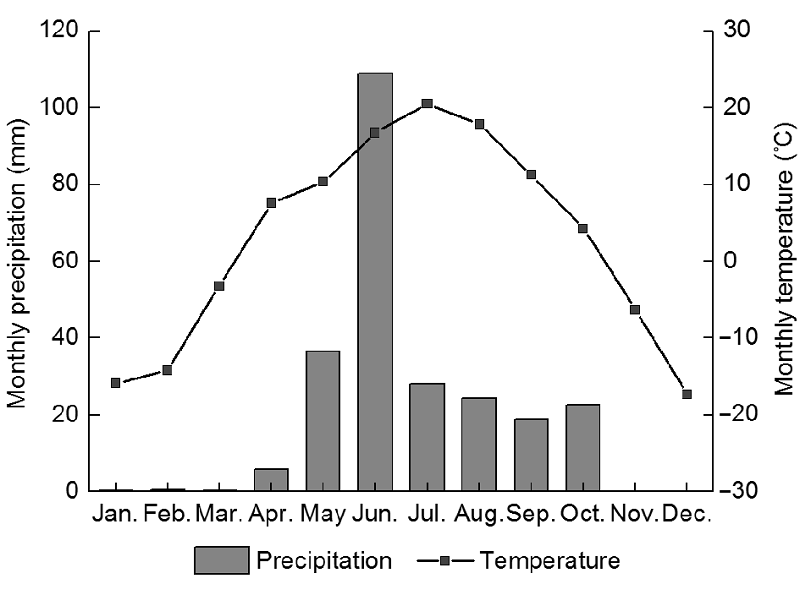
2 结果与讨论2.1 环境条件及群落特性研究区2014年全年平均气温2.6°C, 1月平均气温-19.0°C, 7月平均气温21.4°C。降水的季节性波动较大, 全年总降水量为262 mm, 生长季降水量为228 mm, 占年降水量的87% (图1)。3种不同处理样地土壤营养含量也有所差异(表1)。围封样地(T0)土壤有机质、总氮及总磷的含量最高, 其次为连续放牧样地(T1)。连续放牧样地有效磷含量为2.56 mg·kg-1, 高于围封样地, 春季放牧样地(T2)营养含量最低。研究结果表明, 放牧显著影响放牧样地的植物群落特性 (表2) (P<0.05), T0样地地上生物量为135.83 g∙m-2, 显著高于T1和T2样地, 而T2样地地上生物量显著高于T1样地。立枯和凋落物均表现为T0样地显著高于T1和T2样地, 而T1与T2样地之间无显著差异。T1和T2样地物种丰富度均高于T0样地, 其中T1样地物种丰富度最高(为9), 即放牧增加了物种丰富度。 图1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-307/img_1.png<b>图1</b> 2014年研究区的月降水量和月平均气温<br/><b>Figure 1</b> Monthly total precipitation and mean air temperature in the study sites during 2014 Figure 1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-307/img_1.png<b>图1</b> 2014年研究区的月降水量和月平均气温<br/><b>Figure 1</b> Monthly total precipitation and mean air temperature in the study sites during 2014
图1 2014年研究区的月降水量和月平均气温 Figure 1 Monthly total precipitation and mean air temperature in the study sites during 2014
表1 Table 1 表1 表1 3种样地的土壤特性 Table 1 Soil properties under 3 study sites
Treatments
OM (g∙kg-1)
TN (g∙kg-1)
TP (g∙kg-1)
AP (mg∙kg-1)
T0
19.75
1.50
0.29
2.40
T1
18.85
1.38
0.28
2.56
T2
14.05
1.24
0.30
2.31
T0: Enclosed plot; T1: Continuous grazing plot; T2: Spring grazing plot; OM: Soil organic matter; TN: Total nitrogen; TP: Total phosphorus; AP: Available phosphorus T0: 围封样地; T1: 连续放牧样地; T2: 春季放牧样地; OM: 土壤有机质; TN: 总氮; TP: 总磷; AP: 有效磷
表1 3种样地的土壤特性 Table 1 Soil properties under 3 study sites
表2 Table 2 表2 表2 3种样地群落特性(平均值±标准误, n=9) Table 2 Community characteristics under 3 study sites (means±SE, n=9)
Community characteristics
T0
T1
T2
Aboveground biomass (g∙m-2)
135.83±6.79 a
31.56±2.47 b
55.62±5.20 c
Standing litter (g∙m-2)
63.89±10.27 a
3.78±1.60 b
1.23±0.32 b
Litter (g∙m-2)
61.96±4.09 a
12.44±1.60 b
10.10±2.41 b
Richness
5.89±0.72 a
9.00±0.75 b
7.78±0.43 b
T0, T1 and T2 see Table 1. Different lowercase letters indicate significant differences among the three treatments (LSD test, P<0.05). T0、T1和T2同表1。不同字母表示不同处理间差异显著(LSD检验, P<0.05)。
表2 3种样地群落特性(平均值±标准误, n=9) Table 2 Community characteristics under 3 study sites (means±SE, n=9)
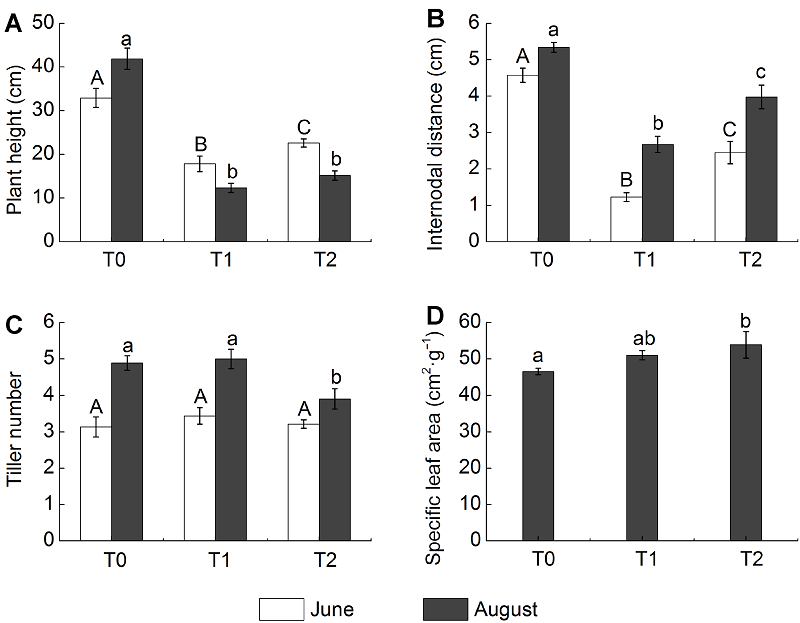
2.2 羊草软性状的响应放牧显著影响了羊草的形态特性, 与围封样地(T0)相比, 类连续放牧(T1)和春季放牧(T2)显著地降低了植物的株高和节间距(图2) (P<0.05)。在6月, T2样地羊草的株高与节间距明显高于T1样地, 但与T0样地无显著差异, 即放牧对羊草的分蘖数无显著影响。在8月, T1和T2样地羊草的株高显著低于T0样地, 但两者间差异不显著, 且T2样地羊草分蘖数显著低于T0与T1样地。放牧导致羊草比叶面积增大, 尽管T1样地羊草比叶面积与T0样地相比差异不显著, 但T2样地羊草比叶面积显著增加(P<0.05)。 图2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-307/img_2.png<b>图2</b> 不同季节放牧下羊草的软性状(平均值±标准误)<br/>T0: 围封样地; T1: 连续放牧样地; T2: 春季放牧样地。不同处理间植物软性状的差异(LSD检验, <i>P</i><0.05), 6月用大写字母表示, 8月用小写字母表示。<br/><b>Figure 2</b> Soft traits of <i>Leymus chinensis</i> under different seasonal grazing treatments (means±SE) <br/>T0: Enclosed plot; T1: Continuous grazing plot; T2: Spring grazing plot. Different letters indicate significant differences among treatments (LSD test, <i>P</i><0.05), with June in capital letters and August in lowercase letters. Figure 2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-307/img_2.png<b>图2</b> 不同季节放牧下羊草的软性状(平均值±标准误)<br/>T0: 围封样地; T1: 连续放牧样地; T2: 春季放牧样地。不同处理间植物软性状的差异(LSD检验, <i>P</i><0.05), 6月用大写字母表示, 8月用小写字母表示。<br/><b>Figure 2</b> Soft traits of <i>Leymus chinensis</i> under different seasonal grazing treatments (means±SE) <br/>T0: Enclosed plot; T1: Continuous grazing plot; T2: Spring grazing plot. Different letters indicate significant differences among treatments (LSD test, <i>P</i><0.05), with June in capital letters and August in lowercase letters.
图2 不同季节放牧下羊草的软性状(平均值±标准误) T0: 围封样地; T1: 连续放牧样地; T2: 春季放牧样地。不同处理间植物软性状的差异(LSD检验, P<0.05), 6月用大写字母表示, 8月用小写字母表示。 Figure 2 Soft traits of Leymus chinensis under different seasonal grazing treatments (means±SE) T0: Enclosed plot; T1: Continuous grazing plot; T2: Spring grazing plot. Different letters indicate significant differences among treatments (LSD test, P<0.05), with June in capital letters and August in lowercase letters.
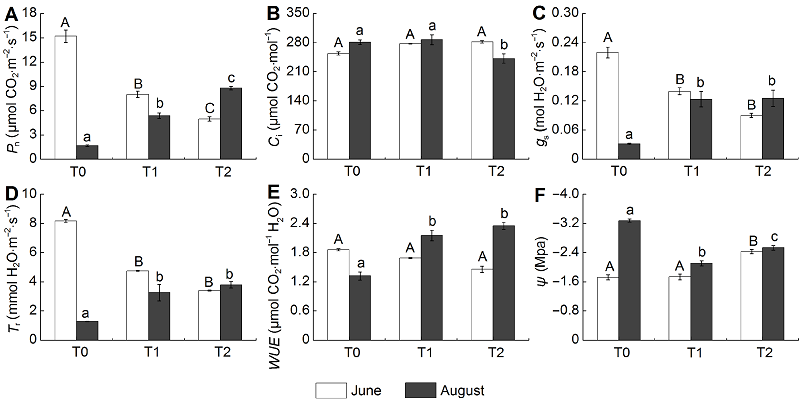
2.3 光合特性和叶片水势的变化放牧极显著影响类连续放牧样地(T1)和春季放牧样地(T2)羊草的气体交换特性(图3) (P<0.01)。在6月, 羊草Pn、Tr和gs都显著低于围封样地(T0) (P<0.05), Ci增加, 但差异不显著, 3种样地WUE无显著差异。T1样地羊草Pn和Tr都高于T2样地, 但两者的gs差异不显著, T2样地羊草叶片ψ最低。8月, 羊草Pn、Tr和gs与6月相比表现出相反的趋势, 即放牧对三者有促进作用, 也提高了羊草WUE (P<0.05)。T2样地羊草Pn最高, 而Ci显著低于T1, 但T1与T2样地羊草Tr和gs无显著差异。 图3https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-307/img_3.png<b>图3</b> 不同季节放牧下羊草的光合特性和叶片水势(平均值±标准误)<br/>T0: 围封样地; T1: 连续放牧样地; T2: 春季放牧样地; <i>P</i><sub>n</sub>: 净光合速率; <i>C</i><sub>i</sub>: 胞间CO<sub>2</sub>浓度; <i>g</i><sub>s</sub>: 气孔导度; <i>T</i><sub>r</sub>: 蒸腾速率; <i>WUE</i>: 水分利用效率; <i>ψ</i>: 叶片水势。羊草不同处理间的差异(LSD检验, <i>P</i><0.05), 6月用大写字母表示, 8月用小写字母表示。<br/><b>Figure 3</b> Photosynthetic characteristics and water potential of <i>Leymus chinensis </i>under different seasonal grazing treatments (means±SE) <br/>T0: Enclosed plot; T1: Continuous grazing plot; T2: Spring grazing plot; <i>P</i><sub>n</sub>: Net photosynthetic rate; <i>C</i><sub>i</sub>: Intercellular CO<sub>2</sub> concentration; <i>g</i><sub>s</sub>: Stomatal conductance; <i>T</i><sub>r</sub>: Transpiration rate; <i>WUE</i>: Water use efficiency; <i>ψ</i>: Water potential. Different letters represent significant differences among treatments (LSD test,<i> P</i><0.05), with June in capital letters and August in lowercase letters. Figure 3https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-307/img_3.png<b>图3</b> 不同季节放牧下羊草的光合特性和叶片水势(平均值±标准误)<br/>T0: 围封样地; T1: 连续放牧样地; T2: 春季放牧样地; <i>P</i><sub>n</sub>: 净光合速率; <i>C</i><sub>i</sub>: 胞间CO<sub>2</sub>浓度; <i>g</i><sub>s</sub>: 气孔导度; <i>T</i><sub>r</sub>: 蒸腾速率; <i>WUE</i>: 水分利用效率; <i>ψ</i>: 叶片水势。羊草不同处理间的差异(LSD检验, <i>P</i><0.05), 6月用大写字母表示, 8月用小写字母表示。<br/><b>Figure 3</b> Photosynthetic characteristics and water potential of <i>Leymus chinensis </i>under different seasonal grazing treatments (means±SE) <br/>T0: Enclosed plot; T1: Continuous grazing plot; T2: Spring grazing plot; <i>P</i><sub>n</sub>: Net photosynthetic rate; <i>C</i><sub>i</sub>: Intercellular CO<sub>2</sub> concentration; <i>g</i><sub>s</sub>: Stomatal conductance; <i>T</i><sub>r</sub>: Transpiration rate; <i>WUE</i>: Water use efficiency; <i>ψ</i>: Water potential. Different letters represent significant differences among treatments (LSD test,<i> P</i><0.05), with June in capital letters and August in lowercase letters.
图3 不同季节放牧下羊草的光合特性和叶片水势(平均值±标准误) T0: 围封样地; T1: 连续放牧样地; T2: 春季放牧样地; Pn: 净光合速率; Ci: 胞间CO2浓度; gs: 气孔导度; Tr: 蒸腾速率; WUE: 水分利用效率; ψ: 叶片水势。羊草不同处理间的差异(LSD检验, P<0.05), 6月用大写字母表示, 8月用小写字母表示。 Figure 3 Photosynthetic characteristics and water potential of Leymus chinensis under different seasonal grazing treatments (means±SE) T0: Enclosed plot; T1: Continuous grazing plot; T2: Spring grazing plot; Pn: Net photosynthetic rate; Ci: Intercellular CO2 concentration; gs: Stomatal conductance; Tr: Transpiration rate; WUE: Water use efficiency; ψ: Water potential. Different letters represent significant differences among treatments (LSD test, P<0.05), with June in capital letters and August in lowercase letters.
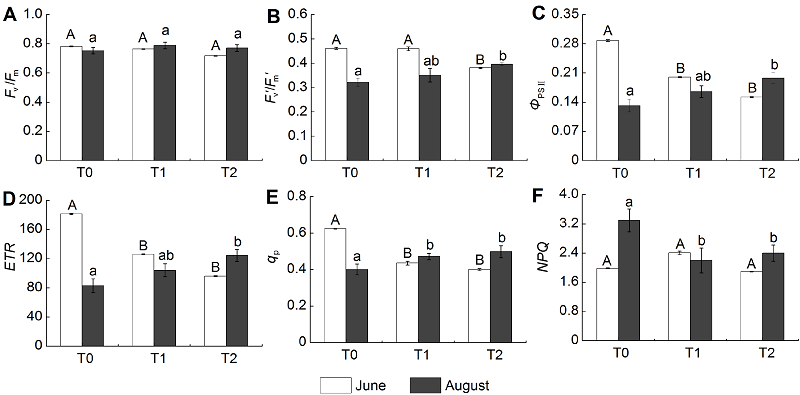
2.4 叶绿素荧光特性不同放牧处理影响羊草的叶绿素荧光特性(图4)。在6月和8月, 围封样地(T0)、类连续放牧样地(T1)和春季放牧样地(T2)之间羊草的Fv/Fm无显著差异, 但T2羊草Fv/Fm较低。6月, T0样地羊草ETR、ΦPSII和qp显著高于T1和T2样地, 即放牧显著降低了羊草的ETR、ΦPSII和qp (P<0.05), T2样地羊草以上3个指标与T1样地相比差异不显著。8月, T0样地羊草ETR、ΦPSII和qp均表现为低于T1和T2样地, 即放牧提高了羊草的ETR、ΦPSII和qp, 但显著降低了NPQ, 其中T1与T0样地羊草ETR和ΦPSII无显著差异, 而T1与T2样地羊草qp和NPQ差异不显著。 图4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-307/img_4.png<b>图4</b> 不同季节放牧下羊草的叶绿素荧光特性(平均值±标准误)<br/>T0: 围封样地; T1: 连续放牧样地; T2: 春季放牧样地; <i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>: PSII最大光化学效率; <i>F</i><sub>v</sub><i>ʹ</i>/<i>F</i><sub>m</sub><i>ʹ</i>: PSII激发能捕获效率; <i>Φ</i><sub>PSII</sub>: 实际光化学效率; <i>ETR</i>: 电子传递效率; <i>q</i><sub>p</sub>: 光化学猝灭系数; <i>NPQ</i>: 非光化学猝灭系数。不同处理下羊草叶绿素荧光特性的差异(LSD检验, <i>P</i><0.05), 6月用大写字母表示, 8月用小写字母表示。<br/><b>Figure 4</b> ChlorophyII fluorescence characteristics of <i>Leymus chinensis</i> under different seasonal grazing treatments (means± SE)<br/>T0: Enclosed plot; T1: Continuous grazing plot; T2: Spring grazing plot; <i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>: Maximal quantum yield of PSII photochemistry; <i>F</i><sub>v</sub><i>ʹ</i>/<i>F</i><sub>m</sub><i>ʹ</i>: The energy harvesting efficiency of PSII; <i>Φ</i><sub>PSII</sub>: Effective quantum yield of PSII photochemistry; <i>ETR</i>: Electron transport rate; <i>q</i><sub>p</sub>: Photochemical quenching coefficient; <i>NPQ</i>: Non-photochemical quenching coefficient. Different letters represent significant differences among treatments (LSD test,<i> P</i><0.05), with June in capital letters and August in lowercase letters. Figure 4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-307/img_4.png<b>图4</b> 不同季节放牧下羊草的叶绿素荧光特性(平均值±标准误)<br/>T0: 围封样地; T1: 连续放牧样地; T2: 春季放牧样地; <i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>: PSII最大光化学效率; <i>F</i><sub>v</sub><i>ʹ</i>/<i>F</i><sub>m</sub><i>ʹ</i>: PSII激发能捕获效率; <i>Φ</i><sub>PSII</sub>: 实际光化学效率; <i>ETR</i>: 电子传递效率; <i>q</i><sub>p</sub>: 光化学猝灭系数; <i>NPQ</i>: 非光化学猝灭系数。不同处理下羊草叶绿素荧光特性的差异(LSD检验, <i>P</i><0.05), 6月用大写字母表示, 8月用小写字母表示。<br/><b>Figure 4</b> ChlorophyII fluorescence characteristics of <i>Leymus chinensis</i> under different seasonal grazing treatments (means± SE)<br/>T0: Enclosed plot; T1: Continuous grazing plot; T2: Spring grazing plot; <i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>: Maximal quantum yield of PSII photochemistry; <i>F</i><sub>v</sub><i>ʹ</i>/<i>F</i><sub>m</sub><i>ʹ</i>: The energy harvesting efficiency of PSII; <i>Φ</i><sub>PSII</sub>: Effective quantum yield of PSII photochemistry; <i>ETR</i>: Electron transport rate; <i>q</i><sub>p</sub>: Photochemical quenching coefficient; <i>NPQ</i>: Non-photochemical quenching coefficient. Different letters represent significant differences among treatments (LSD test,<i> P</i><0.05), with June in capital letters and August in lowercase letters.
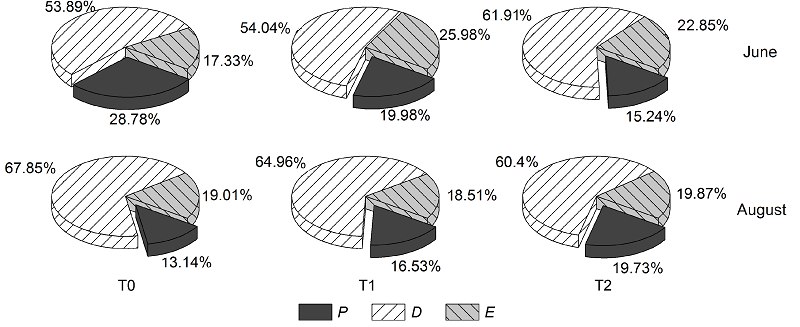
图5 不同季节放牧下羊草的能量分配 P: 光化学反应; D: 热耗散; E: 过剩能量 Figure 5 Energy partition of Leymus chinensis under different seasonal grazing treatments P: Photosynthetic electron transport; D: Thermal energy dissipation; E: Excess
2.6 膜脂过氧化与抗氧化酶活性不同季节放牧处理影响羊草的抗氧化酶系统(图6)。在6月, 围封样地(T0)和连续放牧样地(T1)羊草MDA含量无显著差异, 春季放牧样地(T2)羊草MDA含量显著增加(P<0.05)。此时, 与T0样地相比, T1和T2样地羊草SOD活性显著降低, CAT活性出现升高的趋势。在8月, T1和T2样地羊草MDA含量低于T0样地, 即放牧干扰下, 羊草MDA含量有降低的趋势, T2最低。此时T2样地羊草SOD活性有所降低, CAT活性也较低。而T1样地羊草MDA含量有所降低, 但与T2相比无显著差异, SOD活性降低, 但CAT活性显著升高(P<0.05)。 图6https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-307/img_6.png<b>图6</b> 不同季节放牧下羊草丙二醛含量及抗氧化酶活性(平均值± 标准误)<br/>T0: 围封样地; T1: 连续放牧样地; T2: 春季放牧样地; MDA: 丙二醛; SOD: 超氧化物歧化酶; CAT: 过氧化氢酶。羊草不同处理间的差异(LSD检验, <i>P</i><0.05), 6月用大写字母表示, 8月用小写字母表示。<br/><b>Figure 6</b> The content of malondialdehyde and activities of superoxide dismutase and catalase of <i>Leymus chinensis</i> un- der different seasonal grazing treatments (means±SE)<br/>T0: Enclosed plot; T1: Continuous grazing plot; T2: Spring grazing plot; MDA: Malondialdehyde; SOD: Superoxide dismutase; CAT: Catalase. Different letters represent significant differences among treatments (LSD test, <i>P</i><0.05), with June in capital letters and August in lowercase letters. Figure 6https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-307/img_6.png<b>图6</b> 不同季节放牧下羊草丙二醛含量及抗氧化酶活性(平均值± 标准误)<br/>T0: 围封样地; T1: 连续放牧样地; T2: 春季放牧样地; MDA: 丙二醛; SOD: 超氧化物歧化酶; CAT: 过氧化氢酶。羊草不同处理间的差异(LSD检验, <i>P</i><0.05), 6月用大写字母表示, 8月用小写字母表示。<br/><b>Figure 6</b> The content of malondialdehyde and activities of superoxide dismutase and catalase of <i>Leymus chinensis</i> un- der different seasonal grazing treatments (means±SE)<br/>T0: Enclosed plot; T1: Continuous grazing plot; T2: Spring grazing plot; MDA: Malondialdehyde; SOD: Superoxide dismutase; CAT: Catalase. Different letters represent significant differences among treatments (LSD test, <i>P</i><0.05), with June in capital letters and August in lowercase letters.
图6 不同季节放牧下羊草丙二醛含量及抗氧化酶活性(平均值± 标准误) T0: 围封样地; T1: 连续放牧样地; T2: 春季放牧样地; MDA: 丙二醛; SOD: 超氧化物歧化酶; CAT: 过氧化氢酶。羊草不同处理间的差异(LSD检验, P<0.05), 6月用大写字母表示, 8月用小写字母表示。 Figure 6 The content of malondialdehyde and activities of superoxide dismutase and catalase of Leymus chinensis un- der different seasonal grazing treatments (means±SE) T0: Enclosed plot; T1: Continuous grazing plot; T2: Spring grazing plot; MDA: Malondialdehyde; SOD: Superoxide dismutase; CAT: Catalase. Different letters represent significant differences among treatments (LSD test, P<0.05), with June in capital letters and August in lowercase letters.
2.7 根茎叶渗透调节物质的变化放牧显著影响羊草根茎叶中渗透调节物质的含量(P<0.05) (表3)。6月, 围封样地(T0)羊草叶、茎和根中可溶性糖的含量最高, 春季放牧样地(T2)叶和茎中可溶性糖含量均高于连续放牧样地(T1), 根中两样地则无显著差异。T1样地羊草根和茎中可溶性蛋白与脯氨酸含量显著高于T0与T2样地。8月, T0样地羊草可溶性糖、可溶性蛋白和脯氨酸的含量均较高, T1样地羊草可溶性糖含量与T2相比差异不显著。T1样地羊草茎和根中脯氨酸含量显著高于T2 (P<0.05)。 表3 Table 3 表3 表3 不同季节放牧下羊草渗透调节物质的含量(平均值±标准误) Table 3 Substances contents of osmotic adjustment of Leymus chinensis under different seasonal grazing treatments (means± SE)
June
August
T0
T1
T2
T0
T1
T2
Soluble sugar (mg∙g-1)
Leaf
25.60±1.19 Aa
6.61±0.05 Ba
20.24±2.29 Ca
25.87±0.55 Aa
21.38±2.11 ABa
19.22±3.41 Ba
Stem
18.72±4.52 Ab
23.56±0.60 Bb
17.75±0.96 Aa
49.27±0.76 Ab
30.82±0.69 Bb
26.93±2.04 Bb
Root
11.42±0.68 Ac
7.37±0.64 Aa
6.87±0.87 Ab
23.45±0.55 Aa
11.12±0.33 Bc
5.86±0.33 Cc
Soluble protein (mg∙g-1)
Leaf
0.97±0.07 Aa
1.44±0.12 Bb
2.25±0.11 Ca
1.37±0.06 Aa
0.34±0.06 Ba
0.49±0.01 Ba
Stem
0.64±0.01 Aa
1.09±0.08 Bb
0.78±0.01 Ab
0.67±0.00 ABb
0.89±0.09 Ab
0.35±0.08 Ba
Root
1.17±0.18 Aa
1.96±0.05 Bb
0.59±0.02 Cb
7.11±0.12 Ac
6.53±0.12 Bc
6.20±0.41 Bb
Proline (μg∙g-1)
Leaf
18.64±0.54 Aa
16.71±0.48 Aa
24.18±0.70 Ba
26.73±0.77 Aa
7.30±0.21 Ba
15.73±0.89 Ca
Stem
9.79±0.28 Ab
13.55±0.39 Bb
8.44±0.24 Cb
24.72±1.84 Aa
55.38±1.60 Bb
35.91±1.43 Cb
Root
5.91±0.17 Ac
11.33±0.33 Bc
3.79±0.11 Cc
72.38±2.09 Ab
24.27±0.70 Bc
16.69±0.48 Ca
T0: Enclosed plot; T1: Continuous grazing plot; T2: Spring grazing plot. Different capital letters indicate significant differences among different plots in same month. Different lowercase letters indicate significant differences among root, stem and leaf in same plot. T0: 围封样地; T1: 连续放牧样地; T2: 春季放牧样地。大写字母表示相同月份内不同样地间差异显著, 小写字母表示同一样地根、茎和叶间差异显著。
表3 不同季节放牧下羊草渗透调节物质的含量(平均值±标准误) Table 3 Substances contents of osmotic adjustment of Leymus chinensis under different seasonal grazing treatments (means± SE)
AdlerPB, MilchunasDG, SalaOE, BurkeIC, LauenrothWK (2005). Plant traits and ecosystem grazing effects: comparison of US sagebrush steppe and Patagonian step- pe.Ecol Appl 15, 774-792. [本文引用: 1]
[26]
AnH, LiGQ (2014). Differential effects of grazing on plant functional traits in the desert grassland.Pol J Ecol 62, 239-251. [本文引用: 1]
[27]
BaiYF, HanXG, WuJG, ChenZZ, LiLH (2004). Ecosystem stability and compensatory effects in the Inner Mongolia grassland.Nature 431, 181-184. [本文引用: 1]
[28]
BajjiM, LuttsS, KinetJM (2001). Water deficit effects on solute contribution to osmotic adjustment as a function of leaf ageing in three durum wheat (Triticum durum) cultivars performing differently in arid conditions. Plant Sci 160, 669-681. [本文引用: 1]
[29]
BargerNN, OjimaDS, BelnapJ, WangSP, WangYF, ChenZ (2004). Changes in plant functional groups, litter quality, and soil carbon and nitrogen mineralization with sheep grazing in an Inner Mongolian grassland.Rangeland Ecol Manag 57, 613-619. [本文引用: 1]
[30]
BatesLS, WaldrenRP, TeareID (1973). Rapid determination of free proline for water stress studies.Plant Soil 39, 205-207. [本文引用: 1]
[31]
BenczeS, BambergerZ, JandaT, BallaK, BedőZ, VeiszO (2011). Drought tolerance in cereals in terms of water retention, photosynthesis and antioxidant enzyme activities.Cent Eur J Biol 6, 376-387. [本文引用: 1]
BradfordMM, WilliamsWL (1977). Protein-assay reagent and method. USA 05/694668. 1977-05-17. [本文引用: 1]
[34]
BriskeDD (1996). Strategies of plant survival in grazed systems: a functional interpretation. In: The Ecology and Management of Grazing Systems. Wallingford: CAB International. pp. 37-67. [本文引用: 1]
[35]
BryantJP, KleinDR (1983). Carbon/nutrient balance of boreal plants in relation to vertebrate herbivory. Oikos 40, 357-368. [本文引用: 1]
ChenSP, BaiYF, LinGH, LiangY, HanXG (2005). Effects of grazing on photosynthetic characteristics of major step- pe species in the Xilin River Basin, Inner Mongolia, China.Photosynthetica 43, 559-565. [本文引用: 1]
[38]
ChristineHF, GranhamN (2000). Oxygen processing in photosynthesis: regulation and signaling. New Phytol 146, 359-388. [本文引用: 1]
[39]
ClairborneA (1985). Catalase activity. In: Greenwald RA, ed. Handbook of Methods for Oxygen Radical Research. Boca Raton: CRC Press. pp. 283-284. [本文引用: 1]
[40]
CornelissenJHC, LavorelS, GarnierE, DiazS, BuchmannN, GurvichDE, PausasJG (2003). A handbook of protocols for standardised and easy measurement of plant functional traits worldwide.Aust J Bot 51, 335-380. [本文引用: 1]
Demmig-AdamsB, AdamsIIIWW, BarkerDH, LoganBA, BowlingDR, VerhoevenAS (1996). Using chlorophyll fluorescence to assess the fraction of absorbed light allocated to thermal dissipation of excess excitation. Physiol Plantarum 98, 253-264. [本文引用: 2]
[43]
DhindsaRS, MatoweW (1981). Drought tolerance in two mosses: correlated with enzymatic defence against lipid peroxidation.J Exp Bot 32, 79-91. [本文引用: 1]
[44]
DíazS, Noy-MeirI, CabidoM (2001). Can grazing response of herbaceous plants be predicted from simple vegetative traits.J Appl Ecol 38, 497-508. [本文引用: 1]
[45]
DonaghyDJ, TurnerLR, LanePA, RawnsleyRP (2007). Patterns of leaf and root regrowth, and allocation of water-soluble carbohydrate reserves following defoliation of plants of prairie grass (Bromus willdenowii Kunth.). Grass Forage Sci 62, 497-506. [本文引用: 2]
[46]
DonaldDD, JohnCB (1984). Impacts of grazing intensity and specialed grazing systems on the use and value of rangeland-summary and recommendations. In: Developing Strategies for Rangeland Management. Colorodo: Westview Press. pp. 880-882. [本文引用: 1]
[47]
EarlHJ, TollenaarM (1999). Using chlorophyII fluorometry to compare photosynthetic performance of commercial maize (Zea mays L.) hybrids in the fields in the field. Field Crop Res 61, 201-210. [本文引用: 1]
[48]
FahnestockJT, DetlingJK (2000). Morphological and phy- siological responses of perennial grasses to long-term grazing in the Pryor Mountains, Montana. Am Midl Nat 143, 312-320.
FoyerCH, DescourviresP, KunertKJ (1994). Protection against oxygen radicals: an important defence mechanism studied in transgenic plants. Plant Cell Environ 17, 507-523. [本文引用: 1]
[51]
FunkJL, CornwellWK (2013). Leaf traits within communities: context may affect the mapping of traits to function.Ecology 94, 1893-1897. [本文引用: 1]
GuoX, WangRQ, ChangRY (2014). Effects of nitrogen addition on growth and photosynthetic characteristics of Acer truncatum seedlings. Dendrobiology 72, 151-161. [本文引用: 1]
[54]
HeY, ZhuZJ, YangJ, NiXL, ZhuB (2009). Grafting increases the salt tolerance of tomato by improvement of photosynthesis and enhancement of antioxidant enzymes activity.Environ Exp Bot 66, 270-278. [本文引用: 1]
[55]
KlimešováJ, LatzelV, de BelloF, van GroenendaelJM (2008). Plant functional traits in studies of vegetation changes in response to grazing and mowing: towards a use of more specific traits.Preslia 80, 245-253. [本文引用: 1]
[56]
LarcherW (2003). Carbon utilization and dry matter production. In: Physiological Plant Ecology, 4th edn. Heidelberg: Springer. pp. 71-72. [本文引用: 1]
LiFM, GuoAH, WeiH (1999). Effects of clear plastic film mulch on yield of spring wheat.Field Crop Res 63, 79-86. [本文引用: 1]
[59]
LiSY, VerburgPH, LüSH, WuJL, LiXB (2012). Spatial analysis of the driving factors of grassland degradation under conditions of climate change and intensive use in Inner Mongolia, China.Reg Environ Change 12, 461-474. [本文引用: 1]
[60]
LiangEY, WangYF, XuY, LiuB, ShaoXM (2010). Growth variation in Abies georgei var. smithii along altitudinal gradients in the Sygera Mountains, southeastern Tibetan. Trees 24, 363-373. [本文引用: 1]
[61]
MatysikJ, AliaBB, MohantyP (2002). Molecular mechanisms of quenching of reaction oxygen species by proline under stress in plants. Curr Sci 82, 525-532. [本文引用: 1]
McCormickJI, VirgonaJM, KirkegaardJA (2013). Regrowth of spring canola (Brassica napus) after defoliation. Plant Soil 372, 655-668. [本文引用: 3]
[64]
MooneyKA, HalitschkeR, KesslerA, AgrawalAA (2010). Evolutionary trade-offs in plants mediate the strength of trophic cascades.Science 327, 1642-1644. [本文引用: 1]
[65]
NavasML, RoumetC, BellmannA, LaurentG, GarnierE (2010). Suites of plant traits in species from different stages of a Mediterranean secondary succession.Plant Biol 12, 183-196. [本文引用: 1]
[66]
OgawaA, YamauchiA (2006). Root osmotic adjustment under osmotic stress in maize seedlings. 2. Mode of accumulation of several solutes for osmotic adjustment in the root.Plant Prod Sci 9, 39-46. [本文引用: 1]
[67]
OrtDR (2001). When there is too much light.Plant Physiol 125, 29-32. [本文引用: 1]
[68]
PengY, JiangGM, LiuXH, NiuSL, LiuMZ, BiswasDK (2007). Photosynthesis, transpiration and water use efficiency of four plant species with grazing intensities in hunshandak sandland, China.J Arid Environ 70, 304-315. [本文引用: 1]
[69]
PootP, LambersH (2003). Growth responses to waterlogging and drainage of woody Hakea (Proteaceae) seedlings, originating from contrasting habitats in south-wes- tern Australia.Plant Soil 253, 57-70. [本文引用: 1]
[70]
ShahbaMA, AbbasMS (2014). Drought resistance strategies of seashore paspalum cultivars at different mowing heights.Hortscience 49, 221-229. [本文引用: 1]
[71]
SmithMD, KnappAK (2003). Dominant species maintain ecosystem function with non-random species loss.Ecol Lett 6, 509-517. [本文引用: 1]
[72]
StaalduinenMVA, AntenNPR (2005). Differences in the compensatory growth of two co-occurring grass species in relation to water availability.Oecologia 146, 190-199. [本文引用: 2]
[73]
StearnsSC (1992). The evolution of life histories. New York: Oxford University Press. [本文引用: 1]
[74]
SunJ, JiaYX, GuoSR, LiJ, ShuS (2010). Resistance of spinach plants to seawater stress is correlated with higher activity of xanthophyll cycle and better maintenance of chlorophyll metabolism.Photosynthetica 48, 567-579. [本文引用: 1]
[75]
TongC, WuJ, YongS, YangJ, YongW (2004). A landscapescale assessment of steppe degradation in the Xilin River Basin, Inner Mongolia, China.J Arid Environ 59, 133-149. [本文引用: 1]
[76]
VeskPA, LeishmanMR, WestobyM (2004). Simple traits do not predict grazing response in Australian dry shrublands and woodlands.J Appl Ecol 41, 22-31. [本文引用: 1]
[77]
WangSP, WangYF (2001). Study on over-compensation growth of Cleistogens squarrosa population in Inner Mon- golia steppe. Acta Bot Sin 43, 413-418. [本文引用: 1]
XiuH (2015). Changes of protective enzyme activity and MDA content in leaves of Agropyron cristatum under gra- zing stress. Agr Sci Technol 16, 22-24. [本文引用: 1]
[82]
XuMY, XieF, WangK (2014). Response of vegetation and soil carbon and nitrogen storage to grazing intensity in semi-arid grasslands in the agro-pastoral zone of Northern China. PLoS One 9, e96604. [本文引用: 1]
[83]
YanL, ZhouG, ZhangF (2013). Effects of different grazing intensities on grassland production in China: a meta- analysis.PLoS One 8, e81466. [本文引用: 1]
[84]
ZaiXM, ZhuSN, QinP, WangXY, CheL, LuoFX (2012). Effect of glomus mosseae on chlorophyll content, chlorophyll fluorescence parameters, and chloroplast ultrastructure of beach plum (Prunus maritima) under NaCl stress. Photosynthetica 50, 323-328. [本文引用: 1]
[85]
ZhangZP, MiaoMM, WangCL (2015). Effects of ala on photosynthesis, antioxidant enzyme activity, and gene expression, and regulation of proline accumulation in tomato seedlings under NaCl stress.J Plant Growth Regul 34, 637-650. [本文引用: 1]
[86]
ZhaoW, ChenSP, HanXG, LinGH (2009). Effects of long-term grazing on the morphological and functional traits of Leymus chinensis in the semiarid grassland of Inner Mongolia, China. Ecol Res 24, 99-108. [本文引用: 3]
[87]
ZhengSX, LanZC, LiWH, ShaoRX, ShanYM, WanHW, TaubeF, BaiYF (2011). Differential responses of plant functional trait to grazing between two contrasting dominant C3 and C4 species in a typical steppe of Inner Mongolia, China. Plant Soil 340, 141-155. [本文引用: 3]
[88]
ZhouRL, ZhaoHL (2004). Seasonal pattern of antioxidant enzyme system in the roots of perennial forage grasses grown in Alpine habitat, related to freezing tolerance.Phy- siol Plantarum 121, 399-408. [本文引用: 2]
不同时期放牧对典型草原群落地上生产力的影响 1 2015
... 锡林郭勒盟草原是典型的半干旱草原, 放牧是其传统的土地利用方式.但过度放牧已导致70%的草地处于不同程度的退化中, 大大减少了草地的生产力, 草地退化已经成为重要的经济和生态问题(Tong et al., 2004).优势种是草地生态群落的重要组成部分, 虽然物种数量较少, 但在群落生物量中的分配比例较大, 并在很大程度上决定了草地生态系统的结构和功能(Smith and Knapp, 2003; Bai et al., 2004).羊草(Leymus chinensis)是我国温带草地生态系统的主要优势种, 为多年生禾本科赖草属植物, 草质好、适口性强, 其根茎主要分布在地下10 cm, 根茎发达且繁殖能力强, 有较强的耐牧性(Staalduinen and Anten, 2005).由于草地植物自身具有季节生长节律和再生性(Donald and John, 1984; 李建龙和许鹏, 1991), 传统固定放牧时间长度会导致早期放牧区的可利用牧草的生物量少于后期放牧区, 牧草利用率高, 采食强度大, 放牧后地上现存量低, 不利于草地植物的再生及草地的可持续利用(李勤奋等, 2004; 赵康和宝音陶格涛, 2014; 敖敦高娃和宝音陶格涛, 2015).本研究以优势种羊草为研究对象, 在内蒙古温带典型草原以群落留茬高度6 cm定为终牧期, 并设置了围封、类连续放牧和春季放牧3种处理, 通过测定不同生长季及不同处理下羊草形态和光合生理、抗氧化系统以及渗透调节物等生理特性, 探讨优势种羊草根茎叶功能性状对季节放牧的响应及其权衡.本研究旨在分析优势种对季节放牧的形态软性状的响应及适应机制, 同时探讨羊草叶片光合特性与根茎叶光合产物积累对放牧的响应及适应机制, 以期进一步揭示不同季节放牧下羊草软硬性状的权衡. ...
... 放牧干扰降低了群落的生物量, 但也增加了群落的生物多样性(Zhao et al., 2009; Zheng et al., 2011).放牧干扰下优势种羊草生物量明显降低, 其相对地上生物量所占比例也明显降低.其原因为羊草草质好, 适口性强, 更易被啃食(Chen et al., 2005).放牧显著降低了羊草的软性状(株高和节间距), 羊草表现出矮小化特征, 这也是一种避牧性策略, 而羊草的矮小化是导致地上生物量和草原生产力降低的直接因素(王炜等, 2000; Zheng et al., 2011; 李西良等, 2014).类连续放牧样地羊草株高较春季放牧样地低, 可能是由于羊草改变了其地上与地下资源的分配比例(尹丽等, 2012).放牧干扰使得地上生物量被啃食, 地下储存减少, 影响春季植物的再生, 从而导致植株矮小(白永飞等, 1996).植物株高不仅与其避牧性机制有关, 还与植物的竞争光能力密切相关(Briske, 1996; Wes- toby, 1998; Cornelissen et al., 2003).春季放牧样地与类连续放牧样地羊草相比有较高的株高和较大的比叶面积, 导致春季放牧样地羊草能接受更多的光能.8月, 放牧干扰下的羊草光合速率高于围封样地.这是由于放牧干扰下, 养分通过粪便重新回到土壤中, 从而使得羊草具有较长的生长周期.在牧后恢复期, 羊草地下根茎系统更倾向于快速消耗土壤中的养分来进行补偿性生长, 也导致放牧样地土壤养分相对较低(Bryant and Klein, 1983; Staalduinen and Anten, 2005).春季放牧样地土壤养分最少, 但在8月该样地羊草比叶面积较大, 叶片较薄.较薄的植物叶片需要较少的光合组织就能增加光合能力, 因此, 在缺乏养分的条件下, 植物往往通过增大比叶面积而实现快速生长(Guo et al., 2014). ...
大豆植株不同冠层种子活力及其萌发中抗氧化酶活性 1 2016
... 在环境胁迫下, 植物PSII吸收的光能不能通过ETR传递出去用于光化学反应(P), 激发态的叶绿素分子会与O2反应, 产生大量的单线态氧, 虽然单线态氧的产生吸收了多余的电子, 减缓了光抑制, 但单线态氧的积累不能及时被SOD及CAT等清除, 也会破坏细胞膜(Christine and Graham, 2000; He et al., 2009; Bencze et al., 2011).MDA是细胞膜不饱和脂肪酸分解的产物, 反映细胞膜受伤害的程度(Zhou and Zhao, 2004).6月, 春季放牧样地羊草MDA含量较高, 表明细胞膜发生了膜脂过氧化, 但此时羊草SOD活性较低, 清除自由基的能力较弱.因此, 叶绿素分子吸收的光能大多用于热耗散(D), 光合速率降低.SOD可以将自由基转化为H2O2, H2O2对植物细胞具有毒害作用, 而CAT可以有效将H2O2转化为H2O和O2 (Bowler et al., 1992; Foyer et al., 1994; 陈思羽等, 2016).8月, 春季放牧样地羊草SOD和CAT活性较高, MDA含量最低, 表明抗氧化酶有效地清除了自由基, 保护细胞膜.类连续放牧样地羊草MDA含量出现降低的趋势, SOD活性显著降低, 但CAT活性明显提高, 表明类连续放牧样地羊草可能是依赖于谷胱甘肽和抗坏血酸等抗氧化剂来清除自由基(Zhang et al., 2015). ...
植物叶经济谱的研究进展 1 2014
... 由于可利用资源总量有限, 植物会在功能性状之间进行资源优化配置, 即权衡策略(Stearns, 1992; 张大勇, 2004; 陈莹婷和许振柱, 2014).植物根据所处生境的资源条件, 调整、转换或补偿自身功能, 以平衡生存、生长及繁殖三大目的, 最终表现在植物器官的组织结构及其生理性状上(Navas et al., 2010; Whitman and Aarssen, 2010). ...
不同放牧制度下短花针茅荒漠草原碳平衡研究 1 2010
... 放牧是草地利用的基本形式, 也是干扰草地生产力和植被动态最主要的方式之一(侯向阳和徐海红, 2010; Yan et al., 2013).但长期过度放牧会导致植被盖度减小和草地沙化, 使草地生产力持续衰减, 严重影响草地生态系统的结构和功能(Barger et al., 2004; Li et al., 2012; Xu et al., 2014).植物功能性状能够响应生存环境的变化并(或)对生态系统功能有一定影响(Díaz et al., 2001), 是植物在特定环境下生存策略的重要表现, 为预测放牧对生态系统的影响提供了一种有效的工具(Adler et al., 2005; Klimešová et al., 2008; Mooney et al., 2010).因此, 研究放牧利用下植物功能性状的变化具有重要意义. ...
... 锡林郭勒盟草原是典型的半干旱草原, 放牧是其传统的土地利用方式.但过度放牧已导致70%的草地处于不同程度的退化中, 大大减少了草地的生产力, 草地退化已经成为重要的经济和生态问题(Tong et al., 2004).优势种是草地生态群落的重要组成部分, 虽然物种数量较少, 但在群落生物量中的分配比例较大, 并在很大程度上决定了草地生态系统的结构和功能(Smith and Knapp, 2003; Bai et al., 2004).羊草(Leymus chinensis)是我国温带草地生态系统的主要优势种, 为多年生禾本科赖草属植物, 草质好、适口性强, 其根茎主要分布在地下10 cm, 根茎发达且繁殖能力强, 有较强的耐牧性(Staalduinen and Anten, 2005).由于草地植物自身具有季节生长节律和再生性(Donald and John, 1984; 李建龙和许鹏, 1991), 传统固定放牧时间长度会导致早期放牧区的可利用牧草的生物量少于后期放牧区, 牧草利用率高, 采食强度大, 放牧后地上现存量低, 不利于草地植物的再生及草地的可持续利用(李勤奋等, 2004; 赵康和宝音陶格涛, 2014; 敖敦高娃和宝音陶格涛, 2015).本研究以优势种羊草为研究对象, 在内蒙古温带典型草原以群落留茬高度6 cm定为终牧期, 并设置了围封、类连续放牧和春季放牧3种处理, 通过测定不同生长季及不同处理下羊草形态和光合生理、抗氧化系统以及渗透调节物等生理特性, 探讨优势种羊草根茎叶功能性状对季节放牧的响应及其权衡.本研究旨在分析优势种对季节放牧的形态软性状的响应及适应机制, 同时探讨羊草叶片光合特性与根茎叶光合产物积累对放牧的响应及适应机制, 以期进一步揭示不同季节放牧下羊草软硬性状的权衡. ...
1 2005
... 放牧是草地利用的基本形式, 也是干扰草地生产力和植被动态最主要的方式之一(侯向阳和徐海红, 2010; Yan et al., 2013).但长期过度放牧会导致植被盖度减小和草地沙化, 使草地生产力持续衰减, 严重影响草地生态系统的结构和功能(Barger et al., 2004; Li et al., 2012; Xu et al., 2014).植物功能性状能够响应生存环境的变化并(或)对生态系统功能有一定影响(Díaz et al., 2001), 是植物在特定环境下生存策略的重要表现, 为预测放牧对生态系统的影响提供了一种有效的工具(Adler et al., 2005; Klimešová et al., 2008; Mooney et al., 2010).因此, 研究放牧利用下植物功能性状的变化具有重要意义. ...
1 2014
... 在特定环境下, 植物功能性状间具有权衡关系(Wright et al., 2004; Lavorel, 2013; Funk and Cornwell, 2013).放牧利用下, 植被盖度减小, 植株整体呈现矮小化, 生产力衰减, 而这些现象是一系列植物功能性状相互权衡综合变化的结果(王炜等, 2000; An and Li, 2014; 李西良等, 2015).目前, 关于植物功能性状权衡方面的研究主要集中于通过软性状(soft traits)来评估植物对环境的响应与适应策略.随着放牧强度增加, 植物表现出株高降低、节间距缩短、叶面积减少和生物量降低的特征(Zhao et al., 2009; Zheng et al., 2011).因为高大、叶片大的植株更易于被啃食, 植株会通过矮化和匍匐生长等一些形态性状变化降低牲畜的选择性(Vesk et al., 2004).与此同时, 植株的分蘖能力会相对增加, 以促进被啃食后的快速生长(Zhao et al., 2009).Fahnestock和Detling (2000)认为, 放牧对优势种的改变主要是对其形态特性改变的结果而不是生理特性的改变.但越来越多的研究表明, 放牧可以影响牧后残留组织光系统II (PSII)活性氧的产生与清除、气体交换速率以及光合产物的积累, 并且大多数硬性状(hard traits)的研究仅停留在叶片水平(Donaghy et al., 2007; Xiu, 2015).植物的软性状与不易测量的硬性状密切相关, 如光合速率可以影响碳水化合物的积累, 从而影响生物量和叶面积, 而株高和叶面积也会影响光合作用中光的捕获(Wang and Wang, 2001; Peng et al., 2007; 晏欣等, 2013).然而这些研究大多基于放牧强度, 对于季节放牧下植株硬性状和软性状的变化及两者权衡关系报道较少. ...
1 2004
... 锡林郭勒盟草原是典型的半干旱草原, 放牧是其传统的土地利用方式.但过度放牧已导致70%的草地处于不同程度的退化中, 大大减少了草地的生产力, 草地退化已经成为重要的经济和生态问题(Tong et al., 2004).优势种是草地生态群落的重要组成部分, 虽然物种数量较少, 但在群落生物量中的分配比例较大, 并在很大程度上决定了草地生态系统的结构和功能(Smith and Knapp, 2003; Bai et al., 2004).羊草(Leymus chinensis)是我国温带草地生态系统的主要优势种, 为多年生禾本科赖草属植物, 草质好、适口性强, 其根茎主要分布在地下10 cm, 根茎发达且繁殖能力强, 有较强的耐牧性(Staalduinen and Anten, 2005).由于草地植物自身具有季节生长节律和再生性(Donald and John, 1984; 李建龙和许鹏, 1991), 传统固定放牧时间长度会导致早期放牧区的可利用牧草的生物量少于后期放牧区, 牧草利用率高, 采食强度大, 放牧后地上现存量低, 不利于草地植物的再生及草地的可持续利用(李勤奋等, 2004; 赵康和宝音陶格涛, 2014; 敖敦高娃和宝音陶格涛, 2015).本研究以优势种羊草为研究对象, 在内蒙古温带典型草原以群落留茬高度6 cm定为终牧期, 并设置了围封、类连续放牧和春季放牧3种处理, 通过测定不同生长季及不同处理下羊草形态和光合生理、抗氧化系统以及渗透调节物等生理特性, 探讨优势种羊草根茎叶功能性状对季节放牧的响应及其权衡.本研究旨在分析优势种对季节放牧的形态软性状的响应及适应机制, 同时探讨羊草叶片光合特性与根茎叶光合产物积累对放牧的响应及适应机制, 以期进一步揭示不同季节放牧下羊草软硬性状的权衡. ...
1 2001
... 可溶性糖是光合作用的直接产物, 是植物生长发育的主要能量来源(Li et al., 1999; 刘浩荣等, 2007; McCormick et al., 2013).放牧干扰影响了植物体内可溶性糖的产生与储存.在牧后恢复期, 植物首先会利用残茬和根中的水溶性碳水化合物, 促进叶片快速再生, 提高植物的光合能力; 其次利用茎和根中储存的能量物质.在6月和8月放牧前, 羊草茎中可溶性糖含量显著高于根, 说明牧后恢复一段时期后, 茎中储存了一定的能量(Donaghy et al., 2007; McCormick et al., 2013).6月, 春季放牧样地羊草茎中可溶性糖含量显著降低, 这是由于春季放牧样地羊草光合器官受损使净光合作用降低所致; 而类连续放牧样地羊草较高的光合作用使其茎中积累了较多的可溶性糖.在季节放牧下, 6月和8月植物叶片水势较低, 因此, 羊草处于干旱和放牧双重胁迫下.在胁迫环境中, 植物会通过提高渗透调节物质(如可溶性蛋白和脯氨酸等) 含量提高渗透势, 增加细胞液浓度以对细胞起到保护作用(Bajji et al., 2001; Matysik et al., 2002; 尹本丰和张元明, 2015).类连续放牧样地羊草叶片脯氨酸和可溶性蛋白含量较少, 而根与茎中含量较高.单立山等(2015)在探讨土壤水分胁迫对红砂(Reaumuria songarica)幼苗生长和渗透调节物质的影响中发现, 土壤水分胁迫下脯氨酸和可溶性蛋白在根中积累较多.这是由于在渗透胁迫发生时, 根比叶片反应更快 (Ogawa and Yamauchi, 2006), 并且植物更倾向于降低生命代谢器官的资源分配率, 将有限的资源尽可能分配到茎和根部来实现更大程度的保水与吸水, 以抵御干旱逆境, 从而维持生存(尹丽等, 2012).本研究中, 春季放牧样地羊草可溶性蛋白和脯氨酸较多地分配在叶片中, 而类连续放牧样地羊草则在根和茎中分配较多.相对于春季放牧样地, 类连续放牧样地羊草渗透调节物质和可溶性糖积累较多, 因此, 在牧后恢复期或降水量较多时, 类连续放牧样地羊草能更快速恢复生长(Shahba and Abbas, 2014). ...
1 2004
... 放牧是草地利用的基本形式, 也是干扰草地生产力和植被动态最主要的方式之一(侯向阳和徐海红, 2010; Yan et al., 2013).但长期过度放牧会导致植被盖度减小和草地沙化, 使草地生产力持续衰减, 严重影响草地生态系统的结构和功能(Barger et al., 2004; Li et al., 2012; Xu et al., 2014).植物功能性状能够响应生存环境的变化并(或)对生态系统功能有一定影响(Díaz et al., 2001), 是植物在特定环境下生存策略的重要表现, 为预测放牧对生态系统的影响提供了一种有效的工具(Adler et al., 2005; Klimešová et al., 2008; Mooney et al., 2010).因此, 研究放牧利用下植物功能性状的变化具有重要意义. ...
1 1973
... 分别于6月和8月测定光合特性的当天上午9:00- 11:00, 采集羊草的根、茎和叶, 分别称重后迅速放入液氮中保存, 带回实验室用于生化指标的测定.可溶性糖含量采用蒽酮比色法测定(王学奎, 2006).可溶性蛋白含量采用考马斯亮蓝染色法测定(Bradford and Williams, 1997).脯氨酸含量采用茚三酮显色法测定(Bates et al., 1973). ...
1 2011
... 在环境胁迫下, 植物PSII吸收的光能不能通过ETR传递出去用于光化学反应(P), 激发态的叶绿素分子会与O2反应, 产生大量的单线态氧, 虽然单线态氧的产生吸收了多余的电子, 减缓了光抑制, 但单线态氧的积累不能及时被SOD及CAT等清除, 也会破坏细胞膜(Christine and Graham, 2000; He et al., 2009; Bencze et al., 2011).MDA是细胞膜不饱和脂肪酸分解的产物, 反映细胞膜受伤害的程度(Zhou and Zhao, 2004).6月, 春季放牧样地羊草MDA含量较高, 表明细胞膜发生了膜脂过氧化, 但此时羊草SOD活性较低, 清除自由基的能力较弱.因此, 叶绿素分子吸收的光能大多用于热耗散(D), 光合速率降低.SOD可以将自由基转化为H2O2, H2O2对植物细胞具有毒害作用, 而CAT可以有效将H2O2转化为H2O和O2 (Bowler et al., 1992; Foyer et al., 1994; 陈思羽等, 2016).8月, 春季放牧样地羊草SOD和CAT活性较高, MDA含量最低, 表明抗氧化酶有效地清除了自由基, 保护细胞膜.类连续放牧样地羊草MDA含量出现降低的趋势, SOD活性显著降低, 但CAT活性明显提高, 表明类连续放牧样地羊草可能是依赖于谷胱甘肽和抗坏血酸等抗氧化剂来清除自由基(Zhang et al., 2015). ...
1 1992
... 在环境胁迫下, 植物PSII吸收的光能不能通过ETR传递出去用于光化学反应(P), 激发态的叶绿素分子会与O2反应, 产生大量的单线态氧, 虽然单线态氧的产生吸收了多余的电子, 减缓了光抑制, 但单线态氧的积累不能及时被SOD及CAT等清除, 也会破坏细胞膜(Christine and Graham, 2000; He et al., 2009; Bencze et al., 2011).MDA是细胞膜不饱和脂肪酸分解的产物, 反映细胞膜受伤害的程度(Zhou and Zhao, 2004).6月, 春季放牧样地羊草MDA含量较高, 表明细胞膜发生了膜脂过氧化, 但此时羊草SOD活性较低, 清除自由基的能力较弱.因此, 叶绿素分子吸收的光能大多用于热耗散(D), 光合速率降低.SOD可以将自由基转化为H2O2, H2O2对植物细胞具有毒害作用, 而CAT可以有效将H2O2转化为H2O和O2 (Bowler et al., 1992; Foyer et al., 1994; 陈思羽等, 2016).8月, 春季放牧样地羊草SOD和CAT活性较高, MDA含量最低, 表明抗氧化酶有效地清除了自由基, 保护细胞膜.类连续放牧样地羊草MDA含量出现降低的趋势, SOD活性显著降低, 但CAT活性明显提高, 表明类连续放牧样地羊草可能是依赖于谷胱甘肽和抗坏血酸等抗氧化剂来清除自由基(Zhang et al., 2015). ...
1 1977
... 分别于6月和8月测定光合特性的当天上午9:00- 11:00, 采集羊草的根、茎和叶, 分别称重后迅速放入液氮中保存, 带回实验室用于生化指标的测定.可溶性糖含量采用蒽酮比色法测定(王学奎, 2006).可溶性蛋白含量采用考马斯亮蓝染色法测定(Bradford and Williams, 1997).脯氨酸含量采用茚三酮显色法测定(Bates et al., 1973). ...
1 1996
... 放牧干扰降低了群落的生物量, 但也增加了群落的生物多样性(Zhao et al., 2009; Zheng et al., 2011).放牧干扰下优势种羊草生物量明显降低, 其相对地上生物量所占比例也明显降低.其原因为羊草草质好, 适口性强, 更易被啃食(Chen et al., 2005).放牧显著降低了羊草的软性状(株高和节间距), 羊草表现出矮小化特征, 这也是一种避牧性策略, 而羊草的矮小化是导致地上生物量和草原生产力降低的直接因素(王炜等, 2000; Zheng et al., 2011; 李西良等, 2014).类连续放牧样地羊草株高较春季放牧样地低, 可能是由于羊草改变了其地上与地下资源的分配比例(尹丽等, 2012).放牧干扰使得地上生物量被啃食, 地下储存减少, 影响春季植物的再生, 从而导致植株矮小(白永飞等, 1996).植物株高不仅与其避牧性机制有关, 还与植物的竞争光能力密切相关(Briske, 1996; Wes- toby, 1998; Cornelissen et al., 2003).春季放牧样地与类连续放牧样地羊草相比有较高的株高和较大的比叶面积, 导致春季放牧样地羊草能接受更多的光能.8月, 放牧干扰下的羊草光合速率高于围封样地.这是由于放牧干扰下, 养分通过粪便重新回到土壤中, 从而使得羊草具有较长的生长周期.在牧后恢复期, 羊草地下根茎系统更倾向于快速消耗土壤中的养分来进行补偿性生长, 也导致放牧样地土壤养分相对较低(Bryant and Klein, 1983; Staalduinen and Anten, 2005).春季放牧样地土壤养分最少, 但在8月该样地羊草比叶面积较大, 叶片较薄.较薄的植物叶片需要较少的光合组织就能增加光合能力, 因此, 在缺乏养分的条件下, 植物往往通过增大比叶面积而实现快速生长(Guo et al., 2014). ...
1 1983
... 放牧干扰降低了群落的生物量, 但也增加了群落的生物多样性(Zhao et al., 2009; Zheng et al., 2011).放牧干扰下优势种羊草生物量明显降低, 其相对地上生物量所占比例也明显降低.其原因为羊草草质好, 适口性强, 更易被啃食(Chen et al., 2005).放牧显著降低了羊草的软性状(株高和节间距), 羊草表现出矮小化特征, 这也是一种避牧性策略, 而羊草的矮小化是导致地上生物量和草原生产力降低的直接因素(王炜等, 2000; Zheng et al., 2011; 李西良等, 2014).类连续放牧样地羊草株高较春季放牧样地低, 可能是由于羊草改变了其地上与地下资源的分配比例(尹丽等, 2012).放牧干扰使得地上生物量被啃食, 地下储存减少, 影响春季植物的再生, 从而导致植株矮小(白永飞等, 1996).植物株高不仅与其避牧性机制有关, 还与植物的竞争光能力密切相关(Briske, 1996; Wes- toby, 1998; Cornelissen et al., 2003).春季放牧样地与类连续放牧样地羊草相比有较高的株高和较大的比叶面积, 导致春季放牧样地羊草能接受更多的光能.8月, 放牧干扰下的羊草光合速率高于围封样地.这是由于放牧干扰下, 养分通过粪便重新回到土壤中, 从而使得羊草具有较长的生长周期.在牧后恢复期, 羊草地下根茎系统更倾向于快速消耗土壤中的养分来进行补偿性生长, 也导致放牧样地土壤养分相对较低(Bryant and Klein, 1983; Staalduinen and Anten, 2005).春季放牧样地土壤养分最少, 但在8月该样地羊草比叶面积较大, 叶片较薄.较薄的植物叶片需要较少的光合组织就能增加光合能力, 因此, 在缺乏养分的条件下, 植物往往通过增大比叶面积而实现快速生长(Guo et al., 2014). ...
... 于2014年6月和8月放牧前, 在无风晴天的上午9:00- 11:00, 利用LI-6400 (LI-COR, Lincoln, USA)光合测定系统测定气体交换参数.每次测定3-5株, 每株植物测定3-5个长势相近的叶片, 设9次重复.测定指标包括: 净光合速率(net photosynthetic rate, Pn)、蒸腾速率(transpiration rate, Tr)、胞间二氧化碳浓度(inter- cellular CO2 concentration, Ci)、气孔导度(stomatal conductance, gs)以及计算所得水分利用效率(WUE= Pn/Tr).同时测定叶绿素荧光参数, 测前暗适应半小时, 测得最大荧光(maximal fluorescence yield of the dark-adapted state, Fm)和最小荧光(minimal fluorescence yield of the dark-adapted state, F0).在光下Pn稳定后测得光下最大荧光(maximal fluorescence yield of the light-adapted state, Fmʹ)和稳态荧光(steady-state fluorescence yield, Fs).通过计算得到以下参数: 最大光量子效率(Fv/Fm=(Fm-F0)/Fm), PSII激发能捕获效率Fvʹ/Fmʹ=(Fmʹ-F0ʹ)/Fmʹ, 实际量子效率(ΦPSII=(Fmʹ-Fs)/Fmʹ), 光化学猝灭系数(qP=(Fmʹ- Fs)/(Fmʹ-F0ʹ)) (许大全, 2013)和电子传递速率(ETR= ΦPSII×PPFD×0.5×α) (Maxwell and Johnson, 2000), 能量分配计算得光化学反应P=FvʹFmʹ×qpʹ, 热耗散D=1-(Fvʹ/Fmʹ), 激发能E=Fvʹ/Fmʹ×(1-qp) (Demmig- Admas et al., 1996). ... ... 6月是羊草生长速率最快的季节, 因此, 围封样地羊草有较高的净光合速率, 而放牧样地羊草Pn降低, gs降低, 但Ci上升, 即放牧样地羊草光合速率的降低是由非气孔因素导致的(Farquhar and Sharkey, 1982; 白永飞和徐志信, 1994; 张仁和等, 2011).Fv/Fm反映了PSII潜在的量子效率, 用于指示植物光合性能, 其值在0.75-0.85之间, 表明植物没有受到环境胁迫(厉广辉等, 2014; Catoni et al., 2015).类连续放牧样地羊草Pn显著高于春季放牧样地, 可能是由于春季放牧样地羊草Fv/Fm小于0.75, 说明其光合机构受到一定的损伤, 导致光抑制(Sun et al., 2010; 张仁和等, 2011).此时羊草PSII吸收的过剩光能会通过叶黄素循环和跨类囊体膜ΔpH增加热耗散(D), 减小光合反应中心P的份额.因此, 春季放牧样地羊草的Pn较小(Demmig-Admas et al., 1996; Ort, 2001).8月, 羊草吸收的光能在能量分配中热耗散(D)所占的份额最大, 表明此时羊草将吸收的光能大多通过热能形式散失.放牧样地羊草叶片水势较高, gs也较高, 植物光合作用过程有足够的CO2供应.同时, 其qp也相对较大, PSII反应中心的开放程度较高, ETR和ΦPSII较大, 即植物吸收的光能进入PSII反应中心的P较多, 电子传递产生的ATP和NADPH也较多, 从而加强了植物对碳的固定和同化作用, 使Pn升高(Cosgrove and Borowitzka, 2010; 贾永霞等, 2011).而类连续放牧样地羊草净光合速率低于春季放牧样地, 可能是由于其叶片水势、qp、ETR和ΦPSII较低所致. ...
1 1981
... 分别于6月和8月放牧前测定叶片气体交换参数的当天上午9:00-11:00, 采集羊草的根、茎和叶, 迅速放入液氮中保存, 带回实验室用于生化指标的测定.丙二醛(malondialdehyde, MDA)含量采用硫代巴比妥酸法测定(Dhindsa and Matowe, 1981).超氧化物歧化酶(superoxide dismutase, SOD)活性采用氮蓝四唑(NBT)法测定(Giannopolitis and Ries, 1977).过氧化氢酶(catalase, CAT)活性采用紫外分光光度法测定(Clairborne, 1985). ...
1 2001
... 放牧是草地利用的基本形式, 也是干扰草地生产力和植被动态最主要的方式之一(侯向阳和徐海红, 2010; Yan et al., 2013).但长期过度放牧会导致植被盖度减小和草地沙化, 使草地生产力持续衰减, 严重影响草地生态系统的结构和功能(Barger et al., 2004; Li et al., 2012; Xu et al., 2014).植物功能性状能够响应生存环境的变化并(或)对生态系统功能有一定影响(Díaz et al., 2001), 是植物在特定环境下生存策略的重要表现, 为预测放牧对生态系统的影响提供了一种有效的工具(Adler et al., 2005; Klimešová et al., 2008; Mooney et al., 2010).因此, 研究放牧利用下植物功能性状的变化具有重要意义. ...
2 2007
... 在特定环境下, 植物功能性状间具有权衡关系(Wright et al., 2004; Lavorel, 2013; Funk and Cornwell, 2013).放牧利用下, 植被盖度减小, 植株整体呈现矮小化, 生产力衰减, 而这些现象是一系列植物功能性状相互权衡综合变化的结果(王炜等, 2000; An and Li, 2014; 李西良等, 2015).目前, 关于植物功能性状权衡方面的研究主要集中于通过软性状(soft traits)来评估植物对环境的响应与适应策略.随着放牧强度增加, 植物表现出株高降低、节间距缩短、叶面积减少和生物量降低的特征(Zhao et al., 2009; Zheng et al., 2011).因为高大、叶片大的植株更易于被啃食, 植株会通过矮化和匍匐生长等一些形态性状变化降低牲畜的选择性(Vesk et al., 2004).与此同时, 植株的分蘖能力会相对增加, 以促进被啃食后的快速生长(Zhao et al., 2009).Fahnestock和Detling (2000)认为, 放牧对优势种的改变主要是对其形态特性改变的结果而不是生理特性的改变.但越来越多的研究表明, 放牧可以影响牧后残留组织光系统II (PSII)活性氧的产生与清除、气体交换速率以及光合产物的积累, 并且大多数硬性状(hard traits)的研究仅停留在叶片水平(Donaghy et al., 2007; Xiu, 2015).植物的软性状与不易测量的硬性状密切相关, 如光合速率可以影响碳水化合物的积累, 从而影响生物量和叶面积, 而株高和叶面积也会影响光合作用中光的捕获(Wang and Wang, 2001; Peng et al., 2007; 晏欣等, 2013).然而这些研究大多基于放牧强度, 对于季节放牧下植株硬性状和软性状的变化及两者权衡关系报道较少. ... ... 可溶性糖是光合作用的直接产物, 是植物生长发育的主要能量来源(Li et al., 1999; 刘浩荣等, 2007; McCormick et al., 2013).放牧干扰影响了植物体内可溶性糖的产生与储存.在牧后恢复期, 植物首先会利用残茬和根中的水溶性碳水化合物, 促进叶片快速再生, 提高植物的光合能力; 其次利用茎和根中储存的能量物质.在6月和8月放牧前, 羊草茎中可溶性糖含量显著高于根, 说明牧后恢复一段时期后, 茎中储存了一定的能量(Donaghy et al., 2007; McCormick et al., 2013).6月, 春季放牧样地羊草茎中可溶性糖含量显著降低, 这是由于春季放牧样地羊草光合器官受损使净光合作用降低所致; 而类连续放牧样地羊草较高的光合作用使其茎中积累了较多的可溶性糖.在季节放牧下, 6月和8月植物叶片水势较低, 因此, 羊草处于干旱和放牧双重胁迫下.在胁迫环境中, 植物会通过提高渗透调节物质(如可溶性蛋白和脯氨酸等) 含量提高渗透势, 增加细胞液浓度以对细胞起到保护作用(Bajji et al., 2001; Matysik et al., 2002; 尹本丰和张元明, 2015).类连续放牧样地羊草叶片脯氨酸和可溶性蛋白含量较少, 而根与茎中含量较高.单立山等(2015)在探讨土壤水分胁迫对红砂(Reaumuria songarica)幼苗生长和渗透调节物质的影响中发现, 土壤水分胁迫下脯氨酸和可溶性蛋白在根中积累较多.这是由于在渗透胁迫发生时, 根比叶片反应更快 (Ogawa and Yamauchi, 2006), 并且植物更倾向于降低生命代谢器官的资源分配率, 将有限的资源尽可能分配到茎和根部来实现更大程度的保水与吸水, 以抵御干旱逆境, 从而维持生存(尹丽等, 2012).本研究中, 春季放牧样地羊草可溶性蛋白和脯氨酸较多地分配在叶片中, 而类连续放牧样地羊草则在根和茎中分配较多.相对于春季放牧样地, 类连续放牧样地羊草渗透调节物质和可溶性糖积累较多, 因此, 在牧后恢复期或降水量较多时, 类连续放牧样地羊草能更快速恢复生长(Shahba and Abbas, 2014). ...
1 1984
... 锡林郭勒盟草原是典型的半干旱草原, 放牧是其传统的土地利用方式.但过度放牧已导致70%的草地处于不同程度的退化中, 大大减少了草地的生产力, 草地退化已经成为重要的经济和生态问题(Tong et al., 2004).优势种是草地生态群落的重要组成部分, 虽然物种数量较少, 但在群落生物量中的分配比例较大, 并在很大程度上决定了草地生态系统的结构和功能(Smith and Knapp, 2003; Bai et al., 2004).羊草(Leymus chinensis)是我国温带草地生态系统的主要优势种, 为多年生禾本科赖草属植物, 草质好、适口性强, 其根茎主要分布在地下10 cm, 根茎发达且繁殖能力强, 有较强的耐牧性(Staalduinen and Anten, 2005).由于草地植物自身具有季节生长节律和再生性(Donald and John, 1984; 李建龙和许鹏, 1991), 传统固定放牧时间长度会导致早期放牧区的可利用牧草的生物量少于后期放牧区, 牧草利用率高, 采食强度大, 放牧后地上现存量低, 不利于草地植物的再生及草地的可持续利用(李勤奋等, 2004; 赵康和宝音陶格涛, 2014; 敖敦高娃和宝音陶格涛, 2015).本研究以优势种羊草为研究对象, 在内蒙古温带典型草原以群落留茬高度6 cm定为终牧期, 并设置了围封、类连续放牧和春季放牧3种处理, 通过测定不同生长季及不同处理下羊草形态和光合生理、抗氧化系统以及渗透调节物等生理特性, 探讨优势种羊草根茎叶功能性状对季节放牧的响应及其权衡.本研究旨在分析优势种对季节放牧的形态软性状的响应及适应机制, 同时探讨羊草叶片光合特性与根茎叶光合产物积累对放牧的响应及适应机制, 以期进一步揭示不同季节放牧下羊草软硬性状的权衡. ...
1 1999
... 植物在不同的环境中生存并与环境相适应, 形成了不同的生理生态学特性(Liang et al., 2010).在放牧干扰下, 植物光合作用、蒸腾作用和暗呼吸发生变化, 体内的营养物质重新分配, 促进了植物的再生长, 进一步提高了牧草的耐牧性(晏欣等, 2013).叶绿素荧光参数可以直接或间接反映光合作用过程、受胁迫时光合能力变化和植物光合器官损伤状况, 因而被广泛地应用于植物光合作用研究中(Earl and Tollenaar, 1999; Larcher, 2003; Zai et al., 2012; 李辉等, 2016). ...
... 在环境胁迫下, 植物PSII吸收的光能不能通过ETR传递出去用于光化学反应(P), 激发态的叶绿素分子会与O2反应, 产生大量的单线态氧, 虽然单线态氧的产生吸收了多余的电子, 减缓了光抑制, 但单线态氧的积累不能及时被SOD及CAT等清除, 也会破坏细胞膜(Christine and Graham, 2000; He et al., 2009; Bencze et al., 2011).MDA是细胞膜不饱和脂肪酸分解的产物, 反映细胞膜受伤害的程度(Zhou and Zhao, 2004).6月, 春季放牧样地羊草MDA含量较高, 表明细胞膜发生了膜脂过氧化, 但此时羊草SOD活性较低, 清除自由基的能力较弱.因此, 叶绿素分子吸收的光能大多用于热耗散(D), 光合速率降低.SOD可以将自由基转化为H2O2, H2O2对植物细胞具有毒害作用, 而CAT可以有效将H2O2转化为H2O和O2 (Bowler et al., 1992; Foyer et al., 1994; 陈思羽等, 2016).8月, 春季放牧样地羊草SOD和CAT活性较高, MDA含量最低, 表明抗氧化酶有效地清除了自由基, 保护细胞膜.类连续放牧样地羊草MDA含量出现降低的趋势, SOD活性显著降低, 但CAT活性明显提高, 表明类连续放牧样地羊草可能是依赖于谷胱甘肽和抗坏血酸等抗氧化剂来清除自由基(Zhang et al., 2015). ...
1 2013
... 在特定环境下, 植物功能性状间具有权衡关系(Wright et al., 2004; Lavorel, 2013; Funk and Cornwell, 2013).放牧利用下, 植被盖度减小, 植株整体呈现矮小化, 生产力衰减, 而这些现象是一系列植物功能性状相互权衡综合变化的结果(王炜等, 2000; An and Li, 2014; 李西良等, 2015).目前, 关于植物功能性状权衡方面的研究主要集中于通过软性状(soft traits)来评估植物对环境的响应与适应策略.随着放牧强度增加, 植物表现出株高降低、节间距缩短、叶面积减少和生物量降低的特征(Zhao et al., 2009; Zheng et al., 2011).因为高大、叶片大的植株更易于被啃食, 植株会通过矮化和匍匐生长等一些形态性状变化降低牲畜的选择性(Vesk et al., 2004).与此同时, 植株的分蘖能力会相对增加, 以促进被啃食后的快速生长(Zhao et al., 2009).Fahnestock和Detling (2000)认为, 放牧对优势种的改变主要是对其形态特性改变的结果而不是生理特性的改变.但越来越多的研究表明, 放牧可以影响牧后残留组织光系统II (PSII)活性氧的产生与清除、气体交换速率以及光合产物的积累, 并且大多数硬性状(hard traits)的研究仅停留在叶片水平(Donaghy et al., 2007; Xiu, 2015).植物的软性状与不易测量的硬性状密切相关, 如光合速率可以影响碳水化合物的积累, 从而影响生物量和叶面积, 而株高和叶面积也会影响光合作用中光的捕获(Wang and Wang, 2001; Peng et al., 2007; 晏欣等, 2013).然而这些研究大多基于放牧强度, 对于季节放牧下植株硬性状和软性状的变化及两者权衡关系报道较少. ...
1 1977
... 分别于6月和8月放牧前测定叶片气体交换参数的当天上午9:00-11:00, 采集羊草的根、茎和叶, 迅速放入液氮中保存, 带回实验室用于生化指标的测定.丙二醛(malondialdehyde, MDA)含量采用硫代巴比妥酸法测定(Dhindsa and Matowe, 1981).超氧化物歧化酶(superoxide dismutase, SOD)活性采用氮蓝四唑(NBT)法测定(Giannopolitis and Ries, 1977).过氧化氢酶(catalase, CAT)活性采用紫外分光光度法测定(Clairborne, 1985). ...
1 2014
... 放牧干扰降低了群落的生物量, 但也增加了群落的生物多样性(Zhao et al., 2009; Zheng et al., 2011).放牧干扰下优势种羊草生物量明显降低, 其相对地上生物量所占比例也明显降低.其原因为羊草草质好, 适口性强, 更易被啃食(Chen et al., 2005).放牧显著降低了羊草的软性状(株高和节间距), 羊草表现出矮小化特征, 这也是一种避牧性策略, 而羊草的矮小化是导致地上生物量和草原生产力降低的直接因素(王炜等, 2000; Zheng et al., 2011; 李西良等, 2014).类连续放牧样地羊草株高较春季放牧样地低, 可能是由于羊草改变了其地上与地下资源的分配比例(尹丽等, 2012).放牧干扰使得地上生物量被啃食, 地下储存减少, 影响春季植物的再生, 从而导致植株矮小(白永飞等, 1996).植物株高不仅与其避牧性机制有关, 还与植物的竞争光能力密切相关(Briske, 1996; Wes- toby, 1998; Cornelissen et al., 2003).春季放牧样地与类连续放牧样地羊草相比有较高的株高和较大的比叶面积, 导致春季放牧样地羊草能接受更多的光能.8月, 放牧干扰下的羊草光合速率高于围封样地.这是由于放牧干扰下, 养分通过粪便重新回到土壤中, 从而使得羊草具有较长的生长周期.在牧后恢复期, 羊草地下根茎系统更倾向于快速消耗土壤中的养分来进行补偿性生长, 也导致放牧样地土壤养分相对较低(Bryant and Klein, 1983; Staalduinen and Anten, 2005).春季放牧样地土壤养分最少, 但在8月该样地羊草比叶面积较大, 叶片较薄.较薄的植物叶片需要较少的光合组织就能增加光合能力, 因此, 在缺乏养分的条件下, 植物往往通过增大比叶面积而实现快速生长(Guo et al., 2014). ...
1 2009
... 在环境胁迫下, 植物PSII吸收的光能不能通过ETR传递出去用于光化学反应(P), 激发态的叶绿素分子会与O2反应, 产生大量的单线态氧, 虽然单线态氧的产生吸收了多余的电子, 减缓了光抑制, 但单线态氧的积累不能及时被SOD及CAT等清除, 也会破坏细胞膜(Christine and Graham, 2000; He et al., 2009; Bencze et al., 2011).MDA是细胞膜不饱和脂肪酸分解的产物, 反映细胞膜受伤害的程度(Zhou and Zhao, 2004).6月, 春季放牧样地羊草MDA含量较高, 表明细胞膜发生了膜脂过氧化, 但此时羊草SOD活性较低, 清除自由基的能力较弱.因此, 叶绿素分子吸收的光能大多用于热耗散(D), 光合速率降低.SOD可以将自由基转化为H2O2, H2O2对植物细胞具有毒害作用, 而CAT可以有效将H2O2转化为H2O和O2 (Bowler et al., 1992; Foyer et al., 1994; 陈思羽等, 2016).8月, 春季放牧样地羊草SOD和CAT活性较高, MDA含量最低, 表明抗氧化酶有效地清除了自由基, 保护细胞膜.类连续放牧样地羊草MDA含量出现降低的趋势, SOD活性显著降低, 但CAT活性明显提高, 表明类连续放牧样地羊草可能是依赖于谷胱甘肽和抗坏血酸等抗氧化剂来清除自由基(Zhang et al., 2015). ...
1 2008
... 放牧是草地利用的基本形式, 也是干扰草地生产力和植被动态最主要的方式之一(侯向阳和徐海红, 2010; Yan et al., 2013).但长期过度放牧会导致植被盖度减小和草地沙化, 使草地生产力持续衰减, 严重影响草地生态系统的结构和功能(Barger et al., 2004; Li et al., 2012; Xu et al., 2014).植物功能性状能够响应生存环境的变化并(或)对生态系统功能有一定影响(Díaz et al., 2001), 是植物在特定环境下生存策略的重要表现, 为预测放牧对生态系统的影响提供了一种有效的工具(Adler et al., 2005; Klimešová et al., 2008; Mooney et al., 2010).因此, 研究放牧利用下植物功能性状的变化具有重要意义. ...
1 2003
... 植物在不同的环境中生存并与环境相适应, 形成了不同的生理生态学特性(Liang et al., 2010).在放牧干扰下, 植物光合作用、蒸腾作用和暗呼吸发生变化, 体内的营养物质重新分配, 促进了植物的再生长, 进一步提高了牧草的耐牧性(晏欣等, 2013).叶绿素荧光参数可以直接或间接反映光合作用过程、受胁迫时光合能力变化和植物光合器官损伤状况, 因而被广泛地应用于植物光合作用研究中(Earl and Tollenaar, 1999; Larcher, 2003; Zai et al., 2012; 李辉等, 2016). ...
1 2013
... 在特定环境下, 植物功能性状间具有权衡关系(Wright et al., 2004; Lavorel, 2013; Funk and Cornwell, 2013).放牧利用下, 植被盖度减小, 植株整体呈现矮小化, 生产力衰减, 而这些现象是一系列植物功能性状相互权衡综合变化的结果(王炜等, 2000; An and Li, 2014; 李西良等, 2015).目前, 关于植物功能性状权衡方面的研究主要集中于通过软性状(soft traits)来评估植物对环境的响应与适应策略.随着放牧强度增加, 植物表现出株高降低、节间距缩短、叶面积减少和生物量降低的特征(Zhao et al., 2009; Zheng et al., 2011).因为高大、叶片大的植株更易于被啃食, 植株会通过矮化和匍匐生长等一些形态性状变化降低牲畜的选择性(Vesk et al., 2004).与此同时, 植株的分蘖能力会相对增加, 以促进被啃食后的快速生长(Zhao et al., 2009).Fahnestock和Detling (2000)认为, 放牧对优势种的改变主要是对其形态特性改变的结果而不是生理特性的改变.但越来越多的研究表明, 放牧可以影响牧后残留组织光系统II (PSII)活性氧的产生与清除、气体交换速率以及光合产物的积累, 并且大多数硬性状(hard traits)的研究仅停留在叶片水平(Donaghy et al., 2007; Xiu, 2015).植物的软性状与不易测量的硬性状密切相关, 如光合速率可以影响碳水化合物的积累, 从而影响生物量和叶面积, 而株高和叶面积也会影响光合作用中光的捕获(Wang and Wang, 2001; Peng et al., 2007; 晏欣等, 2013).然而这些研究大多基于放牧强度, 对于季节放牧下植株硬性状和软性状的变化及两者权衡关系报道较少. ...
1 1999
... 可溶性糖是光合作用的直接产物, 是植物生长发育的主要能量来源(Li et al., 1999; 刘浩荣等, 2007; McCormick et al., 2013).放牧干扰影响了植物体内可溶性糖的产生与储存.在牧后恢复期, 植物首先会利用残茬和根中的水溶性碳水化合物, 促进叶片快速再生, 提高植物的光合能力; 其次利用茎和根中储存的能量物质.在6月和8月放牧前, 羊草茎中可溶性糖含量显著高于根, 说明牧后恢复一段时期后, 茎中储存了一定的能量(Donaghy et al., 2007; McCormick et al., 2013).6月, 春季放牧样地羊草茎中可溶性糖含量显著降低, 这是由于春季放牧样地羊草光合器官受损使净光合作用降低所致; 而类连续放牧样地羊草较高的光合作用使其茎中积累了较多的可溶性糖.在季节放牧下, 6月和8月植物叶片水势较低, 因此, 羊草处于干旱和放牧双重胁迫下.在胁迫环境中, 植物会通过提高渗透调节物质(如可溶性蛋白和脯氨酸等) 含量提高渗透势, 增加细胞液浓度以对细胞起到保护作用(Bajji et al., 2001; Matysik et al., 2002; 尹本丰和张元明, 2015).类连续放牧样地羊草叶片脯氨酸和可溶性蛋白含量较少, 而根与茎中含量较高.单立山等(2015)在探讨土壤水分胁迫对红砂(Reaumuria songarica)幼苗生长和渗透调节物质的影响中发现, 土壤水分胁迫下脯氨酸和可溶性蛋白在根中积累较多.这是由于在渗透胁迫发生时, 根比叶片反应更快 (Ogawa and Yamauchi, 2006), 并且植物更倾向于降低生命代谢器官的资源分配率, 将有限的资源尽可能分配到茎和根部来实现更大程度的保水与吸水, 以抵御干旱逆境, 从而维持生存(尹丽等, 2012).本研究中, 春季放牧样地羊草可溶性蛋白和脯氨酸较多地分配在叶片中, 而类连续放牧样地羊草则在根和茎中分配较多.相对于春季放牧样地, 类连续放牧样地羊草渗透调节物质和可溶性糖积累较多, 因此, 在牧后恢复期或降水量较多时, 类连续放牧样地羊草能更快速恢复生长(Shahba and Abbas, 2014). ...
1 2012
... 放牧是草地利用的基本形式, 也是干扰草地生产力和植被动态最主要的方式之一(侯向阳和徐海红, 2010; Yan et al., 2013).但长期过度放牧会导致植被盖度减小和草地沙化, 使草地生产力持续衰减, 严重影响草地生态系统的结构和功能(Barger et al., 2004; Li et al., 2012; Xu et al., 2014).植物功能性状能够响应生存环境的变化并(或)对生态系统功能有一定影响(Díaz et al., 2001), 是植物在特定环境下生存策略的重要表现, 为预测放牧对生态系统的影响提供了一种有效的工具(Adler et al., 2005; Klimešová et al., 2008; Mooney et al., 2010).因此, 研究放牧利用下植物功能性状的变化具有重要意义. ...
1 2010
... 植物在不同的环境中生存并与环境相适应, 形成了不同的生理生态学特性(Liang et al., 2010).在放牧干扰下, 植物光合作用、蒸腾作用和暗呼吸发生变化, 体内的营养物质重新分配, 促进了植物的再生长, 进一步提高了牧草的耐牧性(晏欣等, 2013).叶绿素荧光参数可以直接或间接反映光合作用过程、受胁迫时光合能力变化和植物光合器官损伤状况, 因而被广泛地应用于植物光合作用研究中(Earl and Tollenaar, 1999; Larcher, 2003; Zai et al., 2012; 李辉等, 2016). ...
1 2002
... 可溶性糖是光合作用的直接产物, 是植物生长发育的主要能量来源(Li et al., 1999; 刘浩荣等, 2007; McCormick et al., 2013).放牧干扰影响了植物体内可溶性糖的产生与储存.在牧后恢复期, 植物首先会利用残茬和根中的水溶性碳水化合物, 促进叶片快速再生, 提高植物的光合能力; 其次利用茎和根中储存的能量物质.在6月和8月放牧前, 羊草茎中可溶性糖含量显著高于根, 说明牧后恢复一段时期后, 茎中储存了一定的能量(Donaghy et al., 2007; McCormick et al., 2013).6月, 春季放牧样地羊草茎中可溶性糖含量显著降低, 这是由于春季放牧样地羊草光合器官受损使净光合作用降低所致; 而类连续放牧样地羊草较高的光合作用使其茎中积累了较多的可溶性糖.在季节放牧下, 6月和8月植物叶片水势较低, 因此, 羊草处于干旱和放牧双重胁迫下.在胁迫环境中, 植物会通过提高渗透调节物质(如可溶性蛋白和脯氨酸等) 含量提高渗透势, 增加细胞液浓度以对细胞起到保护作用(Bajji et al., 2001; Matysik et al., 2002; 尹本丰和张元明, 2015).类连续放牧样地羊草叶片脯氨酸和可溶性蛋白含量较少, 而根与茎中含量较高.单立山等(2015)在探讨土壤水分胁迫对红砂(Reaumuria songarica)幼苗生长和渗透调节物质的影响中发现, 土壤水分胁迫下脯氨酸和可溶性蛋白在根中积累较多.这是由于在渗透胁迫发生时, 根比叶片反应更快 (Ogawa and Yamauchi, 2006), 并且植物更倾向于降低生命代谢器官的资源分配率, 将有限的资源尽可能分配到茎和根部来实现更大程度的保水与吸水, 以抵御干旱逆境, 从而维持生存(尹丽等, 2012).本研究中, 春季放牧样地羊草可溶性蛋白和脯氨酸较多地分配在叶片中, 而类连续放牧样地羊草则在根和茎中分配较多.相对于春季放牧样地, 类连续放牧样地羊草渗透调节物质和可溶性糖积累较多, 因此, 在牧后恢复期或降水量较多时, 类连续放牧样地羊草能更快速恢复生长(Shahba and Abbas, 2014). ...
1 2000
... 于2014年6月和8月放牧前, 在无风晴天的上午9:00- 11:00, 利用LI-6400 (LI-COR, Lincoln, USA)光合测定系统测定气体交换参数.每次测定3-5株, 每株植物测定3-5个长势相近的叶片, 设9次重复.测定指标包括: 净光合速率(net photosynthetic rate, Pn)、蒸腾速率(transpiration rate, Tr)、胞间二氧化碳浓度(inter- cellular CO2 concentration, Ci)、气孔导度(stomatal conductance, gs)以及计算所得水分利用效率(WUE= Pn/Tr).同时测定叶绿素荧光参数, 测前暗适应半小时, 测得最大荧光(maximal fluorescence yield of the dark-adapted state, Fm)和最小荧光(minimal fluorescence yield of the dark-adapted state, F0).在光下Pn稳定后测得光下最大荧光(maximal fluorescence yield of the light-adapted state, Fmʹ)和稳态荧光(steady-state fluorescence yield, Fs).通过计算得到以下参数: 最大光量子效率(Fv/Fm=(Fm-F0)/Fm), PSII激发能捕获效率Fvʹ/Fmʹ=(Fmʹ-F0ʹ)/Fmʹ, 实际量子效率(ΦPSII=(Fmʹ-Fs)/Fmʹ), 光化学猝灭系数(qP=(Fmʹ- Fs)/(Fmʹ-F0ʹ)) (许大全, 2013)和电子传递速率(ETR= ΦPSII×PPFD×0.5×α) (Maxwell and Johnson, 2000), 能量分配计算得光化学反应P=FvʹFmʹ×qpʹ, 热耗散D=1-(Fvʹ/Fmʹ), 激发能E=Fvʹ/Fmʹ×(1-qp) (Demmig- Admas et al., 1996). ...
... 放牧是草地利用的基本形式, 也是干扰草地生产力和植被动态最主要的方式之一(侯向阳和徐海红, 2010; Yan et al., 2013).但长期过度放牧会导致植被盖度减小和草地沙化, 使草地生产力持续衰减, 严重影响草地生态系统的结构和功能(Barger et al., 2004; Li et al., 2012; Xu et al., 2014).植物功能性状能够响应生存环境的变化并(或)对生态系统功能有一定影响(Díaz et al., 2001), 是植物在特定环境下生存策略的重要表现, 为预测放牧对生态系统的影响提供了一种有效的工具(Adler et al., 2005; Klimešová et al., 2008; Mooney et al., 2010).因此, 研究放牧利用下植物功能性状的变化具有重要意义. ...
1 2010
... 由于可利用资源总量有限, 植物会在功能性状之间进行资源优化配置, 即权衡策略(Stearns, 1992; 张大勇, 2004; 陈莹婷和许振柱, 2014).植物根据所处生境的资源条件, 调整、转换或补偿自身功能, 以平衡生存、生长及繁殖三大目的, 最终表现在植物器官的组织结构及其生理性状上(Navas et al., 2010; Whitman and Aarssen, 2010). ...
1 2006
... 可溶性糖是光合作用的直接产物, 是植物生长发育的主要能量来源(Li et al., 1999; 刘浩荣等, 2007; McCormick et al., 2013).放牧干扰影响了植物体内可溶性糖的产生与储存.在牧后恢复期, 植物首先会利用残茬和根中的水溶性碳水化合物, 促进叶片快速再生, 提高植物的光合能力; 其次利用茎和根中储存的能量物质.在6月和8月放牧前, 羊草茎中可溶性糖含量显著高于根, 说明牧后恢复一段时期后, 茎中储存了一定的能量(Donaghy et al., 2007; McCormick et al., 2013).6月, 春季放牧样地羊草茎中可溶性糖含量显著降低, 这是由于春季放牧样地羊草光合器官受损使净光合作用降低所致; 而类连续放牧样地羊草较高的光合作用使其茎中积累了较多的可溶性糖.在季节放牧下, 6月和8月植物叶片水势较低, 因此, 羊草处于干旱和放牧双重胁迫下.在胁迫环境中, 植物会通过提高渗透调节物质(如可溶性蛋白和脯氨酸等) 含量提高渗透势, 增加细胞液浓度以对细胞起到保护作用(Bajji et al., 2001; Matysik et al., 2002; 尹本丰和张元明, 2015).类连续放牧样地羊草叶片脯氨酸和可溶性蛋白含量较少, 而根与茎中含量较高.单立山等(2015)在探讨土壤水分胁迫对红砂(Reaumuria songarica)幼苗生长和渗透调节物质的影响中发现, 土壤水分胁迫下脯氨酸和可溶性蛋白在根中积累较多.这是由于在渗透胁迫发生时, 根比叶片反应更快 (Ogawa and Yamauchi, 2006), 并且植物更倾向于降低生命代谢器官的资源分配率, 将有限的资源尽可能分配到茎和根部来实现更大程度的保水与吸水, 以抵御干旱逆境, 从而维持生存(尹丽等, 2012).本研究中, 春季放牧样地羊草可溶性蛋白和脯氨酸较多地分配在叶片中, 而类连续放牧样地羊草则在根和茎中分配较多.相对于春季放牧样地, 类连续放牧样地羊草渗透调节物质和可溶性糖积累较多, 因此, 在牧后恢复期或降水量较多时, 类连续放牧样地羊草能更快速恢复生长(Shahba and Abbas, 2014). ...
... 锡林郭勒盟草原是典型的半干旱草原, 放牧是其传统的土地利用方式.但过度放牧已导致70%的草地处于不同程度的退化中, 大大减少了草地的生产力, 草地退化已经成为重要的经济和生态问题(Tong et al., 2004).优势种是草地生态群落的重要组成部分, 虽然物种数量较少, 但在群落生物量中的分配比例较大, 并在很大程度上决定了草地生态系统的结构和功能(Smith and Knapp, 2003; Bai et al., 2004).羊草(Leymus chinensis)是我国温带草地生态系统的主要优势种, 为多年生禾本科赖草属植物, 草质好、适口性强, 其根茎主要分布在地下10 cm, 根茎发达且繁殖能力强, 有较强的耐牧性(Staalduinen and Anten, 2005).由于草地植物自身具有季节生长节律和再生性(Donald and John, 1984; 李建龙和许鹏, 1991), 传统固定放牧时间长度会导致早期放牧区的可利用牧草的生物量少于后期放牧区, 牧草利用率高, 采食强度大, 放牧后地上现存量低, 不利于草地植物的再生及草地的可持续利用(李勤奋等, 2004; 赵康和宝音陶格涛, 2014; 敖敦高娃和宝音陶格涛, 2015).本研究以优势种羊草为研究对象, 在内蒙古温带典型草原以群落留茬高度6 cm定为终牧期, 并设置了围封、类连续放牧和春季放牧3种处理, 通过测定不同生长季及不同处理下羊草形态和光合生理、抗氧化系统以及渗透调节物等生理特性, 探讨优势种羊草根茎叶功能性状对季节放牧的响应及其权衡.本研究旨在分析优势种对季节放牧的形态软性状的响应及适应机制, 同时探讨羊草叶片光合特性与根茎叶光合产物积累对放牧的响应及适应机制, 以期进一步揭示不同季节放牧下羊草软硬性状的权衡. ...
1 2004
... 在特定环境下, 植物功能性状间具有权衡关系(Wright et al., 2004; Lavorel, 2013; Funk and Cornwell, 2013).放牧利用下, 植被盖度减小, 植株整体呈现矮小化, 生产力衰减, 而这些现象是一系列植物功能性状相互权衡综合变化的结果(王炜等, 2000; An and Li, 2014; 李西良等, 2015).目前, 关于植物功能性状权衡方面的研究主要集中于通过软性状(soft traits)来评估植物对环境的响应与适应策略.随着放牧强度增加, 植物表现出株高降低、节间距缩短、叶面积减少和生物量降低的特征(Zhao et al., 2009; Zheng et al., 2011).因为高大、叶片大的植株更易于被啃食, 植株会通过矮化和匍匐生长等一些形态性状变化降低牲畜的选择性(Vesk et al., 2004).与此同时, 植株的分蘖能力会相对增加, 以促进被啃食后的快速生长(Zhao et al., 2009).Fahnestock和Detling (2000)认为, 放牧对优势种的改变主要是对其形态特性改变的结果而不是生理特性的改变.但越来越多的研究表明, 放牧可以影响牧后残留组织光系统II (PSII)活性氧的产生与清除、气体交换速率以及光合产物的积累, 并且大多数硬性状(hard traits)的研究仅停留在叶片水平(Donaghy et al., 2007; Xiu, 2015).植物的软性状与不易测量的硬性状密切相关, 如光合速率可以影响碳水化合物的积累, 从而影响生物量和叶面积, 而株高和叶面积也会影响光合作用中光的捕获(Wang and Wang, 2001; Peng et al., 2007; 晏欣等, 2013).然而这些研究大多基于放牧强度, 对于季节放牧下植株硬性状和软性状的变化及两者权衡关系报道较少. ...
1 2001
... 在特定环境下, 植物功能性状间具有权衡关系(Wright et al., 2004; Lavorel, 2013; Funk and Cornwell, 2013).放牧利用下, 植被盖度减小, 植株整体呈现矮小化, 生产力衰减, 而这些现象是一系列植物功能性状相互权衡综合变化的结果(王炜等, 2000; An and Li, 2014; 李西良等, 2015).目前, 关于植物功能性状权衡方面的研究主要集中于通过软性状(soft traits)来评估植物对环境的响应与适应策略.随着放牧强度增加, 植物表现出株高降低、节间距缩短、叶面积减少和生物量降低的特征(Zhao et al., 2009; Zheng et al., 2011).因为高大、叶片大的植株更易于被啃食, 植株会通过矮化和匍匐生长等一些形态性状变化降低牲畜的选择性(Vesk et al., 2004).与此同时, 植株的分蘖能力会相对增加, 以促进被啃食后的快速生长(Zhao et al., 2009).Fahnestock和Detling (2000)认为, 放牧对优势种的改变主要是对其形态特性改变的结果而不是生理特性的改变.但越来越多的研究表明, 放牧可以影响牧后残留组织光系统II (PSII)活性氧的产生与清除、气体交换速率以及光合产物的积累, 并且大多数硬性状(hard traits)的研究仅停留在叶片水平(Donaghy et al., 2007; Xiu, 2015).植物的软性状与不易测量的硬性状密切相关, 如光合速率可以影响碳水化合物的积累, 从而影响生物量和叶面积, 而株高和叶面积也会影响光合作用中光的捕获(Wang and Wang, 2001; Peng et al., 2007; 晏欣等, 2013).然而这些研究大多基于放牧强度, 对于季节放牧下植株硬性状和软性状的变化及两者权衡关系报道较少. ...
1 1998
... 放牧干扰降低了群落的生物量, 但也增加了群落的生物多样性(Zhao et al., 2009; Zheng et al., 2011).放牧干扰下优势种羊草生物量明显降低, 其相对地上生物量所占比例也明显降低.其原因为羊草草质好, 适口性强, 更易被啃食(Chen et al., 2005).放牧显著降低了羊草的软性状(株高和节间距), 羊草表现出矮小化特征, 这也是一种避牧性策略, 而羊草的矮小化是导致地上生物量和草原生产力降低的直接因素(王炜等, 2000; Zheng et al., 2011; 李西良等, 2014).类连续放牧样地羊草株高较春季放牧样地低, 可能是由于羊草改变了其地上与地下资源的分配比例(尹丽等, 2012).放牧干扰使得地上生物量被啃食, 地下储存减少, 影响春季植物的再生, 从而导致植株矮小(白永飞等, 1996).植物株高不仅与其避牧性机制有关, 还与植物的竞争光能力密切相关(Briske, 1996; Wes- toby, 1998; Cornelissen et al., 2003).春季放牧样地与类连续放牧样地羊草相比有较高的株高和较大的比叶面积, 导致春季放牧样地羊草能接受更多的光能.8月, 放牧干扰下的羊草光合速率高于围封样地.这是由于放牧干扰下, 养分通过粪便重新回到土壤中, 从而使得羊草具有较长的生长周期.在牧后恢复期, 羊草地下根茎系统更倾向于快速消耗土壤中的养分来进行补偿性生长, 也导致放牧样地土壤养分相对较低(Bryant and Klein, 1983; Staalduinen and Anten, 2005).春季放牧样地土壤养分最少, 但在8月该样地羊草比叶面积较大, 叶片较薄.较薄的植物叶片需要较少的光合组织就能增加光合能力, 因此, 在缺乏养分的条件下, 植物往往通过增大比叶面积而实现快速生长(Guo et al., 2014). ...
1 2010
... 由于可利用资源总量有限, 植物会在功能性状之间进行资源优化配置, 即权衡策略(Stearns, 1992; 张大勇, 2004; 陈莹婷和许振柱, 2014).植物根据所处生境的资源条件, 调整、转换或补偿自身功能, 以平衡生存、生长及繁殖三大目的, 最终表现在植物器官的组织结构及其生理性状上(Navas et al., 2010; Whitman and Aarssen, 2010). ...
1 2004
... 在特定环境下, 植物功能性状间具有权衡关系(Wright et al., 2004; Lavorel, 2013; Funk and Cornwell, 2013).放牧利用下, 植被盖度减小, 植株整体呈现矮小化, 生产力衰减, 而这些现象是一系列植物功能性状相互权衡综合变化的结果(王炜等, 2000; An and Li, 2014; 李西良等, 2015).目前, 关于植物功能性状权衡方面的研究主要集中于通过软性状(soft traits)来评估植物对环境的响应与适应策略.随着放牧强度增加, 植物表现出株高降低、节间距缩短、叶面积减少和生物量降低的特征(Zhao et al., 2009; Zheng et al., 2011).因为高大、叶片大的植株更易于被啃食, 植株会通过矮化和匍匐生长等一些形态性状变化降低牲畜的选择性(Vesk et al., 2004).与此同时, 植株的分蘖能力会相对增加, 以促进被啃食后的快速生长(Zhao et al., 2009).Fahnestock和Detling (2000)认为, 放牧对优势种的改变主要是对其形态特性改变的结果而不是生理特性的改变.但越来越多的研究表明, 放牧可以影响牧后残留组织光系统II (PSII)活性氧的产生与清除、气体交换速率以及光合产物的积累, 并且大多数硬性状(hard traits)的研究仅停留在叶片水平(Donaghy et al., 2007; Xiu, 2015).植物的软性状与不易测量的硬性状密切相关, 如光合速率可以影响碳水化合物的积累, 从而影响生物量和叶面积, 而株高和叶面积也会影响光合作用中光的捕获(Wang and Wang, 2001; Peng et al., 2007; 晏欣等, 2013).然而这些研究大多基于放牧强度, 对于季节放牧下植株硬性状和软性状的变化及两者权衡关系报道较少. ...
1 2015
... 在特定环境下, 植物功能性状间具有权衡关系(Wright et al., 2004; Lavorel, 2013; Funk and Cornwell, 2013).放牧利用下, 植被盖度减小, 植株整体呈现矮小化, 生产力衰减, 而这些现象是一系列植物功能性状相互权衡综合变化的结果(王炜等, 2000; An and Li, 2014; 李西良等, 2015).目前, 关于植物功能性状权衡方面的研究主要集中于通过软性状(soft traits)来评估植物对环境的响应与适应策略.随着放牧强度增加, 植物表现出株高降低、节间距缩短、叶面积减少和生物量降低的特征(Zhao et al., 2009; Zheng et al., 2011).因为高大、叶片大的植株更易于被啃食, 植株会通过矮化和匍匐生长等一些形态性状变化降低牲畜的选择性(Vesk et al., 2004).与此同时, 植株的分蘖能力会相对增加, 以促进被啃食后的快速生长(Zhao et al., 2009).Fahnestock和Detling (2000)认为, 放牧对优势种的改变主要是对其形态特性改变的结果而不是生理特性的改变.但越来越多的研究表明, 放牧可以影响牧后残留组织光系统II (PSII)活性氧的产生与清除、气体交换速率以及光合产物的积累, 并且大多数硬性状(hard traits)的研究仅停留在叶片水平(Donaghy et al., 2007; Xiu, 2015).植物的软性状与不易测量的硬性状密切相关, 如光合速率可以影响碳水化合物的积累, 从而影响生物量和叶面积, 而株高和叶面积也会影响光合作用中光的捕获(Wang and Wang, 2001; Peng et al., 2007; 晏欣等, 2013).然而这些研究大多基于放牧强度, 对于季节放牧下植株硬性状和软性状的变化及两者权衡关系报道较少. ...
1 2014
... 放牧是草地利用的基本形式, 也是干扰草地生产力和植被动态最主要的方式之一(侯向阳和徐海红, 2010; Yan et al., 2013).但长期过度放牧会导致植被盖度减小和草地沙化, 使草地生产力持续衰减, 严重影响草地生态系统的结构和功能(Barger et al., 2004; Li et al., 2012; Xu et al., 2014).植物功能性状能够响应生存环境的变化并(或)对生态系统功能有一定影响(Díaz et al., 2001), 是植物在特定环境下生存策略的重要表现, 为预测放牧对生态系统的影响提供了一种有效的工具(Adler et al., 2005; Klimešová et al., 2008; Mooney et al., 2010).因此, 研究放牧利用下植物功能性状的变化具有重要意义. ...
1 2013
... 放牧是草地利用的基本形式, 也是干扰草地生产力和植被动态最主要的方式之一(侯向阳和徐海红, 2010; Yan et al., 2013).但长期过度放牧会导致植被盖度减小和草地沙化, 使草地生产力持续衰减, 严重影响草地生态系统的结构和功能(Barger et al., 2004; Li et al., 2012; Xu et al., 2014).植物功能性状能够响应生存环境的变化并(或)对生态系统功能有一定影响(Díaz et al., 2001), 是植物在特定环境下生存策略的重要表现, 为预测放牧对生态系统的影响提供了一种有效的工具(Adler et al., 2005; Klimešová et al., 2008; Mooney et al., 2010).因此, 研究放牧利用下植物功能性状的变化具有重要意义. ...
1 2012
... 植物在不同的环境中生存并与环境相适应, 形成了不同的生理生态学特性(Liang et al., 2010).在放牧干扰下, 植物光合作用、蒸腾作用和暗呼吸发生变化, 体内的营养物质重新分配, 促进了植物的再生长, 进一步提高了牧草的耐牧性(晏欣等, 2013).叶绿素荧光参数可以直接或间接反映光合作用过程、受胁迫时光合能力变化和植物光合器官损伤状况, 因而被广泛地应用于植物光合作用研究中(Earl and Tollenaar, 1999; Larcher, 2003; Zai et al., 2012; 李辉等, 2016). ...
1 2015
... 在环境胁迫下, 植物PSII吸收的光能不能通过ETR传递出去用于光化学反应(P), 激发态的叶绿素分子会与O2反应, 产生大量的单线态氧, 虽然单线态氧的产生吸收了多余的电子, 减缓了光抑制, 但单线态氧的积累不能及时被SOD及CAT等清除, 也会破坏细胞膜(Christine and Graham, 2000; He et al., 2009; Bencze et al., 2011).MDA是细胞膜不饱和脂肪酸分解的产物, 反映细胞膜受伤害的程度(Zhou and Zhao, 2004).6月, 春季放牧样地羊草MDA含量较高, 表明细胞膜发生了膜脂过氧化, 但此时羊草SOD活性较低, 清除自由基的能力较弱.因此, 叶绿素分子吸收的光能大多用于热耗散(D), 光合速率降低.SOD可以将自由基转化为H2O2, H2O2对植物细胞具有毒害作用, 而CAT可以有效将H2O2转化为H2O和O2 (Bowler et al., 1992; Foyer et al., 1994; 陈思羽等, 2016).8月, 春季放牧样地羊草SOD和CAT活性较高, MDA含量最低, 表明抗氧化酶有效地清除了自由基, 保护细胞膜.类连续放牧样地羊草MDA含量出现降低的趋势, SOD活性显著降低, 但CAT活性明显提高, 表明类连续放牧样地羊草可能是依赖于谷胱甘肽和抗坏血酸等抗氧化剂来清除自由基(Zhang et al., 2015). ...
3 2009
... 在特定环境下, 植物功能性状间具有权衡关系(Wright et al., 2004; Lavorel, 2013; Funk and Cornwell, 2013).放牧利用下, 植被盖度减小, 植株整体呈现矮小化, 生产力衰减, 而这些现象是一系列植物功能性状相互权衡综合变化的结果(王炜等, 2000; An and Li, 2014; 李西良等, 2015).目前, 关于植物功能性状权衡方面的研究主要集中于通过软性状(soft traits)来评估植物对环境的响应与适应策略.随着放牧强度增加, 植物表现出株高降低、节间距缩短、叶面积减少和生物量降低的特征(Zhao et al., 2009; Zheng et al., 2011).因为高大、叶片大的植株更易于被啃食, 植株会通过矮化和匍匐生长等一些形态性状变化降低牲畜的选择性(Vesk et al., 2004).与此同时, 植株的分蘖能力会相对增加, 以促进被啃食后的快速生长(Zhao et al., 2009).Fahnestock和Detling (2000)认为, 放牧对优势种的改变主要是对其形态特性改变的结果而不是生理特性的改变.但越来越多的研究表明, 放牧可以影响牧后残留组织光系统II (PSII)活性氧的产生与清除、气体交换速率以及光合产物的积累, 并且大多数硬性状(hard traits)的研究仅停留在叶片水平(Donaghy et al., 2007; Xiu, 2015).植物的软性状与不易测量的硬性状密切相关, 如光合速率可以影响碳水化合物的积累, 从而影响生物量和叶面积, 而株高和叶面积也会影响光合作用中光的捕获(Wang and Wang, 2001; Peng et al., 2007; 晏欣等, 2013).然而这些研究大多基于放牧强度, 对于季节放牧下植株硬性状和软性状的变化及两者权衡关系报道较少. ... ... ).与此同时, 植株的分蘖能力会相对增加, 以促进被啃食后的快速生长(Zhao et al., 2009).Fahnestock和Detling (2000)认为, 放牧对优势种的改变主要是对其形态特性改变的结果而不是生理特性的改变.但越来越多的研究表明, 放牧可以影响牧后残留组织光系统II (PSII)活性氧的产生与清除、气体交换速率以及光合产物的积累, 并且大多数硬性状(hard traits)的研究仅停留在叶片水平(Donaghy et al., 2007; Xiu, 2015).植物的软性状与不易测量的硬性状密切相关, 如光合速率可以影响碳水化合物的积累, 从而影响生物量和叶面积, 而株高和叶面积也会影响光合作用中光的捕获(Wang and Wang, 2001; Peng et al., 2007; 晏欣等, 2013).然而这些研究大多基于放牧强度, 对于季节放牧下植株硬性状和软性状的变化及两者权衡关系报道较少. ... ... 放牧干扰降低了群落的生物量, 但也增加了群落的生物多样性(Zhao et al., 2009; Zheng et al., 2011).放牧干扰下优势种羊草生物量明显降低, 其相对地上生物量所占比例也明显降低.其原因为羊草草质好, 适口性强, 更易被啃食(Chen et al., 2005).放牧显著降低了羊草的软性状(株高和节间距), 羊草表现出矮小化特征, 这也是一种避牧性策略, 而羊草的矮小化是导致地上生物量和草原生产力降低的直接因素(王炜等, 2000; Zheng et al., 2011; 李西良等, 2014).类连续放牧样地羊草株高较春季放牧样地低, 可能是由于羊草改变了其地上与地下资源的分配比例(尹丽等, 2012).放牧干扰使得地上生物量被啃食, 地下储存减少, 影响春季植物的再生, 从而导致植株矮小(白永飞等, 1996).植物株高不仅与其避牧性机制有关, 还与植物的竞争光能力密切相关(Briske, 1996; Wes- toby, 1998; Cornelissen et al., 2003).春季放牧样地与类连续放牧样地羊草相比有较高的株高和较大的比叶面积, 导致春季放牧样地羊草能接受更多的光能.8月, 放牧干扰下的羊草光合速率高于围封样地.这是由于放牧干扰下, 养分通过粪便重新回到土壤中, 从而使得羊草具有较长的生长周期.在牧后恢复期, 羊草地下根茎系统更倾向于快速消耗土壤中的养分来进行补偿性生长, 也导致放牧样地土壤养分相对较低(Bryant and Klein, 1983; Staalduinen and Anten, 2005).春季放牧样地土壤养分最少, 但在8月该样地羊草比叶面积较大, 叶片较薄.较薄的植物叶片需要较少的光合组织就能增加光合能力, 因此, 在缺乏养分的条件下, 植物往往通过增大比叶面积而实现快速生长(Guo et al., 2014). ...
3 2011
... 在特定环境下, 植物功能性状间具有权衡关系(Wright et al., 2004; Lavorel, 2013; Funk and Cornwell, 2013).放牧利用下, 植被盖度减小, 植株整体呈现矮小化, 生产力衰减, 而这些现象是一系列植物功能性状相互权衡综合变化的结果(王炜等, 2000; An and Li, 2014; 李西良等, 2015).目前, 关于植物功能性状权衡方面的研究主要集中于通过软性状(soft traits)来评估植物对环境的响应与适应策略.随着放牧强度增加, 植物表现出株高降低、节间距缩短、叶面积减少和生物量降低的特征(Zhao et al., 2009; Zheng et al., 2011).因为高大、叶片大的植株更易于被啃食, 植株会通过矮化和匍匐生长等一些形态性状变化降低牲畜的选择性(Vesk et al., 2004).与此同时, 植株的分蘖能力会相对增加, 以促进被啃食后的快速生长(Zhao et al., 2009).Fahnestock和Detling (2000)认为, 放牧对优势种的改变主要是对其形态特性改变的结果而不是生理特性的改变.但越来越多的研究表明, 放牧可以影响牧后残留组织光系统II (PSII)活性氧的产生与清除、气体交换速率以及光合产物的积累, 并且大多数硬性状(hard traits)的研究仅停留在叶片水平(Donaghy et al., 2007; Xiu, 2015).植物的软性状与不易测量的硬性状密切相关, 如光合速率可以影响碳水化合物的积累, 从而影响生物量和叶面积, 而株高和叶面积也会影响光合作用中光的捕获(Wang and Wang, 2001; Peng et al., 2007; 晏欣等, 2013).然而这些研究大多基于放牧强度, 对于季节放牧下植株硬性状和软性状的变化及两者权衡关系报道较少. ... ... 放牧干扰降低了群落的生物量, 但也增加了群落的生物多样性(Zhao et al., 2009; Zheng et al., 2011).放牧干扰下优势种羊草生物量明显降低, 其相对地上生物量所占比例也明显降低.其原因为羊草草质好, 适口性强, 更易被啃食(Chen et al., 2005).放牧显著降低了羊草的软性状(株高和节间距), 羊草表现出矮小化特征, 这也是一种避牧性策略, 而羊草的矮小化是导致地上生物量和草原生产力降低的直接因素(王炜等, 2000; Zheng et al., 2011; 李西良等, 2014).类连续放牧样地羊草株高较春季放牧样地低, 可能是由于羊草改变了其地上与地下资源的分配比例(尹丽等, 2012).放牧干扰使得地上生物量被啃食, 地下储存减少, 影响春季植物的再生, 从而导致植株矮小(白永飞等, 1996).植物株高不仅与其避牧性机制有关, 还与植物的竞争光能力密切相关(Briske, 1996; Wes- toby, 1998; Cornelissen et al., 2003).春季放牧样地与类连续放牧样地羊草相比有较高的株高和较大的比叶面积, 导致春季放牧样地羊草能接受更多的光能.8月, 放牧干扰下的羊草光合速率高于围封样地.这是由于放牧干扰下, 养分通过粪便重新回到土壤中, 从而使得羊草具有较长的生长周期.在牧后恢复期, 羊草地下根茎系统更倾向于快速消耗土壤中的养分来进行补偿性生长, 也导致放牧样地土壤养分相对较低(Bryant and Klein, 1983; Staalduinen and Anten, 2005).春季放牧样地土壤养分最少, 但在8月该样地羊草比叶面积较大, 叶片较薄.较薄的植物叶片需要较少的光合组织就能增加光合能力, 因此, 在缺乏养分的条件下, 植物往往通过增大比叶面积而实现快速生长(Guo et al., 2014). ... ... ; Zheng et al., 2011; 李西良等, 2014).类连续放牧样地羊草株高较春季放牧样地低, 可能是由于羊草改变了其地上与地下资源的分配比例(尹丽等, 2012).放牧干扰使得地上生物量被啃食, 地下储存减少, 影响春季植物的再生, 从而导致植株矮小(白永飞等, 1996).植物株高不仅与其避牧性机制有关, 还与植物的竞争光能力密切相关(Briske, 1996; Wes- toby, 1998; Cornelissen et al., 2003).春季放牧样地与类连续放牧样地羊草相比有较高的株高和较大的比叶面积, 导致春季放牧样地羊草能接受更多的光能.8月, 放牧干扰下的羊草光合速率高于围封样地.这是由于放牧干扰下, 养分通过粪便重新回到土壤中, 从而使得羊草具有较长的生长周期.在牧后恢复期, 羊草地下根茎系统更倾向于快速消耗土壤中的养分来进行补偿性生长, 也导致放牧样地土壤养分相对较低(Bryant and Klein, 1983; Staalduinen and Anten, 2005).春季放牧样地土壤养分最少, 但在8月该样地羊草比叶面积较大, 叶片较薄.较薄的植物叶片需要较少的光合组织就能增加光合能力, 因此, 在缺乏养分的条件下, 植物往往通过增大比叶面积而实现快速生长(Guo et al., 2014). ...
2 2004
... 在环境胁迫下, 植物PSII吸收的光能不能通过ETR传递出去用于光化学反应(P), 激发态的叶绿素分子会与O2反应, 产生大量的单线态氧, 虽然单线态氧的产生吸收了多余的电子, 减缓了光抑制, 但单线态氧的积累不能及时被SOD及CAT等清除, 也会破坏细胞膜(Christine and Graham, 2000; He et al., 2009; Bencze et al., 2011).MDA是细胞膜不饱和脂肪酸分解的产物, 反映细胞膜受伤害的程度(Zhou and Zhao, 2004).6月, 春季放牧样地羊草MDA含量较高, 表明细胞膜发生了膜脂过氧化, 但此时羊草SOD活性较低, 清除自由基的能力较弱.因此, 叶绿素分子吸收的光能大多用于热耗散(D), 光合速率降低.SOD可以将自由基转化为H2O2, H2O2对植物细胞具有毒害作用, 而CAT可以有效将H2O2转化为H2O和O2 (Bowler et al., 1992; Foyer et al., 1994; 陈思羽等, 2016).8月, 春季放牧样地羊草SOD和CAT活性较高, MDA含量最低, 表明抗氧化酶有效地清除了自由基, 保护细胞膜.类连续放牧样地羊草MDA含量出现降低的趋势, SOD活性显著降低, 但CAT活性明显提高, 表明类连续放牧样地羊草可能是依赖于谷胱甘肽和抗坏血酸等抗氧化剂来清除自由基(Zhang et al., 2015). ... ... 在环境胁迫下, 光合作用、能量分配及抗氧化系统之间的动态平衡被打破, 植物重新进行资源分配, 以保证自身生长发育, 达到一种经济策略.在6月, 类连续放牧样地羊草ETR减小, P分配较少, 这可能是导致Pn降低的主要原因(Cosgrove and Borowitzka, 2010).Pn的降低直接导致羊草叶片中可溶性糖含量较少, 但其茎中储存了大量的可溶性糖, 可溶性蛋白和脯氨酸也较多分配于根与茎中(McCormick et al., 2013).因此, 此时类连续放牧样地羊草通过降低生命代谢器官的资源分配率, 将有限的资源充分分配到茎和根部来实现更大程度的保水、吸水, 以抵御干旱逆境, 有利于羊草牧后再生(尹丽等, 2012).春季放牧样地羊草Fv/Fm比值较低, MDA含量较高, 表明激活的叶绿素分子与氧气反应, 产生大量自由基, 对细胞膜造成了损伤, 导致其Pn降低(Zhou et al., 2004; 张仁和等, 2011).为了减少自由基的产生, 羊草将吸收的光能较多地用于热耗散(D), 而大量积累的自由基也促使CAT活性提高, 以保护植物细胞, 达到一个新的平衡状态.此时, 羊草的光合产物在叶片积累多, 根、茎较少, 不利于羊草根、茎吸水, 也是导致其叶片水势较低的原因, 这都不利于有机物质的储存与羊草牧后再生.8月, 围封样地羊草的可溶性糖含量较高, 但Pn较低, 表明可能是由于可溶性糖的大量积累以负反馈的形式使Pn降低(白永飞和徐志信, 1994; Poot and Lambers, 2003; 刘浩荣等, 2007).类连续放牧样地羊草净光合速率有所升高, 且其MDA含量有所降低, 是由于其CAT活性较高, 此时羊草吸收的光能较多用于光化学反应(P), 减少了活性氧的产生.与6月相似, 羊草将较多的光合产物及渗透调节物质分配于根和茎中.春季放牧样地羊草SOD和CAT活性较高, MDA含量最低, 表明抗氧化酶有效清除了自由基, 且Fv/Fm在正常范围, 细胞膜未受到伤害.PSII吸收的光能较多地用于光化学反应, ETR较高, 导致其Pn较高.其光合直接产物、可溶性蛋白和脯氨酸分配于叶片相对较多, 这与6月的情况类似. ...
 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}