郭金 ,
杨小艳 ,
邓洪平 * , ,
黄琴 ,
李运婷 ,
张华雨 西南大学生命科学学院, 三峡库区生态环境教育部重点实验室, 重庆 400715
Guo Jin ,
Yang Xiaoyan ,
Deng Hongping * ,
Huang Qin ,
Li Yunting ,
Zhang Huayu Key Laboratory of Eco-environment in Three Gorges Reservoir Region (Ministry of Education), School of Life Sciences, Southwest University, Chongqing 400715, China 引用本文 郭金 ,
杨小艳 ,
邓洪平 ,
黄琴 ,
李运婷 ,
张华雨 . 细枝柃性别表达特性及资源分配. , 2017, 52(2): 202-209
贡献者
* 通讯作者。E-mail: denghp@swu.edu.cn 基金资助
科技部国家科技基础平台国家标本平台-教学标本子平台运行服务项目(No.2005DKA21403-JK)和国家自然基金(No.31600491) ;
接受日期:2016-01-13接受日期:2016-09-4网络出版日期:2017-04-10
-->Copyright
20172010 《植物学报》编辑部
History
Received :
Accepted :
Online :
摘要: 已有的资料将柃木属(Eurya )描述为严格的雌雄异株植物, 性别变异现象极为少见。目前仅在柃木(E. japonica )和钝叶柃(E. obtusifolia )等少数种类中报道过两性花的存在。近几年笔者发现细枝柃(E. loquaiana )存在性别变异现象, 性别变异株上具有不同性别类型的花。该文从单花和植株水平分析了细枝柃的性别表达特性, 并对不同类型花的花部构件生物量分配进行比较分析。结果表明, 细枝柃具有6种类型的花, 从单花水平上看, 细枝柃性别有雌性、雄性及两性3种类型; 细枝柃性别在植株水平上体现较为复杂, 有雌株, 雄株, 雌花和两性花同株, 雄花和两性花同株, 雌雄异花同株及雌花、雄花、两性花同株6种类型; 在细枝柃花部构件生物量分配中, 雄花(包括雄株花和变异株雄花)花部构件生物量分配中雄蕊生物量的分配低于雌花(包括雌株花和变异株雌花)中雌蕊生物量的分配; 两性花中, 雄蕊生物量分配低于雌蕊, 这是其优化资源分配的手段, 进而获取最大适合度收益。 关键词 :
细枝柃 ;
性别表达 ;
资源分配 Abstract :
The genus of Eurya Thunb. was described as strictly dioecious, and the gender variation was extremely rare. Until now, bisexual flowers had only been reported in a few species such as E. japonica and E. obtusifolia . We recently found gender variation in E. loquaiana with different types of flowers. Here, we analyzed sex expression characteristics at the flower and plant levels and compared reproduction allocation between different kinds of flowers. E. loquaiana had six types of flowers: pistillate flower, staminate flower, pistillate flower with staminode, staminate flower with pistillode, hermaphrodite flower, and sterile flower. At the flower level, the genders of E. loquaiana were female, male, and bisexual; the gender at the plant level was complicated. E. loquaiana had six gender types: gynoecius and roesious, gynomonoecy, and romonoecy, monoecious and trimonoecious; Stamen biomass allocation was less in male flowers (including the male plant flowers and gender variant plants) than pistil biomass allocation in female flowers (including the female plant and gender variant plants). In bisexual flowers, the stamen biomass allocation was less than the pistil biomass allocation. This is a means to optimize its resource allocation and thus obtain the most fitness benefits. Key words :
Eurya loquaiana ;
sex expression ;
resource allocation 植物的性别通常受遗传控制(
Meagher, 1988 ;
Dellaporta and Urrea, 1993 )。植物的性别分化可能发生于花发育的早期或后期(
Caporali et al., 1994 ;
Hormaza and Polito, 1996 ), 当性别分化发生相对 较晚时, 其它性别的性器官可能会有表达(
Mayer and Charlesworth, 1991 )。然而, 在完全雌雄异株物种的自然种群中, 两性个体较为罕见。Rottenberg (1998)在调查以色列41种雌雄异株植物的12个种群时, 发现它们之中出现两性个体的概率低于0.1%。在某些雌雄异株植物中偶尔会出现相对性别特征的现象, 称为性别变异(gender variation)。常用leaky dioecy (
Humeau et al.,1999 ;
Venkatasamy et al., 2007 )、inconstancy (
Sarkissian et al., 2001 )、exceptional bisexual individual (
Rottenberg, 2000 )、subdioecy (
Hader and Barrett, 2006 )、near-dioecy (
Solomon, 1985 )及paradioecy (
Standley, 1985 )等词描述出现性别变异现象的雌雄异株植物或种群。
柃木属(
Eurya )隶属山茶科(Theaceae), 约130种, 是Thunberg于1783年根据日本产的
Eurya japonica 为模式创立的新属(
林来官, 1998 )。已有的研究资料将柃木属描述为严格的雌雄异株植物, 性别变异现象极为少见, 偶有关于柃木属植物性别变异现象的报道。Vesque (1895)发现柃木属存在性别变异现象。日本****Murata (1991)发现柃木(
E. japonica )中存在两性花。王茜等(2012)在调查中发现钝叶柃(
E. obtusifolia )中存在性别变异现象, 并对其不同性别花的花部形态与传粉特征进行了研究。 近几年, 笔者通过野外调查发现细枝柃(
E. loqu- aiana )中存在性别变异现象, 性别变异株上具有不同性别类型的花。为探究细枝柃的性别表达特性, 本研究从单花和植株两个层面上对其性别表达进行探讨, 并分析花部构件的资源分配差异, 以揭示性别差异的生态适应机制。同时依据性别变异株上花的类型初步探讨柃木属性别进化的途径, 为柃木属的性别演化规律研究提供基础资料。
1 材料与方法1.1 研究材料以重庆市分布的山茶科柃木属植物细枝柃(
Eurya loquaiana Dunn.)为研究对象, 其形态特征与《中国植物志》及地方植物志(
熊济华, 1990 ;
陈封怀, 1991 )中的描述基本一致, 但部分植株表现出明显的性别变异现象, 即性别表达偏离正常的雌株和雄株。在缙云山调查的300余株细枝柃中, 性别变异株的比例达23.08%。
1.2 研究地点2013-2014年间, 对重庆市北碚区、大足区、江津区、涪陵区、巴南区、丰都县及城口县的7个研究样点进行野外调查, 仅在缙云山发现存在性别变异的细枝柃植株, 故将研究地点设在缙云山。该地区具有亚热带季风湿润气候特征, 年平均气温13.6°C。结合调查区实际情况, 实验共设4个20 m×20 m的样地(
表1 ), 主要位于针阔混交林的林中或林缘。
表1 Table 1 表1 表1 缙云山细枝柃性别变异株的分布
Table 1 Distribution of sexual variant plants of
Eurya loquaiana at Jinyun Mountain
Site No. Geographic (N) Coordinates (E) Alt. (m) SM1 29°49′48″ 106°22′44″ 859 SM2 29°50′09″ 106°22′41″ 753 SM3 29°50′05″ 106°22′36″ 753 FX1 29°49′37″ 106°23′31″ 890
表1
缙云山细枝柃性别变异株的分布
Table 1
Distribution of sexual variant plants of
Eurya loquaiana at Jinyun Mountain
1.3 方法1.3.1 性别变异株的调查与标记
2013年11月初于细枝柃花期, 对缙云山的细枝柃花性别进行初步确定。观察方法: 从每株树东、西、南、北4个方向随机抽取树冠中的20朵花, 进行形态学观察并做好标记。
1.3.2 取样
对所标记的变异株进行观察记录, 取回新鲜花朵及待开花蕾置于冰盒内, 带回实验室于NikonSM21000体视镜下解剖并做详细观察记录。部分样品用FAA固定液长期保存。与此同时, 以相同方法分别取回雌株和雄株样品作比较分析。
1.3.3 花性别的确定
分别解剖从雌株、雄株及变异株上取回的样品, 观察其花部结构, 特别注意生殖构件的结构与发育程度。若雌蕊在形态上完善, 柱头具有可授性, 子房内具正常胚珠, 则视为具有雌性功能; 若雄蕊在形态上完善, 花粉粒具有活力, 则视为具有雄性功能。
柱头可授性的检测采用联苯胺-过氧化氢法(
张志良等, 2009 )。花粉活力的测定采用TTC (2, 3, 5-氯化三苯基四氮唑)法(
张志良等, 2009 )。
1.3.4 植株性别的确定
变异株性别功能的确定采用
Lloyd等(1980) 的方法。用以下公式对性别进行量化:
G i=
g i/(
g i+
a i)
其中,
G i表示雌属性,
g i表示具胚珠的花数,
a i表示能产生花粉的花数, 雄属性
A i=1-
G i。
G i>0.5则表示雌属性大于雄属性。
1.3.5 性别资源投资
在细枝柃开花高峰期, 从4个样地随机选取雌株、雄株及性别变异株各5株, 分别进行编号。从每个植株上随机选取20朵花, 分别测量其花冠大小、雌蕊及雄蕊长度, 以研究雌雄蕊大小与生物量分配间的相关性。然后将花部结构分为雌蕊、雄蕊以及其余部分, 根据植株性别进行详细编号, 放入80°C烘箱烘至干重不再变化后称重, 用于分析花部构件的生物量分配。
2 结果与讨论2.1 性别类型细枝柃具有6种类型的花(附录1): 雌花、雄花、具退化雌蕊的雄花、具退化雄蕊的雌花、两性花以及不育花, 其中以雌花、雄花和两性花类型较多, 各花的特征比较见
表2 。在细枝柃6种类型的花中, 雌花和具退化雄蕊的雌花具有雌性功能; 雄花及具退化雌蕊的雄花具有雄性功能; 两性花具两性功能; 不育花无性别功能。
表2 Table 2 表2 表2 细枝柃不同性别植株上花的特征
Table 2 The flower characteristics in different gender plants of
Eurya loquaiana Flower type Petal Style Ovary Ovule number Stamens number Anther Filament The flower of the female plant The brim coils instead outwards 3 gap in general Peak green, 21-45 - - - The pistillate flower of the gender variant plant The brim coils instead outwards 1-4 gap Pink with chalky white, peak green,1-3 loculus 12-27 - - - The pistillate flower with staminode The brim coils instead outwards slightly 2-3 gap, 4 gap rarely Pink or peak green,1-3 loculus 1-30 1-8 Similar to anther spots, shrinking, no pollen Filiform, thre- adiness The hermaphrodite flower The brim coils instead outwards slightly 1-3 gap Pink or peak green, 1-3 loculus 1-29 5-10 Linear-hastate, orange Tenuous The flower of the male plant No crolling - - - 8-16 Linear-hastate, orange Tenuous The staminate flower of the gender variant plant No crolling - - - 8-10 Linear-hastate, orange Tenuous The staminate flower with pistillode The brim coils instead outwards slightly Filiform Growing No 6-12 Linear-hastate, orange Tenuous The sterile flower The brim coils instead outwards slightly Filiform, 1-2 gap No growth No 1-6 Similar to anther spots, shrinking, no pollen Filiform, threadiness
表2
细枝柃不同性别植株上花的特征
Table 2
The flower characteristics in different gender plants of
Eurya loquaiana 2.2 缙云山细枝柃性别变异株的分布2013-2014年间, 采用样线法对缙云山细枝柃群落展开调查, 共调查细枝柃300余株, 发现性别变异株41株(
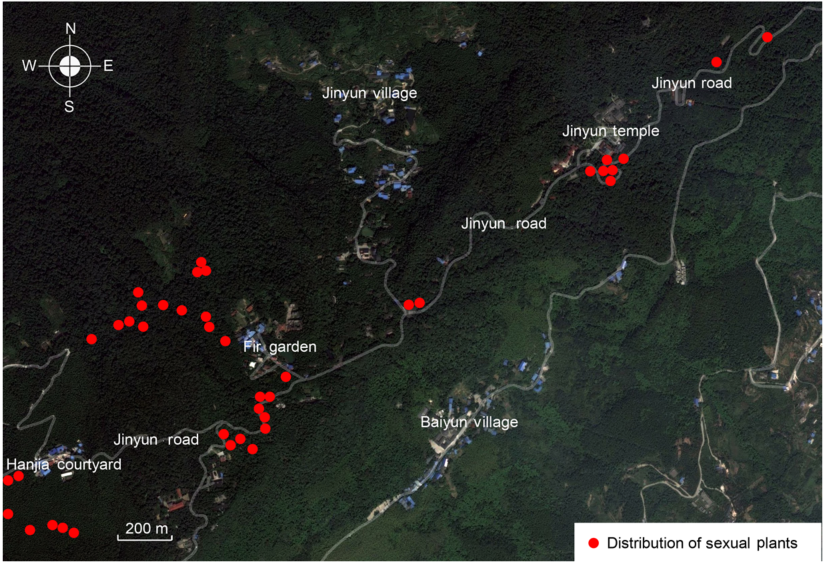
图1 ; 附录2, 附录3), 主要集中分布于沙木园和复兴寺两地, 部分变异株零散分布于竹楼至三角花园段缙云路旁。其中, 杉木园分布24株, 复兴寺分布7株, 竹楼至三角花园段缙云路旁分布10株。这些性别变异株大多混生于雌、雄株之间, 它们之间可能存在着基因交流。缙云山细枝柃性别变异株不仅分布集中而且变异比例较大, 在所设4个集中分布地20 m×20 m样方中, 性别变异株比例可达23.08%。
图1 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-202/img_1.png <b>图1</b> 缙云山细枝柃性别变异株分布信息<br/><b>Figure 1</b> Distribution of sexual plants of <i>Eurya loquaiana</i> at Jinyun Mountain Figure 1 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-202/img_1.png <b>图1</b> 缙云山细枝柃性别变异株分布信息<br/><b>Figure 1</b> Distribution of sexual plants of <i>Eurya loquaiana</i> at Jinyun Mountain 在新窗口打开 下载 下载原图ZIP 生成PPT 图1
缙云山细枝柃性别变异株分布信息
Figure 1
Distribution of sexual plants of
Eurya loquaiana at Jinyun Mountain
2.3 变异株的性别表达细枝柃性别变异株在形态上与雌株、雄株有着很多共同的特征(
表2 ), 但并不是所有变异株都能够同时具有雌性和雄性功能。因性别变异株常在同一植株内开出多种不同类型的花, 故其性别表达比较复杂。为确定变异株的性别类型, 对性别变异株上的花进行取样观察。最后, 确定细枝柃性别在植株水平上共有6种类型: 雌株, 雄株, 雌花、两性花同株, 雄花、两性花同株, 雌雄异花同株, 雌花、雄花及两性花同株。其中, 以雌花、两性花同株类型最多, 为14株, 占总株数的34.51%。
2.4 性别资源分配植物固着生长的习性决定其不能主动地寻找配偶, 为能在不同的环境条件下实现雌雄配子的最大适合度, 植物在长期的进化过程中演化出类型多样的繁殖系统和繁殖策略(
Barrett, 2000 )。植物在生活史中, 通过资源的最佳分配格局并凭借特有的繁殖属性来适应特定环境, 以提高物种适合度(
Eckert and Barrett, 1994 ;
傅国立, 2000 )。根据资源利用假说, 植物可通过调控分配给繁殖构件和营养构件的相对资源比例来保障繁殖成功(
Charlesworth and Charlesworth, 1987 ;
Zhang and Jiang, 2000 )。再分配理论指出, 资源分配模式的改变能够促进雌雄异株植物的进化(
Givnish, 1980 )。研究结果表明, 细枝柃不同类型花的雌、雄蕊特征存在一定差异(
表3 )。变异株雄花的雄蕊显著小于雄株花与性别变异株两性花, 变异株两性花雄蕊小于雄株花。变异株雌花的雌蕊显著小于雌株花与性别变异株两性花, 变异株两性花雌蕊小于雌株花, 但差异不明显。
细枝柃花部构件各生物量见
表4 。结果表明, 雄株花总生物量均显著高于变异株花与雌株花, 变异株上雄花生物量显著高于两性花与雌花。不同类型植株, 雄花生物量大于雌株花与变异株花, 两性花生物量介于两者之间。性别变异株上雄花生物量最大, 雌花生物量最小, 两性花介于两者之间。
细枝柃花部构件各生物量的分配见
表5 。结果表明, 雌株上雌花与性别变异株上的雌花雌蕊生物量的分配均高于两性花, 雌株花又高于变异株雌花; 雄株上雄花与性别变异株上雄花雄蕊生物量的分配亦均高于两性花, 且雄株花高于变异株雄花。
表3 Table 3 表3 表3 细枝柃花部雌雄蕊的基本特征
Table 3 Pistil and stamen basic characteristics of
Eurya loquaiana Flower type Pistil length (mm) Ovary size Stamen length Anther size (length×width) The flower of the female plant 4.77±0.37 1.32±0.11×1.10±0.07 / / The flower of the male plant / / 3.48±0.21 1.28±0.18×0.55±0.10 The pistillate flower of the gender variant plant 4.24±0.28* 1.38±0.16×0.92±0.08* / / The staminate flower of the gender variant plant / / 2.8±0.27* 0.93±0.15×0.45±0.08* The hermaphrodite flower of the gender variant plant 4.38±0.47 1.51±0.58×0.95±0.41 3.01±0.31 1.14±0.18×0.50±0.12
* Significant differences at P <0.05; / indicate null of data (the same as below). * 差异显著(P <0.05); / 表示没有数据(下表同)。 表3
细枝柃花部雌雄蕊的基本特征
Table 3
Pistil and stamen basic characteristics of
Eurya loquaiana 表4 Table 4 表4 表4 细枝柃花部各构件生物量
Table 4 Module biomass of
Eurya loquaiana Flower type The biomass of pistil -1 ) The biomass of -1 ) The biomass of the rest-1 ) The total biomass -1 ) The flower of the female plant 0.0349 / 0.1815 0.2164 The flower of the male plant / 0.0503 0.2948 0.3451 The pistillate flower of the gender variant plant / 0.0226 0.1775 0.2001 The staminate flower of the gender variant plant 0.0182 / 0.1009 0.1191 The hermaphrodite flower of the gender variant plant 0.0171 0.0155 0.1357 0.1683
表4
细枝柃花部各构件生物量
Table 4
Module biomass of
Eurya loquaiana 表5 Table 5 表5 表5 细枝柃花部各构件生物量分配
Table 5 Module biomass allocation of
Eurya loquaiana Flower type The ratio of pistil (%) The ratio of stamen (%) The flower of the female plant 16.13 / The flower of the male plant / 14.58 The pistillate flower of the gender variant plant / 11.29 The staminate flower of the gender variant plant 15.28 / The hermaphrodite flower of the gender variant plant 10.16 9.21
表5
细枝柃花部各构件生物量分配
Table 5
Module biomass allocation of
Eurya loquaiana 2.5 讨论与结论2.5.1 性别表达规律
有关细枝柃的性别, 植物志记载为雌雄异株, 但笔者发现细枝柃植株花具有各种性别类型。总体而言, 细枝柃的性别表达规律具有以下几个特点。
(1) 细枝柃具有6种类型的花: 雌花、雄花、具退化雌蕊的雄花、具退化雄蕊的雌花、两性花及不育花。从单花水平上看, 细枝柃性别有雌性、雄性及两性3种类型。
(2) 细枝柃性别在植株水平上体现较为复杂, 有雌株, 雄株, 雌花、两性花同株, 雄花、两性花同株,雌雄异花同株, 雌花、雄花、两性花同株共6种类型。
(3) 细枝柃野外种群中, 性别变异株相对较少, 以典型的雌株和雄株为主要组成部分。不同种群有着不同的性别组合方式, 无性别变异株的种群, 其性别可直接描述为雌雄异株; 含性别变异株的种群, 性别表达十分复杂, 较难进行描述。
(4) 在已发现的41株变异株中, 未发现单纯的两性花植株, 具有两性功能的植株主要是因为同时具有雌花、两性花或雄花、两性花或雌花、雄花、两性花。
细枝柃的性别表达特性, 是细枝柃性别决定的外在体现及其性别进化的结果。目前, 已在10种柃木属植物中发现性别变异, 有7种为林来官在整理标本时发现的, 但未进行详尽描述(
王茜等, 2012 )。Murata (1991)对柃木中出现的性别变异进行了详细描述。王茜(2012)对钝叶柃性别变异性别表达特性进行了描述。本研究的观察结果与他们的描述有许多相似之处。
2.5.2 不同类型花的生殖构件特征
细枝柃不同类型花的生殖构件特征存在以下差异。
(1) 雄株花生物量明显大于雌株花和变异株花, 雄花(包括雄株花和变异株雄花)中雄蕊的生物量分配量低于雌花(包括雌株花和变异株雌花)中雌蕊的生物量分配。
(2) 雄株花雄蕊大小与生物量均大于变异株雄花和两性花的相应指标; 雌株花雌蕊大小与生物量均大于变异株雌花和两性花的相应指标。
(3) 在性别变异株上, 雄花生物量最大, 雌花最小, 两性花介于两者之间。两性花中, 雄蕊生物量分配低于雌蕊生物量分配, 但差异不明显。
这些差异表明, 在细枝柃花期, 雄株用于生殖的资源投资大于雌株。雌株在此时投入给生殖的资源弱于雄株。若投入给花瓣较多的资源, 收益不大, 因雌株在后期孕育果实和种子时需要投资大量的能量。为达到资源的优化配置, 雌株将在花冠上的投资节约下来, 以孕育果实和种子(
Miller and Venable, 2003 ;
张大勇, 2004 )。在细枝柃各花部构件生物量分配中, 雄花(包括雄株花和变异株雄花)花部构件生物量分配中雄蕊生物量的分配低于雌花(包括雌株花和变异株雌花)中雌蕊生物量的分配; 两性花中, 雄蕊生物量分配低于雌蕊生物量分配, 这是其优化资源分配的手段, 进而获取最大适合度收益。
2.5.3 性别变异株上花的变化过程
从细枝柃性别变异株上各花的形态与性别功能, 可推测变异株上花的变化过程为: 雌花→具退化雄蕊的雌花→两性花→具退化雌蕊的雄花→雄花。根据目前已经得到证实的5条由两性状态向雌雄异株进化的途径(
张大勇, 2004 ), 推测柃木属植物向雌雄异株进化需经历雌全株阶段。在本研究所调查的41株变异株中, 以雌全株比例最多, 与该推断相符。有研究证明, 在干旱胁迫下, 雌雄异株植物种群内的性别格局会发生改变, 种群中的雄株数会增加, 致使雌雄比例下降(
Borthwick and Scully, 1954 ;
McArthur, 1977 ;
Dafni and Shmida, 2002 ;
Nanami et al., 2004 ;
Adam et al., 2011 )。细枝柃性别变异株上花的变化过程预示着变异株有转变为雄株的趋势, 若变化稳定持续, 变异株将会转变为雄株, 最终雄株数量增加, 雌雄比例下降, 但这一结论需经连续多年的观察证实。
植物的性别表达是遗传物质和环境因子共同作用的结果。与遗传控制不同, 环境因子可以通过调控特异基因的表达来诱导植物性别的转换(
Solomon, 1985 )。在苔藓、蕨类和种子植物类群中, 关于环境改变导致植物性别发生转变已有报道(
Golenberg and West, 2013 )。被子植物中, 性别转变现象发生于雌雄异株与雌雄异花同株植物类群的概率大于雌雄同株植物类群(
Korpelainen and Labile, 1998 )。植物雌雄个体或雌雄器官之间存在着深刻的代谢差异, 环境条件的改变使植物的代谢途径或强度发生朝另一种性别发生方向改变(
曹宗巽, 1965 )。基因突变导致机体不适或退化及种群资源限制也能使植物性别发生变异(
向远寅, 1984 )。本研究中, 缙云山细枝柃种群中, 雌株占优势, 雌性性别有向雄性性别转化的趋势, 因此推测其性别发生变异可能是资源限制所致。缙云山气候环境特殊, 细枝柃花期常逢阴雨, 传粉效率受影响。根据种群繁殖理论, 细枝柃为扩大种群, 需调整雌雄比例。后续我们将利用生态学、分子生物学、细胞学及解剖学等手段作进一步研究。
附录1 细枝柃不同性别花的形态 Appendix 1 The flower morphology in different gender plant of
Eurya loquaiana http://www.chinbullbotany.com/fileup/PDF/t16007-1.pdf 附录2 细枝柃性别变异株花的形态 Appendix 2 The flower morphology of sexual variant plants of
Eurya loquaiana http://www.chinbullbotany.com/fileup/PDF/t16007-2.pdf 附录3 缙云山细枝柃性别变异株性别表达信息 Appendix 3 Sex expression information of sexual variant plants of
Eurya loquaiana at Jinyun Mountain
http://www.chinbullbotany.com/fileup/PDF/t16007-3.pdf The authors have declared that no competing interests exist. 参考文献 文献选项 原文顺序 文献年度倒序 文中引用次数倒序 被引期刊影响因子 [1] 曹宗巽 (1965 ). 植物的性别分化及其控制. 生物学通报 2, 6 -9 .[2] 陈封怀 (1991). 广东植物志(第一卷) . 广州: 广东科技出版社. pp. 838 -843 .[3] 傅国立 (2000). 中国高等植物 . 青岛: 青岛出版社. pp. 511 -513 .[4] 林来官 (1998). 中国植物志(第50卷第1分册) . 北京: 科学出版社. pp. 170 -176 .[5] 王茜 , 邓洪平 , 丁博 , 周光林 (2012 ). 钝叶柃不同性别花的花部形态与传粉特征比较. 生态学报 12, 3921 -3930 .[6] 向远寅 (1984 ). 生物的性别与性别转变. 新疆林业 2, 44 -50 .[7] 熊济华 (1990). 四川植物志(第8卷) . 成都: 四川民族出版社. pp. 215 .[8] 张大勇 (2004). 植物生活史进化与繁殖生态学 . 北京: 科学出版社. pp. 325 -326 .[9] 张志良 , 瞿伟菁 , 李小方 (2009). 植物生理学实验指导 . 北京: 高等教育出版社. pp. 223 -225 .[10] Adam H Collin M Richaud F Beulé T Cros D 2011 ). Environmental regulation of sex determination in oil palm: current knowledge and insights from other species.Ann Bot-London 1529 -1537 .[11] Barrett SCH 2000 ). The evolution and function of stylar polymorphisms in flowering plants.Ann Bot-London 253 -265 .[12] Borthwick HA Scully NJ 1954 ). Photoperiodic responses of hemp.Int J Plant Sci 14 -29 .[13] Caporali E Carboni A Galli MG Rossi G Spada A 1994 ). Development of male and female flower inAsparagus officinalis . Search for point of transition from hermaphroditic to unisexual development pathway. Sex Plant Reprod 239 -249 .[14] Charlesworth D Charlesworth B 1987 ). The effect of investment in attractive structures on allocation to male and female functions in plants.Evolution 948 -968 .[15] Dafni A Shmida A 2002 ). Andromonecy inColchicum stevenii C. Koch (Liliaceae)—frequency, phenology, and reserve allocation. Israel J Plant Sci 51 -57 .[16] Dellaporta SL Urrea A 1993 ). Sex determination in flowe- ring plants.Plant Cell 124 -125 .[17] Eckert CG Barrett SCH 1994 ). Tristyly, self-compatibility and floral variation in Decodon verticillatus (Lythraceae). Biol J Linn Soc 1 -30 .[18] Givnish TJ 1980 ). Ecological constraints on the evolution of breeding systems in seed plants—dioecy and dispersal in Gymnosperms.Evolution 959 -972 .[19] Golenberg EM West NW 2013 ). Hormonal interactions and gene regulation can link monoecy and environmental plasticity to the evolution of dioecy in plants. Am J Bot 1022 -1037 .[20] Hader LD Barrett SCH 2006). Ecology and evolution of flowers. New York: Oxford University Press. pp. 50 -85 .[21] Hormaza JI Polito VS 1996 ). Pistillate and staminate flower development in dioeciousPistacia vera Anacardiaceae). Am J Bot 83, 759 -766 .[22] Humeau L Pailler T Thompson JD 1999 ). Cryptic dioecy and leaky dioecy in edemic species of Dombeya (Sterculiaceae) on La Réunion. Am J Bot 1437 -1447 .[23] Korpelainen H 1998 ). Labile sex expression in plants.Biol Rev 157 -180 .[24] Lloyd DG 1980 ). Sexual strategies in plants. III. A quantitative method for describing the gender of plants.New Zeal J Bot 103 -108 .[25] Mayer SS Charlesworth D 1991 ). Cryptic dioecy in flowe- ring plants.Trend Ecol Evol 320 -325 .[26] McArthur ED 1977 ). Environmentally induced changes of sex expression inAtriplex canescens . Heredity 97 -103 .[27] Meagher TR Sex determination in plants. New York: Oxford University Press. pp. 113 -125 .[28] Miller JS Venable DL 2003 ). Floral morphometrics and the evolution of sexual dimorphism inLycium (Solanaceae). Evolution 74 -86 .[29] Murata H 1991 ). Variation of sex expression and flower structure in a population ofEurya japonica Thunb. J Jap Bot 229 -234 .[30] Nanami S Hideyuki K Takuo Y 2004 ). Sex change towards female in dyingAcer rufinerve trees. Ann Bot 733 -740 .[31] Rottenberg A 1998 ). Sex ratio and gender stability in the dioecious plants of Israel.Bot J Linn Soc 137 -148 .[32] Rottenberg A 2000 ). Fertility of exceptional bisexual individuals in four dioecious plant species.Sex Plant Reprod 219 -211 .[33] Sarkissian TS Barrett SCH Harder LD 2001 ). Gender variation inSagittaria latifolia (Alismataceae): is size all that matters? Ecology 360 -373 .[34] Solomon BP 1985 ). Environmentally influenced changes in sex expression in an andromonoecious plant.Ecology 1321 -1332 .[35] Standley LA 1985 ). Paradioecy and gender ratios inCarex macrocephala (Cyperaceae). Am Midl Nat 283 -286 .[36] Venkatasamy S Khittoo G Keeley S 2007 ). Leaky dioecy inDiospyros (Ebenaceae) endemic to the Island of Mauritius. Plant Ecol 139 -146 .[37] Vesque MJ 1895 ). Revision of genusEurya Thunb. Bulletin Société Botanique France 160 .[38] Zhang DY Jiang XH 2000 ). Costly solicitation, timing of offspring onflict, and resource allocation in plants.Ann Bot 123 -131 .
植物的性别分化及其控制
1
1965
... 植物的性别表达是遗传物质和环境因子共同作用的结果.与遗传控制不同, 环境因子可以通过调控特异基因的表达来诱导植物性别的转换(
Solomon, 1985 ).在苔藓、蕨类和种子植物类群中, 关于环境改变导致植物性别发生转变已有报道(
Golenberg and West, 2013 ).被子植物中, 性别转变现象发生于雌雄异株与雌雄异花同株植物类群的概率大于雌雄同株植物类群(
Korpelainen and Labile, 1998 ).植物雌雄个体或雌雄器官之间存在着深刻的代谢差异, 环境条件的改变使植物的代谢途径或强度发生朝另一种性别发生方向改变(
曹宗巽, 1965 ).基因突变导致机体不适或退化及种群资源限制也能使植物性别发生变异(
向远寅, 1984 ).本研究中, 缙云山细枝柃种群中, 雌株占优势, 雌性性别有向雄性性别转化的趋势, 因此推测其性别发生变异可能是资源限制所致.缙云山气候环境特殊, 细枝柃花期常逢阴雨, 传粉效率受影响.根据种群繁殖理论, 细枝柃为扩大种群, 需调整雌雄比例.后续我们将利用生态学、分子生物学、细胞学及解剖学等手段作进一步研究. ...
1
... 以重庆市分布的山茶科柃木属植物细枝柃(
Eurya loquaiana Dunn.)为研究对象, 其形态特征与《中国植物志》及地方植物志(
熊济华, 1990 ;
陈封怀, 1991 )中的描述基本一致, 但部分植株表现出明显的性别变异现象, 即性别表达偏离正常的雌株和雄株.在缙云山调查的300余株细枝柃中, 性别变异株的比例达23.08%. ...
1
... 植物固着生长的习性决定其不能主动地寻找配偶, 为能在不同的环境条件下实现雌雄配子的最大适合度, 植物在长期的进化过程中演化出类型多样的繁殖系统和繁殖策略(
Barrett, 2000 ).植物在生活史中, 通过资源的最佳分配格局并凭借特有的繁殖属性来适应特定环境, 以提高物种适合度(
Eckert and Barrett, 1994 ;
傅国立, 2000 ).根据资源利用假说, 植物可通过调控分配给繁殖构件和营养构件的相对资源比例来保障繁殖成功(
Charlesworth and Charlesworth, 1987 ;
Zhang and Jiang, 2000 ).再分配理论指出, 资源分配模式的改变能够促进雌雄异株植物的进化(
Givnish, 1980 ).研究结果表明, 细枝柃不同类型花的雌、雄蕊特征存在一定差异(
表3 ).变异株雄花的雄蕊显著小于雄株花与性别变异株两性花, 变异株两性花雄蕊小于雄株花.变异株雌花的雌蕊显著小于雌株花与性别变异株两性花, 变异株两性花雌蕊小于雌株花, 但差异不明显. ...
1
... 柃木属(
Eurya )隶属山茶科(Theaceae), 约130种, 是Thunberg于1783年根据日本产的
Eurya japonica 为模式创立的新属(
林来官, 1998 ).已有的研究资料将柃木属描述为严格的雌雄异株植物, 性别变异现象极为少见, 偶有关于柃木属植物性别变异现象的报道.Vesque (1895)发现柃木属存在性别变异现象.日本****Murata (1991)发现柃木(
E. japonica )中存在两性花.王茜等(2012)在调查中发现钝叶柃(
E. obtusifolia )中存在性别变异现象, 并对其不同性别花的花部形态与传粉特征进行了研究. 近几年, 笔者通过野外调查发现细枝柃(
E. loqu- aiana )中存在性别变异现象, 性别变异株上具有不同性别类型的花.为探究细枝柃的性别表达特性, 本研究从单花和植株两个层面上对其性别表达进行探讨, 并分析花部构件的资源分配差异, 以揭示性别差异的生态适应机制.同时依据性别变异株上花的类型初步探讨柃木属性别进化的途径, 为柃木属的性别演化规律研究提供基础资料. ...
钝叶柃不同性别花的花部形态与传粉特征比较
1
2012
... 细枝柃的性别表达特性, 是细枝柃性别决定的外在体现及其性别进化的结果.目前, 已在10种柃木属植物中发现性别变异, 有7种为林来官在整理标本时发现的, 但未进行详尽描述(
王茜等, 2012 ).Murata (1991)对柃木中出现的性别变异进行了详细描述.王茜(2012)对钝叶柃性别变异性别表达特性进行了描述.本研究的观察结果与他们的描述有许多相似之处. ...
生物的性别与性别转变
1
1984
... 植物的性别表达是遗传物质和环境因子共同作用的结果.与遗传控制不同, 环境因子可以通过调控特异基因的表达来诱导植物性别的转换(
Solomon, 1985 ).在苔藓、蕨类和种子植物类群中, 关于环境改变导致植物性别发生转变已有报道(
Golenberg and West, 2013 ).被子植物中, 性别转变现象发生于雌雄异株与雌雄异花同株植物类群的概率大于雌雄同株植物类群(
Korpelainen and Labile, 1998 ).植物雌雄个体或雌雄器官之间存在着深刻的代谢差异, 环境条件的改变使植物的代谢途径或强度发生朝另一种性别发生方向改变(
曹宗巽, 1965 ).基因突变导致机体不适或退化及种群资源限制也能使植物性别发生变异(
向远寅, 1984 ).本研究中, 缙云山细枝柃种群中, 雌株占优势, 雌性性别有向雄性性别转化的趋势, 因此推测其性别发生变异可能是资源限制所致.缙云山气候环境特殊, 细枝柃花期常逢阴雨, 传粉效率受影响.根据种群繁殖理论, 细枝柃为扩大种群, 需调整雌雄比例.后续我们将利用生态学、分子生物学、细胞学及解剖学等手段作进一步研究. ...
1
... 以重庆市分布的山茶科柃木属植物细枝柃(
Eurya loquaiana Dunn.)为研究对象, 其形态特征与《中国植物志》及地方植物志(
熊济华, 1990 ;
陈封怀, 1991 )中的描述基本一致, 但部分植株表现出明显的性别变异现象, 即性别表达偏离正常的雌株和雄株.在缙云山调查的300余株细枝柃中, 性别变异株的比例达23.08%. ...
2
... 这些差异表明, 在细枝柃花期, 雄株用于生殖的资源投资大于雌株.雌株在此时投入给生殖的资源弱于雄株.若投入给花瓣较多的资源, 收益不大, 因雌株在后期孕育果实和种子时需要投资大量的能量.为达到资源的优化配置, 雌株将在花冠上的投资节约下来, 以孕育果实和种子(
Miller and Venable, 2003 ;
张大勇, 2004 ).在细枝柃各花部构件生物量分配中, 雄花(包括雄株花和变异株雄花)花部构件生物量分配中雄蕊生物量的分配低于雌花(包括雌株花和变异株雌花)中雌蕊生物量的分配; 两性花中, 雄蕊生物量分配低于雌蕊生物量分配, 这是其优化资源分配的手段, 进而获取最大适合度收益. ...
... 从细枝柃性别变异株上各花的形态与性别功能, 可推测变异株上花的变化过程为: 雌花→具退化雄蕊的雌花→两性花→具退化雌蕊的雄花→雄花.根据目前已经得到证实的5条由两性状态向雌雄异株进化的途径(
张大勇, 2004 ), 推测柃木属植物向雌雄异株进化需经历雌全株阶段.在本研究所调查的41株变异株中, 以雌全株比例最多, 与该推断相符.有研究证明, 在干旱胁迫下, 雌雄异株植物种群内的性别格局会发生改变, 种群中的雄株数会增加, 致使雌雄比例下降(
Borthwick and Scully, 1954 ;
McArthur, 1977 ;
Dafni and Shmida, 2002 ;
Nanami et al., 2004 ;
Adam et al., 2011 ).细枝柃性别变异株上花的变化过程预示着变异株有转变为雄株的趋势, 若变化稳定持续, 变异株将会转变为雄株, 最终雄株数量增加, 雌雄比例下降, 但这一结论需经连续多年的观察证实. ...
2
... 柱头可授性的检测采用联苯胺-过氧化氢法(
张志良等, 2009 ).花粉活力的测定采用TTC (2, 3, 5-氯化三苯基四氮唑)法(
张志良等, 2009 ). ...
... ).花粉活力的测定采用TTC (2, 3, 5-氯化三苯基四氮唑)法(
张志良等, 2009 ). ...
1
2011
... 从细枝柃性别变异株上各花的形态与性别功能, 可推测变异株上花的变化过程为: 雌花→具退化雄蕊的雌花→两性花→具退化雌蕊的雄花→雄花.根据目前已经得到证实的5条由两性状态向雌雄异株进化的途径(
张大勇, 2004 ), 推测柃木属植物向雌雄异株进化需经历雌全株阶段.在本研究所调查的41株变异株中, 以雌全株比例最多, 与该推断相符.有研究证明, 在干旱胁迫下, 雌雄异株植物种群内的性别格局会发生改变, 种群中的雄株数会增加, 致使雌雄比例下降(
Borthwick and Scully, 1954 ;
McArthur, 1977 ;
Dafni and Shmida, 2002 ;
Nanami et al., 2004 ;
Adam et al., 2011 ).细枝柃性别变异株上花的变化过程预示着变异株有转变为雄株的趋势, 若变化稳定持续, 变异株将会转变为雄株, 最终雄株数量增加, 雌雄比例下降, 但这一结论需经连续多年的观察证实. ...
1
2000
... 植物固着生长的习性决定其不能主动地寻找配偶, 为能在不同的环境条件下实现雌雄配子的最大适合度, 植物在长期的进化过程中演化出类型多样的繁殖系统和繁殖策略(
Barrett, 2000 ).植物在生活史中, 通过资源的最佳分配格局并凭借特有的繁殖属性来适应特定环境, 以提高物种适合度(
Eckert and Barrett, 1994 ;
傅国立, 2000 ).根据资源利用假说, 植物可通过调控分配给繁殖构件和营养构件的相对资源比例来保障繁殖成功(
Charlesworth and Charlesworth, 1987 ;
Zhang and Jiang, 2000 ).再分配理论指出, 资源分配模式的改变能够促进雌雄异株植物的进化(
Givnish, 1980 ).研究结果表明, 细枝柃不同类型花的雌、雄蕊特征存在一定差异(
表3 ).变异株雄花的雄蕊显著小于雄株花与性别变异株两性花, 变异株两性花雄蕊小于雄株花.变异株雌花的雌蕊显著小于雌株花与性别变异株两性花, 变异株两性花雌蕊小于雌株花, 但差异不明显. ...
1
1954
... 从细枝柃性别变异株上各花的形态与性别功能, 可推测变异株上花的变化过程为: 雌花→具退化雄蕊的雌花→两性花→具退化雌蕊的雄花→雄花.根据目前已经得到证实的5条由两性状态向雌雄异株进化的途径(
张大勇, 2004 ), 推测柃木属植物向雌雄异株进化需经历雌全株阶段.在本研究所调查的41株变异株中, 以雌全株比例最多, 与该推断相符.有研究证明, 在干旱胁迫下, 雌雄异株植物种群内的性别格局会发生改变, 种群中的雄株数会增加, 致使雌雄比例下降(
Borthwick and Scully, 1954 ;
McArthur, 1977 ;
Dafni and Shmida, 2002 ;
Nanami et al., 2004 ;
Adam et al., 2011 ).细枝柃性别变异株上花的变化过程预示着变异株有转变为雄株的趋势, 若变化稳定持续, 变异株将会转变为雄株, 最终雄株数量增加, 雌雄比例下降, 但这一结论需经连续多年的观察证实. ...
1
1994
... 植物的性别通常受遗传控制(
Meagher, 1988 ;
Dellaporta and Urrea, 1993 ).植物的性别分化可能发生于花发育的早期或后期(
Caporali et al., 1994 ;
Hormaza and Polito, 1996 ), 当性别分化发生相对 较晚时, 其它性别的性器官可能会有表达(
Mayer and Charlesworth, 1991 ).然而, 在完全雌雄异株物种的自然种群中, 两性个体较为罕见.Rottenberg (1998)在调查以色列41种雌雄异株植物的12个种群时, 发现它们之中出现两性个体的概率低于0.1%.在某些雌雄异株植物中偶尔会出现相对性别特征的现象, 称为性别变异(gender variation).常用leaky dioecy (
Humeau et al.,1999 ;
Venkatasamy et al., 2007 )、inconstancy (
Sarkissian et al., 2001 )、exceptional bisexual individual (
Rottenberg, 2000 )、subdioecy (
Hader and Barrett, 2006 )、near-dioecy (
Solomon, 1985 )及paradioecy (
Standley, 1985 )等词描述出现性别变异现象的雌雄异株植物或种群. ...
1
1987
... 植物固着生长的习性决定其不能主动地寻找配偶, 为能在不同的环境条件下实现雌雄配子的最大适合度, 植物在长期的进化过程中演化出类型多样的繁殖系统和繁殖策略(
Barrett, 2000 ).植物在生活史中, 通过资源的最佳分配格局并凭借特有的繁殖属性来适应特定环境, 以提高物种适合度(
Eckert and Barrett, 1994 ;
傅国立, 2000 ).根据资源利用假说, 植物可通过调控分配给繁殖构件和营养构件的相对资源比例来保障繁殖成功(
Charlesworth and Charlesworth, 1987 ;
Zhang and Jiang, 2000 ).再分配理论指出, 资源分配模式的改变能够促进雌雄异株植物的进化(
Givnish, 1980 ).研究结果表明, 细枝柃不同类型花的雌、雄蕊特征存在一定差异(
表3 ).变异株雄花的雄蕊显著小于雄株花与性别变异株两性花, 变异株两性花雄蕊小于雄株花.变异株雌花的雌蕊显著小于雌株花与性别变异株两性花, 变异株两性花雌蕊小于雌株花, 但差异不明显. ...
1
2002
... 从细枝柃性别变异株上各花的形态与性别功能, 可推测变异株上花的变化过程为: 雌花→具退化雄蕊的雌花→两性花→具退化雌蕊的雄花→雄花.根据目前已经得到证实的5条由两性状态向雌雄异株进化的途径(
张大勇, 2004 ), 推测柃木属植物向雌雄异株进化需经历雌全株阶段.在本研究所调查的41株变异株中, 以雌全株比例最多, 与该推断相符.有研究证明, 在干旱胁迫下, 雌雄异株植物种群内的性别格局会发生改变, 种群中的雄株数会增加, 致使雌雄比例下降(
Borthwick and Scully, 1954 ;
McArthur, 1977 ;
Dafni and Shmida, 2002 ;
Nanami et al., 2004 ;
Adam et al., 2011 ).细枝柃性别变异株上花的变化过程预示着变异株有转变为雄株的趋势, 若变化稳定持续, 变异株将会转变为雄株, 最终雄株数量增加, 雌雄比例下降, 但这一结论需经连续多年的观察证实. ...
1
1993
... 植物的性别通常受遗传控制(
Meagher, 1988 ;
Dellaporta and Urrea, 1993 ).植物的性别分化可能发生于花发育的早期或后期(
Caporali et al., 1994 ;
Hormaza and Polito, 1996 ), 当性别分化发生相对 较晚时, 其它性别的性器官可能会有表达(
Mayer and Charlesworth, 1991 ).然而, 在完全雌雄异株物种的自然种群中, 两性个体较为罕见.Rottenberg (1998)在调查以色列41种雌雄异株植物的12个种群时, 发现它们之中出现两性个体的概率低于0.1%.在某些雌雄异株植物中偶尔会出现相对性别特征的现象, 称为性别变异(gender variation).常用leaky dioecy (
Humeau et al.,1999 ;
Venkatasamy et al., 2007 )、inconstancy (
Sarkissian et al., 2001 )、exceptional bisexual individual (
Rottenberg, 2000 )、subdioecy (
Hader and Barrett, 2006 )、near-dioecy (
Solomon, 1985 )及paradioecy (
Standley, 1985 )等词描述出现性别变异现象的雌雄异株植物或种群. ...
1
1994
... 植物固着生长的习性决定其不能主动地寻找配偶, 为能在不同的环境条件下实现雌雄配子的最大适合度, 植物在长期的进化过程中演化出类型多样的繁殖系统和繁殖策略(
Barrett, 2000 ).植物在生活史中, 通过资源的最佳分配格局并凭借特有的繁殖属性来适应特定环境, 以提高物种适合度(
Eckert and Barrett, 1994 ;
傅国立, 2000 ).根据资源利用假说, 植物可通过调控分配给繁殖构件和营养构件的相对资源比例来保障繁殖成功(
Charlesworth and Charlesworth, 1987 ;
Zhang and Jiang, 2000 ).再分配理论指出, 资源分配模式的改变能够促进雌雄异株植物的进化(
Givnish, 1980 ).研究结果表明, 细枝柃不同类型花的雌、雄蕊特征存在一定差异(
表3 ).变异株雄花的雄蕊显著小于雄株花与性别变异株两性花, 变异株两性花雄蕊小于雄株花.变异株雌花的雌蕊显著小于雌株花与性别变异株两性花, 变异株两性花雌蕊小于雌株花, 但差异不明显. ...
1
1980
... 植物固着生长的习性决定其不能主动地寻找配偶, 为能在不同的环境条件下实现雌雄配子的最大适合度, 植物在长期的进化过程中演化出类型多样的繁殖系统和繁殖策略(
Barrett, 2000 ).植物在生活史中, 通过资源的最佳分配格局并凭借特有的繁殖属性来适应特定环境, 以提高物种适合度(
Eckert and Barrett, 1994 ;
傅国立, 2000 ).根据资源利用假说, 植物可通过调控分配给繁殖构件和营养构件的相对资源比例来保障繁殖成功(
Charlesworth and Charlesworth, 1987 ;
Zhang and Jiang, 2000 ).再分配理论指出, 资源分配模式的改变能够促进雌雄异株植物的进化(
Givnish, 1980 ).研究结果表明, 细枝柃不同类型花的雌、雄蕊特征存在一定差异(
表3 ).变异株雄花的雄蕊显著小于雄株花与性别变异株两性花, 变异株两性花雄蕊小于雄株花.变异株雌花的雌蕊显著小于雌株花与性别变异株两性花, 变异株两性花雌蕊小于雌株花, 但差异不明显. ...
1
2013
... 植物的性别表达是遗传物质和环境因子共同作用的结果.与遗传控制不同, 环境因子可以通过调控特异基因的表达来诱导植物性别的转换(
Solomon, 1985 ).在苔藓、蕨类和种子植物类群中, 关于环境改变导致植物性别发生转变已有报道(
Golenberg and West, 2013 ).被子植物中, 性别转变现象发生于雌雄异株与雌雄异花同株植物类群的概率大于雌雄同株植物类群(
Korpelainen and Labile, 1998 ).植物雌雄个体或雌雄器官之间存在着深刻的代谢差异, 环境条件的改变使植物的代谢途径或强度发生朝另一种性别发生方向改变(
曹宗巽, 1965 ).基因突变导致机体不适或退化及种群资源限制也能使植物性别发生变异(
向远寅, 1984 ).本研究中, 缙云山细枝柃种群中, 雌株占优势, 雌性性别有向雄性性别转化的趋势, 因此推测其性别发生变异可能是资源限制所致.缙云山气候环境特殊, 细枝柃花期常逢阴雨, 传粉效率受影响.根据种群繁殖理论, 细枝柃为扩大种群, 需调整雌雄比例.后续我们将利用生态学、分子生物学、细胞学及解剖学等手段作进一步研究. ...
1
... 植物的性别通常受遗传控制(
Meagher, 1988 ;
Dellaporta and Urrea, 1993 ).植物的性别分化可能发生于花发育的早期或后期(
Caporali et al., 1994 ;
Hormaza and Polito, 1996 ), 当性别分化发生相对 较晚时, 其它性别的性器官可能会有表达(
Mayer and Charlesworth, 1991 ).然而, 在完全雌雄异株物种的自然种群中, 两性个体较为罕见.Rottenberg (1998)在调查以色列41种雌雄异株植物的12个种群时, 发现它们之中出现两性个体的概率低于0.1%.在某些雌雄异株植物中偶尔会出现相对性别特征的现象, 称为性别变异(gender variation).常用leaky dioecy (
Humeau et al.,1999 ;
Venkatasamy et al., 2007 )、inconstancy (
Sarkissian et al., 2001 )、exceptional bisexual individual (
Rottenberg, 2000 )、subdioecy (
Hader and Barrett, 2006 )、near-dioecy (
Solomon, 1985 )及paradioecy (
Standley, 1985 )等词描述出现性别变异现象的雌雄异株植物或种群. ...
1
1996
... 植物的性别通常受遗传控制(
Meagher, 1988 ;
Dellaporta and Urrea, 1993 ).植物的性别分化可能发生于花发育的早期或后期(
Caporali et al., 1994 ;
Hormaza and Polito, 1996 ), 当性别分化发生相对 较晚时, 其它性别的性器官可能会有表达(
Mayer and Charlesworth, 1991 ).然而, 在完全雌雄异株物种的自然种群中, 两性个体较为罕见.Rottenberg (1998)在调查以色列41种雌雄异株植物的12个种群时, 发现它们之中出现两性个体的概率低于0.1%.在某些雌雄异株植物中偶尔会出现相对性别特征的现象, 称为性别变异(gender variation).常用leaky dioecy (
Humeau et al.,1999 ;
Venkatasamy et al., 2007 )、inconstancy (
Sarkissian et al., 2001 )、exceptional bisexual individual (
Rottenberg, 2000 )、subdioecy (
Hader and Barrett, 2006 )、near-dioecy (
Solomon, 1985 )及paradioecy (
Standley, 1985 )等词描述出现性别变异现象的雌雄异株植物或种群. ...
1
1999
... 植物的性别通常受遗传控制(
Meagher, 1988 ;
Dellaporta and Urrea, 1993 ).植物的性别分化可能发生于花发育的早期或后期(
Caporali et al., 1994 ;
Hormaza and Polito, 1996 ), 当性别分化发生相对 较晚时, 其它性别的性器官可能会有表达(
Mayer and Charlesworth, 1991 ).然而, 在完全雌雄异株物种的自然种群中, 两性个体较为罕见.Rottenberg (1998)在调查以色列41种雌雄异株植物的12个种群时, 发现它们之中出现两性个体的概率低于0.1%.在某些雌雄异株植物中偶尔会出现相对性别特征的现象, 称为性别变异(gender variation).常用leaky dioecy (
Humeau et al.,1999 ;
Venkatasamy et al., 2007 )、inconstancy (
Sarkissian et al., 2001 )、exceptional bisexual individual (
Rottenberg, 2000 )、subdioecy (
Hader and Barrett, 2006 )、near-dioecy (
Solomon, 1985 )及paradioecy (
Standley, 1985 )等词描述出现性别变异现象的雌雄异株植物或种群. ...
1
1998
... 植物的性别表达是遗传物质和环境因子共同作用的结果.与遗传控制不同, 环境因子可以通过调控特异基因的表达来诱导植物性别的转换(
Solomon, 1985 ).在苔藓、蕨类和种子植物类群中, 关于环境改变导致植物性别发生转变已有报道(
Golenberg and West, 2013 ).被子植物中, 性别转变现象发生于雌雄异株与雌雄异花同株植物类群的概率大于雌雄同株植物类群(
Korpelainen and Labile, 1998 ).植物雌雄个体或雌雄器官之间存在着深刻的代谢差异, 环境条件的改变使植物的代谢途径或强度发生朝另一种性别发生方向改变(
曹宗巽, 1965 ).基因突变导致机体不适或退化及种群资源限制也能使植物性别发生变异(
向远寅, 1984 ).本研究中, 缙云山细枝柃种群中, 雌株占优势, 雌性性别有向雄性性别转化的趋势, 因此推测其性别发生变异可能是资源限制所致.缙云山气候环境特殊, 细枝柃花期常逢阴雨, 传粉效率受影响.根据种群繁殖理论, 细枝柃为扩大种群, 需调整雌雄比例.后续我们将利用生态学、分子生物学、细胞学及解剖学等手段作进一步研究. ...
1
1980
... 变异株性别功能的确定采用
Lloyd等(1980) 的方法.用以下公式对性别进行量化: ...
1
1991
... 植物的性别通常受遗传控制(
Meagher, 1988 ;
Dellaporta and Urrea, 1993 ).植物的性别分化可能发生于花发育的早期或后期(
Caporali et al., 1994 ;
Hormaza and Polito, 1996 ), 当性别分化发生相对 较晚时, 其它性别的性器官可能会有表达(
Mayer and Charlesworth, 1991 ).然而, 在完全雌雄异株物种的自然种群中, 两性个体较为罕见.Rottenberg (1998)在调查以色列41种雌雄异株植物的12个种群时, 发现它们之中出现两性个体的概率低于0.1%.在某些雌雄异株植物中偶尔会出现相对性别特征的现象, 称为性别变异(gender variation).常用leaky dioecy (
Humeau et al.,1999 ;
Venkatasamy et al., 2007 )、inconstancy (
Sarkissian et al., 2001 )、exceptional bisexual individual (
Rottenberg, 2000 )、subdioecy (
Hader and Barrett, 2006 )、near-dioecy (
Solomon, 1985 )及paradioecy (
Standley, 1985 )等词描述出现性别变异现象的雌雄异株植物或种群. ...
1
1977
... 从细枝柃性别变异株上各花的形态与性别功能, 可推测变异株上花的变化过程为: 雌花→具退化雄蕊的雌花→两性花→具退化雌蕊的雄花→雄花.根据目前已经得到证实的5条由两性状态向雌雄异株进化的途径(
张大勇, 2004 ), 推测柃木属植物向雌雄异株进化需经历雌全株阶段.在本研究所调查的41株变异株中, 以雌全株比例最多, 与该推断相符.有研究证明, 在干旱胁迫下, 雌雄异株植物种群内的性别格局会发生改变, 种群中的雄株数会增加, 致使雌雄比例下降(
Borthwick and Scully, 1954 ;
McArthur, 1977 ;
Dafni and Shmida, 2002 ;
Nanami et al., 2004 ;
Adam et al., 2011 ).细枝柃性别变异株上花的变化过程预示着变异株有转变为雄株的趋势, 若变化稳定持续, 变异株将会转变为雄株, 最终雄株数量增加, 雌雄比例下降, 但这一结论需经连续多年的观察证实. ...
1
... 植物的性别通常受遗传控制(
Meagher, 1988 ;
Dellaporta and Urrea, 1993 ).植物的性别分化可能发生于花发育的早期或后期(
Caporali et al., 1994 ;
Hormaza and Polito, 1996 ), 当性别分化发生相对 较晚时, 其它性别的性器官可能会有表达(
Mayer and Charlesworth, 1991 ).然而, 在完全雌雄异株物种的自然种群中, 两性个体较为罕见.Rottenberg (1998)在调查以色列41种雌雄异株植物的12个种群时, 发现它们之中出现两性个体的概率低于0.1%.在某些雌雄异株植物中偶尔会出现相对性别特征的现象, 称为性别变异(gender variation).常用leaky dioecy (
Humeau et al.,1999 ;
Venkatasamy et al., 2007 )、inconstancy (
Sarkissian et al., 2001 )、exceptional bisexual individual (
Rottenberg, 2000 )、subdioecy (
Hader and Barrett, 2006 )、near-dioecy (
Solomon, 1985 )及paradioecy (
Standley, 1985 )等词描述出现性别变异现象的雌雄异株植物或种群. ...
1
2003
... 这些差异表明, 在细枝柃花期, 雄株用于生殖的资源投资大于雌株.雌株在此时投入给生殖的资源弱于雄株.若投入给花瓣较多的资源, 收益不大, 因雌株在后期孕育果实和种子时需要投资大量的能量.为达到资源的优化配置, 雌株将在花冠上的投资节约下来, 以孕育果实和种子(
Miller and Venable, 2003 ;
张大勇, 2004 ).在细枝柃各花部构件生物量分配中, 雄花(包括雄株花和变异株雄花)花部构件生物量分配中雄蕊生物量的分配低于雌花(包括雌株花和变异株雌花)中雌蕊生物量的分配; 两性花中, 雄蕊生物量分配低于雌蕊生物量分配, 这是其优化资源分配的手段, 进而获取最大适合度收益. ...
1991
1
2004
... 从细枝柃性别变异株上各花的形态与性别功能, 可推测变异株上花的变化过程为: 雌花→具退化雄蕊的雌花→两性花→具退化雌蕊的雄花→雄花.根据目前已经得到证实的5条由两性状态向雌雄异株进化的途径(
张大勇, 2004 ), 推测柃木属植物向雌雄异株进化需经历雌全株阶段.在本研究所调查的41株变异株中, 以雌全株比例最多, 与该推断相符.有研究证明, 在干旱胁迫下, 雌雄异株植物种群内的性别格局会发生改变, 种群中的雄株数会增加, 致使雌雄比例下降(
Borthwick and Scully, 1954 ;
McArthur, 1977 ;
Dafni and Shmida, 2002 ;
Nanami et al., 2004 ;
Adam et al., 2011 ).细枝柃性别变异株上花的变化过程预示着变异株有转变为雄株的趋势, 若变化稳定持续, 变异株将会转变为雄株, 最终雄株数量增加, 雌雄比例下降, 但这一结论需经连续多年的观察证实. ...
1998
1
2000
... 植物的性别通常受遗传控制(
Meagher, 1988 ;
Dellaporta and Urrea, 1993 ).植物的性别分化可能发生于花发育的早期或后期(
Caporali et al., 1994 ;
Hormaza and Polito, 1996 ), 当性别分化发生相对 较晚时, 其它性别的性器官可能会有表达(
Mayer and Charlesworth, 1991 ).然而, 在完全雌雄异株物种的自然种群中, 两性个体较为罕见.Rottenberg (1998)在调查以色列41种雌雄异株植物的12个种群时, 发现它们之中出现两性个体的概率低于0.1%.在某些雌雄异株植物中偶尔会出现相对性别特征的现象, 称为性别变异(gender variation).常用leaky dioecy (
Humeau et al.,1999 ;
Venkatasamy et al., 2007 )、inconstancy (
Sarkissian et al., 2001 )、exceptional bisexual individual (
Rottenberg, 2000 )、subdioecy (
Hader and Barrett, 2006 )、near-dioecy (
Solomon, 1985 )及paradioecy (
Standley, 1985 )等词描述出现性别变异现象的雌雄异株植物或种群. ...
1
2001
... 植物的性别通常受遗传控制(
Meagher, 1988 ;
Dellaporta and Urrea, 1993 ).植物的性别分化可能发生于花发育的早期或后期(
Caporali et al., 1994 ;
Hormaza and Polito, 1996 ), 当性别分化发生相对 较晚时, 其它性别的性器官可能会有表达(
Mayer and Charlesworth, 1991 ).然而, 在完全雌雄异株物种的自然种群中, 两性个体较为罕见.Rottenberg (1998)在调查以色列41种雌雄异株植物的12个种群时, 发现它们之中出现两性个体的概率低于0.1%.在某些雌雄异株植物中偶尔会出现相对性别特征的现象, 称为性别变异(gender variation).常用leaky dioecy (
Humeau et al.,1999 ;
Venkatasamy et al., 2007 )、inconstancy (
Sarkissian et al., 2001 )、exceptional bisexual individual (
Rottenberg, 2000 )、subdioecy (
Hader and Barrett, 2006 )、near-dioecy (
Solomon, 1985 )及paradioecy (
Standley, 1985 )等词描述出现性别变异现象的雌雄异株植物或种群. ...
2
1985
... 植物的性别通常受遗传控制(
Meagher, 1988 ;
Dellaporta and Urrea, 1993 ).植物的性别分化可能发生于花发育的早期或后期(
Caporali et al., 1994 ;
Hormaza and Polito, 1996 ), 当性别分化发生相对 较晚时, 其它性别的性器官可能会有表达(
Mayer and Charlesworth, 1991 ).然而, 在完全雌雄异株物种的自然种群中, 两性个体较为罕见.Rottenberg (1998)在调查以色列41种雌雄异株植物的12个种群时, 发现它们之中出现两性个体的概率低于0.1%.在某些雌雄异株植物中偶尔会出现相对性别特征的现象, 称为性别变异(gender variation).常用leaky dioecy (
Humeau et al.,1999 ;
Venkatasamy et al., 2007 )、inconstancy (
Sarkissian et al., 2001 )、exceptional bisexual individual (
Rottenberg, 2000 )、subdioecy (
Hader and Barrett, 2006 )、near-dioecy (
Solomon, 1985 )及paradioecy (
Standley, 1985 )等词描述出现性别变异现象的雌雄异株植物或种群. ...
... 植物的性别表达是遗传物质和环境因子共同作用的结果.与遗传控制不同, 环境因子可以通过调控特异基因的表达来诱导植物性别的转换(
Solomon, 1985 ).在苔藓、蕨类和种子植物类群中, 关于环境改变导致植物性别发生转变已有报道(
Golenberg and West, 2013 ).被子植物中, 性别转变现象发生于雌雄异株与雌雄异花同株植物类群的概率大于雌雄同株植物类群(
Korpelainen and Labile, 1998 ).植物雌雄个体或雌雄器官之间存在着深刻的代谢差异, 环境条件的改变使植物的代谢途径或强度发生朝另一种性别发生方向改变(
曹宗巽, 1965 ).基因突变导致机体不适或退化及种群资源限制也能使植物性别发生变异(
向远寅, 1984 ).本研究中, 缙云山细枝柃种群中, 雌株占优势, 雌性性别有向雄性性别转化的趋势, 因此推测其性别发生变异可能是资源限制所致.缙云山气候环境特殊, 细枝柃花期常逢阴雨, 传粉效率受影响.根据种群繁殖理论, 细枝柃为扩大种群, 需调整雌雄比例.后续我们将利用生态学、分子生物学、细胞学及解剖学等手段作进一步研究. ...
1
1985
... 植物的性别通常受遗传控制(
Meagher, 1988 ;
Dellaporta and Urrea, 1993 ).植物的性别分化可能发生于花发育的早期或后期(
Caporali et al., 1994 ;
Hormaza and Polito, 1996 ), 当性别分化发生相对 较晚时, 其它性别的性器官可能会有表达(
Mayer and Charlesworth, 1991 ).然而, 在完全雌雄异株物种的自然种群中, 两性个体较为罕见.Rottenberg (1998)在调查以色列41种雌雄异株植物的12个种群时, 发现它们之中出现两性个体的概率低于0.1%.在某些雌雄异株植物中偶尔会出现相对性别特征的现象, 称为性别变异(gender variation).常用leaky dioecy (
Humeau et al.,1999 ;
Venkatasamy et al., 2007 )、inconstancy (
Sarkissian et al., 2001 )、exceptional bisexual individual (
Rottenberg, 2000 )、subdioecy (
Hader and Barrett, 2006 )、near-dioecy (
Solomon, 1985 )及paradioecy (
Standley, 1985 )等词描述出现性别变异现象的雌雄异株植物或种群. ...
1
2007
... 植物的性别通常受遗传控制(
Meagher, 1988 ;
Dellaporta and Urrea, 1993 ).植物的性别分化可能发生于花发育的早期或后期(
Caporali et al., 1994 ;
Hormaza and Polito, 1996 ), 当性别分化发生相对 较晚时, 其它性别的性器官可能会有表达(
Mayer and Charlesworth, 1991 ).然而, 在完全雌雄异株物种的自然种群中, 两性个体较为罕见.Rottenberg (1998)在调查以色列41种雌雄异株植物的12个种群时, 发现它们之中出现两性个体的概率低于0.1%.在某些雌雄异株植物中偶尔会出现相对性别特征的现象, 称为性别变异(gender variation).常用leaky dioecy (
Humeau et al.,1999 ;
Venkatasamy et al., 2007 )、inconstancy (
Sarkissian et al., 2001 )、exceptional bisexual individual (
Rottenberg, 2000 )、subdioecy (
Hader and Barrett, 2006 )、near-dioecy (
Solomon, 1985 )及paradioecy (
Standley, 1985 )等词描述出现性别变异现象的雌雄异株植物或种群. ...
1895
1
2000
... 植物固着生长的习性决定其不能主动地寻找配偶, 为能在不同的环境条件下实现雌雄配子的最大适合度, 植物在长期的进化过程中演化出类型多样的繁殖系统和繁殖策略(
Barrett, 2000 ).植物在生活史中, 通过资源的最佳分配格局并凭借特有的繁殖属性来适应特定环境, 以提高物种适合度(
Eckert and Barrett, 1994 ;
傅国立, 2000 ).根据资源利用假说, 植物可通过调控分配给繁殖构件和营养构件的相对资源比例来保障繁殖成功(
Charlesworth and Charlesworth, 1987 ;
Zhang and Jiang, 2000 ).再分配理论指出, 资源分配模式的改变能够促进雌雄异株植物的进化(
Givnish, 1980 ).研究结果表明, 细枝柃不同类型花的雌、雄蕊特征存在一定差异(
表3 ).变异株雄花的雄蕊显著小于雄株花与性别变异株两性花, 变异株两性花雄蕊小于雄株花.变异株雌花的雌蕊显著小于雌株花与性别变异株两性花, 变异株两性花雌蕊小于雌株花, 但差异不明显. ...
 ,
, 

{kind=link}
{kind=link}