Effects of competition and climate on tree radial growth of Pinus sibirica in Altai Mountains, Xinjiang, China
Jian KANG1,2,3, Han-Xue LIANG1,3, Shao-Wei JIANG1,2,3, Huo-Xing ZHU1,3, Peng ZHOU1,2,3, Jian-Guo HUANG,1,2,3,*1Key Laboratory of Vegetation Restoration and Management of Degraded Ecosystems, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China 2College of Resources and Environment, University of Chinese Academy of Sciences, Beijing 100049, China 3Core Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China
National Natural Science Foundation of China(41661144007) National Natural Science Foundation of China(4171101346) National Natural Science Foundation of China(41701047)
Abstract Aims As an important part of the ecosystems of Central Asia and the whole world, forest growth dynamics of the boreal forest in Altai Mountains can affect global thermal radiation, carbon balance and so on. Therefore, it is crucial to explore the main factors affecting tree radial growth of the boreal forest in Altai Mountains. Methods We selected Pinus sibirica in Kanas National Nature Reserve of Xinjiang as the target tree species, and established tree-ring chronology of P. sibirica. To explore the influence of competition and climate on radial growth of P. sibirica in the Altai Mountains, Xinjiang, we analyzed the relationships among the cumulated basal area increment (BAI), competition index and climate factors at different time intervals by applying various methods like linear mixed effect model and correlation analysis. Important findings The results of the linear mixed effect model showed that: (1) the BAI over the past 30 years can be best predicted by the sun of competitors’ diameter at breast height (SDBH). (2) Significant positive correlations were found between the standard chronology, and mean air temperature, mean maximum air temperature and mean minimum air temperature in March. (3) The highest value of the cumulated BAI was observed when air temperature was from 0 to 5 °C, and the competition index was below 100. The lowest value of the cumulative growth occurs when air temperature reaches -10 °C and the competition index exceeds 300. Tree radial growth of subject trees was influenced by both the diameter at breast height of the competing trees and the early growing season air temperature. Competition plays a more important role in affecting radial growth of P. sibirica than climate factors in this area. Therefore, our results would provide a scientific basis for forest management of P. sibirica in Kanas National Nature Reserve of Xinjiang. Keywords:competition;radial growth;climate;Pinus sibirica
PDF (1409KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 康剑, 梁寒雪, 蒋少伟, 朱火星, 周鹏, 黄建国. 竞争和气候对新疆阿尔泰山西伯利亚五针松树木径向生长的影响. 植物生态学报, 2020, 44(12): 1195-1202. DOI: 10.17521/cjpe.2020.0224 KANG Jian, LIANG Han-Xue, JIANG Shao-Wei, ZHU Huo-Xing, ZHOU Peng, HUANG Jian-Guo. Effects of competition and climate on tree radial growth of Pinus sibirica in Altai Mountains, Xinjiang, China. Chinese Journal of Plant Ecology, 2020, 44(12): 1195-1202. DOI: 10.17521/cjpe.2020.0224
森林生态系统在全球碳循环中起着至关重要的作用(Pan et al., 2011)。作为陆地生态系统中最大的生物群落之一, 北半球中高纬度的北方森林生态系统的动态发展对全球变化具有重要意义(Kerhoulas & Kane, 2012)。树木个体是森林生态系统的重要组成部分, 其生长除了受到自身遗传因素的影响, 还会受到诸多外界因素的影响(Kawata, 1997)。其中, 气候因子对于树木径向生长有显著的影响。水热条件的变化对北方森林树木径向生长的影响较大。研究发现蒙古中北部半干旱地区的西伯利亚落叶松(Larix sibirica)、西伯利亚五针松(Pinus sibirica)、欧洲赤松(Pinus sylvestris)树木晚材的生长主要和当年的生长季气温相关(de Grandpré et al., 2011)。尚华明等(2010)对哈萨克斯坦境内的阿尔泰山南坡西伯利亚落叶松研究发现, 树轮宽度与6月份气温显著正相关。伴随气候变化带来的降水增加, 中国西北部的祁连山区青海云杉(Picea crassifolia)的生长期延长, 其年径向生长量近年来明显增加(Gao et al., 2018)。然而, 研究者通过对阿尔泰山中东部的西伯利亚落叶松进行树轮气候学研究, 发现西伯利亚落叶松生长量与当年6月降水量呈负相关关系(黄力平等, 2015)。
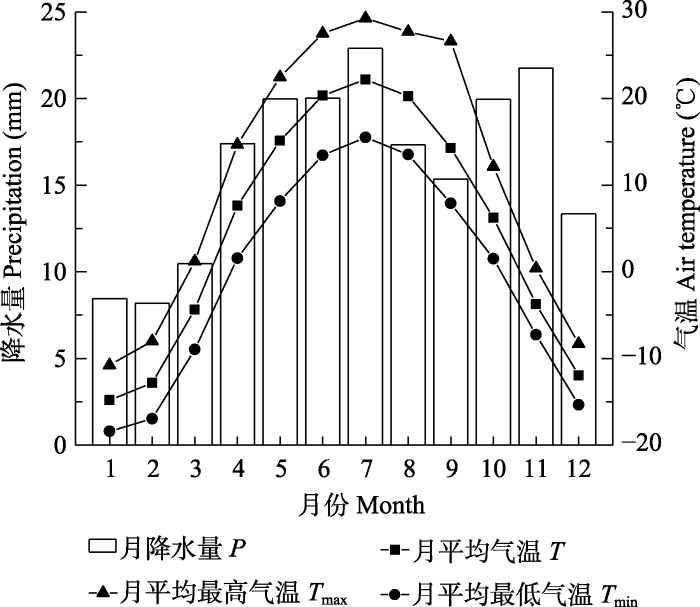
Fig. 1Variation trend of monthly mean air temperature (T), monthly mean minimum air temperature (Tmin), monthly mean maximum temperature (Tmax) and monthly precipitation (P) in the research area of Altai Mountains, Xinjiang from 1958 to 2017.
Table 3 表3 表3新疆阿尔泰山西伯利亚五针松竞争指数与累积断面积生长量的线性混合效应模型结果 Table 3Results of the linear mixed-effects models of competitive indices and cumulated basal area increments of Pinus sibirica in Altai Mountains, Xinjiang
时间间隔 Time interval (a)
竞争指数 Competition indices
a
b
临界R2 Marginal R2
条件R2 Conditional R2
5
N
0.098 3
0.000 0
0.09
0.59
SDBH
0.082 9
-0.000 1
0.12
0.6
SBA
0.072 0
-0.000 4
0.08
0.52
10
N
0.204 5
-0.000 1
0.12
0.57
SDBH
0.169 1
-0.000 3
0.16
0.59
SBA
0.145 0
-0.000 9
0.11
0.49
15
N
0.328 1
-0.000 1
0.15
0.58
SDBH
0.265 7
-0.000 5
0.20
0.60
SBA
0.226 7
-0.001 4
0.14
0.49
20
N
0.409 0
-0.000 2
0.13
0.52
SDBH
0.338 6
-0.000 6
0.18
0.55
SBA
0.290 6
-0.001 9
0.13
0.46
25
N
0.486 2
-0.000 2
0.13
0.51
SDBH
0.404 9
-0.000 7
0.18
0.54
SBA
0.349 4
-0.002 2
0.14
0.45
30
N
0.578 1
-0.000 2
0.15
0.47
SDBH
0.484 7
-0.000 9
0.22
0.62
SBA
0.416 5
-0.002 8
0.17
0.41
a、b, 混合效应中的固定效应参数。加粗部分为拟合效果好的混合效应模型结果。N, 竞争树密度; SBA, 竞争树胸高断面积和; SDBH, 竞争树胸径和。 a, b, fixed effect parameters in the mixed effect model. Bold parts are the results of mixed effect model with good fitting effect. N, density of competitors; SBA, sum of competitors’ stand basal area; SDBH, sum of competitors’ diameter at breast height.
Fig. 2Correlation between ring width chronology of Pinus sibirica and monthly climate factors. p8-p12 represents August to December of the previous year. P, monthly precipitation; T, monthly mean air temperature; Tmax, monthly mean maximum air temperature; Tmin, monthly mean minimum air temperature. * p < 0.05.
Fig. 3Relationship between cumulated basal area increments (BAI) of Pinus sibirica in Altai Mountains, Xinjiang and air temperature (T) and competition index (CI).
3 讨论
3.1 竞争对树木生长的影响
结果表明, 混合效应模型在过去15-30年的拟合效果更好。该结果与其他区域的研究结果具有一致性。研究发现, 中国亚热带地区的马尾松(Pinus massoniana)的生长量在15-25年时与竞争指数的拟合效果较好, 其中, 过去20年的生长量与竞争指数的模型拟合最佳(Liang et al., 2019)。在加拿大西部的研究中也有类似的结果, 研究发现该区域的山杨(Populus tremuloides)的生长量在过去10-25年尺度上与竞争指数达到模型拟合效果最优(Huang et al., 2013)。本研究中的西伯利亚五针松是暗针叶林的主要建群种, 其生长寿命可达400年以上, 径向生长的速生期在50-120年, 连年生长量的最高值出现在90年, 且树木生长的前50年是生长缓慢期(但新球和洪加凤, 1993)。在15-30年这个区间内, 西伯利亚五针松为幼龄林时期, 还处在林冠遮阴阶段, 营养空间较小、生长缓慢(但新球和洪加凤, 1993)。此前的研究表明, 目标树的径级越大, 其对竞争的敏感度会越低(Gómez-Aparicio et al., 2011)。因此, 15-30年的树木径向生长量对于竞争有着更强的敏感性。
deGrandpré L, TardifJC, HesslA, PedersonN, ConciatoriF, GreenTR, OyunsanaaB, BaatarbilegN (2011). Seasonal shift in the climate responses of Pinus sibirica, Pinus sylvestris, and Larix sibirica trees from semi-arid, north- central Mongolia Canadian Journal of Forest Research, 41, 1242-1255. [本文引用: 2]
DongLH, LiFR, JiaWW (2013). Effects of tree competition on biomass and biomass models of Pinus koraiensis plantation Journal of Beijing Forestry University, 35(6), 15-22. [本文引用: 1]
DulamsurenC, KhishigjargalM, LeuschnerC, HauckM (2014). Response of tree-ring width to climate warming and selective logging in larch forests of the Mongolian Altai Journal of Plant Ecology, 7, 24-38. [本文引用: 1]
GaoLL, GouXH, DengY, WangZQ, GuF, WangF (2018). Increased growth of Qinghai spruce in northwestern China during the recent warming hiatus Agricultural and Forest Meteorology, 260- 261, 9-16. [本文引用: 1]
Gómez-AparicioL, García-ValdésR, Ruíz-BenitoP, ZavalaMA (2011). Disentangling the relative importance of climate, size and competition on tree growth in Iberian forests: implications for forest management under global change Global Change Biology, 17, 2400-2414. [本文引用: 1]
HanDX, JinGZ (2017). Influences of topography and competition on DBH growth in different growth stages in a typical mixed broadleaved-Korean pine forest, northeastern China Journal of Beijing Forestry University, 39(1), 9-19. [本文引用: 1]
HolmesRL (1983). Computer-assisted quality control in tree-ring dating and measurement Tree-Ring Bulletin, 43, 69-78. [本文引用: 1]
HuangJG, StadtKJ, DawsonA, ComeauPG (2013). Modelling growth-competition relationships in trembling aspen and white spruce mixed boreal forests of Western Canada PLOS ONE, 8, e77607. DOI: 10.1371/journal.pone.0077607. URLPMID:24204891 [本文引用: 2]
HuangLP, GaoYQ, LiY, ZhangTW, HuDY, WangL (2015). Growth of Siberia larch in the middle east of Altay Mountains and its response to climate change Arid Land Geography, 38, 1169-1178. [本文引用: 1]
KangJ, JiangSW, HuangJG (2020). Radial growth response of four dominant tree species to climate factors in the Sayan Range of the Altai Mountains, Russia Acta Ecologica Sinica, 40, 6135-6146. [本文引用: 2]
KangYC, LiuBH, RenJY, TanKL (2019). Effect of competition on tree ring growth and climate factors of Pinus koraiensis Journal of Northeast Forestry University, 47(11), 6-10. [本文引用: 1]
KerhoulasLP, KaneJM (2012). Sensitivity of ring growth and carbon allocation to climatic variation vary within ponderosa pine trees Tree Physiology, 32, 14-23. URLPMID:22094578 [本文引用: 1]
KharukVI, RansonKJ, ImST, DvinskayaML (2009). Response of Pinus sibirica and Larix sibirica to climate change in southern Siberian alpine forest-tundra ecotone Scandinavian Journal of Forest Research, 24, 130-139. [本文引用: 1]
KwonS, PanLL, ShiZJ, YangXH, ZhangX, LiuYS, ZhangKB (2019). Radial growth of Mongolian pine and its response to climate at different competition intensities Chinese Journal of Ecology, 38, 1962-1972. [本文引用: 1]
LiZS, LiuGH, FuBJ, ZhangQB, HuCJ, LuoSZ (2011). Influence of different detrending methods on climate signal in tree-ring chronologies in Wolong National Natural Reserve, western Sichuan, China Chinese Journal of Plant Ecology, 35, 707-721. [本文引用: 1]
LiangHX, HuangJG, MaQQ, LiJY, WangZ, GuoXL, ZhuHX, JiangSW, ZhouP, YuBY, LuoDW (2019). Contributions of competition and climate on radial growth of Pinus massoniana in subtropics of China Agricultural and Forest Meteorology, 274, 7-17. [本文引用: 3]
LiuB, PanCD, LiGH, YuGB, ZhangF, GuoK, ZouZY (2019). Quantitative classification and sequencing of communities in pyrogenic succession of Kanas Taiga Ecology and Environmental Sciences, 28, 1961-1973. [本文引用: 2]
MaSY (2016). Research on the Response of Radial Growth to Thinning and Climatic Factors for Platycladus orientalis Plantation in Beijing Master degree dissertation, Beijing Forestry University, Beijing. [本文引用: 1]
NikolaevaSA, SavchukDA, BocharovAY (2015). Influence of different factors on tree growth of Pinus sibirica in the highlands of the central Altai Mountains Journal of Siberian Federal University Biology, 8, 299-318. [本文引用: 1]
PanY, BirdseyRA, FangJY, HoughtonR, KauppiPE, KurzWA, PhillipsOL, ShvidenkoA, LewisSL, CanadellJG, CiaisP, JacksonRB, PacalaSW, McGuireAD, PiaoSL, RautiainenA, SitchS, HayesD (2011). A large and persistent carbon sink in the world’s forests Science, 333, 988-993. DOI:10.1126/science.1201609URLPMID:21764754 [本文引用: 1] The terrestrial carbon sink has been large in recent decades, but its size and location remain uncertain. Using forest inventory data and long-term ecosystem carbon studies, we estimate a total forest sink of 2.4 +/- 0.4 petagrams of carbon per year (Pg C year(-1)) globally for 1990 to 2007. We also estimate a source of 1.3 +/- 0.7 Pg C year(-1) from tropical land-use change, consisting of a gross tropical deforestation emission of 2.9 +/- 0.5 Pg C year(-1) partially compensated by a carbon sink in tropical forest regrowth of 1.6 +/- 0.5 Pg C year(-1). Together, the fluxes comprise a net global forest sink of 1.1 +/- 0.8 Pg C year(-1), with tropical estimates having the largest uncertainties. Our total forest sink estimate is equivalent in magnitude to the terrestrial sink deduced from fossil fuel emissions and land-use change sources minus ocean and atmospheric sinks.
ShahS (2019). The Growth-climate Relationships and Potential Distribution of Siberian Pine (Pinus sibirica) in Northeast China PhD dissertation, Beijing Forestry University, Beijing. [本文引用: 1]
ShangHM, WeiWS, YuanYJ, YuSL, ZhangTW, AizemaitiW, LiXJ (2010). Response of tree ring width to recent climate change, south slope of Altai Mountains Acta Ecologica Sinica, 30, 2246-2253. [本文引用: 1]
StadtKJ, HustonC, CoatesKD, FengZL, DaleMRT, LieffersVJ (2007). Evaluation of competition and light estimation indices for predicting diameter growth in mature boreal mixed forests Annals of Forest Science, 64, 477-490. [本文引用: 1]
SidorovaOV, SaurerM, MyglanVS, EichlerA, SchwikowskiM, KirdyanovAV, BryukhanovaMV, GerasimovaOV, KaluginIA, DaryinAV, SiegwolfRTW (2012). A multi-proxy approach for revealing recent climatic changes in the Russian Altai Climate Dynamics, 38, 175-188. [本文引用: 1]
VelisevichSN, KozlovDS (2006). Effects of temperature and precipitation on radial growth of Siberian larch in ecotopes with optimal, insufficient, and excessive soil moistening Russian Journal of Ecology, 37, 241-246. [本文引用: 1]
WangZQ, WuGS, WangJB (2000). Application of competition index in assessing intraspecific and interspecific spatial relations between Manchurian ash and dahurian larch Chinese Journal of Applied Ecology, 11, 641-645. URLPMID:11767512 [本文引用: 1] The competition among trees within a stand includes aboveground and underground parts. How to determine the zone of competition influence is important for predicting tree growth. We divide the zone into two parts: one part is defined as inside radius, the distance from nearest competitors to subject tree, in which, the competition influence is the sum of both aboveground and underground; the another part is defined as outside radius, the distance of competitors to subject tree, where the competition influence comes mostly from underground. Thus, the competition index, based upon the divided two radii, could be decomposed into different components. We used the data of manchurian ash(MA) (Fraxinus mandshurica) and dahurian larch(DL) (Larix gmelini) from pure and mixed stands to analyze the competition influence of aboveground and underground in inside and of underground in outside radius. The results show that for MA in pure stand, the competition effect was 59.06% from inside radius, and 40.94% from outside radius. For DL in pure stand, the competition effects in both radii were 71.31% and 28.69%, respectively. There was a stronger competition influence in inside than in outside radius. Underground competitions were the important issues for both species in pure stands. In a mixed stand of MA and DL, there were two species in both radii, and the effects of competition came from intraspecific and interspecific competitors. For subject tree of MA, the effect of interspecific competition was 49.55%, and that of intraspecific was 50.45%. For subject tree of DL, the effects were 81.11% and 18.89%, respectively. DL displayed a stronger interspecific competition than MA in the zone of competition influence, and was a main effect of competition on MA. For subject tree of MA, the effect of competition from inside radius was 25.33%, which was the sum of competition influence from aboveground and underground, and about 74.67% was from outside radius. For subject tree of DL, both in inside and outside radii were 55.37% and 44.64%, respectively. Comparing with the information from both species and two radii, the root-root competition from underground was an important component. DL grew well, and could reduce the impact of intraspecific competition for MA when both species mixed in a stand. [ 王政权, 吴巩胜, 王军邦 (2000). 利用竞争指数评价水曲柳落叶松种内种间空间竞争关系 应用生态学报, 11, 641-645.] PMID:11767512 [本文引用: 1]
WigleyTML, BriffaKR, JonesPD (1984). On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology Journal of Climate and Applied Meteorology, 23, 201-213. [本文引用: 1]
XuanHC, GuoMZ, GaoLS, FanCY (2020). Effect of competition environment changes on the radial growth of Pinus koraiensis and Fraxinus mandshurica in mixed coniferous-broad-leaved forest Acta Ecologica Sinica, 40, 4087-4093. [本文引用: 1]
YuDP, WangSZ, TangLN, DaiLM, WangQL, WangSX (2005). Relationship between tree-ring chronology of Larix olgensis in Changbai Mountains and the climate change Chinese Journal of Applied Ecology, 16, 14-20. [本文引用: 1]
ZangC, BiondiF (2015). Treeclim: an R package for the numerical calibration of proxy-climate relationships Ecography, 38, 431-436. [本文引用: 1]
ZhangDL, FengZD, YangYP, LanB, RanM, MuGJ (2018). Peat δ13C celluose-recorded wetting trend during the past 8000 years in the southern Altai Mountains, northern Xinjiang, NW China Journal of Asian Earth Sciences, 156, 174-179. [本文引用: 1]
ZhangJ, HuangS, HeFL (2015). Half-century evidence from western Canada shows forest dynamics are primarily driven by competition followed by climate Proceedings of the National Academy of Sciences of the United States of America, 112, 4009-4014. DOI:10.1073/pnas.1420844112URLPMID:25775576 [本文引用: 1] Tree mortality, growth, and recruitment are essential components of forest dynamics and resiliency, for which there is great concern as climate change progresses at high latitudes. Tree mortality has been observed to increase over the past decades in many regions, but the causes of this increase are not well understood, and we know even less about long-term changes in growth and recruitment rates. Using a dataset of long-term (1958-2009) observations on 1,680 permanent sample plots from undisturbed natural forests in western Canada, we found that tree demographic rates have changed markedly over the last five decades. We observed a widespread, significant increase in tree mortality, a significant decrease in tree growth, and a similar but weaker trend of decreasing recruitment. However, these changes varied widely across tree size, forest age, ecozones, and species. We found that competition was the primary factor causing the long-term changes in tree mortality, growth, and recruitment. Regional climate had a weaker yet still significant effect on tree mortality, but little effect on tree growth and recruitment. This finding suggests that internal community-level processes-more so than external climatic factors-are driving forest dynamics.
ZouWT, ZengWS, ZhangLJ, ZengM (2015). Modeling crown biomass for four pine species in China Forests, 6, 433-449. [本文引用: 1]
Disentangling the relative importance of climate, size and competition on tree growth in Iberian forests: implications for forest management under global change 1 2011
Evaluation of competition and light estimation indices for predicting diameter growth in mature boreal mixed forests 1 2007
... 以每株目标树为中心, 将其半径4 m范围内的所有立木作为竞争树, 测量并记录所有树的胸径(Zou et al., 2015).利用公式计算以下3种竞争指数用来表征目标树受到的竞争强度(Stadt et al., 2007).包括竞争树密度(N, 株·hm-2)、竞争树胸径和(SDBH, m·hm-2)、竞争树胸高断面积和(SBA, m2·hm-2) (Huang et al., 2013).单木竞争指数公式为: ...
A multi-proxy approach for revealing recent climatic changes in the Russian Altai 1 2012
... 西比利亚红松的树木径向生长主要受到3月份气温的促进作用, 这一结果与同在喀纳斯保护区开展的西伯利亚五针松径向生长与气候因子关系的研究结果一致(Shah, 2019).但与阿尔泰山其他区域的研究结果不一致.阿尔泰山中部的西伯利亚五针松与气候的响应研究表明, 6月份气温对树木径向生长起直接作用(Kharuk et al., 2009; Nikolaeva et al., 2015).在俄罗斯境内阿尔泰山的研究发现, 西伯利亚五针松的树木径向生长主要受到3月降水量和7月气温的积极影响(康剑等, 2020).造成研究结果不一致的原因可能是小气候环境的差异所导致的.阿尔泰山由于地处温带与北寒带的过渡地带, 且山脉地形复杂多变, 其南北两侧气候差异较大(Sidorova et al., 2012; Zhang et al., 2018).本研究区域位于中国境内的阿尔泰山区域, 属于阿尔泰山中段南坡, 因此, 局部区域的小气候环境与其他区域存在差异.3月正值早春季节, 为树木的生长季前期, 此时研究区寒冷且多积雪, 土壤冻结, 不利于树木根系的吸收与活动.因此, 气温升高有利于土壤解冻, 同时导致融雪发生, 从而加快树木根系开始恢复生理活动, 进而促进树木径向生长(于大炮等, 2005). ...
Effects of temperature and precipitation on radial growth of Siberian larch in ecotopes with optimal, insufficient, and excessive soil moistening 1 2006
 ,1,2,3,*
,1,2,3,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT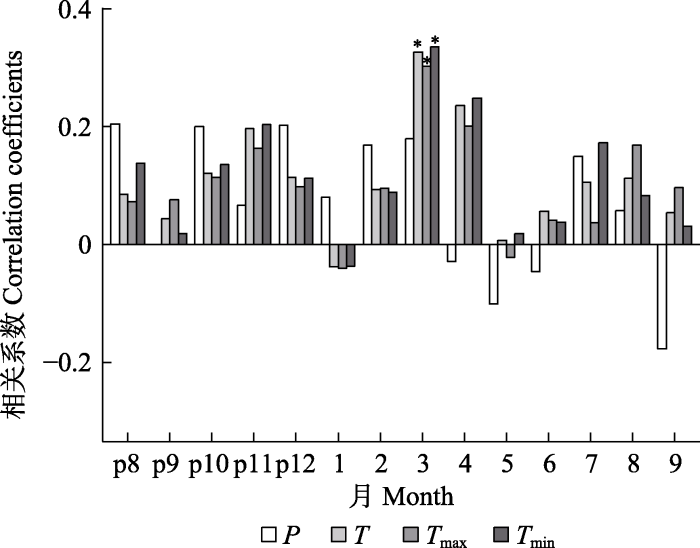 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT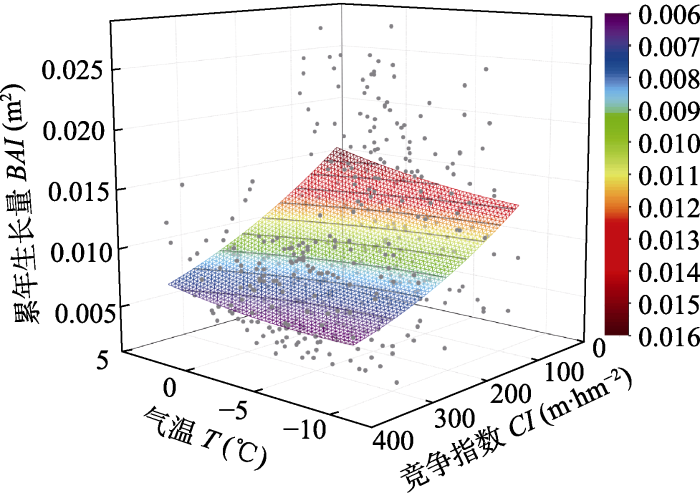 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}