Ecophysiological adaptability of four tree species in the southern subtropical evergreen broad-leaved forest to warming
Xu LI,, Ting WU, Yan CHENG, Na-Dan TAN, Fen JIANG, Shi-Zhong LIU, Guo-Wei CHU, Ze MENG, Ju-Xiu LIU,*Key Laboratory of Vegetation Restoration and Management of Degraded Ecosystems, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China
National Natural Science Foundation of China(41977287) National Natural Science Foundation of China(41991285) Special Forestry Project of Guangdong Province (Monitoring and Research on the Impact of Environmental Change on wild Plant Diversity)
Abstract Aims The subject of this study was to investigate warming effects on leaf stomatal traits, anatomical structure and photosynthetic traits of four common tree species in subtropical evergreen broad-leaved forest of southern China, and to compare their physiological adaptability to warming. Our study aims to provide a theoretical basis for better predicting the tree growth of native forests in a warming climate. Methods One-year-old seedlings of Syzygium rehderianum, Ormosia pinnata, Castanopsis hystrix and Schima superba were selected and exposed to two levels of temperature (ambient temperature and infrared heater warming). Leaf stomatal traits, anatomical structure and photosynthetic characteristics were measured to represent the abilities of stomatal regulation, leaf tissue regulation and nutrient maintenance, respectively. Important findings For Syzygium rehderianum, warming decreased its leaf sponge tissue thickness, photosynthetic nitrogen-use efficiency (PNUE) and photosynthetic phosphorous-use efficiency (PPUE). Seedling of O. pinnata exposed to warming showed increased stomatal conductance, photosynthetic rate, PNUE and PPUE, but decreased stomatal density, leaf thickness and palisade tissue thickness. For C. hystrix, warming decreased the stomata size, but did not affect its photosynthetic rate. Seedling of Schima superba exposed to warming showed lower stomata density, leaf palisade tissue thickness, photosynthetic rate, PNUE and PPUE, but higher stomata size. These results suggested that O. pinnata, Syzygium rehderianum and Schima superba could reduce their leaf thickness to acclimate to warming conditions. The abilities of stomatal regulation, nutrient maintenance, photosynthetic rate and PNUE varied among these tree species. Warming would be beneficial for the growth of O. pinnata due to increased photosynthetic rate, PNUE and PPUE, while not for Syzygium rehderianum and Schima superba, the two dominant tree species of native forests. This study indicated that, with projected climate change, O. pinnata may replace Syzygium rehderianum and Schima superba as a new dominant tree species in the subtropical evergreen broad-leaved forest for its stronger adaptability to warming. Keywords:infrared heating;stomatal size;stomatal density;leaf anatomical structure;photosynthesis;subtropical tree species
PDF (2289KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 李旭, 吴婷, 程严, 谭钠丹, 蒋芬, 刘世忠, 褚国伟, 孟泽, 刘菊秀. 南亚热带常绿阔叶林4个树种对增温的生理生态适应能力比较. 植物生态学报, 2020, 44(12): 1203-1214. DOI: 10.17521/cjpe.2020.0318 LI Xu, WU Ting, CHENG Yan, TAN Na-Dan, JIANG Fen, LIU Shi-Zhong, CHU Guo-Wei, MENG Ze, LIU Ju-Xiu. Ecophysiological adaptability of four tree species in the southern subtropical evergreen broad-leaved forest to warming. Chinese Journal of Plant Ecology, 2020, 44(12): 1203-1214. DOI: 10.17521/cjpe.2020.0318
CT, 角质层厚度; LT, 叶片厚度; LET, 下表皮厚度; PT, 栅栏组织; ST, 海绵组织厚度; UET, 上表皮厚度。 Fig. 1Leaf anatomical structure and stomatal characteristics of four tree species in southern subtropical evergreen broad-leaved forest.
新窗口打开|下载原图ZIP|生成PPT 图3增温对南亚热带常绿阔叶林4个树种的气孔导度(Gs)、气孔密度(SD)和气孔大小(SS)的影响(平均值+标准误差, n = 6)。*, p < 0.05; **, p < 0.01; ***, p < 0.001。
Fig. 3Effects of warming on stomatal conductance (Gs), stomatal density (SD), and stomatal size (SS) of four tree species in southern subtropical evergreen broad-leaved forest (mean + SE, n = 6).*, p < 0.05; **, p < 0.01; ***, p < 0.001.
Table 1 表1 表1增温(W)、树种(S)及其交互作用对南亚热带常绿阔叶林4个树种叶片解剖结构和光合特性的双因素方差分析 Table 1Effects of warming (W), tree species (S) and their interactions on the leaf anatomical structure and photosynthetic characters of four tree species in southern subtropical evergreen broad-leaved forest
新窗口打开|下载原图ZIP|生成PPT 图4增温对南亚热带常绿阔叶林4个树种叶片厚度(LT)、栅栏组织厚度(PT)和海绵组织厚度(ST)的影响的箱型图。图中的数据节点依次为上限、上四分位数、中位数、下四分位数和下限。圆圈为数据点。*, p < 0.05; **, p < 0.01; ***, p < 0.001; n = 9。
Fig. 4Box plots illustrating the effects of warming on leaf thickness (LT), palisade tissue thickness (PT) and sponge tissue thickness (ST) of four tree species in southern subtropical evergreen broad-leaved forest. Whiskers of box plots indicates upper extreme, upper quartile, median, lower quartile and lower extreme, respectively. Each circle represents one individual tree. *, p < 0.05; **, p < 0.01; ***, p < 0.001; n = 9.
图5
新窗口打开|下载原图ZIP|生成PPT 图5增温对南亚热带常绿阔叶林4个树种叶片上、下表皮厚度(UET、LET)和角质层厚度(CT)的影响的箱型图。图中的数据节点依次为上限、上四分位数、中位数、下四分位数和下限。圆圈为数据点。*, p < 0.05; **, p < 0.01; ***, p < 0.001; n = 9。
Fig. 5Box plots illustrating the effects of warming on leaf upper epidermis thickness (UET), lower epidermis thickness (LET) and cuticle thickness (CT) of four tree species in southern subtropical evergreen broad-leaved forest. Whiskers of box plots indicates upper extreme, upper quartile, median, lower quartile and lower extreme, respectively. Each circle represents one individual tree. *, p < 0.05; **, p < 0.01; ***, p < 0.001; n = 9.
新窗口打开|下载原图ZIP|生成PPT 图6增温对南亚热带常绿阔叶林4个树种的光合速率(Pn)、蒸腾速率(Tr)和水分利用效率(WUE)的影响(平均值+标准误差, n = 6)。*, p < 0.05; **, p < 0.01; ***, p < 0.001。
Fig. 6Effects of warming on photosynthetic rate (Pn), transpiration rate (Tr), and water use efficiency (WUE) of four tree species in southern subtropical evergreen broad-leaved forest in 2018 and 2019 (mean + SE, n = 6). *, p < 0.05; **, p < 0.01; ***, p < 0.001.
图7
新窗口打开|下载原图ZIP|生成PPT 图7增温对南亚热带常绿阔叶林4个树种的光合氮利用效率(PNUE)和光合磷利用效率(PPUE)的影响(平均值+标准误差, n = 6)。*, p < 0.05; **, p < 0.01; ***, p < 0.001。
Fig. 7Effects of warming on the photosynthetic nitrogen-use efficiency (PNUE) and photosynthetic phosphorous-use efficiency (PPUE) of four tree species in southern subtropical evergreen broad-leaved forest in 2018 and 2019 (mean + SE, n = 6). *, p < 0.05; **, p < 0.01; ***, p < 0.001.
以Pn与Gs线性回归的斜率作为植物的光合敏感性(Wu et al., 2018)。回归分析表明增温改变了4个树种的光合敏感性(表2), 增温下海南红豆和红锥的光合敏感性由显著相关(p < 0.05)变为不相关, 增温下红枝蒲桃和木荷的光合敏感性有降低的趋势, 但未达到显著水平。
Table 2 表2 表2增温和对照条件下南亚热带常绿阔叶林4个树种光合速率和气孔导度的关系(一元线性回归) Table 2Relationship between photosynthesis rate and stomatal conductance under control and warming environments of four tree species in southern subtropical evergreen broad-leaved forest (unary linear regression)
Table 3 表3 表3南亚热带常绿阔叶林4个树种叶片解剖结构特征与光合特征相关关系 Table 3Correlations between leaf anatomical structure and photosynthetic characters of four tree species in southern subtropical evergreen broad-leaved forest
BremnerJ, MulvaneyC (1982). Nitrogen-total//Page AL. Methods of Soil Analysis, Part 2: Chemical and Microbiological Properties 2nd ed. American Society of Agronomy, Soil Science Society of America, Madison. [本文引用: 1]
CavaleriMA, ReedSC, SmithWK, WoodTE (2015). Urgent need for warming experiments in tropical forests Global Change Biology, 21, 2111-2121. DOI:10.1111/gcb.12860URLPMID:25641092 [本文引用: 1] Although tropical forests account for only a fraction of the planet's terrestrial surface, they exchange more carbon dioxide with the atmosphere than any other biome on Earth, and thus play a disproportionate role in the global climate. In the next 20 years, the tropics will experience unprecedented warming, yet there is exceedingly high uncertainty about their potential responses to this imminent climatic change. Here, we prioritize research approaches given both funding and logistical constraints in order to resolve major uncertainties about how tropical forests function and also to improve predictive capacity of earth system models. We investigate overall model uncertainty of tropical latitudes and explore the scientific benefits and inevitable trade-offs inherent in large-scale manipulative field experiments. With a Coupled Model Intercomparison Project Phase 5 analysis, we found that model variability in projected net ecosystem production was nearly 3 times greater in the tropics than for any other latitude. Through a review of the most current literature, we concluded that manipulative warming experiments are vital to accurately predict future tropical forest carbon balance, and we further recommend the establishment of a network of comparable studies spanning gradients of precipitation, edaphic qualities, plant types, and/or land use change. We provide arguments for long-term, single-factor warming experiments that incorporate warming of the most biogeochemically active ecosystem components (i.e. leaves, roots, soil microbes). Hypothesis testing of underlying mechanisms should be a priority, along with improving model parameterization and constraints. No single tropical forest is representative of all tropical forests; therefore logistical feasibility should be the most important consideration for locating large-scale manipulative experiments. Above all, we advocate for multi-faceted research programs, and we offer arguments for what we consider the most powerful and urgent way forward in order to improve our understanding of tropical forest responses to climate change.
ChenL, WenYG, ZengJ, WangH, WangJX, DellB, LiuSR (2019). Differential responses of net N mineralization and nitrification to throughfall reduction in a Castanopsis hystrix plantation in Southern China Forest Ecosystems, 6, 1-11. [本文引用: 1]
DingYH, WangHJ (2016). Newly acquired knowledge on the scientific issues related to climate change over the recent 100 years in China Chinese Science Bulletin, 61, 1029-1041. [本文引用: 1]
DusengeME, WayDA (2017). Warming puts the squeeze on photosynthesis—Lessons from tropical trees Journal of Experimental Botany, 68, 2073-2077. [本文引用: 1]
EllsworthDS, ReichPB (1996). Photosynthesis and leaf nitrogen in five Amazonian tree species during early secondary succession Ecology, 77, 581-594. [本文引用: 1]
EscuderoA, MediavillaS (2003). Decline in photosynthetic nitrogen use efficiency with leaf age and nitrogen resorption as determinants of leaf life span Journal of Ecology, 91, 880-889. [本文引用: 1]
FengYL, FuGL, ZhengYL (2008). Specific leaf area relates to the differences in leaf construction cost, photosynthesis, nitrogen allocation, and use efficiencies between invasive and noninvasive alien congeners Planta, 228, 383-390. URLPMID:18392694 [本文引用: 1]
FranksPJ, BeerlingDJ (2009). Maximum leaf conductance driven by CO2 effects on stomatal size and density over geologic time Proceedings of the National Academy of Sciences of the United States of America, 106, 10343-10347. [本文引用: 3]
FranksPJ, FarquharGD (2007). The mechanical diversity of stomata and its significance in gas-exchange control Plant Physiology, 143, 78-87. URLPMID:17114276 [本文引用: 2]
HepworthC, Doheny-AdamsT, HuntL, CameronDD, GrayJE (2015). Manipulating stomatal density enhances drought tolerance without deleterious effect on nutrient uptake New Phytologist, 208, 336-341. [本文引用: 1]
HetheringtonAM, WoodwardFI (2003). The role of stomata in sensing and driving environmental change Nature, 424, 901-908. URLPMID:12931178 [本文引用: 2]
HuJY, GuoK, DongM (2008). Variation of leaf structure of two dominant species in alpine grassland and the relationship between leaf structure and ecological factors Chinese Journal of Plant Ecology, 32, 370-378. [本文引用: 1]
HuangJ, ChenC, ZhangWX, DingCJ, SuXH, HuangQJ (2017). Effects of drought stress on anatomical structure and photosynthetic characteristics of transgenic JERF36 Populus alba × P. berolinensis seedling leaves Scientia Silvae Sinicae, 53(5), 8-15. [本文引用: 1]
JamesSA, BellDT (2000). Influence of light availability on leaf structure and growth of two Eucalyptus globulus ssp. globulus provenances Tree Physiology, 20, 1007-1018. URLPMID:11305455 [本文引用: 1]
LiDJ, ZhouXH, WuLY, ZhouJZ, LuoYQ (2013). Contrasting responses of heterotrophic and autotrophic respiration to experimental warming in a winter annual-dominated prairie Global Change Biology, 19, 3553-3564. URLPMID:23728995 [本文引用: 1]
LiW, FuZ, HaoXZ, LiQY, ZhangCP (2020). Leaf anatomical structure and photosynthetic characteristics of Megaskepasma erythrochlamys and Pachystachys lutea in greenhouse Chinese Agricultural Science Bulletin, 36, 58-61. [本文引用: 1]
LiuL (2016). Study on the Photosynthetic Characteristics of Liangshan Introduced Olive (Olea europaea L.) Cultivars Master degree dissertation, Sichuan Agricultural University, Ya’an, Sichuan. [本文引用: 1]
LiuJX, HuangWJ, ZhouGY, ZhangDQ, LiuSZ, LiYY (2013). Nitrogen to phosphorus ratios of tree species in response to elevated carbon dioxide and nitrogen addition in subtropical forests Global Change Biology, 19, 208-216. URLPMID:23504732 [本文引用: 1]
LiuJX, LiYL, LiuSZ, LiYY, ChuGW, MengZ, ZhangDQ (2013). An introduction to an experimental design for studying effects of air temperature rise on model forest ecosystems Chinese Journal of Plant Ecology, 37, 558-565. [本文引用: 3]
Méndez-AlonzoR, EwersFW, JacobsenAL, PrattRB, ScoffoniC, BartlettMK, SackL (2019). Covariation between leaf hydraulics and biomechanics is driven by leaf density in Mediterranean shrubs Trees, 33, 507-519. [本文引用: 1]
NiuSL, HanXG, MaKP, WanSQ (2007). Field facilities in global warming and terrestrial ecosystem research Journal of Plant Ecology (Chinese Version), 31, 262-271. [本文引用: 1]
ReichPB, WaltersMB, EllsworthDS, UhlC (1994). Photosynthesis-nitrogen relations in Amazonian tree species Oecologia, 97, 62-72. [本文引用: 1]
RobinsonDE, WagnerRG, BellFW, SwantonCJ (2001). Photosynthesis, nitrogen-use efficiency, and water-use efficiency of jack pine seedlings in competition with four boreal forest plant species Canadian Journal of Forest Research, 31, 2014-2025. DOI:10.1139/x01-133URL [本文引用: 1]
RoyerDL, MillerIM, PeppeDJ, HickeyLJ (2010). Leaf economic traits from fossils support a weedy habit for early angiosperms American Journal of Botany, 97, 438-445. URLPMID:21622407 [本文引用: 1]
SackL, CowanPD, JaikumarN, HolbrookNM (2003). The ‘hydrology’ of leaves: co-ordination of structure and function in temperate woody species Plant, Cell & Environment, 26, 1343-1356. [本文引用: 1]
ScoffoniC, RawlsM, McKownA, CochardH, SackL (2011). Decline of leaf hydraulic conductance with dehydration: relationship to leaf size and venation architecture Plant Physiology, 156, 832-843. DOI:10.1104/pp.111.173856URLPMID:21511989 [本文引用: 1]
SigurdssonBD, MedhurstJL, WallinG, EggertssonO, LinderS (2013). Growth of mature boreal Norway spruce was not affected by elevated [CO2] and/or air temperature unless nutrient availability was improved Tree Physiology, 33, 1192-1205. URLPMID:23878169 [本文引用: 1]
SlotM, WinterK (2017). Photosynthetic acclimation to warming in tropical forest tree seedlings Journal of Experimental Botany, 68, 2275-2284. URLPMID:28453647 [本文引用: 3]
StinzianoJR, HünerNPA, WayDA (2015). Warming delays autumn declines in photosynthetic capacity in a boreal conifer, Norway spruce (Picea abies) Tree Physiology, 35, 1303-1313. [本文引用: 1]
SuJ, SunB, WangDZ (1995). The biological characteristics and ornamental value of Ormosia pinnata Forest Research, 8, 677-681. [本文引用: 1]
TanakaY, SuganoSS, ShimadaT, Hara-NishimuraI (2013). Enhancement of leaf photosynthetic capacity through increased stomatal density in Arabidopsis. New Phytologist, 198, 757-764. [本文引用: 1]
TangB, YinCY, WangYJ, SunYY, LiuQ (2016). Positive effects of night warming on physiology of coniferous trees in late growing season: leaf and root Acta Oecologica, 73, 21-30. [本文引用: 1]
UpadhyayRK, SoniDK, SinghR, DwivediUN, PathreUV, NathP, SaneAP (2013). SIERF36, an EAR-motif-containing ERF gene from tomato, alters stomatal density and modulates photosynthesis and growth Journal of Experimental Botany, 64, 3237-3247. URLPMID:23840010 [本文引用: 1]
vanOmmen Kloeke AEE, DoumaJC, Ordo?ezJC, ReichPB, vanBodegom PM (2012). Global quantification of contrasting leaf life span strategies for deciduous and evergreen species in response to environmental conditions Global Ecology and Biogeography, 21, 224-235. [本文引用: 1]
WuGL, LiuH, HuaL, LuoQ, LinYX, HePC, FengSW, LiuJX, YeQ (2018). Differential responses of stomata and photosynthesis to elevated temperature in two co-occurring subtropical forest tree species Frontiers in Plant Science, 9, 467. DOI: 10.3389/fpls.2018.00467. URLPMID:29740458 [本文引用: 5]
YeWM, XiongDC, YangZJ, ZhuYG, ZhangQF, LiuXF, LinWS, XuC, ZhangJ, YangYS (2019). Effect of soil warming on growth and photosynthetic characteristics of Cunninghamia lanceolata saplings Acta Ecologica Sinica, 39, 2501-2509. [本文引用: 2]
YeWM (2019). Effects of Simulated Warming on Photosynthetic Characteristics and Product Distribution of Cunninghamia lanceolata Master degree dissertation, Fujian Normal University, Fuzhou. [本文引用: 1]
ZhengYP, XuM, HouRX, ShenRC, QiuS, OuyangZ (2013). Effects of experimental warming on stomatal traits in leaves of maize (Zea may L.) Ecology and Evolution, 3, 3095-3111. [本文引用: 1]
ZhouGY, PengCH, LiYL, LiuSZ, ZhangQM, TangXL, LiuJX, YanJH, ZhangDQ, ChuGW (2013). A climate change-induced threat to the ecological resilience of a subtropical monsoon evergreen broad-leaved forest in Southern China Global Change Biology, 19, 1197-1210. URLPMID:23504896 [本文引用: 2]
ZhouGY, WeiXH, WuYP, LiuSG, HuangYH, YanJH, ZhangDQ, ZhangQM, LiuJX, MengZ, WangCL, ChuGW, LiuSZ, TangXL, LiuXD (2011). Quantifying the hydrological responses to climate change in an intact forested small watershed in Southern China Global Change Biology, 17, 3736-3746. [本文引用: 1]
Nitrogen-total//Page AL. Methods of Soil Analysis, Part 2: Chemical and Microbiological Properties 1 1982
Photosynthesis and leaf nitrogen in five Amazonian tree species during early secondary succession 1 1996
... 由于在自然生态系统中常常缺乏可利用的N, 对植物的生长造成限制(Onoda et al., 2017), 因此具有较高PNUE的植物常常具有更高的生长速率(Robinson et al., 2001)及在生态系统中较强的竞争能力, 同时高PNUE也有利于植物的进化(Reich et al., 1994).而较低PNUE的植物叶片则常常具有较高的耐受环境胁迫的能力(Ellsworth & Reich, 1996; van Ommen et al., 2012)及较高的叶片寿命(Onoda et al., 2017), 能够使得叶片在整个生命周期中充分利用其内部的N进行光合作用(Escudero & Mediavilla, 2003).我们发现增温条件下红枝蒲桃和木荷的PNUE和PPUE降低, 红锥的PNUE和PPUE无显著变化, 而海南红豆的PNUE和PPUE升高, 这表明红枝蒲桃和木荷为适应高温环境, 采取了降低PNUE和PPUE的策略, 而海南红豆却表现出对高温环境的良好适应性, 在未来气候变暖的情况下在该生态系统中可能具有较强的竞争能力.基于鼎湖山站长期的原位监测也发现, 在全球气候变化下南亚热带常绿阔叶林有向灌丛化演替的趋势(Zhou et al., 2013).此外, 由于海南红豆是豆科植物, 较好的N吸收能力也可能是其PNUE升高的原因(Liu et al., 2013). ...
Decline in photosynthetic nitrogen use efficiency with leaf age and nitrogen resorption as determinants of leaf life span 1 2003
... 由于在自然生态系统中常常缺乏可利用的N, 对植物的生长造成限制(Onoda et al., 2017), 因此具有较高PNUE的植物常常具有更高的生长速率(Robinson et al., 2001)及在生态系统中较强的竞争能力, 同时高PNUE也有利于植物的进化(Reich et al., 1994).而较低PNUE的植物叶片则常常具有较高的耐受环境胁迫的能力(Ellsworth & Reich, 1996; van Ommen et al., 2012)及较高的叶片寿命(Onoda et al., 2017), 能够使得叶片在整个生命周期中充分利用其内部的N进行光合作用(Escudero & Mediavilla, 2003).我们发现增温条件下红枝蒲桃和木荷的PNUE和PPUE降低, 红锥的PNUE和PPUE无显著变化, 而海南红豆的PNUE和PPUE升高, 这表明红枝蒲桃和木荷为适应高温环境, 采取了降低PNUE和PPUE的策略, 而海南红豆却表现出对高温环境的良好适应性, 在未来气候变暖的情况下在该生态系统中可能具有较强的竞争能力.基于鼎湖山站长期的原位监测也发现, 在全球气候变化下南亚热带常绿阔叶林有向灌丛化演替的趋势(Zhou et al., 2013).此外, 由于海南红豆是豆科植物, 较好的N吸收能力也可能是其PNUE升高的原因(Liu et al., 2013). ...
Specific leaf area relates to the differences in leaf construction cost, photosynthesis, nitrogen allocation, and use efficiencies between invasive and noninvasive alien congeners 1 2008
 ,, 吴婷, 程严, 谭钠丹, 蒋芬, 刘世忠, 褚国伟, 孟泽, 刘菊秀
,, 吴婷, 程严, 谭钠丹, 蒋芬, 刘世忠, 褚国伟, 孟泽, 刘菊秀 ,*中国科学院华南植物园退化生态系统植被恢复与管理重点实验室, 广州 510650
,*中国科学院华南植物园退化生态系统植被恢复与管理重点实验室, 广州 510650
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT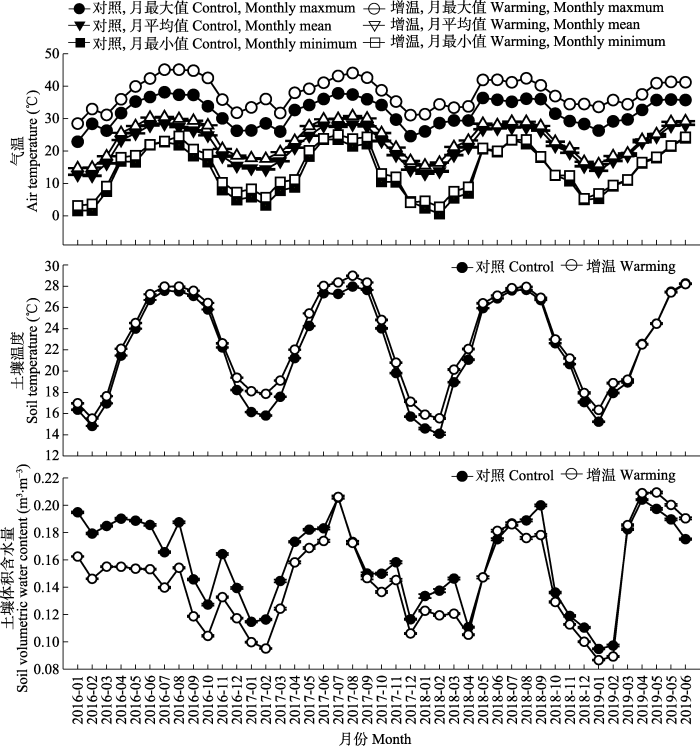 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT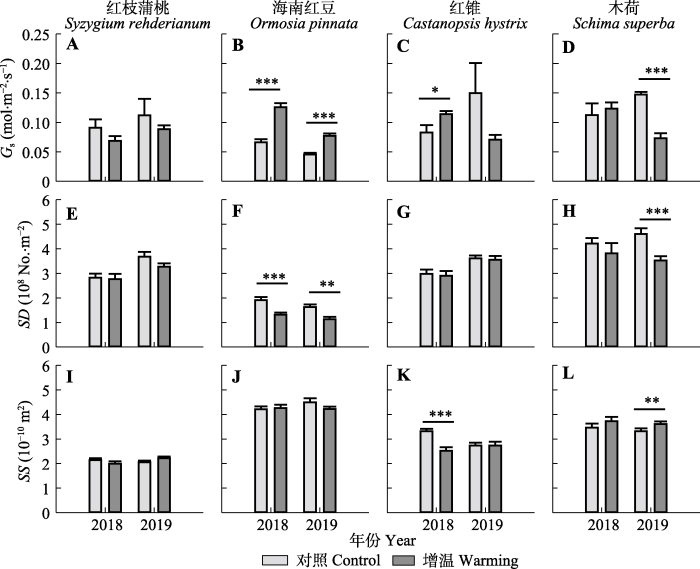 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT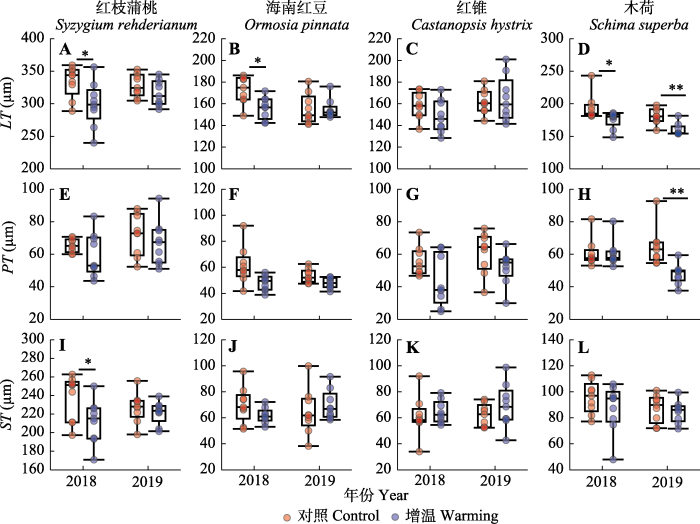 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT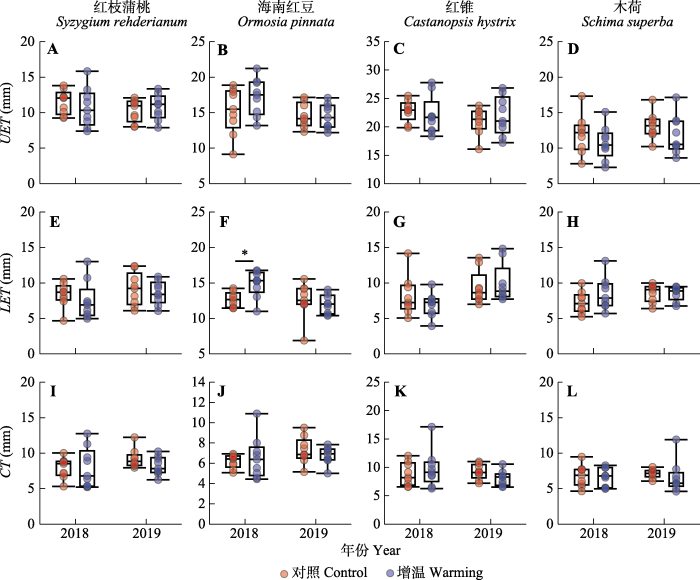 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT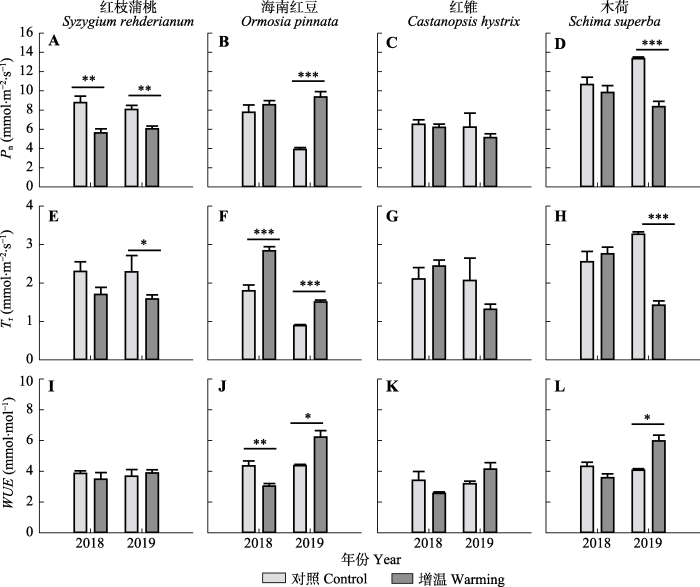 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT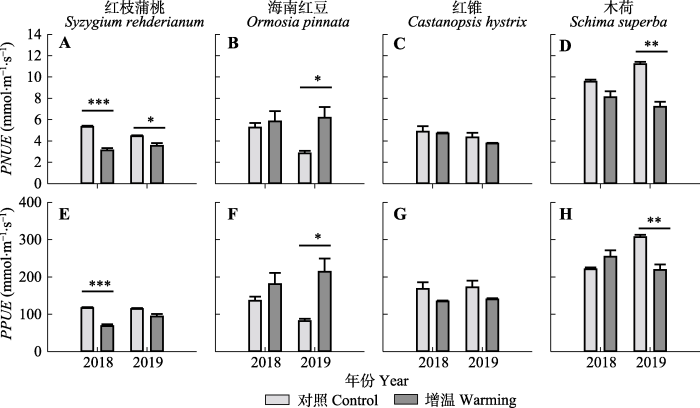 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}