<script type="text/javascript" src="https://cdn.bootcss.com/mathjax/2.7.2-beta.0/MathJax.js?config=TeX-AMS-MML_HTMLorMML"></script>
<script type='text/x-mathjax-config'>
MathJax.Hub.Config({
extensions: ["tex2jax.js"],
jax: ["input/TeX", "output/HTML-CSS"],
tex2jax: {inlineMath: [ ['$','$'], ["\\(","\\)"] ],displayMath: [ ['$$','$$'], ["\\[","\\]"] ],processEscapes: true},
"HTML-CSS": { availableFonts: ["TeX"] },
TeX: {equationNumbers: {autoNumber: ["none"], useLabelIds: true}},
"HTML-CSS": {linebreaks: {automatic: true}},
SVG: {linebreaks: {automatic: true}}
});
</script>
白天道, 余春兰, 甘泽朝, 赖海荣, 杨隐超, 黄厚宸, 蒋维昕
, * 广西大学林学院, 国家林业与草原局中南速生材繁育重点实验室, 南宁 530004
Association of cone and seed traits of Pinus yunnanensis var. tenuifolia with geo-meteorological factors Tian-Dao BAI, Chun-Lan YU, Ze-Chao GAN, Hai-Rong LAI, Yin-Chao YANG, Hou-Chen HUANG, Wei-Xin JIANG
, * College of Forestry, Guangxi University, Key Laboratory of National Forestry and Grassland Administration for Fast-growing Wood Breeding and Cultivation in Central and South China, Nanning 530004, China
通讯作者: *(
jwx_1985@163.com )
编委: 罗天祥
责任编辑: 李敏
收稿日期: 2020-08-6
接受日期: 2020-10-26
网络出版日期: 2020-12-20
基金资助: 广西自然科学基金 (2018GXNSFBA281110 )国家自然科学基金 (31400575 )
Corresponding authors: *(
jwx_1985@163.com )
Received: 2020-08-6
Accepted: 2020-10-26
Online: 2020-12-20
Fund supported: Natural Science Foundation of Guangxi (2018GXNSFBA281110 )National Natural Science Foundation of China (31400575). (31400575 )
摘要 细叶云南松(
Pinus yunnanensis var.
tenuifolia )是分布于滇、黔、桂交界处的一个云南松变种, 其自然生境独特, 属典型的干热河谷气候, 具有重要的生态、经济价值。该研究以分布于南盘江-红水河流域沿线的8个细叶云南松天然种群为材料, 采用描述性统计及巢式方差分析比较种群间及种群内种实性状差异, 运用Pearson相关及典型相关分析探究种实性状与地理气象因子间的相关性, 最后对种群进行主成分聚类和Mantel检验, 揭示其种实性状地理变异模式。结果显示: 细叶云南松11个种实性状在种群间和种群内都存在极显著差异(
p < 0.001), 变异丰富。种实性状以种群内变异为主(平均表型分化系数
VST = 18.65%), 球果性状种群间分化(24.22%-39.88%)高于种子及种翅性状(4.14%-13.80%), 表明球果性状受到更强的环境选择。尽管部分相关系数未达显著水平, 但整体上种实性状与经纬度、年平均气温呈正相关关系, 与相对湿度、年降水量呈负相关关系, 表明细叶云南松种实性状受到地理隔离、湿度和温度的协同选择作用, 使其能较好适应干热环境。主成分聚类将参试种群划分为3类, 位于东部的罗甸伍家坟(WJ)、罗甸大亭(DT)种群聚为一类, 其种实形态较大, 位于西南部的兴义坝汪(BW)种群单独一类, 其种实形态较小, 其他种群聚为一类, 种实形态介于前两类之间。总体上, 种实性状值有自西向东递增的趋势。Mantel检验表明, 参试种群存在明显的空间结构, 主要体现为渐变群模式。
关键词: 细叶云南松 ;
球果性状 ;
种子性状 ;
地理变异 ;
生态适应性 ;
遗传资源 Abstract Aims Pinus yunnanensis var.
tenuifolia is an ecologically and economically important timber tree located at the junction of Yunnan, Guizhou and Guangxi, China. The natural distribution area represents a typical habitat of hot-dry valley. This paper aimed to describe the association between variation patterns of cone and seed traits (CST) and the geo-meteorological factors, explore its ecological adaptability, and provide a reference for the genetic resources conservation, evaluation and utilization.
Methods We sampled eight wild populations of
P. yunnanensis var.
tenuifolia distributed along the Nanpan-Hongshui River basin. The CST among and within populations were analyzed via discriptive statistics and nested ANOVA. Correlations between CST and geo-meteorological were evaluated based on Pearson and canonical correlation coefficients. Principal component analysis and Mantel test were applied to reveal the geographic variation pattern.
Important findings Abundant variations of eleven CST among and within populations were indicated by the extremely significant difference of nested ANOVA results (
p < 0.001). The variation within population was the main source (the average coefficient of phenotypic differentiation
VST = 18.65%), and generally, the
VST of cone traits (24.22%-39.88%) were larger than those of seed and wing traits (4.14%-13.80%), indicating more environment selection pressure on the cone traits. Majority of CST was positively correlated (though part of variable-pairs statistically insignificant) with latitude and longitude, annual average temperature, and negatively correlated with relative humidity and annual rainfall. This result suggests that the CST of
P. yunnanensis var.
tenuifolia is subjected to strong environmental selection, especially to temperature and relative humidity, prompting the species to adapt the hot-dry environment. Eight natural populations of
P. yunnanensis var.
tenuifolia were divided into three groups via principal component and cluster analysis. The two Luodian populations (WJ and DT) in the east were grouped into one group, and their cones and seeds were large. A population (BW) of Xingyi in the southwest belonged to a separate group, and its cones and seeds were small. The other populations were grouped together, and their CST fell between the first two groups. On the whole, the CST in
P. yunnanensis var.
tenuifolia increased from west to east. Mantel test exhibited a significant spatial structure among populations, and the variation pattern of populations was consistent with that of the cline.
Keywords: Pinus yunnanensis var. tenuifolia ;
cone trait ;
seed trait ;
geographic variation ;
ecological adaptivity ;
genetic resources PDF (3051KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 白天道, 余春兰, 甘泽朝, 赖海荣, 杨隐超, 黄厚宸, 蒋维昕. 细叶云南松种实性状变异与地理气象因子的关联. 植物生态学报, 2020, 44(12): 1224-1235. DOI:
10.17521/cjpe.2020.0269 BAI Tian-Dao, YU Chun-Lan, GAN Ze-Chao, LAI Hai-Rong, YANG Yin-Chao, HUANG Hou-Chen, JIANG Wei-Xin.
Association of cone and seed traits of Pinus yunnanensis var. tenuifolia with geo-meteorological factors .
Chinese Journal of Plant Ecology , 2020, 44(12): 1224-1235. DOI:
10.17521/cjpe.2020.0269 细叶云南松(
Pinus yunnanensis var.
tenuifolia )是云南松(
P. yunnanensis )因南亚热带独特的河谷气候长期影响及适应下逐渐形成的一个地理变种, 是我国特有种, 其天然分布区域狭窄, 仅限于黔、滇、桂交界处的南盘江流域、北盘江流域以及桂西北的红水河流域, 属于微域分布的植被类型(
徐学良, 1983 ;
王献溥, 1991 )。细叶云南松生境狭窄, 属于年平均气温较高, 降水量小于蒸发量, 总体较为干旱的河谷区域, 焚风效应明显, 气候独特。独特的生境使细叶云南松具有较高的生态研究价值(
李治基和王献溥, 1981 )。此外, 细叶云南松干型直、树体高大, 出材率高, 木材强度和质量较好, 具有较高的开发利用前景。但近年来, 由于该区域其他经济林树种(油茶(
Camellia oleifera )、八角(
Illicium verum )等)的大力发展, 对细叶云南松生境的人为干扰急剧加重, 大量细叶云南松林遭到蚕食、破坏, 原有的诸多大面积连续分布林分已消失殆尽, 呈零星点状分布(
吴东山等, 2016 )。针对细叶云南松的遗传资源调查、收集、保存、评价迫在眉睫。
要开展细叶云南松遗传资源的收集和保护, 首先要掌握其生态特性及种群遗传变异规律。目前, 针对细叶云南松天然林的研究主要集中在林分结构、林下土壤特性等方面(
潘婷, 2018 ;
Yu et al ., 2018 ;
李远发等, 2019 ;
廖良宁等, 2019 )。针对细叶云南松遗传资源评价的研究相对较少, 仅见
杨章旗等(2014) 对广西西北部3个种群进行了SSR遗传多样性分析, 认为遗传多样性与欧洲栓皮栎(
Quercus suber )、黄杉(
Pseudotsuga menziesii )等相比普遍不高, 但种群间基因流动频繁, 分化较小。表型多样性是遗传与环境共同作用的结果, 研究不同地理环境条件下的天然种群的表型性状分化程度和变异规律, 不仅可在一定程度上反映其遗传变异水平, 也可以有效揭示生物对不同环境的适应, 进而探究其发展演化(
Donohue et al ., 2001 ;
Etterson, 2004 ;
Gitonga et al ., 2008 ;
Shirley & Vesk, 2009 )。种实性状在物种自然演化过程中具有较高的遗传稳定性, 不易受短期环境变化的影响。同时, 作为主要繁殖器官, 种实性状对物种的定居、繁衍及分布具有重要影响(
Cilas et al ., 2009 )。研究表明, 果实形态、种子大小、质量、种翅的有无等都会影响其种子传播媒介(风、动物等)的传播效率, 进而影响物种的分布范围(
Wall, 2008 ;
Zhang et al ., 2020 )。本文通过对细叶云南松自然分布区的8个天然种群开展种实性状分析, 以揭示细叶云南松种实性状在空间分布上的表型分化和变异规律, 并探究其与细叶云南松特殊生境的关系, 为进一步开展细叶云南松的种质资源保护与利用提供参考。
1 材料和方法 1.1 样本采集 根据中国植物志(
http://www.cvh.ac.cn/ )中细叶云南松的分布记录及实地调研情况, 本试验采集的样品集中分布于南盘江-红水河流域贵州境内的8个代表性细叶云南松天然种群, 采样时间为2019年10月中旬, 种群信息详见
表1 。采样方法: 每个种群选择15株生长正常、结实较多、树干通直、无明显病虫害的植株作为采种母树, 分别测量母树胸径、海拔、经纬度等信息。为保证采种母树的代表性, 降低采种母树间的亲缘关系带来的系统误差, 每个种群内采种单株间距在80 m以上。使用高空采样器(20 m)采集树冠中上部的发育正常的成熟球果, 每株随机摘取球果20-30个, 装入布袋(35 cm × 25 cm)带回实验室。
Table 1 表1 表1 细叶云南松采样种群的地理位置和气象因子概况
Table 1
Locations and meteorological factors of the sampling populations of Pinus yunnanensis var. tenuifolia 种群 经度 纬度 海拔高度 年平均气温 年平均相对 年降水量 地貌 坡度 土壤类型 林分类型 林分平均DBH ± SD (cm) 兴义坝汪 Bawang, Xingyi (BW) 104.83° 24.70° 872.1 17.1 79.6 1 301.7 山地峡谷 Mountain gulch 10-15 砖红壤 Latosol 松栎混交林 26.03 ± 7.94 册亨弼佑 Biyou, Ceheng (BY) 106.00° 24.78° 701.0 18.7 78.8 1 244.7 山地峡谷 Mountain gulch 25-30 砖红壤 Latosol 松栎混交林 23.61 ± 8.44 兴义岔江 Chajiang, Xingyi (CJ) 104.68° 25.07° 1 250.8 16.0 79.8 1 413.0 山地峡谷 Mountain gulch 15-20 砖红壤 Latosol 松栎混交林 24.94 ± 9.79 望谟大观 Daguan, Wangmo 106.27° 25.08° 870.0 18.7 78.1 1 211.6 山地峡谷 Mountain gulch 10-15 砖红壤 Latosol 松栎混交林 32.64 ± 11.28 罗甸大亭 Dating, Luodian (DT) 106.93° 25.30° 1 087.1 17.8 78.6 1 157.0 山地峡谷 Mountain gulch 15-20 砖红壤 Latosol 纯林 24.56 ± 5.73 安龙钱相 Qianxiang, Anlong (QX) 105.48° 25.20° 1 378.9 15.4 79.4 1 278.9 山地峡谷 Mountain gulch 5-10 砖红壤 Latosol 纯林 20.65 ± 8.23 安龙新桥 Xinqiao, Anlong (XQ) 105.28° 25.15° 1 315.0 16.1 79.6 1 266.9 山地峡谷 Mountain gulch 10-15 砖红壤 Latosol 松栎混交林 26.29 ± 13.19 罗甸伍家坟 Wujiafen, Luodian (WJ) 106.93° 25.48° 776.2 17.5 78.7 1 157.3 山地峡谷 Mountain gulch 25-30 砖红壤 Latosol 纯林 35.74 ± 8.38
采样点年平均气温、相对湿度和年降水量通过克里格插值法获得, 原始数据为中国气象数据网(http://data.cma.cn/ )发布的采样点周边区域77个县/市气象站1981-2010年累计的平均值。 Values of mean annual air temperature, relative humidity and mean annual precipitation were estimated (Kriging interpolation method) using the climate data during 1981 to 2010 from 77 climatic stations around the sampling locations. The original data were downloaded from http://data.cma.cn/ . DBH, diameter at breast height. 新窗口打开 |
下载CSV 1.2 性状测定 球果采集后, 于一周内完成球果形态指标测定; 后将球果暴晒干燥后脱粒, 再分别测量种子的相关性状。球果测定: 全测每个单株所采集的球果, 用电子天平称量单个球果鲜质量, 精确到0.01 g; 用游标卡尺测量球果长度及宽度, 精确到0.01 mm。种子及种翅测定: 每个单株随机选择50粒种子进行测定, 利用游标卡尺测定种子长度及宽度, 同时测定对应种子的种翅长度、宽度, 精确到0.01 mm; 种子千粒质量测定按二百粒法(去除种翅后, 按四分法选择200粒种子称质量, 4次重复, 换算为种子千粒质量), 测量精度为0.01 g。
1.3 数据处理 表型变异分析: 利用R 3.6.1 (
R Core Team, 2018 )计算各性状的种群平均值、标准偏差、种群内及种群间变异系数; 基于巢氏方差分析模型分析种群间及种群内差异, 线性模型为:
Yijk =
μ +
τi +
δj ( i ) +
εijk , 其中,
Yijk 为第
i 个种群第
j 个家系第
k 个观测值,
μ 为总平均值,
τi 为种群间效应值,
δj ( i ) 为种群内单株(家系)随机效应值,
εijk 为随机误差(
Logan, 2010 ); 利用表型分化系数(
VST )来反映种群间及种群内的表型
分化状况, 其计算公式为: $V_{S T}=\frac{\sigma_{t / s}^{2}}{\sigma_{t / s}^{2}+\sigma_{s}^{2}}$, 式中,$ \sigma_{t / s}^{2}$为种群间方差分量,$ \sigma_{s}^{2}$为种群内(家系)方差分量(
葛颂等, 1988 )。
表型与环境相关分析: 基于采样区域及其周边气象站观测数据(附录I, II), 利用R相关数据包基于空间插值法(Kriging interpolation)获得各采样点气象数据估计值(
Bivand et al ., 2013 ;
Graler et al ., 2016 ;
Tennekes, 2018 )。利用R数据包Hmisc 4.4计算Pearson相关系数及显著性
p 值(
Harrell & Dupont, 2020 ), 并利用corrplot 0.84绘制相关性图(
Wei & Simko, 2017 )。采样地地理气象因子与参试性状间的典型相关分析采用CCA 1.2数据包(
González & Déjean, 2012 ), 并采用ggplot2绘图(
Wickham, 2016 )。
地理变异规律分析: 采用数据包factoextra进行种实性状主成分分析, 获得综合反映种实性状的代表性主成分, 并以采样区域的气象插值数据作为背景, 分析种群主成分得分及典型相关性状值在采样区域内的变异趋势。采用平均轮廓系数确定最佳类别数
K, 并基于
K -质心法(
K -Medoids)进行种群聚类(
Kassambara, 2017 )。采用Mantel检验分析种群间表型差异随地理距离的变化趋势(
Jombart, 2008 )。
2 结果 2.1 细叶云南松天然种群间和种群内种实性状变异特征 细叶云南松天然种群表型性状在种群间和种群内(植株间)两个层次的方差分析显示(
表2 ), 种群的球果长(
CL )、球果宽(
CW )、球果长宽比(
CR )、球果鲜质量(
FM )、种子长(
SL )、种子宽(
SW )、种子长宽比(
SR )、种子千粒质量(
TSM )、种翅长(
WL )、种翅宽(
WW )、种翅长宽比(
WR )共11个性状在种群间、种群内均存在极显著差异(
p < 0.001)。表明细叶云南松种实形态在种群间及种群内个体间均存在广泛差异。
Table 2 表2 表2 细叶云南松天然种群间和种群内的种实性状巢氏方差分析
Table 2
Nested-ANOVA of cone and seed traits among and within populations of Pinus yunnanensis var. tenuifolia 性状Trait 均方 Mean square F 种群间 Among populations 种群内 Within populations 随机误差 Random error 种群间 Among populations 种群内 Within populations CL 4 460.408 831.114 42.857 104.075*** 19.393*** CW 596.236 173.065 5.813 102.560*** 29.770*** CR 0.995 0.240 0.017 58.159*** 14.041*** FM 11 240.191 2 093.936 81.514 137.893*** 25.688*** SL 27.586 11.767 0.122 226.300*** 96.532*** SW 5.184 2.726 0.065 79.562*** 41.835*** SR 16.907 7.154 0.102 166.553*** 70.471*** TSM 97.442 48.754 0.418 233.050*** 116.603*** WL 647.766 335.707 2.428 266.774*** 138.257*** WW 25.997 19.652 0.375 69.362*** 52.433*** WR 1.045 0.658 0.020 52.216*** 32.874***
***, α = 0.001。CL, 球果长; CR, 果实长宽比; CW, 球果宽; FM, 球果鲜质量; SL, 种子长; SR , 种子长宽比; SW, 种子宽; TSM , 种子千粒质量; WL, 种翅长; WR, 种翅长宽比; WW, 种翅宽。 ***, α = 0.001. CL, cone length; CR, cone aspect ratio; CW, cone width; FM, fresh cone mass; SL, seed length; SR, seed aspect ratio; SW, seed width; TSM, thousand-seeds mass; WL, seed-wing length; WR, seed-wing aspect ratio; WW, seed-wing width. 新窗口打开 |
下载CSV 多重比较(
表3 )表明, 细叶云南松天然种群种实性状在种群间存在着显著的差异。在球果性状方面, 罗甸伍家坟(WJ)种群
CL 、
CW 及
FM 均较大, 其次为罗甸大亭(DT)种群, 而兴义坝汪(BW)种群球果相对较小较轻; 在种子性状(
SL 、
SW 及
TSW )方面也有相似趋势, 仍然以DT、WJ种群种子性状值较大, BW、安龙钱相(QX)种群种子性状值较小。种翅性状上,
WL 以WJ种群最大,
WW 则以望谟大观(DG)种群最大, BW最小。在长宽比上,
CR 、
SR 及
WR 均以WJ种群最大, 以QX最小。综合所有性状, WJ种群各性状值普遍较大, BW种群总体较小。
Table 3 表3 表3 细叶云南松天然种群种实性状平均值、标准偏差及种群间Duncan’s多重比较
Table 3
Mean, standard deviation (SD ) and Duncan’s multiple range test of cone and seed traits of Pinus yunnanensis var. tenuifolia 种群 CL (mm) SD )CW (mm) SD )CR (SD )FM (g) SD )SL (mm) SD )SW (mm) SD )SR (SD )TSM (g) SD )WL (mm) SD )WW (mm) SD )WR (SD )兴义坝汪 53.72g 31.52d 1.70e 26.07f 5.54c 3.18d 1.76b 11.88e 17.70e 5.78e 3.11d 册亨弼佑 63.10de 34.18c 1.85b 40.37d 5.79b 3.41b 1.70c 14.81bcd 19.66c 6.00d 3.32b 兴义岔江 60.81e 34.78c 1.75de 39.62d 5.48c 3.40b 1.63d 14.91bc 19.70c 6.32c 3.16d 望谟大观 63.61cd 36.51b 1.75de 44.69c 5.77b 3.41b 1.70c 14.42cd 19.49c 6.49a 3.03e 罗甸大亭 67.83b 38.18a 1.78cd 50.32b 6.09a 3.49a 1.75b 16.78a 20.38b 6.37abc 3.23c 安龙钱相 57.51f 35.06c 1.63f 33.86e 5.36d 3.19d 1.69c 13.23cde 18.09de 6.45ab 2.83f 罗甸伍家坟 72.34a 37.22ab 1.94a 54.62a 6.04a 3.39b 1.79a 16.48ab 21.31a 6.28c 3.41a 安龙新桥 66.16bc 36.35b 1.82bc 47.80bc 5.54c 3.28c 1.70c 13.07de 18.27d 6.34bc 2.90f
下划线数值为对应指标种群最大平均值。平均值后的字母代表在α = 0.05水平上的种群间多重比较结果。 Values with underline denote the maximum of trait mean of each population. Letters after the means denote the significance of the multiple comparisons among populations. 新窗口打开 |
下载CSV 由性状种群内变异系数(
表4 )可知, 不同性状变异水平有较大差异,
FM 种群平均变异系数最大(36.64%),
SW 最小(9.98%)。不同种群内变异系数也有明显不同, BW种群种子、种翅性状变异较大, DT种群则在多个性状上变异较小。从种群间变异系数来看, 变异系数最大仍然为
FM (21.91%), 但最小为
SR (2.91%), 总体上, 种群间、种群内均以质量性状(单果鲜质量、种子千粒质量)变异系数较大, 长度性状(
CL 、
SL 、
WL )次之, 宽度性状(
SW 、
WW )变异相对最小。
表4 表4 细叶云南松种群种实表型性状的变异系数
Table 4 Coefficient of variation (
CV ) of cone and seed traits from populations of
Pinus yunnanensis var.
tenuifolia 种群 表型变异系数 CV (%) CL CW CR FM SL SW SR TSM WL WW WR Mean 兴义坝汪 17.94 12.91 10.59 45.80 13.54 11.95 15.91 22.98 17.97 17.47 18.65 18.70 册亨弼佑 15.47 11.41 10.81 35.00 9.84 10.56 8.82 19.78 14.90 15.67 15.36 15.24 兴义岔江 19.29 15.30 12.00 45.38 9.12 10.29 11.04 28.44 14.87 15.66 14.56 17.81 望谟大观 14.51 12.46 11.43 33.25 11.27 9.38 10.59 22.05 17.34 14.33 16.17 15.71 罗甸大亭 10.33 10.79 8.43 29.33 9.69 7.45 10.29 16.81 13.35 8.95 17.65 13.01 安龙钱相 21.60 14.60 9.82 39.46 10.82 10.97 7.69 33.71 10.50 11.94 12.37 16.68 罗甸伍家坟 15.32 10.80 8.25 33.98 8.11 8.26 8.94 16.44 13.94 10.99 14.37 13.58 安龙新桥 14.99 10.07 9.89 30.92 9.75 10.98 8.82 28.23 15.27 13.09 11.38 14.85 平均值 Mean 16.18 12.29 10.15 36.64 10.27 9.98 10.27 23.56 14.77 13.51 15.07 15.70 种群间变异系数 CVA 9.31 5.85 5.33 21.91 4.67 3.39 2.91 11.67 6.37 3.87 6.38 7.42
下划线数值为对应性状种群变异系数最大值。种群间变异系数(CVA )为种群间性状标准偏差与总均值的比。 Values with underline denote the maximum of variation coefficient (CV ). CV among populations (CVA ) were calculated as the standard deviation between the trait mean of populations divided by the total mean of all populations. 新窗口打开 |
下载CSV 2.2 细叶云南松种群间表型分化 细叶云南松表型性状方差分量和
VST 显示(
表5 ), 细叶云南松果实及种子性状
VST 的变异幅度为4.14%-39.88%, 表明不同性状在种群间和种群内个体间的方差分量占比有较大差异。其中球果性状(
CL 、
CW 、
CR 及
FM )在种群间的方差占比普遍较高, 种群间分化较大。而种子、种翅性状种群间分化明显低于球果性状, 其中又以翅宽(
WW )在种群间分化最小(
VST = 4.14%)。
Table 5 表5 表5 细叶云南松方差分量及表型分化系数
Table 5
Variance components and differentiation coefficient of phenotypic traits of Pinus yunnanensis var. tenuifolia 性状 方差分量 Variance component 方差分量百分比 Percentage of variance component (%) 表型分化系数 种群间 种群内 随机误差 种群间 种群内 随机误差 CL 45.405 5 68.437 6 42.767 7 28.99 43.70 27.31 39.88 CW 4.573 8 14.312 0 5.805 9 18.52 57.96 23.51 24.22 CR 0.011 0 0.017 6 0.017 0 24.04 38.65 37.31 38.35 FM 85.877 4 161.015 3 81.274 8 26.17 49.06 24.77 34.79 SL 0.037 4 0.233 9 0.121 9 9.52 59.48 31.00 13.80 SW 0.005 6 0.053 4 0.065 2 4.54 43.00 52.46 9.55 SR 0.000 9 0.012 8 0.020 0 2.53 38.06 59.41 6.23 TSM 1.427 1 12.075 3 0.418 1 10.25 86.74 3.00 10.57 WL 0.729 8 6.661 7 2.428 1 7.43 67.84 24.73 9.87 WW 0.016 6 0.383 8 0.374 8 2.14 49.51 48.35 4.14 WR 0.022 6 0.141 1 0.101 5 8.52 53.21 38.27 13.80 平均值Mean 12.97 53.38 33.65 18.65
性状缩写见表2 。 Trait abbreviations see Table 2 . 新窗口打开 |
下载CSV 总体来看, 长度(
CL 、
SL )及质量
(FM 、
TSM )性状在种群间分化比宽度性状(
CW 、
SW 及
WW )要大。以多性状表型分化系数均值(18.65%)为参照, 发现所有种子、种翅相关性状的
VST 均小于平均值, 而球果相关性状
VST 均高于平均值, 这可能意味着球果相关表型性状受环境因子的影响更大, 因种群间地理环境差异较大, 使不同种群间存在较大分化, 而同一林分环境相对一致, 使得其单株间的球果性状变异较小。
2.3 细叶云南松天然种群种实性状和地理生态因子相关性 为进一步分析细叶云南松种实性状和地理气候因子的关系, 对11个种实性状与采样地主要地理气候因子进行了Pearson相关分析(
图1 )。结果表明,
CL 、
CW 、
FM 与纬度(
Lat )、经度(
Lon )呈显著(
p < 0.05)或极显著(
p < 0.01)正相关关系, 与海拔(
Alt )、年降水量(
MAP )、年平均气温(
MAT )及相对湿度(
MRH )相关性不显著。
SL 与
Lon 、
MAT (极)显著正相关(
p < 0.05), 与
MAP 、
MRH 显著负相关(
p < 0.05)。
WL 与
Lon 显著正相关。
WW 与
Lat 显著正相关。
WR 与
Alt 显著负相关。
TSM 与
Lon 显著正相关。需指出的是, 尽管部分测定指标与
MAP 、
MAT 及
MRH 相关性未达显著水平,但总体趋势一致, 即种实性状与经纬度、
MAT 主要呈正相关关系, 而与
MRH 、
MAP 呈负相关关系。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1细叶云南松表型性状与地理气候因子之间的Pearson相关性。 *、**分别表示在95%、99%置信水平下显著相关。
Alt, 海拔;
CL, 果长;
CR, 球果长宽比;
CW, 果宽;
FM, 单果鲜质量;
Lat, 纬度;
Lon, 经度;
MAP, 年降水量;
MAT, 年平均气温;
MRH, 相对湿度;
SL, 种子长;
SR, 种子长宽比;
SW, 种子宽;
TSM, 种子千粒质量;
WL, 种翅长;
WR, 种翅长宽比;
WW, 种翅宽。
Fig. 1Pearson correlation between phenotypic traits and geo-meteorological parameters of Pinus yunnanensis var. tenuifolia. * and ** denote significant correlation between variables at
α = 0.05 and
α = 0.01.
Alt, altitude;
CL, cone length;
CR, cone aspect ratio;
CW, cone width;
FM, fresh cone mass;
Lat, latitude;
Lon, longitude;
MAP, mean annual precipitation;
MAT, mean annual air temperature;
MRH, mean relative humidity;
SL, seed length;
SR, seed aspect ratio;
SW, seed width;
TSM, thousand-seeds mass;
WL, seed-wing length;
WR, seed-wing aspect ratio;
WW, seed-wing width.
种实性状与地理气象因子间的典型相关分析结果(
图2 )显示, 地理气象因子均与典型变量1或2有较高的相关性。其中
Lon 、
Lat 、
MAP 和
MRH 与典型变量1相关性较高,
Alt 、
MAT 与变量2相关性较高。且从向量夹角来看,
Lon 、
Lat 与
MAP 和
MRH 呈负相关关系, 与
MAT 呈正相关关系。从种实性状来看, 除
SR 外, 其他指标均与典型变量1或2有较高相关性, 其中大部分种实性状与变量1高度相关, 而
WR 、
WW 与变量2相关性更大, 绝大部分种实性状分布相对集中, 表明彼此间存在明显的正相关关系。结合种实性状与地理气象因子来看, 种实性状值均随
Lat 、
Lon 增加而增大, 随
MRH 、
MAP 的增大而减小。这与Pearson相关分析结果一致。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2种实性状与地理气候因子的典型相关性。 Alt, 海拔;
CL, 果长;
CR, 球果长宽比;
CW, 果宽;
FM, 单果鲜质量;
Lat, 纬度;
Lon, 经度;
MAP, 年降水量;
MAT, 年平均气温;
MRH, 相对湿度;
SL, 种子长;
SR, 种子长宽比;
SW, 种子宽;
TSM, 种子千粒质量;
WL, 种翅长;
WR, 种翅长宽比;
WW, 种翅宽。
Fig. 2Canonical correlation analysis (CCA) between phenotypic traits and geo-meteorological factors. Alt, altitude;
CL, cone length;
CR, cone aspect ratio;
CW, cone width;
FM, fresh cone mass;
Lat, latitude;
Lon, longitude;
MAP, mean annual precipitation;
MAT, mean annual air temperature;
MRH, mean relative humidity;
SL, seed length;
SR, seed aspect ratio;
SW, seed width;
TSM, thousand-seeds mass;
WL, seed-wing length;
WR, seed-wing aspect ratio;
WW, seed-wing width.
2.4 细叶云南松天然种群主成分及聚类分析 为进一步呈现种群种实性状随经纬度及主要气象因子的变异规律, 将种实性状利用主成分分析进行降维, 发现第一主成分(PC1)能较大程度反映细叶云南松种实性状综合特征(方差贡献率63.98%)(附录III, IV)。将种群
SL 值(与
MAT 、
MAP 及
MRH 均显著相关)、种群PC1得分(附录V)分别与通过空间插值获得的采样区域的气象因子梯度进行对应分析(
图3 ), 发现
SL 有随
MAT 增加而增大趋势, 但两者不完全吻合, 西部4个种群(CJ、XQ、QX及BW)位于
MAT 较低区域,
SL 值亦相对较小, 中部两个种群(BY、DG)
MAT 最高, 而
SL 处于中等, 位于东部的WJ、DT具有中度
MAT, 但
SL 最大。基于种群PC1值与
MAT 的对应分析亦有相似结果。从
MAP 来看,
SL 及PC1值的增大与
MAP 下降的趋势较为一致, 西部种群
SL 和PC1均较小,
MAP 较大, 中部种群
SL 、PC1及
MAP 均为中等, 东部种群
SL 、PC1较大,
MAP 较小。在
MRH 梯度图中,
SL 和PC1随
MRH 的增大而减小。从经纬度来看, 除QX外, 种实性状有自西向东、自南向北逐渐增大的趋势。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3细叶云南松8个天然种群种实性状随地理气象因子的变化模式。 MAP, 年降水量;
MAT, 年平均气温;
MRH, 相对湿度;
SL, 种子长; PC1为各种群种实性状主成分分析中第一主成分得分值(第一主成分贡献率63.98%)。种群缩写(BW、BY、CJ、DG、DT、QX、WJ、XQ)同
表1。Fig. 3Distribution model of the seed and cone traits of Pinus yunnanensis var. tenuifolia with the geo-meteorological factors. MAP, mean annual precipitation; MAT, mean annual air temperature; MRH, mean relative humidity; SL, seed length; PC1 (63.98%), the score of the first principal component of each population based on principal component analysis of seed and cone traits. See Table1 for population abbreviation (BW, BY, CJ, DG, DT, QX, WJ, XQ).K )为3 (图4A )。以聚类数K = 3为基准, 基于前两个主成分(累计方差贡献率83.87%)对种群进行聚类(图4B ), 结果显示位于东部的DT、WJ种群聚为一类, 其种实性状特点为球果及种子大而重, 种翅较长。位于西南部的BW种群单独聚为一类, 其种实性状特点为球果、种子及种翅小、质量轻。其他种群(QX、XQ、CJ、BY、DG)聚为一类, 其种实性状值介于前两类之间。图4 新窗口打开 |下载原图ZIP |生成PPT 图4细叶云南松天然种群种实性状的主成分聚类。 K -Medoids法)。种群缩写(BW、BY、CJ、DG、DT、QX、WJ、XQ)同表1。Fig. 4Principal component and cluster analysis based on the phenotypic traits of eight natural populations in Pinus yunnanensis var. tenuifolia. K -Medoids algorithm. See Table1 for population abbreviation (BW, BY, CJ, DG, DT, QX, WJ, XQ).图5A )显示, 表型距离与地理距离间相关值(0.562 8)显著偏离零假设(无空间结构)下的拟合值(p = 0.009), 表明种群种实性状符合距离隔离模式(IBD), 种群间存在明显的空间结构。性状的表型距离随地理距离的增大而增大, 二维核密度估计图显示表型距离为单核斑块, 证明种群空间结构近似符合渐变群(cline)模式(图5B )。图5 新窗口打开 |下载原图ZIP |生成PPT 图5基于表型距离与地理距离(欧式距离)的Mantel检验。 p = 0.009)。B, 表型距离与地理距离的相关性图, 彩色斑块为基于距离值的二维核密度估计结果, 单核表明符合渐变群模式。Fig. 5Mantel test between morpho-distance and geo-distance (Euclidean distance). 3 讨论 3.1 细叶云南松表型变异及分化 表型多样性是遗传多样性和环境多样性的综合表现。对种实性状进行种群差异及多样性研究, 可在一定程度上反映该物种的遗传变异大小和分布规律, 同时还能反映种实形态变异对长期环境的适应关系(Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 )。本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%)。这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化。总体上, 细叶云南松种群间总体分化水平属于中低水平。这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 )。同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现。3.2 细叶云南松种实形态地理变异规律及其生态适应性 不少研究表明, 海拔、年平均气温和经纬度是影响种实表型性状变异的主要因素(Shen et al ., 2015 ; 刘雄盛等, 2017 ; 何庆海等, 2018 ; Wu et al ., 2018 ; 刘莹等, 2019 )。细叶云南松种实性状与地理气象因子相关分析结果可以看出, 绝大部分性状与地理气象因子的相关性表现出一致的趋势, 主要表现为与经纬度、年平均气温呈正相关关系, 与相对湿度、年降水量和海拔呈负相关关系, 这意味着随降水量和相对湿度下降, 年平均气温升高, 其球果、种子、种翅及千粒质量有增大趋势。这与同属的白皮松(李斌等, 2002 ), 油松(姬明飞等, 2013 )等有所不同。这说明不同的生境下的树种种实性状受到环境的选择压力不同, 其表型性状变异方向亦不同(Businsky et al ., 2014 )。水分和温度是影响种子萌发、苗木生长的重要环境因素(McLaren & McDonald 2003 ; Bazin et al ., 2011 ), 因而其对种子及苗木生态适应性不可避免具有重要选择作用。一些研究(Andersson, 1996 ; Simons & Johnston, 2000 )表明, 果实及种子大小、千粒质量与生活力和幼苗生长适应性有显著的正相关关系, 换言之, 较大果实及种子有利于其繁殖。因此, 综合以上可以推测, 在南盘江、红水河沿线特殊的干热河谷气候长期作用下(李治基和王献溥, 1981 ), 细叶云南松逐步演化出了适应干旱、高温生境的种实性状, 进而为其在特殊生境下的繁衍、更新奠定了基础。基于相关分析显著的性状(SL )和种群代表性主成分得分(PC1)与地理气象因子的对应分析发现, SL 、PC1随经纬度呈现渐变趋势, 地理距离较近的种群种实形态更为相似, 这一方面可能源于地理距离较近种群间基因流动更为频繁, 遗传相似度更高, 另一方面其环境因素更为相似, 对种实性状选择作用趋于一致(图3 )。此外, SL 、PC1随MAP 、MRH 的变化比随MAT 的变化具有更高的一致性, 表明该区域的干旱生境相对于温度对细叶云南松种实性状可能具有更强的选择作用。图3 )来看, 分组呈西南-东北走向, 基本符合距离渐变模式, 但也有例外, 如QX种群种实较小, 与组内其他种群差异较大, 推测与该种群位于人为活动较频繁区域, 受到较强的人为干扰和破坏(如拔大毛式择伐)有关, 其样株胸径普遍小于其他种群也间接印证了该推测(表1 )。总体上, 种群间地理距离越大, 分化越大。Mantel检验也证实了这一结论(图5 )。综上可以推测, 地理距离造成的基因流大小及自然环境差异(湿度、温度等)的协同作用是引起细叶云南松表型分化的主要原因。尽管近年来受到其他经济林木种植的冲击, 使得细叶云南松生境受到较大程度的破坏, 呈现碎片化, 但因时间相对较短, 未对其现有种群分化水平造成明显影响, 使其遗传分化仍维持在中低水平。但随着人为干扰的进一步加剧, 种群规模缩小, 种群间的隔离增大, 种群内近交或自交加剧, 其种群分化水平、遗传漂变很可能进一步增大。附录I 细叶云南松天然种群采样点及用于空间插值分析的气象站点分布图 Supplement I Map of sampling sites and the weather stations for spatial interpolation of eight natural populations in Pinus yunnanensis var. tenuifolia https://www.plant-ecology.com/fileup/1005-264X/PDF/cjpe.2020.0269-S1.pdf 附录II 用于空间插值分析的气象站点基本信息 Supplement II Basic information of the climatic stations for spatial interpolation https://www.plant-ecology.com/fileup/1005-264X/PDF/cjpe.2020.0269-S2.pdf 附录III 细叶云南松种实性状在各主成分(PC1-PC7)中的贡献值 Supplement III Contribution of the seed and cone traits of Pinus yunnanensis var. tenuifolia to principle components (PC1-PC7) https://www.plant-ecology.com/fileup/1005-264X/PDF/cjpe.2020.0269-S3.pdf 附录IV 细叶云南松种实性状主成分特征值及方差贡献率 Supplement IV Eigenvalues and the percentages of variance of principle components of the seed and cone traits of Pinus yunnanensis var. tenuifolia https://www.plant-ecology.com/fileup/1005-264X/PDF/cjpe.2020.0269-S4.pdf 附录V 细叶云南松种实性状种群主成分得分值 Supplement V Scores of principle components of the populations of the seed and cone traits of Pinus yunnanensis var. tenuifolia https://www.plant-ecology.com/fileup/1005-264X/PDF/cjpe.2020.0269-S5.pdf 致谢 感谢贵州省望谟县林业局、兴义市林业局、安龙县林业局、罗甸县林业局和册亨县林业局在样本采集工作中给予的帮助。[1] Andersson S 1996 ). Seed size as a determinant of germination rate in Crepis tectorum (Asteraceae): evidence from a seed burial experimentBotany 568 -572 . [本文引用: 1] [2] Bazin J Batlla D Dussert S El-Maarouf-Bouteau H Bailly C 2011 ). Role of relative humidity, temperature, and water status in dormancy alleviation of sunflower seeds during dry after-ripeningJournal of Experimental Botany 627 -640 . URL PMID:20978143 [本文引用: 1] [3] Bivand RS Pebesma E Gomez-Rubio V 2013 ). Applied Spatial Data Analysis with R 2nd ed. Springer, New York . [本文引用: 1] [4] Businsky R Frantík T Vít P 2014 ). Morphological evaluation of the Pinus kesiyacomplex (Pinaceae)Plant Systematics & Evolution 273 -285 . [本文引用: 1] [5] Cilas C Machado R Motamayor JC 2009 ). Relations between several traits linked to sexual plant reproduction in Theobroma cacao L.: number of ovules per ovary, number of seeds per pod, and seed weightTree Genetics Genomes 219 -226 . [本文引用: 1] [6] Diao SF Shao WH Jiang JM Dong RX Sun HG 2014 ). Phenotypic diversity in natural populations of Sapindus mukorossi based on fruit and seed traitsActa Ecologica Sinica 1451 -1460 . [本文引用: 1] [ 刁松锋 , 邵文豪 , 姜景民 , 董汝湘 , 孙洪刚 (2014 ). 基于种实性状的无患子天然群体表型多样性研究生态学报 , 34, 1451 -1460 .] [本文引用: 1] [7] Donohue K Pyle EH Messiqua D Heschel MS Schmitt J 2001 ). Adaptive divergence in plasticity in natural populations of Impatiens capensis and its consequences for performance in novel habitatsEvolution 692 -702 . URL PMID:11392387 [本文引用: 1] [8] Etterson JR 2004 ). Evolutionary potential of Chamaecrista fasciculata in relation to climate change. I. Clinal parterns of selection along an environmental gradient in the great plantsEvolution 1446 -1456 . 10.1111/j.0014-3820.2004.tb01726.x URL PMID:15341148 [本文引用: 1] [9] Feng QH Shi ZM Xu ZJR Miao N Tang JC Liu XL Zhang L 2017 ). Phenotypic variations in cones and seeds of natural Cupressus chengiana populations in ChinaChinese Journal of Applied Ecology 748 -756 . 10.13287/j.1001-9332.201703.001 URL PMID:29740999 [本文引用: 1] [ 冯秋红 , 史作民 , 徐峥静茹 , 缪宁 , 唐敬超 , 刘兴良 , 张雷 (2017 ). 岷江柏天然种群种实表型变异特征应用生态学报 , 28, 748 -756 .] 29740999 [本文引用: 1] [10] Ge S Wang MX Chen YW 1988 ). An analysis of population genetic structure of masson pine by isozyme techniqueScientia Silvae Sinicae 399 -409 . [本文引用: 1] [ 葛颂 , 王明庥 , 陈岳武 (1988 ). 用同工酶研究马尾松群体的遗传结构林业科学 , 24, 399 -409 .] [本文引用: 1] [11] Gitonga L Kahangi E Muigai A Ngamau K Gichuki S Cheluget W Wepukhulu S 2008 ). Assessment of phenotypic diversity of macadamia (Macadamia spp.) germplasm in Kenya using leaf and fruit morphologyAfrican Journal of Plant Science 86 -93 . [本文引用: 2] [12] González I Déjean S 2012 ). CCA: canonical correlation analysis. R package version 1.2 . [2020 -08-06]. https:// CRAN.R-project.org/package=CCA. URL [本文引用: 1] [13] Graler B Pebesma E Heuvelink GB 2016 ). Spatio-temporal interpolation using gstatR Journal 204 -218 . [本文引用: 1] [14] Gu YJ Luo JX Wu YW Cao XJ 2009 ). Phenotypic diversity in natural population of Picea balfouriana in Sichuan, ChinaChinese Journal of Plant Ecology 291 -301 . [本文引用: 1] [ 辜云杰 , 罗建勋 , 吴远伟 , 曹小军 (2009 ). 川西云杉天然种群表型多样性植物生态学报 , 33, 291 -301 .] [本文引用: 1] [15] Harrell Jr FE Dupont C 2020 ). Harrell Miscellaneous. R package version 4.4-02020 -08-06]. https://CRAN.R-project. org/package=Hmisc. URL [本文引用: 1] [16] He QH Yang SZ Li YG Shen X Liu XH 2018 ). Phenotypic variations in seed and fruit traits of Liquidambar formosana populationsChinese Journal of Plant Ecology 752 -763 . [本文引用: 1] [ 何庆海 , 杨少宗 , 李因刚 , 沈鑫 , 柳新红 (2018 ). 枫香树种群种子与果实表型性状变异分析植物生态学报 , 42, 752 -763 .] [本文引用: 1] [17] Ji MF Zhang XW Han J Ding DL 2013 ). Phenotypic diversity of cone and seed in natural Pinus tabuliformis populations in ChinaActa Bot Boreali-Occidential Sinica 1898 -1905 . [本文引用: 2] [ 姬明飞 , 张晓玮 , 韩瑾 , 丁东粮 (2013 ). 油松天然群体的种实性状表型多样性研究西北植物学报 , 33, 1898 -1905 .] [本文引用: 2] [18] Jombart T 2008 ). adegenet: a R package for the multivariate analysis of genetic markersBioinformatics 1403 -1405 . URL PMID:18397895 [本文引用: 1] [19] Kassambara A 2017 ). Practical guide to cluster analysis in R: unsupervised machine learning . [2020 -08-06]. http://www. sthda.com. URL [本文引用: 1] [20] Li B Gu WC Lu BM 2002 ). A study on phenotypic diversity of seeds and cones characteristics in Pinus bungeana Biodiversity Science 181 -188 . [本文引用: 2] [ 李斌 , 顾万春 , 卢宝明 (2002 ). 白皮松天然群体种实性状表型多样性研究生物多样性 , 10, 51 -58 .] [本文引用: 2] [21] Li SF Su JR Liu WD Lang XD Zhang ZJ Su L Jia CXZ Yang HJ 2013 ). Phenotypic variations in cones and seeds of natural Pinus kesiya var. langbianensis populations in Yunnan Province, ChinaChinese Journal of Plant Ecology 998 -1009 . [本文引用: 1] [ 李帅锋 , 苏建荣 , 刘万德 , 郎学东 , 张志钧 , 苏磊 , 贾呈鑫卓 , 杨华景 (2013 ). 思茅松天然群体种实表型变异植物生态学报 , 37, 998 -1009 .] [本文引用: 1] [22] Li YF He JA Yu SF Liao LN Wang HX Ye SM 2019 ). Spatial patterns of trees in a south subtropical Pinus yunnanensis var. tenuifolia forest after selective logging of large-sized treesChinese Journal of Ecology 3585 -3592 . [本文引用: 1] [ 李远发 , 何吉安 , 喻素芳 , 廖良宁 , 王宏翔 , 叶绍明 (2019 ). 南亚热带细叶云南松林大径木择伐后的空间格局生态学杂志 , 38, 3585 -3592 .] [本文引用: 1] [23] Li ZJ Wang XP 1981 ). The distribution of Pinus yunnanensis var. tenuifolia in relation to the environmental conditionsActa Phytoecologica et Geobotanica Sinica 28 -37 . [本文引用: 3] [ 李治基 , 王献溥 (1981 ). 广西细叶云南松地理分布和环境的关系植物生态学报 , 5, 28 -37 .] [本文引用: 3] [24] Liao LN Lu ZJ Li YF Ye SM Yu SF 2019 ). Litter and soil nutrient characteristics of Pinus yunnanensis var. tenuifolia natural forest in northwest GuangxiJournal of Northwest Forestry University 31 -38 . [本文引用: 1] [ 廖良宁 , 卢姿瑾 , 李远发 , 叶绍明 , 喻素芳 (2019 ). 桂西北细叶云南松天然林凋落物及土壤养分特征西北林学院学报 , 34, 31 -38 .] [本文引用: 1] [25] Liu XS Jiang Y Huang RL Liu F Xiao YF Jiang Y Wei SX 2017 ). Variation in traits of cone and seed of Keteleeria fortunei var. cyclolepis and its relationship with environmental factorsGuihaia 118 -126 . [本文引用: 1] [ 刘雄盛 , 蒋燚 , 黄荣林 , 刘菲 , 肖玉菲 , 姜英 , 韦铄星 (2017 ). 江南油杉种实性状变异及其与环境因子的关系广西植物 , 37, 118 -126 .] [本文引用: 1] [26] Liu Y Ma WJ Yang GJ Wang JC Yi F Wang YT Wang JH Wang CY 2019 ). Geographical variation of the seed and germination characteristics of natural populations of Sophora moorcroftiana Journal of Northwest Forestry University 22 -29 . [本文引用: 1] [ 刘莹 , 麻文俊 , 杨桂娟 , 王俊臣 , 易飞 , 王玉婷 , 王军辉 , 王春燕 (2019 ). 砂生槐天然种群种实形态和萌发特征地理变异西北林学院学报 , 34, 22 -29 .] [本文引用: 1] [27] Logan M 2010 ). Biostatistical Design and Analysis Using R: A Practical Guide Wiley-Blackwell, West Sussex, UK . [本文引用: 1] [28] Luo JX Gu WC 2005 ). Study on phenotypic diversity of natural population in Picea asperata Scientia Silvae Sinicae 2 ), 66 -73 . [本文引用: 1] [ 罗建勋 , 顾万春 (2005 ). 云杉天然群体表型多样性研究林业科学 , 41(2 ), 66 -73 .] [本文引用: 1] [29] McLaren KP McDonald MA 2003 ). The effects of moisture and shade on seed germination and seedling survival in a tropical dry forest in JamaicaForest Ecology and Management 61 -75 . [本文引用: 1] [30] Mosseler LMOCA Rajora OP 2007 ). Extensive long-distance pollen dispersal in a fragmented landscape maintains genetic diversity in white spruceJournal of Heredity 640 -645 . [本文引用: 1] [31] Pan T 2018 ). Study on Structure Characteristics of the Main Tree Species of Pinus yunnanensis var. tenuifolia Secondary Forest in the Nanpan River Basin Master degree dissertation, Guangxi University, Nanning . [本文引用: 1] [ 潘婷 (2018 ). 南盘江流域细叶云南松次生林主要树种结构特征研究硕士学位论文, 广西大学, 南宁 .] [本文引用: 1] [32] R Core Team 2018 ). R: a Language and Environment for Statistical Computing R Foundation for Statistical Computing, Vienna, Austria . [本文引用: 1] [33] Shen L Xu R Sai Liu S Jun C Xu C Xie C Liu T 2015 ). Phenotypic variation of seed traits of Haloxylon ammodendron and its affecting factorsBiochemical Systematics and Ecology 81 -87 . [本文引用: 1] [34] Shirley L Vesk P 2009 ). Eucalyptus phylogenetics: How traits, evolution, and the environment influence the distribution of a diverse genus94th ESA Annual Convention, Austin, USA . [本文引用: 1] [35] Simons AM Johnston MO 2000 ). Variation in seed traits of Lobelia inflata (Campanulaceae): sources and fitness consequencesAmerican Journal of Botany 124 -132 . URL PMID:10636835 [本文引用: 1] [36] Sun C Wang J Duan J Zhao G Weng X Jia L 2017 ). Association of fruit and seed traits of Sapindus mukorossi germplasm with environmental factors in southern ChinaForests 491 . DOI: 10.3390/f8120491 . [本文引用: 1] [37] Tennekes M 2018 ). tmap: thematic maps in RJournal of Statistical Software 1 -39 . 10.18637/jss.v084.i01 URL PMID:30450020 [本文引用: 1] [38] Wall SBV 2008 ). On the relative contributions of wind vs. animals to seed dispersal of four Sierra Nevada pinesEcology 1837 -1849 . URL PMID:18705371 [本文引用: 1] [39] Wang XP 1991 ). The phytocoenological features of Pinus yunnanensis var. tenuifolia forest in GuangxiBulletin of Botanical Research 91 -103 . [本文引用: 1] [ 王献溥 (1991 ). 广西细叶云南松林的群落学特点植物研究 , 11, 91 -103 .] [本文引用: 1] [40] Wei T Simko V 2017 ). R package “corrplot”: visualization of a Correlation Matrix (Version 0.84) . [2020 -08-06]. https:// github.com/taiyun/corrplot. URL [本文引用: 1] [41] Wickham H 2016 ). ggplot2: Elegant Graphics for Data Analysis Springer-Verlag, New York . [本文引用: 1] [42] Wu DS Tang X Cen ZM Xu CL Yang ZQ 2016 ). Dynamic analysis of Pinus yunnanensis var. tenuifolia forest resources in GuangxiGuangxi Forestry Science 316 -321 . [本文引用: 1] [ 吴东山 , 唐鑫 , 岑祖明 , 许春林 , 杨章旗 (2016 ). 广西细叶云南松森林资源动态分析广西林业科学 , 45, 316 -321 .] [本文引用: 1] [43] Wu H Meng H Wang S Wei X Jiang M 2018 ). Geographic patterns and environmental drivers of seed traits of a relict tree speciesForest Ecology and Management 59 -68 . [本文引用: 1] [44] Xu XL 1983 ). Geographical distribution and growth characteristics of Pinus yunnanensis var. tenuifolia forest in Guizhou ProvinceGuizhou Science 94 -98 . [本文引用: 2] [ 徐学良 (1983 ). 细叶云南松林在贵州的地理分布和生长特性贵州科学 , 94 -98 .] [本文引用: 2] [45] Xu Y Deng LL Zhou L Cai NH Luo YR Li DL Duan AA Xu YL 2015 ). Seeds and cones phenotypic variation among populations of Pinus yunnanensis at different altitudesSeed 70 -74 . [本文引用: 1] [ 徐杨 , 邓丽丽 , 周丽 , 蔡年辉 , 罗一然 , 李德龙 , 段安安 , 许玉兰 (2015 ). 云南松不同海拔天然群体种实性状表型多样性研究种子 , 34, 70 -74 .] [本文引用: 1] [46] Yang ZQ Feng YH Wu DS 2014 ). Analysis of genetic diversity of Pinus yunnanensis var. tenuifolia nature populations by SSR markerGuihaia 10 -14 . [本文引用: 1] [ 杨章旗 , 冯源恒 , 吴东山 (2014 ). 细叶云南松天然种源林遗传多样性的SSR分析广西植物 , 34, 10 -14 .] [本文引用: 1] [47] Yu S She G Ye S Zhou X Li Y 2018 ). Characteristics of soil microbial biomass and community composition in Pinus yunnanensis var. tenuifolia secondary forests view supplementary materialJournal of Sustainable Forestry 753 -770 . [本文引用: 1] [48] Zhang MY Su CX Lu CH 2020 ). Cones structure and seed traits of four species of large-seeded pines: adaptation to animal-mediated dispersalEcology and Evolution 5293 -5301 . URL PMID:32607152 [本文引用: 1] Crepis tectorum (Asteraceae): evidence from a seed burial experimentShen et al ., 2015 ; 刘雄盛等, 2017 ; 何庆海等, 2018 ; Wu et al ., 2018 ; 刘莹等, 2019 ).细叶云南松种实性状与地理气象因子相关分析结果可以看出, 绝大部分性状与地理气象因子的相关性表现出一致的趋势, 主要表现为与经纬度、年平均气温呈正相关关系, 与相对湿度、年降水量和海拔呈负相关关系, 这意味着随降水量和相对湿度下降, 年平均气温升高, 其球果、种子、种翅及千粒质量有增大趋势.这与同属的白皮松(李斌等, 2002 ), 油松(姬明飞等, 2013 )等有所不同.这说明不同的生境下的树种种实性状受到环境的选择压力不同, 其表型性状变异方向亦不同(Businsky et al ., 2014 ).水分和温度是影响种子萌发、苗木生长的重要环境因素(McLaren & McDonald 2003 ; Bazin et al ., 2011 ), 因而其对种子及苗木生态适应性不可避免具有重要选择作用.一些研究(Andersson, 1996 ; Simons & Johnston, 2000 )表明, 果实及种子大小、千粒质量与生活力和幼苗生长适应性有显著的正相关关系, 换言之, 较大果实及种子有利于其繁殖.因此, 综合以上可以推测, 在南盘江、红水河沿线特殊的干热河谷气候长期作用下(李治基和王献溥, 1981 ), 细叶云南松逐步演化出了适应干旱、高温生境的种实性状, 进而为其在特殊生境下的繁衍、更新奠定了基础.基于相关分析显著的性状(SL )和种群代表性主成分得分(PC1)与地理气象因子的对应分析发现, SL 、PC1随经纬度呈现渐变趋势, 地理距离较近的种群种实形态更为相似, 这一方面可能源于地理距离较近种群间基因流动更为频繁, 遗传相似度更高, 另一方面其环境因素更为相似, 对种实性状选择作用趋于一致(图3 ).此外, SL 、PC1随MAP 、MRH 的变化比随MAT 的变化具有更高的一致性, 表明该区域的干旱生境相对于温度对细叶云南松种实性状可能具有更强的选择作用. ...Shen et al ., 2015 ; 刘雄盛等, 2017 ; 何庆海等, 2018 ; Wu et al ., 2018 ; 刘莹等, 2019 ).细叶云南松种实性状与地理气象因子相关分析结果可以看出, 绝大部分性状与地理气象因子的相关性表现出一致的趋势, 主要表现为与经纬度、年平均气温呈正相关关系, 与相对湿度、年降水量和海拔呈负相关关系, 这意味着随降水量和相对湿度下降, 年平均气温升高, 其球果、种子、种翅及千粒质量有增大趋势.这与同属的白皮松(李斌等, 2002 ), 油松(姬明飞等, 2013 )等有所不同.这说明不同的生境下的树种种实性状受到环境的选择压力不同, 其表型性状变异方向亦不同(Businsky et al ., 2014 ).水分和温度是影响种子萌发、苗木生长的重要环境因素(McLaren & McDonald 2003 ; Bazin et al ., 2011 ), 因而其对种子及苗木生态适应性不可避免具有重要选择作用.一些研究(Andersson, 1996 ; Simons & Johnston, 2000 )表明, 果实及种子大小、千粒质量与生活力和幼苗生长适应性有显著的正相关关系, 换言之, 较大果实及种子有利于其繁殖.因此, 综合以上可以推测, 在南盘江、红水河沿线特殊的干热河谷气候长期作用下(李治基和王献溥, 1981 ), 细叶云南松逐步演化出了适应干旱、高温生境的种实性状, 进而为其在特殊生境下的繁衍、更新奠定了基础.基于相关分析显著的性状(SL )和种群代表性主成分得分(PC1)与地理气象因子的对应分析发现, SL 、PC1随经纬度呈现渐变趋势, 地理距离较近的种群种实形态更为相似, 这一方面可能源于地理距离较近种群间基因流动更为频繁, 遗传相似度更高, 另一方面其环境因素更为相似, 对种实性状选择作用趋于一致(图3 ).此外, SL 、PC1随MAP 、MRH 的变化比随MAT 的变化具有更高的一致性, 表明该区域的干旱生境相对于温度对细叶云南松种实性状可能具有更强的选择作用. ...Applied Spatial Data Analysis with R Bivand et al ., 2013 ; Graler et al ., 2016 ; Tennekes, 2018 ).利用R数据包Hmisc 4.4计算Pearson相关系数及显著性p 值(Harrell & Dupont, 2020 ), 并利用corrplot 0.84绘制相关性图(Wei & Simko, 2017 ).采样地地理气象因子与参试性状间的典型相关分析采用CCA 1.2数据包(González & Déjean, 2012 ), 并采用ggplot2绘图(Wickham, 2016 ). ...Pinus kesiyacomplex (Pinaceae)Shen et al ., 2015 ; 刘雄盛等, 2017 ; 何庆海等, 2018 ; Wu et al ., 2018 ; 刘莹等, 2019 ).细叶云南松种实性状与地理气象因子相关分析结果可以看出, 绝大部分性状与地理气象因子的相关性表现出一致的趋势, 主要表现为与经纬度、年平均气温呈正相关关系, 与相对湿度、年降水量和海拔呈负相关关系, 这意味着随降水量和相对湿度下降, 年平均气温升高, 其球果、种子、种翅及千粒质量有增大趋势.这与同属的白皮松(李斌等, 2002 ), 油松(姬明飞等, 2013 )等有所不同.这说明不同的生境下的树种种实性状受到环境的选择压力不同, 其表型性状变异方向亦不同(Businsky et al ., 2014 ).水分和温度是影响种子萌发、苗木生长的重要环境因素(McLaren & McDonald 2003 ; Bazin et al ., 2011 ), 因而其对种子及苗木生态适应性不可避免具有重要选择作用.一些研究(Andersson, 1996 ; Simons & Johnston, 2000 )表明, 果实及种子大小、千粒质量与生活力和幼苗生长适应性有显著的正相关关系, 换言之, 较大果实及种子有利于其繁殖.因此, 综合以上可以推测, 在南盘江、红水河沿线特殊的干热河谷气候长期作用下(李治基和王献溥, 1981 ), 细叶云南松逐步演化出了适应干旱、高温生境的种实性状, 进而为其在特殊生境下的繁衍、更新奠定了基础.基于相关分析显著的性状(SL )和种群代表性主成分得分(PC1)与地理气象因子的对应分析发现, SL 、PC1随经纬度呈现渐变趋势, 地理距离较近的种群种实形态更为相似, 这一方面可能源于地理距离较近种群间基因流动更为频繁, 遗传相似度更高, 另一方面其环境因素更为相似, 对种实性状选择作用趋于一致(图3 ).此外, SL 、PC1随MAP 、MRH 的变化比随MAT 的变化具有更高的一致性, 表明该区域的干旱生境相对于温度对细叶云南松种实性状可能具有更强的选择作用. ...Theobroma cacao L.: number of ovules per ovary, number of seeds per pod, and seed weight潘婷, 2018 ; Yu et al ., 2018 ; 李远发等, 2019 ; 廖良宁等, 2019 ).针对细叶云南松遗传资源评价的研究相对较少, 仅见杨章旗等(2014) 对广西西北部3个种群进行了SSR遗传多样性分析, 认为遗传多样性与欧洲栓皮栎(Quercus suber )、黄杉(Pseudotsuga menziesii )等相比普遍不高, 但种群间基因流动频繁, 分化较小.表型多样性是遗传与环境共同作用的结果, 研究不同地理环境条件下的天然种群的表型性状分化程度和变异规律, 不仅可在一定程度上反映其遗传变异水平, 也可以有效揭示生物对不同环境的适应, 进而探究其发展演化(Donohue et al ., 2001 ; Etterson, 2004 ; Gitonga et al ., 2008 ; Shirley & Vesk, 2009 ).种实性状在物种自然演化过程中具有较高的遗传稳定性, 不易受短期环境变化的影响.同时, 作为主要繁殖器官, 种实性状对物种的定居、繁衍及分布具有重要影响(Cilas et al ., 2009 ).研究表明, 果实形态、种子大小、质量、种翅的有无等都会影响其种子传播媒介(风、动物等)的传播效率, 进而影响物种的分布范围(Wall, 2008 ; Zhang et al ., 2020 ).本文通过对细叶云南松自然分布区的8个天然种群开展种实性状分析, 以揭示细叶云南松种实性状在空间分布上的表型分化和变异规律, 并探究其与细叶云南松特殊生境的关系, 为进一步开展细叶云南松的种质资源保护与利用提供参考. ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...Impatiens capensis and its consequences for performance in novel habitats潘婷, 2018 ; Yu et al ., 2018 ; 李远发等, 2019 ; 廖良宁等, 2019 ).针对细叶云南松遗传资源评价的研究相对较少, 仅见杨章旗等(2014) 对广西西北部3个种群进行了SSR遗传多样性分析, 认为遗传多样性与欧洲栓皮栎(Quercus suber )、黄杉(Pseudotsuga menziesii )等相比普遍不高, 但种群间基因流动频繁, 分化较小.表型多样性是遗传与环境共同作用的结果, 研究不同地理环境条件下的天然种群的表型性状分化程度和变异规律, 不仅可在一定程度上反映其遗传变异水平, 也可以有效揭示生物对不同环境的适应, 进而探究其发展演化(Donohue et al ., 2001 ; Etterson, 2004 ; Gitonga et al ., 2008 ; Shirley & Vesk, 2009 ).种实性状在物种自然演化过程中具有较高的遗传稳定性, 不易受短期环境变化的影响.同时, 作为主要繁殖器官, 种实性状对物种的定居、繁衍及分布具有重要影响(Cilas et al ., 2009 ).研究表明, 果实形态、种子大小、质量、种翅的有无等都会影响其种子传播媒介(风、动物等)的传播效率, 进而影响物种的分布范围(Wall, 2008 ; Zhang et al ., 2020 ).本文通过对细叶云南松自然分布区的8个天然种群开展种实性状分析, 以揭示细叶云南松种实性状在空间分布上的表型分化和变异规律, 并探究其与细叶云南松特殊生境的关系, 为进一步开展细叶云南松的种质资源保护与利用提供参考. ...Chamaecrista fasciculata in relation to climate change. I. Clinal parterns of selection along an environmental gradient in the great plants潘婷, 2018 ; Yu et al ., 2018 ; 李远发等, 2019 ; 廖良宁等, 2019 ).针对细叶云南松遗传资源评价的研究相对较少, 仅见杨章旗等(2014) 对广西西北部3个种群进行了SSR遗传多样性分析, 认为遗传多样性与欧洲栓皮栎(Quercus suber )、黄杉(Pseudotsuga menziesii )等相比普遍不高, 但种群间基因流动频繁, 分化较小.表型多样性是遗传与环境共同作用的结果, 研究不同地理环境条件下的天然种群的表型性状分化程度和变异规律, 不仅可在一定程度上反映其遗传变异水平, 也可以有效揭示生物对不同环境的适应, 进而探究其发展演化(Donohue et al ., 2001 ; Etterson, 2004 ; Gitonga et al ., 2008 ; Shirley & Vesk, 2009 ).种实性状在物种自然演化过程中具有较高的遗传稳定性, 不易受短期环境变化的影响.同时, 作为主要繁殖器官, 种实性状对物种的定居、繁衍及分布具有重要影响(Cilas et al ., 2009 ).研究表明, 果实形态、种子大小、质量、种翅的有无等都会影响其种子传播媒介(风、动物等)的传播效率, 进而影响物种的分布范围(Wall, 2008 ; Zhang et al ., 2020 ).本文通过对细叶云南松自然分布区的8个天然种群开展种实性状分析, 以揭示细叶云南松种实性状在空间分布上的表型分化和变异规律, 并探究其与细叶云南松特殊生境的关系, 为进一步开展细叶云南松的种质资源保护与利用提供参考. ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...葛颂等, 1988 ). ...葛颂等, 1988 ). ...Macadamia spp.) germplasm in Kenya using leaf and fruit morphology潘婷, 2018 ; Yu et al ., 2018 ; 李远发等, 2019 ; 廖良宁等, 2019 ).针对细叶云南松遗传资源评价的研究相对较少, 仅见杨章旗等(2014) 对广西西北部3个种群进行了SSR遗传多样性分析, 认为遗传多样性与欧洲栓皮栎(Quercus suber )、黄杉(Pseudotsuga menziesii )等相比普遍不高, 但种群间基因流动频繁, 分化较小.表型多样性是遗传与环境共同作用的结果, 研究不同地理环境条件下的天然种群的表型性状分化程度和变异规律, 不仅可在一定程度上反映其遗传变异水平, 也可以有效揭示生物对不同环境的适应, 进而探究其发展演化(Donohue et al ., 2001 ; Etterson, 2004 ; Gitonga et al ., 2008 ; Shirley & Vesk, 2009 ).种实性状在物种自然演化过程中具有较高的遗传稳定性, 不易受短期环境变化的影响.同时, 作为主要繁殖器官, 种实性状对物种的定居、繁衍及分布具有重要影响(Cilas et al ., 2009 ).研究表明, 果实形态、种子大小、质量、种翅的有无等都会影响其种子传播媒介(风、动物等)的传播效率, 进而影响物种的分布范围(Wall, 2008 ; Zhang et al ., 2020 ).本文通过对细叶云南松自然分布区的8个天然种群开展种实性状分析, 以揭示细叶云南松种实性状在空间分布上的表型分化和变异规律, 并探究其与细叶云南松特殊生境的关系, 为进一步开展细叶云南松的种质资源保护与利用提供参考. ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...Bivand et al ., 2013 ; Graler et al ., 2016 ; Tennekes, 2018 ).利用R数据包Hmisc 4.4计算Pearson相关系数及显著性p 值(Harrell & Dupont, 2020 ), 并利用corrplot 0.84绘制相关性图(Wei & Simko, 2017 ).采样地地理气象因子与参试性状间的典型相关分析采用CCA 1.2数据包(González & Déjean, 2012 ), 并采用ggplot2绘图(Wickham, 2016 ). ...Bivand et al ., 2013 ; Graler et al ., 2016 ; Tennekes, 2018 ).利用R数据包Hmisc 4.4计算Pearson相关系数及显著性p 值(Harrell & Dupont, 2020 ), 并利用corrplot 0.84绘制相关性图(Wei & Simko, 2017 ).采样地地理气象因子与参试性状间的典型相关分析采用CCA 1.2数据包(González & Déjean, 2012 ), 并采用ggplot2绘图(Wickham, 2016 ). ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...Bivand et al ., 2013 ; Graler et al ., 2016 ; Tennekes, 2018 ).利用R数据包Hmisc 4.4计算Pearson相关系数及显著性p 值(Harrell & Dupont, 2020 ), 并利用corrplot 0.84绘制相关性图(Wei & Simko, 2017 ).采样地地理气象因子与参试性状间的典型相关分析采用CCA 1.2数据包(González & Déjean, 2012 ), 并采用ggplot2绘图(Wickham, 2016 ). ...Shen et al ., 2015 ; 刘雄盛等, 2017 ; 何庆海等, 2018 ; Wu et al ., 2018 ; 刘莹等, 2019 ).细叶云南松种实性状与地理气象因子相关分析结果可以看出, 绝大部分性状与地理气象因子的相关性表现出一致的趋势, 主要表现为与经纬度、年平均气温呈正相关关系, 与相对湿度、年降水量和海拔呈负相关关系, 这意味着随降水量和相对湿度下降, 年平均气温升高, 其球果、种子、种翅及千粒质量有增大趋势.这与同属的白皮松(李斌等, 2002 ), 油松(姬明飞等, 2013 )等有所不同.这说明不同的生境下的树种种实性状受到环境的选择压力不同, 其表型性状变异方向亦不同(Businsky et al ., 2014 ).水分和温度是影响种子萌发、苗木生长的重要环境因素(McLaren & McDonald 2003 ; Bazin et al ., 2011 ), 因而其对种子及苗木生态适应性不可避免具有重要选择作用.一些研究(Andersson, 1996 ; Simons & Johnston, 2000 )表明, 果实及种子大小、千粒质量与生活力和幼苗生长适应性有显著的正相关关系, 换言之, 较大果实及种子有利于其繁殖.因此, 综合以上可以推测, 在南盘江、红水河沿线特殊的干热河谷气候长期作用下(李治基和王献溥, 1981 ), 细叶云南松逐步演化出了适应干旱、高温生境的种实性状, 进而为其在特殊生境下的繁衍、更新奠定了基础.基于相关分析显著的性状(SL )和种群代表性主成分得分(PC1)与地理气象因子的对应分析发现, SL 、PC1随经纬度呈现渐变趋势, 地理距离较近的种群种实形态更为相似, 这一方面可能源于地理距离较近种群间基因流动更为频繁, 遗传相似度更高, 另一方面其环境因素更为相似, 对种实性状选择作用趋于一致(图3 ).此外, SL 、PC1随MAP 、MRH 的变化比随MAT 的变化具有更高的一致性, 表明该区域的干旱生境相对于温度对细叶云南松种实性状可能具有更强的选择作用. ...Shen et al ., 2015 ; 刘雄盛等, 2017 ; 何庆海等, 2018 ; Wu et al ., 2018 ; 刘莹等, 2019 ).细叶云南松种实性状与地理气象因子相关分析结果可以看出, 绝大部分性状与地理气象因子的相关性表现出一致的趋势, 主要表现为与经纬度、年平均气温呈正相关关系, 与相对湿度、年降水量和海拔呈负相关关系, 这意味着随降水量和相对湿度下降, 年平均气温升高, 其球果、种子、种翅及千粒质量有增大趋势.这与同属的白皮松(李斌等, 2002 ), 油松(姬明飞等, 2013 )等有所不同.这说明不同的生境下的树种种实性状受到环境的选择压力不同, 其表型性状变异方向亦不同(Businsky et al ., 2014 ).水分和温度是影响种子萌发、苗木生长的重要环境因素(McLaren & McDonald 2003 ; Bazin et al ., 2011 ), 因而其对种子及苗木生态适应性不可避免具有重要选择作用.一些研究(Andersson, 1996 ; Simons & Johnston, 2000 )表明, 果实及种子大小、千粒质量与生活力和幼苗生长适应性有显著的正相关关系, 换言之, 较大果实及种子有利于其繁殖.因此, 综合以上可以推测, 在南盘江、红水河沿线特殊的干热河谷气候长期作用下(李治基和王献溥, 1981 ), 细叶云南松逐步演化出了适应干旱、高温生境的种实性状, 进而为其在特殊生境下的繁衍、更新奠定了基础.基于相关分析显著的性状(SL )和种群代表性主成分得分(PC1)与地理气象因子的对应分析发现, SL 、PC1随经纬度呈现渐变趋势, 地理距离较近的种群种实形态更为相似, 这一方面可能源于地理距离较近种群间基因流动更为频繁, 遗传相似度更高, 另一方面其环境因素更为相似, 对种实性状选择作用趋于一致(图3 ).此外, SL 、PC1随MAP 、MRH 的变化比随MAT 的变化具有更高的一致性, 表明该区域的干旱生境相对于温度对细叶云南松种实性状可能具有更强的选择作用. ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...Shen et al ., 2015 ; 刘雄盛等, 2017 ; 何庆海等, 2018 ; Wu et al ., 2018 ; 刘莹等, 2019 ).细叶云南松种实性状与地理气象因子相关分析结果可以看出, 绝大部分性状与地理气象因子的相关性表现出一致的趋势, 主要表现为与经纬度、年平均气温呈正相关关系, 与相对湿度、年降水量和海拔呈负相关关系, 这意味着随降水量和相对湿度下降, 年平均气温升高, 其球果、种子、种翅及千粒质量有增大趋势.这与同属的白皮松(李斌等, 2002 ), 油松(姬明飞等, 2013 )等有所不同.这说明不同的生境下的树种种实性状受到环境的选择压力不同, 其表型性状变异方向亦不同(Businsky et al ., 2014 ).水分和温度是影响种子萌发、苗木生长的重要环境因素(McLaren & McDonald 2003 ; Bazin et al ., 2011 ), 因而其对种子及苗木生态适应性不可避免具有重要选择作用.一些研究(Andersson, 1996 ; Simons & Johnston, 2000 )表明, 果实及种子大小、千粒质量与生活力和幼苗生长适应性有显著的正相关关系, 换言之, 较大果实及种子有利于其繁殖.因此, 综合以上可以推测, 在南盘江、红水河沿线特殊的干热河谷气候长期作用下(李治基和王献溥, 1981 ), 细叶云南松逐步演化出了适应干旱、高温生境的种实性状, 进而为其在特殊生境下的繁衍、更新奠定了基础.基于相关分析显著的性状(SL )和种群代表性主成分得分(PC1)与地理气象因子的对应分析发现, SL 、PC1随经纬度呈现渐变趋势, 地理距离较近的种群种实形态更为相似, 这一方面可能源于地理距离较近种群间基因流动更为频繁, 遗传相似度更高, 另一方面其环境因素更为相似, 对种实性状选择作用趋于一致(图3 ).此外, SL 、PC1随MAP 、MRH 的变化比随MAT 的变化具有更高的一致性, 表明该区域的干旱生境相对于温度对细叶云南松种实性状可能具有更强的选择作用. ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...Shen et al ., 2015 ; 刘雄盛等, 2017 ; 何庆海等, 2018 ; Wu et al ., 2018 ; 刘莹等, 2019 ).细叶云南松种实性状与地理气象因子相关分析结果可以看出, 绝大部分性状与地理气象因子的相关性表现出一致的趋势, 主要表现为与经纬度、年平均气温呈正相关关系, 与相对湿度、年降水量和海拔呈负相关关系, 这意味着随降水量和相对湿度下降, 年平均气温升高, 其球果、种子、种翅及千粒质量有增大趋势.这与同属的白皮松(李斌等, 2002 ), 油松(姬明飞等, 2013 )等有所不同.这说明不同的生境下的树种种实性状受到环境的选择压力不同, 其表型性状变异方向亦不同(Businsky et al ., 2014 ).水分和温度是影响种子萌发、苗木生长的重要环境因素(McLaren & McDonald 2003 ; Bazin et al ., 2011 ), 因而其对种子及苗木生态适应性不可避免具有重要选择作用.一些研究(Andersson, 1996 ; Simons & Johnston, 2000 )表明, 果实及种子大小、千粒质量与生活力和幼苗生长适应性有显著的正相关关系, 换言之, 较大果实及种子有利于其繁殖.因此, 综合以上可以推测, 在南盘江、红水河沿线特殊的干热河谷气候长期作用下(李治基和王献溥, 1981 ), 细叶云南松逐步演化出了适应干旱、高温生境的种实性状, 进而为其在特殊生境下的繁衍、更新奠定了基础.基于相关分析显著的性状(SL )和种群代表性主成分得分(PC1)与地理气象因子的对应分析发现, SL 、PC1随经纬度呈现渐变趋势, 地理距离较近的种群种实形态更为相似, 这一方面可能源于地理距离较近种群间基因流动更为频繁, 遗传相似度更高, 另一方面其环境因素更为相似, 对种实性状选择作用趋于一致(图3 ).此外, SL 、PC1随MAP 、MRH 的变化比随MAT 的变化具有更高的一致性, 表明该区域的干旱生境相对于温度对细叶云南松种实性状可能具有更强的选择作用. ...K, 并基于K -质心法(K -Medoids)进行种群聚类(Kassambara, 2017 ).采用Mantel检验分析种群间表型差异随地理距离的变化趋势(Jombart, 2008 ). ...K, 并基于K -质心法(K -Medoids)进行种群聚类(Kassambara, 2017 ).采用Mantel检验分析种群间表型差异随地理距离的变化趋势(Jombart, 2008 ). ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...Shen et al ., 2015 ; 刘雄盛等, 2017 ; 何庆海等, 2018 ; Wu et al ., 2018 ; 刘莹等, 2019 ).细叶云南松种实性状与地理气象因子相关分析结果可以看出, 绝大部分性状与地理气象因子的相关性表现出一致的趋势, 主要表现为与经纬度、年平均气温呈正相关关系, 与相对湿度、年降水量和海拔呈负相关关系, 这意味着随降水量和相对湿度下降, 年平均气温升高, 其球果、种子、种翅及千粒质量有增大趋势.这与同属的白皮松(李斌等, 2002 ), 油松(姬明飞等, 2013 )等有所不同.这说明不同的生境下的树种种实性状受到环境的选择压力不同, 其表型性状变异方向亦不同(Businsky et al ., 2014 ).水分和温度是影响种子萌发、苗木生长的重要环境因素(McLaren & McDonald 2003 ; Bazin et al ., 2011 ), 因而其对种子及苗木生态适应性不可避免具有重要选择作用.一些研究(Andersson, 1996 ; Simons & Johnston, 2000 )表明, 果实及种子大小、千粒质量与生活力和幼苗生长适应性有显著的正相关关系, 换言之, 较大果实及种子有利于其繁殖.因此, 综合以上可以推测, 在南盘江、红水河沿线特殊的干热河谷气候长期作用下(李治基和王献溥, 1981 ), 细叶云南松逐步演化出了适应干旱、高温生境的种实性状, 进而为其在特殊生境下的繁衍、更新奠定了基础.基于相关分析显著的性状(SL )和种群代表性主成分得分(PC1)与地理气象因子的对应分析发现, SL 、PC1随经纬度呈现渐变趋势, 地理距离较近的种群种实形态更为相似, 这一方面可能源于地理距离较近种群间基因流动更为频繁, 遗传相似度更高, 另一方面其环境因素更为相似, 对种实性状选择作用趋于一致(图3 ).此外, SL 、PC1随MAP 、MRH 的变化比随MAT 的变化具有更高的一致性, 表明该区域的干旱生境相对于温度对细叶云南松种实性状可能具有更强的选择作用. ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...Shen et al ., 2015 ; 刘雄盛等, 2017 ; 何庆海等, 2018 ; Wu et al ., 2018 ; 刘莹等, 2019 ).细叶云南松种实性状与地理气象因子相关分析结果可以看出, 绝大部分性状与地理气象因子的相关性表现出一致的趋势, 主要表现为与经纬度、年平均气温呈正相关关系, 与相对湿度、年降水量和海拔呈负相关关系, 这意味着随降水量和相对湿度下降, 年平均气温升高, 其球果、种子、种翅及千粒质量有增大趋势.这与同属的白皮松(李斌等, 2002 ), 油松(姬明飞等, 2013 )等有所不同.这说明不同的生境下的树种种实性状受到环境的选择压力不同, 其表型性状变异方向亦不同(Businsky et al ., 2014 ).水分和温度是影响种子萌发、苗木生长的重要环境因素(McLaren & McDonald 2003 ; Bazin et al ., 2011 ), 因而其对种子及苗木生态适应性不可避免具有重要选择作用.一些研究(Andersson, 1996 ; Simons & Johnston, 2000 )表明, 果实及种子大小、千粒质量与生活力和幼苗生长适应性有显著的正相关关系, 换言之, 较大果实及种子有利于其繁殖.因此, 综合以上可以推测, 在南盘江、红水河沿线特殊的干热河谷气候长期作用下(李治基和王献溥, 1981 ), 细叶云南松逐步演化出了适应干旱、高温生境的种实性状, 进而为其在特殊生境下的繁衍、更新奠定了基础.基于相关分析显著的性状(SL )和种群代表性主成分得分(PC1)与地理气象因子的对应分析发现, SL 、PC1随经纬度呈现渐变趋势, 地理距离较近的种群种实形态更为相似, 这一方面可能源于地理距离较近种群间基因流动更为频繁, 遗传相似度更高, 另一方面其环境因素更为相似, 对种实性状选择作用趋于一致(图3 ).此外, SL 、PC1随MAP 、MRH 的变化比随MAT 的变化具有更高的一致性, 表明该区域的干旱生境相对于温度对细叶云南松种实性状可能具有更强的选择作用. ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...潘婷, 2018 ; Yu et al ., 2018 ; 李远发等, 2019 ; 廖良宁等, 2019 ).针对细叶云南松遗传资源评价的研究相对较少, 仅见杨章旗等(2014) 对广西西北部3个种群进行了SSR遗传多样性分析, 认为遗传多样性与欧洲栓皮栎(Quercus suber )、黄杉(Pseudotsuga menziesii )等相比普遍不高, 但种群间基因流动频繁, 分化较小.表型多样性是遗传与环境共同作用的结果, 研究不同地理环境条件下的天然种群的表型性状分化程度和变异规律, 不仅可在一定程度上反映其遗传变异水平, 也可以有效揭示生物对不同环境的适应, 进而探究其发展演化(Donohue et al ., 2001 ; Etterson, 2004 ; Gitonga et al ., 2008 ; Shirley & Vesk, 2009 ).种实性状在物种自然演化过程中具有较高的遗传稳定性, 不易受短期环境变化的影响.同时, 作为主要繁殖器官, 种实性状对物种的定居、繁衍及分布具有重要影响(Cilas et al ., 2009 ).研究表明, 果实形态、种子大小、质量、种翅的有无等都会影响其种子传播媒介(风、动物等)的传播效率, 进而影响物种的分布范围(Wall, 2008 ; Zhang et al ., 2020 ).本文通过对细叶云南松自然分布区的8个天然种群开展种实性状分析, 以揭示细叶云南松种实性状在空间分布上的表型分化和变异规律, 并探究其与细叶云南松特殊生境的关系, 为进一步开展细叶云南松的种质资源保护与利用提供参考. ...潘婷, 2018 ; Yu et al ., 2018 ; 李远发等, 2019 ; 廖良宁等, 2019 ).针对细叶云南松遗传资源评价的研究相对较少, 仅见杨章旗等(2014) 对广西西北部3个种群进行了SSR遗传多样性分析, 认为遗传多样性与欧洲栓皮栎(Quercus suber )、黄杉(Pseudotsuga menziesii )等相比普遍不高, 但种群间基因流动频繁, 分化较小.表型多样性是遗传与环境共同作用的结果, 研究不同地理环境条件下的天然种群的表型性状分化程度和变异规律, 不仅可在一定程度上反映其遗传变异水平, 也可以有效揭示生物对不同环境的适应, 进而探究其发展演化(Donohue et al ., 2001 ; Etterson, 2004 ; Gitonga et al ., 2008 ; Shirley & Vesk, 2009 ).种实性状在物种自然演化过程中具有较高的遗传稳定性, 不易受短期环境变化的影响.同时, 作为主要繁殖器官, 种实性状对物种的定居、繁衍及分布具有重要影响(Cilas et al ., 2009 ).研究表明, 果实形态、种子大小、质量、种翅的有无等都会影响其种子传播媒介(风、动物等)的传播效率, 进而影响物种的分布范围(Wall, 2008 ; Zhang et al ., 2020 ).本文通过对细叶云南松自然分布区的8个天然种群开展种实性状分析, 以揭示细叶云南松种实性状在空间分布上的表型分化和变异规律, 并探究其与细叶云南松特殊生境的关系, 为进一步开展细叶云南松的种质资源保护与利用提供参考. ...Pinus yunnanensis var. tenuifolia )是云南松(P. yunnanensis )因南亚热带独特的河谷气候长期影响及适应下逐渐形成的一个地理变种, 是我国特有种, 其天然分布区域狭窄, 仅限于黔、滇、桂交界处的南盘江流域、北盘江流域以及桂西北的红水河流域, 属于微域分布的植被类型(徐学良, 1983 ; 王献溥, 1991 ).细叶云南松生境狭窄, 属于年平均气温较高, 降水量小于蒸发量, 总体较为干旱的河谷区域, 焚风效应明显, 气候独特.独特的生境使细叶云南松具有较高的生态研究价值(李治基和王献溥, 1981 ).此外, 细叶云南松干型直、树体高大, 出材率高, 木材强度和质量较好, 具有较高的开发利用前景.但近年来, 由于该区域其他经济林树种(油茶(Camellia oleifera )、八角(Illicium verum )等)的大力发展, 对细叶云南松生境的人为干扰急剧加重, 大量细叶云南松林遭到蚕食、破坏, 原有的诸多大面积连续分布林分已消失殆尽, 呈零星点状分布(吴东山等, 2016 ).针对细叶云南松的遗传资源调查、收集、保存、评价迫在眉睫. ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...Shen et al ., 2015 ; 刘雄盛等, 2017 ; 何庆海等, 2018 ; Wu et al ., 2018 ; 刘莹等, 2019 ).细叶云南松种实性状与地理气象因子相关分析结果可以看出, 绝大部分性状与地理气象因子的相关性表现出一致的趋势, 主要表现为与经纬度、年平均气温呈正相关关系, 与相对湿度、年降水量和海拔呈负相关关系, 这意味着随降水量和相对湿度下降, 年平均气温升高, 其球果、种子、种翅及千粒质量有增大趋势.这与同属的白皮松(李斌等, 2002 ), 油松(姬明飞等, 2013 )等有所不同.这说明不同的生境下的树种种实性状受到环境的选择压力不同, 其表型性状变异方向亦不同(Businsky et al ., 2014 ).水分和温度是影响种子萌发、苗木生长的重要环境因素(McLaren & McDonald 2003 ; Bazin et al ., 2011 ), 因而其对种子及苗木生态适应性不可避免具有重要选择作用.一些研究(Andersson, 1996 ; Simons & Johnston, 2000 )表明, 果实及种子大小、千粒质量与生活力和幼苗生长适应性有显著的正相关关系, 换言之, 较大果实及种子有利于其繁殖.因此, 综合以上可以推测, 在南盘江、红水河沿线特殊的干热河谷气候长期作用下(李治基和王献溥, 1981 ), 细叶云南松逐步演化出了适应干旱、高温生境的种实性状, 进而为其在特殊生境下的繁衍、更新奠定了基础.基于相关分析显著的性状(SL )和种群代表性主成分得分(PC1)与地理气象因子的对应分析发现, SL 、PC1随经纬度呈现渐变趋势, 地理距离较近的种群种实形态更为相似, 这一方面可能源于地理距离较近种群间基因流动更为频繁, 遗传相似度更高, 另一方面其环境因素更为相似, 对种实性状选择作用趋于一致(图3 ).此外, SL 、PC1随MAP 、MRH 的变化比随MAT 的变化具有更高的一致性, 表明该区域的干旱生境相对于温度对细叶云南松种实性状可能具有更强的选择作用. ...Pinus yunnanensis var. tenuifolia )是云南松(P. yunnanensis )因南亚热带独特的河谷气候长期影响及适应下逐渐形成的一个地理变种, 是我国特有种, 其天然分布区域狭窄, 仅限于黔、滇、桂交界处的南盘江流域、北盘江流域以及桂西北的红水河流域, 属于微域分布的植被类型(徐学良, 1983 ; 王献溥, 1991 ).细叶云南松生境狭窄, 属于年平均气温较高, 降水量小于蒸发量, 总体较为干旱的河谷区域, 焚风效应明显, 气候独特.独特的生境使细叶云南松具有较高的生态研究价值(李治基和王献溥, 1981 ).此外, 细叶云南松干型直、树体高大, 出材率高, 木材强度和质量较好, 具有较高的开发利用前景.但近年来, 由于该区域其他经济林树种(油茶(Camellia oleifera )、八角(Illicium verum )等)的大力发展, 对细叶云南松生境的人为干扰急剧加重, 大量细叶云南松林遭到蚕食、破坏, 原有的诸多大面积连续分布林分已消失殆尽, 呈零星点状分布(吴东山等, 2016 ).针对细叶云南松的遗传资源调查、收集、保存、评价迫在眉睫. ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...Shen et al ., 2015 ; 刘雄盛等, 2017 ; 何庆海等, 2018 ; Wu et al ., 2018 ; 刘莹等, 2019 ).细叶云南松种实性状与地理气象因子相关分析结果可以看出, 绝大部分性状与地理气象因子的相关性表现出一致的趋势, 主要表现为与经纬度、年平均气温呈正相关关系, 与相对湿度、年降水量和海拔呈负相关关系, 这意味着随降水量和相对湿度下降, 年平均气温升高, 其球果、种子、种翅及千粒质量有增大趋势.这与同属的白皮松(李斌等, 2002 ), 油松(姬明飞等, 2013 )等有所不同.这说明不同的生境下的树种种实性状受到环境的选择压力不同, 其表型性状变异方向亦不同(Businsky et al ., 2014 ).水分和温度是影响种子萌发、苗木生长的重要环境因素(McLaren & McDonald 2003 ; Bazin et al ., 2011 ), 因而其对种子及苗木生态适应性不可避免具有重要选择作用.一些研究(Andersson, 1996 ; Simons & Johnston, 2000 )表明, 果实及种子大小、千粒质量与生活力和幼苗生长适应性有显著的正相关关系, 换言之, 较大果实及种子有利于其繁殖.因此, 综合以上可以推测, 在南盘江、红水河沿线特殊的干热河谷气候长期作用下(李治基和王献溥, 1981 ), 细叶云南松逐步演化出了适应干旱、高温生境的种实性状, 进而为其在特殊生境下的繁衍、更新奠定了基础.基于相关分析显著的性状(SL )和种群代表性主成分得分(PC1)与地理气象因子的对应分析发现, SL 、PC1随经纬度呈现渐变趋势, 地理距离较近的种群种实形态更为相似, 这一方面可能源于地理距离较近种群间基因流动更为频繁, 遗传相似度更高, 另一方面其环境因素更为相似, 对种实性状选择作用趋于一致(图3 ).此外, SL 、PC1随MAP 、MRH 的变化比随MAT 的变化具有更高的一致性, 表明该区域的干旱生境相对于温度对细叶云南松种实性状可能具有更强的选择作用. ...潘婷, 2018 ; Yu et al ., 2018 ; 李远发等, 2019 ; 廖良宁等, 2019 ).针对细叶云南松遗传资源评价的研究相对较少, 仅见杨章旗等(2014) 对广西西北部3个种群进行了SSR遗传多样性分析, 认为遗传多样性与欧洲栓皮栎(Quercus suber )、黄杉(Pseudotsuga menziesii )等相比普遍不高, 但种群间基因流动频繁, 分化较小.表型多样性是遗传与环境共同作用的结果, 研究不同地理环境条件下的天然种群的表型性状分化程度和变异规律, 不仅可在一定程度上反映其遗传变异水平, 也可以有效揭示生物对不同环境的适应, 进而探究其发展演化(Donohue et al ., 2001 ; Etterson, 2004 ; Gitonga et al ., 2008 ; Shirley & Vesk, 2009 ).种实性状在物种自然演化过程中具有较高的遗传稳定性, 不易受短期环境变化的影响.同时, 作为主要繁殖器官, 种实性状对物种的定居、繁衍及分布具有重要影响(Cilas et al ., 2009 ).研究表明, 果实形态、种子大小、质量、种翅的有无等都会影响其种子传播媒介(风、动物等)的传播效率, 进而影响物种的分布范围(Wall, 2008 ; Zhang et al ., 2020 ).本文通过对细叶云南松自然分布区的8个天然种群开展种实性状分析, 以揭示细叶云南松种实性状在空间分布上的表型分化和变异规律, 并探究其与细叶云南松特殊生境的关系, 为进一步开展细叶云南松的种质资源保护与利用提供参考. ...潘婷, 2018 ; Yu et al ., 2018 ; 李远发等, 2019 ; 廖良宁等, 2019 ).针对细叶云南松遗传资源评价的研究相对较少, 仅见杨章旗等(2014) 对广西西北部3个种群进行了SSR遗传多样性分析, 认为遗传多样性与欧洲栓皮栎(Quercus suber )、黄杉(Pseudotsuga menziesii )等相比普遍不高, 但种群间基因流动频繁, 分化较小.表型多样性是遗传与环境共同作用的结果, 研究不同地理环境条件下的天然种群的表型性状分化程度和变异规律, 不仅可在一定程度上反映其遗传变异水平, 也可以有效揭示生物对不同环境的适应, 进而探究其发展演化(Donohue et al ., 2001 ; Etterson, 2004 ; Gitonga et al ., 2008 ; Shirley & Vesk, 2009 ).种实性状在物种自然演化过程中具有较高的遗传稳定性, 不易受短期环境变化的影响.同时, 作为主要繁殖器官, 种实性状对物种的定居、繁衍及分布具有重要影响(Cilas et al ., 2009 ).研究表明, 果实形态、种子大小、质量、种翅的有无等都会影响其种子传播媒介(风、动物等)的传播效率, 进而影响物种的分布范围(Wall, 2008 ; Zhang et al ., 2020 ).本文通过对细叶云南松自然分布区的8个天然种群开展种实性状分析, 以揭示细叶云南松种实性状在空间分布上的表型分化和变异规律, 并探究其与细叶云南松特殊生境的关系, 为进一步开展细叶云南松的种质资源保护与利用提供参考. ...Shen et al ., 2015 ; 刘雄盛等, 2017 ; 何庆海等, 2018 ; Wu et al ., 2018 ; 刘莹等, 2019 ).细叶云南松种实性状与地理气象因子相关分析结果可以看出, 绝大部分性状与地理气象因子的相关性表现出一致的趋势, 主要表现为与经纬度、年平均气温呈正相关关系, 与相对湿度、年降水量和海拔呈负相关关系, 这意味着随降水量和相对湿度下降, 年平均气温升高, 其球果、种子、种翅及千粒质量有增大趋势.这与同属的白皮松(李斌等, 2002 ), 油松(姬明飞等, 2013 )等有所不同.这说明不同的生境下的树种种实性状受到环境的选择压力不同, 其表型性状变异方向亦不同(Businsky et al ., 2014 ).水分和温度是影响种子萌发、苗木生长的重要环境因素(McLaren & McDonald 2003 ; Bazin et al ., 2011 ), 因而其对种子及苗木生态适应性不可避免具有重要选择作用.一些研究(Andersson, 1996 ; Simons & Johnston, 2000 )表明, 果实及种子大小、千粒质量与生活力和幼苗生长适应性有显著的正相关关系, 换言之, 较大果实及种子有利于其繁殖.因此, 综合以上可以推测, 在南盘江、红水河沿线特殊的干热河谷气候长期作用下(李治基和王献溥, 1981 ), 细叶云南松逐步演化出了适应干旱、高温生境的种实性状, 进而为其在特殊生境下的繁衍、更新奠定了基础.基于相关分析显著的性状(SL )和种群代表性主成分得分(PC1)与地理气象因子的对应分析发现, SL 、PC1随经纬度呈现渐变趋势, 地理距离较近的种群种实形态更为相似, 这一方面可能源于地理距离较近种群间基因流动更为频繁, 遗传相似度更高, 另一方面其环境因素更为相似, 对种实性状选择作用趋于一致(图3 ).此外, SL 、PC1随MAP 、MRH 的变化比随MAT 的变化具有更高的一致性, 表明该区域的干旱生境相对于温度对细叶云南松种实性状可能具有更强的选择作用. ...Shen et al ., 2015 ; 刘雄盛等, 2017 ; 何庆海等, 2018 ; Wu et al ., 2018 ; 刘莹等, 2019 ).细叶云南松种实性状与地理气象因子相关分析结果可以看出, 绝大部分性状与地理气象因子的相关性表现出一致的趋势, 主要表现为与经纬度、年平均气温呈正相关关系, 与相对湿度、年降水量和海拔呈负相关关系, 这意味着随降水量和相对湿度下降, 年平均气温升高, 其球果、种子、种翅及千粒质量有增大趋势.这与同属的白皮松(李斌等, 2002 ), 油松(姬明飞等, 2013 )等有所不同.这说明不同的生境下的树种种实性状受到环境的选择压力不同, 其表型性状变异方向亦不同(Businsky et al ., 2014 ).水分和温度是影响种子萌发、苗木生长的重要环境因素(McLaren & McDonald 2003 ; Bazin et al ., 2011 ), 因而其对种子及苗木生态适应性不可避免具有重要选择作用.一些研究(Andersson, 1996 ; Simons & Johnston, 2000 )表明, 果实及种子大小、千粒质量与生活力和幼苗生长适应性有显著的正相关关系, 换言之, 较大果实及种子有利于其繁殖.因此, 综合以上可以推测, 在南盘江、红水河沿线特殊的干热河谷气候长期作用下(李治基和王献溥, 1981 ), 细叶云南松逐步演化出了适应干旱、高温生境的种实性状, 进而为其在特殊生境下的繁衍、更新奠定了基础.基于相关分析显著的性状(SL )和种群代表性主成分得分(PC1)与地理气象因子的对应分析发现, SL 、PC1随经纬度呈现渐变趋势, 地理距离较近的种群种实形态更为相似, 这一方面可能源于地理距离较近种群间基因流动更为频繁, 遗传相似度更高, 另一方面其环境因素更为相似, 对种实性状选择作用趋于一致(图3 ).此外, SL 、PC1随MAP 、MRH 的变化比随MAT 的变化具有更高的一致性, 表明该区域的干旱生境相对于温度对细叶云南松种实性状可能具有更强的选择作用. ...Shen et al ., 2015 ; 刘雄盛等, 2017 ; 何庆海等, 2018 ; Wu et al ., 2018 ; 刘莹等, 2019 ).细叶云南松种实性状与地理气象因子相关分析结果可以看出, 绝大部分性状与地理气象因子的相关性表现出一致的趋势, 主要表现为与经纬度、年平均气温呈正相关关系, 与相对湿度、年降水量和海拔呈负相关关系, 这意味着随降水量和相对湿度下降, 年平均气温升高, 其球果、种子、种翅及千粒质量有增大趋势.这与同属的白皮松(李斌等, 2002 ), 油松(姬明飞等, 2013 )等有所不同.这说明不同的生境下的树种种实性状受到环境的选择压力不同, 其表型性状变异方向亦不同(Businsky et al ., 2014 ).水分和温度是影响种子萌发、苗木生长的重要环境因素(McLaren & McDonald 2003 ; Bazin et al ., 2011 ), 因而其对种子及苗木生态适应性不可避免具有重要选择作用.一些研究(Andersson, 1996 ; Simons & Johnston, 2000 )表明, 果实及种子大小、千粒质量与生活力和幼苗生长适应性有显著的正相关关系, 换言之, 较大果实及种子有利于其繁殖.因此, 综合以上可以推测, 在南盘江、红水河沿线特殊的干热河谷气候长期作用下(李治基和王献溥, 1981 ), 细叶云南松逐步演化出了适应干旱、高温生境的种实性状, 进而为其在特殊生境下的繁衍、更新奠定了基础.基于相关分析显著的性状(SL )和种群代表性主成分得分(PC1)与地理气象因子的对应分析发现, SL 、PC1随经纬度呈现渐变趋势, 地理距离较近的种群种实形态更为相似, 这一方面可能源于地理距离较近种群间基因流动更为频繁, 遗传相似度更高, 另一方面其环境因素更为相似, 对种实性状选择作用趋于一致(图3 ).此外, SL 、PC1随MAP 、MRH 的变化比随MAT 的变化具有更高的一致性, 表明该区域的干旱生境相对于温度对细叶云南松种实性状可能具有更强的选择作用. ...Shen et al ., 2015 ; 刘雄盛等, 2017 ; 何庆海等, 2018 ; Wu et al ., 2018 ; 刘莹等, 2019 ).细叶云南松种实性状与地理气象因子相关分析结果可以看出, 绝大部分性状与地理气象因子的相关性表现出一致的趋势, 主要表现为与经纬度、年平均气温呈正相关关系, 与相对湿度、年降水量和海拔呈负相关关系, 这意味着随降水量和相对湿度下降, 年平均气温升高, 其球果、种子、种翅及千粒质量有增大趋势.这与同属的白皮松(李斌等, 2002 ), 油松(姬明飞等, 2013 )等有所不同.这说明不同的生境下的树种种实性状受到环境的选择压力不同, 其表型性状变异方向亦不同(Businsky et al ., 2014 ).水分和温度是影响种子萌发、苗木生长的重要环境因素(McLaren & McDonald 2003 ; Bazin et al ., 2011 ), 因而其对种子及苗木生态适应性不可避免具有重要选择作用.一些研究(Andersson, 1996 ; Simons & Johnston, 2000 )表明, 果实及种子大小、千粒质量与生活力和幼苗生长适应性有显著的正相关关系, 换言之, 较大果实及种子有利于其繁殖.因此, 综合以上可以推测, 在南盘江、红水河沿线特殊的干热河谷气候长期作用下(李治基和王献溥, 1981 ), 细叶云南松逐步演化出了适应干旱、高温生境的种实性状, 进而为其在特殊生境下的繁衍、更新奠定了基础.基于相关分析显著的性状(SL )和种群代表性主成分得分(PC1)与地理气象因子的对应分析发现, SL 、PC1随经纬度呈现渐变趋势, 地理距离较近的种群种实形态更为相似, 这一方面可能源于地理距离较近种群间基因流动更为频繁, 遗传相似度更高, 另一方面其环境因素更为相似, 对种实性状选择作用趋于一致(图3 ).此外, SL 、PC1随MAP 、MRH 的变化比随MAT 的变化具有更高的一致性, 表明该区域的干旱生境相对于温度对细叶云南松种实性状可能具有更强的选择作用. ...Biostatistical Design and Analysis Using R: A Practical Guide R Core Team, 2018 )计算各性状的种群平均值、标准偏差、种群内及种群间变异系数; 基于巢氏方差分析模型分析种群间及种群内差异, 线性模型为: Yijk = μ + τi + δj ( i ) + εijk , 其中, Yijk 为第i 个种群第j 个家系第k 个观测值, μ 为总平均值, τi 为种群间效应值, δj ( i ) 为种群内单株(家系)随机效应值, εijk 为随机误差(Logan, 2010 ); 利用表型分化系数(VST )来反映种群间及种群内的表型 ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...Shen et al ., 2015 ; 刘雄盛等, 2017 ; 何庆海等, 2018 ; Wu et al ., 2018 ; 刘莹等, 2019 ).细叶云南松种实性状与地理气象因子相关分析结果可以看出, 绝大部分性状与地理气象因子的相关性表现出一致的趋势, 主要表现为与经纬度、年平均气温呈正相关关系, 与相对湿度、年降水量和海拔呈负相关关系, 这意味着随降水量和相对湿度下降, 年平均气温升高, 其球果、种子、种翅及千粒质量有增大趋势.这与同属的白皮松(李斌等, 2002 ), 油松(姬明飞等, 2013 )等有所不同.这说明不同的生境下的树种种实性状受到环境的选择压力不同, 其表型性状变异方向亦不同(Businsky et al ., 2014 ).水分和温度是影响种子萌发、苗木生长的重要环境因素(McLaren & McDonald 2003 ; Bazin et al ., 2011 ), 因而其对种子及苗木生态适应性不可避免具有重要选择作用.一些研究(Andersson, 1996 ; Simons & Johnston, 2000 )表明, 果实及种子大小、千粒质量与生活力和幼苗生长适应性有显著的正相关关系, 换言之, 较大果实及种子有利于其繁殖.因此, 综合以上可以推测, 在南盘江、红水河沿线特殊的干热河谷气候长期作用下(李治基和王献溥, 1981 ), 细叶云南松逐步演化出了适应干旱、高温生境的种实性状, 进而为其在特殊生境下的繁衍、更新奠定了基础.基于相关分析显著的性状(SL )和种群代表性主成分得分(PC1)与地理气象因子的对应分析发现, SL 、PC1随经纬度呈现渐变趋势, 地理距离较近的种群种实形态更为相似, 这一方面可能源于地理距离较近种群间基因流动更为频繁, 遗传相似度更高, 另一方面其环境因素更为相似, 对种实性状选择作用趋于一致(图3 ).此外, SL 、PC1随MAP 、MRH 的变化比随MAT 的变化具有更高的一致性, 表明该区域的干旱生境相对于温度对细叶云南松种实性状可能具有更强的选择作用. ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...潘婷, 2018 ; Yu et al ., 2018 ; 李远发等, 2019 ; 廖良宁等, 2019 ).针对细叶云南松遗传资源评价的研究相对较少, 仅见杨章旗等(2014) 对广西西北部3个种群进行了SSR遗传多样性分析, 认为遗传多样性与欧洲栓皮栎(Quercus suber )、黄杉(Pseudotsuga menziesii )等相比普遍不高, 但种群间基因流动频繁, 分化较小.表型多样性是遗传与环境共同作用的结果, 研究不同地理环境条件下的天然种群的表型性状分化程度和变异规律, 不仅可在一定程度上反映其遗传变异水平, 也可以有效揭示生物对不同环境的适应, 进而探究其发展演化(Donohue et al ., 2001 ; Etterson, 2004 ; Gitonga et al ., 2008 ; Shirley & Vesk, 2009 ).种实性状在物种自然演化过程中具有较高的遗传稳定性, 不易受短期环境变化的影响.同时, 作为主要繁殖器官, 种实性状对物种的定居、繁衍及分布具有重要影响(Cilas et al ., 2009 ).研究表明, 果实形态、种子大小、质量、种翅的有无等都会影响其种子传播媒介(风、动物等)的传播效率, 进而影响物种的分布范围(Wall, 2008 ; Zhang et al ., 2020 ).本文通过对细叶云南松自然分布区的8个天然种群开展种实性状分析, 以揭示细叶云南松种实性状在空间分布上的表型分化和变异规律, 并探究其与细叶云南松特殊生境的关系, 为进一步开展细叶云南松的种质资源保护与利用提供参考. ...潘婷, 2018 ; Yu et al ., 2018 ; 李远发等, 2019 ; 廖良宁等, 2019 ).针对细叶云南松遗传资源评价的研究相对较少, 仅见杨章旗等(2014) 对广西西北部3个种群进行了SSR遗传多样性分析, 认为遗传多样性与欧洲栓皮栎(Quercus suber )、黄杉(Pseudotsuga menziesii )等相比普遍不高, 但种群间基因流动频繁, 分化较小.表型多样性是遗传与环境共同作用的结果, 研究不同地理环境条件下的天然种群的表型性状分化程度和变异规律, 不仅可在一定程度上反映其遗传变异水平, 也可以有效揭示生物对不同环境的适应, 进而探究其发展演化(Donohue et al ., 2001 ; Etterson, 2004 ; Gitonga et al ., 2008 ; Shirley & Vesk, 2009 ).种实性状在物种自然演化过程中具有较高的遗传稳定性, 不易受短期环境变化的影响.同时, 作为主要繁殖器官, 种实性状对物种的定居、繁衍及分布具有重要影响(Cilas et al ., 2009 ).研究表明, 果实形态、种子大小、质量、种翅的有无等都会影响其种子传播媒介(风、动物等)的传播效率, 进而影响物种的分布范围(Wall, 2008 ; Zhang et al ., 2020 ).本文通过对细叶云南松自然分布区的8个天然种群开展种实性状分析, 以揭示细叶云南松种实性状在空间分布上的表型分化和变异规律, 并探究其与细叶云南松特殊生境的关系, 为进一步开展细叶云南松的种质资源保护与利用提供参考. ...R: a Language and Environment for Statistical Computing R Core Team, 2018 )计算各性状的种群平均值、标准偏差、种群内及种群间变异系数; 基于巢氏方差分析模型分析种群间及种群内差异, 线性模型为: Yijk = μ + τi + δj ( i ) + εijk , 其中, Yijk 为第i 个种群第j 个家系第k 个观测值, μ 为总平均值, τi 为种群间效应值, δj ( i ) 为种群内单株(家系)随机效应值, εijk 为随机误差(Logan, 2010 ); 利用表型分化系数(VST )来反映种群间及种群内的表型 ...Haloxylon ammodendron and its affecting factorsShen et al ., 2015 ; 刘雄盛等, 2017 ; 何庆海等, 2018 ; Wu et al ., 2018 ; 刘莹等, 2019 ).细叶云南松种实性状与地理气象因子相关分析结果可以看出, 绝大部分性状与地理气象因子的相关性表现出一致的趋势, 主要表现为与经纬度、年平均气温呈正相关关系, 与相对湿度、年降水量和海拔呈负相关关系, 这意味着随降水量和相对湿度下降, 年平均气温升高, 其球果、种子、种翅及千粒质量有增大趋势.这与同属的白皮松(李斌等, 2002 ), 油松(姬明飞等, 2013 )等有所不同.这说明不同的生境下的树种种实性状受到环境的选择压力不同, 其表型性状变异方向亦不同(Businsky et al ., 2014 ).水分和温度是影响种子萌发、苗木生长的重要环境因素(McLaren & McDonald 2003 ; Bazin et al ., 2011 ), 因而其对种子及苗木生态适应性不可避免具有重要选择作用.一些研究(Andersson, 1996 ; Simons & Johnston, 2000 )表明, 果实及种子大小、千粒质量与生活力和幼苗生长适应性有显著的正相关关系, 换言之, 较大果实及种子有利于其繁殖.因此, 综合以上可以推测, 在南盘江、红水河沿线特殊的干热河谷气候长期作用下(李治基和王献溥, 1981 ), 细叶云南松逐步演化出了适应干旱、高温生境的种实性状, 进而为其在特殊生境下的繁衍、更新奠定了基础.基于相关分析显著的性状(SL )和种群代表性主成分得分(PC1)与地理气象因子的对应分析发现, SL 、PC1随经纬度呈现渐变趋势, 地理距离较近的种群种实形态更为相似, 这一方面可能源于地理距离较近种群间基因流动更为频繁, 遗传相似度更高, 另一方面其环境因素更为相似, 对种实性状选择作用趋于一致(图3 ).此外, SL 、PC1随MAP 、MRH 的变化比随MAT 的变化具有更高的一致性, 表明该区域的干旱生境相对于温度对细叶云南松种实性状可能具有更强的选择作用. ...Eucalyptus phylogenetics: How traits, evolution, and the environment influence the distribution of a diverse genus潘婷, 2018 ; Yu et al ., 2018 ; 李远发等, 2019 ; 廖良宁等, 2019 ).针对细叶云南松遗传资源评价的研究相对较少, 仅见杨章旗等(2014) 对广西西北部3个种群进行了SSR遗传多样性分析, 认为遗传多样性与欧洲栓皮栎(Quercus suber )、黄杉(Pseudotsuga menziesii )等相比普遍不高, 但种群间基因流动频繁, 分化较小.表型多样性是遗传与环境共同作用的结果, 研究不同地理环境条件下的天然种群的表型性状分化程度和变异规律, 不仅可在一定程度上反映其遗传变异水平, 也可以有效揭示生物对不同环境的适应, 进而探究其发展演化(Donohue et al ., 2001 ; Etterson, 2004 ; Gitonga et al ., 2008 ; Shirley & Vesk, 2009 ).种实性状在物种自然演化过程中具有较高的遗传稳定性, 不易受短期环境变化的影响.同时, 作为主要繁殖器官, 种实性状对物种的定居、繁衍及分布具有重要影响(Cilas et al ., 2009 ).研究表明, 果实形态、种子大小、质量、种翅的有无等都会影响其种子传播媒介(风、动物等)的传播效率, 进而影响物种的分布范围(Wall, 2008 ; Zhang et al ., 2020 ).本文通过对细叶云南松自然分布区的8个天然种群开展种实性状分析, 以揭示细叶云南松种实性状在空间分布上的表型分化和变异规律, 并探究其与细叶云南松特殊生境的关系, 为进一步开展细叶云南松的种质资源保护与利用提供参考. ...Lobelia inflata (Campanulaceae): sources and fitness consequencesShen et al ., 2015 ; 刘雄盛等, 2017 ; 何庆海等, 2018 ; Wu et al ., 2018 ; 刘莹等, 2019 ).细叶云南松种实性状与地理气象因子相关分析结果可以看出, 绝大部分性状与地理气象因子的相关性表现出一致的趋势, 主要表现为与经纬度、年平均气温呈正相关关系, 与相对湿度、年降水量和海拔呈负相关关系, 这意味着随降水量和相对湿度下降, 年平均气温升高, 其球果、种子、种翅及千粒质量有增大趋势.这与同属的白皮松(李斌等, 2002 ), 油松(姬明飞等, 2013 )等有所不同.这说明不同的生境下的树种种实性状受到环境的选择压力不同, 其表型性状变异方向亦不同(Businsky et al ., 2014 ).水分和温度是影响种子萌发、苗木生长的重要环境因素(McLaren & McDonald 2003 ; Bazin et al ., 2011 ), 因而其对种子及苗木生态适应性不可避免具有重要选择作用.一些研究(Andersson, 1996 ; Simons & Johnston, 2000 )表明, 果实及种子大小、千粒质量与生活力和幼苗生长适应性有显著的正相关关系, 换言之, 较大果实及种子有利于其繁殖.因此, 综合以上可以推测, 在南盘江、红水河沿线特殊的干热河谷气候长期作用下(李治基和王献溥, 1981 ), 细叶云南松逐步演化出了适应干旱、高温生境的种实性状, 进而为其在特殊生境下的繁衍、更新奠定了基础.基于相关分析显著的性状(SL )和种群代表性主成分得分(PC1)与地理气象因子的对应分析发现, SL 、PC1随经纬度呈现渐变趋势, 地理距离较近的种群种实形态更为相似, 这一方面可能源于地理距离较近种群间基因流动更为频繁, 遗传相似度更高, 另一方面其环境因素更为相似, 对种实性状选择作用趋于一致(图3 ).此外, SL 、PC1随MAP 、MRH 的变化比随MAT 的变化具有更高的一致性, 表明该区域的干旱生境相对于温度对细叶云南松种实性状可能具有更强的选择作用. ...Sapindus mukorossi germplasm with environmental factors in southern ChinaGitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...Bivand et al ., 2013 ; Graler et al ., 2016 ; Tennekes, 2018 ).利用R数据包Hmisc 4.4计算Pearson相关系数及显著性p 值(Harrell & Dupont, 2020 ), 并利用corrplot 0.84绘制相关性图(Wei & Simko, 2017 ).采样地地理气象因子与参试性状间的典型相关分析采用CCA 1.2数据包(González & Déjean, 2012 ), 并采用ggplot2绘图(Wickham, 2016 ). ...潘婷, 2018 ; Yu et al ., 2018 ; 李远发等, 2019 ; 廖良宁等, 2019 ).针对细叶云南松遗传资源评价的研究相对较少, 仅见杨章旗等(2014) 对广西西北部3个种群进行了SSR遗传多样性分析, 认为遗传多样性与欧洲栓皮栎(Quercus suber )、黄杉(Pseudotsuga menziesii )等相比普遍不高, 但种群间基因流动频繁, 分化较小.表型多样性是遗传与环境共同作用的结果, 研究不同地理环境条件下的天然种群的表型性状分化程度和变异规律, 不仅可在一定程度上反映其遗传变异水平, 也可以有效揭示生物对不同环境的适应, 进而探究其发展演化(Donohue et al ., 2001 ; Etterson, 2004 ; Gitonga et al ., 2008 ; Shirley & Vesk, 2009 ).种实性状在物种自然演化过程中具有较高的遗传稳定性, 不易受短期环境变化的影响.同时, 作为主要繁殖器官, 种实性状对物种的定居、繁衍及分布具有重要影响(Cilas et al ., 2009 ).研究表明, 果实形态、种子大小、质量、种翅的有无等都会影响其种子传播媒介(风、动物等)的传播效率, 进而影响物种的分布范围(Wall, 2008 ; Zhang et al ., 2020 ).本文通过对细叶云南松自然分布区的8个天然种群开展种实性状分析, 以揭示细叶云南松种实性状在空间分布上的表型分化和变异规律, 并探究其与细叶云南松特殊生境的关系, 为进一步开展细叶云南松的种质资源保护与利用提供参考. ...Pinus yunnanensis var. tenuifolia )是云南松(P. yunnanensis )因南亚热带独特的河谷气候长期影响及适应下逐渐形成的一个地理变种, 是我国特有种, 其天然分布区域狭窄, 仅限于黔、滇、桂交界处的南盘江流域、北盘江流域以及桂西北的红水河流域, 属于微域分布的植被类型(徐学良, 1983 ; 王献溥, 1991 ).细叶云南松生境狭窄, 属于年平均气温较高, 降水量小于蒸发量, 总体较为干旱的河谷区域, 焚风效应明显, 气候独特.独特的生境使细叶云南松具有较高的生态研究价值(李治基和王献溥, 1981 ).此外, 细叶云南松干型直、树体高大, 出材率高, 木材强度和质量较好, 具有较高的开发利用前景.但近年来, 由于该区域其他经济林树种(油茶(Camellia oleifera )、八角(Illicium verum )等)的大力发展, 对细叶云南松生境的人为干扰急剧加重, 大量细叶云南松林遭到蚕食、破坏, 原有的诸多大面积连续分布林分已消失殆尽, 呈零星点状分布(吴东山等, 2016 ).针对细叶云南松的遗传资源调查、收集、保存、评价迫在眉睫. ...Pinus yunnanensis var. tenuifolia )是云南松(P. yunnanensis )因南亚热带独特的河谷气候长期影响及适应下逐渐形成的一个地理变种, 是我国特有种, 其天然分布区域狭窄, 仅限于黔、滇、桂交界处的南盘江流域、北盘江流域以及桂西北的红水河流域, 属于微域分布的植被类型(徐学良, 1983 ; 王献溥, 1991 ).细叶云南松生境狭窄, 属于年平均气温较高, 降水量小于蒸发量, 总体较为干旱的河谷区域, 焚风效应明显, 气候独特.独特的生境使细叶云南松具有较高的生态研究价值(李治基和王献溥, 1981 ).此外, 细叶云南松干型直、树体高大, 出材率高, 木材强度和质量较好, 具有较高的开发利用前景.但近年来, 由于该区域其他经济林树种(油茶(Camellia oleifera )、八角(Illicium verum )等)的大力发展, 对细叶云南松生境的人为干扰急剧加重, 大量细叶云南松林遭到蚕食、破坏, 原有的诸多大面积连续分布林分已消失殆尽, 呈零星点状分布(吴东山等, 2016 ).针对细叶云南松的遗传资源调查、收集、保存、评价迫在眉睫. ...Bivand et al ., 2013 ; Graler et al ., 2016 ; Tennekes, 2018 ).利用R数据包Hmisc 4.4计算Pearson相关系数及显著性p 值(Harrell & Dupont, 2020 ), 并利用corrplot 0.84绘制相关性图(Wei & Simko, 2017 ).采样地地理气象因子与参试性状间的典型相关分析采用CCA 1.2数据包(González & Déjean, 2012 ), 并采用ggplot2绘图(Wickham, 2016 ). ...ggplot2: Elegant Graphics for Data Analysis Bivand et al ., 2013 ; Graler et al ., 2016 ; Tennekes, 2018 ).利用R数据包Hmisc 4.4计算Pearson相关系数及显著性p 值(Harrell & Dupont, 2020 ), 并利用corrplot 0.84绘制相关性图(Wei & Simko, 2017 ).采样地地理气象因子与参试性状间的典型相关分析采用CCA 1.2数据包(González & Déjean, 2012 ), 并采用ggplot2绘图(Wickham, 2016 ). ...Pinus yunnanensis var. tenuifolia )是云南松(P. yunnanensis )因南亚热带独特的河谷气候长期影响及适应下逐渐形成的一个地理变种, 是我国特有种, 其天然分布区域狭窄, 仅限于黔、滇、桂交界处的南盘江流域、北盘江流域以及桂西北的红水河流域, 属于微域分布的植被类型(徐学良, 1983 ; 王献溥, 1991 ).细叶云南松生境狭窄, 属于年平均气温较高, 降水量小于蒸发量, 总体较为干旱的河谷区域, 焚风效应明显, 气候独特.独特的生境使细叶云南松具有较高的生态研究价值(李治基和王献溥, 1981 ).此外, 细叶云南松干型直、树体高大, 出材率高, 木材强度和质量较好, 具有较高的开发利用前景.但近年来, 由于该区域其他经济林树种(油茶(Camellia oleifera )、八角(Illicium verum )等)的大力发展, 对细叶云南松生境的人为干扰急剧加重, 大量细叶云南松林遭到蚕食、破坏, 原有的诸多大面积连续分布林分已消失殆尽, 呈零星点状分布(吴东山等, 2016 ).针对细叶云南松的遗传资源调查、收集、保存、评价迫在眉睫. ...Pinus yunnanensis var. tenuifolia )是云南松(P. yunnanensis )因南亚热带独特的河谷气候长期影响及适应下逐渐形成的一个地理变种, 是我国特有种, 其天然分布区域狭窄, 仅限于黔、滇、桂交界处的南盘江流域、北盘江流域以及桂西北的红水河流域, 属于微域分布的植被类型(徐学良, 1983 ; 王献溥, 1991 ).细叶云南松生境狭窄, 属于年平均气温较高, 降水量小于蒸发量, 总体较为干旱的河谷区域, 焚风效应明显, 气候独特.独特的生境使细叶云南松具有较高的生态研究价值(李治基和王献溥, 1981 ).此外, 细叶云南松干型直、树体高大, 出材率高, 木材强度和质量较好, 具有较高的开发利用前景.但近年来, 由于该区域其他经济林树种(油茶(Camellia oleifera )、八角(Illicium verum )等)的大力发展, 对细叶云南松生境的人为干扰急剧加重, 大量细叶云南松林遭到蚕食、破坏, 原有的诸多大面积连续分布林分已消失殆尽, 呈零星点状分布(吴东山等, 2016 ).针对细叶云南松的遗传资源调查、收集、保存、评价迫在眉睫. ...Shen et al ., 2015 ; 刘雄盛等, 2017 ; 何庆海等, 2018 ; Wu et al ., 2018 ; 刘莹等, 2019 ).细叶云南松种实性状与地理气象因子相关分析结果可以看出, 绝大部分性状与地理气象因子的相关性表现出一致的趋势, 主要表现为与经纬度、年平均气温呈正相关关系, 与相对湿度、年降水量和海拔呈负相关关系, 这意味着随降水量和相对湿度下降, 年平均气温升高, 其球果、种子、种翅及千粒质量有增大趋势.这与同属的白皮松(李斌等, 2002 ), 油松(姬明飞等, 2013 )等有所不同.这说明不同的生境下的树种种实性状受到环境的选择压力不同, 其表型性状变异方向亦不同(Businsky et al ., 2014 ).水分和温度是影响种子萌发、苗木生长的重要环境因素(McLaren & McDonald 2003 ; Bazin et al ., 2011 ), 因而其对种子及苗木生态适应性不可避免具有重要选择作用.一些研究(Andersson, 1996 ; Simons & Johnston, 2000 )表明, 果实及种子大小、千粒质量与生活力和幼苗生长适应性有显著的正相关关系, 换言之, 较大果实及种子有利于其繁殖.因此, 综合以上可以推测, 在南盘江、红水河沿线特殊的干热河谷气候长期作用下(李治基和王献溥, 1981 ), 细叶云南松逐步演化出了适应干旱、高温生境的种实性状, 进而为其在特殊生境下的繁衍、更新奠定了基础.基于相关分析显著的性状(SL )和种群代表性主成分得分(PC1)与地理气象因子的对应分析发现, SL 、PC1随经纬度呈现渐变趋势, 地理距离较近的种群种实形态更为相似, 这一方面可能源于地理距离较近种群间基因流动更为频繁, 遗传相似度更高, 另一方面其环境因素更为相似, 对种实性状选择作用趋于一致(图3 ).此外, SL 、PC1随MAP 、MRH 的变化比随MAT 的变化具有更高的一致性, 表明该区域的干旱生境相对于温度对细叶云南松种实性状可能具有更强的选择作用. ...Pinus yunnanensis var. tenuifolia )是云南松(P. yunnanensis )因南亚热带独特的河谷气候长期影响及适应下逐渐形成的一个地理变种, 是我国特有种, 其天然分布区域狭窄, 仅限于黔、滇、桂交界处的南盘江流域、北盘江流域以及桂西北的红水河流域, 属于微域分布的植被类型(徐学良, 1983 ; 王献溥, 1991 ).细叶云南松生境狭窄, 属于年平均气温较高, 降水量小于蒸发量, 总体较为干旱的河谷区域, 焚风效应明显, 气候独特.独特的生境使细叶云南松具有较高的生态研究价值(李治基和王献溥, 1981 ).此外, 细叶云南松干型直、树体高大, 出材率高, 木材强度和质量较好, 具有较高的开发利用前景.但近年来, 由于该区域其他经济林树种(油茶(Camellia oleifera )、八角(Illicium verum )等)的大力发展, 对细叶云南松生境的人为干扰急剧加重, 大量细叶云南松林遭到蚕食、破坏, 原有的诸多大面积连续分布林分已消失殆尽, 呈零星点状分布(吴东山等, 2016 ).针对细叶云南松的遗传资源调查、收集、保存、评价迫在眉睫. ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...Pinus yunnanensis var. tenuifolia )是云南松(P. yunnanensis )因南亚热带独特的河谷气候长期影响及适应下逐渐形成的一个地理变种, 是我国特有种, 其天然分布区域狭窄, 仅限于黔、滇、桂交界处的南盘江流域、北盘江流域以及桂西北的红水河流域, 属于微域分布的植被类型(徐学良, 1983 ; 王献溥, 1991 ).细叶云南松生境狭窄, 属于年平均气温较高, 降水量小于蒸发量, 总体较为干旱的河谷区域, 焚风效应明显, 气候独特.独特的生境使细叶云南松具有较高的生态研究价值(李治基和王献溥, 1981 ).此外, 细叶云南松干型直、树体高大, 出材率高, 木材强度和质量较好, 具有较高的开发利用前景.但近年来, 由于该区域其他经济林树种(油茶(Camellia oleifera )、八角(Illicium verum )等)的大力发展, 对细叶云南松生境的人为干扰急剧加重, 大量细叶云南松林遭到蚕食、破坏, 原有的诸多大面积连续分布林分已消失殆尽, 呈零星点状分布(吴东山等, 2016 ).针对细叶云南松的遗传资源调查、收集、保存、评价迫在眉睫. ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...Gitonga et al ., 2008 ; 刁松锋等, 2014 ; Sun et al ., 2017 ).本研究发现细叶云南松种实性状在种群间、种群内均存在极显著差异(p < 0.001), 细叶云南松种实性状变异以种群内为主(VST = 18.65%).这与其近缘种云南松种实性状的研究结果一致(VST = 18.54%)(徐杨等, 2015 ), 高于思茅松(Pinus kesiya var. langbianensis )种群分化(11.95%)(李帅锋等, 2013 ), 低于其他近源科属植物, 如白皮松(P. bungeana ) (22.8%)(李斌等, 2002 ), 油松(P. tabuliformis )(38.97%) (姬明飞等, 2013 ), 云杉(Picea asperata )(30.99%)(罗建勋和顾万春, 2005 )、川西云杉(Picea balfouriana ) (36.53%)(辜云杰等, 2009 ), 岷江柏(Cupressus chengiana )(43.4%)(冯秋红等, 2017 )等的种群分化.总体上, 细叶云南松种群间总体分化水平属于中低水平.这可能因早年细叶云南松在该区域呈连续分布(李治基和王献溥, 1981 ; 徐学良, 1983 ), 种群间地理隔离不明显, 加之风媒植物传粉距离较远, 基因交流较为频繁, 降低了种群间的分化(Mosseler & Rajora, 2007 ).同时, 由于种实主要采集自成年母树而非幼林, 从遗传上说, 其变异情况主要反映的是多年前的情况, 近年来人为干扰对种群分化造成的影响尚未体现. ...潘婷, 2018 ; Yu et al ., 2018 ; 李远发等, 2019 ; 廖良宁等, 2019 ).针对细叶云南松遗传资源评价的研究相对较少, 仅见杨章旗等(2014) 对广西西北部3个种群进行了SSR遗传多样性分析, 认为遗传多样性与欧洲栓皮栎(Quercus suber )、黄杉(Pseudotsuga menziesii )等相比普遍不高, 但种群间基因流动频繁, 分化较小.表型多样性是遗传与环境共同作用的结果, 研究不同地理环境条件下的天然种群的表型性状分化程度和变异规律, 不仅可在一定程度上反映其遗传变异水平, 也可以有效揭示生物对不同环境的适应, 进而探究其发展演化(Donohue et al ., 2001 ; Etterson, 2004 ; Gitonga et al ., 2008 ; Shirley & Vesk, 2009 ).种实性状在物种自然演化过程中具有较高的遗传稳定性, 不易受短期环境变化的影响.同时, 作为主要繁殖器官, 种实性状对物种的定居、繁衍及分布具有重要影响(Cilas et al ., 2009 ).研究表明, 果实形态、种子大小、质量、种翅的有无等都会影响其种子传播媒介(风、动物等)的传播效率, 进而影响物种的分布范围(Wall, 2008 ; Zhang et al ., 2020 ).本文通过对细叶云南松自然分布区的8个天然种群开展种实性状分析, 以揭示细叶云南松种实性状在空间分布上的表型分化和变异规律, 并探究其与细叶云南松特殊生境的关系, 为进一步开展细叶云南松的种质资源保护与利用提供参考. ...潘婷, 2018 ; Yu et al ., 2018 ; 李远发等, 2019 ; 廖良宁等, 2019 ).针对细叶云南松遗传资源评价的研究相对较少, 仅见杨章旗等(2014) 对广西西北部3个种群进行了SSR遗传多样性分析, 认为遗传多样性与欧洲栓皮栎(Quercus suber )、黄杉(Pseudotsuga menziesii )等相比普遍不高, 但种群间基因流动频繁, 分化较小.表型多样性是遗传与环境共同作用的结果, 研究不同地理环境条件下的天然种群的表型性状分化程度和变异规律, 不仅可在一定程度上反映其遗传变异水平, 也可以有效揭示生物对不同环境的适应, 进而探究其发展演化(Donohue et al ., 2001 ; Etterson, 2004 ; Gitonga et al ., 2008 ; Shirley & Vesk, 2009 ).种实性状在物种自然演化过程中具有较高的遗传稳定性, 不易受短期环境变化的影响.同时, 作为主要繁殖器官, 种实性状对物种的定居、繁衍及分布具有重要影响(Cilas et al ., 2009 ).研究表明, 果实形态、种子大小、质量、种翅的有无等都会影响其种子传播媒介(风、动物等)的传播效率, 进而影响物种的分布范围(Wall, 2008 ; Zhang et al ., 2020 ).本文通过对细叶云南松自然分布区的8个天然种群开展种实性状分析, 以揭示细叶云南松种实性状在空间分布上的表型分化和变异规律, 并探究其与细叶云南松特殊生境的关系, 为进一步开展细叶云南松的种质资源保护与利用提供参考. ...Pinus yunnanensis var. tenuifolia secondary forests view supplementary material潘婷, 2018 ; Yu et al ., 2018 ; 李远发等, 2019 ; 廖良宁等, 2019 ).针对细叶云南松遗传资源评价的研究相对较少, 仅见杨章旗等(2014) 对广西西北部3个种群进行了SSR遗传多样性分析, 认为遗传多样性与欧洲栓皮栎(Quercus suber )、黄杉(Pseudotsuga menziesii )等相比普遍不高, 但种群间基因流动频繁, 分化较小.表型多样性是遗传与环境共同作用的结果, 研究不同地理环境条件下的天然种群的表型性状分化程度和变异规律, 不仅可在一定程度上反映其遗传变异水平, 也可以有效揭示生物对不同环境的适应, 进而探究其发展演化(Donohue et al ., 2001 ; Etterson, 2004 ; Gitonga et al ., 2008 ; Shirley & Vesk, 2009 ).种实性状在物种自然演化过程中具有较高的遗传稳定性, 不易受短期环境变化的影响.同时, 作为主要繁殖器官, 种实性状对物种的定居、繁衍及分布具有重要影响(Cilas et al ., 2009 ).研究表明, 果实形态、种子大小、质量、种翅的有无等都会影响其种子传播媒介(风、动物等)的传播效率, 进而影响物种的分布范围(Wall, 2008 ; Zhang et al ., 2020 ).本文通过对细叶云南松自然分布区的8个天然种群开展种实性状分析, 以揭示细叶云南松种实性状在空间分布上的表型分化和变异规律, 并探究其与细叶云南松特殊生境的关系, 为进一步开展细叶云南松的种质资源保护与利用提供参考. ...潘婷, 2018 ; Yu et al ., 2018 ; 李远发等, 2019 ; 廖良宁等, 2019 ).针对细叶云南松遗传资源评价的研究相对较少, 仅见杨章旗等(2014) 对广西西北部3个种群进行了SSR遗传多样性分析, 认为遗传多样性与欧洲栓皮栎(Quercus suber )、黄杉(Pseudotsuga menziesii )等相比普遍不高, 但种群间基因流动频繁, 分化较小.表型多样性是遗传与环境共同作用的结果, 研究不同地理环境条件下的天然种群的表型性状分化程度和变异规律, 不仅可在一定程度上反映其遗传变异水平, 也可以有效揭示生物对不同环境的适应, 进而探究其发展演化(Donohue et al ., 2001 ; Etterson, 2004 ; Gitonga et al ., 2008 ; Shirley & Vesk, 2009 ).种实性状在物种自然演化过程中具有较高的遗传稳定性, 不易受短期环境变化的影响.同时, 作为主要繁殖器官, 种实性状对物种的定居、繁衍及分布具有重要影响(Cilas et al ., 2009 ).研究表明, 果实形态、种子大小、质量、种翅的有无等都会影响其种子传播媒介(风、动物等)的传播效率, 进而影响物种的分布范围(Wall, 2008 ; Zhang et al ., 2020 ).本文通过对细叶云南松自然分布区的8个天然种群开展种实性状分析, 以揭示细叶云南松种实性状在空间分布上的表型分化和变异规律, 并探究其与细叶云南松特殊生境的关系, 为进一步开展细叶云南松的种质资源保护与利用提供参考. ...  ,*广西大学林学院, 国家林业与草原局中南速生材繁育重点实验室, 南宁 530004
,*广西大学林学院, 国家林业与草原局中南速生材繁育重点实验室, 南宁 530004
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT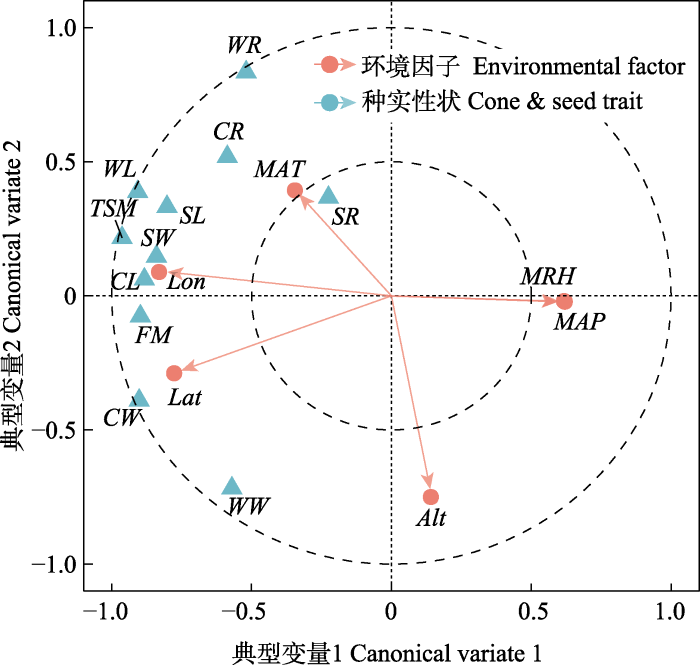 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT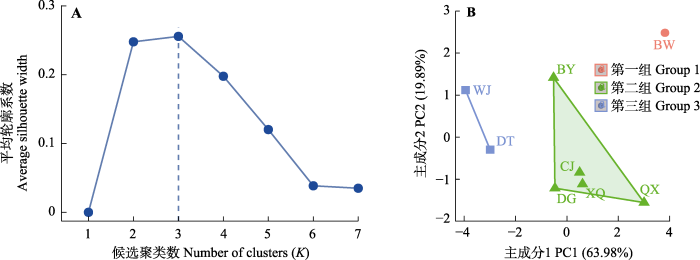 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT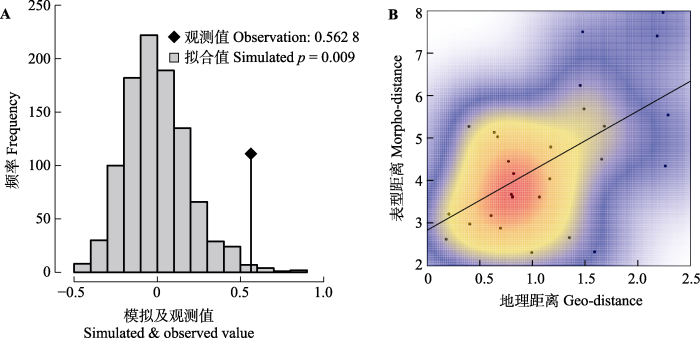 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}