Abstract Aims Seed emergence is a key stage of plant life history and thus of important impacts on interspecific relationships and community composition. Increasing evidence has shown that seed mass determines (or strongly affects) seed emergence within a species, but it has seldom been tested across species at different depths of soil burial. Accordingly, the purpose of this study was to examine the influence of soil burial depths on the relationship between seed mass and seed emergence. Methods We performed a full-factorial experiment. Two factors included plant species (i.e. Achillea millefolium, Achnatherum sibiricum, Chenopodium glaucum, Centaurea maculosa, Medicago falcata, Poa pratensis, P. secunda, Solidago canadensis, Stipa bungeana and Vulpia octoflora) and soil burial depth (i.e. 0, 1, 2, 4 and 8 cm). Each combination was replicated five times, totaling 250 pots (10 species × 5 burial depths × 5 replicates). For each species, seed mass was determined prior to sowing at a given soil burial depth. We watered the pots and recorded seed emergence every day for 30 days. We determined the first date of seed emergence, and calculated the overall seed emergence, the tolerance to soil burial, and T50 (i.e. the number of days in the presence of 50% emergence). Seed mass and seed emergence were analyzed using an analysis of variance, and the relationships between two variables were tested using correlation analyses. Important findings We found that: (1) The optimum soil burial depth for seed emergence varied among the species, and for most of tested species this optimum depth ranged from 0 to 2 cm (i.e. shallow soil); six species had the maximum seed emergence at 1 cm soil burial depth, and no species had the maximum seed emergence at 4 or 8 cm. (2) There was a positive correlation between seed mass and the tolerance to soil burial across the 10 tested species, specifically, the tolerance to soil burial and the optimum depths increased with seed mass. (3) At deeper soil burial depths (i.e. 4 or 8 cm), there were significantly positive correlations among seed mass, seed emergence, and T50; in contrast, such correlations did not occur at shallower burial depths (i.e. 0, 1 or 2 cm). Keywords:soil burial depth;seed mass;seed emergence rate;emergence speed;tolerance to soil burial
Fig. 1The mass per seed of the ten species used in the experiment (mean ± SE). Different lowercase letters indicate significant differences in mass per seed among species (p < 0.05). Am, Achillea millefolium; As, Achnatherum sibiricum; Cg, Chenopodium glaucum; Cm, Centaurea maculosa; Mf, Medicago falcata; Pp, Poa pratensis; Ps, Poa secunda; Sb, Stipa bungeana; Sc, Solidago canadensis; Vo, Vulpia octoflora.
2.2 种子出苗
2.2.1 首次出苗时间
种子质量(F = 40.49, p < 0.001)、土埋深度(F = 48.26, p < 0.001)以及两者交互作用(F = 4.95, p < 0.001)显著影响种子首次出苗时间。总体上, 种子首次出苗时间随着土壤深度增加而延长(图2), 例如, 长芒草种子首次出苗时间随土壤深度的增加依次为: 6、7、8、9和10天; 草地早熟禾种子首次出苗时间随土壤深度的增加分别为7、9、10和21天(8 cm土埋时出苗率为0, 所以没有首次出苗时间)。种子质量的增大可使较深土埋种子的首次出苗时间提前。
2.2.2 最大出苗率
种子质量(F = 43.91, p < 0.001)、土埋深度(F = 190.32, p < 0.001)以及两者交互作用(F = 17.15, p < 0.001)显著影响种子出苗率。不同植物的种子出苗率存在较大差异, 羽茅、灰绿藜和加拿大一枝黄花种子最大出苗率相对较低, 分别为22%、8%和19%; 而千叶蓍、草地早熟禾、Poa secunda和Vulpia octoflora具有较高的最大出苗率, 分别为82%、77%、75%和89%; 长芒草、野苜蓿和Centaurea maculosa则介于两者之间, 分别为47%、51%和61% (图2)。
Fig. 3Relationship between seed mass and the tolerance to soil burial. Each point represents 10 species (mean ± SE). r represents the correlation coefficient between two variables, and the p represents probability.
Fig. 4Illustrations of seed mass vs. seed emergence and seed mass vs. T50 at five different soil burial depths. The ten data points per panel represent the different plant species (mean ± SE).
Fig. 5Relationships between seed emergence and T50 at five different soil burial depths. Each point per panel represents one tested plant species (mean ± SE).
. GaoRR, ZhaoRH, HuangZY, YangXJ, WeiXY, HeZ, WalckJL (2018). Soil temperature and moisture regulate seed dormancy cycling of a dune annual in a temperate desert Environmental and Experimental Botany, 155, 688-694. DOI:10.1016/j.envexpbot.2018.08.010URL [本文引用: 1]
. GardarinA, DürrC, ManninoMR, BussetH, ColbachN (2010). Seed mortality in the soil is related to seed coat thickness Seed Science Research, 20, 243-256. DOI:10.1017/S0960258510000255URL [本文引用: 1]
. GuilleminJP, ChauvelB (2011). Effects of the seed weight and burial depth on the seed behavior of common ragweed (Ambrosia artemisiifolia). Weed Biology and Management, 11, 217-223. DOI:10.1111/j.1445-6664.2011.00423.xURL [本文引用: 1] Common ragweed (Ambrosia artemisiifolia L.) is one of the annual plants that were described recently as invasive weeds in Europe. This species is described as an invasive plant that produces seeds that are highly variable. Its production of variably sized seeds is regarded as promoting its spread in different environments. Experiments were carried out to determine the influence of the seed weight and temperature on germination and the influence of the seed weight and burial depth on seedling emergence. The seeds were divided into a number of classes of weight and the seed weight effect on germination was evaluated by Petri dish assays. In another experiment, the seeds were buried at different depths in a clay soil/sand mix to estimate the burial effect on germination and seedling emergence. The germination level of A. artemisiifolia was high overall, between 76.8% and 94.2%. The seed germination was modified by temperature but it was not influenced by the seed weight. The amounts of germination and seedling emergence were greater for the seeds on the soil surface and decreased with an increasing burial depth, from 2 to 8 cm. No germination or emergence was observed for the seeds that were buried at 10 and 12 cm. The lightest seeds were more sensitive to burial. A greater level of seedling emergence for those seeds that were placed near the soil surface could explain the success of this species in open habitats, where the probability of deeper burial is low. After high seed production, the management of A. artemisiifolia in fields could be partly achieved through soil tillage, burying seeds below 10 cm, and not carrying out deep soil tillage the following year.
. GuoCR, LuJQ, YangDZ, ZhaoLP (2009). Impacts of burial and insect infection on germination and seedling growth of acorns ofQuercus variabilis. Forest Ecology and Management, 258, 1497-1502. DOI:10.1016/j.foreco.2009.05.044URL [本文引用: 5]
. HarperJL ( 2010). Biology of Plant Populations. Blackburn Press, United States. [本文引用: 5]
. HarperJL, BentonRA (1966). The behaviour of seeds in soil: The germination of seeds on the surface of a water supplying substrate Journal of Ecology, 54, 151-166. DOI:10.2307/2257664URL [本文引用: 2]
. HeYH, ZhaoHL, LiuXP, ZhangTH ( 2008). The effect of seed size variation of Caragana microphylla on seed germination and seedling growth. Seed, 27(8), 10-13. [本文引用: 1]
. JavaidMM, TanveerA (2014). Germination ecology of Emex spinosa and Emex australis, invasive weeds of winter crops. Weed Research, 54, 565-575. DOI:10.1111/wre.12111URL [本文引用: 1] Emex spinosa and Emex australis are invasive dicotyledonous weeds. The effects of various environmental factors on the germination of these weeds were investigated under laboratory and glasshouse conditions. Germination response of both species was lower at warmer temperature, and maximum germination was recorded at 20/12 degrees C (day/night). Light stimulated germination in both species, but considerable germination also occurred under darkness. More than 80% of E.spinosa seeds germinated at pH between 6 and 9, whereas E.australis seeds germination was considerably decreased at pH 9. Emex spinosa was fairly tolerant to salinity as compared with E.australis and germination (21%) of E.spinosa occurred even at 200mm NaCl. Both species were sensitive to osmotic stress, but E.spinosa tolerated more osmotic stress than E.australis. Temperature above 20/12 degrees C (day/night) and low osmotic potential increased time to start germination and mean germination time (MGT), as well as decreased germination index (GI) of both species. Darkness resulted in increased MGT and decreased GI in both species when compared with 10h photoperiod. Salt stress strongly increased time to obtain 50% germination and reduced GI of both species. In both species, an increasing burial depth decreased emergence percentage and emergence index and increased time to start emergence, although some seed emerged even at 10cm burial depth. It was concluded that both species can germinate over a wide range of environmental conditions. However, E.australis was more sensitive under adverse environmental conditions compared with E.spinosa. This information on germination ecology may aid in developing tools and strategies for management.
. KondoT, SatoC (2007). Effects of temperature, light, storage conditions, sowing time, and burial depth on the seed germination of Cardiocrinum cordatum var. glehnii(Liliaceae). Landscape and Ecological Engineering, 3, 89-97. DOI:10.1007/s11355-007-0022-3URL [本文引用: 1] In this study, we conducted experiments to accumulate practical information on the propagation and establishment of a population of Cardiocrinum cordatum var. glehnii by seed sowing. C. cordatum var. glehnii seeds require approximately 19months from seed dispersal to cotyledon emergence in the field. However, the period from seed dispersal to radicle emergence was shortened to approximately 7–8months by the temperature transition of 25/15°C (60days)→15/5°C (30days)→0°C (120days)→15/5°C (i.e., 15/5°C represents alternating temperature treatment wherein the seeds were placed at 15°C for 12h during the day and then at 5°C for 12h during the night). More than 90% of the seeds, which were stored dry at 5°C for 12months and sown in pots in the field, showed cotyledon emergence, whereas in seeds stored dry at 25°C, dry at room temperature, and non-dry at room temperature, cotyledon emergence was decreased by less than 1%. More than 88% of the seeds that were stored dry at 5°C and sown in the field in October 2002 immediately after collecting, November, and from April to July 2003 showed cotyledon emergence in spring 2004. However, seeds sown in August, September, and October 2003 showed cotyledon emergences of 57.6%, 0%, and 0% in spring 2004, respectively. Seeds collected in October 2002 and sown until July 2003 in the field received adequate high temperature in summer, moderate temperature in autumn, and cold temperature in winter; therefore, the percentage of cotyledon emergence was high in spring 2004. On the other hand, seeds sown in August 2003 or later could not receive enough high temperature; thus, cotyledons emerged from only a few seeds.
. LiQY, ZhaoWZ, FangHY (2006). Effects of sand burial depth and seed mass on seedling emergence and growth of Nitraria sphaerocarpa. Plant Ecology, 185, 191-198. DOI:10.1007/s11258-005-9094-zURL [本文引用: 6] A greenhouse experiment was conducted to test the effects of sand burial depth and seed mass on seedling emergence and growth of Nitraria sphaerocarpa. Seeds of Nitraria sphaerocarpa were sorted into three size-classes (large, medium, small) and artificially buried at 0, 1, 2, 3, 4, 5 and 6cm depths in plastic pots filled with unsterilized sand. In the seven treatments, the percent emergence, seedling mass and seedling height, significantly affected by both burial depth and seed size, were highest at the optimal burial depth of 2cm burial depth, and decreased with increasing burial depth in each seed size-class. Although seedling mass was usually greatest for large seeds and least for small seeds at each burial depth, little difference was observed in seedling height at shallow burial depths of 0–3cm. In each seed size-class, with increasing burial depth, both root-mass ratio and aboveground stem-mass ratio decreased, while belowground stem-mass ratio increased. In each burial depth, with decreasing seed size, belowground stem-mass ratio increased, while root-mass ratio decreased.
. LiZ, FuAL, ShaWL, ZhengXH, YangG ( 2004). The influence of sowing depth to the germination of Caretoides ewermanniana(Stschegl. ex Losinsk.) Botsch. et Ikonn. Grass-Feeding Livestock, ( 4), 66-67. URL [本文引用: 1]
. LiuGX, HanJG, ZhaoX ( 2007). Effects of sowing depth and seed source on seedling emergence of Leymus chinensis. Agricultural Research in the Arid Areas, 25(5), 69-74. [本文引用: 1]
. AhmedS, Ope?aJL, ChauhanBS (2015). Seed germination ecology of doveweed (Murdannia nudiflora) and its implication for management in dry-seeded rice. Weed Science, 63, 491-501. DOI:10.1614/WS-D-14-00115.1URL [本文引用: 1]
. AminiR, GholamiF, GhanepourS (2017). Effects of environmental factors and burial depth on seed germination and emergence of two populations of Caucalis platycarpos. Weed Research, 57, 247-256. DOI:10.1111/wre.2017.57.issue-4URL [本文引用: 2]
. AminiR, MobliA, GhanepourS (2016). Effect of environmental factors on seed germination and emergence of Lepidium vesicarium Plant Species Biology, 31, 178-187. DOI:10.1111/psbi.2016.31.issue-3URL [本文引用: 3]
. Bani-AameurF, Sipple-MichmerhuizenJ (2001). Germination and seedling survival of argan (Argania spinosa) under experimental saline conditions. Journal of Arid Environments, 49, 533-540. DOI:10.1006/jare.2001.0804URL [本文引用: 1]
. MüllerFL, RaittL, CysterL (2018). Impact of seed burial depth on the emergence and growth of annual forage legumes South African Journal of Plant and Soil, 35, 71-74. DOI:10.1080/02571862.2017.1319505URL [本文引用: 4]
. NieCL, ZhengYR ( 2005). Effects of water supply and sand burial on seed germination and seedling emergence of four dominant psammophytes in the Ordos Plateau Acta Phytoecologica Sinica, 29, 32-41. URL [本文引用: 1] 羊柴 (Hedysarum leave)、柠条 (Caragana korshinskii)、籽蒿 (Artemisia sphaerocephala)与油蒿 (Artemisia ordosica)为鄂尔多斯高原广泛分布的植物种,也是该地区飞播选用的主要植物种。由于飞播为鄂尔多斯高原植被恢复的重要手段,而飞播后植物种子的出苗率与土壤水分及沙埋密切相关。因此,上述植物种子萌发与出苗对水分和沙埋反应的研究对当地退化生态系统的恢复具有重要意义。通过温室控制实验,对4种植物萌发特性及出苗与水分和沙埋深度的关系进行研究,阐明了种子萌发最适合的水分条件和沙埋深度。结果表明,4种植物种子萌发与出苗最适宜的供水量都接近于当地种子萌发时期的平均降水量, 最适宜的沙埋深度为0.5~1 cm, 过多的水分和过深的沙埋会降低种子萌发与出苗,反映了4种植物对沙区环境的生态适应。4种植物最适宜的供水量均为123 ml,羊柴、柠条、籽蒿和油蒿的最适沙埋深度分别为:0.5、1.0、0.5和0.5 cm。根据实验结果及当地气候特点,鄂尔多斯地区的飞播时间应由6月初提前至5月中下旬,以提高飞播植物的出苗率。
. BaskinCC, BaskinJM ( 1998). Seeds. Ecology, Biogeography, and Evolution of Dormancy and Germination. Academic Press, San Diego, USA. 212-534. [本文引用: 10]
. ChenH, MaunMA (1999). Effects of sand burial depth on seed germination and seedling emergence of Cirsium pitcheri. Plant Ecology, 140, 53-60. DOI:10.1023/A:1009779613847URL [本文引用: 4]
. NobleIR (1989). Attributes of invaders and the invading process: Terrestrial and vascular plants. In: Drake JA, Mooney HA, di Castri F, Groves RH, Kruger FJ, Rejmanek M, Williamson MH eds. Biological Invasions: A Global Perspective. Wiley, Chichester. 301-313. [本文引用: 1]
. ChenZH, PengJF, ZhangDM, ZhaoJG (2002). Seed germination and storage of woody species in the lower subtropical forest Acta Botanica Sinica, 44, 1469-1476. [本文引用: 2]
. PazH, MazerSJ, Martinez-RamosM (2005). Comparative ecology of seed mass in Psychotria(Rubiaceae): Within- and between-species effects of seed mass on early performance. Functional Ecology, 19, 707-718. DOI:10.1111/fec.2005.19.issue-4URL [本文引用: 1]
. PengHJ ( 2001). Effects of seed size and seedling depth on emergence of six perennial grasses Pratacultural Science, 18(6), 30-35. [本文引用: 1]
. RenJ, TaoL, LiuXM (2002). Effect of sand burial depth on seed germination and seedling emergence of Calligonum L. species. Journal of Arid Environments, 51, 603-611. URLPMID:15624824 [本文引用: 1] This paper studied the effects of sand-burying on the seed germination and seedling emergence of six dominance plants in Keerqin Sandy Land. Six treatments of artificially sand-burying to depths 0, 2, 4, 6, 8, 10 and 12 cm were designed. The results showed that the seed germination of Agriophyllum squarrosum was significantly different (P &lt; 0.05) between different treatments, but that of Artemesia halodendron was only significantly different (P&lt; 0.05) between 2 cm and other burial depths. For other four plants, their seed germination was significantly different (P &lt; 0.05) only between 0 cm and other burial depths. Sand-burying significantly affected the seedling emergence of all plant species. The emergence rate decreased with increasing burial depth. Higher emergence percentages were obtained at 2 and 4 cm burial depths. The species with larger seeds emerged from deeper depth, and the maximum burial depth of seedling emergence was Sophora flavescdens &gt; Atraphaxis manshurica &gt; or = Agriophyllum squarrosum &gt; Hedysarum fruticosum &gt; Bassia dasyphylla &gt; Artemesia halodendron.
. SeiwaK, WatanabeA, SaitohT, KannoH, AkasakaS (2002). Effects of burying depth and seed size on seedling establishment of Japanese chestnuts, Castanea crenata. Forest Ecology and Management, 164, 149-156. DOI:10.1016/S0378-1127(01)00607-7URL [本文引用: 1]
. SimonsAM, JohnstonMO (2000). Variation in seed traits of Lobelia inflata(Campanulaceae): Sources and fitness consequences. American Journal of Botany, 87, 124-132. URLPMID:10636835 [本文引用: 1] Seed germination constitutes an important event in the life cycle of plants. Two related seed traits affect fitness: seed size and the timing of seed germination. In three sets of experiments, we (1) partition the sources of seed-size variance in Lobelia inflata into components attributable to fruit size, relative fruit position, and parental identity; (2) examine the influence of pregermination conditions and seed size on time to germination; and (3) assess the fitness consequences of seed size and germination timing under seminatural, harsh conditions. Seed-size variance is attributable to both parental identity and fruit position within an individual. Distal fruits produce larger but fewer seeds. No significant correlation exists between fruit size and seed size, but a trade-off is found between the number and size of seeds contained in a fruit after correcting for fruit size. The timing of germination is influenced by seed size, light conditions before winter, and winter duration. Germination timing influences survival, and despite small seed size in this species (2 × 10 g/seed), seed size has a persistent and significant association with both final plant size and the probability of survival to autumn.
. StantonML (1984). Seed variation in wild radish: Effect of seed size on components of seedling an adult fitness Ecology, 65, 1105-1112. DOI:10.2307/1938318URL [本文引用: 1]
. WangGH, YuKL, GouQQ (2018). Effects of sand burial disturbance on establishment of three desert shrub species in the margin of oasis in northwestern China Ecological Research, 34, 127-135. DOI:10.1111/ere.2019.34.issue-1URL [本文引用: 3]
. WinnAA (1988). Ecological and evolutionary consequences of seed size in Prunella vulgaris. Ecology, 69, 1537-1544. DOI:10.2307/1941651URL [本文引用: 2]
. WuGL, DuGZ ( 2008). Relationships between seed size and seedling growth strategy of herbaceous plant Chinese Journal of Applied Ecology, 19, 191-197. URLPMID:18419094 [本文引用: 1] Seed size and seedling recruitment strategy are of importance in the life-history strategy of plant. In this paper, the current ecological researches at home and aboard on the relationships between seed size and seedling growth were reviewed from the aspects of the effects of seed size on seed germination and seedling emergence, the relationships between seed size and seedling growth traits, and the relationships between seed size and seedling survival and competition ability. Some suggestions on future researches in this field were put forward. There were likely different relationships between seed size and seedling growth in different microenvironments and vegetation types, and the effects of seed size on seedling growth could result in different contributions of different seed-size species to the seedling recruitment of vegetation. The large-scale community level and the small-scale intra- and inter-species level researches on this issue should be strengthened, which would have significance for the recruitment and renewing of natural vegetation. [ 武高林, 杜国祯 ( 2008). 植物种子大小与幼苗生长策略研究进展 应用生态学报, 19, 191-197.] URLPMID:18419094 [本文引用: 1] Seed size and seedling recruitment strategy are of importance in the life-history strategy of plant. In this paper, the current ecological researches at home and aboard on the relationships between seed size and seedling growth were reviewed from the aspects of the effects of seed size on seed germination and seedling emergence, the relationships between seed size and seedling growth traits, and the relationships between seed size and seedling survival and competition ability. Some suggestions on future researches in this field were put forward. There were likely different relationships between seed size and seedling growth in different microenvironments and vegetation types, and the effects of seed size on seedling growth could result in different contributions of different seed-size species to the seedling recruitment of vegetation. The large-scale community level and the small-scale intra- and inter-species level researches on this issue should be strengthened, which would have significance for the recruitment and renewing of natural vegetation.
. WuX, LiJ, XuHL, DongLY (2015). Factors affecting seed germination and seedling emergence of Asia minor bluegrass (Polypogon fugax). Weed Science, 63, 440-447. DOI:10.1614/WS-D-14-00093.1URL [本文引用: 1]
. YangHL, CaoZP, DongM, YeYZ, HuangZY ( 2007). Effects of sand burying on caryopsis germination and seedling growth of Bromus inermis Leyss. Chinese Journal of Applied Ecology, 18, 2438-2443. URLPMID:18260444 [本文引用: 1] This paper studied the effects of different depth sand burying on the caryopsis germination, seedling emergence and growth of Bromus inermis Leyss., a perennial gramineous grass dominated on Othidag sandy land. The results showed that the maximum burying depth for B. inermis caryopsis germination and seedling emergence was 12 cm and 8 cm, respectively. Below the thresholds, the percentages of caryopsis germination and seedling emergence decreased with increasing depth of sand burying. All the 1- and 2-week-old seedlings of B. inermis could survive when the sand burying depth was up to 33% of their height, and their biomass was no less than the unburied seedlings. When the burying depth increased to 66% of their height, 70% of 1-week-old seedlings and 25% of 2-week-old seedlings could survive, but their growth was impaired. No seedlings could survive when they were wholly buried in sand. When buried by sand, the seedlings allocated more biomass for the development of their aboveground part, which might be an important adaptation strategy of B. inermis to sand burying. [ 杨慧玲, 曹志平, 董鸣, 叶永忠, 黄振英 ( 2007). 沙埋对无芒雀麦种子萌发和幼苗生长的影响 应用生态学报, 18, 2438-2443.] URLPMID:18260444 [本文引用: 1] This paper studied the effects of different depth sand burying on the caryopsis germination, seedling emergence and growth of Bromus inermis Leyss., a perennial gramineous grass dominated on Othidag sandy land. The results showed that the maximum burying depth for B. inermis caryopsis germination and seedling emergence was 12 cm and 8 cm, respectively. Below the thresholds, the percentages of caryopsis germination and seedling emergence decreased with increasing depth of sand burying. All the 1- and 2-week-old seedlings of B. inermis could survive when the sand burying depth was up to 33% of their height, and their biomass was no less than the unburied seedlings. When the burying depth increased to 66% of their height, 70% of 1-week-old seedlings and 25% of 2-week-old seedlings could survive, but their growth was impaired. No seedlings could survive when they were wholly buried in sand. When buried by sand, the seedlings allocated more biomass for the development of their aboveground part, which might be an important adaptation strategy of B. inermis to sand burying.
. YeXH, LiLL, BaskinCC, BaskinJM, DuJ, HuangZY (2019 a). Sand burial helps regulate timing of seed germination of a dominant herb in an inland dune ecosystem with a semiarid temperate climate Science of the Total Environment, 680, 44-50. DOI:10.1016/j.scitotenv.2019.05.087URLPMID:31100667 [本文引用: 1] Sand burial plays important role in the life history of species in sandy areas of arid and semi-arid temperate regions, by affecting seed germination, seedling growth and survival and plant community. However, few studies have focused on the effects of sand burial on the dynamics of seed dormancy in such areas. In this study, seed germination characteristics of Allium tenuissimum, a dominant perennial herb in the dune ecosystem in Ordos Plateau in northern China, was investigated. Dormancy dynamics were monitored for seeds buried at sand depths of 0, 2 and 5?cm in the natural habitats for 13?months. Freshly matured seeds of A. tenuissimum were conditionally dormant and germinated to high percentages at high but not low temperatures. Germination percentages were increased by incubation at summer temperature (15/25?°C) and dry storage, but GA3 and cold stratification had no significant effects. These results suggested that seeds of A. tenuissimum were conditionally dormant, and incubation under warm, wet (but not cold, wet) conditions and dry after-ripening resulted in complete dormancy releasing. Seeds buried at 2 and 5?cm in the field for 13?months exhibited seasonal changes in germination, with an increase in spring and summer followed by decrease in autumn and winter. However, seeds on the soil surface (0?cm) gradually came out of dormancy beginning in January, and germinated percentage was highest in July, after which remained high. The peak of nondormancy for seeds buried in the field was in the summer, which corresponds to the period of highest annual precipitation. Sand burial helped mediate seeds dormancy dynamics of A. tenuissimum via seasonal dormancy cycling. Our finding contributes to a better understanding of how timing of seedling establishment is controlled in a semiarid dune ecosystem in temperate climate.
. YeXH, LiuZL, GaoSQ, CuiQG, LiuGF, DuJ, DongM, HuangZY, CornelissenJHC (2016). Differential plant species responses to interactions of sand burial, precipitation enhancement and climatic variation promote co-existence in Chinese steppe vegetation Journal of Vegetation Science, 28, 139-148. DOI:10.1111/jvs.2017.28.issue-1URL [本文引用: 1]
. YeXH, LiuZL, ZhangSD, GaoSQ, LiuGF, CuiQG, DuJ, HuangZY, CornelissenJHC (2019b). Experimental sand burial and precipitation enhancement alter plant and soil carbon allocation in a semi-arid steppe in north China Science of the Total Environment, 651, 3099-3106. DOI:10.1016/j.scitotenv.2018.10.208URLPMID:30463160 [本文引用: 1] Sand burial is a common phenomenon in inland semi-arid and arid areas, affecting plant growth and even plant community structure. Precipitation regime, including the variation of precipitation intensity and frequency, also drives community structure and functions in such areas. However, few studies have focused on the combined effect of sand burial and changed precipitation regime on community function, specifically its role in carbon storage. A 2-yr field experiment with factorial treatments of precipitation (control, slight enhancement and strong enhancement) and sand addition (control and 5?cm sand addition) was conducted to test the responses of plant and soil carbon content in a semi-arid typical steppe in N China. Results showed that sand burial had no significant effect on plant carbon density, but significantly changed the allocation of plant carbon from aboveground to belowground; these responses differed among species and life forms in the community. Precipitation enhancement had no significant effects on plant carbon and its allocation, perhaps because effects of precipitation on plants are due more to precipitation frequency than to precipitation intensity per event. Sand burial and precipitation enhancement decreased soil carbon, especially soil organic carbon, and promoted soil carbon to be distributed deeper down the soil profile. These findings will help to understand how sand deposition affects plant and soil carbon storage and their allocation in plant communities under a changing precipitation regime, and more generally, to understand carbon storage dynamics in early-successional sandy ecosystems in the context of global change.
. ZhongJD, ZhouXX, LiXL, YuanCC, LiuJX, LiuWG ( 2017). Effects of different seeding depths on seedling emergence and growth of Bidens alba, Praxelis clematidea and Ageratum conyzoides. Chinese Journal of Tropical Agriculture, 37(1), 1-6. [本文引用: 1]
. ZhuYJ, YangXJ, BaskinCC, BaskinJM, DongM, HuangZY (2014). Effects of amount and frequency of precipitation and sand burial on seed germination, seedling emergence and survival of the dune grass Leymus secalinus in semiarid China. Plant and Soil, 374, 399-409. DOI:10.1007/s11104-013-1892-9URL [本文引用: 1] Seed germination and seedling emergence are vulnerable to water stress in arid environments. When precipitation is low and unpredictable during the early growing season, seeds near the sand surface often suffer from hydration/dehydration during germination. We investigated the responses of seedling emergence and survival of a sand dune grass with high sand stabilization value to amount and frequency of precipitation and depth of burial in sand. Effects of amount and frequency of precipitation, burial and hydration/dehydration on seedling emergence of Leymus secalinus, were examined using standard procedures. Seedling emergence was affected by amount and frequency of monthly precipitation and depth of burial, and it decreased as precipitation frequency decreased with same amount of precipitation. Highest emergence percentage was obtained with 100 or 150 mm precipitation at 1-4 cm depth. Hydration/dehydration treatments decreased germination and increased dormancy percentage. Young seedlings with root lengths of 0-1 mm desiccated up to 30 days revived after rehydration. Seedling emergence of L. secalinus is adapted to 150 mm monthly precipitation with frequency of 10-30 times per month, 1-4 cm burial depth and dehydration interval of 1-2 days. Alteration of amount and/or frequency of precipitation caused by climate change could markedly affect seedling emergence and population regeneration of this species.
. DolanRW (1984). The effect of seed size and maternal source on individual size in a population of Ludwigia leptocarpa(Onagraceae). American Journal of Botany, 71, 1302-1307. DOI:10.1002/ajb2.1984.71.issue-9URL [本文引用: 1]
Soil temperature and moisture regulate seed dormancy cycling of a dune annual in a temperate desert 1 2018
... 种子出苗是植物生活史中极为重要的一个阶段(Harper & Benton, 1966; Harper, 2010; Ahmed et al., 2015).这个阶段对植物种群数量增长、种间竞争、群落组成、食物链等具有重要的调控作用(Stanton, 1984; Bani-Aameur & Sipple-Michmerhuizen, 2001; Ye et al., 2016).种子出苗代表着由对环境条件耐受性最强阶段(种子休眠)向最脆弱、最敏感阶段的高风险转变(Harper & Benton, 1966; Harper, 2010), 极易受到众多外界环境因素, 如温度、光照、土壤含水量、氧气含量和土壤埋藏深度等(Wu et al., 2015; Amini et al., 2016; Gao et al., 2018; Ye et al., 2019b)的影响.因此, 研究种子出苗过程及其驱动因素具有重要的生态学意义. ...
Seed mortality in the soil is related to seed coat thickness 1 2010
Impacts of burial and insect infection on germination and seedling growth of acorns ofQuercus variabilis. 5 2009
... 种子出苗受遗传和环境的共同影响(Baskin & Baskin, 1998; Li et al., 2006).种子大小是植物相对稳定的一个内在特征, 它与种子传播、萌发、出土和幼苗竞争等过程密切相关(武高林和杜国祯, 2008).比如, 较大种子总体上比较小种子具有更高的出苗率(Winn, 1988), 幼苗具有更大的竞争力和更高的存活力(Simons & Johnston, 2000; Li et al., 2006).当然也有相反的报道(Dolan, 1984).种子质量是衡量种子大小的重要指标, 种子质量与耐土壤埋藏能力之间存在正相关关系(Chen & Maun, 1999; Chen et al., 2002).较大质量种子往往储存更多的物质和能量, 因而可能从更深的土壤中顺利出苗(聂春雷和郑元润, 2005; Li et al., 2006; Guo et al., 2009; Müller et al., 2018).也有研究发现种子大小与出苗率之间没有直接的相关性(何玉惠等, 2008). ...
... 种子萌发后能否顺利出土不仅与自身储存的物质和能量有关, 还取决于它在土壤中的位置(Baskin & Baskin, 1998).换言之, 土壤埋藏深度是决定种子命运的关键性因素.一方面, 适当深度的土壤埋藏有助于为种子创造适宜的温度、水分等出苗条件(Baskin & Baskin, 1998; Li et al., 2006; Guo et al., 2009).另一方面, 埋藏过深会对种子出苗产生阻碍或抑制(Baskin & Baskin, 1998; 李柱等, 2004; Li et al., 2006; Guo et al., 2009).种子出苗率随埋藏深度的增加而逐渐减小(Amini et al., 2017), 这与种子出苗前储存物质逐渐耗尽有关(Javaid & Tanveer, 2014; Amini et al., 2016).最适宜种子出苗的埋藏深度往往位于较浅土层, 并随土埋深度的增加而降低(Amini et al., 2017; Wang et al., 2018). ...
... ; Guo et al., 2009).种子出苗率随埋藏深度的增加而逐渐减小(Amini et al., 2017), 这与种子出苗前储存物质逐渐耗尽有关(Javaid & Tanveer, 2014; Amini et al., 2016).最适宜种子出苗的埋藏深度往往位于较浅土层, 并随土埋深度的增加而降低(Amini et al., 2017; Wang et al., 2018). ...
... 我们的结果表明, 在物种水平, 种子质量依然是决定种子出苗的一个重要内在因素.这与以前大量的种内研究结果(Winn, 1988; Baskin & Baskin, 1998; Li et al., 2006; Guo et al., 2009; Müller et al., 2018)一致.有利于不同物种种子出苗的适宜土埋深度各不相同, 较浅的土壤埋藏深度对于种子出苗更加有利(Wang et al., 2018).我们发现, 10种植物种子出苗的最适土壤条件都在较浅土层, 特别是在1 cm的土层; 土壤表面撒种或是较深土层播种会在一定程度上降低种子的出苗率.这些结果与Harper和Benton (1966)的研究结果相一致.最适出苗的土埋深度与种子出苗所需的氧气、光照、温度和水分等条件有关(Kondo & Sato, 2007).本实验中有3个物种在土壤表层播种时拥有最高的出苗率, 这与以前类似研究的结果(杨慧玲等, 2007; Guo et al., 2009; Zhu et al., 2014)不同.产生不同结果的可能原因与物种自身繁殖特性及栽培基质有关, 如千叶蓍和加拿大一枝黄花, 其种子质量较小且以风媒传播为主, 这种繁殖特性决定了它们在土壤表层时出苗率最大.再者, 较多前人的研究以沙子作为实验基质, 其浅层的保水性能差于土壤, 从而可能在一定程度上因无法提供连续而充足的水分供应而降低了种子的出苗率.最适宜种子出苗的土埋深度在物种间存在明显差异, 这说明土埋深度在调节种子出苗时机方面具有重要作用.这种意义已经被最新的研究所证实(Ye et al., 2019a).目前对这方面的机理研究依然不足, 这将是未来研究的一个重点. ...
... ; Guo et al., 2009; Zhu et al., 2014)不同.产生不同结果的可能原因与物种自身繁殖特性及栽培基质有关, 如千叶蓍和加拿大一枝黄花, 其种子质量较小且以风媒传播为主, 这种繁殖特性决定了它们在土壤表层时出苗率最大.再者, 较多前人的研究以沙子作为实验基质, 其浅层的保水性能差于土壤, 从而可能在一定程度上因无法提供连续而充足的水分供应而降低了种子的出苗率.最适宜种子出苗的土埋深度在物种间存在明显差异, 这说明土埋深度在调节种子出苗时机方面具有重要作用.这种意义已经被最新的研究所证实(Ye et al., 2019a).目前对这方面的机理研究依然不足, 这将是未来研究的一个重点. ...
5
... 种子出苗是植物生活史中极为重要的一个阶段(Harper & Benton, 1966; Harper, 2010; Ahmed et al., 2015).这个阶段对植物种群数量增长、种间竞争、群落组成、食物链等具有重要的调控作用(Stanton, 1984; Bani-Aameur & Sipple-Michmerhuizen, 2001; Ye et al., 2016).种子出苗代表着由对环境条件耐受性最强阶段(种子休眠)向最脆弱、最敏感阶段的高风险转变(Harper & Benton, 1966; Harper, 2010), 极易受到众多外界环境因素, 如温度、光照、土壤含水量、氧气含量和土壤埋藏深度等(Wu et al., 2015; Amini et al., 2016; Gao et al., 2018; Ye et al., 2019b)的影响.因此, 研究种子出苗过程及其驱动因素具有重要的生态学意义. ...
... ; Harper, 2010), 极易受到众多外界环境因素, 如温度、光照、土壤含水量、氧气含量和土壤埋藏深度等(Wu et al., 2015; Amini et al., 2016; Gao et al., 2018; Ye et al., 2019b)的影响.因此, 研究种子出苗过程及其驱动因素具有重要的生态学意义. ...
... 我们的结果表明, 在物种水平, 种子质量依然是决定种子出苗的一个重要内在因素.这与以前大量的种内研究结果(Winn, 1988; Baskin & Baskin, 1998; Li et al., 2006; Guo et al., 2009; Müller et al., 2018)一致.有利于不同物种种子出苗的适宜土埋深度各不相同, 较浅的土壤埋藏深度对于种子出苗更加有利(Wang et al., 2018).我们发现, 10种植物种子出苗的最适土壤条件都在较浅土层, 特别是在1 cm的土层; 土壤表面撒种或是较深土层播种会在一定程度上降低种子的出苗率.这些结果与Harper和Benton (1966)的研究结果相一致.最适出苗的土埋深度与种子出苗所需的氧气、光照、温度和水分等条件有关(Kondo & Sato, 2007).本实验中有3个物种在土壤表层播种时拥有最高的出苗率, 这与以前类似研究的结果(杨慧玲等, 2007; Guo et al., 2009; Zhu et al., 2014)不同.产生不同结果的可能原因与物种自身繁殖特性及栽培基质有关, 如千叶蓍和加拿大一枝黄花, 其种子质量较小且以风媒传播为主, 这种繁殖特性决定了它们在土壤表层时出苗率最大.再者, 较多前人的研究以沙子作为实验基质, 其浅层的保水性能差于土壤, 从而可能在一定程度上因无法提供连续而充足的水分供应而降低了种子的出苗率.最适宜种子出苗的土埋深度在物种间存在明显差异, 这说明土埋深度在调节种子出苗时机方面具有重要作用.这种意义已经被最新的研究所证实(Ye et al., 2019a).目前对这方面的机理研究依然不足, 这将是未来研究的一个重点. ...
The behaviour of seeds in soil: The germination of seeds on the surface of a water supplying substrate 2 1966
... 种子出苗是植物生活史中极为重要的一个阶段(Harper & Benton, 1966; Harper, 2010; Ahmed et al., 2015).这个阶段对植物种群数量增长、种间竞争、群落组成、食物链等具有重要的调控作用(Stanton, 1984; Bani-Aameur & Sipple-Michmerhuizen, 2001; Ye et al., 2016).种子出苗代表着由对环境条件耐受性最强阶段(种子休眠)向最脆弱、最敏感阶段的高风险转变(Harper & Benton, 1966; Harper, 2010), 极易受到众多外界环境因素, 如温度、光照、土壤含水量、氧气含量和土壤埋藏深度等(Wu et al., 2015; Amini et al., 2016; Gao et al., 2018; Ye et al., 2019b)的影响.因此, 研究种子出苗过程及其驱动因素具有重要的生态学意义. ...
... ).种子出苗代表着由对环境条件耐受性最强阶段(种子休眠)向最脆弱、最敏感阶段的高风险转变(Harper & Benton, 1966; Harper, 2010), 极易受到众多外界环境因素, 如温度、光照、土壤含水量、氧气含量和土壤埋藏深度等(Wu et al., 2015; Amini et al., 2016; Gao et al., 2018; Ye et al., 2019b)的影响.因此, 研究种子出苗过程及其驱动因素具有重要的生态学意义. ...
小叶锦鸡儿种子大小变异对萌发和幼苗生长的影响 1 2008
... 种子出苗受遗传和环境的共同影响(Baskin & Baskin, 1998; Li et al., 2006).种子大小是植物相对稳定的一个内在特征, 它与种子传播、萌发、出土和幼苗竞争等过程密切相关(武高林和杜国祯, 2008).比如, 较大种子总体上比较小种子具有更高的出苗率(Winn, 1988), 幼苗具有更大的竞争力和更高的存活力(Simons & Johnston, 2000; Li et al., 2006).当然也有相反的报道(Dolan, 1984).种子质量是衡量种子大小的重要指标, 种子质量与耐土壤埋藏能力之间存在正相关关系(Chen & Maun, 1999; Chen et al., 2002).较大质量种子往往储存更多的物质和能量, 因而可能从更深的土壤中顺利出苗(聂春雷和郑元润, 2005; Li et al., 2006; Guo et al., 2009; Müller et al., 2018).也有研究发现种子大小与出苗率之间没有直接的相关性(何玉惠等, 2008). ...
小叶锦鸡儿种子大小变异对萌发和幼苗生长的影响 1 2008
... 种子出苗受遗传和环境的共同影响(Baskin & Baskin, 1998; Li et al., 2006).种子大小是植物相对稳定的一个内在特征, 它与种子传播、萌发、出土和幼苗竞争等过程密切相关(武高林和杜国祯, 2008).比如, 较大种子总体上比较小种子具有更高的出苗率(Winn, 1988), 幼苗具有更大的竞争力和更高的存活力(Simons & Johnston, 2000; Li et al., 2006).当然也有相反的报道(Dolan, 1984).种子质量是衡量种子大小的重要指标, 种子质量与耐土壤埋藏能力之间存在正相关关系(Chen & Maun, 1999; Chen et al., 2002).较大质量种子往往储存更多的物质和能量, 因而可能从更深的土壤中顺利出苗(聂春雷和郑元润, 2005; Li et al., 2006; Guo et al., 2009; Müller et al., 2018).也有研究发现种子大小与出苗率之间没有直接的相关性(何玉惠等, 2008). ...
Germination ecology of Emex spinosa and Emex australis, invasive weeds of winter crops. 1 2014
... 种子萌发后能否顺利出土不仅与自身储存的物质和能量有关, 还取决于它在土壤中的位置(Baskin & Baskin, 1998).换言之, 土壤埋藏深度是决定种子命运的关键性因素.一方面, 适当深度的土壤埋藏有助于为种子创造适宜的温度、水分等出苗条件(Baskin & Baskin, 1998; Li et al., 2006; Guo et al., 2009).另一方面, 埋藏过深会对种子出苗产生阻碍或抑制(Baskin & Baskin, 1998; 李柱等, 2004; Li et al., 2006; Guo et al., 2009).种子出苗率随埋藏深度的增加而逐渐减小(Amini et al., 2017), 这与种子出苗前储存物质逐渐耗尽有关(Javaid & Tanveer, 2014; Amini et al., 2016).最适宜种子出苗的埋藏深度往往位于较浅土层, 并随土埋深度的增加而降低(Amini et al., 2017; Wang et al., 2018). ...
Effects of temperature, light, storage conditions, sowing time, and burial depth on the seed germination of Cardiocrinum cordatum var. glehnii(Liliaceae). 1 2007
... 我们的结果表明, 在物种水平, 种子质量依然是决定种子出苗的一个重要内在因素.这与以前大量的种内研究结果(Winn, 1988; Baskin & Baskin, 1998; Li et al., 2006; Guo et al., 2009; Müller et al., 2018)一致.有利于不同物种种子出苗的适宜土埋深度各不相同, 较浅的土壤埋藏深度对于种子出苗更加有利(Wang et al., 2018).我们发现, 10种植物种子出苗的最适土壤条件都在较浅土层, 特别是在1 cm的土层; 土壤表面撒种或是较深土层播种会在一定程度上降低种子的出苗率.这些结果与Harper和Benton (1966)的研究结果相一致.最适出苗的土埋深度与种子出苗所需的氧气、光照、温度和水分等条件有关(Kondo & Sato, 2007).本实验中有3个物种在土壤表层播种时拥有最高的出苗率, 这与以前类似研究的结果(杨慧玲等, 2007; Guo et al., 2009; Zhu et al., 2014)不同.产生不同结果的可能原因与物种自身繁殖特性及栽培基质有关, 如千叶蓍和加拿大一枝黄花, 其种子质量较小且以风媒传播为主, 这种繁殖特性决定了它们在土壤表层时出苗率最大.再者, 较多前人的研究以沙子作为实验基质, 其浅层的保水性能差于土壤, 从而可能在一定程度上因无法提供连续而充足的水分供应而降低了种子的出苗率.最适宜种子出苗的土埋深度在物种间存在明显差异, 这说明土埋深度在调节种子出苗时机方面具有重要作用.这种意义已经被最新的研究所证实(Ye et al., 2019a).目前对这方面的机理研究依然不足, 这将是未来研究的一个重点. ...
Effects of sand burial depth and seed mass on seedling emergence and growth of Nitraria sphaerocarpa. 6 2006
... 种子出苗受遗传和环境的共同影响(Baskin & Baskin, 1998; Li et al., 2006).种子大小是植物相对稳定的一个内在特征, 它与种子传播、萌发、出土和幼苗竞争等过程密切相关(武高林和杜国祯, 2008).比如, 较大种子总体上比较小种子具有更高的出苗率(Winn, 1988), 幼苗具有更大的竞争力和更高的存活力(Simons & Johnston, 2000; Li et al., 2006).当然也有相反的报道(Dolan, 1984).种子质量是衡量种子大小的重要指标, 种子质量与耐土壤埋藏能力之间存在正相关关系(Chen & Maun, 1999; Chen et al., 2002).较大质量种子往往储存更多的物质和能量, 因而可能从更深的土壤中顺利出苗(聂春雷和郑元润, 2005; Li et al., 2006; Guo et al., 2009; Müller et al., 2018).也有研究发现种子大小与出苗率之间没有直接的相关性(何玉惠等, 2008). ...
... ; Li et al., 2006).当然也有相反的报道(Dolan, 1984).种子质量是衡量种子大小的重要指标, 种子质量与耐土壤埋藏能力之间存在正相关关系(Chen & Maun, 1999; Chen et al., 2002).较大质量种子往往储存更多的物质和能量, 因而可能从更深的土壤中顺利出苗(聂春雷和郑元润, 2005; Li et al., 2006; Guo et al., 2009; Müller et al., 2018).也有研究发现种子大小与出苗率之间没有直接的相关性(何玉惠等, 2008). ...
... ; Li et al., 2006; Guo et al., 2009; Müller et al., 2018).也有研究发现种子大小与出苗率之间没有直接的相关性(何玉惠等, 2008). ...
... 种子萌发后能否顺利出土不仅与自身储存的物质和能量有关, 还取决于它在土壤中的位置(Baskin & Baskin, 1998).换言之, 土壤埋藏深度是决定种子命运的关键性因素.一方面, 适当深度的土壤埋藏有助于为种子创造适宜的温度、水分等出苗条件(Baskin & Baskin, 1998; Li et al., 2006; Guo et al., 2009).另一方面, 埋藏过深会对种子出苗产生阻碍或抑制(Baskin & Baskin, 1998; 李柱等, 2004; Li et al., 2006; Guo et al., 2009).种子出苗率随埋藏深度的增加而逐渐减小(Amini et al., 2017), 这与种子出苗前储存物质逐渐耗尽有关(Javaid & Tanveer, 2014; Amini et al., 2016).最适宜种子出苗的埋藏深度往往位于较浅土层, 并随土埋深度的增加而降低(Amini et al., 2017; Wang et al., 2018). ...
... ; Li et al., 2006; Guo et al., 2009).种子出苗率随埋藏深度的增加而逐渐减小(Amini et al., 2017), 这与种子出苗前储存物质逐渐耗尽有关(Javaid & Tanveer, 2014; Amini et al., 2016).最适宜种子出苗的埋藏深度往往位于较浅土层, 并随土埋深度的增加而降低(Amini et al., 2017; Wang et al., 2018). ...
... 我们的结果表明, 在物种水平, 种子质量依然是决定种子出苗的一个重要内在因素.这与以前大量的种内研究结果(Winn, 1988; Baskin & Baskin, 1998; Li et al., 2006; Guo et al., 2009; Müller et al., 2018)一致.有利于不同物种种子出苗的适宜土埋深度各不相同, 较浅的土壤埋藏深度对于种子出苗更加有利(Wang et al., 2018).我们发现, 10种植物种子出苗的最适土壤条件都在较浅土层, 特别是在1 cm的土层; 土壤表面撒种或是较深土层播种会在一定程度上降低种子的出苗率.这些结果与Harper和Benton (1966)的研究结果相一致.最适出苗的土埋深度与种子出苗所需的氧气、光照、温度和水分等条件有关(Kondo & Sato, 2007).本实验中有3个物种在土壤表层播种时拥有最高的出苗率, 这与以前类似研究的结果(杨慧玲等, 2007; Guo et al., 2009; Zhu et al., 2014)不同.产生不同结果的可能原因与物种自身繁殖特性及栽培基质有关, 如千叶蓍和加拿大一枝黄花, 其种子质量较小且以风媒传播为主, 这种繁殖特性决定了它们在土壤表层时出苗率最大.再者, 较多前人的研究以沙子作为实验基质, 其浅层的保水性能差于土壤, 从而可能在一定程度上因无法提供连续而充足的水分供应而降低了种子的出苗率.最适宜种子出苗的土埋深度在物种间存在明显差异, 这说明土埋深度在调节种子出苗时机方面具有重要作用.这种意义已经被最新的研究所证实(Ye et al., 2019a).目前对这方面的机理研究依然不足, 这将是未来研究的一个重点. ...
播种深度对心叶驼绒藜出苗的影响 1 2004
... 种子萌发后能否顺利出土不仅与自身储存的物质和能量有关, 还取决于它在土壤中的位置(Baskin & Baskin, 1998).换言之, 土壤埋藏深度是决定种子命运的关键性因素.一方面, 适当深度的土壤埋藏有助于为种子创造适宜的温度、水分等出苗条件(Baskin & Baskin, 1998; Li et al., 2006; Guo et al., 2009).另一方面, 埋藏过深会对种子出苗产生阻碍或抑制(Baskin & Baskin, 1998; 李柱等, 2004; Li et al., 2006; Guo et al., 2009).种子出苗率随埋藏深度的增加而逐渐减小(Amini et al., 2017), 这与种子出苗前储存物质逐渐耗尽有关(Javaid & Tanveer, 2014; Amini et al., 2016).最适宜种子出苗的埋藏深度往往位于较浅土层, 并随土埋深度的增加而降低(Amini et al., 2017; Wang et al., 2018). ...
播种深度对心叶驼绒藜出苗的影响 1 2004
... 种子萌发后能否顺利出土不仅与自身储存的物质和能量有关, 还取决于它在土壤中的位置(Baskin & Baskin, 1998).换言之, 土壤埋藏深度是决定种子命运的关键性因素.一方面, 适当深度的土壤埋藏有助于为种子创造适宜的温度、水分等出苗条件(Baskin & Baskin, 1998; Li et al., 2006; Guo et al., 2009).另一方面, 埋藏过深会对种子出苗产生阻碍或抑制(Baskin & Baskin, 1998; 李柱等, 2004; Li et al., 2006; Guo et al., 2009).种子出苗率随埋藏深度的增加而逐渐减小(Amini et al., 2017), 这与种子出苗前储存物质逐渐耗尽有关(Javaid & Tanveer, 2014; Amini et al., 2016).最适宜种子出苗的埋藏深度往往位于较浅土层, 并随土埋深度的增加而降低(Amini et al., 2017; Wang et al., 2018). ...
Seed germination ecology of doveweed (Murdannia nudiflora) and its implication for management in dry-seeded rice. 1 2015
... 种子出苗是植物生活史中极为重要的一个阶段(Harper & Benton, 1966; Harper, 2010; Ahmed et al., 2015).这个阶段对植物种群数量增长、种间竞争、群落组成、食物链等具有重要的调控作用(Stanton, 1984; Bani-Aameur & Sipple-Michmerhuizen, 2001; Ye et al., 2016).种子出苗代表着由对环境条件耐受性最强阶段(种子休眠)向最脆弱、最敏感阶段的高风险转变(Harper & Benton, 1966; Harper, 2010), 极易受到众多外界环境因素, 如温度、光照、土壤含水量、氧气含量和土壤埋藏深度等(Wu et al., 2015; Amini et al., 2016; Gao et al., 2018; Ye et al., 2019b)的影响.因此, 研究种子出苗过程及其驱动因素具有重要的生态学意义. ...
Effects of environmental factors and burial depth on seed germination and emergence of two populations of Caucalis platycarpos. 2 2017
... 种子萌发后能否顺利出土不仅与自身储存的物质和能量有关, 还取决于它在土壤中的位置(Baskin & Baskin, 1998).换言之, 土壤埋藏深度是决定种子命运的关键性因素.一方面, 适当深度的土壤埋藏有助于为种子创造适宜的温度、水分等出苗条件(Baskin & Baskin, 1998; Li et al., 2006; Guo et al., 2009).另一方面, 埋藏过深会对种子出苗产生阻碍或抑制(Baskin & Baskin, 1998; 李柱等, 2004; Li et al., 2006; Guo et al., 2009).种子出苗率随埋藏深度的增加而逐渐减小(Amini et al., 2017), 这与种子出苗前储存物质逐渐耗尽有关(Javaid & Tanveer, 2014; Amini et al., 2016).最适宜种子出苗的埋藏深度往往位于较浅土层, 并随土埋深度的增加而降低(Amini et al., 2017; Wang et al., 2018). ...
... ).最适宜种子出苗的埋藏深度往往位于较浅土层, 并随土埋深度的增加而降低(Amini et al., 2017; Wang et al., 2018). ...
Effect of environmental factors on seed germination and emergence of Lepidium vesicarium 3 2016
... 种子出苗是植物生活史中极为重要的一个阶段(Harper & Benton, 1966; Harper, 2010; Ahmed et al., 2015).这个阶段对植物种群数量增长、种间竞争、群落组成、食物链等具有重要的调控作用(Stanton, 1984; Bani-Aameur & Sipple-Michmerhuizen, 2001; Ye et al., 2016).种子出苗代表着由对环境条件耐受性最强阶段(种子休眠)向最脆弱、最敏感阶段的高风险转变(Harper & Benton, 1966; Harper, 2010), 极易受到众多外界环境因素, 如温度、光照、土壤含水量、氧气含量和土壤埋藏深度等(Wu et al., 2015; Amini et al., 2016; Gao et al., 2018; Ye et al., 2019b)的影响.因此, 研究种子出苗过程及其驱动因素具有重要的生态学意义. ...
... 种子萌发后能否顺利出土不仅与自身储存的物质和能量有关, 还取决于它在土壤中的位置(Baskin & Baskin, 1998).换言之, 土壤埋藏深度是决定种子命运的关键性因素.一方面, 适当深度的土壤埋藏有助于为种子创造适宜的温度、水分等出苗条件(Baskin & Baskin, 1998; Li et al., 2006; Guo et al., 2009).另一方面, 埋藏过深会对种子出苗产生阻碍或抑制(Baskin & Baskin, 1998; 李柱等, 2004; Li et al., 2006; Guo et al., 2009).种子出苗率随埋藏深度的增加而逐渐减小(Amini et al., 2017), 这与种子出苗前储存物质逐渐耗尽有关(Javaid & Tanveer, 2014; Amini et al., 2016).最适宜种子出苗的埋藏深度往往位于较浅土层, 并随土埋深度的增加而降低(Amini et al., 2017; Wang et al., 2018). ...
Germination and seedling survival of argan (Argania spinosa) under experimental saline conditions. 1 2001
... 种子出苗是植物生活史中极为重要的一个阶段(Harper & Benton, 1966; Harper, 2010; Ahmed et al., 2015).这个阶段对植物种群数量增长、种间竞争、群落组成、食物链等具有重要的调控作用(Stanton, 1984; Bani-Aameur & Sipple-Michmerhuizen, 2001; Ye et al., 2016).种子出苗代表着由对环境条件耐受性最强阶段(种子休眠)向最脆弱、最敏感阶段的高风险转变(Harper & Benton, 1966; Harper, 2010), 极易受到众多外界环境因素, 如温度、光照、土壤含水量、氧气含量和土壤埋藏深度等(Wu et al., 2015; Amini et al., 2016; Gao et al., 2018; Ye et al., 2019b)的影响.因此, 研究种子出苗过程及其驱动因素具有重要的生态学意义. ...
Impact of seed burial depth on the emergence and growth of annual forage legumes 4 2018
... 种子出苗受遗传和环境的共同影响(Baskin & Baskin, 1998; Li et al., 2006).种子大小是植物相对稳定的一个内在特征, 它与种子传播、萌发、出土和幼苗竞争等过程密切相关(武高林和杜国祯, 2008).比如, 较大种子总体上比较小种子具有更高的出苗率(Winn, 1988), 幼苗具有更大的竞争力和更高的存活力(Simons & Johnston, 2000; Li et al., 2006).当然也有相反的报道(Dolan, 1984).种子质量是衡量种子大小的重要指标, 种子质量与耐土壤埋藏能力之间存在正相关关系(Chen & Maun, 1999; Chen et al., 2002).较大质量种子往往储存更多的物质和能量, 因而可能从更深的土壤中顺利出苗(聂春雷和郑元润, 2005; Li et al., 2006; Guo et al., 2009; Müller et al., 2018).也有研究发现种子大小与出苗率之间没有直接的相关性(何玉惠等, 2008). ...
... 我们的结果表明, 在物种水平, 种子质量依然是决定种子出苗的一个重要内在因素.这与以前大量的种内研究结果(Winn, 1988; Baskin & Baskin, 1998; Li et al., 2006; Guo et al., 2009; Müller et al., 2018)一致.有利于不同物种种子出苗的适宜土埋深度各不相同, 较浅的土壤埋藏深度对于种子出苗更加有利(Wang et al., 2018).我们发现, 10种植物种子出苗的最适土壤条件都在较浅土层, 特别是在1 cm的土层; 土壤表面撒种或是较深土层播种会在一定程度上降低种子的出苗率.这些结果与Harper和Benton (1966)的研究结果相一致.最适出苗的土埋深度与种子出苗所需的氧气、光照、温度和水分等条件有关(Kondo & Sato, 2007).本实验中有3个物种在土壤表层播种时拥有最高的出苗率, 这与以前类似研究的结果(杨慧玲等, 2007; Guo et al., 2009; Zhu et al., 2014)不同.产生不同结果的可能原因与物种自身繁殖特性及栽培基质有关, 如千叶蓍和加拿大一枝黄花, 其种子质量较小且以风媒传播为主, 这种繁殖特性决定了它们在土壤表层时出苗率最大.再者, 较多前人的研究以沙子作为实验基质, 其浅层的保水性能差于土壤, 从而可能在一定程度上因无法提供连续而充足的水分供应而降低了种子的出苗率.最适宜种子出苗的土埋深度在物种间存在明显差异, 这说明土埋深度在调节种子出苗时机方面具有重要作用.这种意义已经被最新的研究所证实(Ye et al., 2019a).目前对这方面的机理研究依然不足, 这将是未来研究的一个重点. ...
... 然而, 许多研究表明植物种子质量与出苗率之间没有相关性(Chen & Maun, 1999; Chen et al., 2002; Paz et al., 2005).这一结论是基于选择同种植物不同个体的种子作为质量梯度得出的, 而不是基于不同物种间种子质量与出苗率的关系, 也没有将两者的关系在不同土壤埋藏深度下进行对比分析(Peng, 2001; Müller et al., 2018).本实验通过探究10种不同植物种子质量与出苗率在5个不同土壤埋藏深度下的相关性, 发现在较深土埋条件下, 单粒种子质量同出苗率间存在正相关关系, 但这种相关关系在较浅土埋条件下不存在.这一发现表明, 土壤埋藏深度强烈影响物种水平的“种子质量-种子出苗关系”.在较深土埋条件下, 具有较大质量种子的物种其出苗率要高于种子质量较小的物种, 这可能与较大种子储存有更多的营养物质及种子自身存在生物量分配的权衡有关(Baskin & Baskin, 1998).在较深埋藏条件下, 种子出苗过程中较大比例的能量储备将被耗尽, 它们对未来生长的初始投资较少, 而将更多的生物量分配给地下部分的生长(Seiwa et al., 2002).较小种子可能由于分配给地下茎生长的能量较少而不能支撑幼苗从较深土层中出苗(Wang et al., 2018).综上所述, 在较浅土层中种子的出苗可能是一个随机过程, 受种子大小等自身特征的影响相对较小; 而在较深土层中种子出苗可能是一个定向过程, 受自身遗传特征的影响相对较大, 其出苗过程在一定程度上受种子大小的调控. ...
鄂尔多斯高原4种主要沙生植物种子萌发与出苗对水分和沙埋的响应 1 2005
... 种子出苗受遗传和环境的共同影响(Baskin & Baskin, 1998; Li et al., 2006).种子大小是植物相对稳定的一个内在特征, 它与种子传播、萌发、出土和幼苗竞争等过程密切相关(武高林和杜国祯, 2008).比如, 较大种子总体上比较小种子具有更高的出苗率(Winn, 1988), 幼苗具有更大的竞争力和更高的存活力(Simons & Johnston, 2000; Li et al., 2006).当然也有相反的报道(Dolan, 1984).种子质量是衡量种子大小的重要指标, 种子质量与耐土壤埋藏能力之间存在正相关关系(Chen & Maun, 1999; Chen et al., 2002).较大质量种子往往储存更多的物质和能量, 因而可能从更深的土壤中顺利出苗(聂春雷和郑元润, 2005; Li et al., 2006; Guo et al., 2009; Müller et al., 2018).也有研究发现种子大小与出苗率之间没有直接的相关性(何玉惠等, 2008). ...
10
... 种子出苗受遗传和环境的共同影响(Baskin & Baskin, 1998; Li et al., 2006).种子大小是植物相对稳定的一个内在特征, 它与种子传播、萌发、出土和幼苗竞争等过程密切相关(武高林和杜国祯, 2008).比如, 较大种子总体上比较小种子具有更高的出苗率(Winn, 1988), 幼苗具有更大的竞争力和更高的存活力(Simons & Johnston, 2000; Li et al., 2006).当然也有相反的报道(Dolan, 1984).种子质量是衡量种子大小的重要指标, 种子质量与耐土壤埋藏能力之间存在正相关关系(Chen & Maun, 1999; Chen et al., 2002).较大质量种子往往储存更多的物质和能量, 因而可能从更深的土壤中顺利出苗(聂春雷和郑元润, 2005; Li et al., 2006; Guo et al., 2009; Müller et al., 2018).也有研究发现种子大小与出苗率之间没有直接的相关性(何玉惠等, 2008). ...
... 种子萌发后能否顺利出土不仅与自身储存的物质和能量有关, 还取决于它在土壤中的位置(Baskin & Baskin, 1998).换言之, 土壤埋藏深度是决定种子命运的关键性因素.一方面, 适当深度的土壤埋藏有助于为种子创造适宜的温度、水分等出苗条件(Baskin & Baskin, 1998; Li et al., 2006; Guo et al., 2009).另一方面, 埋藏过深会对种子出苗产生阻碍或抑制(Baskin & Baskin, 1998; 李柱等, 2004; Li et al., 2006; Guo et al., 2009).种子出苗率随埋藏深度的增加而逐渐减小(Amini et al., 2017), 这与种子出苗前储存物质逐渐耗尽有关(Javaid & Tanveer, 2014; Amini et al., 2016).最适宜种子出苗的埋藏深度往往位于较浅土层, 并随土埋深度的增加而降低(Amini et al., 2017; Wang et al., 2018). ...
... ).换言之, 土壤埋藏深度是决定种子命运的关键性因素.一方面, 适当深度的土壤埋藏有助于为种子创造适宜的温度、水分等出苗条件(Baskin & Baskin, 1998; Li et al., 2006; Guo et al., 2009).另一方面, 埋藏过深会对种子出苗产生阻碍或抑制(Baskin & Baskin, 1998; 李柱等, 2004; Li et al., 2006; Guo et al., 2009).种子出苗率随埋藏深度的增加而逐渐减小(Amini et al., 2017), 这与种子出苗前储存物质逐渐耗尽有关(Javaid & Tanveer, 2014; Amini et al., 2016).最适宜种子出苗的埋藏深度往往位于较浅土层, 并随土埋深度的增加而降低(Amini et al., 2017; Wang et al., 2018). ...
... ).另一方面, 埋藏过深会对种子出苗产生阻碍或抑制(Baskin & Baskin, 1998; 李柱等, 2004; Li et al., 2006; Guo et al., 2009).种子出苗率随埋藏深度的增加而逐渐减小(Amini et al., 2017), 这与种子出苗前储存物质逐渐耗尽有关(Javaid & Tanveer, 2014; Amini et al., 2016).最适宜种子出苗的埋藏深度往往位于较浅土层, 并随土埋深度的增加而降低(Amini et al., 2017; Wang et al., 2018). ...
Effects of burying depth and seed size on seedling establishment of Japanese chestnuts, Castanea crenata. 1 2002
... 然而, 许多研究表明植物种子质量与出苗率之间没有相关性(Chen & Maun, 1999; Chen et al., 2002; Paz et al., 2005).这一结论是基于选择同种植物不同个体的种子作为质量梯度得出的, 而不是基于不同物种间种子质量与出苗率的关系, 也没有将两者的关系在不同土壤埋藏深度下进行对比分析(Peng, 2001; Müller et al., 2018).本实验通过探究10种不同植物种子质量与出苗率在5个不同土壤埋藏深度下的相关性, 发现在较深土埋条件下, 单粒种子质量同出苗率间存在正相关关系, 但这种相关关系在较浅土埋条件下不存在.这一发现表明, 土壤埋藏深度强烈影响物种水平的“种子质量-种子出苗关系”.在较深土埋条件下, 具有较大质量种子的物种其出苗率要高于种子质量较小的物种, 这可能与较大种子储存有更多的营养物质及种子自身存在生物量分配的权衡有关(Baskin & Baskin, 1998).在较深埋藏条件下, 种子出苗过程中较大比例的能量储备将被耗尽, 它们对未来生长的初始投资较少, 而将更多的生物量分配给地下部分的生长(Seiwa et al., 2002).较小种子可能由于分配给地下茎生长的能量较少而不能支撑幼苗从较深土层中出苗(Wang et al., 2018).综上所述, 在较浅土层中种子的出苗可能是一个随机过程, 受种子大小等自身特征的影响相对较小; 而在较深土层中种子出苗可能是一个定向过程, 受自身遗传特征的影响相对较大, 其出苗过程在一定程度上受种子大小的调控. ...
Variation in seed traits of Lobelia inflata(Campanulaceae): Sources and fitness consequences. 1 2000
... 种子出苗受遗传和环境的共同影响(Baskin & Baskin, 1998; Li et al., 2006).种子大小是植物相对稳定的一个内在特征, 它与种子传播、萌发、出土和幼苗竞争等过程密切相关(武高林和杜国祯, 2008).比如, 较大种子总体上比较小种子具有更高的出苗率(Winn, 1988), 幼苗具有更大的竞争力和更高的存活力(Simons & Johnston, 2000; Li et al., 2006).当然也有相反的报道(Dolan, 1984).种子质量是衡量种子大小的重要指标, 种子质量与耐土壤埋藏能力之间存在正相关关系(Chen & Maun, 1999; Chen et al., 2002).较大质量种子往往储存更多的物质和能量, 因而可能从更深的土壤中顺利出苗(聂春雷和郑元润, 2005; Li et al., 2006; Guo et al., 2009; Müller et al., 2018).也有研究发现种子大小与出苗率之间没有直接的相关性(何玉惠等, 2008). ...
Seed variation in wild radish: Effect of seed size on components of seedling an adult fitness 1 1984
... 种子出苗是植物生活史中极为重要的一个阶段(Harper & Benton, 1966; Harper, 2010; Ahmed et al., 2015).这个阶段对植物种群数量增长、种间竞争、群落组成、食物链等具有重要的调控作用(Stanton, 1984; Bani-Aameur & Sipple-Michmerhuizen, 2001; Ye et al., 2016).种子出苗代表着由对环境条件耐受性最强阶段(种子休眠)向最脆弱、最敏感阶段的高风险转变(Harper & Benton, 1966; Harper, 2010), 极易受到众多外界环境因素, 如温度、光照、土壤含水量、氧气含量和土壤埋藏深度等(Wu et al., 2015; Amini et al., 2016; Gao et al., 2018; Ye et al., 2019b)的影响.因此, 研究种子出苗过程及其驱动因素具有重要的生态学意义. ...
Effects of sand burial disturbance on establishment of three desert shrub species in the margin of oasis in northwestern China 3 2018
... 种子萌发后能否顺利出土不仅与自身储存的物质和能量有关, 还取决于它在土壤中的位置(Baskin & Baskin, 1998).换言之, 土壤埋藏深度是决定种子命运的关键性因素.一方面, 适当深度的土壤埋藏有助于为种子创造适宜的温度、水分等出苗条件(Baskin & Baskin, 1998; Li et al., 2006; Guo et al., 2009).另一方面, 埋藏过深会对种子出苗产生阻碍或抑制(Baskin & Baskin, 1998; 李柱等, 2004; Li et al., 2006; Guo et al., 2009).种子出苗率随埋藏深度的增加而逐渐减小(Amini et al., 2017), 这与种子出苗前储存物质逐渐耗尽有关(Javaid & Tanveer, 2014; Amini et al., 2016).最适宜种子出苗的埋藏深度往往位于较浅土层, 并随土埋深度的增加而降低(Amini et al., 2017; Wang et al., 2018). ...
... 我们的结果表明, 在物种水平, 种子质量依然是决定种子出苗的一个重要内在因素.这与以前大量的种内研究结果(Winn, 1988; Baskin & Baskin, 1998; Li et al., 2006; Guo et al., 2009; Müller et al., 2018)一致.有利于不同物种种子出苗的适宜土埋深度各不相同, 较浅的土壤埋藏深度对于种子出苗更加有利(Wang et al., 2018).我们发现, 10种植物种子出苗的最适土壤条件都在较浅土层, 特别是在1 cm的土层; 土壤表面撒种或是较深土层播种会在一定程度上降低种子的出苗率.这些结果与Harper和Benton (1966)的研究结果相一致.最适出苗的土埋深度与种子出苗所需的氧气、光照、温度和水分等条件有关(Kondo & Sato, 2007).本实验中有3个物种在土壤表层播种时拥有最高的出苗率, 这与以前类似研究的结果(杨慧玲等, 2007; Guo et al., 2009; Zhu et al., 2014)不同.产生不同结果的可能原因与物种自身繁殖特性及栽培基质有关, 如千叶蓍和加拿大一枝黄花, 其种子质量较小且以风媒传播为主, 这种繁殖特性决定了它们在土壤表层时出苗率最大.再者, 较多前人的研究以沙子作为实验基质, 其浅层的保水性能差于土壤, 从而可能在一定程度上因无法提供连续而充足的水分供应而降低了种子的出苗率.最适宜种子出苗的土埋深度在物种间存在明显差异, 这说明土埋深度在调节种子出苗时机方面具有重要作用.这种意义已经被最新的研究所证实(Ye et al., 2019a).目前对这方面的机理研究依然不足, 这将是未来研究的一个重点. ...
... 然而, 许多研究表明植物种子质量与出苗率之间没有相关性(Chen & Maun, 1999; Chen et al., 2002; Paz et al., 2005).这一结论是基于选择同种植物不同个体的种子作为质量梯度得出的, 而不是基于不同物种间种子质量与出苗率的关系, 也没有将两者的关系在不同土壤埋藏深度下进行对比分析(Peng, 2001; Müller et al., 2018).本实验通过探究10种不同植物种子质量与出苗率在5个不同土壤埋藏深度下的相关性, 发现在较深土埋条件下, 单粒种子质量同出苗率间存在正相关关系, 但这种相关关系在较浅土埋条件下不存在.这一发现表明, 土壤埋藏深度强烈影响物种水平的“种子质量-种子出苗关系”.在较深土埋条件下, 具有较大质量种子的物种其出苗率要高于种子质量较小的物种, 这可能与较大种子储存有更多的营养物质及种子自身存在生物量分配的权衡有关(Baskin & Baskin, 1998).在较深埋藏条件下, 种子出苗过程中较大比例的能量储备将被耗尽, 它们对未来生长的初始投资较少, 而将更多的生物量分配给地下部分的生长(Seiwa et al., 2002).较小种子可能由于分配给地下茎生长的能量较少而不能支撑幼苗从较深土层中出苗(Wang et al., 2018).综上所述, 在较浅土层中种子的出苗可能是一个随机过程, 受种子大小等自身特征的影响相对较小; 而在较深土层中种子出苗可能是一个定向过程, 受自身遗传特征的影响相对较大, 其出苗过程在一定程度上受种子大小的调控. ...
Ecological and evolutionary consequences of seed size in Prunella vulgaris. 2 1988
... 种子出苗受遗传和环境的共同影响(Baskin & Baskin, 1998; Li et al., 2006).种子大小是植物相对稳定的一个内在特征, 它与种子传播、萌发、出土和幼苗竞争等过程密切相关(武高林和杜国祯, 2008).比如, 较大种子总体上比较小种子具有更高的出苗率(Winn, 1988), 幼苗具有更大的竞争力和更高的存活力(Simons & Johnston, 2000; Li et al., 2006).当然也有相反的报道(Dolan, 1984).种子质量是衡量种子大小的重要指标, 种子质量与耐土壤埋藏能力之间存在正相关关系(Chen & Maun, 1999; Chen et al., 2002).较大质量种子往往储存更多的物质和能量, 因而可能从更深的土壤中顺利出苗(聂春雷和郑元润, 2005; Li et al., 2006; Guo et al., 2009; Müller et al., 2018).也有研究发现种子大小与出苗率之间没有直接的相关性(何玉惠等, 2008). ...
... 我们的结果表明, 在物种水平, 种子质量依然是决定种子出苗的一个重要内在因素.这与以前大量的种内研究结果(Winn, 1988; Baskin & Baskin, 1998; Li et al., 2006; Guo et al., 2009; Müller et al., 2018)一致.有利于不同物种种子出苗的适宜土埋深度各不相同, 较浅的土壤埋藏深度对于种子出苗更加有利(Wang et al., 2018).我们发现, 10种植物种子出苗的最适土壤条件都在较浅土层, 特别是在1 cm的土层; 土壤表面撒种或是较深土层播种会在一定程度上降低种子的出苗率.这些结果与Harper和Benton (1966)的研究结果相一致.最适出苗的土埋深度与种子出苗所需的氧气、光照、温度和水分等条件有关(Kondo & Sato, 2007).本实验中有3个物种在土壤表层播种时拥有最高的出苗率, 这与以前类似研究的结果(杨慧玲等, 2007; Guo et al., 2009; Zhu et al., 2014)不同.产生不同结果的可能原因与物种自身繁殖特性及栽培基质有关, 如千叶蓍和加拿大一枝黄花, 其种子质量较小且以风媒传播为主, 这种繁殖特性决定了它们在土壤表层时出苗率最大.再者, 较多前人的研究以沙子作为实验基质, 其浅层的保水性能差于土壤, 从而可能在一定程度上因无法提供连续而充足的水分供应而降低了种子的出苗率.最适宜种子出苗的土埋深度在物种间存在明显差异, 这说明土埋深度在调节种子出苗时机方面具有重要作用.这种意义已经被最新的研究所证实(Ye et al., 2019a).目前对这方面的机理研究依然不足, 这将是未来研究的一个重点. ...
植物种子大小与幼苗生长策略研究进展 1 2008
... 种子出苗受遗传和环境的共同影响(Baskin & Baskin, 1998; Li et al., 2006).种子大小是植物相对稳定的一个内在特征, 它与种子传播、萌发、出土和幼苗竞争等过程密切相关(武高林和杜国祯, 2008).比如, 较大种子总体上比较小种子具有更高的出苗率(Winn, 1988), 幼苗具有更大的竞争力和更高的存活力(Simons & Johnston, 2000; Li et al., 2006).当然也有相反的报道(Dolan, 1984).种子质量是衡量种子大小的重要指标, 种子质量与耐土壤埋藏能力之间存在正相关关系(Chen & Maun, 1999; Chen et al., 2002).较大质量种子往往储存更多的物质和能量, 因而可能从更深的土壤中顺利出苗(聂春雷和郑元润, 2005; Li et al., 2006; Guo et al., 2009; Müller et al., 2018).也有研究发现种子大小与出苗率之间没有直接的相关性(何玉惠等, 2008). ...
植物种子大小与幼苗生长策略研究进展 1 2008
... 种子出苗受遗传和环境的共同影响(Baskin & Baskin, 1998; Li et al., 2006).种子大小是植物相对稳定的一个内在特征, 它与种子传播、萌发、出土和幼苗竞争等过程密切相关(武高林和杜国祯, 2008).比如, 较大种子总体上比较小种子具有更高的出苗率(Winn, 1988), 幼苗具有更大的竞争力和更高的存活力(Simons & Johnston, 2000; Li et al., 2006).当然也有相反的报道(Dolan, 1984).种子质量是衡量种子大小的重要指标, 种子质量与耐土壤埋藏能力之间存在正相关关系(Chen & Maun, 1999; Chen et al., 2002).较大质量种子往往储存更多的物质和能量, 因而可能从更深的土壤中顺利出苗(聂春雷和郑元润, 2005; Li et al., 2006; Guo et al., 2009; Müller et al., 2018).也有研究发现种子大小与出苗率之间没有直接的相关性(何玉惠等, 2008). ...
Factors affecting seed germination and seedling emergence of Asia minor bluegrass (Polypogon fugax). 1 2015
... 种子出苗是植物生活史中极为重要的一个阶段(Harper & Benton, 1966; Harper, 2010; Ahmed et al., 2015).这个阶段对植物种群数量增长、种间竞争、群落组成、食物链等具有重要的调控作用(Stanton, 1984; Bani-Aameur & Sipple-Michmerhuizen, 2001; Ye et al., 2016).种子出苗代表着由对环境条件耐受性最强阶段(种子休眠)向最脆弱、最敏感阶段的高风险转变(Harper & Benton, 1966; Harper, 2010), 极易受到众多外界环境因素, 如温度、光照、土壤含水量、氧气含量和土壤埋藏深度等(Wu et al., 2015; Amini et al., 2016; Gao et al., 2018; Ye et al., 2019b)的影响.因此, 研究种子出苗过程及其驱动因素具有重要的生态学意义. ...
沙埋对无芒雀麦种子萌发和幼苗生长的影响 1 2007
... 我们的结果表明, 在物种水平, 种子质量依然是决定种子出苗的一个重要内在因素.这与以前大量的种内研究结果(Winn, 1988; Baskin & Baskin, 1998; Li et al., 2006; Guo et al., 2009; Müller et al., 2018)一致.有利于不同物种种子出苗的适宜土埋深度各不相同, 较浅的土壤埋藏深度对于种子出苗更加有利(Wang et al., 2018).我们发现, 10种植物种子出苗的最适土壤条件都在较浅土层, 特别是在1 cm的土层; 土壤表面撒种或是较深土层播种会在一定程度上降低种子的出苗率.这些结果与Harper和Benton (1966)的研究结果相一致.最适出苗的土埋深度与种子出苗所需的氧气、光照、温度和水分等条件有关(Kondo & Sato, 2007).本实验中有3个物种在土壤表层播种时拥有最高的出苗率, 这与以前类似研究的结果(杨慧玲等, 2007; Guo et al., 2009; Zhu et al., 2014)不同.产生不同结果的可能原因与物种自身繁殖特性及栽培基质有关, 如千叶蓍和加拿大一枝黄花, 其种子质量较小且以风媒传播为主, 这种繁殖特性决定了它们在土壤表层时出苗率最大.再者, 较多前人的研究以沙子作为实验基质, 其浅层的保水性能差于土壤, 从而可能在一定程度上因无法提供连续而充足的水分供应而降低了种子的出苗率.最适宜种子出苗的土埋深度在物种间存在明显差异, 这说明土埋深度在调节种子出苗时机方面具有重要作用.这种意义已经被最新的研究所证实(Ye et al., 2019a).目前对这方面的机理研究依然不足, 这将是未来研究的一个重点. ...
沙埋对无芒雀麦种子萌发和幼苗生长的影响 1 2007
... 我们的结果表明, 在物种水平, 种子质量依然是决定种子出苗的一个重要内在因素.这与以前大量的种内研究结果(Winn, 1988; Baskin & Baskin, 1998; Li et al., 2006; Guo et al., 2009; Müller et al., 2018)一致.有利于不同物种种子出苗的适宜土埋深度各不相同, 较浅的土壤埋藏深度对于种子出苗更加有利(Wang et al., 2018).我们发现, 10种植物种子出苗的最适土壤条件都在较浅土层, 特别是在1 cm的土层; 土壤表面撒种或是较深土层播种会在一定程度上降低种子的出苗率.这些结果与Harper和Benton (1966)的研究结果相一致.最适出苗的土埋深度与种子出苗所需的氧气、光照、温度和水分等条件有关(Kondo & Sato, 2007).本实验中有3个物种在土壤表层播种时拥有最高的出苗率, 这与以前类似研究的结果(杨慧玲等, 2007; Guo et al., 2009; Zhu et al., 2014)不同.产生不同结果的可能原因与物种自身繁殖特性及栽培基质有关, 如千叶蓍和加拿大一枝黄花, 其种子质量较小且以风媒传播为主, 这种繁殖特性决定了它们在土壤表层时出苗率最大.再者, 较多前人的研究以沙子作为实验基质, 其浅层的保水性能差于土壤, 从而可能在一定程度上因无法提供连续而充足的水分供应而降低了种子的出苗率.最适宜种子出苗的土埋深度在物种间存在明显差异, 这说明土埋深度在调节种子出苗时机方面具有重要作用.这种意义已经被最新的研究所证实(Ye et al., 2019a).目前对这方面的机理研究依然不足, 这将是未来研究的一个重点. ...
a). Sand burial helps regulate timing of seed germination of a dominant herb in an inland dune ecosystem with a semiarid temperate climate 1 2019
... 我们的结果表明, 在物种水平, 种子质量依然是决定种子出苗的一个重要内在因素.这与以前大量的种内研究结果(Winn, 1988; Baskin & Baskin, 1998; Li et al., 2006; Guo et al., 2009; Müller et al., 2018)一致.有利于不同物种种子出苗的适宜土埋深度各不相同, 较浅的土壤埋藏深度对于种子出苗更加有利(Wang et al., 2018).我们发现, 10种植物种子出苗的最适土壤条件都在较浅土层, 特别是在1 cm的土层; 土壤表面撒种或是较深土层播种会在一定程度上降低种子的出苗率.这些结果与Harper和Benton (1966)的研究结果相一致.最适出苗的土埋深度与种子出苗所需的氧气、光照、温度和水分等条件有关(Kondo & Sato, 2007).本实验中有3个物种在土壤表层播种时拥有最高的出苗率, 这与以前类似研究的结果(杨慧玲等, 2007; Guo et al., 2009; Zhu et al., 2014)不同.产生不同结果的可能原因与物种自身繁殖特性及栽培基质有关, 如千叶蓍和加拿大一枝黄花, 其种子质量较小且以风媒传播为主, 这种繁殖特性决定了它们在土壤表层时出苗率最大.再者, 较多前人的研究以沙子作为实验基质, 其浅层的保水性能差于土壤, 从而可能在一定程度上因无法提供连续而充足的水分供应而降低了种子的出苗率.最适宜种子出苗的土埋深度在物种间存在明显差异, 这说明土埋深度在调节种子出苗时机方面具有重要作用.这种意义已经被最新的研究所证实(Ye et al., 2019a).目前对这方面的机理研究依然不足, 这将是未来研究的一个重点. ...
Differential plant species responses to interactions of sand burial, precipitation enhancement and climatic variation promote co-existence in Chinese steppe vegetation 1 2016
... 种子出苗是植物生活史中极为重要的一个阶段(Harper & Benton, 1966; Harper, 2010; Ahmed et al., 2015).这个阶段对植物种群数量增长、种间竞争、群落组成、食物链等具有重要的调控作用(Stanton, 1984; Bani-Aameur & Sipple-Michmerhuizen, 2001; Ye et al., 2016).种子出苗代表着由对环境条件耐受性最强阶段(种子休眠)向最脆弱、最敏感阶段的高风险转变(Harper & Benton, 1966; Harper, 2010), 极易受到众多外界环境因素, 如温度、光照、土壤含水量、氧气含量和土壤埋藏深度等(Wu et al., 2015; Amini et al., 2016; Gao et al., 2018; Ye et al., 2019b)的影响.因此, 研究种子出苗过程及其驱动因素具有重要的生态学意义. ...
Experimental sand burial and precipitation enhancement alter plant and soil carbon allocation in a semi-arid steppe in north China 1 2019
... 种子出苗是植物生活史中极为重要的一个阶段(Harper & Benton, 1966; Harper, 2010; Ahmed et al., 2015).这个阶段对植物种群数量增长、种间竞争、群落组成、食物链等具有重要的调控作用(Stanton, 1984; Bani-Aameur & Sipple-Michmerhuizen, 2001; Ye et al., 2016).种子出苗代表着由对环境条件耐受性最强阶段(种子休眠)向最脆弱、最敏感阶段的高风险转变(Harper & Benton, 1966; Harper, 2010), 极易受到众多外界环境因素, 如温度、光照、土壤含水量、氧气含量和土壤埋藏深度等(Wu et al., 2015; Amini et al., 2016; Gao et al., 2018; Ye et al., 2019b)的影响.因此, 研究种子出苗过程及其驱动因素具有重要的生态学意义. ...
Effects of amount and frequency of precipitation and sand burial on seed germination, seedling emergence and survival of the dune grass Leymus secalinus in semiarid China. 1 2014
... 我们的结果表明, 在物种水平, 种子质量依然是决定种子出苗的一个重要内在因素.这与以前大量的种内研究结果(Winn, 1988; Baskin & Baskin, 1998; Li et al., 2006; Guo et al., 2009; Müller et al., 2018)一致.有利于不同物种种子出苗的适宜土埋深度各不相同, 较浅的土壤埋藏深度对于种子出苗更加有利(Wang et al., 2018).我们发现, 10种植物种子出苗的最适土壤条件都在较浅土层, 特别是在1 cm的土层; 土壤表面撒种或是较深土层播种会在一定程度上降低种子的出苗率.这些结果与Harper和Benton (1966)的研究结果相一致.最适出苗的土埋深度与种子出苗所需的氧气、光照、温度和水分等条件有关(Kondo & Sato, 2007).本实验中有3个物种在土壤表层播种时拥有最高的出苗率, 这与以前类似研究的结果(杨慧玲等, 2007; Guo et al., 2009; Zhu et al., 2014)不同.产生不同结果的可能原因与物种自身繁殖特性及栽培基质有关, 如千叶蓍和加拿大一枝黄花, 其种子质量较小且以风媒传播为主, 这种繁殖特性决定了它们在土壤表层时出苗率最大.再者, 较多前人的研究以沙子作为实验基质, 其浅层的保水性能差于土壤, 从而可能在一定程度上因无法提供连续而充足的水分供应而降低了种子的出苗率.最适宜种子出苗的土埋深度在物种间存在明显差异, 这说明土埋深度在调节种子出苗时机方面具有重要作用.这种意义已经被最新的研究所证实(Ye et al., 2019a).目前对这方面的机理研究依然不足, 这将是未来研究的一个重点. ...
The effect of seed size and maternal source on individual size in a population of Ludwigia leptocarpa(Onagraceae). 1 1984
... 种子出苗受遗传和环境的共同影响(Baskin & Baskin, 1998; Li et al., 2006).种子大小是植物相对稳定的一个内在特征, 它与种子传播、萌发、出土和幼苗竞争等过程密切相关(武高林和杜国祯, 2008).比如, 较大种子总体上比较小种子具有更高的出苗率(Winn, 1988), 幼苗具有更大的竞争力和更高的存活力(Simons & Johnston, 2000; Li et al., 2006).当然也有相反的报道(Dolan, 1984).种子质量是衡量种子大小的重要指标, 种子质量与耐土壤埋藏能力之间存在正相关关系(Chen & Maun, 1999; Chen et al., 2002).较大质量种子往往储存更多的物质和能量, 因而可能从更深的土壤中顺利出苗(聂春雷和郑元润, 2005; Li et al., 2006; Guo et al., 2009; Müller et al., 2018).也有研究发现种子大小与出苗率之间没有直接的相关性(何玉惠等, 2008). ...
 ,*中国科学院大学资源与环境学院, 北京 101408
,*中国科学院大学资源与环境学院, 北京 101408
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT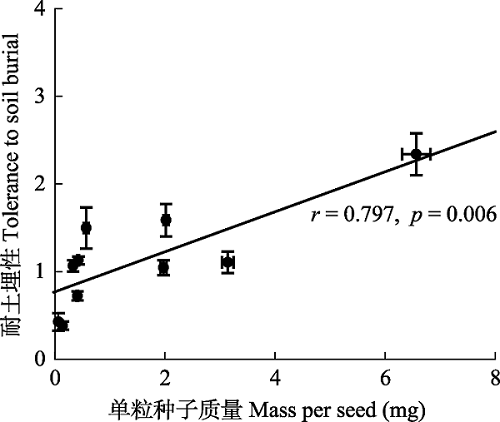 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}