Season dynamics of carbon use efficiency and its influencing factors in the old-growth Abies fabri forest in Gongga Mountain, western Sichuan, China
Shu-Miao SHU,1,2, Wan-Ze ZHU,1,*, Fei RAN1, Shou-Qin SUN1, Yuan-Yuan ZHANG1,21Institute of Mountain Hazards and Environment, Chinese Academy of Sciences, Chengdu 610041, China 2University of Chinese Academy of Sciences, Beijing 100049, China
National Key R&D Program of China(2017YFC0505004) Major Scientific and Technological Projects of Environmental Governance and Ecological Protection in Sichuan Province(2018SZDZX0031)
Abstract Aims Carbon use efficiency (CUE), an important function parameter, reflects the carbon sequestration capacity of forest ecosystems. It is useful in analyzing the temporal dynamics of carbon budgets at the organ, individual and community scales. It can help to determine and predict the carbon sink/source of terrestrial ecosystems, which is a matter of widespread concern. Methods Using the biometric method, we measured and calculated the respiration and net productivity dynamics of different fir organs from an old-growth Abies fabri forest on Gongga Mountain in the eastern Qinghai-Xizang Plateau, China. We studied the CUE dynamics of the tree layer and its organs and analyzed their influencing factors. We also estimated the CUE of whole trees of different diameters at breast-height (DBH) classes. Important findings (1) Monthly respiration rates in both the tree layer and its organs are positively related to temperature, and fine roots have the highest respiration rate of all. There is no significant difference in the annual respiration of whole trees with different DBH classes, and the small DBH trees (30-40 cm) have the minimum annual stem respiration. (2) The monthly net primary productivity (NPP) of the fine root and whole stem in the tree layer increases with temperature, with the fine root accounting for the largest proportion. The small trees have the greatest annual NPP, and their needle NPP is also significantly higher than that of the medium DBH (50- 60 cm) and large DBH (75-90 cm) trees. (3) The CUE of the tree layer and its organs are mostly among 0.30 to 0.60. The monthly changes in the CUE of both fine roots and stems are similar, and their CUE increases with temperature. The CUE of trees and their organs all decrease significantly with tree growth. (4) The CUE of both the stems and fine roots is positively related with the air and soil temperature, while precipitaion has a positive effect on needle CUE. Fine root CUE has negative and positive effects on stem CUE and needle CUE, respectively. The tree layer CUE depends mainly on stem and fine root CUE. These results indicate that old-growth forests have strong and sustainable carbon sink functions, and play an important role in regional carbon storage and the carbon cycle of the forest ecosystem. Keywords:carbon use efficiency;net primary productivity;respiration carbon consumption;old Abies fabri forest;Gongga Mountain
PDF (1404KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 舒树淼, 朱万泽, 冉飞, 孙守琴, 张元媛. 贡嘎山峨眉冷杉成熟林碳利用效率季节动态及其影响因子. 植物生态学报, 2020, 44(11): 1127-1137. DOI: 10.17521/cjpe.2019.0289 SHU Shu-Miao, ZHU Wan-Ze, RAN Fei, SUN Shou-Qin, ZHANG Yuan-Yuan. Season dynamics of carbon use efficiency and its influencing factors in the old-growth Abies fabri forest in Gongga Mountain, western Sichuan, China. Chinese Journal of Plant Ecology, 2020, 44(11): 1127-1137. DOI: 10.17521/cjpe.2019.0289
森林在碳汇中发挥着不可替代的作用, 占陆地面积27.51%的森林年固碳量约为整个陆地圈的2/3 (Dixon et al., 1994; 项文化等, 2003)。森林固碳强度及动态是预测森林固碳潜力的重要内容。植被生长是森林固碳的主要原因, 净初级生产力(NPP)是反映森林群落或生态系统固碳强度的直观指标(Zhou et al., 2010; Williams et al., 2014)。然而单纯的NPP不足以反映群落或生态系统的固碳能力, 这是因为植被生长依赖于构建成本, 存在一定的呼吸消耗(Gifford, 2003)。例如, 为了应对环境变化, 植被会调整其生长策略(Jar?u?ka & Barna, 2011), 在胁迫环境下, 植被可能将更多的碳用于维护与防御, 导致呼吸速率提高。尽管植被NPP可能会保持不变, 但投入到生长的资源占比却明显减少, 意味着植被固碳能力的降低。被定义为NPP/GPP或1 - Ra/GPP (即净生产力与总初级生产力(GPP)之比, Ra为自养呼吸速率)的碳利用效率(CUE)可以直接反映植被的固碳能力。不仅如此, 该指标还适用于描述和比较任意时间段内不同器官、植物个体和群落的固碳规律(朱万泽, 2013)。CUE的引入为了解植被碳源/汇功能的变化提供了一个新的视角和切入点。
Fig. 1Research route of seasonal dynamics of carbon use efficiency (CUE) in an old-growth Abies fabri forest on Gongga Mountain. NPP, net primary productivity.
Fig. 2Respiration of the tree layer and trees with different diameter classes in an old-growth Abies fabri forest on Gongga Mountain. Different lowercase letters indicate significant difference at p < 0.05 level for the respiration amount of same organ under different diameter classes.
Fig. 3Net primary productivity of the tree layer and trees with different diameter classes in an old-growth Abies fabri forest on Gongga Mountain. Different lowercase letters indicate significant difference at p < 0.05 level for the net primary productivity of same organ under different diameter classes.
Fig. 4Carbon use efficiency (CUE) of the tree layer and trees with different diameter classes in an old-growth Abies fabri forest on Gongga Mountain. Different lowercase letters indicate significant difference at p < 0.05 level for the CUE of same organ under different diameter classes.
Fig. 5Influencing factors of carbon use efficiency (CUE) in tree layer and organs of an old-growth Abies fabri forest. Values associated with solid arrows are standardized path coefficients, indicating positive or negative effects. Values associated with the rectangles are R2, indicating the proportion of variation explained by relationships with other variables. Small circles represent residuals. Values associated with two way arrow indicate the correlation. The dotted line indicates the preset path.
Table 1 表1 表1峨眉冷杉成熟林乔木层碳利用效率(CUE)结构方程模型标准化影响系数 Table 1Standardized influence coefficients of the structural equation model (SEM) for the arbor layer carbon use efficiency (CUE) of Abies fabri old-growth forest
影响因素 Impact factor
标准化总影响系数 Standardized total influence coefficients
标准化直接影响系数 Standardized direct influence coefficients
AnX, ChenYM, TangYK (2017). Factors affecting the spatial variation of carbon use efficiency and carbon fluxes in east Asian forest and grassland Research of Soil and Water Conservation, 24(5), 79-87. [本文引用: 1]
AshrafM, AhmadA, McNeillyT (2001). Growth and photosynthetic characteristics in pearl millet under water stress and different potassium supply Photosynthetica, 39, 389-394. [本文引用: 1]
ChambersJQ, TribuzyES, ToledoLC, CrispimBF, HiguchiN, dos SantosJ, AraújoAC, KruijtB, NobreAD, TrumboreSE (2004). Respiration from a tropical forest ecosystem: partitioning of sources and low carbon use efficiency Ecological Applications, 14, 72-88. [本文引用: 3]
ChantumaP, LacointeA, KasemsapP, ThanisawanyangkuraS, GohetE, ClémentA, GuilliotA, AméglioT, ThalerP (2009). Carbohydrate storage in wood and bark of rubber trees submitted to different level of C demand induced by latex tapping Tree Physiology, 29, 1021-1031. DOI:10.1093/treephys/tpp043URLPMID:19556234 [本文引用: 1] When the current level of carbohydrates produced by photosynthesis is not enough to meet the C demand for maintenance, growth or metabolism, trees use stored carbohydrates. In rubber trees (Hevea brasiliensis Muell. Arg.), however, a previous study (Silpi U., A. Lacointe, P. Kasemsap, S. Thanisawanyangkura, P. Chantuma, E. Gohet, N. Musigamart, A. Clement, T. Ameglio and P. Thaler. 2007. Carbohydrate reserves as a competing sink: evidence from tapping the rubber tree. Tree Physiol. 27:881-889) showed that the additional sink created by latex tapping results not in a decrease, but in an increase in the non-structural carbohydrate (NSC) storage in trunk wood. In this study, the response of NSC storage to latex tapping was further investigated to better understand the trade-off between latex regeneration, biomass and storage. Three tapping systems were compared to the untapped Control for 2 years. Soluble sugars and starch were analyzed in bark and wood on both sides of the trunk, from 50 to 200 cm from the ground. The results confirmed over the 2 years that tapped trees stored more NSC, mainly starch, than untapped Control. Moreover, a double cut alternative tapping system, which produced a higher latex yield than conventional systems, led to even higher NSC concentrations. In all tapped trees, the increase in storage occurred together with a reduction in trunk radial growth. This was interpreted as a shift in carbon allocation toward the creation of reserves, at the expense of growth, to cover the increased risk induced by tapping (repeated wounding and loss of C in latex). Starch was lower in bark than in wood, whereas it was the contrary for soluble sugars. The resulting NSC was twice as low and less variable in bark than in wood. Although latex regeneration occurs in the bark, changes related to latex tapping were more marked in wood than in bark. From seasonal dynamics and differences between the two sides of the trunk in response to tapping, we concluded that starch in wood behaved as the long-term reserve compartment at the whole trunk level, whereas starch in bark was a local buffer. Soluble sugars behaved like an intermediate, ready-to-use compartment in both wood and bark. Finally, the dynamics of carbohydrate reserves appears a relevant parameter to assess the long-term performance of latex tapping systems.
ChenGS, YangYS, GuoJF, XieJS, YangZJ (2011). Relationships between carbon allocation and partitioning of soil respiration across world mature forests Plant Ecology, 212, 195-206. [本文引用: 1]
ChenGS, HobbieSE, ReichPB, YangYS, RobinsonD (2019). Allometry of fine roots in forest ecosystems Ecology Letters, 22, 322-331. URLPMID:30488519 [本文引用: 2]
de LuciaEH, DrakeJE, ThomasRB, Gonzalez-MelerM (2007). Forest carbon use efficiency: Is respiration a constant fraction of gross primary production? Global Change Biology, 13, 1157-1167. [本文引用: 1]
DixonRK, SolomonAM, BrownS, HoughtonRA, TrexierMC, WisniewskiJ (1994). Carbon pools and flux of global forest ecosystems Science, 263, 185-190. URLPMID:17839174 [本文引用: 1]
DuZ, GanSS, HuJ (2014). Characteristics and protection strategy of forest resources in the alpine region of southwest China Forest Resources Management, (z1), 27-31. [本文引用: 1]
MalhiY, Arag?oL, MetcalfeD, PAIVAR, QuesadaC, AlmeidaS, AndersonL, BrandoP, ChambersJ, CostaA, HutyraL, SouzaP, Pati?oS, PyleE, RobertsonA, TeixeiraL (2009). Comprehensive assessment of carbon productivity, allocation and storage in three Amazonian forests Global Change Biology, 15, 1255-1274. [本文引用: 1]
MencucciniM, Martínez-VilaltaJ, VanderkleinD, HamidH, A, KorakakiE, LeeS, MichielsB (2005). Size-mediated ageing reduces vigour in trees Ecology Letters, 11, 1183-1190. [本文引用: 1]
GaoSP, LiJX, XuMC, ChenX, DaiJ (2007). Leaf N and P stoichiometry of common species in successional stages of the evergreen broad-leaved forest in Tiantong National Forest Park, Zhejiang Province, China Acta Ecologica Sinica, 27, 947-952. [本文引用: 1]
GiffordRM (2003). Plant respiration in productivity models: conceptualisation, representation and issues for global terrestrial carbon-cycle research Functional Plant Biology, 30, 171-186. URLPMID:32689003 [本文引用: 2]
HuZY, WangGX, SunXY, WangJ, ChenXP, SongCL, SongXY, LinS (2019). Variations in belowground carbon use strategies under different climatic conditions Agricultural and Forest Meteorology, 268, 32-39. [本文引用: 1]
Jar?u?kaB, BarnaM (2011). Plasticity in above-ground biomass allocation in Fagus sylvatica L. saplings in response to light availability Annals of Forest Research, 54, 151-160. [本文引用: 1]
KalynAL, van ReesKCJ (2006). Contribution of fine roots to ecosystem biomass and net primary production in black spruce, aspen, and jack pine forests in Saskatchewan Agricultural and Forest Meteorology, 140, 236-243. [本文引用: 1]
LiuT, SunSQ, QiuY (2017). Dynamics and differences in the decomposition of litters from three dominating plants in subalpine ecosystems in Western Sichuan China Mountain Research, 35, 663-668. [本文引用: 1]
LiuZG, QiuFY (1986). The main vegetation types and their distribution in the Gongga Mountainous region Acta Phytoecologica et Geobotanica Sinica, (1), 28-36. [本文引用: 1]
LuyssaertS, SchulzeED, B?rnerA, KnohlA, Hessenm?llerD, LawBE, CiaisP, GraceJ (2008). Old-growth forests as global carbon sinks Nature, 455, 213-215. DOI:10.1038/nature07276URLPMID:18784722 [本文引用: 1] Old-growth forests remove carbon dioxide from the atmosphere at rates that vary with climate and nitrogen deposition. The sequestered carbon dioxide is stored in live woody tissues and slowly decomposing organic matter in litter and soil. Old-growth forests therefore serve as a global carbon dioxide sink, but they are not protected by international treaties, because it is generally thought that ageing forests cease to accumulate carbon. Here we report a search of literature and databases for forest carbon-flux estimates. We find that in forests between 15 and 800 years of age, net ecosystem productivity (the net carbon balance of the forest including soils) is usually positive. Our results demonstrate that old-growth forests can continue to accumulate carbon, contrary to the long-standing view that they are carbon neutral. Over 30 per cent of the global forest area is unmanaged primary forest, and this area contains the remaining old-growth forests. Half of the primary forests (6 x 10(8) hectares) are located in the boreal and temperate regions of the Northern Hemisphere. On the basis of our analysis, these forests alone sequester about 1.3 +/- 0.5 gigatonnes of carbon per year. Thus, our findings suggest that 15 per cent of the global forest area, which is currently not considered when offsetting increasing atmospheric carbon dioxide concentrations, provides at least 10 per cent of the global net ecosystem productivity. Old-growth forests accumulate carbon for centuries and contain large quantities of it. We expect, however, that much of this carbon, even soil carbon, will move back to the atmosphere if these forests are disturbed.
M?kel?A, ValentineHT (2001). The ratio of NPP to GPP: evidence of change over the course of stand development Tree Physiology, 21, 1015-1030. URLPMID:11560815 [本文引用: 1]
MalhiY (2012). The productivity, metabolism and carbon cycle of tropical forest vegetation Journal of Ecology, 100, 65-75. [本文引用: 1]
MalhiY, DoughtyC, GalbraithD (2011). The allocation of ecosystem net primary productivity in tropical forests Philosophical Transactions of the Royal Society B, 366, 3225-3245. [本文引用: 1]
MetcalfeDB, MeirP, Arag?oLEOC, Lobo-do-ValeR, GalbraithD, FisherRA, ChavesMM, MarocoJP, da CostaACL, de AlmeidaSS, BragaAP, Gon?alvesPHL, de AthaydesJ, da CostaM, PortelaTTB, de OliveiraAAR, MalhiY, WilliamsM (2010). Shifts in plant respiration and carbon use efficiency at a large-scale drought experiment in the eastern Amazon New Phytologist, 187, 608-621. [本文引用: 1]
MolesAT, WartonDI, WarmanL, SwensonNG, LaffanSW, ZanneAE, PitmanA, HemmingsFA, LeishmanMR (2009). Global patterns in plant height Journal of Ecology, 97, 923-932. [本文引用: 1]
MollierA, PellerinS (1999). Maize root system growth and development as influenced by phosphorous deficiency Journal of Experimental Botany, 50, 487-497. [本文引用: 1]
PengL, PengJH, SunSQ, WuYH, HeG (2015). Characteristics of mountain ecosystem soil respiration along an elevation gradient on Gongga Mountain Journal of Mountain Science, 33, 696-702. [本文引用: 1]
RaichJW, NadelhofferKJ (1989). Belowground carbon allocation in forest ecosystems: global trends Ecology, 70, 1346-1354. [本文引用: 1]
ShuSM, ZhuWZ, WangWZ, JiaM, ZhangYY, ShengZL (2019). Effects of tree size heterogeneity on carbon sink in old forests Forest Ecology and Management, 432, 637-648. [本文引用: 5]
WaringRH, LandsbergJJ, WilliamsM (1998). Net primary production of forests: a constant fraction of gross primary production? Tree Physiology, 18, 129-134. URLPMID:12651397 [本文引用: 1]
WilliamsCA, CollatzGJ, MasekJ, HuangC, GowardSN (2014). Impacts of disturbance history on forest carbon stocks and fluxes: merging satellite disturbance mapping with forest inventory data in a carbon cycle model framework Remote Sensing of Environment, 151, 57-71. [本文引用: 1]
WuML (2010). Structural Equation Model: Operation and Application of AMOS. Chongqing University Press, Chongqing. [本文引用: 1]
XiangWH, TianDL, YanWD (2003). Review of researches on forest biomass and productivity Central South Forest Inventory and Planning, 22(3), 57-64. [本文引用: 1]
ZhangFY, QuanQ, MaFF, TianDS, ZhouQP, NiuSL (2019). Differential responses of ecosystem carbon flux components to experimental precipitation gradient in an alpine meadow Functional Ecology, 33, 889-900. [本文引用: 2]
ZhaoG (2015). The Breathing Dynamics of the Emei Fir Trunks of Gongga Mountain and Its Influence Factors. Master degree dissertation, University of the Chinese Academy of Sciences, Beijing. [本文引用: 2]
ZhouT, ShiPJ, JiaGS, LiXJ, LuoYQ (2010). Spatial patterns of ecosystem carbon residence time in Chinese forests Science China Earth Sciences, 53, 1229-1240. [本文引用: 1]
ZhuWZ (2013). Advances in the carbon use efficiency of forest Chinese Journal of Plant Ecology, 37, 1043-1058. [本文引用: 3]
... 近年来有关CUE的研究主要集中在群落和生态系统尺度, 包括森林与草地植被生态系统(安相等, 2017; Shu et al., 2019; Zhang et al., 2019).这些研究揭示了CUE可能是由植被类型、环境条件和不同生长阶段共同决定的(朱万泽, 2013; Shu et al., 2019; Zhang et al., 2019).不仅如此, 森林发育过程中的树木大小分布也在很大程度上直接决定了群落和生态系统CUE的动态(Shu et al., 2019).在较长的时间尺度上, 群落CUE随树木个体的生长逐渐降低, 之后由于外界干扰的影响, 群落CUE可能会由于内部物种稳定的大小结构趋近于某一定值.这些结果既支持了群落CUE动态变化假说(Gifford, 2003; de Lucia et al., 2007), 也印证了CUE变化的最终稳定性观点, 证实了成熟森林可能普遍为碳汇的结论(Luyssaert et al., 2008; 朱万泽, 2020).然而, 森林生态系统不同组分CUE及其变化规律依然少有关注.不同组分如叶片、树干和树根间CUE有何联系? 它们对林分CUE的贡献如何等都值得进一步研究.这些问题的理解, 不仅有利于认识群落和生态系统的碳分配规律, 还可深入了解树木的生活史策略, 对理解气候变化下森林生态系统固碳能力的响应与适应亦有重要意义. ...
东亚森林、草地碳利用效率及碳通量空间变化的影响因素分析 1 2017
... 近年来有关CUE的研究主要集中在群落和生态系统尺度, 包括森林与草地植被生态系统(安相等, 2017; Shu et al., 2019; Zhang et al., 2019).这些研究揭示了CUE可能是由植被类型、环境条件和不同生长阶段共同决定的(朱万泽, 2013; Shu et al., 2019; Zhang et al., 2019).不仅如此, 森林发育过程中的树木大小分布也在很大程度上直接决定了群落和生态系统CUE的动态(Shu et al., 2019).在较长的时间尺度上, 群落CUE随树木个体的生长逐渐降低, 之后由于外界干扰的影响, 群落CUE可能会由于内部物种稳定的大小结构趋近于某一定值.这些结果既支持了群落CUE动态变化假说(Gifford, 2003; de Lucia et al., 2007), 也印证了CUE变化的最终稳定性观点, 证实了成熟森林可能普遍为碳汇的结论(Luyssaert et al., 2008; 朱万泽, 2020).然而, 森林生态系统不同组分CUE及其变化规律依然少有关注.不同组分如叶片、树干和树根间CUE有何联系? 它们对林分CUE的贡献如何等都值得进一步研究.这些问题的理解, 不仅有利于认识群落和生态系统的碳分配规律, 还可深入了解树木的生活史策略, 对理解气候变化下森林生态系统固碳能力的响应与适应亦有重要意义. ...
Growth and photosynthetic characteristics in pearl millet under water stress and different potassium supply 1 2001
Forest carbon use efficiency: Is respiration a constant fraction of gross primary production? 1 2007
... 近年来有关CUE的研究主要集中在群落和生态系统尺度, 包括森林与草地植被生态系统(安相等, 2017; Shu et al., 2019; Zhang et al., 2019).这些研究揭示了CUE可能是由植被类型、环境条件和不同生长阶段共同决定的(朱万泽, 2013; Shu et al., 2019; Zhang et al., 2019).不仅如此, 森林发育过程中的树木大小分布也在很大程度上直接决定了群落和生态系统CUE的动态(Shu et al., 2019).在较长的时间尺度上, 群落CUE随树木个体的生长逐渐降低, 之后由于外界干扰的影响, 群落CUE可能会由于内部物种稳定的大小结构趋近于某一定值.这些结果既支持了群落CUE动态变化假说(Gifford, 2003; de Lucia et al., 2007), 也印证了CUE变化的最终稳定性观点, 证实了成熟森林可能普遍为碳汇的结论(Luyssaert et al., 2008; 朱万泽, 2020).然而, 森林生态系统不同组分CUE及其变化规律依然少有关注.不同组分如叶片、树干和树根间CUE有何联系? 它们对林分CUE的贡献如何等都值得进一步研究.这些问题的理解, 不仅有利于认识群落和生态系统的碳分配规律, 还可深入了解树木的生活史策略, 对理解气候变化下森林生态系统固碳能力的响应与适应亦有重要意义. ...
Carbon pools and flux of global forest ecosystems 1 1994
... 森林在碳汇中发挥着不可替代的作用, 占陆地面积27.51%的森林年固碳量约为整个陆地圈的2/3 (Dixon et al., 1994; 项文化等, 2003).森林固碳强度及动态是预测森林固碳潜力的重要内容.植被生长是森林固碳的主要原因, 净初级生产力(NPP)是反映森林群落或生态系统固碳强度的直观指标(Zhou et al., 2010; Williams et al., 2014).然而单纯的NPP不足以反映群落或生态系统的固碳能力, 这是因为植被生长依赖于构建成本, 存在一定的呼吸消耗(Gifford, 2003).例如, 为了应对环境变化, 植被会调整其生长策略(Jar?u?ka & Barna, 2011), 在胁迫环境下, 植被可能将更多的碳用于维护与防御, 导致呼吸速率提高.尽管植被NPP可能会保持不变, 但投入到生长的资源占比却明显减少, 意味着植被固碳能力的降低.被定义为NPP/GPP或1 - Ra/GPP (即净生产力与总初级生产力(GPP)之比, Ra为自养呼吸速率)的碳利用效率(CUE)可以直接反映植被的固碳能力.不仅如此, 该指标还适用于描述和比较任意时间段内不同器官、植物个体和群落的固碳规律(朱万泽, 2013).CUE的引入为了解植被碳源/汇功能的变化提供了一个新的视角和切入点. ...
Plant respiration in productivity models: conceptualisation, representation and issues for global terrestrial carbon-cycle research 2 2003
... 森林在碳汇中发挥着不可替代的作用, 占陆地面积27.51%的森林年固碳量约为整个陆地圈的2/3 (Dixon et al., 1994; 项文化等, 2003).森林固碳强度及动态是预测森林固碳潜力的重要内容.植被生长是森林固碳的主要原因, 净初级生产力(NPP)是反映森林群落或生态系统固碳强度的直观指标(Zhou et al., 2010; Williams et al., 2014).然而单纯的NPP不足以反映群落或生态系统的固碳能力, 这是因为植被生长依赖于构建成本, 存在一定的呼吸消耗(Gifford, 2003).例如, 为了应对环境变化, 植被会调整其生长策略(Jar?u?ka & Barna, 2011), 在胁迫环境下, 植被可能将更多的碳用于维护与防御, 导致呼吸速率提高.尽管植被NPP可能会保持不变, 但投入到生长的资源占比却明显减少, 意味着植被固碳能力的降低.被定义为NPP/GPP或1 - Ra/GPP (即净生产力与总初级生产力(GPP)之比, Ra为自养呼吸速率)的碳利用效率(CUE)可以直接反映植被的固碳能力.不仅如此, 该指标还适用于描述和比较任意时间段内不同器官、植物个体和群落的固碳规律(朱万泽, 2013).CUE的引入为了解植被碳源/汇功能的变化提供了一个新的视角和切入点. ...
... 近年来有关CUE的研究主要集中在群落和生态系统尺度, 包括森林与草地植被生态系统(安相等, 2017; Shu et al., 2019; Zhang et al., 2019).这些研究揭示了CUE可能是由植被类型、环境条件和不同生长阶段共同决定的(朱万泽, 2013; Shu et al., 2019; Zhang et al., 2019).不仅如此, 森林发育过程中的树木大小分布也在很大程度上直接决定了群落和生态系统CUE的动态(Shu et al., 2019).在较长的时间尺度上, 群落CUE随树木个体的生长逐渐降低, 之后由于外界干扰的影响, 群落CUE可能会由于内部物种稳定的大小结构趋近于某一定值.这些结果既支持了群落CUE动态变化假说(Gifford, 2003; de Lucia et al., 2007), 也印证了CUE变化的最终稳定性观点, 证实了成熟森林可能普遍为碳汇的结论(Luyssaert et al., 2008; 朱万泽, 2020).然而, 森林生态系统不同组分CUE及其变化规律依然少有关注.不同组分如叶片、树干和树根间CUE有何联系? 它们对林分CUE的贡献如何等都值得进一步研究.这些问题的理解, 不仅有利于认识群落和生态系统的碳分配规律, 还可深入了解树木的生活史策略, 对理解气候变化下森林生态系统固碳能力的响应与适应亦有重要意义. ...
Variations in belowground carbon use strategies under different climatic conditions 1 2019
... 近年来有关CUE的研究主要集中在群落和生态系统尺度, 包括森林与草地植被生态系统(安相等, 2017; Shu et al., 2019; Zhang et al., 2019).这些研究揭示了CUE可能是由植被类型、环境条件和不同生长阶段共同决定的(朱万泽, 2013; Shu et al., 2019; Zhang et al., 2019).不仅如此, 森林发育过程中的树木大小分布也在很大程度上直接决定了群落和生态系统CUE的动态(Shu et al., 2019).在较长的时间尺度上, 群落CUE随树木个体的生长逐渐降低, 之后由于外界干扰的影响, 群落CUE可能会由于内部物种稳定的大小结构趋近于某一定值.这些结果既支持了群落CUE动态变化假说(Gifford, 2003; de Lucia et al., 2007), 也印证了CUE变化的最终稳定性观点, 证实了成熟森林可能普遍为碳汇的结论(Luyssaert et al., 2008; 朱万泽, 2020).然而, 森林生态系统不同组分CUE及其变化规律依然少有关注.不同组分如叶片、树干和树根间CUE有何联系? 它们对林分CUE的贡献如何等都值得进一步研究.这些问题的理解, 不仅有利于认识群落和生态系统的碳分配规律, 还可深入了解树木的生活史策略, 对理解气候变化下森林生态系统固碳能力的响应与适应亦有重要意义. ...
The ratio of NPP to GPP: evidence of change over the course of stand development 1 2001
... 两年的观测结果表明, 川西贡嘎山峨眉冷杉成熟林乔木层仍具有较高的CUE, 分别为0.43 (2016年)和0.41 (2017年)(图4), 表明了这一地区的峨眉冷杉成熟林具有较强的碳汇能力.该结果接近Shu等(2019)基于相同林分通量计算的CUE (0.401), 并支持了Gifford等(2003)森林CUE在0.40-0.45的结论.从植被类型上看, 川西亚高山针叶林CUE介于温带(0.50左右)(Waring et al., 1998; Chambers et al., 2004)和热带阔叶林(0.30-0.40)(Chambers et al., 2004; Malhi et al., 2009, 2011; Metcalfe et al., 2010; Malhi, 2012)之间.乔木层不同器官CUE也都保持较高水平.针叶(0.30或0.47)和树干CUE (0.40或0.31)与中部热带森林(0.23和0.43)(Chambers et al., 2004)接近, 细根CUE (0.55或0.57)略高于温带和热带森林(0.43-0.54)(Chen et al., 2011).林分中树木个体CUE存在明显差异.大径级树木CUE普遍要比小径级树木低0.20, 与林分生长发育过程中, 中幼林CUE高于成熟林的观点相似(M?kel? & Valentine, 2001; Shu et al., 2019).树干和针叶CUE的下降是引起树木CUE下降的主要原因.值得注意的是, 大径级冷杉(300年以上) CUE依然可以保持在0.35左右, 暗示峨眉冷杉树木CUE的降低是一个非常漫长的过程.林分的碳汇功能主要由具有较强固碳能力和最多数量(N = 13)的中径级树木所决定.不仅如此, 大径级树木的树干和细根生产力也没有明显下降(图3).稳定的林分结构维持了峨眉冷杉成熟林碳汇功能的长期稳定.来自林分的枯木质残体及生物量周转最终会以有机碳的形式进入土壤, 由于有机碳的增加会导致异养呼吸速率的不断减小(Zhou et al., 2006), 则峨眉冷杉成熟森林生态系统也会表现出稳定且长期的明显碳汇. ...
The productivity, metabolism and carbon cycle of tropical forest vegetation 1 2012
... 两年的观测结果表明, 川西贡嘎山峨眉冷杉成熟林乔木层仍具有较高的CUE, 分别为0.43 (2016年)和0.41 (2017年)(图4), 表明了这一地区的峨眉冷杉成熟林具有较强的碳汇能力.该结果接近Shu等(2019)基于相同林分通量计算的CUE (0.401), 并支持了Gifford等(2003)森林CUE在0.40-0.45的结论.从植被类型上看, 川西亚高山针叶林CUE介于温带(0.50左右)(Waring et al., 1998; Chambers et al., 2004)和热带阔叶林(0.30-0.40)(Chambers et al., 2004; Malhi et al., 2009, 2011; Metcalfe et al., 2010; Malhi, 2012)之间.乔木层不同器官CUE也都保持较高水平.针叶(0.30或0.47)和树干CUE (0.40或0.31)与中部热带森林(0.23和0.43)(Chambers et al., 2004)接近, 细根CUE (0.55或0.57)略高于温带和热带森林(0.43-0.54)(Chen et al., 2011).林分中树木个体CUE存在明显差异.大径级树木CUE普遍要比小径级树木低0.20, 与林分生长发育过程中, 中幼林CUE高于成熟林的观点相似(M?kel? & Valentine, 2001; Shu et al., 2019).树干和针叶CUE的下降是引起树木CUE下降的主要原因.值得注意的是, 大径级冷杉(300年以上) CUE依然可以保持在0.35左右, 暗示峨眉冷杉树木CUE的降低是一个非常漫长的过程.林分的碳汇功能主要由具有较强固碳能力和最多数量(N = 13)的中径级树木所决定.不仅如此, 大径级树木的树干和细根生产力也没有明显下降(图3).稳定的林分结构维持了峨眉冷杉成熟林碳汇功能的长期稳定.来自林分的枯木质残体及生物量周转最终会以有机碳的形式进入土壤, 由于有机碳的增加会导致异养呼吸速率的不断减小(Zhou et al., 2006), 则峨眉冷杉成熟森林生态系统也会表现出稳定且长期的明显碳汇. ...
The allocation of ecosystem net primary productivity in tropical forests 1 2011
... 两年的观测结果表明, 川西贡嘎山峨眉冷杉成熟林乔木层仍具有较高的CUE, 分别为0.43 (2016年)和0.41 (2017年)(图4), 表明了这一地区的峨眉冷杉成熟林具有较强的碳汇能力.该结果接近Shu等(2019)基于相同林分通量计算的CUE (0.401), 并支持了Gifford等(2003)森林CUE在0.40-0.45的结论.从植被类型上看, 川西亚高山针叶林CUE介于温带(0.50左右)(Waring et al., 1998; Chambers et al., 2004)和热带阔叶林(0.30-0.40)(Chambers et al., 2004; Malhi et al., 2009, 2011; Metcalfe et al., 2010; Malhi, 2012)之间.乔木层不同器官CUE也都保持较高水平.针叶(0.30或0.47)和树干CUE (0.40或0.31)与中部热带森林(0.23和0.43)(Chambers et al., 2004)接近, 细根CUE (0.55或0.57)略高于温带和热带森林(0.43-0.54)(Chen et al., 2011).林分中树木个体CUE存在明显差异.大径级树木CUE普遍要比小径级树木低0.20, 与林分生长发育过程中, 中幼林CUE高于成熟林的观点相似(M?kel? & Valentine, 2001; Shu et al., 2019).树干和针叶CUE的下降是引起树木CUE下降的主要原因.值得注意的是, 大径级冷杉(300年以上) CUE依然可以保持在0.35左右, 暗示峨眉冷杉树木CUE的降低是一个非常漫长的过程.林分的碳汇功能主要由具有较强固碳能力和最多数量(N = 13)的中径级树木所决定.不仅如此, 大径级树木的树干和细根生产力也没有明显下降(图3).稳定的林分结构维持了峨眉冷杉成熟林碳汇功能的长期稳定.来自林分的枯木质残体及生物量周转最终会以有机碳的形式进入土壤, 由于有机碳的增加会导致异养呼吸速率的不断减小(Zhou et al., 2006), 则峨眉冷杉成熟森林生态系统也会表现出稳定且长期的明显碳汇. ...
Shifts in plant respiration and carbon use efficiency at a large-scale drought experiment in the eastern Amazon 1 2010
... 两年的观测结果表明, 川西贡嘎山峨眉冷杉成熟林乔木层仍具有较高的CUE, 分别为0.43 (2016年)和0.41 (2017年)(图4), 表明了这一地区的峨眉冷杉成熟林具有较强的碳汇能力.该结果接近Shu等(2019)基于相同林分通量计算的CUE (0.401), 并支持了Gifford等(2003)森林CUE在0.40-0.45的结论.从植被类型上看, 川西亚高山针叶林CUE介于温带(0.50左右)(Waring et al., 1998; Chambers et al., 2004)和热带阔叶林(0.30-0.40)(Chambers et al., 2004; Malhi et al., 2009, 2011; Metcalfe et al., 2010; Malhi, 2012)之间.乔木层不同器官CUE也都保持较高水平.针叶(0.30或0.47)和树干CUE (0.40或0.31)与中部热带森林(0.23和0.43)(Chambers et al., 2004)接近, 细根CUE (0.55或0.57)略高于温带和热带森林(0.43-0.54)(Chen et al., 2011).林分中树木个体CUE存在明显差异.大径级树木CUE普遍要比小径级树木低0.20, 与林分生长发育过程中, 中幼林CUE高于成熟林的观点相似(M?kel? & Valentine, 2001; Shu et al., 2019).树干和针叶CUE的下降是引起树木CUE下降的主要原因.值得注意的是, 大径级冷杉(300年以上) CUE依然可以保持在0.35左右, 暗示峨眉冷杉树木CUE的降低是一个非常漫长的过程.林分的碳汇功能主要由具有较强固碳能力和最多数量(N = 13)的中径级树木所决定.不仅如此, 大径级树木的树干和细根生产力也没有明显下降(图3).稳定的林分结构维持了峨眉冷杉成熟林碳汇功能的长期稳定.来自林分的枯木质残体及生物量周转最终会以有机碳的形式进入土壤, 由于有机碳的增加会导致异养呼吸速率的不断减小(Zhou et al., 2006), 则峨眉冷杉成熟森林生态系统也会表现出稳定且长期的明显碳汇. ...
Global patterns in plant height 1 2009
... 由叶-导管-根组成的连续体通常被考虑为具有稳定关系的模块结构(Chen et al., 2019).然而在林分尺度下, 尤其是成熟林不同器官CUE的联系以及对林分尺度CUE的影响尚不明确.叶片作为树木摄取能量与资源的主要器官, 其资源利用策略影响其他器官和树木CUE.然而, 成熟林的叶片生长不再是资源投入的重点.研究表明, 成熟林的地下碳分配(包括根系生长、呼吸和分泌物)占总碳的25%-63% (Raich & Nadelhoffer, 1989), 其中细根周转是消耗地下NPP的主要途径(Kalyn & van Rees, 2006), 暗示对地下资源(如矿质元素)的摄取与利用可能是成熟林维持自身功能, 尤其是叶片生长和光合的必要条件, 因此, 细根CUE的改变可能会影响叶片CUE.作为树木生物量的主要构成部分, 树干不仅对树木光竞争具有重要作用(Moles et al., 2009), 而且也是储存和运输光合产物及水分的主要场所(Chantuma et al., 2009).在成熟林中, 光竞争趋于缓和, 树干已有较强的运输能力, 其生长可能较少地影响其他器官, 因而树干CUE的变化可能是相对独立的. ...
Maize root system growth and development as influenced by phosphorous deficiency 1 1999
Belowground carbon allocation in forest ecosystems: global trends 1 1989
... 由叶-导管-根组成的连续体通常被考虑为具有稳定关系的模块结构(Chen et al., 2019).然而在林分尺度下, 尤其是成熟林不同器官CUE的联系以及对林分尺度CUE的影响尚不明确.叶片作为树木摄取能量与资源的主要器官, 其资源利用策略影响其他器官和树木CUE.然而, 成熟林的叶片生长不再是资源投入的重点.研究表明, 成熟林的地下碳分配(包括根系生长、呼吸和分泌物)占总碳的25%-63% (Raich & Nadelhoffer, 1989), 其中细根周转是消耗地下NPP的主要途径(Kalyn & van Rees, 2006), 暗示对地下资源(如矿质元素)的摄取与利用可能是成熟林维持自身功能, 尤其是叶片生长和光合的必要条件, 因此, 细根CUE的改变可能会影响叶片CUE.作为树木生物量的主要构成部分, 树干不仅对树木光竞争具有重要作用(Moles et al., 2009), 而且也是储存和运输光合产物及水分的主要场所(Chantuma et al., 2009).在成熟林中, 光竞争趋于缓和, 树干已有较强的运输能力, 其生长可能较少地影响其他器官, 因而树干CUE的变化可能是相对独立的. ...
Effects of tree size heterogeneity on carbon sink in old forests 5 2019
... 近年来有关CUE的研究主要集中在群落和生态系统尺度, 包括森林与草地植被生态系统(安相等, 2017; Shu et al., 2019; Zhang et al., 2019).这些研究揭示了CUE可能是由植被类型、环境条件和不同生长阶段共同决定的(朱万泽, 2013; Shu et al., 2019; Zhang et al., 2019).不仅如此, 森林发育过程中的树木大小分布也在很大程度上直接决定了群落和生态系统CUE的动态(Shu et al., 2019).在较长的时间尺度上, 群落CUE随树木个体的生长逐渐降低, 之后由于外界干扰的影响, 群落CUE可能会由于内部物种稳定的大小结构趋近于某一定值.这些结果既支持了群落CUE动态变化假说(Gifford, 2003; de Lucia et al., 2007), 也印证了CUE变化的最终稳定性观点, 证实了成熟森林可能普遍为碳汇的结论(Luyssaert et al., 2008; 朱万泽, 2020).然而, 森林生态系统不同组分CUE及其变化规律依然少有关注.不同组分如叶片、树干和树根间CUE有何联系? 它们对林分CUE的贡献如何等都值得进一步研究.这些问题的理解, 不仅有利于认识群落和生态系统的碳分配规律, 还可深入了解树木的生活史策略, 对理解气候变化下森林生态系统固碳能力的响应与适应亦有重要意义. ...
... ; Shu et al., 2019; Zhang et al., 2019).不仅如此, 森林发育过程中的树木大小分布也在很大程度上直接决定了群落和生态系统CUE的动态(Shu et al., 2019).在较长的时间尺度上, 群落CUE随树木个体的生长逐渐降低, 之后由于外界干扰的影响, 群落CUE可能会由于内部物种稳定的大小结构趋近于某一定值.这些结果既支持了群落CUE动态变化假说(Gifford, 2003; de Lucia et al., 2007), 也印证了CUE变化的最终稳定性观点, 证实了成熟森林可能普遍为碳汇的结论(Luyssaert et al., 2008; 朱万泽, 2020).然而, 森林生态系统不同组分CUE及其变化规律依然少有关注.不同组分如叶片、树干和树根间CUE有何联系? 它们对林分CUE的贡献如何等都值得进一步研究.这些问题的理解, 不仅有利于认识群落和生态系统的碳分配规律, 还可深入了解树木的生活史策略, 对理解气候变化下森林生态系统固碳能力的响应与适应亦有重要意义. ...
... 的动态(Shu et al., 2019).在较长的时间尺度上, 群落CUE随树木个体的生长逐渐降低, 之后由于外界干扰的影响, 群落CUE可能会由于内部物种稳定的大小结构趋近于某一定值.这些结果既支持了群落CUE动态变化假说(Gifford, 2003; de Lucia et al., 2007), 也印证了CUE变化的最终稳定性观点, 证实了成熟森林可能普遍为碳汇的结论(Luyssaert et al., 2008; 朱万泽, 2020).然而, 森林生态系统不同组分CUE及其变化规律依然少有关注.不同组分如叶片、树干和树根间CUE有何联系? 它们对林分CUE的贡献如何等都值得进一步研究.这些问题的理解, 不仅有利于认识群落和生态系统的碳分配规律, 还可深入了解树木的生活史策略, 对理解气候变化下森林生态系统固碳能力的响应与适应亦有重要意义. ...
... 研究地位于在中国科学院贡嘎山高山生态系统观测试验站(简称贡嘎山站), 林分分层明显, 其中乔木层占比巨大, 决定了整个林分的碳收支(Shu et al., 2019), 并几乎都由峨眉冷杉构成.峨眉冷杉平均年龄为120 a, 平均树高为25 m, 平均胸径为47.3 cm, 郁闭度0.7左右, 密度151株·hm-2; 鲜有糙皮桦(Betula utilis)、香桦(Betula insignis)、江南花楸(Sorbus hemsleyi)等阔叶树种.除峨眉冷杉幼树外, 灌木层主要是冷箭竹(Bashania fangiana).草本层主要有鹿药(Smilacina japonica)、窄瓣鹿药(Smilacina paniculata)和紫花碎米荠(Cardamine tangutorum), 以及山羽藓(Abietinella abietina)和锦丝藓(Actinothuidium hookeri)等.峨眉冷杉的生长季为5-10月, 非生长季为11月至次年4月. ...
... 两年的观测结果表明, 川西贡嘎山峨眉冷杉成熟林乔木层仍具有较高的CUE, 分别为0.43 (2016年)和0.41 (2017年)(图4), 表明了这一地区的峨眉冷杉成熟林具有较强的碳汇能力.该结果接近Shu等(2019)基于相同林分通量计算的CUE (0.401), 并支持了Gifford等(2003)森林CUE在0.40-0.45的结论.从植被类型上看, 川西亚高山针叶林CUE介于温带(0.50左右)(Waring et al., 1998; Chambers et al., 2004)和热带阔叶林(0.30-0.40)(Chambers et al., 2004; Malhi et al., 2009, 2011; Metcalfe et al., 2010; Malhi, 2012)之间.乔木层不同器官CUE也都保持较高水平.针叶(0.30或0.47)和树干CUE (0.40或0.31)与中部热带森林(0.23和0.43)(Chambers et al., 2004)接近, 细根CUE (0.55或0.57)略高于温带和热带森林(0.43-0.54)(Chen et al., 2011).林分中树木个体CUE存在明显差异.大径级树木CUE普遍要比小径级树木低0.20, 与林分生长发育过程中, 中幼林CUE高于成熟林的观点相似(M?kel? & Valentine, 2001; Shu et al., 2019).树干和针叶CUE的下降是引起树木CUE下降的主要原因.值得注意的是, 大径级冷杉(300年以上) CUE依然可以保持在0.35左右, 暗示峨眉冷杉树木CUE的降低是一个非常漫长的过程.林分的碳汇功能主要由具有较强固碳能力和最多数量(N = 13)的中径级树木所决定.不仅如此, 大径级树木的树干和细根生产力也没有明显下降(图3).稳定的林分结构维持了峨眉冷杉成熟林碳汇功能的长期稳定.来自林分的枯木质残体及生物量周转最终会以有机碳的形式进入土壤, 由于有机碳的增加会导致异养呼吸速率的不断减小(Zhou et al., 2006), 则峨眉冷杉成熟森林生态系统也会表现出稳定且长期的明显碳汇. ...
Net primary production of forests: a constant fraction of gross primary production? 1 1998
... 两年的观测结果表明, 川西贡嘎山峨眉冷杉成熟林乔木层仍具有较高的CUE, 分别为0.43 (2016年)和0.41 (2017年)(图4), 表明了这一地区的峨眉冷杉成熟林具有较强的碳汇能力.该结果接近Shu等(2019)基于相同林分通量计算的CUE (0.401), 并支持了Gifford等(2003)森林CUE在0.40-0.45的结论.从植被类型上看, 川西亚高山针叶林CUE介于温带(0.50左右)(Waring et al., 1998; Chambers et al., 2004)和热带阔叶林(0.30-0.40)(Chambers et al., 2004; Malhi et al., 2009, 2011; Metcalfe et al., 2010; Malhi, 2012)之间.乔木层不同器官CUE也都保持较高水平.针叶(0.30或0.47)和树干CUE (0.40或0.31)与中部热带森林(0.23和0.43)(Chambers et al., 2004)接近, 细根CUE (0.55或0.57)略高于温带和热带森林(0.43-0.54)(Chen et al., 2011).林分中树木个体CUE存在明显差异.大径级树木CUE普遍要比小径级树木低0.20, 与林分生长发育过程中, 中幼林CUE高于成熟林的观点相似(M?kel? & Valentine, 2001; Shu et al., 2019).树干和针叶CUE的下降是引起树木CUE下降的主要原因.值得注意的是, 大径级冷杉(300年以上) CUE依然可以保持在0.35左右, 暗示峨眉冷杉树木CUE的降低是一个非常漫长的过程.林分的碳汇功能主要由具有较强固碳能力和最多数量(N = 13)的中径级树木所决定.不仅如此, 大径级树木的树干和细根生产力也没有明显下降(图3).稳定的林分结构维持了峨眉冷杉成熟林碳汇功能的长期稳定.来自林分的枯木质残体及生物量周转最终会以有机碳的形式进入土壤, 由于有机碳的增加会导致异养呼吸速率的不断减小(Zhou et al., 2006), 则峨眉冷杉成熟森林生态系统也会表现出稳定且长期的明显碳汇. ...
Impacts of disturbance history on forest carbon stocks and fluxes: merging satellite disturbance mapping with forest inventory data in a carbon cycle model framework 1 2014
... 森林在碳汇中发挥着不可替代的作用, 占陆地面积27.51%的森林年固碳量约为整个陆地圈的2/3 (Dixon et al., 1994; 项文化等, 2003).森林固碳强度及动态是预测森林固碳潜力的重要内容.植被生长是森林固碳的主要原因, 净初级生产力(NPP)是反映森林群落或生态系统固碳强度的直观指标(Zhou et al., 2010; Williams et al., 2014).然而单纯的NPP不足以反映群落或生态系统的固碳能力, 这是因为植被生长依赖于构建成本, 存在一定的呼吸消耗(Gifford, 2003).例如, 为了应对环境变化, 植被会调整其生长策略(Jar?u?ka & Barna, 2011), 在胁迫环境下, 植被可能将更多的碳用于维护与防御, 导致呼吸速率提高.尽管植被NPP可能会保持不变, 但投入到生长的资源占比却明显减少, 意味着植被固碳能力的降低.被定义为NPP/GPP或1 - Ra/GPP (即净生产力与总初级生产力(GPP)之比, Ra为自养呼吸速率)的碳利用效率(CUE)可以直接反映植被的固碳能力.不仅如此, 该指标还适用于描述和比较任意时间段内不同器官、植物个体和群落的固碳规律(朱万泽, 2013).CUE的引入为了解植被碳源/汇功能的变化提供了一个新的视角和切入点. ...
Differential responses of ecosystem carbon flux components to experimental precipitation gradient in an alpine meadow 2 2019
... 近年来有关CUE的研究主要集中在群落和生态系统尺度, 包括森林与草地植被生态系统(安相等, 2017; Shu et al., 2019; Zhang et al., 2019).这些研究揭示了CUE可能是由植被类型、环境条件和不同生长阶段共同决定的(朱万泽, 2013; Shu et al., 2019; Zhang et al., 2019).不仅如此, 森林发育过程中的树木大小分布也在很大程度上直接决定了群落和生态系统CUE的动态(Shu et al., 2019).在较长的时间尺度上, 群落CUE随树木个体的生长逐渐降低, 之后由于外界干扰的影响, 群落CUE可能会由于内部物种稳定的大小结构趋近于某一定值.这些结果既支持了群落CUE动态变化假说(Gifford, 2003; de Lucia et al., 2007), 也印证了CUE变化的最终稳定性观点, 证实了成熟森林可能普遍为碳汇的结论(Luyssaert et al., 2008; 朱万泽, 2020).然而, 森林生态系统不同组分CUE及其变化规律依然少有关注.不同组分如叶片、树干和树根间CUE有何联系? 它们对林分CUE的贡献如何等都值得进一步研究.这些问题的理解, 不仅有利于认识群落和生态系统的碳分配规律, 还可深入了解树木的生活史策略, 对理解气候变化下森林生态系统固碳能力的响应与适应亦有重要意义. ...
... ; Zhang et al., 2019).不仅如此, 森林发育过程中的树木大小分布也在很大程度上直接决定了群落和生态系统CUE的动态(Shu et al., 2019).在较长的时间尺度上, 群落CUE随树木个体的生长逐渐降低, 之后由于外界干扰的影响, 群落CUE可能会由于内部物种稳定的大小结构趋近于某一定值.这些结果既支持了群落CUE动态变化假说(Gifford, 2003; de Lucia et al., 2007), 也印证了CUE变化的最终稳定性观点, 证实了成熟森林可能普遍为碳汇的结论(Luyssaert et al., 2008; 朱万泽, 2020).然而, 森林生态系统不同组分CUE及其变化规律依然少有关注.不同组分如叶片、树干和树根间CUE有何联系? 它们对林分CUE的贡献如何等都值得进一步研究.这些问题的理解, 不仅有利于认识群落和生态系统的碳分配规律, 还可深入了解树木的生活史策略, 对理解气候变化下森林生态系统固碳能力的响应与适应亦有重要意义. ...
Spatial patterns of ecosystem carbon residence time in Chinese forests 1 2010
... 森林在碳汇中发挥着不可替代的作用, 占陆地面积27.51%的森林年固碳量约为整个陆地圈的2/3 (Dixon et al., 1994; 项文化等, 2003).森林固碳强度及动态是预测森林固碳潜力的重要内容.植被生长是森林固碳的主要原因, 净初级生产力(NPP)是反映森林群落或生态系统固碳强度的直观指标(Zhou et al., 2010; Williams et al., 2014).然而单纯的NPP不足以反映群落或生态系统的固碳能力, 这是因为植被生长依赖于构建成本, 存在一定的呼吸消耗(Gifford, 2003).例如, 为了应对环境变化, 植被会调整其生长策略(Jar?u?ka & Barna, 2011), 在胁迫环境下, 植被可能将更多的碳用于维护与防御, 导致呼吸速率提高.尽管植被NPP可能会保持不变, 但投入到生长的资源占比却明显减少, 意味着植被固碳能力的降低.被定义为NPP/GPP或1 - Ra/GPP (即净生产力与总初级生产力(GPP)之比, Ra为自养呼吸速率)的碳利用效率(CUE)可以直接反映植被的固碳能力.不仅如此, 该指标还适用于描述和比较任意时间段内不同器官、植物个体和群落的固碳规律(朱万泽, 2013).CUE的引入为了解植被碳源/汇功能的变化提供了一个新的视角和切入点. ...
森林碳利用效率研究进展 3 2013
... 森林在碳汇中发挥着不可替代的作用, 占陆地面积27.51%的森林年固碳量约为整个陆地圈的2/3 (Dixon et al., 1994; 项文化等, 2003).森林固碳强度及动态是预测森林固碳潜力的重要内容.植被生长是森林固碳的主要原因, 净初级生产力(NPP)是反映森林群落或生态系统固碳强度的直观指标(Zhou et al., 2010; Williams et al., 2014).然而单纯的NPP不足以反映群落或生态系统的固碳能力, 这是因为植被生长依赖于构建成本, 存在一定的呼吸消耗(Gifford, 2003).例如, 为了应对环境变化, 植被会调整其生长策略(Jar?u?ka & Barna, 2011), 在胁迫环境下, 植被可能将更多的碳用于维护与防御, 导致呼吸速率提高.尽管植被NPP可能会保持不变, 但投入到生长的资源占比却明显减少, 意味着植被固碳能力的降低.被定义为NPP/GPP或1 - Ra/GPP (即净生产力与总初级生产力(GPP)之比, Ra为自养呼吸速率)的碳利用效率(CUE)可以直接反映植被的固碳能力.不仅如此, 该指标还适用于描述和比较任意时间段内不同器官、植物个体和群落的固碳规律(朱万泽, 2013).CUE的引入为了解植被碳源/汇功能的变化提供了一个新的视角和切入点. ...
... 近年来有关CUE的研究主要集中在群落和生态系统尺度, 包括森林与草地植被生态系统(安相等, 2017; Shu et al., 2019; Zhang et al., 2019).这些研究揭示了CUE可能是由植被类型、环境条件和不同生长阶段共同决定的(朱万泽, 2013; Shu et al., 2019; Zhang et al., 2019).不仅如此, 森林发育过程中的树木大小分布也在很大程度上直接决定了群落和生态系统CUE的动态(Shu et al., 2019).在较长的时间尺度上, 群落CUE随树木个体的生长逐渐降低, 之后由于外界干扰的影响, 群落CUE可能会由于内部物种稳定的大小结构趋近于某一定值.这些结果既支持了群落CUE动态变化假说(Gifford, 2003; de Lucia et al., 2007), 也印证了CUE变化的最终稳定性观点, 证实了成熟森林可能普遍为碳汇的结论(Luyssaert et al., 2008; 朱万泽, 2020).然而, 森林生态系统不同组分CUE及其变化规律依然少有关注.不同组分如叶片、树干和树根间CUE有何联系? 它们对林分CUE的贡献如何等都值得进一步研究.这些问题的理解, 不仅有利于认识群落和生态系统的碳分配规律, 还可深入了解树木的生活史策略, 对理解气候变化下森林生态系统固碳能力的响应与适应亦有重要意义. ...
... 森林在碳汇中发挥着不可替代的作用, 占陆地面积27.51%的森林年固碳量约为整个陆地圈的2/3 (Dixon et al., 1994; 项文化等, 2003).森林固碳强度及动态是预测森林固碳潜力的重要内容.植被生长是森林固碳的主要原因, 净初级生产力(NPP)是反映森林群落或生态系统固碳强度的直观指标(Zhou et al., 2010; Williams et al., 2014).然而单纯的NPP不足以反映群落或生态系统的固碳能力, 这是因为植被生长依赖于构建成本, 存在一定的呼吸消耗(Gifford, 2003).例如, 为了应对环境变化, 植被会调整其生长策略(Jar?u?ka & Barna, 2011), 在胁迫环境下, 植被可能将更多的碳用于维护与防御, 导致呼吸速率提高.尽管植被NPP可能会保持不变, 但投入到生长的资源占比却明显减少, 意味着植被固碳能力的降低.被定义为NPP/GPP或1 - Ra/GPP (即净生产力与总初级生产力(GPP)之比, Ra为自养呼吸速率)的碳利用效率(CUE)可以直接反映植被的固碳能力.不仅如此, 该指标还适用于描述和比较任意时间段内不同器官、植物个体和群落的固碳规律(朱万泽, 2013).CUE的引入为了解植被碳源/汇功能的变化提供了一个新的视角和切入点. ...
... 近年来有关CUE的研究主要集中在群落和生态系统尺度, 包括森林与草地植被生态系统(安相等, 2017; Shu et al., 2019; Zhang et al., 2019).这些研究揭示了CUE可能是由植被类型、环境条件和不同生长阶段共同决定的(朱万泽, 2013; Shu et al., 2019; Zhang et al., 2019).不仅如此, 森林发育过程中的树木大小分布也在很大程度上直接决定了群落和生态系统CUE的动态(Shu et al., 2019).在较长的时间尺度上, 群落CUE随树木个体的生长逐渐降低, 之后由于外界干扰的影响, 群落CUE可能会由于内部物种稳定的大小结构趋近于某一定值.这些结果既支持了群落CUE动态变化假说(Gifford, 2003; de Lucia et al., 2007), 也印证了CUE变化的最终稳定性观点, 证实了成熟森林可能普遍为碳汇的结论(Luyssaert et al., 2008; 朱万泽, 2020).然而, 森林生态系统不同组分CUE及其变化规律依然少有关注.不同组分如叶片、树干和树根间CUE有何联系? 它们对林分CUE的贡献如何等都值得进一步研究.这些问题的理解, 不仅有利于认识群落和生态系统的碳分配规律, 还可深入了解树木的生活史策略, 对理解气候变化下森林生态系统固碳能力的响应与适应亦有重要意义. ...
... 近年来有关CUE的研究主要集中在群落和生态系统尺度, 包括森林与草地植被生态系统(安相等, 2017; Shu et al., 2019; Zhang et al., 2019).这些研究揭示了CUE可能是由植被类型、环境条件和不同生长阶段共同决定的(朱万泽, 2013; Shu et al., 2019; Zhang et al., 2019).不仅如此, 森林发育过程中的树木大小分布也在很大程度上直接决定了群落和生态系统CUE的动态(Shu et al., 2019).在较长的时间尺度上, 群落CUE随树木个体的生长逐渐降低, 之后由于外界干扰的影响, 群落CUE可能会由于内部物种稳定的大小结构趋近于某一定值.这些结果既支持了群落CUE动态变化假说(Gifford, 2003; de Lucia et al., 2007), 也印证了CUE变化的最终稳定性观点, 证实了成熟森林可能普遍为碳汇的结论(Luyssaert et al., 2008; 朱万泽, 2020).然而, 森林生态系统不同组分CUE及其变化规律依然少有关注.不同组分如叶片、树干和树根间CUE有何联系? 它们对林分CUE的贡献如何等都值得进一步研究.这些问题的理解, 不仅有利于认识群落和生态系统的碳分配规律, 还可深入了解树木的生活史策略, 对理解气候变化下森林生态系统固碳能力的响应与适应亦有重要意义. ...
成熟森林固碳研究进展 1 2020
... 近年来有关CUE的研究主要集中在群落和生态系统尺度, 包括森林与草地植被生态系统(安相等, 2017; Shu et al., 2019; Zhang et al., 2019).这些研究揭示了CUE可能是由植被类型、环境条件和不同生长阶段共同决定的(朱万泽, 2013; Shu et al., 2019; Zhang et al., 2019).不仅如此, 森林发育过程中的树木大小分布也在很大程度上直接决定了群落和生态系统CUE的动态(Shu et al., 2019).在较长的时间尺度上, 群落CUE随树木个体的生长逐渐降低, 之后由于外界干扰的影响, 群落CUE可能会由于内部物种稳定的大小结构趋近于某一定值.这些结果既支持了群落CUE动态变化假说(Gifford, 2003; de Lucia et al., 2007), 也印证了CUE变化的最终稳定性观点, 证实了成熟森林可能普遍为碳汇的结论(Luyssaert et al., 2008; 朱万泽, 2020).然而, 森林生态系统不同组分CUE及其变化规律依然少有关注.不同组分如叶片、树干和树根间CUE有何联系? 它们对林分CUE的贡献如何等都值得进一步研究.这些问题的理解, 不仅有利于认识群落和生态系统的碳分配规律, 还可深入了解树木的生活史策略, 对理解气候变化下森林生态系统固碳能力的响应与适应亦有重要意义. ...
 ,1,2, 朱万泽
,1,2, 朱万泽 ,1,*, 冉飞1, 孙守琴1, 张元媛1,2
,1,*, 冉飞1, 孙守琴1, 张元媛1,2
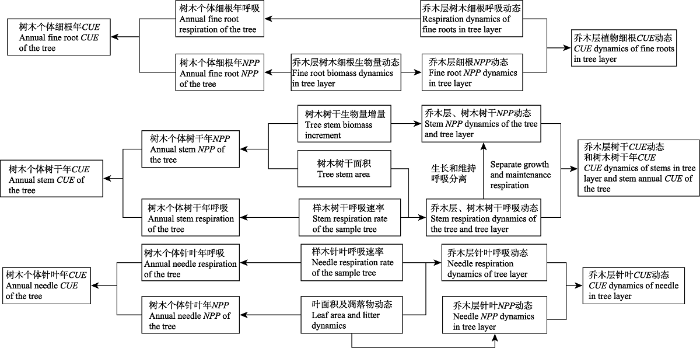 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT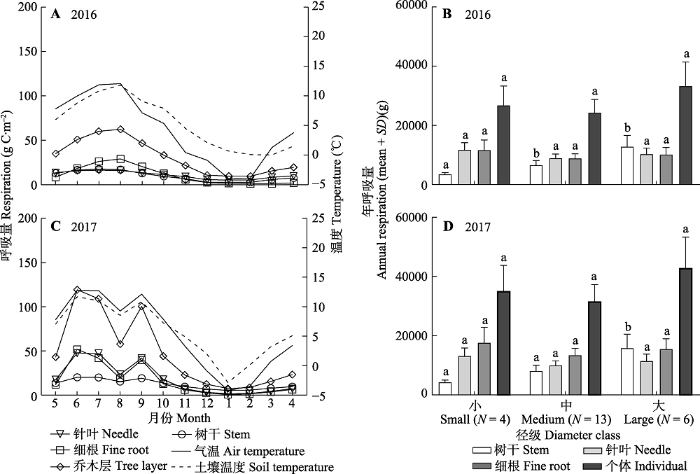 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT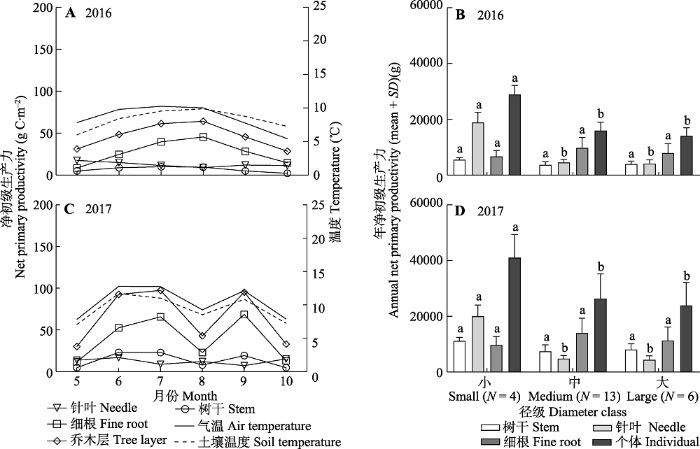 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT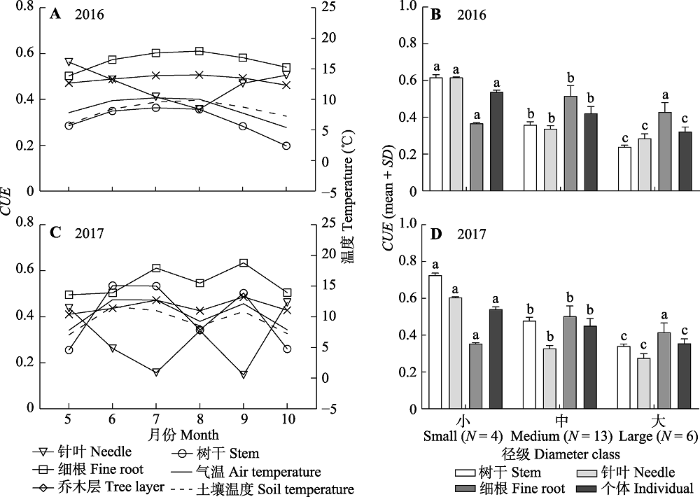 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT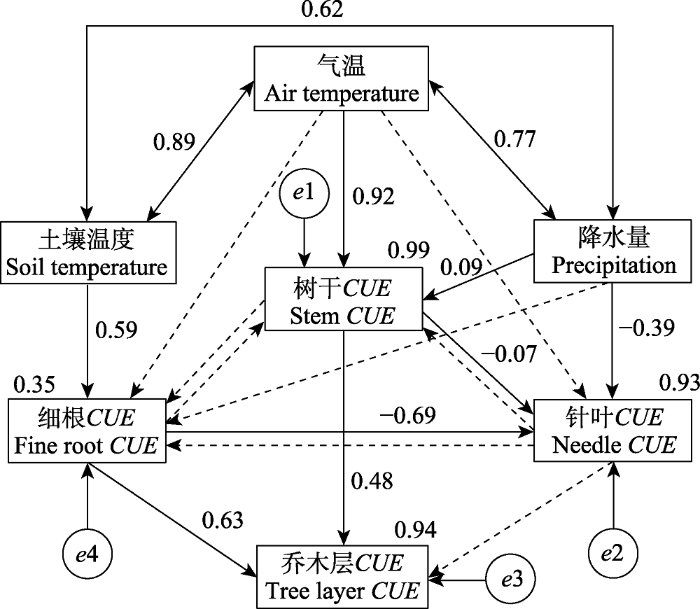 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}