拓锋
1 , 2 , 刘贤德
, 1 , 2 , * , 刘润红
3 , 赵维俊
2 , 敬文茂
2 , 马剑
2 , 武秀荣
2 , 赵晶忠
2 , 马雪娥
2 1 甘肃农业大学林学院, 兰州 7300702 甘肃省祁连山水源涵养林研究院(甘肃祁连山森林生态系统国家定位观测研究站, 甘肃祁连山森林生态监测与评估国际科技合作基地, 甘肃省森林生态与冻土水文水资源重点实验室, 甘肃省祁连山森林生态系统野外科学观测研究站), 甘肃张掖 7340003 兰州大学生命科学学院, 兰州 730000Spatial distribution patterns and association of Picea crassifolia population in Dayekou Basin of Qilian Mountains, northwestern China Feng TA
1 , 2 , Xian-De LIU
, 1 , 2 , * , Run-Hong LIU
3 , Wei-Jun ZHAO
2 , Wen-Mao JING
2 , Jian MA
2 , Xiu-Rong WU
2 , Jing-Zhong ZHAO
2 , Xue-E MA
2 1 College of Forestry, Gansu Agricultural University, Lanzhou 730070, China2 Gansu Province Qilian Water Resource Conservation Forest Research Institute (Gansu Qilian Mountain Forest Eco-system of the State Research Station, Gansu Qilian Mountain Forest Ecological Monitoring and Evaluation International Science and Technology Cooperation Base, Key Laboratory of Hydrology and Water Resources of Forest Ecology and Frozen Soil of Gansu Province, Gansu Qilian Mountain Forest Ecosystem Observation and Research Station), Zhangye, Gansu 734000, China3 School of Life Sciences, Lanzhou University, Lanzhou 730000, China通讯作者: *
liuxiande666@163.com 编委: 李镇清
责任编辑: 李敏
收稿日期: 2020-06-1
接受日期: 2020-06-1
网络出版日期: 2020-11-20
基金资助: 甘肃省自然科学基金 (18JR4RA002 )
Corresponding authors: *
liuxiande666@163.com Received: 2020-06-1
Accepted: 2020-06-1
Online: 2020-11-20
Fund supported: Natural Science Foundation of Gansu Province (18JR4RA002 )
摘要 研究青海云杉(
Picea crassifolia )种群不同发育阶段的空间分布格局及其关联性, 能揭示其种群的发展规律, 推断其潜在的生态学过程或影响机制, 进而为该种群的配置、种植和森林的经营与管理等提供指导。该研究基于祁连山大野口流域10.2 hm
2 青海云杉动态监测样地调查资料, 分析了青海云杉群落的物种组成和径级结构, 并采用点格局方法, 通过单变量成对相关函数和双变量成对相关函数对不同年龄阶段青海云杉种群的空间分布格局及其空间关联性进行分析。结果表明: (1)青海云杉种群的径级结构呈倒“J”型分布, 属增长型种群, 种群自然更新状况良好。(2)青海云杉种群4个径级的个体在小尺度上呈聚集分布, 但随着空间尺度的增大, 聚集强度逐渐减弱, 趋向随机分布; 较小径级的个体常呈聚集分布, 而随着径级增大, 也趋向于随机分布。(3)青海云杉大树个体在小尺度(<4 m)范围内与其他径级个体都呈负关联, 随着尺度的增加, 大树与幼树表现出正关联且关联性随尺度的增加而减弱, 而与小树、中树则表现为无关联或者微弱的负关联; 径级相近的个体空间关联性为正关联或无关联, 随着个体径级间差距增大, 空间关联性转变为无关联或负关联。研究表明, 祁连山大野口流域青海云杉种群的分布格局及其关联性随空间尺度和发育阶段而变化, 这是由种群自身的生物生态学特性、种内种间关系以及环境条件共同作用的结果。
关键词: 祁连山 ;
青海云杉种群 ;
成对相关函数 ;
径级结构 ;
空间分布格局 ;
空间关联 Abstract Aims Exploring the spatial distribution pattern and the association among different developmental stages of
Picea crassifolia populations can reveal the succession patterns of the populations, infer their potential ecological mechanisms, and thus provide guidance for the configuration, planting, forest management and management of the populations.
Methods Based on the survey data of a
P. crassifolia dynamic monitoring plot (10.2 hm
2 ) in Dayekou basin, Qilian Mountains, China. The species composition and diameter structure of the community were analyzed, and the point pattern method was used to analyze the spatial distribution pattern and spatial correlation of
P. crassifolia populations at different age stages through univariate pairwise correlation functions and bivariate pairwise correlation functions.
Important findings The results showed that: (1) The diameter class structure of
P. crassifolia population was an inverted “J” shape, indicating that the population was a growing population and the population was in good natural renewal status. (2) The individuals of four diameter classes in the
P. crassifolia population were aggregated at a small scale, but as the spatial scale increased, the aggregation intensity gradually weakened and tended to be randomly distributed. Moreover, individuals of smaller diameter classes used to be aggregated, but as the diameter increased, they also tended to be randomly distributed. (3) The big trees in the
P. crassifolia population were negatively correlated with other diameter classes at a small scale (<4 m). As the scale increased, the big trees and young trees showed a positive association and the correlation decreased with increasing scale. However, as the scale increased, big trees showed no or weakly negative correlation with small and medium trees. In addition, the spatial association of individuals with similar diameter classes showed positive or no association. As the difference between individual diameter classes increased, their spatial association changed to no or negative association. In conclusion, the spatial distribution patterns and association of
P. crassifolia population in Dayekou Basin of Qilian Mountains varied with the spatial scale and developmental stage, which was the result of the combined effects of the population’s bioecological characteristics, intraspecific and interspecific relationships, and environmental conditions.
Keywords: Qilian Mountains ;
Picea crassifolia population;
pairwise correlation function ;
diameter structure ;
spatial distribution patterns ;
spatial association PDF (1626KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 拓锋, 刘贤德, 刘润红, 赵维俊, 敬文茂, 马剑, 武秀荣, 赵晶忠, 马雪娥. 祁连山大野口流域青海云杉种群空间格局及其 关联性. 植物生态学报, 2020, 44(11): 1172-1183. DOI:
10.17521/cjpe.2020.0177 TA Feng, LIU Xian-De, LIU Run-Hong, ZHAO Wei-Jun, JING Wen-Mao, MA Jian, WU Xiu-Rong, ZHAO Jing-Zhong, MA Xue-E.
Spatial distribution patterns and association of Picea crassifolia population in Dayekou Basin of Qilian Mountains, northwestern China .
Chinese Journal of Plant Ecology , 2020, 44(11): 1172-1183. DOI:
10.17521/cjpe.2020.0177 种群的空间格局是指特定时间内, 种群所有个体在其水平生活空间内的分布状况及其属性在空间上的配置特征, 往往是由种群自身生物学特性、种内种间竞争、环境条件等因素综合作用的结果(
林玥, 2008 ;
Liu et al ., 2020 )。种群的空间分布格局和空间关联性是种群空间格局研究的2个主要内容, 也是种群生态关系在空间格局上的两种表现形式(
张金屯和孟东平, 2004 )。大量研究表明种群的空间分布格局与空间关联性关系密切, 聚集分布和空间正关联表明种群内部正向(互惠)的生态关系, 规则分布和空间负关联则表明种群内部负向(互斥)的生态关系, 随机分布和空间无关联则代表着种群内部不存在明确的生态关系(
杨洪晓, 2006 )。因此, 研究植物种群的空间分布格局及其关联性不仅能够了解种群的空间水平结构与生物、生态学习性(如生活史策略、是否喜光或耐阴等)、种群间相互作用、种群与环境因子的相互作用规律(如植物与生长环境间的适合度、生境异质性等)以及种群在群落中的地位和生态学功能, 更重要的是可以揭示这种分布格局及其关联性的潜在成因, 进而推演种群动态变化的生态学过程(如种内和种间竞争、种子扩散和干扰等), 深化对群落结构的认识, 揭示植物群落的生物多样性形成和维持机制(
McDonald et al ., 2003 ;
Hou et al , 2004 ;
Wiegand & Moloney, 2004 ;
Nathan, 2006 ;
Wang et al ., 2010 ;
Guo et al ., 2013 )。
空间分布格局的研究方法众多, 早期的研究方法主要有: 以频度/密度为基础的样方法、以距离为基础的无样地法、方差区组分析法、双向轨迹方差分析、
χ 2 检验和分形理论等(
张金屯, 1998 ;
王本洋和余世孝, 2005 )。传统的样方取样和分析方法常基于单一尺度(固定大小样方)的若干样方, 常采用
χ 2 分析法和方差/均值比率法等统计分析方法, 然而种群空间分布格局及其关联性对种群密度和空间尺度均有很强的依赖性, 在不同的尺度上往往表现出不同的类型, 故早期的取样和分析方法只能研究特定尺度下种群的空间分布格局特征, 难以全面地反映森林内物种的空间分布特点和种间关系(
He et al ., 1997 ;
李立等, 2010 )。鉴于上述研究方法的局限性,
Ripley (1977) 和
Diggle (1983) 分别提出和完善了点格局分析法, 以分析各种尺度上的空间分布格局。该方法以数字化的植物个体位置信息为基本数据, 将每个个体都视为二维空间中的一个点, 所有个体组成了植物种的空间分布图, 然后在空间分布图的基础上进行种群空间格局分析。其优点是能最大限度地利用点与点之间的距离, 不仅能够提供全面的空间格局信息, 而且具有较强的检验能力, 克服了传统研究方法中尺度单一等不足, 可以分析任意尺度下种群的空间格局特征及种内、种间关系(
Wiegand & Moloney, 2004 ;
Schleicher et al ., 2011 ;
沈志强等, 2016 ;
涂洪润等, 2019 )。因此, 空间点格局分析法在种群空间格局分布特征及潜在生态过程假说的研究方面得到了广泛应用。
祁连山是中国西部的主要山脉之一, 位于青藏高原、蒙古高原、黄土高原三大高原交汇带, 是石羊河、黑河、疏勒河等6大内陆河和黄河流域的重要水源产流地, 维系着河西乃至西部地区生存与发展的命脉, 在国家生态建设中具有十分重要的战略地位(
Gao et al ., 2013 ;
Zhu et al ., 2017 )。祁连山山地森林的主要建群种——青海云杉(
Picea crassifolia ), 为松科云杉属(
Picea )乔木, 是中国青藏高原东北边缘特有树种, 多以纯林形式存在, 呈带状或斑块状分布于海拔2 300-3 300 m的阴坡和半阴坡, 分布面积占祁连山水源涵养林面积的24.74%, 占乔木林面积的75.72%, 对于维系祁连山的生物多样性、水源涵养功能乃至区域生态平衡都具有至关重要的作用(
田风霞等, 2012 ;
何芳兰等, 2016 ;
张华等, 2016 )。部分****(
赵丽琼等, 2010 ;
张华等, 2016 )虽然已对青海云杉种群空间格局及其关联性进行了研究, 但是已有研究的空间尺度相对较小, 仅能揭示小尺度下的种群空间分布格局, 难以客观地反映青海云杉种群空间分布的特点和种内关系。鉴于此, 本研究基于祁连山大野口流域10.2 hm
2 青海云杉动态监测样地数据, 采用空间点格局分析方法中的成对相关函数对不同径级青海云杉种群的空间分布格局及其关联性进行分析, 旨在阐明: (1)祁连山大野口流域青海云杉种群不同径级个体的空间分布格局及其关联性如何随空间尺度进行变化; (2)不同径级个体的空间分布格局及其关联性形成的潜在生态学过程与影响机制。上述科学问题的合理解答, 能从空间格局的角度客观地揭示青海云杉种群发展规律, 可以为制定该种群的配置、种植与保护管理措施提供指导, 并对祁连山地区的植被恢复与重建、生物多样性保护等具有重要的意义和价值。
1 材料和方法 1.1 研究样地概况 研究样地位于祁连山中段北坡大野口流域青海云杉林内, 地理位置为100.25° E, 38.53° N。样地最低海拔2 856 m, 最高海拔2 962 m, 最大高差106 m, 坡向西北, 坡度13°。根据祁连山水源涵养林研究院的资料, 样地所在的大野口流域气候属于温带大陆性高寒半湿润山地气候, 雨热同期, 冬季寒冷漫长, 夏季短暂, 年平均气温5.4 ℃, 最冷月(1月)平均气温为-12.5 ℃, 最热月(7月)平均气温为19.6 ℃, 年降水量为300-500 mm, 主要集中在6-9月, 年平均相对湿度60% (1994-2004年), 年蒸发量为1 488 mm。样地的成土母质为钙质岩, 土壤类型为山地森林灰褐土, 土壤pH 6.3-7.9, 厚度约1 m。林分起源为天然次生林, 为青海云杉纯林, 林分结构简单, 郁闭度较高, 林下光照不足。青海云杉群落成层现象明显, 可明显划分为乔木层、灌木层、草本层和苔藓层4个层次(
赵丽琼等, 2010 )。其中, 乔木树种组成单一, 除青海云杉外, 只有少量祁连圆柏(
Juniperus przewalskii ); 灌木生长发育不良, 物种较少, 主要有金露梅(
Potentilla fruticosa )、银露梅(
Potentilla glabra )、鬼箭锦鸡儿(
Caragana jubata )、吉拉柳(
Salix gilashanica )、鲜黄小檗(
Berberis diaphana )和高山绣线菊(
Spiraea alpina ); 草本植物物种较丰富, 主要有黑穗薹草(
Carex atrata )、珠芽蓼(
Polygonum viviparum )、针茅(
Stipa capillata )和马蔺(
Iris lactea )等典型高山草甸植物。苔藓层发育良好, 盖度达80%以上, 厚度约10 cm, 主要物种有山羽藓(
Abietinella abietina )、欧灰藓(
Hypnum cupressiforme )和匐灯藓(
Plagiomnium cuspidatum )等(
李效雄等, 2013 )。
1.2 样地设置与调查 参照CTFS (Center for Tropical Forest Science)和中国森林生物多样性监测网络(CForBio)的大样地建立与监测技术规范(
Condit, 1995 )(
http://www.ctfs.si.edu/ ), 于2010-2011年在祁连山国家级自然保护区大野口流域的相对平坦的青海云杉林内建立了一块面积10.2 hm
2 的青海云杉动态监测样地。整个样地呈长方形, 以样地西北角为原点, 南北方向为
x 轴(长340 m), 东西方向为
y 轴(宽300 m), 利用全站仪将整个样地划分成255个20 m × 20 m的样方(附录I), 进一步将每个20 m × 20 m的样方细分成16个5 m × 5 m的小样方, 对小样方内所有胸径(
DBH ) ≥ 1.0 cm的木本个体及其分支进行涂漆, 挂牌标记以便进行永久监测, 测量其相对坐标(
x ,
y )、胸径、冠幅、枝下高, 记录生长状况(存活、倒伏或枯立)等信息。采用全站仪测定每个样方的海拔高度, 并绘制等高线地形图(附录II)。首次调查结果表明, 样地内共记录到
DBH ≥ 1.0 cm的活立木19 927株, 枯死木411株(
李效雄等, 2013 )。
1.3 统计与分析 1.3.1 种群径级划分
虽然径级和龄级之间存在差异, 但在同样的环境条件下, 树木的径级和龄级对生境条件的响应规律具有一致性。青海云杉寿命很长, 在100-140年以后生长才缓慢下来, 少数林木在300-400年仍能持续缓慢生长。已有研究表明, 其胸径和树龄具有明显的线性关系,
y = 0.2133
x - 1.1136,
R 2 = 0.984 9 (
y 为胸径,
x 树龄)。如: 当胸径为5.0、12.5以及22.5 cm时分别大致对应树龄30、64和111年(
杨文娟, 2018 )。因此可以用胸径来反映种群的生活史阶段(
Frost & Rydin, 2000 ;
李立等, 2010 )。从而该研究采用径级结构(胸径大小)代替年龄结构, 分析不同径级青海云杉种群分布格局及其关联性的变化。该研究参照李妍(2013)对青海云杉种群径级结构的划分标准, 同时结合样地内青海云杉胸径的实际情况以及青海云杉的生活史特点, 依据青海云杉胸径大小将其划分为: 径级I (幼树, 1 cm ≤
DBH < 5.0 cm)、径级II (小树, 5.0 cm ≤
DBH < 12.5 cm)、径级III (中树, 12.5 cm ≤
DBH < 22.5 cm)、径级IV (大树,
DBH ≥ 22.5 cm) 4个不同径级。
1.3.2 种群空间分布格局
该研究以
Ripley (1977) 提出的Ripley’s
K (
K (
r ))函数为基础, 采用单变量成对相关函数(
g (
r )), 以样地中每个植株的空间坐标为基础, 分析不同径级青海云杉个体在不同尺度下的分布格局。其中,
K (
r )函数的计算公式(
Besag, 1977 ;
Haase, 1995 )如下:
(1) $K(r)=\frac{A}{{{n}^{2}}}\sum\limits_{i=1}^{n}{\sum\limits_{j=1}^{n}{\frac{{{I}_{r}}({{u}_{ij}})}{{{W}_{ij}}}}} (i\ne j)$ 式中:
r 为空间尺度, 即以目标树为圆点的取样圆半径;
A 为研究样地面积;
n 为样地内某一植物种群的个体总数;
uij 为个体
i 到个体
j 之间的距离;
Wij 为边界效应校正系数, 是指以点
i 为圆心、
uij 为半径的圆落在样地面积
A 中的弧长与其自身圆周长的比值;
I 为指示函数, 当
uij >
r 时,
Ir (
uij ) = 0; 当
uij ≤
r 时,
Ir (
uij ) = 1。
g (
r )由
K 函数衍生而来。与
K (
r )相比,
g (
r )能有效消除小尺度累积效应的缺点, 在评估聚集程度方面的研究中得到了广泛的应用(
Diggle, 2003 )。
g (
r )与
K 函数的关系为:
(2) $g(r)=\frac{1}{\text{2 }\!\!\pi\!\!\text{ }r}\cdot \frac{dK(r)}{dr}$ 式中: 若
g (
r ) = 1, 种群在
r 尺度上呈随机分布; 若
g (
r ) < 1, 种群在
r 尺度呈规则分布; 若
g (
r ) > 1, 种群在
r 尺度呈聚集分布。
1.3.3 空间关联性分析
两个径级之间的空间关联性分析, 实际上是2个径级之间的多变量点格局分析, 也叫多元点格局分析(
张金屯, 2004 )。该研究采用双变量成对相关函数(
g 12 (
r )), 分析不同径级青海云杉个体的空间关联性。其计算公式如下所示:
(3) ${{g}_{12}}(r)=\frac{1}{2\pi r}\cdot \frac{d{{K}_{12}}(r)}{d(r)}$ 式中: 当
g 12 (
r ) = 1时, 2个不同径级个体在
r 尺度下无显著关联性;
g 12 (
r ) < 1时, 2个径级个体在
r 尺度呈显著负关联;
g 12 (
r ) > 1时, 2个不同径级个体在
r 尺度呈显著正关联。
由于
g (
r )的非聚集属性, 利用它进行点格局分析时需利用零模型来观察实际过程和理论过程之间的差异(
Stoyan & Penttinen, 2000 )。祁连山大野口流域青海云杉林样地地势相对平坦, 地形因子对于种群空间格局及其关联性影响较弱, 因此采用完全空间随机零模型作为零模型, 即假设物种的空间分布完全随机, 没有潜在的生物学过程发挥作用, 观察青海云杉种群不同径级个体的空间分布格局及其关联性与匀质泊松分布是否存在差异(
张觅等, 2014 )。空间尺度设定为0-60 m, 步长为1 m。采用蒙特卡罗(Monte-Carlo)随机模拟999次产生置信度为99%的包迹线, 以检验物种分布格局及关联性是否显著。在单变量分析中, 若实际分布的
g (
r )值落在包迹线之内, 则符合零模型, 为随机分布; 若在上包迹线以上, 则呈聚集分布; 若在下包迹线以下, 则为规则分布。在双变量分析中, 若实际分布的
g 12 (
r )值落在包迹线之间, 表示接受零假设, 说明不同径级间个体相互独立, 无空间相关性; 若在上包迹线以上, 则两径级间为正相关; 若落在下包迹线以下, 则两径级间为负相关关系(
Hao et al ., 2017 )。该研究所有统计与分析均在R 4.0.1软件中“spatstat”程序包(
Baddeley & Turner, 2005 )的相关函数中完成。
2 结果和分析 2.1 青海云杉种群及其群落的径级结构 青海云杉种群空间分布及其群落的径级结构如
图1 和
图2 所示, 在10.2 hm
2 青海云杉群落样地内共有
DBH ≥ 1 cm的植株个体19 927株, 隶属于3科3属3种, 密度为1 954株·hm
-2 , 平均胸径为10.1 cm, 径级I、II、III和IV的个体数分别为8 539、5 444、3 493和2 451株, 分别约占总个体数的42.85%、27.32%、17.53%和12.3%。其中, 青海云杉个体19 726株, 密度为1 933株·hm
-2 , 种群平均胸径为10.1 cm, 径级I、II、III和IV的个体数分别为8 489、5 367、3 428和2 442株, 分别约占总个体数的42.6%、26.93%、17.2%和12.25%。其他物种4个径级之和只占到总体个数的1%左右。从个体数量和胸径等数据来看, 青海云杉均占有绝对优势, 因此该样地是以青海云杉为建群种的单优植物群落。此外, 样地内有大量小径级个体储备, 个体数量随径级增大而逐渐减少, 种群的径级结构近似于倒“J”型, 属增长型种群, 种群更新良好。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1祁连山大野口流域10.2 hm2 森林动态监测样地青海云杉种群不同径级个体的空间分布图。I, 径级I (幼树); II, 径级II (小树); III, 径级III (中树); IV, 径级IV (大树)。 Fig. 1Spatial distribution of Picea crassifolia individuals with different diameter at breast height (DBH ) in the forest dynamics plot of 10.2 hm2 in Dayekou Basin of Qilian Mountains, northwestern China. I, diameter class I (saplings); II, diameter class II (small trees); III, diameter class III (medium trees); IV, diameter class IV (big trees). 图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2祁连山大野口流域10.2 hm2 森林动态监测样地内各物种的径级结构。按胸径划分个体大小。I, 1-5 cm; II, 5-12.5 cm; III, 12.5-22.5 cm; IV, ≥22.5 cm。 Fig. 2Size structure of all individuals in the forest dynamics plot of 10.2 hm2 in Dayekou Basin of Qilian Mountains, northwestern China. The size of individuals was estimated by their diameter at breast height (DBH ). I, 1-5 cm; II, 5-12.5 cm; III, 12.5-22.5 cm; IV, ≥22.5 cm. 2.2 不同径级青海云杉种群的空间分布格局 g (
r )函数分析结果(
图3 )表明, 随尺度变化, 4个不同径级青海云杉种群的空间分布格局有所差异。样地中处于径级I的青海云杉幼苗在0-46 m尺度上呈聚集分布, 在46-60 m的尺度上逐渐趋于随机分布; 小树和中树在0-60 m的尺度上均呈现聚集分布; 大树在0-2 m尺度上呈规则分布, 2-8 m的尺度上呈随机分布, 8-15 m的尺度上呈聚集分布, 在15-60 m的尺度上呈随机分布。总体来看, 不同径级的青海云杉个体在较小尺度内均表现为聚集分布, 随着空间尺度的增加, 呈现由聚集分布趋向于随机分布。此外, 较小径级的个体常呈聚集分布, 而随着径级增大, 呈现由聚集分布趋向于随机分布。从青海云杉群落及其种群的散点分布图(
图1 )也可以看出, 不同径级的青海云杉在样地内多呈聚集分布或随机分布。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3祁连山大野口流域10.2 hm2 森林动态监测样地内青海云杉种群不同径级个体间的空间格局。A, 径级I (幼树)。B, 径级II (小树)。C, 径级III (中树)。D, 径级IV (大树)。r , 以目标树为圆点的取样圆半径; ?g obs (r ), 单变量成对相关函数g (r )的函数值; g theo (r ), g (r )函数的期望值; 灰色阴影部分为99%的置信区间。 Fig. 3Spatial distribution pattern of Picea crassifolia individuals with different diameter at breast height (DBH ) in the forest dynamics plot in Dayekou Basin of Qilian Mountains, northwestern China. A, DBH class I (saplings). B, DBH class II (small trees). C, DBH class III (medium trees). D, DBH class IV (big trees). r, the radius of the sampling circle with the target tree as the centroid; ?g obs (r ), the function value of the pair-correlation function (g (r )); g theo (r ), the expected value of the g (r ) function; the gray shaded part is the 99% confidence interval. 2.3 青海云杉种群不同径级间的空间关联 由青海云杉种群不同径级间的空间关联图(
图4 )可知, 青海云杉种群4个不同径级个体间的空间关联性格局各异。样地中处于幼树与小树的青海云杉个体在0-19 m的尺度上呈正关联, 19-43 m表现为微弱的正关联, 大于43 m后无关联; 幼树与中树在0-1 m呈正关联, 1-2 m无关联, 2-34 m为负关联, 大于34 m后无关联; 幼树和大树在0-5 m内呈负关联, 5-7 m无关联, 7-8 m呈正关联, 9-12 m又呈现无关联, 大于12 m后呈正关联且随着尺度增大关联性减弱; 小树与中树在0-60 m的尺度上都呈正关联, 且随着尺度的增加关联性减弱; 小树和大树在0-10 m上呈负关联, 10-28 m上无关联, 大于28 m后呈负关联; 中树和大树在0-4 m呈负关联, 在4-25 m无关联, 大于25 m后基本呈负关联。从整体上来看, 在小尺度(<4 m)范围内大树个体与其他径级个体都呈负关联, 随着尺度的增大, 大树与幼树表现出正关联且关联性随尺度的增大而减弱, 而与小树、中树则表现为无关联或者微弱的负关联; 在所有尺度内, 径级相近的个体空间关联性呈正关联或无关联, 随着个体径级间差距增大, 它们之间的空间关联性转变为无关联或呈负关联。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4祁连山大野口流域10.2 hm2 森林动态监测样地内青海云杉种群不同径级个体间的空间关联性。A, 幼树与小树。B, 幼树与中树。C, 幼树与大树。D, 小树与中树。E, 小树与大树。F, 中树与大树。r, 以目标树为圆点的取样圆半径; ?g obs (r ), 双变量成对相关函数g 12 (r )的函数值; g theo (r ), g 12 (r )函数的期望值; 灰色阴影部分为99%的置信区间; 1-4表示4个径级。 Fig. 4Spatial association of Picea crassifolia individuals with different diameter at breast height in the forest dynamics plot in Dayekou Basin of Qilian Mountains, northwestern China. A, Saplings and small trees. B, Saplings and medium trees. C, Saplings and big trees. D, Small trees and medium trees. E, Small trees and big trees. F, Medium trees and big trees. r, the radius of the sampling circle with the target tree as the dot; ?g obs (r ), the function value of the bivariate pair-correlation function (g 12 (r )); g theo (r ), the theoretical value of the g 12 (r ) function; the gray shaded part is the 99% confidence interval; 1-4 indicate four different diameter classes. 3 讨论 3.1 青海云杉种群的径级结构 植物种群的径级结构是指不同径级组的个体在种群内所占比例和配置情况(
田润炜等, 2016 ), 是种群的生殖特性与外界环境相互作用的结果, 在一定程度上体现了种群的径级结构特征和生长发育状况, 进而可以预测种群和群落的动态发展趋势(
涂洪润等, 2019 )。本研究发现, 样地内青海云杉种群的幼苗和小树个体储备较为丰富, 其种群径级结构呈倒“J”型分布, 这说明该种群属增长型种群, 种群天然更新能力良好, 只要未来不遭受强烈干扰, 种群数量会保持逐渐增长。这与
李效雄等(2012) 和
张华等(2016) 分别对祁连山大野口流域和天老池小流域的青海云杉种群结构的研究结果一致。这可能是由青海云杉种群的生物学特性与环境因素共同作用的结果。已有研究表明, 森林更新过程中最重要和最敏感的阶段是幼苗阶段(
李晓笑等, 2011 ;
赵阳等, 2018 ), 林内有限的资源和空间往往成为种群更新的重要限制因素(
项小燕等, 2016 )。青海云杉种群的幼苗和小树个体数量多主要是由其生殖特性决定, 其种群可以产生大量高质量种子, 自然萌发形成的幼苗大都集中分布在林窗下, 形成大量小径级个体, 占据有利资源, 这是其种群在群落中持续处于优势地位的主要原因。随着小径级个体的生长, 它们对光照、水分、土壤养分等环境资源和空间需求随之增加, 由于环境资源和空间有限, 种内和种间竞争加剧, 出现强烈的自疏和他疏现象, 竞争能力弱的幼苗和小树在竞争中会大量死亡, 只有少量小径级个体能够成功转化为大径级个体, 造成样地内仅存在少量的中树和大树, 这成为该青海云杉种群更新和发展的“瓶颈” (
张婕等, 2014 ;
白卫国等, 2017 ;
于世川等, 2017 )。因此, 要实现种群数量的稳定增长, 必须加强对种群结构的保护, 尤其要重视对幼苗和小树及其生存环境的保护和改善。
3.2 青海云杉种群不同径级个体的空间分布格局 森林的空间格局与空间尺度密切相关, 物种的空间分布格局随着尺度的变化而变化(
Condit et al ., 2000 ;
杨华等, 2014 )。一般来说, 小尺度上的物种分布格局主要受种群自身生物学特性(如种内种间竞争、种子扩散限制和幼苗补充过程等)影响, 而大尺度上物种的空间分布则主要受生境异质性(如光照、土壤养分、水分和温度等)影响(
Shen et al ., 2013 )。本研究表明, 青海云杉种群不同径级的青海云杉个体在较小尺度内均表现为聚集分布, 随着空间尺度的增加, 呈现由聚集分布到随机分布的变化趋势。这一结果与国内外大多数有关种群空间格局的研究结果一致(
刘建泉, 2004 ;
Guo et al ., 2013 ;
张觅等, 2014 ;
张华等, 2016 )。这是因为同一种群的不同个体对生境条件的需求基本上是一致的(
张金屯和孟东平, 2004 ), 且聚集分布有利于种群个体相互庇护, 从而抵御外界不利环境, 以提高存活率(
Diggle, 1981 )。此外, 生境的异质性容易引起种群聚集生长, 同时由于植株对光照、水分、养分等资源的竞争较大(
李海东等, 2011 ), 导致聚集强度随着空间尺度的增大而呈现下降趋势, 表现为在较小尺度上植株倾向于聚集分布逐渐过渡为在较大尺度上的随机分布。
通常认为, 由于物种的生态学和生物学特性、种子扩散限制和生境异质性等机制的影响, 较小径级的物种常呈聚集分布, 尤其在小尺度上更加明显(
祝燕等, 2011 )。而随着径级增大, 种内和种间为争夺有限资源和空间的竞争也大大增强, 常呈随机或规则分布, 聚集强度也随径级的增加而降低(
Condit et al ., 2000 )。本研究结果也符合这一规律。样地内的青海云杉种群的幼树、小树和中树这类低龄级个体多呈聚集分布, 这一方面可能是由于青海云杉的种子具有种翅, 主要依靠风力传播, 使得大多数种子散落在母树周围一定范围内且随着距离的增大其种子传播的数量逐渐下降, 这些生物学特性使其形成聚集分布的空间分布特征。实地调查过程中也发现, 青海云杉幼苗大都集中分布于林窗下, 在阴蔽的林冠下, 出现幼苗和小树死亡现象, 在地被层特别是苔藓层较厚的地方, 幼苗较少。另一方面, 低龄级种群个体较小, 处在群落的下层, 对光、热、水和土壤养分等生态因子的竞争能力较弱, 聚集可以提高其竞争能力以及抵御外界不良环境, 提高成活率。此外, 小生境的异质作用导致幼树与小树分布不均匀, 如局部地形、土壤和植被等环境因子的差异, 以及采伐、自然灾害等形成的林窗, 都为低龄级个体的大量繁殖和生长创造了良好的条件(
张华等, 2016 )。随着种群年龄的增长, 个体对光照、水分、营养等资源和空间的需求不断增加, 种内和种间竞争加剧而出现强烈的自疏和他疏现象, 导致大量林木死亡, 种群密度下降, 聚集强度逐渐减弱, 种群由聚集分布向随机分布或规则分布转变。此外, 大树主要呈随机分布, 这可能是因为大树位于林冠层, 高度较高, 树冠基本定型, 根系也伸展到一定程度, 种内竞争加剧, 自疏效应使得种群的聚集分布格局被打破, 由聚集分布向随机分布转变。
3.3 青海云杉种群不同径级个体的空间关联性分析 种群内不同径级个体的空间关联性描述了某一特定的时间内植物种群组成个体之间的空间分布关系及功能关系(
Shields et al ., 2014 ;
王慧杰等, 2016 )。这种关系是种群内不同径级个体间、种群与环境长期相互作用的结果, 通过它可以揭示种群的现状并预测种群发展趋势(
涂洪润等, 2019 )。空间正关联反映了种群内部正向(互惠)的生态关系, 空间负关联意味着种群内部负向(互斥)的生态关系, 空间无关联则反映了种群内部无明确的生态关系(
张金屯, 1998 )。该研究发现小尺度(<4 m)下大树个体与其他径级个体都呈负关联, 随着尺度的增大, 大树与幼树表现出正关联且关联性随尺度的增大而减弱, 而与小树、中树则表现为无关联或者微弱的负关联。小尺度下大树与其他径级个体都呈负关联, 表明在一定范围内其他径级个体都受到大树的排斥, 这可能是因为大树个体对土壤水分、光等资源的需求量更大, 竞争能力更强, 从而使得其周围其他径级个体的分布密度非常小(
张华等, 2016 )。也有研究表明青海云杉凋落物对种子的萌发和幼苗的生长有自毒作用(
王丽红等, 2017 ), 而大树周围凋落物一般也较为丰富, 导致大树周围种子和幼苗的生长受到抑制。实际调查中也发现在大树周围尤其是林冠下其他径级个体数量很少。大树个体和幼树个体在较大尺度上表现出正关联且随着尺度的增大, 关联性逐渐减弱, 这很可能是由种子扩散限制所引起的(
梁爽等, 2014 ), 大树产生的种子数量较多, 幼树的分布在一定程度上能反映种子的分布, 大树和幼树的关联性变化特点与青海云杉种子主要通过风力传播的特点具有一致性。此外, 在较大尺度下, 大树个体与小树、中树个体都表现为无关联或者微弱的负关联, 一方面表明随着尺度的增大, 大树对小树和中树的影响逐渐降低, 竞争趋于和缓, 另一方面随着径级的增大, 大树与其他径级个体关联性也发生变化, 说明不同径级青海云杉空间关联性随着发育阶段而变化, 这可能是由种群自身的生物生态学特性、种内种间关系以及环境条件共同作用的结果。
该研究还发现径级相近的个体空间关联性呈正关联或无关联, 随着个体径级间差距增大, 它们之间的空间关联性转变为无关联或负关联, 这说明同一物种不同径级的个体之间同时存在竞争、依赖以及无明确的生态关系, 这是由于物种具有一致的生境偏好所造成的(
杨洪晓等, 2006 )。径级相近个体的空间关联性呈正关联, 表明它们相伴出现, 对生境的选择具有一致性, 在空间分布上是相互依存的。这可能是由于它们生长发育阶段相近, 对环境资源的需求具有相似性和生态位存在重叠, 但小径级个体相对幼小, 对光照、水分和土壤养分等资源的竞争能力相对较弱, 需要借助高大植株的庇护, 才得以提高存活率, 而较大径级的植株个体可以改善样地的小生境, 使得邻近径级的个体更适宜生存, 彼此共享资源(
涂洪润等, 2019 )。同时, 由于物种专一性的资源分布不均, 种内竞争具有非对称性的特点, 也就是说当径级相近时, 不同径级间关系较协调(
Getzin et al ., 2008 )。而无关联则说明个体间互不干扰, 个体在前期发育过程中已经占据适宜的生态位, 与种内其他径级个体间的关联性不高, 能各自生长发育(
涂洪润等, 2019 )。随着个体间径级差距增加, 它们之间的关联性转变成无关联甚至负关联, 说明随着个体间径级差距的加大, 它们的空间正关联关系逐渐减弱, 当个体间差异较大时, 大径级个体在竞争中处于优势地位, 可能导致小径级个体的生长发育迟滞或者死亡, 使其周围小径级个体很少(
Kenkel, 1988 ), 这也间接证实了随着个体大小的生长变化, 青海云杉种群的空间分布存在一个从聚集分布到随机分布的自疏过程。
此外, 不同径级的青海云杉个体在较小尺度内均表现为聚集分布, 随着空间尺度的增加, 呈现由聚集分布趋向于随机分布, 这说明祁连山大野口流域青海云杉种群的分布格局及其关联性随空间尺度及发育阶段而变化, 且种群的空间分布格局与其空间关联性存在一定的关系。不同径级青海云杉种群的空间分布格局及其关联性的变化, 使得青海云杉种群能充分利用样地内有限的资源, 有利于青海云杉种群的生存和稳定发展, 同时也对整个青海云杉群落的生存和发展起到了促进作用。
4 结论 综上所述, 该研究首次基于10.2 hm
2 青海云杉动态监测样地调查资料, 分析了祁连山大野口流域青海云杉种群不同径级个体的空间格局及其关联性。研究发现, 祁连山大野口流域青海云杉种群的分布格局及其关联性随空间尺度及发育阶段而变化, 这是由种群自身的生物生态学特性、种内种间关系以及环境条件综合作用的结果, 且不同空间尺度及不同发育阶段青海云杉种群分布格局的形成可能存在不同的机制。该研究结果进一步加深了我们对祁连山青海云杉种群空间分布格局及其关联的认识, 进而有利于更好地理解其潜在生态学过程或影响机理。但仅仅靠物种空间格局及其关联性的分析还无法对许多问题给予合理的解释, 因为不一样的生态过程可以产生相同的格局, 而相同的格局也可能是多个不同的生态学过程共同作用的结果。因此, 未来应对固定监测样地内的青海云杉个体进行长期动态监测, 基于长时间序列的群落结构和环境因子数据, 进一步探讨扩散限制、竞争排除和生境过滤等因素对青海云杉林物种共存的影响。
致谢 感谢中国科学院沙漠与沙漠化重点实验室开放基金(KLDD-2019)资助。
附录I 祁连山大野口流域青海云杉10.2 hm
2 森林动态监测样地网格示意图
Supplement I Grid of the 10.2 hm
2 forest dynamics plot of
Picea crassifolia population in Dayekou basin of Qilian Mountains, northwestern China
https://www.plant-ecology.com/fileup/1005-264X/PDF/cjpe.2020.0177-S1.pdf 附录II 祁连山大野口流域青海云杉10.2 hm
2 森林动态监测样地地形图
Supplement II Topography map of the 10.2 hm
2 forest dynamics plot of
Picea crassifolia population in Dayekou basin of Qilian Mountains, northwestern China
https://www.plant-ecology.com/fileup/1005-264X/PDF/cjpe.2020.0177-S2.pdf [1] Baddeley A Turner R 2005 ). spatstat: an R package for analyzing spatial point patternsJournal of Statistical Software 1 -42 . [本文引用: 1] [2] Bai WG Xu HL Wu QB Li TH Li JB Liao GY 2017 ). Ecological study of Keteleeria davidiana populations of Yachang forest farm in Guangxi Zhuang Autonomous RegionJournal of Nanjing Forestry University (Natural Sciences Edition )71 -76 . [本文引用: 1] [ 白卫国 , 徐惠玲 , 吴庆标 , 李铁华 , 李加博 , 廖国勇 (2017 ). 广西雅长林区铁坚油杉种群结构分布研究南京林业大学学报(自然科学版) , 41, 71 -76 .] [本文引用: 1] [3] Besag JE 1977 ). Contribution to the discussion of Dr Ripley’s paperJournal of the Royal Statistical Society B 193 -195 . [本文引用: 1] [4] Condit R 1995 ). Research in large, long-term tropical forest plotsTrends in Ecology & Evolution 18 -22 . URL PMID:
21236939 [本文引用: 1] [5] Condit R Ashton PS Baker P Bunyavejchewin S Gunatilleke S Gunatilleke N Hubbell SP Foster RB Itoh A LaFrankie JV Lee HS Losos E Manokaran N Sukumar R Yamakura T 2000 ). Spatial patterns in the distribution of tropical tree speciesScience 1414 -1418 . URL PMID:
10827950 [本文引用: 2] [6] Diggle PJ 1981 ). Binary mosaics and the spatial pattern of heatherBiometrics 531 -539 . [本文引用: 1] [7] Diggle PJ 2003 ). Statistical Analysis of Spatial Point Patterns [本文引用: 1] [8] Frost I Rydin H 2000 ). Spatial pattern and size distribution of the animal-dispersed tree Quercus robur in two spruce- dominated forestsécoscience 38 -44 . [本文引用: 1] [9] Gao LL Gou XH Deng Y Liu WH Yang MX Zhao ZQ 2013 ). Climate-growth analysis of Qilian juniper across an altitudinal gradient in the central Qilian Mountains, northwest ChinaTrees 379 -388 . [本文引用: 1] [10] Getzin S Wiegand T Wiegand K He F 2008 ). Heterogeneity influences spatial patterns and demographics in forest standsJournal of Ecology 807 -820 . [本文引用: 1] [11] Guo YL Lu JM Franklin SB Wang QG Xu YZ Zhang KH Bao DC Qiao XJ Huang HD Lu ZJ Jiang MX 2013 ). Spatial distribution of tree species in a species-rich subtropical mountain forest in central ChinaCanadian Journal of Forest Research 826 -835 . [本文引用: 2] [12] Haase P 1995 ). Spatial pattern analysis in ecology based on Ripley’s K-function: introduction and methods of edge correctionJournal of Vegetation Science 575 -582 . [本文引用: 1] [13] Hao HM Huang Z Lu R Jia C Liu Y Liu BR Wu GL 2017 ). Patches structure succession based on spatial point pattern features in semi-arid ecosystems of the water-wind erosion crisscross regionGlobal Ecology and Conservation 158 -165 . [本文引用: 1] [14] He FL Legendre P Lafrankie JV 1997 ). Distribution patterns of tree species in a Malaysian tropical rain forestJournal of Vegetation Science 105 -114 . [本文引用: 1] [15] He FL Xu XY Yu QS Liu HJ He FY Ma JM Jin HX 2016 ). A study on the comparison of community structural characteristics and species diversity between plantations and natural forests of Picea crassifolia in Qilian MountainsJournal of Northwest Forestry University 1 -7 . [本文引用: 1] [ 何芳兰 , 徐先英 , 尉秋实 , 刘虎俊 , 贺访印 , 马俊梅 , 金红喜 (2016 ). 祁连山青海云杉人工林与天然林群落结构特征及物种多样性比较研究西北林学院学报 , 31, 1 -7 .] [本文引用: 1] [16] Hou JH Mi XC Liu CR Ma KP 2004 ). Spatial patterns and associations in a Quercus -Betula forest in Northern ChinaJournal of Vegetation Science 407 -414 . [本文引用: 1] [17] Kenkel NC 1988 ). Pattern of self-thinning in jack pine: testing the random mortality hypothesisEcology 1017 -1024 . [本文引用: 1] [18] Li HD Shen WS Fang Y Yan SG Zhang H Zhao W 2011 ). Point pattern analysis of several psammophyte populations in the riparian ecotone in the middle reaches of Yarlung Zangbo River of Tibet, ChinaChinese Journal of Plant Ecology 834 -843 . DOI:
10.3724/SP.J.1258.2011.00834 URL [本文引用: 1] AimsArtemisia wellbyi, A. younghusbandii and Sophora moorcroftiana are the major psammophyte populations on the aeolian desertified land in the riparian ecotone of the middle reaches of the Yarlung Zangbo River. Our objectives were to determine how spatial scales are related to population patterns and how species spatially adapt to semi-arid conditions in the riparian ecotone, as well as to provide theoretical foundation for vegetation restoration on sand dunes in the study area.Methods We selected four representative plant communities under different habitat conditions in the riparian ecotone, measured the position, height and crown diameter of each individual in a 20 m × 30 m plot and studied the population structure, spatial distribution and association of these psammophyte populations using point pattern analysis.Important findings The population structure of A. wellbyi on semi-exposed sandy gravel land and semi-fixed sandy land is that of a growing population, while A. younghusbandii and S. moorcroftiana on fixed sandy land both have the structure of a declining population. Artemisia wellbyi and A. younghusbandii both have a clumped distribution at different scales, while the distribution pattern of S. moorcroftiana changed from clumped to random to clumped to random as the scale increased. Spatial distribution is mainly determined by the smaller size classes, while fluctuations or changes of population distribution are mainly determined by the larger size classes. There is a scale effect among the studied populations, and positive spatial association occurs mainly at certain scales. Spatial association is often affected by plant size and environmental heterogeneity. The spatial association of A. wellbyi and S. moorcroftiana on semi-exposed sandy gravel land or semi-fixed sandy land changes from independent to positive to independent to positive. The spatial association of A. wellbyi and Oxytropis sericopetala on semi-exposed sandy gravel land changes from positive to independent to positive to independent to negative, and that of A. younghusbandii and S. moorcroftiana changes from positive to independent. The degree of aggregation weakens with increasing plant size. The smaller size class has a clumped distribution, and the larger has a random distribution or random and clumped. The positive spatial association between different size classes of the same species appears to weaken when the difference of plant size becomes greater and even changes into negative spatial association, while the spatial association of various size classes at a smaller scale often is positive or independent.]]>
[ 李海东 , 沈渭寿 , 方颖 , 燕守广 , 张慧 , 赵卫 (2011 ). 雅鲁藏布江中游河岸带几种主要沙生植物种群点格局分析植物生态学报 , 35, 834 -843 .] [本文引用: 1] [19] Li L Chen JH Ren HB Mi XC Yu MJ Yang B 2010 ). Spatial patterns of Castanopsis eyrei and Schima superba in mid-subtropical broad-leaved evergreen forest in Gutianshan National Reserve, ChinaChinese Journal of Plant Ecology 241 -252 . DOI:
10.3773/j.issn.1005-264x.2010.03.001 URL [本文引用: 2] Aims Many mechanisms of forest patterns have been examined in tropical rain forest; however, there are only a handful of similar studies on subtropical broad-leaved evergreen forest. Our objective is to analyze the main mechanisms of distribution pattern of Castanopsis eyrei and Schima superba in GTS (Gutianshan) forest plot.Methods Based on a stem map of a 24 hm2 permanent plot in the middle subtropical broad-leaved evergreen forest at Gutianshan National Reserve, we analyzed the distribution pattern of the dominant species C. eyrei and S.superba in saplings, juvenile trees, adult trees, large trees and old trees using spatial point pattern analysis. We also examined spatial associations among different growth stages.Important findingsCastanopsis eyrei and S. superba had a clustered distribution across a range of scales (0–100 m). Saplings, juvenile trees and adult trees tended to be more clumped than big trees. Old trees tended to besomewhat more clumped than big trees. Our results suggested that the two species have obvious habitat preferences at larger scales. For C. eyrei, saplings were significantly positively associated with juvenile trees at scales≤ 100 m, as were saplings and adult trees, juvenile trees and adult trees, adult trees and big trees. Generally there was no correlation between big trees and old trees at scales ≤ 10 m, whereas their association tended to be positive at larger scales (10–100 m). Generally there were negative or no correlations between other size classes. For S. superba, the saplings were significantly positively associated with juvenile trees at scales ≤ 100 m, as were saplings and adult trees, juvenile trees and adult trees. Generally there were negative or no correlations between juvenile trees and old trees, adult trees and old trees, big trees and old trees, saplings and big trees, juvenile trees and big trees, adult trees and big trees. Generally there was negative correlation between saplings and old trees at scales ≤ 20 m, whereas their association tended to be positively at larger scales (25–100 m). These results suggested that dominant species facilitated coexistence of other species through emptying space for colonization of other species, which is probably attributed to density dependence or the Janzen-Connell effect. Both processes depended on species in intensity and acting time.]]>
[ 李立 , 陈建华 , 任海保 , 米湘成 , 于明坚 , 杨波 (2010 ). 古田山常绿阔叶林优势树种甜槠和木荷的空间格局分析植物生态学报 , 34, 241 -252 .] [本文引用: 2] [20] Li XX Liu XD Zhao WJ 2013 ). Community structure of a dynamical plot of Picea crassifolia forest in Qilian Mountains, ChinaJournal of Desert Research 94 -100 . DOI:
10.7522/j.issn.1000-694X.2013.00013 URL [本文引用: 2] According to Chinese forest ecosystem dynamic monitoring plot setting method and international forest ecology large plot construction technical standards, a 10.2 hm2 Picea crassifolia forest dynamic monitoring plot was established in the Qilian Mountains Nature Reserve in 2010 and 2011. The 19 927 woody plants with DBH (diameter at breast height) greater than 1 cm at the plot were surveyed based on positioning survey method, and the community characteristic was analyzed. (1) The layer structure difference is obvious for the P. crassifolia community, and it can be divided into tree layer, shrub layer, herb layer and bryophyta layer. The tree layer is the main layer of the community, and its vertical structure is more complex than the shrub layer and herb layer, and the bryophyta layer is also well developed. (2) The diameter structure of the P. crassifolia community is obviously an "inverted-J" form, the DBH of the individuals is concentrated in 1-5 cm and 21-25 cm, which account for 91.42% of the total individuals, so regeneration of the P. crassifolia population is good. (3) The height structure of the P. crassifolia individuals with DBH greater than 1 cm is single-peak form, and the height of the main individuals is less than 6 m, which account for more than 60.00% of the total individuals. So the height of the P. crassifolia individuals is relatively low, and young trees are much, mid trees hold a certain proportion, and big trees are little. There is a significantly quadratic function relation between the tree height and the DBH of the P. crassifolia (P<0.05). (4) The spatial distribution of the P. crassifolia individuals displays obviously a clustering distribution pattern. The point pattern analysis of small trees, mid trees and big trees of the P. crassifolia community shows that the clump intensity of the P. crassifolia population reduces with the increasing of the population age, and the congregation distribution turns into random distribution, which displays obviously a diffusion trend. There is no obvious spatial heterogeneity for spatial distribution of the P. crassifolia individuals, and 2 534 big trees of the P. crassifolia community are randomly distributed in the large plot, showing no significant human interference.
[ 李效雄 , 刘贤德 , 赵维俊 (2013 ). 祁连山青海云杉林动态监测样地群落特征中国沙漠 , 33, 94 -100 .] [本文引用: 2] [21] Li XX Wang QC Cui GF Yang CD 2011 ). Structure and dynamic characteristics of a wild population of endangered Abies fanjingshanensisActa Botanica Boreali-Occidentalia Sinica 1479 -1486 . [本文引用: 1] [ 李晓笑 , 王清春 , 崔国发 , 杨传东 (2011 ). 濒危植物梵净山冷杉野生种群结构及动态特征西北植物学报 , 31, 1479 -1486 .] [本文引用: 1] [22] Li Y 2013 ). Study on Picea crassifolia Dynamics and Regeneration in the Eastern Slope of Helan Mountain 13 -14 . [ 李妍 (2013 ). 贺兰山东坡青海云杉种群动态及更新特征研究 . 硕士学位论文, 西北农林科技大学, 陕西杨凌 . 13 -14 .] [23] Liang S Xu H Lin JY Li YD Lin MX 2014 ). Spatial distribution pattern of the dominant species Gironniera subaequalis in tropical montane rainforest of Jianfengling, Hainan Island, ChinaChinese Journal of Plant Ecology 1273 -1282 . [本文引用: 1] [ 梁爽 , 许涵 , 林家怡 , 李意德 , 林明献 (2014 ). 尖峰岭热带山地雨林优势树种白颜树空间分布格局植物生态学报 , 38, 1273 -1282 .] [本文引用: 1] [24] Lin Y Ren JY Yue M 2008 ). Population structure and spatial analysis of Betula albo-sinensis at Taibai Mountain, northwestern ChinaJournal of Plant Ecology (Chinese Version )1335 -1345 . [本文引用: 1] [ 林玥 , 任坚毅 , 岳明 (2008 ). 太白山红桦种群结构与空间分析植物生态学报 , 32, 1335 -1345 .] [本文引用: 1] [25] Liu JQ 2004 ). Spatial pattern of Picea crassifolia population in Qilianshan nature conservationJournal of Northwest Forestry University 152 -155 . [本文引用: 1] [ 刘建泉 (2004 ). 祁连山保护区青海云杉种群分布格局的研究西北林学院学报 , 19, 152 -155 .] [本文引用: 1] [26] Liu PC Wang WD Bai ZQ Guo ZJ Ren W Huang JH Xu Y Yao J Ding Y Zang RG 2020 ). Competition and facilitation co-regulate the spatial patterns of boreal tree species in Kanas of Xinjiang, northwest ChinaForest Ecology and Management 118167 . DOI: 10.1016/j.foreco. 2020.118167 . [本文引用: 1] [27] McDonald RI Peet RK Urban DL 2003 ). Spatial pattern of Quercus regeneration limitation and Acer rubrum invasion in a Piedmont forestJournal of Vegetation Science 441 -450 . DOI:
10.1658/1100-9233(2003)014[0441:SPOQRL]2.0.CO;2 URL [本文引用: 1] [28] Nathan R 2006 ). Long-distance dispersal of plantsScience 786 -788 . URL PMID:
16902126 [本文引用: 1] [29] Ripley BD 1976 ). The second-order analysis of stationary point processesJournal of Applied Probability 255 -266 . [30] Schleicher J Wiegand K Ward D 2011 ). Changes of woody plant interaction and spatial distribution between rocky and sandy soil areas in a semi-arid savanna, South AfricaJournal of Arid Environments 270 -278 . DOI:
10.1016/j.jaridenv.2010.10.003 URL [本文引用: 1] [31] Shen GC He FL Waagepetersen R Sun IF Hao ZQ Chen ZS Yu MJ 2013 ). Quantifying effects of habitat heterogeneity and other clustering processes on spatial distributions of tree speciesEcology 2436 -2443 . URL PMID:
24400495 [本文引用: 1] [32] Shen ZQ Hua M Dan Q Lu J Fang JP 2016 ). Spatial pattern analysis and associations of Quercus aquifolioides population at different growth stages in Southeast Tibet, ChinaChinese Journal of Applied Ecology 387 -394 . URL PMID:
27396109 [本文引用: 1] [ 沈志强 , 华敏 , 丹曲 , 卢杰 , 方江平 (2016 ). 藏东南川滇高山栎种群不同生长阶段的空间格局与关联性应用生态学报 , 27, 387 -394 .] PMID:
27396109 [本文引用: 1] [33] Shields JM Jenkins MA Saunders MR Zhang H Jenkins LH Parks AM 2014 ). Age distribution and spatial patterning of an invasive shrub in secondary hardwood forestsForest Science 830 -840 . [本文引用: 1] [34] Stoyan D Penttinen A 2000 ). Recent applications of point process methods in forestry statisticsStatistical Science 61 -78 . [本文引用: 1] [35] Tian FX Zhao CY Feng ZD Peng SZ Peng HH 2012 ). Eco-hydrological effects of Qinghai spruce (Picea crassifolia ) canopy and its influence factors in the Qilian MountainsActa Ecologica Sinica 62 -72 . [本文引用: 1] [ 田风霞 , 赵传燕 , 冯兆东 , 彭守璋 , 彭焕华 (2012 ). 祁连山青海云杉林冠生态水文效应及其影响因素生态学报 , 32, 62 -72 .] [本文引用: 1] [36] Tian RW Cai XB Liu LY Jiang XH Tursun B Abrahman M 2016 ). Characteristics of the age structure and dynamics of Malus sieversii populationActa Botanica Boreali- Occidentalia Sinica 811 -817 . [本文引用: 1] [ 田润炜 , 蔡新斌 , 刘丽燕 , 江晓珩 , 布早拉木?吐尔逊 , 买尔燕古丽?阿不都热合曼 (2016 ). 新疆野苹果种群年龄结构特征与动态分析西北植物学报 , 36, 811 -817 .] [本文引用: 1] [37] Tu HR Li JF Liu RH Liang SC Lan ZN Zhang XY Kang XD Jiang Y 2019 ). Spatial distribution patterns and association of Loropetalum chinense population in karst hills of Guilin, Southwest ChinaChinese Journal of Applied Ecology 2621 -2630 . DOI:
10.13287/j.1001-9332.201908.016 URL PMID:
31418186 [本文引用: 5] The spatial distribution and association of populations can reflect succession patterns and its adaptation strategies to the change of environmental factors, with important significance for vegetation restoration, reconstruction, and biodiversity conservation. We used the point-pattern analytical method to examine the spatial distribution and its association of the Loropetalum chinense population in karst hills of Guilin, China, based on field surveys. On the basis of Ripley K function, we used the pair-correlation function statistic derived to explore difference in the distribution patterns.We used Ripley L function to examine the spatial associations among the three diameter classes. The results showed that the diameter class structure of the population exhibited an irregular inverted
[ 涂洪润 , 李娇凤 , 刘润红 , 梁士楚 , 兰泽南 , 章欣仪 , 康馨丹 , 姜勇 (2019 ). 桂林岩溶石山檵木种群空间格局及其关联性应用生态学报 , 30, 2621 -2630 .] PMID:
31418186 [本文引用: 5] [38] Wang BY Yu SX 2005 ). Multi-scale analyses of population distribution patternsActa Phytoecologica Sinica 235 -241 . [本文引用: 1] [ 王本洋 , 余世孝 (2005 ). 种群分布格局的多尺度分析植物生态学报 , 29, 235 -241 .] [本文引用: 1] [39] Wang HJ Chang SL Zhang YT Xie J He P Song CC Sun XJ 2016 ). Density-dependent effects in Picea schrenkiana forests in Tianshan MountainsBiodiversity Science 252 -261 . DOI:
10.17520/biods.2015282 URL [本文引用: 1] [ 王慧杰 , 常顺利 , 张毓涛 , 谢锦 , 何平 , 宋成程 , 孙雪娇 (2016 ). 天山雪岭云杉森林群落的密度制约效应生物多样性 , 24, 252 -261 .] [本文引用: 1] [40] Wang LH Zhao DP Chen GL 2017 ). Autotoxicity of litters from Picea crassifolia on its seed germination and seedling growthMolecular Plant Breeding 4275 -4281 . [本文引用: 1] [ 王丽红 , 赵东平 , 陈贵林 (2017 ). 青海云杉凋落物对种子萌发和幼苗生长的自毒作用分子植物育种 , 15, 4275 -4281 .] [本文引用: 1] [41] Wang XG Ye J Li BH Zhang J Lin F Hao ZQ 2010 ). Spatial distributions of species in an old-growth temperate forest, northeastern ChinaCanadian Journal of Forest Research 1011 -1019 . [本文引用: 1] [42] Wiegand T Moloney K 2004 ). Rings, circles, and null-models for point pattern analysis in ecologyOikos 209 -229 . [本文引用: 2] [43] Xiang XY Wu GL Duan RY Wang ZG Zhang ZX Wang GY Zhang XP 2016 ). Studies on population structure and dynamics of Pinus dabeshanensisResources and Environment in the Yangtze Basin 55 -62 . [本文引用: 1] [ 项小燕 , 吴甘霖 , 段仁燕 , 王志高 , 张中信 , 王广艳 , 张小平 (2016 ). 大别山五针松种群结构及动态研究长江流域资源与环境 , 25, 55 -62 .] [本文引用: 1] [44] Yang H Li YL Shen L Kang XG Yue G Wang Y 2014 ). Spatial distribution patterns of seedling and sapling in a spruce-fir forest in the Changbai Mountains area in northeastern ChinaActa Ecologica Sinica 7311 -7319 . [本文引用: 1] [ 杨华 , 李艳丽 , 沈林 , 亢新刚 , 岳刚 , 王妍 (2014 ). 长白山云冷杉林幼苗幼树空间分布格局及其更新特征生态学报 , 34, 7311 -7319 .] [本文引用: 1] [45] Yang HX Zhang JT Wu B Li XS Zhang YY 2006 ). Point pattern analysis of Artemisia ordosica population in the Mu Us sandy landJournal of Plant Ecology (Chinese Version )563 -570 . [本文引用: 2] [ 杨洪晓 , 张金屯 , 吴波 , 李晓松 , 张友炎 (2006 ). 毛乌素沙地油蒿种群点格局分析植物生态学报 , 30, 563 -570 .] [本文引用: 2] [46] Yang WJ 2018 ). Spatial Distribution, Structural Characteristics and Evapotranspiration of Qinghai Spruce Forests in the Qilian Mountains, Northwest China 56 -57 . [本文引用: 1] [ 杨文娟 (2018 ). 祁连山青海云杉林空间分布和结构特征及蒸散研究 . 博士学位论文, 中国林业科学研究院, 北京 . 56 -57 .] [本文引用: 1] [47] Yu SC Zhang WH You JJ Deng L Huang CZ Xing ZL Fan RR He T 2017 ). Effect of thinning on Quercus wutaishanica trees form quality by the analytic hierarchy process in Huanglong MountainsScientia Silvae Sinicae 11 ), 104 -113 . [本文引用: 1] [ 于世川 , 张文辉 , 尤健健 , 邓磊 , 黄财智 , 邢忠利 , 樊蓉蓉 , 何婷 (2017 ). 抚育间伐对黄龙山辽东栎林木形质的影响林业科学 , 53(11 ), 104 -113 .] [本文引用: 1] [48] Zhang H Huang Q Zhao CY Zhang L Xu CH Kang YR 2016 ). Spatial pattern analysis of individuals in different age-classes of Picea crassifoliaChinese Journal of Ecology 1182 -1187 . [本文引用: 6] [ 张华 , 黄琴 , 赵传燕 , 张兰 , 徐春华 , 康雅茸 (2016 ). 青海云杉不同龄级立木的点格局分析生态学杂志 , 35, 1182 -1187 .] [本文引用: 6] [49] Zhang J Shangguan TL Duan YH Guo W Liu WH Guo DG 2014 ). Age structure and dynamics of Quercus wutaishanica population in Lingkong Mountain of Shanxi Province, ChinaChinese Journal of Applied Ecology 3125 -3130 . URL PMID:
25898607 [本文引用: 1] Using the plant survivorship theory, the age structure, and the relationship between tree height and diameter (DBH) of Quercus wutaishanica population in Lingkong Mountain were analyzed, and the static life table was compiled and the survival curve plotted. The shuttle shape in age structure of Q. wutaishanica population suggested its temporal stability. The linear regression significantly fitted the positive correlation between tree height and DBH. The maximal life expectancy was observed among the trees beyond the age of the highest mortality and coincided with the lowest point of mortality density, suggesting the strong vitality of the seedlings and young trees that survived in the natural selection and intraspecific competition. The population stability of the Q. wutaishanica population was characterized by the Deevey-II of the survival curve. The dynamic pattern was characterized by the recession in the early phase, growth in the intermediate phase, and stability in the latter phase.
[ 张婕 , 上官铁梁 , 段毅豪 , 郭微 , 刘卫华 , 郭东罡 (2014 ). 灵空山辽东栎种群年龄结构与动态应用生态学报 , 25, 3125 -3130 .] PMID:
25898607 [本文引用: 1] [50] Zhang JT 1998 ). Analysis of spatial point pattern for plant speciesActa Phytoecologica Sinica 344 -349 . [本文引用: 2] [ 张金屯 (1998 ). 植物种群空间分布的点格局分析植物生态学报 , 22, 344 -349 .] [本文引用: 2] [51] Zhang JT Meng DP 2004 ). Spatial pattern analysis of individuals in different age-classes of Larix principis-rupprechtii in Luya Mountain reserve, Shanxi, ChinaActa Ecologica Sinica 35 -40 . [本文引用: 3] [ 张金屯 , 孟东平 (2004 ). 芦芽山华北落叶松林不同龄级立木的点格局分析生态学报 , 24, 35 -40 .] [本文引用: 3] [52] Zhang M Mi XC Jin GZ 2014 ). Composition and spatial patterns of the Liangshui spruce-fir valley forest in the Xiao Hinggan MountainsChinese Science Bulletin 2377 -2391 . [本文引用: 2] [ 张觅 , 米湘成 , 金光泽 (2014 ). 小兴安岭凉水谷地云冷杉林群落组成与空间格局科学通报 , 59, 2377 -2391 .] [本文引用: 2] [53] Zhao LQ Huang HG Liang DS Zhang XL 2010 ). Spatial distribution pattern of Picea crassifolia population in Dayekou, Gansu ProvinceJournal of Beijing Forestry University 4 ), 59 -64 . [本文引用: 2] [ 赵丽琼 , 黄华国 , 梁大双 , 张晓丽 (2010 ). 甘肃大野口青海云杉种群的空间分布格局北京林业大学学报 , 32(4 ), 59 -64 .] [本文引用: 2] [54] Zhao Y Qi R Jiao J Chen XL Cao JH Feng YM Yang MM 2018 ). The population structure and dynamic characteristics of Picea purpurea at the Gahai-zecha areaActa Ecologica Sinica 7447 -7457 . [本文引用: 1] [ 赵阳 , 齐瑞 , 焦健 , 陈学龙 , 曹家豪 , 冯宜明 , 杨萌萌 (2018 ). 尕海-则岔地区紫果云杉种群结构与动态特征生态学报 , 38, 7447 -7457 .] [本文引用: 1] [55] Zhu X He ZB Du J Chen LF Lin PF Li J 2017 ). Temporal variability in soil moisture after thinning in semi-arid Picea crassifolia plantations in northwestern ChinaForest Ecology and Management 273 -285 . [本文引用: 1] [56] Zhu Y Bai F Liu HF Li WC Li L Li GQ Wang SZ Sang WG 2011 ). Population distribution patterns and interspecific spatial associations in warm temperate secondary forests, BeijingBiodiversity Science 252 -259 . [本文引用: 1] [ 祝燕 , 白帆 , 刘海丰 , 李文超 , 李亮 , 李广起 , 王顺忠 , 桑卫国 (2011 ). 北京暖温带次生林种群分布格局与种间空间关联性生物多样性 , 19, 252 -259 .] [本文引用: 1] spatstat: an R package for analyzing spatial point patterns
1
2005
... 由于
g (
r )的非聚集属性, 利用它进行点格局分析时需利用零模型来观察实际过程和理论过程之间的差异(
Stoyan & Penttinen, 2000 ).祁连山大野口流域青海云杉林样地地势相对平坦, 地形因子对于种群空间格局及其关联性影响较弱, 因此采用完全空间随机零模型作为零模型, 即假设物种的空间分布完全随机, 没有潜在的生物学过程发挥作用, 观察青海云杉种群不同径级个体的空间分布格局及其关联性与匀质泊松分布是否存在差异(
张觅等, 2014 ).空间尺度设定为0-60 m, 步长为1 m.采用蒙特卡罗(Monte-Carlo)随机模拟999次产生置信度为99%的包迹线, 以检验物种分布格局及关联性是否显著.在单变量分析中, 若实际分布的
g (
r )值落在包迹线之内, 则符合零模型, 为随机分布; 若在上包迹线以上, 则呈聚集分布; 若在下包迹线以下, 则为规则分布.在双变量分析中, 若实际分布的
g 12 (
r )值落在包迹线之间, 表示接受零假设, 说明不同径级间个体相互独立, 无空间相关性; 若在上包迹线以上, 则两径级间为正相关; 若落在下包迹线以下, 则两径级间为负相关关系(
Hao et al ., 2017 ).该研究所有统计与分析均在R 4.0.1软件中“spatstat”程序包(
Baddeley & Turner, 2005 )的相关函数中完成. ...
广西雅长林区铁坚油杉种群结构分布研究
1
2017
... 植物种群的径级结构是指不同径级组的个体在种群内所占比例和配置情况(
田润炜等, 2016 ), 是种群的生殖特性与外界环境相互作用的结果, 在一定程度上体现了种群的径级结构特征和生长发育状况, 进而可以预测种群和群落的动态发展趋势(
涂洪润等, 2019 ).本研究发现, 样地内青海云杉种群的幼苗和小树个体储备较为丰富, 其种群径级结构呈倒“J”型分布, 这说明该种群属增长型种群, 种群天然更新能力良好, 只要未来不遭受强烈干扰, 种群数量会保持逐渐增长.这与
李效雄等(2012) 和
张华等(2016) 分别对祁连山大野口流域和天老池小流域的青海云杉种群结构的研究结果一致.这可能是由青海云杉种群的生物学特性与环境因素共同作用的结果.已有研究表明, 森林更新过程中最重要和最敏感的阶段是幼苗阶段(
李晓笑等, 2011 ;
赵阳等, 2018 ), 林内有限的资源和空间往往成为种群更新的重要限制因素(
项小燕等, 2016 ).青海云杉种群的幼苗和小树个体数量多主要是由其生殖特性决定, 其种群可以产生大量高质量种子, 自然萌发形成的幼苗大都集中分布在林窗下, 形成大量小径级个体, 占据有利资源, 这是其种群在群落中持续处于优势地位的主要原因.随着小径级个体的生长, 它们对光照、水分、土壤养分等环境资源和空间需求随之增加, 由于环境资源和空间有限, 种内和种间竞争加剧, 出现强烈的自疏和他疏现象, 竞争能力弱的幼苗和小树在竞争中会大量死亡, 只有少量小径级个体能够成功转化为大径级个体, 造成样地内仅存在少量的中树和大树, 这成为该青海云杉种群更新和发展的“瓶颈” (
张婕等, 2014 ;
白卫国等, 2017 ;
于世川等, 2017 ).因此, 要实现种群数量的稳定增长, 必须加强对种群结构的保护, 尤其要重视对幼苗和小树及其生存环境的保护和改善. ...
广西雅长林区铁坚油杉种群结构分布研究
1
2017
... 植物种群的径级结构是指不同径级组的个体在种群内所占比例和配置情况(
田润炜等, 2016 ), 是种群的生殖特性与外界环境相互作用的结果, 在一定程度上体现了种群的径级结构特征和生长发育状况, 进而可以预测种群和群落的动态发展趋势(
涂洪润等, 2019 ).本研究发现, 样地内青海云杉种群的幼苗和小树个体储备较为丰富, 其种群径级结构呈倒“J”型分布, 这说明该种群属增长型种群, 种群天然更新能力良好, 只要未来不遭受强烈干扰, 种群数量会保持逐渐增长.这与
李效雄等(2012) 和
张华等(2016) 分别对祁连山大野口流域和天老池小流域的青海云杉种群结构的研究结果一致.这可能是由青海云杉种群的生物学特性与环境因素共同作用的结果.已有研究表明, 森林更新过程中最重要和最敏感的阶段是幼苗阶段(
李晓笑等, 2011 ;
赵阳等, 2018 ), 林内有限的资源和空间往往成为种群更新的重要限制因素(
项小燕等, 2016 ).青海云杉种群的幼苗和小树个体数量多主要是由其生殖特性决定, 其种群可以产生大量高质量种子, 自然萌发形成的幼苗大都集中分布在林窗下, 形成大量小径级个体, 占据有利资源, 这是其种群在群落中持续处于优势地位的主要原因.随着小径级个体的生长, 它们对光照、水分、土壤养分等环境资源和空间需求随之增加, 由于环境资源和空间有限, 种内和种间竞争加剧, 出现强烈的自疏和他疏现象, 竞争能力弱的幼苗和小树在竞争中会大量死亡, 只有少量小径级个体能够成功转化为大径级个体, 造成样地内仅存在少量的中树和大树, 这成为该青海云杉种群更新和发展的“瓶颈” (
张婕等, 2014 ;
白卫国等, 2017 ;
于世川等, 2017 ).因此, 要实现种群数量的稳定增长, 必须加强对种群结构的保护, 尤其要重视对幼苗和小树及其生存环境的保护和改善. ...
Contribution to the discussion of Dr Ripley’s paper
1
1977
... 该研究以
Ripley (1977) 提出的Ripley’s
K (
K (
r ))函数为基础, 采用单变量成对相关函数(
g (
r )), 以样地中每个植株的空间坐标为基础, 分析不同径级青海云杉个体在不同尺度下的分布格局.其中,
K (
r )函数的计算公式(
Besag, 1977 ;
Haase, 1995 )如下: ...
Research in large, long-term tropical forest plots
1
1995
... 参照CTFS (Center for Tropical Forest Science)和中国森林生物多样性监测网络(CForBio)的大样地建立与监测技术规范(
Condit, 1995 )(
http://www.ctfs.si.edu/ ), 于2010-2011年在祁连山国家级自然保护区大野口流域的相对平坦的青海云杉林内建立了一块面积10.2 hm
2 的青海云杉动态监测样地.整个样地呈长方形, 以样地西北角为原点, 南北方向为
x 轴(长340 m), 东西方向为
y 轴(宽300 m), 利用全站仪将整个样地划分成255个20 m × 20 m的样方(附录I), 进一步将每个20 m × 20 m的样方细分成16个5 m × 5 m的小样方, 对小样方内所有胸径(
DBH ) ≥ 1.0 cm的木本个体及其分支进行涂漆, 挂牌标记以便进行永久监测, 测量其相对坐标(
x ,
y )、胸径、冠幅、枝下高, 记录生长状况(存活、倒伏或枯立)等信息.采用全站仪测定每个样方的海拔高度, 并绘制等高线地形图(附录II).首次调查结果表明, 样地内共记录到
DBH ≥ 1.0 cm的活立木19 927株, 枯死木411株(
李效雄等, 2013 ). ...
Spatial patterns in the distribution of tropical tree species
2
2000
... 森林的空间格局与空间尺度密切相关, 物种的空间分布格局随着尺度的变化而变化(
Condit et al ., 2000 ;
杨华等, 2014 ).一般来说, 小尺度上的物种分布格局主要受种群自身生物学特性(如种内种间竞争、种子扩散限制和幼苗补充过程等)影响, 而大尺度上物种的空间分布则主要受生境异质性(如光照、土壤养分、水分和温度等)影响(
Shen et al ., 2013 ).本研究表明, 青海云杉种群不同径级的青海云杉个体在较小尺度内均表现为聚集分布, 随着空间尺度的增加, 呈现由聚集分布到随机分布的变化趋势.这一结果与国内外大多数有关种群空间格局的研究结果一致(
刘建泉, 2004 ;
Guo et al ., 2013 ;
张觅等, 2014 ;
张华等, 2016 ).这是因为同一种群的不同个体对生境条件的需求基本上是一致的(
张金屯和孟东平, 2004 ), 且聚集分布有利于种群个体相互庇护, 从而抵御外界不利环境, 以提高存活率(
Diggle, 1981 ).此外, 生境的异质性容易引起种群聚集生长, 同时由于植株对光照、水分、养分等资源的竞争较大(
李海东等, 2011 ), 导致聚集强度随着空间尺度的增大而呈现下降趋势, 表现为在较小尺度上植株倾向于聚集分布逐渐过渡为在较大尺度上的随机分布. ...
... 通常认为, 由于物种的生态学和生物学特性、种子扩散限制和生境异质性等机制的影响, 较小径级的物种常呈聚集分布, 尤其在小尺度上更加明显(
祝燕等, 2011 ).而随着径级增大, 种内和种间为争夺有限资源和空间的竞争也大大增强, 常呈随机或规则分布, 聚集强度也随径级的增加而降低(
Condit et al ., 2000 ).本研究结果也符合这一规律.样地内的青海云杉种群的幼树、小树和中树这类低龄级个体多呈聚集分布, 这一方面可能是由于青海云杉的种子具有种翅, 主要依靠风力传播, 使得大多数种子散落在母树周围一定范围内且随着距离的增大其种子传播的数量逐渐下降, 这些生物学特性使其形成聚集分布的空间分布特征.实地调查过程中也发现, 青海云杉幼苗大都集中分布于林窗下, 在阴蔽的林冠下, 出现幼苗和小树死亡现象, 在地被层特别是苔藓层较厚的地方, 幼苗较少.另一方面, 低龄级种群个体较小, 处在群落的下层, 对光、热、水和土壤养分等生态因子的竞争能力较弱, 聚集可以提高其竞争能力以及抵御外界不良环境, 提高成活率.此外, 小生境的异质作用导致幼树与小树分布不均匀, 如局部地形、土壤和植被等环境因子的差异, 以及采伐、自然灾害等形成的林窗, 都为低龄级个体的大量繁殖和生长创造了良好的条件(
张华等, 2016 ).随着种群年龄的增长, 个体对光照、水分、营养等资源和空间的需求不断增加, 种内和种间竞争加剧而出现强烈的自疏和他疏现象, 导致大量林木死亡, 种群密度下降, 聚集强度逐渐减弱, 种群由聚集分布向随机分布或规则分布转变.此外, 大树主要呈随机分布, 这可能是因为大树位于林冠层, 高度较高, 树冠基本定型, 根系也伸展到一定程度, 种内竞争加剧, 自疏效应使得种群的聚集分布格局被打破, 由聚集分布向随机分布转变. ...
Binary mosaics and the spatial pattern of heather
1
1981
... 森林的空间格局与空间尺度密切相关, 物种的空间分布格局随着尺度的变化而变化(
Condit et al ., 2000 ;
杨华等, 2014 ).一般来说, 小尺度上的物种分布格局主要受种群自身生物学特性(如种内种间竞争、种子扩散限制和幼苗补充过程等)影响, 而大尺度上物种的空间分布则主要受生境异质性(如光照、土壤养分、水分和温度等)影响(
Shen et al ., 2013 ).本研究表明, 青海云杉种群不同径级的青海云杉个体在较小尺度内均表现为聚集分布, 随着空间尺度的增加, 呈现由聚集分布到随机分布的变化趋势.这一结果与国内外大多数有关种群空间格局的研究结果一致(
刘建泉, 2004 ;
Guo et al ., 2013 ;
张觅等, 2014 ;
张华等, 2016 ).这是因为同一种群的不同个体对生境条件的需求基本上是一致的(
张金屯和孟东平, 2004 ), 且聚集分布有利于种群个体相互庇护, 从而抵御外界不利环境, 以提高存活率(
Diggle, 1981 ).此外, 生境的异质性容易引起种群聚集生长, 同时由于植株对光照、水分、养分等资源的竞争较大(
李海东等, 2011 ), 导致聚集强度随着空间尺度的增大而呈现下降趋势, 表现为在较小尺度上植株倾向于聚集分布逐渐过渡为在较大尺度上的随机分布. ...
1
2003
...
g (
r )由
K 函数衍生而来.与
K (
r )相比,
g (
r )能有效消除小尺度累积效应的缺点, 在评估聚集程度方面的研究中得到了广泛的应用(
Diggle, 2003 ).
g (
r )与
K 函数的关系为: ...
Spatial pattern and size distribution of the animal-dispersed tree
Quercus robur in two spruce- dominated forests
1
2000
... 虽然径级和龄级之间存在差异, 但在同样的环境条件下, 树木的径级和龄级对生境条件的响应规律具有一致性.青海云杉寿命很长, 在100-140年以后生长才缓慢下来, 少数林木在300-400年仍能持续缓慢生长.已有研究表明, 其胸径和树龄具有明显的线性关系,
y = 0.2133
x - 1.1136,
R 2 = 0.984 9 (
y 为胸径,
x 树龄).如: 当胸径为5.0、12.5以及22.5 cm时分别大致对应树龄30、64和111年(
杨文娟, 2018 ).因此可以用胸径来反映种群的生活史阶段(
Frost & Rydin, 2000 ;
李立等, 2010 ).从而该研究采用径级结构(胸径大小)代替年龄结构, 分析不同径级青海云杉种群分布格局及其关联性的变化.该研究参照李妍(2013)对青海云杉种群径级结构的划分标准, 同时结合样地内青海云杉胸径的实际情况以及青海云杉的生活史特点, 依据青海云杉胸径大小将其划分为: 径级I (幼树, 1 cm ≤
DBH < 5.0 cm)、径级II (小树, 5.0 cm ≤
DBH < 12.5 cm)、径级III (中树, 12.5 cm ≤
DBH < 22.5 cm)、径级IV (大树,
DBH ≥ 22.5 cm) 4个不同径级. ...
Climate-growth analysis of Qilian juniper across an altitudinal gradient in the central Qilian Mountains, northwest China
1
2013
... 祁连山是中国西部的主要山脉之一, 位于青藏高原、蒙古高原、黄土高原三大高原交汇带, 是石羊河、黑河、疏勒河等6大内陆河和黄河流域的重要水源产流地, 维系着河西乃至西部地区生存与发展的命脉, 在国家生态建设中具有十分重要的战略地位(
Gao et al ., 2013 ;
Zhu et al ., 2017 ).祁连山山地森林的主要建群种——青海云杉(
Picea crassifolia ), 为松科云杉属(
Picea )乔木, 是中国青藏高原东北边缘特有树种, 多以纯林形式存在, 呈带状或斑块状分布于海拔2 300-3 300 m的阴坡和半阴坡, 分布面积占祁连山水源涵养林面积的24.74%, 占乔木林面积的75.72%, 对于维系祁连山的生物多样性、水源涵养功能乃至区域生态平衡都具有至关重要的作用(
田风霞等, 2012 ;
何芳兰等, 2016 ;
张华等, 2016 ).部分****(
赵丽琼等, 2010 ;
张华等, 2016 )虽然已对青海云杉种群空间格局及其关联性进行了研究, 但是已有研究的空间尺度相对较小, 仅能揭示小尺度下的种群空间分布格局, 难以客观地反映青海云杉种群空间分布的特点和种内关系.鉴于此, 本研究基于祁连山大野口流域10.2 hm
2 青海云杉动态监测样地数据, 采用空间点格局分析方法中的成对相关函数对不同径级青海云杉种群的空间分布格局及其关联性进行分析, 旨在阐明: (1)祁连山大野口流域青海云杉种群不同径级个体的空间分布格局及其关联性如何随空间尺度进行变化; (2)不同径级个体的空间分布格局及其关联性形成的潜在生态学过程与影响机制.上述科学问题的合理解答, 能从空间格局的角度客观地揭示青海云杉种群发展规律, 可以为制定该种群的配置、种植与保护管理措施提供指导, 并对祁连山地区的植被恢复与重建、生物多样性保护等具有重要的意义和价值. ...
Heterogeneity influences spatial patterns and demographics in forest stands
1
2008
... 该研究还发现径级相近的个体空间关联性呈正关联或无关联, 随着个体径级间差距增大, 它们之间的空间关联性转变为无关联或负关联, 这说明同一物种不同径级的个体之间同时存在竞争、依赖以及无明确的生态关系, 这是由于物种具有一致的生境偏好所造成的(
杨洪晓等, 2006 ).径级相近个体的空间关联性呈正关联, 表明它们相伴出现, 对生境的选择具有一致性, 在空间分布上是相互依存的.这可能是由于它们生长发育阶段相近, 对环境资源的需求具有相似性和生态位存在重叠, 但小径级个体相对幼小, 对光照、水分和土壤养分等资源的竞争能力相对较弱, 需要借助高大植株的庇护, 才得以提高存活率, 而较大径级的植株个体可以改善样地的小生境, 使得邻近径级的个体更适宜生存, 彼此共享资源(
涂洪润等, 2019 ).同时, 由于物种专一性的资源分布不均, 种内竞争具有非对称性的特点, 也就是说当径级相近时, 不同径级间关系较协调(
Getzin et al ., 2008 ).而无关联则说明个体间互不干扰, 个体在前期发育过程中已经占据适宜的生态位, 与种内其他径级个体间的关联性不高, 能各自生长发育(
涂洪润等, 2019 ).随着个体间径级差距增加, 它们之间的关联性转变成无关联甚至负关联, 说明随着个体间径级差距的加大, 它们的空间正关联关系逐渐减弱, 当个体间差异较大时, 大径级个体在竞争中处于优势地位, 可能导致小径级个体的生长发育迟滞或者死亡, 使其周围小径级个体很少(
Kenkel, 1988 ), 这也间接证实了随着个体大小的生长变化, 青海云杉种群的空间分布存在一个从聚集分布到随机分布的自疏过程. ...
Spatial distribution of tree species in a species-rich subtropical mountain forest in central China
2
2013
... 种群的空间格局是指特定时间内, 种群所有个体在其水平生活空间内的分布状况及其属性在空间上的配置特征, 往往是由种群自身生物学特性、种内种间竞争、环境条件等因素综合作用的结果(
林玥, 2008 ;
Liu et al ., 2020 ).种群的空间分布格局和空间关联性是种群空间格局研究的2个主要内容, 也是种群生态关系在空间格局上的两种表现形式(
张金屯和孟东平, 2004 ).大量研究表明种群的空间分布格局与空间关联性关系密切, 聚集分布和空间正关联表明种群内部正向(互惠)的生态关系, 规则分布和空间负关联则表明种群内部负向(互斥)的生态关系, 随机分布和空间无关联则代表着种群内部不存在明确的生态关系(
杨洪晓, 2006 ).因此, 研究植物种群的空间分布格局及其关联性不仅能够了解种群的空间水平结构与生物、生态学习性(如生活史策略、是否喜光或耐阴等)、种群间相互作用、种群与环境因子的相互作用规律(如植物与生长环境间的适合度、生境异质性等)以及种群在群落中的地位和生态学功能, 更重要的是可以揭示这种分布格局及其关联性的潜在成因, 进而推演种群动态变化的生态学过程(如种内和种间竞争、种子扩散和干扰等), 深化对群落结构的认识, 揭示植物群落的生物多样性形成和维持机制(
McDonald et al ., 2003 ;
Hou et al , 2004 ;
Wiegand & Moloney, 2004 ;
Nathan, 2006 ;
Wang et al ., 2010 ;
Guo et al ., 2013 ). ...
... 森林的空间格局与空间尺度密切相关, 物种的空间分布格局随着尺度的变化而变化(
Condit et al ., 2000 ;
杨华等, 2014 ).一般来说, 小尺度上的物种分布格局主要受种群自身生物学特性(如种内种间竞争、种子扩散限制和幼苗补充过程等)影响, 而大尺度上物种的空间分布则主要受生境异质性(如光照、土壤养分、水分和温度等)影响(
Shen et al ., 2013 ).本研究表明, 青海云杉种群不同径级的青海云杉个体在较小尺度内均表现为聚集分布, 随着空间尺度的增加, 呈现由聚集分布到随机分布的变化趋势.这一结果与国内外大多数有关种群空间格局的研究结果一致(
刘建泉, 2004 ;
Guo et al ., 2013 ;
张觅等, 2014 ;
张华等, 2016 ).这是因为同一种群的不同个体对生境条件的需求基本上是一致的(
张金屯和孟东平, 2004 ), 且聚集分布有利于种群个体相互庇护, 从而抵御外界不利环境, 以提高存活率(
Diggle, 1981 ).此外, 生境的异质性容易引起种群聚集生长, 同时由于植株对光照、水分、养分等资源的竞争较大(
李海东等, 2011 ), 导致聚集强度随着空间尺度的增大而呈现下降趋势, 表现为在较小尺度上植株倾向于聚集分布逐渐过渡为在较大尺度上的随机分布. ...
Spatial pattern analysis in ecology based on Ripley’s K-function: introduction and methods of edge correction
1
1995
... 该研究以
Ripley (1977) 提出的Ripley’s
K (
K (
r ))函数为基础, 采用单变量成对相关函数(
g (
r )), 以样地中每个植株的空间坐标为基础, 分析不同径级青海云杉个体在不同尺度下的分布格局.其中,
K (
r )函数的计算公式(
Besag, 1977 ;
Haase, 1995 )如下: ...
Patches structure succession based on spatial point pattern features in semi-arid ecosystems of the water-wind erosion crisscross region
1
2017
... 由于
g (
r )的非聚集属性, 利用它进行点格局分析时需利用零模型来观察实际过程和理论过程之间的差异(
Stoyan & Penttinen, 2000 ).祁连山大野口流域青海云杉林样地地势相对平坦, 地形因子对于种群空间格局及其关联性影响较弱, 因此采用完全空间随机零模型作为零模型, 即假设物种的空间分布完全随机, 没有潜在的生物学过程发挥作用, 观察青海云杉种群不同径级个体的空间分布格局及其关联性与匀质泊松分布是否存在差异(
张觅等, 2014 ).空间尺度设定为0-60 m, 步长为1 m.采用蒙特卡罗(Monte-Carlo)随机模拟999次产生置信度为99%的包迹线, 以检验物种分布格局及关联性是否显著.在单变量分析中, 若实际分布的
g (
r )值落在包迹线之内, 则符合零模型, 为随机分布; 若在上包迹线以上, 则呈聚集分布; 若在下包迹线以下, 则为规则分布.在双变量分析中, 若实际分布的
g 12 (
r )值落在包迹线之间, 表示接受零假设, 说明不同径级间个体相互独立, 无空间相关性; 若在上包迹线以上, 则两径级间为正相关; 若落在下包迹线以下, 则两径级间为负相关关系(
Hao et al ., 2017 ).该研究所有统计与分析均在R 4.0.1软件中“spatstat”程序包(
Baddeley & Turner, 2005 )的相关函数中完成. ...
Distribution patterns of tree species in a Malaysian tropical rain forest
1
1997
... 空间分布格局的研究方法众多, 早期的研究方法主要有: 以频度/密度为基础的样方法、以距离为基础的无样地法、方差区组分析法、双向轨迹方差分析、
χ 2 检验和分形理论等(
张金屯, 1998 ;
王本洋和余世孝, 2005 ).传统的样方取样和分析方法常基于单一尺度(固定大小样方)的若干样方, 常采用
χ 2 分析法和方差/均值比率法等统计分析方法, 然而种群空间分布格局及其关联性对种群密度和空间尺度均有很强的依赖性, 在不同的尺度上往往表现出不同的类型, 故早期的取样和分析方法只能研究特定尺度下种群的空间分布格局特征, 难以全面地反映森林内物种的空间分布特点和种间关系(
He et al ., 1997 ;
李立等, 2010 ).鉴于上述研究方法的局限性,
Ripley (1977) 和
Diggle (1983) 分别提出和完善了点格局分析法, 以分析各种尺度上的空间分布格局.该方法以数字化的植物个体位置信息为基本数据, 将每个个体都视为二维空间中的一个点, 所有个体组成了植物种的空间分布图, 然后在空间分布图的基础上进行种群空间格局分析.其优点是能最大限度地利用点与点之间的距离, 不仅能够提供全面的空间格局信息, 而且具有较强的检验能力, 克服了传统研究方法中尺度单一等不足, 可以分析任意尺度下种群的空间格局特征及种内、种间关系(
Wiegand & Moloney, 2004 ;
Schleicher et al ., 2011 ;
沈志强等, 2016 ;
涂洪润等, 2019 ).因此, 空间点格局分析法在种群空间格局分布特征及潜在生态过程假说的研究方面得到了广泛应用. ...
祁连山青海云杉人工林与天然林群落结构特征及物种多样性比较研究
1
2016
... 祁连山是中国西部的主要山脉之一, 位于青藏高原、蒙古高原、黄土高原三大高原交汇带, 是石羊河、黑河、疏勒河等6大内陆河和黄河流域的重要水源产流地, 维系着河西乃至西部地区生存与发展的命脉, 在国家生态建设中具有十分重要的战略地位(
Gao et al ., 2013 ;
Zhu et al ., 2017 ).祁连山山地森林的主要建群种——青海云杉(
Picea crassifolia ), 为松科云杉属(
Picea )乔木, 是中国青藏高原东北边缘特有树种, 多以纯林形式存在, 呈带状或斑块状分布于海拔2 300-3 300 m的阴坡和半阴坡, 分布面积占祁连山水源涵养林面积的24.74%, 占乔木林面积的75.72%, 对于维系祁连山的生物多样性、水源涵养功能乃至区域生态平衡都具有至关重要的作用(
田风霞等, 2012 ;
何芳兰等, 2016 ;
张华等, 2016 ).部分****(
赵丽琼等, 2010 ;
张华等, 2016 )虽然已对青海云杉种群空间格局及其关联性进行了研究, 但是已有研究的空间尺度相对较小, 仅能揭示小尺度下的种群空间分布格局, 难以客观地反映青海云杉种群空间分布的特点和种内关系.鉴于此, 本研究基于祁连山大野口流域10.2 hm
2 青海云杉动态监测样地数据, 采用空间点格局分析方法中的成对相关函数对不同径级青海云杉种群的空间分布格局及其关联性进行分析, 旨在阐明: (1)祁连山大野口流域青海云杉种群不同径级个体的空间分布格局及其关联性如何随空间尺度进行变化; (2)不同径级个体的空间分布格局及其关联性形成的潜在生态学过程与影响机制.上述科学问题的合理解答, 能从空间格局的角度客观地揭示青海云杉种群发展规律, 可以为制定该种群的配置、种植与保护管理措施提供指导, 并对祁连山地区的植被恢复与重建、生物多样性保护等具有重要的意义和价值. ...
祁连山青海云杉人工林与天然林群落结构特征及物种多样性比较研究
1
2016
... 祁连山是中国西部的主要山脉之一, 位于青藏高原、蒙古高原、黄土高原三大高原交汇带, 是石羊河、黑河、疏勒河等6大内陆河和黄河流域的重要水源产流地, 维系着河西乃至西部地区生存与发展的命脉, 在国家生态建设中具有十分重要的战略地位(
Gao et al ., 2013 ;
Zhu et al ., 2017 ).祁连山山地森林的主要建群种——青海云杉(
Picea crassifolia ), 为松科云杉属(
Picea )乔木, 是中国青藏高原东北边缘特有树种, 多以纯林形式存在, 呈带状或斑块状分布于海拔2 300-3 300 m的阴坡和半阴坡, 分布面积占祁连山水源涵养林面积的24.74%, 占乔木林面积的75.72%, 对于维系祁连山的生物多样性、水源涵养功能乃至区域生态平衡都具有至关重要的作用(
田风霞等, 2012 ;
何芳兰等, 2016 ;
张华等, 2016 ).部分****(
赵丽琼等, 2010 ;
张华等, 2016 )虽然已对青海云杉种群空间格局及其关联性进行了研究, 但是已有研究的空间尺度相对较小, 仅能揭示小尺度下的种群空间分布格局, 难以客观地反映青海云杉种群空间分布的特点和种内关系.鉴于此, 本研究基于祁连山大野口流域10.2 hm
2 青海云杉动态监测样地数据, 采用空间点格局分析方法中的成对相关函数对不同径级青海云杉种群的空间分布格局及其关联性进行分析, 旨在阐明: (1)祁连山大野口流域青海云杉种群不同径级个体的空间分布格局及其关联性如何随空间尺度进行变化; (2)不同径级个体的空间分布格局及其关联性形成的潜在生态学过程与影响机制.上述科学问题的合理解答, 能从空间格局的角度客观地揭示青海云杉种群发展规律, 可以为制定该种群的配置、种植与保护管理措施提供指导, 并对祁连山地区的植被恢复与重建、生物多样性保护等具有重要的意义和价值. ...
Spatial patterns and associations in a
Quercus -
Betula forest in Northern China
1
2004
... 种群的空间格局是指特定时间内, 种群所有个体在其水平生活空间内的分布状况及其属性在空间上的配置特征, 往往是由种群自身生物学特性、种内种间竞争、环境条件等因素综合作用的结果(
林玥, 2008 ;
Liu et al ., 2020 ).种群的空间分布格局和空间关联性是种群空间格局研究的2个主要内容, 也是种群生态关系在空间格局上的两种表现形式(
张金屯和孟东平, 2004 ).大量研究表明种群的空间分布格局与空间关联性关系密切, 聚集分布和空间正关联表明种群内部正向(互惠)的生态关系, 规则分布和空间负关联则表明种群内部负向(互斥)的生态关系, 随机分布和空间无关联则代表着种群内部不存在明确的生态关系(
杨洪晓, 2006 ).因此, 研究植物种群的空间分布格局及其关联性不仅能够了解种群的空间水平结构与生物、生态学习性(如生活史策略、是否喜光或耐阴等)、种群间相互作用、种群与环境因子的相互作用规律(如植物与生长环境间的适合度、生境异质性等)以及种群在群落中的地位和生态学功能, 更重要的是可以揭示这种分布格局及其关联性的潜在成因, 进而推演种群动态变化的生态学过程(如种内和种间竞争、种子扩散和干扰等), 深化对群落结构的认识, 揭示植物群落的生物多样性形成和维持机制(
McDonald et al ., 2003 ;
Hou et al , 2004 ;
Wiegand & Moloney, 2004 ;
Nathan, 2006 ;
Wang et al ., 2010 ;
Guo et al ., 2013 ). ...
Pattern of self-thinning in jack pine: testing the random mortality hypothesis
1
1988
... 该研究还发现径级相近的个体空间关联性呈正关联或无关联, 随着个体径级间差距增大, 它们之间的空间关联性转变为无关联或负关联, 这说明同一物种不同径级的个体之间同时存在竞争、依赖以及无明确的生态关系, 这是由于物种具有一致的生境偏好所造成的(
杨洪晓等, 2006 ).径级相近个体的空间关联性呈正关联, 表明它们相伴出现, 对生境的选择具有一致性, 在空间分布上是相互依存的.这可能是由于它们生长发育阶段相近, 对环境资源的需求具有相似性和生态位存在重叠, 但小径级个体相对幼小, 对光照、水分和土壤养分等资源的竞争能力相对较弱, 需要借助高大植株的庇护, 才得以提高存活率, 而较大径级的植株个体可以改善样地的小生境, 使得邻近径级的个体更适宜生存, 彼此共享资源(
涂洪润等, 2019 ).同时, 由于物种专一性的资源分布不均, 种内竞争具有非对称性的特点, 也就是说当径级相近时, 不同径级间关系较协调(
Getzin et al ., 2008 ).而无关联则说明个体间互不干扰, 个体在前期发育过程中已经占据适宜的生态位, 与种内其他径级个体间的关联性不高, 能各自生长发育(
涂洪润等, 2019 ).随着个体间径级差距增加, 它们之间的关联性转变成无关联甚至负关联, 说明随着个体间径级差距的加大, 它们的空间正关联关系逐渐减弱, 当个体间差异较大时, 大径级个体在竞争中处于优势地位, 可能导致小径级个体的生长发育迟滞或者死亡, 使其周围小径级个体很少(
Kenkel, 1988 ), 这也间接证实了随着个体大小的生长变化, 青海云杉种群的空间分布存在一个从聚集分布到随机分布的自疏过程. ...
雅鲁藏布江中游河岸带几种主要沙生植物种群点格局分析
1
2011
... 森林的空间格局与空间尺度密切相关, 物种的空间分布格局随着尺度的变化而变化(
Condit et al ., 2000 ;
杨华等, 2014 ).一般来说, 小尺度上的物种分布格局主要受种群自身生物学特性(如种内种间竞争、种子扩散限制和幼苗补充过程等)影响, 而大尺度上物种的空间分布则主要受生境异质性(如光照、土壤养分、水分和温度等)影响(
Shen et al ., 2013 ).本研究表明, 青海云杉种群不同径级的青海云杉个体在较小尺度内均表现为聚集分布, 随着空间尺度的增加, 呈现由聚集分布到随机分布的变化趋势.这一结果与国内外大多数有关种群空间格局的研究结果一致(
刘建泉, 2004 ;
Guo et al ., 2013 ;
张觅等, 2014 ;
张华等, 2016 ).这是因为同一种群的不同个体对生境条件的需求基本上是一致的(
张金屯和孟东平, 2004 ), 且聚集分布有利于种群个体相互庇护, 从而抵御外界不利环境, 以提高存活率(
Diggle, 1981 ).此外, 生境的异质性容易引起种群聚集生长, 同时由于植株对光照、水分、养分等资源的竞争较大(
李海东等, 2011 ), 导致聚集强度随着空间尺度的增大而呈现下降趋势, 表现为在较小尺度上植株倾向于聚集分布逐渐过渡为在较大尺度上的随机分布. ...
雅鲁藏布江中游河岸带几种主要沙生植物种群点格局分析
1
2011
... 森林的空间格局与空间尺度密切相关, 物种的空间分布格局随着尺度的变化而变化(
Condit et al ., 2000 ;
杨华等, 2014 ).一般来说, 小尺度上的物种分布格局主要受种群自身生物学特性(如种内种间竞争、种子扩散限制和幼苗补充过程等)影响, 而大尺度上物种的空间分布则主要受生境异质性(如光照、土壤养分、水分和温度等)影响(
Shen et al ., 2013 ).本研究表明, 青海云杉种群不同径级的青海云杉个体在较小尺度内均表现为聚集分布, 随着空间尺度的增加, 呈现由聚集分布到随机分布的变化趋势.这一结果与国内外大多数有关种群空间格局的研究结果一致(
刘建泉, 2004 ;
Guo et al ., 2013 ;
张觅等, 2014 ;
张华等, 2016 ).这是因为同一种群的不同个体对生境条件的需求基本上是一致的(
张金屯和孟东平, 2004 ), 且聚集分布有利于种群个体相互庇护, 从而抵御外界不利环境, 以提高存活率(
Diggle, 1981 ).此外, 生境的异质性容易引起种群聚集生长, 同时由于植株对光照、水分、养分等资源的竞争较大(
李海东等, 2011 ), 导致聚集强度随着空间尺度的增大而呈现下降趋势, 表现为在较小尺度上植株倾向于聚集分布逐渐过渡为在较大尺度上的随机分布. ...
古田山常绿阔叶林优势树种甜槠和木荷的空间格局分析
2
2010
... 空间分布格局的研究方法众多, 早期的研究方法主要有: 以频度/密度为基础的样方法、以距离为基础的无样地法、方差区组分析法、双向轨迹方差分析、
χ 2 检验和分形理论等(
张金屯, 1998 ;
王本洋和余世孝, 2005 ).传统的样方取样和分析方法常基于单一尺度(固定大小样方)的若干样方, 常采用
χ 2 分析法和方差/均值比率法等统计分析方法, 然而种群空间分布格局及其关联性对种群密度和空间尺度均有很强的依赖性, 在不同的尺度上往往表现出不同的类型, 故早期的取样和分析方法只能研究特定尺度下种群的空间分布格局特征, 难以全面地反映森林内物种的空间分布特点和种间关系(
He et al ., 1997 ;
李立等, 2010 ).鉴于上述研究方法的局限性,
Ripley (1977) 和
Diggle (1983) 分别提出和完善了点格局分析法, 以分析各种尺度上的空间分布格局.该方法以数字化的植物个体位置信息为基本数据, 将每个个体都视为二维空间中的一个点, 所有个体组成了植物种的空间分布图, 然后在空间分布图的基础上进行种群空间格局分析.其优点是能最大限度地利用点与点之间的距离, 不仅能够提供全面的空间格局信息, 而且具有较强的检验能力, 克服了传统研究方法中尺度单一等不足, 可以分析任意尺度下种群的空间格局特征及种内、种间关系(
Wiegand & Moloney, 2004 ;
Schleicher et al ., 2011 ;
沈志强等, 2016 ;
涂洪润等, 2019 ).因此, 空间点格局分析法在种群空间格局分布特征及潜在生态过程假说的研究方面得到了广泛应用. ...
... 虽然径级和龄级之间存在差异, 但在同样的环境条件下, 树木的径级和龄级对生境条件的响应规律具有一致性.青海云杉寿命很长, 在100-140年以后生长才缓慢下来, 少数林木在300-400年仍能持续缓慢生长.已有研究表明, 其胸径和树龄具有明显的线性关系,
y = 0.2133
x - 1.1136,
R 2 = 0.984 9 (
y 为胸径,
x 树龄).如: 当胸径为5.0、12.5以及22.5 cm时分别大致对应树龄30、64和111年(
杨文娟, 2018 ).因此可以用胸径来反映种群的生活史阶段(
Frost & Rydin, 2000 ;
李立等, 2010 ).从而该研究采用径级结构(胸径大小)代替年龄结构, 分析不同径级青海云杉种群分布格局及其关联性的变化.该研究参照李妍(2013)对青海云杉种群径级结构的划分标准, 同时结合样地内青海云杉胸径的实际情况以及青海云杉的生活史特点, 依据青海云杉胸径大小将其划分为: 径级I (幼树, 1 cm ≤
DBH < 5.0 cm)、径级II (小树, 5.0 cm ≤
DBH < 12.5 cm)、径级III (中树, 12.5 cm ≤
DBH < 22.5 cm)、径级IV (大树,
DBH ≥ 22.5 cm) 4个不同径级. ...
古田山常绿阔叶林优势树种甜槠和木荷的空间格局分析
2
2010
... 空间分布格局的研究方法众多, 早期的研究方法主要有: 以频度/密度为基础的样方法、以距离为基础的无样地法、方差区组分析法、双向轨迹方差分析、
χ 2 检验和分形理论等(
张金屯, 1998 ;
王本洋和余世孝, 2005 ).传统的样方取样和分析方法常基于单一尺度(固定大小样方)的若干样方, 常采用
χ 2 分析法和方差/均值比率法等统计分析方法, 然而种群空间分布格局及其关联性对种群密度和空间尺度均有很强的依赖性, 在不同的尺度上往往表现出不同的类型, 故早期的取样和分析方法只能研究特定尺度下种群的空间分布格局特征, 难以全面地反映森林内物种的空间分布特点和种间关系(
He et al ., 1997 ;
李立等, 2010 ).鉴于上述研究方法的局限性,
Ripley (1977) 和
Diggle (1983) 分别提出和完善了点格局分析法, 以分析各种尺度上的空间分布格局.该方法以数字化的植物个体位置信息为基本数据, 将每个个体都视为二维空间中的一个点, 所有个体组成了植物种的空间分布图, 然后在空间分布图的基础上进行种群空间格局分析.其优点是能最大限度地利用点与点之间的距离, 不仅能够提供全面的空间格局信息, 而且具有较强的检验能力, 克服了传统研究方法中尺度单一等不足, 可以分析任意尺度下种群的空间格局特征及种内、种间关系(
Wiegand & Moloney, 2004 ;
Schleicher et al ., 2011 ;
沈志强等, 2016 ;
涂洪润等, 2019 ).因此, 空间点格局分析法在种群空间格局分布特征及潜在生态过程假说的研究方面得到了广泛应用. ...
... 虽然径级和龄级之间存在差异, 但在同样的环境条件下, 树木的径级和龄级对生境条件的响应规律具有一致性.青海云杉寿命很长, 在100-140年以后生长才缓慢下来, 少数林木在300-400年仍能持续缓慢生长.已有研究表明, 其胸径和树龄具有明显的线性关系,
y = 0.2133
x - 1.1136,
R 2 = 0.984 9 (
y 为胸径,
x 树龄).如: 当胸径为5.0、12.5以及22.5 cm时分别大致对应树龄30、64和111年(
杨文娟, 2018 ).因此可以用胸径来反映种群的生活史阶段(
Frost & Rydin, 2000 ;
李立等, 2010 ).从而该研究采用径级结构(胸径大小)代替年龄结构, 分析不同径级青海云杉种群分布格局及其关联性的变化.该研究参照李妍(2013)对青海云杉种群径级结构的划分标准, 同时结合样地内青海云杉胸径的实际情况以及青海云杉的生活史特点, 依据青海云杉胸径大小将其划分为: 径级I (幼树, 1 cm ≤
DBH < 5.0 cm)、径级II (小树, 5.0 cm ≤
DBH < 12.5 cm)、径级III (中树, 12.5 cm ≤
DBH < 22.5 cm)、径级IV (大树,
DBH ≥ 22.5 cm) 4个不同径级. ...
祁连山青海云杉林动态监测样地群落特征
2
2013
... 研究样地位于祁连山中段北坡大野口流域青海云杉林内, 地理位置为100.25° E, 38.53° N.样地最低海拔2 856 m, 最高海拔2 962 m, 最大高差106 m, 坡向西北, 坡度13°.根据祁连山水源涵养林研究院的资料, 样地所在的大野口流域气候属于温带大陆性高寒半湿润山地气候, 雨热同期, 冬季寒冷漫长, 夏季短暂, 年平均气温5.4 ℃, 最冷月(1月)平均气温为-12.5 ℃, 最热月(7月)平均气温为19.6 ℃, 年降水量为300-500 mm, 主要集中在6-9月, 年平均相对湿度60% (1994-2004年), 年蒸发量为1 488 mm.样地的成土母质为钙质岩, 土壤类型为山地森林灰褐土, 土壤pH 6.3-7.9, 厚度约1 m.林分起源为天然次生林, 为青海云杉纯林, 林分结构简单, 郁闭度较高, 林下光照不足.青海云杉群落成层现象明显, 可明显划分为乔木层、灌木层、草本层和苔藓层4个层次(
赵丽琼等, 2010 ).其中, 乔木树种组成单一, 除青海云杉外, 只有少量祁连圆柏(
Juniperus przewalskii ); 灌木生长发育不良, 物种较少, 主要有金露梅(
Potentilla fruticosa )、银露梅(
Potentilla glabra )、鬼箭锦鸡儿(
Caragana jubata )、吉拉柳(
Salix gilashanica )、鲜黄小檗(
Berberis diaphana )和高山绣线菊(
Spiraea alpina ); 草本植物物种较丰富, 主要有黑穗薹草(
Carex atrata )、珠芽蓼(
Polygonum viviparum )、针茅(
Stipa capillata )和马蔺(
Iris lactea )等典型高山草甸植物.苔藓层发育良好, 盖度达80%以上, 厚度约10 cm, 主要物种有山羽藓(
Abietinella abietina )、欧灰藓(
Hypnum cupressiforme )和匐灯藓(
Plagiomnium cuspidatum )等(
李效雄等, 2013 ). ...
... 参照CTFS (Center for Tropical Forest Science)和中国森林生物多样性监测网络(CForBio)的大样地建立与监测技术规范(
Condit, 1995 )(
http://www.ctfs.si.edu/ ), 于2010-2011年在祁连山国家级自然保护区大野口流域的相对平坦的青海云杉林内建立了一块面积10.2 hm
2 的青海云杉动态监测样地.整个样地呈长方形, 以样地西北角为原点, 南北方向为
x 轴(长340 m), 东西方向为
y 轴(宽300 m), 利用全站仪将整个样地划分成255个20 m × 20 m的样方(附录I), 进一步将每个20 m × 20 m的样方细分成16个5 m × 5 m的小样方, 对小样方内所有胸径(
DBH ) ≥ 1.0 cm的木本个体及其分支进行涂漆, 挂牌标记以便进行永久监测, 测量其相对坐标(
x ,
y )、胸径、冠幅、枝下高, 记录生长状况(存活、倒伏或枯立)等信息.采用全站仪测定每个样方的海拔高度, 并绘制等高线地形图(附录II).首次调查结果表明, 样地内共记录到
DBH ≥ 1.0 cm的活立木19 927株, 枯死木411株(
李效雄等, 2013 ). ...
祁连山青海云杉林动态监测样地群落特征
2
2013
... 研究样地位于祁连山中段北坡大野口流域青海云杉林内, 地理位置为100.25° E, 38.53° N.样地最低海拔2 856 m, 最高海拔2 962 m, 最大高差106 m, 坡向西北, 坡度13°.根据祁连山水源涵养林研究院的资料, 样地所在的大野口流域气候属于温带大陆性高寒半湿润山地气候, 雨热同期, 冬季寒冷漫长, 夏季短暂, 年平均气温5.4 ℃, 最冷月(1月)平均气温为-12.5 ℃, 最热月(7月)平均气温为19.6 ℃, 年降水量为300-500 mm, 主要集中在6-9月, 年平均相对湿度60% (1994-2004年), 年蒸发量为1 488 mm.样地的成土母质为钙质岩, 土壤类型为山地森林灰褐土, 土壤pH 6.3-7.9, 厚度约1 m.林分起源为天然次生林, 为青海云杉纯林, 林分结构简单, 郁闭度较高, 林下光照不足.青海云杉群落成层现象明显, 可明显划分为乔木层、灌木层、草本层和苔藓层4个层次(
赵丽琼等, 2010 ).其中, 乔木树种组成单一, 除青海云杉外, 只有少量祁连圆柏(
Juniperus przewalskii ); 灌木生长发育不良, 物种较少, 主要有金露梅(
Potentilla fruticosa )、银露梅(
Potentilla glabra )、鬼箭锦鸡儿(
Caragana jubata )、吉拉柳(
Salix gilashanica )、鲜黄小檗(
Berberis diaphana )和高山绣线菊(
Spiraea alpina ); 草本植物物种较丰富, 主要有黑穗薹草(
Carex atrata )、珠芽蓼(
Polygonum viviparum )、针茅(
Stipa capillata )和马蔺(
Iris lactea )等典型高山草甸植物.苔藓层发育良好, 盖度达80%以上, 厚度约10 cm, 主要物种有山羽藓(
Abietinella abietina )、欧灰藓(
Hypnum cupressiforme )和匐灯藓(
Plagiomnium cuspidatum )等(
李效雄等, 2013 ). ...
... 参照CTFS (Center for Tropical Forest Science)和中国森林生物多样性监测网络(CForBio)的大样地建立与监测技术规范(
Condit, 1995 )(
http://www.ctfs.si.edu/ ), 于2010-2011年在祁连山国家级自然保护区大野口流域的相对平坦的青海云杉林内建立了一块面积10.2 hm
2 的青海云杉动态监测样地.整个样地呈长方形, 以样地西北角为原点, 南北方向为
x 轴(长340 m), 东西方向为
y 轴(宽300 m), 利用全站仪将整个样地划分成255个20 m × 20 m的样方(附录I), 进一步将每个20 m × 20 m的样方细分成16个5 m × 5 m的小样方, 对小样方内所有胸径(
DBH ) ≥ 1.0 cm的木本个体及其分支进行涂漆, 挂牌标记以便进行永久监测, 测量其相对坐标(
x ,
y )、胸径、冠幅、枝下高, 记录生长状况(存活、倒伏或枯立)等信息.采用全站仪测定每个样方的海拔高度, 并绘制等高线地形图(附录II).首次调查结果表明, 样地内共记录到
DBH ≥ 1.0 cm的活立木19 927株, 枯死木411株(
李效雄等, 2013 ). ...
濒危植物梵净山冷杉野生种群结构及动态特征
1
2011
... 植物种群的径级结构是指不同径级组的个体在种群内所占比例和配置情况(
田润炜等, 2016 ), 是种群的生殖特性与外界环境相互作用的结果, 在一定程度上体现了种群的径级结构特征和生长发育状况, 进而可以预测种群和群落的动态发展趋势(
涂洪润等, 2019 ).本研究发现, 样地内青海云杉种群的幼苗和小树个体储备较为丰富, 其种群径级结构呈倒“J”型分布, 这说明该种群属增长型种群, 种群天然更新能力良好, 只要未来不遭受强烈干扰, 种群数量会保持逐渐增长.这与
李效雄等(2012) 和
张华等(2016) 分别对祁连山大野口流域和天老池小流域的青海云杉种群结构的研究结果一致.这可能是由青海云杉种群的生物学特性与环境因素共同作用的结果.已有研究表明, 森林更新过程中最重要和最敏感的阶段是幼苗阶段(
李晓笑等, 2011 ;
赵阳等, 2018 ), 林内有限的资源和空间往往成为种群更新的重要限制因素(
项小燕等, 2016 ).青海云杉种群的幼苗和小树个体数量多主要是由其生殖特性决定, 其种群可以产生大量高质量种子, 自然萌发形成的幼苗大都集中分布在林窗下, 形成大量小径级个体, 占据有利资源, 这是其种群在群落中持续处于优势地位的主要原因.随着小径级个体的生长, 它们对光照、水分、土壤养分等环境资源和空间需求随之增加, 由于环境资源和空间有限, 种内和种间竞争加剧, 出现强烈的自疏和他疏现象, 竞争能力弱的幼苗和小树在竞争中会大量死亡, 只有少量小径级个体能够成功转化为大径级个体, 造成样地内仅存在少量的中树和大树, 这成为该青海云杉种群更新和发展的“瓶颈” (
张婕等, 2014 ;
白卫国等, 2017 ;
于世川等, 2017 ).因此, 要实现种群数量的稳定增长, 必须加强对种群结构的保护, 尤其要重视对幼苗和小树及其生存环境的保护和改善. ...
濒危植物梵净山冷杉野生种群结构及动态特征
1
2011
... 植物种群的径级结构是指不同径级组的个体在种群内所占比例和配置情况(
田润炜等, 2016 ), 是种群的生殖特性与外界环境相互作用的结果, 在一定程度上体现了种群的径级结构特征和生长发育状况, 进而可以预测种群和群落的动态发展趋势(
涂洪润等, 2019 ).本研究发现, 样地内青海云杉种群的幼苗和小树个体储备较为丰富, 其种群径级结构呈倒“J”型分布, 这说明该种群属增长型种群, 种群天然更新能力良好, 只要未来不遭受强烈干扰, 种群数量会保持逐渐增长.这与
李效雄等(2012) 和
张华等(2016) 分别对祁连山大野口流域和天老池小流域的青海云杉种群结构的研究结果一致.这可能是由青海云杉种群的生物学特性与环境因素共同作用的结果.已有研究表明, 森林更新过程中最重要和最敏感的阶段是幼苗阶段(
李晓笑等, 2011 ;
赵阳等, 2018 ), 林内有限的资源和空间往往成为种群更新的重要限制因素(
项小燕等, 2016 ).青海云杉种群的幼苗和小树个体数量多主要是由其生殖特性决定, 其种群可以产生大量高质量种子, 自然萌发形成的幼苗大都集中分布在林窗下, 形成大量小径级个体, 占据有利资源, 这是其种群在群落中持续处于优势地位的主要原因.随着小径级个体的生长, 它们对光照、水分、土壤养分等环境资源和空间需求随之增加, 由于环境资源和空间有限, 种内和种间竞争加剧, 出现强烈的自疏和他疏现象, 竞争能力弱的幼苗和小树在竞争中会大量死亡, 只有少量小径级个体能够成功转化为大径级个体, 造成样地内仅存在少量的中树和大树, 这成为该青海云杉种群更新和发展的“瓶颈” (
张婕等, 2014 ;
白卫国等, 2017 ;
于世川等, 2017 ).因此, 要实现种群数量的稳定增长, 必须加强对种群结构的保护, 尤其要重视对幼苗和小树及其生存环境的保护和改善. ...
2013
2013
尖峰岭热带山地雨林优势树种白颜树空间分布格局
1
2014
... 种群内不同径级个体的空间关联性描述了某一特定的时间内植物种群组成个体之间的空间分布关系及功能关系(
Shields et al ., 2014 ;
王慧杰等, 2016 ).这种关系是种群内不同径级个体间、种群与环境长期相互作用的结果, 通过它可以揭示种群的现状并预测种群发展趋势(
涂洪润等, 2019 ).空间正关联反映了种群内部正向(互惠)的生态关系, 空间负关联意味着种群内部负向(互斥)的生态关系, 空间无关联则反映了种群内部无明确的生态关系(
张金屯, 1998 ).该研究发现小尺度(<4 m)下大树个体与其他径级个体都呈负关联, 随着尺度的增大, 大树与幼树表现出正关联且关联性随尺度的增大而减弱, 而与小树、中树则表现为无关联或者微弱的负关联.小尺度下大树与其他径级个体都呈负关联, 表明在一定范围内其他径级个体都受到大树的排斥, 这可能是因为大树个体对土壤水分、光等资源的需求量更大, 竞争能力更强, 从而使得其周围其他径级个体的分布密度非常小(
张华等, 2016 ).也有研究表明青海云杉凋落物对种子的萌发和幼苗的生长有自毒作用(
王丽红等, 2017 ), 而大树周围凋落物一般也较为丰富, 导致大树周围种子和幼苗的生长受到抑制.实际调查中也发现在大树周围尤其是林冠下其他径级个体数量很少.大树个体和幼树个体在较大尺度上表现出正关联且随着尺度的增大, 关联性逐渐减弱, 这很可能是由种子扩散限制所引起的(
梁爽等, 2014 ), 大树产生的种子数量较多, 幼树的分布在一定程度上能反映种子的分布, 大树和幼树的关联性变化特点与青海云杉种子主要通过风力传播的特点具有一致性.此外, 在较大尺度下, 大树个体与小树、中树个体都表现为无关联或者微弱的负关联, 一方面表明随着尺度的增大, 大树对小树和中树的影响逐渐降低, 竞争趋于和缓, 另一方面随着径级的增大, 大树与其他径级个体关联性也发生变化, 说明不同径级青海云杉空间关联性随着发育阶段而变化, 这可能是由种群自身的生物生态学特性、种内种间关系以及环境条件共同作用的结果. ...
尖峰岭热带山地雨林优势树种白颜树空间分布格局
1
2014
... 种群内不同径级个体的空间关联性描述了某一特定的时间内植物种群组成个体之间的空间分布关系及功能关系(
Shields et al ., 2014 ;
王慧杰等, 2016 ).这种关系是种群内不同径级个体间、种群与环境长期相互作用的结果, 通过它可以揭示种群的现状并预测种群发展趋势(
涂洪润等, 2019 ).空间正关联反映了种群内部正向(互惠)的生态关系, 空间负关联意味着种群内部负向(互斥)的生态关系, 空间无关联则反映了种群内部无明确的生态关系(
张金屯, 1998 ).该研究发现小尺度(<4 m)下大树个体与其他径级个体都呈负关联, 随着尺度的增大, 大树与幼树表现出正关联且关联性随尺度的增大而减弱, 而与小树、中树则表现为无关联或者微弱的负关联.小尺度下大树与其他径级个体都呈负关联, 表明在一定范围内其他径级个体都受到大树的排斥, 这可能是因为大树个体对土壤水分、光等资源的需求量更大, 竞争能力更强, 从而使得其周围其他径级个体的分布密度非常小(
张华等, 2016 ).也有研究表明青海云杉凋落物对种子的萌发和幼苗的生长有自毒作用(
王丽红等, 2017 ), 而大树周围凋落物一般也较为丰富, 导致大树周围种子和幼苗的生长受到抑制.实际调查中也发现在大树周围尤其是林冠下其他径级个体数量很少.大树个体和幼树个体在较大尺度上表现出正关联且随着尺度的增大, 关联性逐渐减弱, 这很可能是由种子扩散限制所引起的(
梁爽等, 2014 ), 大树产生的种子数量较多, 幼树的分布在一定程度上能反映种子的分布, 大树和幼树的关联性变化特点与青海云杉种子主要通过风力传播的特点具有一致性.此外, 在较大尺度下, 大树个体与小树、中树个体都表现为无关联或者微弱的负关联, 一方面表明随着尺度的增大, 大树对小树和中树的影响逐渐降低, 竞争趋于和缓, 另一方面随着径级的增大, 大树与其他径级个体关联性也发生变化, 说明不同径级青海云杉空间关联性随着发育阶段而变化, 这可能是由种群自身的生物生态学特性、种内种间关系以及环境条件共同作用的结果. ...
太白山红桦种群结构与空间分析
1
2008
... 种群的空间格局是指特定时间内, 种群所有个体在其水平生活空间内的分布状况及其属性在空间上的配置特征, 往往是由种群自身生物学特性、种内种间竞争、环境条件等因素综合作用的结果(
林玥, 2008 ;
Liu et al ., 2020 ).种群的空间分布格局和空间关联性是种群空间格局研究的2个主要内容, 也是种群生态关系在空间格局上的两种表现形式(
张金屯和孟东平, 2004 ).大量研究表明种群的空间分布格局与空间关联性关系密切, 聚集分布和空间正关联表明种群内部正向(互惠)的生态关系, 规则分布和空间负关联则表明种群内部负向(互斥)的生态关系, 随机分布和空间无关联则代表着种群内部不存在明确的生态关系(
杨洪晓, 2006 ).因此, 研究植物种群的空间分布格局及其关联性不仅能够了解种群的空间水平结构与生物、生态学习性(如生活史策略、是否喜光或耐阴等)、种群间相互作用、种群与环境因子的相互作用规律(如植物与生长环境间的适合度、生境异质性等)以及种群在群落中的地位和生态学功能, 更重要的是可以揭示这种分布格局及其关联性的潜在成因, 进而推演种群动态变化的生态学过程(如种内和种间竞争、种子扩散和干扰等), 深化对群落结构的认识, 揭示植物群落的生物多样性形成和维持机制(
McDonald et al ., 2003 ;
Hou et al , 2004 ;
Wiegand & Moloney, 2004 ;
Nathan, 2006 ;
Wang et al ., 2010 ;
Guo et al ., 2013 ). ...
太白山红桦种群结构与空间分析
1
2008
... 种群的空间格局是指特定时间内, 种群所有个体在其水平生活空间内的分布状况及其属性在空间上的配置特征, 往往是由种群自身生物学特性、种内种间竞争、环境条件等因素综合作用的结果(
林玥, 2008 ;
Liu et al ., 2020 ).种群的空间分布格局和空间关联性是种群空间格局研究的2个主要内容, 也是种群生态关系在空间格局上的两种表现形式(
张金屯和孟东平, 2004 ).大量研究表明种群的空间分布格局与空间关联性关系密切, 聚集分布和空间正关联表明种群内部正向(互惠)的生态关系, 规则分布和空间负关联则表明种群内部负向(互斥)的生态关系, 随机分布和空间无关联则代表着种群内部不存在明确的生态关系(
杨洪晓, 2006 ).因此, 研究植物种群的空间分布格局及其关联性不仅能够了解种群的空间水平结构与生物、生态学习性(如生活史策略、是否喜光或耐阴等)、种群间相互作用、种群与环境因子的相互作用规律(如植物与生长环境间的适合度、生境异质性等)以及种群在群落中的地位和生态学功能, 更重要的是可以揭示这种分布格局及其关联性的潜在成因, 进而推演种群动态变化的生态学过程(如种内和种间竞争、种子扩散和干扰等), 深化对群落结构的认识, 揭示植物群落的生物多样性形成和维持机制(
McDonald et al ., 2003 ;
Hou et al , 2004 ;
Wiegand & Moloney, 2004 ;
Nathan, 2006 ;
Wang et al ., 2010 ;
Guo et al ., 2013 ). ...
祁连山保护区青海云杉种群分布格局的研究
1
2004
... 森林的空间格局与空间尺度密切相关, 物种的空间分布格局随着尺度的变化而变化(
Condit et al ., 2000 ;
杨华等, 2014 ).一般来说, 小尺度上的物种分布格局主要受种群自身生物学特性(如种内种间竞争、种子扩散限制和幼苗补充过程等)影响, 而大尺度上物种的空间分布则主要受生境异质性(如光照、土壤养分、水分和温度等)影响(
Shen et al ., 2013 ).本研究表明, 青海云杉种群不同径级的青海云杉个体在较小尺度内均表现为聚集分布, 随着空间尺度的增加, 呈现由聚集分布到随机分布的变化趋势.这一结果与国内外大多数有关种群空间格局的研究结果一致(
刘建泉, 2004 ;
Guo et al ., 2013 ;
张觅等, 2014 ;
张华等, 2016 ).这是因为同一种群的不同个体对生境条件的需求基本上是一致的(
张金屯和孟东平, 2004 ), 且聚集分布有利于种群个体相互庇护, 从而抵御外界不利环境, 以提高存活率(
Diggle, 1981 ).此外, 生境的异质性容易引起种群聚集生长, 同时由于植株对光照、水分、养分等资源的竞争较大(
李海东等, 2011 ), 导致聚集强度随着空间尺度的增大而呈现下降趋势, 表现为在较小尺度上植株倾向于聚集分布逐渐过渡为在较大尺度上的随机分布. ...
祁连山保护区青海云杉种群分布格局的研究
1
2004
... 森林的空间格局与空间尺度密切相关, 物种的空间分布格局随着尺度的变化而变化(
Condit et al ., 2000 ;
杨华等, 2014 ).一般来说, 小尺度上的物种分布格局主要受种群自身生物学特性(如种内种间竞争、种子扩散限制和幼苗补充过程等)影响, 而大尺度上物种的空间分布则主要受生境异质性(如光照、土壤养分、水分和温度等)影响(
Shen et al ., 2013 ).本研究表明, 青海云杉种群不同径级的青海云杉个体在较小尺度内均表现为聚集分布, 随着空间尺度的增加, 呈现由聚集分布到随机分布的变化趋势.这一结果与国内外大多数有关种群空间格局的研究结果一致(
刘建泉, 2004 ;
Guo et al ., 2013 ;
张觅等, 2014 ;
张华等, 2016 ).这是因为同一种群的不同个体对生境条件的需求基本上是一致的(
张金屯和孟东平, 2004 ), 且聚集分布有利于种群个体相互庇护, 从而抵御外界不利环境, 以提高存活率(
Diggle, 1981 ).此外, 生境的异质性容易引起种群聚集生长, 同时由于植株对光照、水分、养分等资源的竞争较大(
李海东等, 2011 ), 导致聚集强度随着空间尺度的增大而呈现下降趋势, 表现为在较小尺度上植株倾向于聚集分布逐渐过渡为在较大尺度上的随机分布. ...
Competition and facilitation co-regulate the spatial patterns of boreal tree species in Kanas of Xinjiang, northwest China
1
2020
... 种群的空间格局是指特定时间内, 种群所有个体在其水平生活空间内的分布状况及其属性在空间上的配置特征, 往往是由种群自身生物学特性、种内种间竞争、环境条件等因素综合作用的结果(
林玥, 2008 ;
Liu et al ., 2020 ).种群的空间分布格局和空间关联性是种群空间格局研究的2个主要内容, 也是种群生态关系在空间格局上的两种表现形式(
张金屯和孟东平, 2004 ).大量研究表明种群的空间分布格局与空间关联性关系密切, 聚集分布和空间正关联表明种群内部正向(互惠)的生态关系, 规则分布和空间负关联则表明种群内部负向(互斥)的生态关系, 随机分布和空间无关联则代表着种群内部不存在明确的生态关系(
杨洪晓, 2006 ).因此, 研究植物种群的空间分布格局及其关联性不仅能够了解种群的空间水平结构与生物、生态学习性(如生活史策略、是否喜光或耐阴等)、种群间相互作用、种群与环境因子的相互作用规律(如植物与生长环境间的适合度、生境异质性等)以及种群在群落中的地位和生态学功能, 更重要的是可以揭示这种分布格局及其关联性的潜在成因, 进而推演种群动态变化的生态学过程(如种内和种间竞争、种子扩散和干扰等), 深化对群落结构的认识, 揭示植物群落的生物多样性形成和维持机制(
McDonald et al ., 2003 ;
Hou et al , 2004 ;
Wiegand & Moloney, 2004 ;
Nathan, 2006 ;
Wang et al ., 2010 ;
Guo et al ., 2013 ). ...
Spatial pattern of
Quercus regeneration limitation and
Acer rubrum invasion in a Piedmont forest
1
2003
... 种群的空间格局是指特定时间内, 种群所有个体在其水平生活空间内的分布状况及其属性在空间上的配置特征, 往往是由种群自身生物学特性、种内种间竞争、环境条件等因素综合作用的结果(
林玥, 2008 ;
Liu et al ., 2020 ).种群的空间分布格局和空间关联性是种群空间格局研究的2个主要内容, 也是种群生态关系在空间格局上的两种表现形式(
张金屯和孟东平, 2004 ).大量研究表明种群的空间分布格局与空间关联性关系密切, 聚集分布和空间正关联表明种群内部正向(互惠)的生态关系, 规则分布和空间负关联则表明种群内部负向(互斥)的生态关系, 随机分布和空间无关联则代表着种群内部不存在明确的生态关系(
杨洪晓, 2006 ).因此, 研究植物种群的空间分布格局及其关联性不仅能够了解种群的空间水平结构与生物、生态学习性(如生活史策略、是否喜光或耐阴等)、种群间相互作用、种群与环境因子的相互作用规律(如植物与生长环境间的适合度、生境异质性等)以及种群在群落中的地位和生态学功能, 更重要的是可以揭示这种分布格局及其关联性的潜在成因, 进而推演种群动态变化的生态学过程(如种内和种间竞争、种子扩散和干扰等), 深化对群落结构的认识, 揭示植物群落的生物多样性形成和维持机制(
McDonald et al ., 2003 ;
Hou et al , 2004 ;
Wiegand & Moloney, 2004 ;
Nathan, 2006 ;
Wang et al ., 2010 ;
Guo et al ., 2013 ). ...
Long-distance dispersal of plants
1
2006
... 种群的空间格局是指特定时间内, 种群所有个体在其水平生活空间内的分布状况及其属性在空间上的配置特征, 往往是由种群自身生物学特性、种内种间竞争、环境条件等因素综合作用的结果(
林玥, 2008 ;
Liu et al ., 2020 ).种群的空间分布格局和空间关联性是种群空间格局研究的2个主要内容, 也是种群生态关系在空间格局上的两种表现形式(
张金屯和孟东平, 2004 ).大量研究表明种群的空间分布格局与空间关联性关系密切, 聚集分布和空间正关联表明种群内部正向(互惠)的生态关系, 规则分布和空间负关联则表明种群内部负向(互斥)的生态关系, 随机分布和空间无关联则代表着种群内部不存在明确的生态关系(
杨洪晓, 2006 ).因此, 研究植物种群的空间分布格局及其关联性不仅能够了解种群的空间水平结构与生物、生态学习性(如生活史策略、是否喜光或耐阴等)、种群间相互作用、种群与环境因子的相互作用规律(如植物与生长环境间的适合度、生境异质性等)以及种群在群落中的地位和生态学功能, 更重要的是可以揭示这种分布格局及其关联性的潜在成因, 进而推演种群动态变化的生态学过程(如种内和种间竞争、种子扩散和干扰等), 深化对群落结构的认识, 揭示植物群落的生物多样性形成和维持机制(
McDonald et al ., 2003 ;
Hou et al , 2004 ;
Wiegand & Moloney, 2004 ;
Nathan, 2006 ;
Wang et al ., 2010 ;
Guo et al ., 2013 ). ...
The second-order analysis of stationary point processes
1976
Changes of woody plant interaction and spatial distribution between rocky and sandy soil areas in a semi-arid savanna, South Africa
1
2011
... 空间分布格局的研究方法众多, 早期的研究方法主要有: 以频度/密度为基础的样方法、以距离为基础的无样地法、方差区组分析法、双向轨迹方差分析、
χ 2 检验和分形理论等(
张金屯, 1998 ;
王本洋和余世孝, 2005 ).传统的样方取样和分析方法常基于单一尺度(固定大小样方)的若干样方, 常采用
χ 2 分析法和方差/均值比率法等统计分析方法, 然而种群空间分布格局及其关联性对种群密度和空间尺度均有很强的依赖性, 在不同的尺度上往往表现出不同的类型, 故早期的取样和分析方法只能研究特定尺度下种群的空间分布格局特征, 难以全面地反映森林内物种的空间分布特点和种间关系(
He et al ., 1997 ;
李立等, 2010 ).鉴于上述研究方法的局限性,
Ripley (1977) 和
Diggle (1983) 分别提出和完善了点格局分析法, 以分析各种尺度上的空间分布格局.该方法以数字化的植物个体位置信息为基本数据, 将每个个体都视为二维空间中的一个点, 所有个体组成了植物种的空间分布图, 然后在空间分布图的基础上进行种群空间格局分析.其优点是能最大限度地利用点与点之间的距离, 不仅能够提供全面的空间格局信息, 而且具有较强的检验能力, 克服了传统研究方法中尺度单一等不足, 可以分析任意尺度下种群的空间格局特征及种内、种间关系(
Wiegand & Moloney, 2004 ;
Schleicher et al ., 2011 ;
沈志强等, 2016 ;
涂洪润等, 2019 ).因此, 空间点格局分析法在种群空间格局分布特征及潜在生态过程假说的研究方面得到了广泛应用. ...
Quantifying effects of habitat heterogeneity and other clustering processes on spatial distributions of tree species
1
2013
... 森林的空间格局与空间尺度密切相关, 物种的空间分布格局随着尺度的变化而变化(
Condit et al ., 2000 ;
杨华等, 2014 ).一般来说, 小尺度上的物种分布格局主要受种群自身生物学特性(如种内种间竞争、种子扩散限制和幼苗补充过程等)影响, 而大尺度上物种的空间分布则主要受生境异质性(如光照、土壤养分、水分和温度等)影响(
Shen et al ., 2013 ).本研究表明, 青海云杉种群不同径级的青海云杉个体在较小尺度内均表现为聚集分布, 随着空间尺度的增加, 呈现由聚集分布到随机分布的变化趋势.这一结果与国内外大多数有关种群空间格局的研究结果一致(
刘建泉, 2004 ;
Guo et al ., 2013 ;
张觅等, 2014 ;
张华等, 2016 ).这是因为同一种群的不同个体对生境条件的需求基本上是一致的(
张金屯和孟东平, 2004 ), 且聚集分布有利于种群个体相互庇护, 从而抵御外界不利环境, 以提高存活率(
Diggle, 1981 ).此外, 生境的异质性容易引起种群聚集生长, 同时由于植株对光照、水分、养分等资源的竞争较大(
李海东等, 2011 ), 导致聚集强度随着空间尺度的增大而呈现下降趋势, 表现为在较小尺度上植株倾向于聚集分布逐渐过渡为在较大尺度上的随机分布. ...
藏东南川滇高山栎种群不同生长阶段的空间格局与关联性
1
2016
... 空间分布格局的研究方法众多, 早期的研究方法主要有: 以频度/密度为基础的样方法、以距离为基础的无样地法、方差区组分析法、双向轨迹方差分析、
χ 2 检验和分形理论等(
张金屯, 1998 ;
王本洋和余世孝, 2005 ).传统的样方取样和分析方法常基于单一尺度(固定大小样方)的若干样方, 常采用
χ 2 分析法和方差/均值比率法等统计分析方法, 然而种群空间分布格局及其关联性对种群密度和空间尺度均有很强的依赖性, 在不同的尺度上往往表现出不同的类型, 故早期的取样和分析方法只能研究特定尺度下种群的空间分布格局特征, 难以全面地反映森林内物种的空间分布特点和种间关系(
He et al ., 1997 ;
李立等, 2010 ).鉴于上述研究方法的局限性,
Ripley (1977) 和
Diggle (1983) 分别提出和完善了点格局分析法, 以分析各种尺度上的空间分布格局.该方法以数字化的植物个体位置信息为基本数据, 将每个个体都视为二维空间中的一个点, 所有个体组成了植物种的空间分布图, 然后在空间分布图的基础上进行种群空间格局分析.其优点是能最大限度地利用点与点之间的距离, 不仅能够提供全面的空间格局信息, 而且具有较强的检验能力, 克服了传统研究方法中尺度单一等不足, 可以分析任意尺度下种群的空间格局特征及种内、种间关系(
Wiegand & Moloney, 2004 ;
Schleicher et al ., 2011 ;
沈志强等, 2016 ;
涂洪润等, 2019 ).因此, 空间点格局分析法在种群空间格局分布特征及潜在生态过程假说的研究方面得到了广泛应用. ...
藏东南川滇高山栎种群不同生长阶段的空间格局与关联性
1
2016
... 空间分布格局的研究方法众多, 早期的研究方法主要有: 以频度/密度为基础的样方法、以距离为基础的无样地法、方差区组分析法、双向轨迹方差分析、
χ 2 检验和分形理论等(
张金屯, 1998 ;
王本洋和余世孝, 2005 ).传统的样方取样和分析方法常基于单一尺度(固定大小样方)的若干样方, 常采用
χ 2 分析法和方差/均值比率法等统计分析方法, 然而种群空间分布格局及其关联性对种群密度和空间尺度均有很强的依赖性, 在不同的尺度上往往表现出不同的类型, 故早期的取样和分析方法只能研究特定尺度下种群的空间分布格局特征, 难以全面地反映森林内物种的空间分布特点和种间关系(
He et al ., 1997 ;
李立等, 2010 ).鉴于上述研究方法的局限性,
Ripley (1977) 和
Diggle (1983) 分别提出和完善了点格局分析法, 以分析各种尺度上的空间分布格局.该方法以数字化的植物个体位置信息为基本数据, 将每个个体都视为二维空间中的一个点, 所有个体组成了植物种的空间分布图, 然后在空间分布图的基础上进行种群空间格局分析.其优点是能最大限度地利用点与点之间的距离, 不仅能够提供全面的空间格局信息, 而且具有较强的检验能力, 克服了传统研究方法中尺度单一等不足, 可以分析任意尺度下种群的空间格局特征及种内、种间关系(
Wiegand & Moloney, 2004 ;
Schleicher et al ., 2011 ;
沈志强等, 2016 ;
涂洪润等, 2019 ).因此, 空间点格局分析法在种群空间格局分布特征及潜在生态过程假说的研究方面得到了广泛应用. ...
Age distribution and spatial patterning of an invasive shrub in secondary hardwood forests
1
2014
... 种群内不同径级个体的空间关联性描述了某一特定的时间内植物种群组成个体之间的空间分布关系及功能关系(
Shields et al ., 2014 ;
王慧杰等, 2016 ).这种关系是种群内不同径级个体间、种群与环境长期相互作用的结果, 通过它可以揭示种群的现状并预测种群发展趋势(
涂洪润等, 2019 ).空间正关联反映了种群内部正向(互惠)的生态关系, 空间负关联意味着种群内部负向(互斥)的生态关系, 空间无关联则反映了种群内部无明确的生态关系(
张金屯, 1998 ).该研究发现小尺度(<4 m)下大树个体与其他径级个体都呈负关联, 随着尺度的增大, 大树与幼树表现出正关联且关联性随尺度的增大而减弱, 而与小树、中树则表现为无关联或者微弱的负关联.小尺度下大树与其他径级个体都呈负关联, 表明在一定范围内其他径级个体都受到大树的排斥, 这可能是因为大树个体对土壤水分、光等资源的需求量更大, 竞争能力更强, 从而使得其周围其他径级个体的分布密度非常小(
张华等, 2016 ).也有研究表明青海云杉凋落物对种子的萌发和幼苗的生长有自毒作用(
王丽红等, 2017 ), 而大树周围凋落物一般也较为丰富, 导致大树周围种子和幼苗的生长受到抑制.实际调查中也发现在大树周围尤其是林冠下其他径级个体数量很少.大树个体和幼树个体在较大尺度上表现出正关联且随着尺度的增大, 关联性逐渐减弱, 这很可能是由种子扩散限制所引起的(
梁爽等, 2014 ), 大树产生的种子数量较多, 幼树的分布在一定程度上能反映种子的分布, 大树和幼树的关联性变化特点与青海云杉种子主要通过风力传播的特点具有一致性.此外, 在较大尺度下, 大树个体与小树、中树个体都表现为无关联或者微弱的负关联, 一方面表明随着尺度的增大, 大树对小树和中树的影响逐渐降低, 竞争趋于和缓, 另一方面随着径级的增大, 大树与其他径级个体关联性也发生变化, 说明不同径级青海云杉空间关联性随着发育阶段而变化, 这可能是由种群自身的生物生态学特性、种内种间关系以及环境条件共同作用的结果. ...
Recent applications of point process methods in forestry statistics
1
2000
... 由于
g (
r )的非聚集属性, 利用它进行点格局分析时需利用零模型来观察实际过程和理论过程之间的差异(
Stoyan & Penttinen, 2000 ).祁连山大野口流域青海云杉林样地地势相对平坦, 地形因子对于种群空间格局及其关联性影响较弱, 因此采用完全空间随机零模型作为零模型, 即假设物种的空间分布完全随机, 没有潜在的生物学过程发挥作用, 观察青海云杉种群不同径级个体的空间分布格局及其关联性与匀质泊松分布是否存在差异(
张觅等, 2014 ).空间尺度设定为0-60 m, 步长为1 m.采用蒙特卡罗(Monte-Carlo)随机模拟999次产生置信度为99%的包迹线, 以检验物种分布格局及关联性是否显著.在单变量分析中, 若实际分布的
g (
r )值落在包迹线之内, 则符合零模型, 为随机分布; 若在上包迹线以上, 则呈聚集分布; 若在下包迹线以下, 则为规则分布.在双变量分析中, 若实际分布的
g 12 (
r )值落在包迹线之间, 表示接受零假设, 说明不同径级间个体相互独立, 无空间相关性; 若在上包迹线以上, 则两径级间为正相关; 若落在下包迹线以下, 则两径级间为负相关关系(
Hao et al ., 2017 ).该研究所有统计与分析均在R 4.0.1软件中“spatstat”程序包(
Baddeley & Turner, 2005 )的相关函数中完成. ...
祁连山青海云杉林冠生态水文效应及其影响因素
1
2012
... 祁连山是中国西部的主要山脉之一, 位于青藏高原、蒙古高原、黄土高原三大高原交汇带, 是石羊河、黑河、疏勒河等6大内陆河和黄河流域的重要水源产流地, 维系着河西乃至西部地区生存与发展的命脉, 在国家生态建设中具有十分重要的战略地位(
Gao et al ., 2013 ;
Zhu et al ., 2017 ).祁连山山地森林的主要建群种——青海云杉(
Picea crassifolia ), 为松科云杉属(
Picea )乔木, 是中国青藏高原东北边缘特有树种, 多以纯林形式存在, 呈带状或斑块状分布于海拔2 300-3 300 m的阴坡和半阴坡, 分布面积占祁连山水源涵养林面积的24.74%, 占乔木林面积的75.72%, 对于维系祁连山的生物多样性、水源涵养功能乃至区域生态平衡都具有至关重要的作用(
田风霞等, 2012 ;
何芳兰等, 2016 ;
张华等, 2016 ).部分****(
赵丽琼等, 2010 ;
张华等, 2016 )虽然已对青海云杉种群空间格局及其关联性进行了研究, 但是已有研究的空间尺度相对较小, 仅能揭示小尺度下的种群空间分布格局, 难以客观地反映青海云杉种群空间分布的特点和种内关系.鉴于此, 本研究基于祁连山大野口流域10.2 hm
2 青海云杉动态监测样地数据, 采用空间点格局分析方法中的成对相关函数对不同径级青海云杉种群的空间分布格局及其关联性进行分析, 旨在阐明: (1)祁连山大野口流域青海云杉种群不同径级个体的空间分布格局及其关联性如何随空间尺度进行变化; (2)不同径级个体的空间分布格局及其关联性形成的潜在生态学过程与影响机制.上述科学问题的合理解答, 能从空间格局的角度客观地揭示青海云杉种群发展规律, 可以为制定该种群的配置、种植与保护管理措施提供指导, 并对祁连山地区的植被恢复与重建、生物多样性保护等具有重要的意义和价值. ...
祁连山青海云杉林冠生态水文效应及其影响因素
1
2012
... 祁连山是中国西部的主要山脉之一, 位于青藏高原、蒙古高原、黄土高原三大高原交汇带, 是石羊河、黑河、疏勒河等6大内陆河和黄河流域的重要水源产流地, 维系着河西乃至西部地区生存与发展的命脉, 在国家生态建设中具有十分重要的战略地位(
Gao et al ., 2013 ;
Zhu et al ., 2017 ).祁连山山地森林的主要建群种——青海云杉(
Picea crassifolia ), 为松科云杉属(
Picea )乔木, 是中国青藏高原东北边缘特有树种, 多以纯林形式存在, 呈带状或斑块状分布于海拔2 300-3 300 m的阴坡和半阴坡, 分布面积占祁连山水源涵养林面积的24.74%, 占乔木林面积的75.72%, 对于维系祁连山的生物多样性、水源涵养功能乃至区域生态平衡都具有至关重要的作用(
田风霞等, 2012 ;
何芳兰等, 2016 ;
张华等, 2016 ).部分****(
赵丽琼等, 2010 ;
张华等, 2016 )虽然已对青海云杉种群空间格局及其关联性进行了研究, 但是已有研究的空间尺度相对较小, 仅能揭示小尺度下的种群空间分布格局, 难以客观地反映青海云杉种群空间分布的特点和种内关系.鉴于此, 本研究基于祁连山大野口流域10.2 hm
2 青海云杉动态监测样地数据, 采用空间点格局分析方法中的成对相关函数对不同径级青海云杉种群的空间分布格局及其关联性进行分析, 旨在阐明: (1)祁连山大野口流域青海云杉种群不同径级个体的空间分布格局及其关联性如何随空间尺度进行变化; (2)不同径级个体的空间分布格局及其关联性形成的潜在生态学过程与影响机制.上述科学问题的合理解答, 能从空间格局的角度客观地揭示青海云杉种群发展规律, 可以为制定该种群的配置、种植与保护管理措施提供指导, 并对祁连山地区的植被恢复与重建、生物多样性保护等具有重要的意义和价值. ...
新疆野苹果种群年龄结构特征与动态分析
1
2016
... 植物种群的径级结构是指不同径级组的个体在种群内所占比例和配置情况(
田润炜等, 2016 ), 是种群的生殖特性与外界环境相互作用的结果, 在一定程度上体现了种群的径级结构特征和生长发育状况, 进而可以预测种群和群落的动态发展趋势(
涂洪润等, 2019 ).本研究发现, 样地内青海云杉种群的幼苗和小树个体储备较为丰富, 其种群径级结构呈倒“J”型分布, 这说明该种群属增长型种群, 种群天然更新能力良好, 只要未来不遭受强烈干扰, 种群数量会保持逐渐增长.这与
李效雄等(2012) 和
张华等(2016) 分别对祁连山大野口流域和天老池小流域的青海云杉种群结构的研究结果一致.这可能是由青海云杉种群的生物学特性与环境因素共同作用的结果.已有研究表明, 森林更新过程中最重要和最敏感的阶段是幼苗阶段(
李晓笑等, 2011 ;
赵阳等, 2018 ), 林内有限的资源和空间往往成为种群更新的重要限制因素(
项小燕等, 2016 ).青海云杉种群的幼苗和小树个体数量多主要是由其生殖特性决定, 其种群可以产生大量高质量种子, 自然萌发形成的幼苗大都集中分布在林窗下, 形成大量小径级个体, 占据有利资源, 这是其种群在群落中持续处于优势地位的主要原因.随着小径级个体的生长, 它们对光照、水分、土壤养分等环境资源和空间需求随之增加, 由于环境资源和空间有限, 种内和种间竞争加剧, 出现强烈的自疏和他疏现象, 竞争能力弱的幼苗和小树在竞争中会大量死亡, 只有少量小径级个体能够成功转化为大径级个体, 造成样地内仅存在少量的中树和大树, 这成为该青海云杉种群更新和发展的“瓶颈” (
张婕等, 2014 ;
白卫国等, 2017 ;
于世川等, 2017 ).因此, 要实现种群数量的稳定增长, 必须加强对种群结构的保护, 尤其要重视对幼苗和小树及其生存环境的保护和改善. ...
新疆野苹果种群年龄结构特征与动态分析
1
2016
... 植物种群的径级结构是指不同径级组的个体在种群内所占比例和配置情况(
田润炜等, 2016 ), 是种群的生殖特性与外界环境相互作用的结果, 在一定程度上体现了种群的径级结构特征和生长发育状况, 进而可以预测种群和群落的动态发展趋势(
涂洪润等, 2019 ).本研究发现, 样地内青海云杉种群的幼苗和小树个体储备较为丰富, 其种群径级结构呈倒“J”型分布, 这说明该种群属增长型种群, 种群天然更新能力良好, 只要未来不遭受强烈干扰, 种群数量会保持逐渐增长.这与
李效雄等(2012) 和
张华等(2016) 分别对祁连山大野口流域和天老池小流域的青海云杉种群结构的研究结果一致.这可能是由青海云杉种群的生物学特性与环境因素共同作用的结果.已有研究表明, 森林更新过程中最重要和最敏感的阶段是幼苗阶段(
李晓笑等, 2011 ;
赵阳等, 2018 ), 林内有限的资源和空间往往成为种群更新的重要限制因素(
项小燕等, 2016 ).青海云杉种群的幼苗和小树个体数量多主要是由其生殖特性决定, 其种群可以产生大量高质量种子, 自然萌发形成的幼苗大都集中分布在林窗下, 形成大量小径级个体, 占据有利资源, 这是其种群在群落中持续处于优势地位的主要原因.随着小径级个体的生长, 它们对光照、水分、土壤养分等环境资源和空间需求随之增加, 由于环境资源和空间有限, 种内和种间竞争加剧, 出现强烈的自疏和他疏现象, 竞争能力弱的幼苗和小树在竞争中会大量死亡, 只有少量小径级个体能够成功转化为大径级个体, 造成样地内仅存在少量的中树和大树, 这成为该青海云杉种群更新和发展的“瓶颈” (
张婕等, 2014 ;
白卫国等, 2017 ;
于世川等, 2017 ).因此, 要实现种群数量的稳定增长, 必须加强对种群结构的保护, 尤其要重视对幼苗和小树及其生存环境的保护和改善. ...
桂林岩溶石山檵木种群空间格局及其关联性
5
2019
... 空间分布格局的研究方法众多, 早期的研究方法主要有: 以频度/密度为基础的样方法、以距离为基础的无样地法、方差区组分析法、双向轨迹方差分析、
χ 2 检验和分形理论等(
张金屯, 1998 ;
王本洋和余世孝, 2005 ).传统的样方取样和分析方法常基于单一尺度(固定大小样方)的若干样方, 常采用
χ 2 分析法和方差/均值比率法等统计分析方法, 然而种群空间分布格局及其关联性对种群密度和空间尺度均有很强的依赖性, 在不同的尺度上往往表现出不同的类型, 故早期的取样和分析方法只能研究特定尺度下种群的空间分布格局特征, 难以全面地反映森林内物种的空间分布特点和种间关系(
He et al ., 1997 ;
李立等, 2010 ).鉴于上述研究方法的局限性,
Ripley (1977) 和
Diggle (1983) 分别提出和完善了点格局分析法, 以分析各种尺度上的空间分布格局.该方法以数字化的植物个体位置信息为基本数据, 将每个个体都视为二维空间中的一个点, 所有个体组成了植物种的空间分布图, 然后在空间分布图的基础上进行种群空间格局分析.其优点是能最大限度地利用点与点之间的距离, 不仅能够提供全面的空间格局信息, 而且具有较强的检验能力, 克服了传统研究方法中尺度单一等不足, 可以分析任意尺度下种群的空间格局特征及种内、种间关系(
Wiegand & Moloney, 2004 ;
Schleicher et al ., 2011 ;
沈志强等, 2016 ;
涂洪润等, 2019 ).因此, 空间点格局分析法在种群空间格局分布特征及潜在生态过程假说的研究方面得到了广泛应用. ...
... 植物种群的径级结构是指不同径级组的个体在种群内所占比例和配置情况(
田润炜等, 2016 ), 是种群的生殖特性与外界环境相互作用的结果, 在一定程度上体现了种群的径级结构特征和生长发育状况, 进而可以预测种群和群落的动态发展趋势(
涂洪润等, 2019 ).本研究发现, 样地内青海云杉种群的幼苗和小树个体储备较为丰富, 其种群径级结构呈倒“J”型分布, 这说明该种群属增长型种群, 种群天然更新能力良好, 只要未来不遭受强烈干扰, 种群数量会保持逐渐增长.这与
李效雄等(2012) 和
张华等(2016) 分别对祁连山大野口流域和天老池小流域的青海云杉种群结构的研究结果一致.这可能是由青海云杉种群的生物学特性与环境因素共同作用的结果.已有研究表明, 森林更新过程中最重要和最敏感的阶段是幼苗阶段(
李晓笑等, 2011 ;
赵阳等, 2018 ), 林内有限的资源和空间往往成为种群更新的重要限制因素(
项小燕等, 2016 ).青海云杉种群的幼苗和小树个体数量多主要是由其生殖特性决定, 其种群可以产生大量高质量种子, 自然萌发形成的幼苗大都集中分布在林窗下, 形成大量小径级个体, 占据有利资源, 这是其种群在群落中持续处于优势地位的主要原因.随着小径级个体的生长, 它们对光照、水分、土壤养分等环境资源和空间需求随之增加, 由于环境资源和空间有限, 种内和种间竞争加剧, 出现强烈的自疏和他疏现象, 竞争能力弱的幼苗和小树在竞争中会大量死亡, 只有少量小径级个体能够成功转化为大径级个体, 造成样地内仅存在少量的中树和大树, 这成为该青海云杉种群更新和发展的“瓶颈” (
张婕等, 2014 ;
白卫国等, 2017 ;
于世川等, 2017 ).因此, 要实现种群数量的稳定增长, 必须加强对种群结构的保护, 尤其要重视对幼苗和小树及其生存环境的保护和改善. ...
... 种群内不同径级个体的空间关联性描述了某一特定的时间内植物种群组成个体之间的空间分布关系及功能关系(
Shields et al ., 2014 ;
王慧杰等, 2016 ).这种关系是种群内不同径级个体间、种群与环境长期相互作用的结果, 通过它可以揭示种群的现状并预测种群发展趋势(
涂洪润等, 2019 ).空间正关联反映了种群内部正向(互惠)的生态关系, 空间负关联意味着种群内部负向(互斥)的生态关系, 空间无关联则反映了种群内部无明确的生态关系(
张金屯, 1998 ).该研究发现小尺度(<4 m)下大树个体与其他径级个体都呈负关联, 随着尺度的增大, 大树与幼树表现出正关联且关联性随尺度的增大而减弱, 而与小树、中树则表现为无关联或者微弱的负关联.小尺度下大树与其他径级个体都呈负关联, 表明在一定范围内其他径级个体都受到大树的排斥, 这可能是因为大树个体对土壤水分、光等资源的需求量更大, 竞争能力更强, 从而使得其周围其他径级个体的分布密度非常小(
张华等, 2016 ).也有研究表明青海云杉凋落物对种子的萌发和幼苗的生长有自毒作用(
王丽红等, 2017 ), 而大树周围凋落物一般也较为丰富, 导致大树周围种子和幼苗的生长受到抑制.实际调查中也发现在大树周围尤其是林冠下其他径级个体数量很少.大树个体和幼树个体在较大尺度上表现出正关联且随着尺度的增大, 关联性逐渐减弱, 这很可能是由种子扩散限制所引起的(
梁爽等, 2014 ), 大树产生的种子数量较多, 幼树的分布在一定程度上能反映种子的分布, 大树和幼树的关联性变化特点与青海云杉种子主要通过风力传播的特点具有一致性.此外, 在较大尺度下, 大树个体与小树、中树个体都表现为无关联或者微弱的负关联, 一方面表明随着尺度的增大, 大树对小树和中树的影响逐渐降低, 竞争趋于和缓, 另一方面随着径级的增大, 大树与其他径级个体关联性也发生变化, 说明不同径级青海云杉空间关联性随着发育阶段而变化, 这可能是由种群自身的生物生态学特性、种内种间关系以及环境条件共同作用的结果. ...
... 该研究还发现径级相近的个体空间关联性呈正关联或无关联, 随着个体径级间差距增大, 它们之间的空间关联性转变为无关联或负关联, 这说明同一物种不同径级的个体之间同时存在竞争、依赖以及无明确的生态关系, 这是由于物种具有一致的生境偏好所造成的(
杨洪晓等, 2006 ).径级相近个体的空间关联性呈正关联, 表明它们相伴出现, 对生境的选择具有一致性, 在空间分布上是相互依存的.这可能是由于它们生长发育阶段相近, 对环境资源的需求具有相似性和生态位存在重叠, 但小径级个体相对幼小, 对光照、水分和土壤养分等资源的竞争能力相对较弱, 需要借助高大植株的庇护, 才得以提高存活率, 而较大径级的植株个体可以改善样地的小生境, 使得邻近径级的个体更适宜生存, 彼此共享资源(
涂洪润等, 2019 ).同时, 由于物种专一性的资源分布不均, 种内竞争具有非对称性的特点, 也就是说当径级相近时, 不同径级间关系较协调(
Getzin et al ., 2008 ).而无关联则说明个体间互不干扰, 个体在前期发育过程中已经占据适宜的生态位, 与种内其他径级个体间的关联性不高, 能各自生长发育(
涂洪润等, 2019 ).随着个体间径级差距增加, 它们之间的关联性转变成无关联甚至负关联, 说明随着个体间径级差距的加大, 它们的空间正关联关系逐渐减弱, 当个体间差异较大时, 大径级个体在竞争中处于优势地位, 可能导致小径级个体的生长发育迟滞或者死亡, 使其周围小径级个体很少(
Kenkel, 1988 ), 这也间接证实了随着个体大小的生长变化, 青海云杉种群的空间分布存在一个从聚集分布到随机分布的自疏过程. ...
... ).而无关联则说明个体间互不干扰, 个体在前期发育过程中已经占据适宜的生态位, 与种内其他径级个体间的关联性不高, 能各自生长发育(
涂洪润等, 2019 ).随着个体间径级差距增加, 它们之间的关联性转变成无关联甚至负关联, 说明随着个体间径级差距的加大, 它们的空间正关联关系逐渐减弱, 当个体间差异较大时, 大径级个体在竞争中处于优势地位, 可能导致小径级个体的生长发育迟滞或者死亡, 使其周围小径级个体很少(
Kenkel, 1988 ), 这也间接证实了随着个体大小的生长变化, 青海云杉种群的空间分布存在一个从聚集分布到随机分布的自疏过程. ...
桂林岩溶石山檵木种群空间格局及其关联性
5
2019
... 空间分布格局的研究方法众多, 早期的研究方法主要有: 以频度/密度为基础的样方法、以距离为基础的无样地法、方差区组分析法、双向轨迹方差分析、
χ 2 检验和分形理论等(
张金屯, 1998 ;
王本洋和余世孝, 2005 ).传统的样方取样和分析方法常基于单一尺度(固定大小样方)的若干样方, 常采用
χ 2 分析法和方差/均值比率法等统计分析方法, 然而种群空间分布格局及其关联性对种群密度和空间尺度均有很强的依赖性, 在不同的尺度上往往表现出不同的类型, 故早期的取样和分析方法只能研究特定尺度下种群的空间分布格局特征, 难以全面地反映森林内物种的空间分布特点和种间关系(
He et al ., 1997 ;
李立等, 2010 ).鉴于上述研究方法的局限性,
Ripley (1977) 和
Diggle (1983) 分别提出和完善了点格局分析法, 以分析各种尺度上的空间分布格局.该方法以数字化的植物个体位置信息为基本数据, 将每个个体都视为二维空间中的一个点, 所有个体组成了植物种的空间分布图, 然后在空间分布图的基础上进行种群空间格局分析.其优点是能最大限度地利用点与点之间的距离, 不仅能够提供全面的空间格局信息, 而且具有较强的检验能力, 克服了传统研究方法中尺度单一等不足, 可以分析任意尺度下种群的空间格局特征及种内、种间关系(
Wiegand & Moloney, 2004 ;
Schleicher et al ., 2011 ;
沈志强等, 2016 ;
涂洪润等, 2019 ).因此, 空间点格局分析法在种群空间格局分布特征及潜在生态过程假说的研究方面得到了广泛应用. ...
... 植物种群的径级结构是指不同径级组的个体在种群内所占比例和配置情况(
田润炜等, 2016 ), 是种群的生殖特性与外界环境相互作用的结果, 在一定程度上体现了种群的径级结构特征和生长发育状况, 进而可以预测种群和群落的动态发展趋势(
涂洪润等, 2019 ).本研究发现, 样地内青海云杉种群的幼苗和小树个体储备较为丰富, 其种群径级结构呈倒“J”型分布, 这说明该种群属增长型种群, 种群天然更新能力良好, 只要未来不遭受强烈干扰, 种群数量会保持逐渐增长.这与
李效雄等(2012) 和
张华等(2016) 分别对祁连山大野口流域和天老池小流域的青海云杉种群结构的研究结果一致.这可能是由青海云杉种群的生物学特性与环境因素共同作用的结果.已有研究表明, 森林更新过程中最重要和最敏感的阶段是幼苗阶段(
李晓笑等, 2011 ;
赵阳等, 2018 ), 林内有限的资源和空间往往成为种群更新的重要限制因素(
项小燕等, 2016 ).青海云杉种群的幼苗和小树个体数量多主要是由其生殖特性决定, 其种群可以产生大量高质量种子, 自然萌发形成的幼苗大都集中分布在林窗下, 形成大量小径级个体, 占据有利资源, 这是其种群在群落中持续处于优势地位的主要原因.随着小径级个体的生长, 它们对光照、水分、土壤养分等环境资源和空间需求随之增加, 由于环境资源和空间有限, 种内和种间竞争加剧, 出现强烈的自疏和他疏现象, 竞争能力弱的幼苗和小树在竞争中会大量死亡, 只有少量小径级个体能够成功转化为大径级个体, 造成样地内仅存在少量的中树和大树, 这成为该青海云杉种群更新和发展的“瓶颈” (
张婕等, 2014 ;
白卫国等, 2017 ;
于世川等, 2017 ).因此, 要实现种群数量的稳定增长, 必须加强对种群结构的保护, 尤其要重视对幼苗和小树及其生存环境的保护和改善. ...
... 种群内不同径级个体的空间关联性描述了某一特定的时间内植物种群组成个体之间的空间分布关系及功能关系(
Shields et al ., 2014 ;
王慧杰等, 2016 ).这种关系是种群内不同径级个体间、种群与环境长期相互作用的结果, 通过它可以揭示种群的现状并预测种群发展趋势(
涂洪润等, 2019 ).空间正关联反映了种群内部正向(互惠)的生态关系, 空间负关联意味着种群内部负向(互斥)的生态关系, 空间无关联则反映了种群内部无明确的生态关系(
张金屯, 1998 ).该研究发现小尺度(<4 m)下大树个体与其他径级个体都呈负关联, 随着尺度的增大, 大树与幼树表现出正关联且关联性随尺度的增大而减弱, 而与小树、中树则表现为无关联或者微弱的负关联.小尺度下大树与其他径级个体都呈负关联, 表明在一定范围内其他径级个体都受到大树的排斥, 这可能是因为大树个体对土壤水分、光等资源的需求量更大, 竞争能力更强, 从而使得其周围其他径级个体的分布密度非常小(
张华等, 2016 ).也有研究表明青海云杉凋落物对种子的萌发和幼苗的生长有自毒作用(
王丽红等, 2017 ), 而大树周围凋落物一般也较为丰富, 导致大树周围种子和幼苗的生长受到抑制.实际调查中也发现在大树周围尤其是林冠下其他径级个体数量很少.大树个体和幼树个体在较大尺度上表现出正关联且随着尺度的增大, 关联性逐渐减弱, 这很可能是由种子扩散限制所引起的(
梁爽等, 2014 ), 大树产生的种子数量较多, 幼树的分布在一定程度上能反映种子的分布, 大树和幼树的关联性变化特点与青海云杉种子主要通过风力传播的特点具有一致性.此外, 在较大尺度下, 大树个体与小树、中树个体都表现为无关联或者微弱的负关联, 一方面表明随着尺度的增大, 大树对小树和中树的影响逐渐降低, 竞争趋于和缓, 另一方面随着径级的增大, 大树与其他径级个体关联性也发生变化, 说明不同径级青海云杉空间关联性随着发育阶段而变化, 这可能是由种群自身的生物生态学特性、种内种间关系以及环境条件共同作用的结果. ...
... 该研究还发现径级相近的个体空间关联性呈正关联或无关联, 随着个体径级间差距增大, 它们之间的空间关联性转变为无关联或负关联, 这说明同一物种不同径级的个体之间同时存在竞争、依赖以及无明确的生态关系, 这是由于物种具有一致的生境偏好所造成的(
杨洪晓等, 2006 ).径级相近个体的空间关联性呈正关联, 表明它们相伴出现, 对生境的选择具有一致性, 在空间分布上是相互依存的.这可能是由于它们生长发育阶段相近, 对环境资源的需求具有相似性和生态位存在重叠, 但小径级个体相对幼小, 对光照、水分和土壤养分等资源的竞争能力相对较弱, 需要借助高大植株的庇护, 才得以提高存活率, 而较大径级的植株个体可以改善样地的小生境, 使得邻近径级的个体更适宜生存, 彼此共享资源(
涂洪润等, 2019 ).同时, 由于物种专一性的资源分布不均, 种内竞争具有非对称性的特点, 也就是说当径级相近时, 不同径级间关系较协调(
Getzin et al ., 2008 ).而无关联则说明个体间互不干扰, 个体在前期发育过程中已经占据适宜的生态位, 与种内其他径级个体间的关联性不高, 能各自生长发育(
涂洪润等, 2019 ).随着个体间径级差距增加, 它们之间的关联性转变成无关联甚至负关联, 说明随着个体间径级差距的加大, 它们的空间正关联关系逐渐减弱, 当个体间差异较大时, 大径级个体在竞争中处于优势地位, 可能导致小径级个体的生长发育迟滞或者死亡, 使其周围小径级个体很少(
Kenkel, 1988 ), 这也间接证实了随着个体大小的生长变化, 青海云杉种群的空间分布存在一个从聚集分布到随机分布的自疏过程. ...
... ).而无关联则说明个体间互不干扰, 个体在前期发育过程中已经占据适宜的生态位, 与种内其他径级个体间的关联性不高, 能各自生长发育(
涂洪润等, 2019 ).随着个体间径级差距增加, 它们之间的关联性转变成无关联甚至负关联, 说明随着个体间径级差距的加大, 它们的空间正关联关系逐渐减弱, 当个体间差异较大时, 大径级个体在竞争中处于优势地位, 可能导致小径级个体的生长发育迟滞或者死亡, 使其周围小径级个体很少(
Kenkel, 1988 ), 这也间接证实了随着个体大小的生长变化, 青海云杉种群的空间分布存在一个从聚集分布到随机分布的自疏过程. ...
种群分布格局的多尺度分析
1
2005
... 空间分布格局的研究方法众多, 早期的研究方法主要有: 以频度/密度为基础的样方法、以距离为基础的无样地法、方差区组分析法、双向轨迹方差分析、
χ 2 检验和分形理论等(
张金屯, 1998 ;
王本洋和余世孝, 2005 ).传统的样方取样和分析方法常基于单一尺度(固定大小样方)的若干样方, 常采用
χ 2 分析法和方差/均值比率法等统计分析方法, 然而种群空间分布格局及其关联性对种群密度和空间尺度均有很强的依赖性, 在不同的尺度上往往表现出不同的类型, 故早期的取样和分析方法只能研究特定尺度下种群的空间分布格局特征, 难以全面地反映森林内物种的空间分布特点和种间关系(
He et al ., 1997 ;
李立等, 2010 ).鉴于上述研究方法的局限性,
Ripley (1977) 和
Diggle (1983) 分别提出和完善了点格局分析法, 以分析各种尺度上的空间分布格局.该方法以数字化的植物个体位置信息为基本数据, 将每个个体都视为二维空间中的一个点, 所有个体组成了植物种的空间分布图, 然后在空间分布图的基础上进行种群空间格局分析.其优点是能最大限度地利用点与点之间的距离, 不仅能够提供全面的空间格局信息, 而且具有较强的检验能力, 克服了传统研究方法中尺度单一等不足, 可以分析任意尺度下种群的空间格局特征及种内、种间关系(
Wiegand & Moloney, 2004 ;
Schleicher et al ., 2011 ;
沈志强等, 2016 ;
涂洪润等, 2019 ).因此, 空间点格局分析法在种群空间格局分布特征及潜在生态过程假说的研究方面得到了广泛应用. ...
种群分布格局的多尺度分析
1
2005
... 空间分布格局的研究方法众多, 早期的研究方法主要有: 以频度/密度为基础的样方法、以距离为基础的无样地法、方差区组分析法、双向轨迹方差分析、
χ 2 检验和分形理论等(
张金屯, 1998 ;
王本洋和余世孝, 2005 ).传统的样方取样和分析方法常基于单一尺度(固定大小样方)的若干样方, 常采用
χ 2 分析法和方差/均值比率法等统计分析方法, 然而种群空间分布格局及其关联性对种群密度和空间尺度均有很强的依赖性, 在不同的尺度上往往表现出不同的类型, 故早期的取样和分析方法只能研究特定尺度下种群的空间分布格局特征, 难以全面地反映森林内物种的空间分布特点和种间关系(
He et al ., 1997 ;
李立等, 2010 ).鉴于上述研究方法的局限性,
Ripley (1977) 和
Diggle (1983) 分别提出和完善了点格局分析法, 以分析各种尺度上的空间分布格局.该方法以数字化的植物个体位置信息为基本数据, 将每个个体都视为二维空间中的一个点, 所有个体组成了植物种的空间分布图, 然后在空间分布图的基础上进行种群空间格局分析.其优点是能最大限度地利用点与点之间的距离, 不仅能够提供全面的空间格局信息, 而且具有较强的检验能力, 克服了传统研究方法中尺度单一等不足, 可以分析任意尺度下种群的空间格局特征及种内、种间关系(
Wiegand & Moloney, 2004 ;
Schleicher et al ., 2011 ;
沈志强等, 2016 ;
涂洪润等, 2019 ).因此, 空间点格局分析法在种群空间格局分布特征及潜在生态过程假说的研究方面得到了广泛应用. ...
天山雪岭云杉森林群落的密度制约效应
1
2016
... 种群内不同径级个体的空间关联性描述了某一特定的时间内植物种群组成个体之间的空间分布关系及功能关系(
Shields et al ., 2014 ;
王慧杰等, 2016 ).这种关系是种群内不同径级个体间、种群与环境长期相互作用的结果, 通过它可以揭示种群的现状并预测种群发展趋势(
涂洪润等, 2019 ).空间正关联反映了种群内部正向(互惠)的生态关系, 空间负关联意味着种群内部负向(互斥)的生态关系, 空间无关联则反映了种群内部无明确的生态关系(
张金屯, 1998 ).该研究发现小尺度(<4 m)下大树个体与其他径级个体都呈负关联, 随着尺度的增大, 大树与幼树表现出正关联且关联性随尺度的增大而减弱, 而与小树、中树则表现为无关联或者微弱的负关联.小尺度下大树与其他径级个体都呈负关联, 表明在一定范围内其他径级个体都受到大树的排斥, 这可能是因为大树个体对土壤水分、光等资源的需求量更大, 竞争能力更强, 从而使得其周围其他径级个体的分布密度非常小(
张华等, 2016 ).也有研究表明青海云杉凋落物对种子的萌发和幼苗的生长有自毒作用(
王丽红等, 2017 ), 而大树周围凋落物一般也较为丰富, 导致大树周围种子和幼苗的生长受到抑制.实际调查中也发现在大树周围尤其是林冠下其他径级个体数量很少.大树个体和幼树个体在较大尺度上表现出正关联且随着尺度的增大, 关联性逐渐减弱, 这很可能是由种子扩散限制所引起的(
梁爽等, 2014 ), 大树产生的种子数量较多, 幼树的分布在一定程度上能反映种子的分布, 大树和幼树的关联性变化特点与青海云杉种子主要通过风力传播的特点具有一致性.此外, 在较大尺度下, 大树个体与小树、中树个体都表现为无关联或者微弱的负关联, 一方面表明随着尺度的增大, 大树对小树和中树的影响逐渐降低, 竞争趋于和缓, 另一方面随着径级的增大, 大树与其他径级个体关联性也发生变化, 说明不同径级青海云杉空间关联性随着发育阶段而变化, 这可能是由种群自身的生物生态学特性、种内种间关系以及环境条件共同作用的结果. ...
天山雪岭云杉森林群落的密度制约效应
1
2016
... 种群内不同径级个体的空间关联性描述了某一特定的时间内植物种群组成个体之间的空间分布关系及功能关系(
Shields et al ., 2014 ;
王慧杰等, 2016 ).这种关系是种群内不同径级个体间、种群与环境长期相互作用的结果, 通过它可以揭示种群的现状并预测种群发展趋势(
涂洪润等, 2019 ).空间正关联反映了种群内部正向(互惠)的生态关系, 空间负关联意味着种群内部负向(互斥)的生态关系, 空间无关联则反映了种群内部无明确的生态关系(
张金屯, 1998 ).该研究发现小尺度(<4 m)下大树个体与其他径级个体都呈负关联, 随着尺度的增大, 大树与幼树表现出正关联且关联性随尺度的增大而减弱, 而与小树、中树则表现为无关联或者微弱的负关联.小尺度下大树与其他径级个体都呈负关联, 表明在一定范围内其他径级个体都受到大树的排斥, 这可能是因为大树个体对土壤水分、光等资源的需求量更大, 竞争能力更强, 从而使得其周围其他径级个体的分布密度非常小(
张华等, 2016 ).也有研究表明青海云杉凋落物对种子的萌发和幼苗的生长有自毒作用(
王丽红等, 2017 ), 而大树周围凋落物一般也较为丰富, 导致大树周围种子和幼苗的生长受到抑制.实际调查中也发现在大树周围尤其是林冠下其他径级个体数量很少.大树个体和幼树个体在较大尺度上表现出正关联且随着尺度的增大, 关联性逐渐减弱, 这很可能是由种子扩散限制所引起的(
梁爽等, 2014 ), 大树产生的种子数量较多, 幼树的分布在一定程度上能反映种子的分布, 大树和幼树的关联性变化特点与青海云杉种子主要通过风力传播的特点具有一致性.此外, 在较大尺度下, 大树个体与小树、中树个体都表现为无关联或者微弱的负关联, 一方面表明随着尺度的增大, 大树对小树和中树的影响逐渐降低, 竞争趋于和缓, 另一方面随着径级的增大, 大树与其他径级个体关联性也发生变化, 说明不同径级青海云杉空间关联性随着发育阶段而变化, 这可能是由种群自身的生物生态学特性、种内种间关系以及环境条件共同作用的结果. ...
青海云杉凋落物对种子萌发和幼苗生长的自毒作用
1
2017
... 种群内不同径级个体的空间关联性描述了某一特定的时间内植物种群组成个体之间的空间分布关系及功能关系(
Shields et al ., 2014 ;
王慧杰等, 2016 ).这种关系是种群内不同径级个体间、种群与环境长期相互作用的结果, 通过它可以揭示种群的现状并预测种群发展趋势(
涂洪润等, 2019 ).空间正关联反映了种群内部正向(互惠)的生态关系, 空间负关联意味着种群内部负向(互斥)的生态关系, 空间无关联则反映了种群内部无明确的生态关系(
张金屯, 1998 ).该研究发现小尺度(<4 m)下大树个体与其他径级个体都呈负关联, 随着尺度的增大, 大树与幼树表现出正关联且关联性随尺度的增大而减弱, 而与小树、中树则表现为无关联或者微弱的负关联.小尺度下大树与其他径级个体都呈负关联, 表明在一定范围内其他径级个体都受到大树的排斥, 这可能是因为大树个体对土壤水分、光等资源的需求量更大, 竞争能力更强, 从而使得其周围其他径级个体的分布密度非常小(
张华等, 2016 ).也有研究表明青海云杉凋落物对种子的萌发和幼苗的生长有自毒作用(
王丽红等, 2017 ), 而大树周围凋落物一般也较为丰富, 导致大树周围种子和幼苗的生长受到抑制.实际调查中也发现在大树周围尤其是林冠下其他径级个体数量很少.大树个体和幼树个体在较大尺度上表现出正关联且随着尺度的增大, 关联性逐渐减弱, 这很可能是由种子扩散限制所引起的(
梁爽等, 2014 ), 大树产生的种子数量较多, 幼树的分布在一定程度上能反映种子的分布, 大树和幼树的关联性变化特点与青海云杉种子主要通过风力传播的特点具有一致性.此外, 在较大尺度下, 大树个体与小树、中树个体都表现为无关联或者微弱的负关联, 一方面表明随着尺度的增大, 大树对小树和中树的影响逐渐降低, 竞争趋于和缓, 另一方面随着径级的增大, 大树与其他径级个体关联性也发生变化, 说明不同径级青海云杉空间关联性随着发育阶段而变化, 这可能是由种群自身的生物生态学特性、种内种间关系以及环境条件共同作用的结果. ...
青海云杉凋落物对种子萌发和幼苗生长的自毒作用
1
2017
... 种群内不同径级个体的空间关联性描述了某一特定的时间内植物种群组成个体之间的空间分布关系及功能关系(
Shields et al ., 2014 ;
王慧杰等, 2016 ).这种关系是种群内不同径级个体间、种群与环境长期相互作用的结果, 通过它可以揭示种群的现状并预测种群发展趋势(
涂洪润等, 2019 ).空间正关联反映了种群内部正向(互惠)的生态关系, 空间负关联意味着种群内部负向(互斥)的生态关系, 空间无关联则反映了种群内部无明确的生态关系(
张金屯, 1998 ).该研究发现小尺度(<4 m)下大树个体与其他径级个体都呈负关联, 随着尺度的增大, 大树与幼树表现出正关联且关联性随尺度的增大而减弱, 而与小树、中树则表现为无关联或者微弱的负关联.小尺度下大树与其他径级个体都呈负关联, 表明在一定范围内其他径级个体都受到大树的排斥, 这可能是因为大树个体对土壤水分、光等资源的需求量更大, 竞争能力更强, 从而使得其周围其他径级个体的分布密度非常小(
张华等, 2016 ).也有研究表明青海云杉凋落物对种子的萌发和幼苗的生长有自毒作用(
王丽红等, 2017 ), 而大树周围凋落物一般也较为丰富, 导致大树周围种子和幼苗的生长受到抑制.实际调查中也发现在大树周围尤其是林冠下其他径级个体数量很少.大树个体和幼树个体在较大尺度上表现出正关联且随着尺度的增大, 关联性逐渐减弱, 这很可能是由种子扩散限制所引起的(
梁爽等, 2014 ), 大树产生的种子数量较多, 幼树的分布在一定程度上能反映种子的分布, 大树和幼树的关联性变化特点与青海云杉种子主要通过风力传播的特点具有一致性.此外, 在较大尺度下, 大树个体与小树、中树个体都表现为无关联或者微弱的负关联, 一方面表明随着尺度的增大, 大树对小树和中树的影响逐渐降低, 竞争趋于和缓, 另一方面随着径级的增大, 大树与其他径级个体关联性也发生变化, 说明不同径级青海云杉空间关联性随着发育阶段而变化, 这可能是由种群自身的生物生态学特性、种内种间关系以及环境条件共同作用的结果. ...
Spatial distributions of species in an old-growth temperate forest, northeastern China
1
2010
... 种群的空间格局是指特定时间内, 种群所有个体在其水平生活空间内的分布状况及其属性在空间上的配置特征, 往往是由种群自身生物学特性、种内种间竞争、环境条件等因素综合作用的结果(
林玥, 2008 ;
Liu et al ., 2020 ).种群的空间分布格局和空间关联性是种群空间格局研究的2个主要内容, 也是种群生态关系在空间格局上的两种表现形式(
张金屯和孟东平, 2004 ).大量研究表明种群的空间分布格局与空间关联性关系密切, 聚集分布和空间正关联表明种群内部正向(互惠)的生态关系, 规则分布和空间负关联则表明种群内部负向(互斥)的生态关系, 随机分布和空间无关联则代表着种群内部不存在明确的生态关系(
杨洪晓, 2006 ).因此, 研究植物种群的空间分布格局及其关联性不仅能够了解种群的空间水平结构与生物、生态学习性(如生活史策略、是否喜光或耐阴等)、种群间相互作用、种群与环境因子的相互作用规律(如植物与生长环境间的适合度、生境异质性等)以及种群在群落中的地位和生态学功能, 更重要的是可以揭示这种分布格局及其关联性的潜在成因, 进而推演种群动态变化的生态学过程(如种内和种间竞争、种子扩散和干扰等), 深化对群落结构的认识, 揭示植物群落的生物多样性形成和维持机制(
McDonald et al ., 2003 ;
Hou et al , 2004 ;
Wiegand & Moloney, 2004 ;
Nathan, 2006 ;
Wang et al ., 2010 ;
Guo et al ., 2013 ). ...
Rings, circles, and null-models for point pattern analysis in ecology
2
2004
... 种群的空间格局是指特定时间内, 种群所有个体在其水平生活空间内的分布状况及其属性在空间上的配置特征, 往往是由种群自身生物学特性、种内种间竞争、环境条件等因素综合作用的结果(
林玥, 2008 ;
Liu et al ., 2020 ).种群的空间分布格局和空间关联性是种群空间格局研究的2个主要内容, 也是种群生态关系在空间格局上的两种表现形式(
张金屯和孟东平, 2004 ).大量研究表明种群的空间分布格局与空间关联性关系密切, 聚集分布和空间正关联表明种群内部正向(互惠)的生态关系, 规则分布和空间负关联则表明种群内部负向(互斥)的生态关系, 随机分布和空间无关联则代表着种群内部不存在明确的生态关系(
杨洪晓, 2006 ).因此, 研究植物种群的空间分布格局及其关联性不仅能够了解种群的空间水平结构与生物、生态学习性(如生活史策略、是否喜光或耐阴等)、种群间相互作用、种群与环境因子的相互作用规律(如植物与生长环境间的适合度、生境异质性等)以及种群在群落中的地位和生态学功能, 更重要的是可以揭示这种分布格局及其关联性的潜在成因, 进而推演种群动态变化的生态学过程(如种内和种间竞争、种子扩散和干扰等), 深化对群落结构的认识, 揭示植物群落的生物多样性形成和维持机制(
McDonald et al ., 2003 ;
Hou et al , 2004 ;
Wiegand & Moloney, 2004 ;
Nathan, 2006 ;
Wang et al ., 2010 ;
Guo et al ., 2013 ). ...
... 空间分布格局的研究方法众多, 早期的研究方法主要有: 以频度/密度为基础的样方法、以距离为基础的无样地法、方差区组分析法、双向轨迹方差分析、
χ 2 检验和分形理论等(
张金屯, 1998 ;
王本洋和余世孝, 2005 ).传统的样方取样和分析方法常基于单一尺度(固定大小样方)的若干样方, 常采用
χ 2 分析法和方差/均值比率法等统计分析方法, 然而种群空间分布格局及其关联性对种群密度和空间尺度均有很强的依赖性, 在不同的尺度上往往表现出不同的类型, 故早期的取样和分析方法只能研究特定尺度下种群的空间分布格局特征, 难以全面地反映森林内物种的空间分布特点和种间关系(
He et al ., 1997 ;
李立等, 2010 ).鉴于上述研究方法的局限性,
Ripley (1977) 和
Diggle (1983) 分别提出和完善了点格局分析法, 以分析各种尺度上的空间分布格局.该方法以数字化的植物个体位置信息为基本数据, 将每个个体都视为二维空间中的一个点, 所有个体组成了植物种的空间分布图, 然后在空间分布图的基础上进行种群空间格局分析.其优点是能最大限度地利用点与点之间的距离, 不仅能够提供全面的空间格局信息, 而且具有较强的检验能力, 克服了传统研究方法中尺度单一等不足, 可以分析任意尺度下种群的空间格局特征及种内、种间关系(
Wiegand & Moloney, 2004 ;
Schleicher et al ., 2011 ;
沈志强等, 2016 ;
涂洪润等, 2019 ).因此, 空间点格局分析法在种群空间格局分布特征及潜在生态过程假说的研究方面得到了广泛应用. ...
大别山五针松种群结构及动态研究
1
2016
... 植物种群的径级结构是指不同径级组的个体在种群内所占比例和配置情况(
田润炜等, 2016 ), 是种群的生殖特性与外界环境相互作用的结果, 在一定程度上体现了种群的径级结构特征和生长发育状况, 进而可以预测种群和群落的动态发展趋势(
涂洪润等, 2019 ).本研究发现, 样地内青海云杉种群的幼苗和小树个体储备较为丰富, 其种群径级结构呈倒“J”型分布, 这说明该种群属增长型种群, 种群天然更新能力良好, 只要未来不遭受强烈干扰, 种群数量会保持逐渐增长.这与
李效雄等(2012) 和
张华等(2016) 分别对祁连山大野口流域和天老池小流域的青海云杉种群结构的研究结果一致.这可能是由青海云杉种群的生物学特性与环境因素共同作用的结果.已有研究表明, 森林更新过程中最重要和最敏感的阶段是幼苗阶段(
李晓笑等, 2011 ;
赵阳等, 2018 ), 林内有限的资源和空间往往成为种群更新的重要限制因素(
项小燕等, 2016 ).青海云杉种群的幼苗和小树个体数量多主要是由其生殖特性决定, 其种群可以产生大量高质量种子, 自然萌发形成的幼苗大都集中分布在林窗下, 形成大量小径级个体, 占据有利资源, 这是其种群在群落中持续处于优势地位的主要原因.随着小径级个体的生长, 它们对光照、水分、土壤养分等环境资源和空间需求随之增加, 由于环境资源和空间有限, 种内和种间竞争加剧, 出现强烈的自疏和他疏现象, 竞争能力弱的幼苗和小树在竞争中会大量死亡, 只有少量小径级个体能够成功转化为大径级个体, 造成样地内仅存在少量的中树和大树, 这成为该青海云杉种群更新和发展的“瓶颈” (
张婕等, 2014 ;
白卫国等, 2017 ;
于世川等, 2017 ).因此, 要实现种群数量的稳定增长, 必须加强对种群结构的保护, 尤其要重视对幼苗和小树及其生存环境的保护和改善. ...
大别山五针松种群结构及动态研究
1
2016
... 植物种群的径级结构是指不同径级组的个体在种群内所占比例和配置情况(
田润炜等, 2016 ), 是种群的生殖特性与外界环境相互作用的结果, 在一定程度上体现了种群的径级结构特征和生长发育状况, 进而可以预测种群和群落的动态发展趋势(
涂洪润等, 2019 ).本研究发现, 样地内青海云杉种群的幼苗和小树个体储备较为丰富, 其种群径级结构呈倒“J”型分布, 这说明该种群属增长型种群, 种群天然更新能力良好, 只要未来不遭受强烈干扰, 种群数量会保持逐渐增长.这与
李效雄等(2012) 和
张华等(2016) 分别对祁连山大野口流域和天老池小流域的青海云杉种群结构的研究结果一致.这可能是由青海云杉种群的生物学特性与环境因素共同作用的结果.已有研究表明, 森林更新过程中最重要和最敏感的阶段是幼苗阶段(
李晓笑等, 2011 ;
赵阳等, 2018 ), 林内有限的资源和空间往往成为种群更新的重要限制因素(
项小燕等, 2016 ).青海云杉种群的幼苗和小树个体数量多主要是由其生殖特性决定, 其种群可以产生大量高质量种子, 自然萌发形成的幼苗大都集中分布在林窗下, 形成大量小径级个体, 占据有利资源, 这是其种群在群落中持续处于优势地位的主要原因.随着小径级个体的生长, 它们对光照、水分、土壤养分等环境资源和空间需求随之增加, 由于环境资源和空间有限, 种内和种间竞争加剧, 出现强烈的自疏和他疏现象, 竞争能力弱的幼苗和小树在竞争中会大量死亡, 只有少量小径级个体能够成功转化为大径级个体, 造成样地内仅存在少量的中树和大树, 这成为该青海云杉种群更新和发展的“瓶颈” (
张婕等, 2014 ;
白卫国等, 2017 ;
于世川等, 2017 ).因此, 要实现种群数量的稳定增长, 必须加强对种群结构的保护, 尤其要重视对幼苗和小树及其生存环境的保护和改善. ...
长白山云冷杉林幼苗幼树空间分布格局及其更新特征
1
2014
... 森林的空间格局与空间尺度密切相关, 物种的空间分布格局随着尺度的变化而变化(
Condit et al ., 2000 ;
杨华等, 2014 ).一般来说, 小尺度上的物种分布格局主要受种群自身生物学特性(如种内种间竞争、种子扩散限制和幼苗补充过程等)影响, 而大尺度上物种的空间分布则主要受生境异质性(如光照、土壤养分、水分和温度等)影响(
Shen et al ., 2013 ).本研究表明, 青海云杉种群不同径级的青海云杉个体在较小尺度内均表现为聚集分布, 随着空间尺度的增加, 呈现由聚集分布到随机分布的变化趋势.这一结果与国内外大多数有关种群空间格局的研究结果一致(
刘建泉, 2004 ;
Guo et al ., 2013 ;
张觅等, 2014 ;
张华等, 2016 ).这是因为同一种群的不同个体对生境条件的需求基本上是一致的(
张金屯和孟东平, 2004 ), 且聚集分布有利于种群个体相互庇护, 从而抵御外界不利环境, 以提高存活率(
Diggle, 1981 ).此外, 生境的异质性容易引起种群聚集生长, 同时由于植株对光照、水分、养分等资源的竞争较大(
李海东等, 2011 ), 导致聚集强度随着空间尺度的增大而呈现下降趋势, 表现为在较小尺度上植株倾向于聚集分布逐渐过渡为在较大尺度上的随机分布. ...
长白山云冷杉林幼苗幼树空间分布格局及其更新特征
1
2014
... 森林的空间格局与空间尺度密切相关, 物种的空间分布格局随着尺度的变化而变化(
Condit et al ., 2000 ;
杨华等, 2014 ).一般来说, 小尺度上的物种分布格局主要受种群自身生物学特性(如种内种间竞争、种子扩散限制和幼苗补充过程等)影响, 而大尺度上物种的空间分布则主要受生境异质性(如光照、土壤养分、水分和温度等)影响(
Shen et al ., 2013 ).本研究表明, 青海云杉种群不同径级的青海云杉个体在较小尺度内均表现为聚集分布, 随着空间尺度的增加, 呈现由聚集分布到随机分布的变化趋势.这一结果与国内外大多数有关种群空间格局的研究结果一致(
刘建泉, 2004 ;
Guo et al ., 2013 ;
张觅等, 2014 ;
张华等, 2016 ).这是因为同一种群的不同个体对生境条件的需求基本上是一致的(
张金屯和孟东平, 2004 ), 且聚集分布有利于种群个体相互庇护, 从而抵御外界不利环境, 以提高存活率(
Diggle, 1981 ).此外, 生境的异质性容易引起种群聚集生长, 同时由于植株对光照、水分、养分等资源的竞争较大(
李海东等, 2011 ), 导致聚集强度随着空间尺度的增大而呈现下降趋势, 表现为在较小尺度上植株倾向于聚集分布逐渐过渡为在较大尺度上的随机分布. ...
毛乌素沙地油蒿种群点格局分析
2
2006
... 种群的空间格局是指特定时间内, 种群所有个体在其水平生活空间内的分布状况及其属性在空间上的配置特征, 往往是由种群自身生物学特性、种内种间竞争、环境条件等因素综合作用的结果(
林玥, 2008 ;
Liu et al ., 2020 ).种群的空间分布格局和空间关联性是种群空间格局研究的2个主要内容, 也是种群生态关系在空间格局上的两种表现形式(
张金屯和孟东平, 2004 ).大量研究表明种群的空间分布格局与空间关联性关系密切, 聚集分布和空间正关联表明种群内部正向(互惠)的生态关系, 规则分布和空间负关联则表明种群内部负向(互斥)的生态关系, 随机分布和空间无关联则代表着种群内部不存在明确的生态关系(
杨洪晓, 2006 ).因此, 研究植物种群的空间分布格局及其关联性不仅能够了解种群的空间水平结构与生物、生态学习性(如生活史策略、是否喜光或耐阴等)、种群间相互作用、种群与环境因子的相互作用规律(如植物与生长环境间的适合度、生境异质性等)以及种群在群落中的地位和生态学功能, 更重要的是可以揭示这种分布格局及其关联性的潜在成因, 进而推演种群动态变化的生态学过程(如种内和种间竞争、种子扩散和干扰等), 深化对群落结构的认识, 揭示植物群落的生物多样性形成和维持机制(
McDonald et al ., 2003 ;
Hou et al , 2004 ;
Wiegand & Moloney, 2004 ;
Nathan, 2006 ;
Wang et al ., 2010 ;
Guo et al ., 2013 ). ...
... 该研究还发现径级相近的个体空间关联性呈正关联或无关联, 随着个体径级间差距增大, 它们之间的空间关联性转变为无关联或负关联, 这说明同一物种不同径级的个体之间同时存在竞争、依赖以及无明确的生态关系, 这是由于物种具有一致的生境偏好所造成的(
杨洪晓等, 2006 ).径级相近个体的空间关联性呈正关联, 表明它们相伴出现, 对生境的选择具有一致性, 在空间分布上是相互依存的.这可能是由于它们生长发育阶段相近, 对环境资源的需求具有相似性和生态位存在重叠, 但小径级个体相对幼小, 对光照、水分和土壤养分等资源的竞争能力相对较弱, 需要借助高大植株的庇护, 才得以提高存活率, 而较大径级的植株个体可以改善样地的小生境, 使得邻近径级的个体更适宜生存, 彼此共享资源(
涂洪润等, 2019 ).同时, 由于物种专一性的资源分布不均, 种内竞争具有非对称性的特点, 也就是说当径级相近时, 不同径级间关系较协调(
Getzin et al ., 2008 ).而无关联则说明个体间互不干扰, 个体在前期发育过程中已经占据适宜的生态位, 与种内其他径级个体间的关联性不高, 能各自生长发育(
涂洪润等, 2019 ).随着个体间径级差距增加, 它们之间的关联性转变成无关联甚至负关联, 说明随着个体间径级差距的加大, 它们的空间正关联关系逐渐减弱, 当个体间差异较大时, 大径级个体在竞争中处于优势地位, 可能导致小径级个体的生长发育迟滞或者死亡, 使其周围小径级个体很少(
Kenkel, 1988 ), 这也间接证实了随着个体大小的生长变化, 青海云杉种群的空间分布存在一个从聚集分布到随机分布的自疏过程. ...
毛乌素沙地油蒿种群点格局分析
2
2006
... 种群的空间格局是指特定时间内, 种群所有个体在其水平生活空间内的分布状况及其属性在空间上的配置特征, 往往是由种群自身生物学特性、种内种间竞争、环境条件等因素综合作用的结果(
林玥, 2008 ;
Liu et al ., 2020 ).种群的空间分布格局和空间关联性是种群空间格局研究的2个主要内容, 也是种群生态关系在空间格局上的两种表现形式(
张金屯和孟东平, 2004 ).大量研究表明种群的空间分布格局与空间关联性关系密切, 聚集分布和空间正关联表明种群内部正向(互惠)的生态关系, 规则分布和空间负关联则表明种群内部负向(互斥)的生态关系, 随机分布和空间无关联则代表着种群内部不存在明确的生态关系(
杨洪晓, 2006 ).因此, 研究植物种群的空间分布格局及其关联性不仅能够了解种群的空间水平结构与生物、生态学习性(如生活史策略、是否喜光或耐阴等)、种群间相互作用、种群与环境因子的相互作用规律(如植物与生长环境间的适合度、生境异质性等)以及种群在群落中的地位和生态学功能, 更重要的是可以揭示这种分布格局及其关联性的潜在成因, 进而推演种群动态变化的生态学过程(如种内和种间竞争、种子扩散和干扰等), 深化对群落结构的认识, 揭示植物群落的生物多样性形成和维持机制(
McDonald et al ., 2003 ;
Hou et al , 2004 ;
Wiegand & Moloney, 2004 ;
Nathan, 2006 ;
Wang et al ., 2010 ;
Guo et al ., 2013 ). ...
... 该研究还发现径级相近的个体空间关联性呈正关联或无关联, 随着个体径级间差距增大, 它们之间的空间关联性转变为无关联或负关联, 这说明同一物种不同径级的个体之间同时存在竞争、依赖以及无明确的生态关系, 这是由于物种具有一致的生境偏好所造成的(
杨洪晓等, 2006 ).径级相近个体的空间关联性呈正关联, 表明它们相伴出现, 对生境的选择具有一致性, 在空间分布上是相互依存的.这可能是由于它们生长发育阶段相近, 对环境资源的需求具有相似性和生态位存在重叠, 但小径级个体相对幼小, 对光照、水分和土壤养分等资源的竞争能力相对较弱, 需要借助高大植株的庇护, 才得以提高存活率, 而较大径级的植株个体可以改善样地的小生境, 使得邻近径级的个体更适宜生存, 彼此共享资源(
涂洪润等, 2019 ).同时, 由于物种专一性的资源分布不均, 种内竞争具有非对称性的特点, 也就是说当径级相近时, 不同径级间关系较协调(
Getzin et al ., 2008 ).而无关联则说明个体间互不干扰, 个体在前期发育过程中已经占据适宜的生态位, 与种内其他径级个体间的关联性不高, 能各自生长发育(
涂洪润等, 2019 ).随着个体间径级差距增加, 它们之间的关联性转变成无关联甚至负关联, 说明随着个体间径级差距的加大, 它们的空间正关联关系逐渐减弱, 当个体间差异较大时, 大径级个体在竞争中处于优势地位, 可能导致小径级个体的生长发育迟滞或者死亡, 使其周围小径级个体很少(
Kenkel, 1988 ), 这也间接证实了随着个体大小的生长变化, 青海云杉种群的空间分布存在一个从聚集分布到随机分布的自疏过程. ...
1
2018
... 虽然径级和龄级之间存在差异, 但在同样的环境条件下, 树木的径级和龄级对生境条件的响应规律具有一致性.青海云杉寿命很长, 在100-140年以后生长才缓慢下来, 少数林木在300-400年仍能持续缓慢生长.已有研究表明, 其胸径和树龄具有明显的线性关系,
y = 0.2133
x - 1.1136,
R 2 = 0.984 9 (
y 为胸径,
x 树龄).如: 当胸径为5.0、12.5以及22.5 cm时分别大致对应树龄30、64和111年(
杨文娟, 2018 ).因此可以用胸径来反映种群的生活史阶段(
Frost & Rydin, 2000 ;
李立等, 2010 ).从而该研究采用径级结构(胸径大小)代替年龄结构, 分析不同径级青海云杉种群分布格局及其关联性的变化.该研究参照李妍(2013)对青海云杉种群径级结构的划分标准, 同时结合样地内青海云杉胸径的实际情况以及青海云杉的生活史特点, 依据青海云杉胸径大小将其划分为: 径级I (幼树, 1 cm ≤
DBH < 5.0 cm)、径级II (小树, 5.0 cm ≤
DBH < 12.5 cm)、径级III (中树, 12.5 cm ≤
DBH < 22.5 cm)、径级IV (大树,
DBH ≥ 22.5 cm) 4个不同径级. ...
1
2018
... 虽然径级和龄级之间存在差异, 但在同样的环境条件下, 树木的径级和龄级对生境条件的响应规律具有一致性.青海云杉寿命很长, 在100-140年以后生长才缓慢下来, 少数林木在300-400年仍能持续缓慢生长.已有研究表明, 其胸径和树龄具有明显的线性关系,
y = 0.2133
x - 1.1136,
R 2 = 0.984 9 (
y 为胸径,
x 树龄).如: 当胸径为5.0、12.5以及22.5 cm时分别大致对应树龄30、64和111年(
杨文娟, 2018 ).因此可以用胸径来反映种群的生活史阶段(
Frost & Rydin, 2000 ;
李立等, 2010 ).从而该研究采用径级结构(胸径大小)代替年龄结构, 分析不同径级青海云杉种群分布格局及其关联性的变化.该研究参照李妍(2013)对青海云杉种群径级结构的划分标准, 同时结合样地内青海云杉胸径的实际情况以及青海云杉的生活史特点, 依据青海云杉胸径大小将其划分为: 径级I (幼树, 1 cm ≤
DBH < 5.0 cm)、径级II (小树, 5.0 cm ≤
DBH < 12.5 cm)、径级III (中树, 12.5 cm ≤
DBH < 22.5 cm)、径级IV (大树,
DBH ≥ 22.5 cm) 4个不同径级. ...
抚育间伐对黄龙山辽东栎林木形质的影响
1
2017
... 植物种群的径级结构是指不同径级组的个体在种群内所占比例和配置情况(
田润炜等, 2016 ), 是种群的生殖特性与外界环境相互作用的结果, 在一定程度上体现了种群的径级结构特征和生长发育状况, 进而可以预测种群和群落的动态发展趋势(
涂洪润等, 2019 ).本研究发现, 样地内青海云杉种群的幼苗和小树个体储备较为丰富, 其种群径级结构呈倒“J”型分布, 这说明该种群属增长型种群, 种群天然更新能力良好, 只要未来不遭受强烈干扰, 种群数量会保持逐渐增长.这与
李效雄等(2012) 和
张华等(2016) 分别对祁连山大野口流域和天老池小流域的青海云杉种群结构的研究结果一致.这可能是由青海云杉种群的生物学特性与环境因素共同作用的结果.已有研究表明, 森林更新过程中最重要和最敏感的阶段是幼苗阶段(
李晓笑等, 2011 ;
赵阳等, 2018 ), 林内有限的资源和空间往往成为种群更新的重要限制因素(
项小燕等, 2016 ).青海云杉种群的幼苗和小树个体数量多主要是由其生殖特性决定, 其种群可以产生大量高质量种子, 自然萌发形成的幼苗大都集中分布在林窗下, 形成大量小径级个体, 占据有利资源, 这是其种群在群落中持续处于优势地位的主要原因.随着小径级个体的生长, 它们对光照、水分、土壤养分等环境资源和空间需求随之增加, 由于环境资源和空间有限, 种内和种间竞争加剧, 出现强烈的自疏和他疏现象, 竞争能力弱的幼苗和小树在竞争中会大量死亡, 只有少量小径级个体能够成功转化为大径级个体, 造成样地内仅存在少量的中树和大树, 这成为该青海云杉种群更新和发展的“瓶颈” (
张婕等, 2014 ;
白卫国等, 2017 ;
于世川等, 2017 ).因此, 要实现种群数量的稳定增长, 必须加强对种群结构的保护, 尤其要重视对幼苗和小树及其生存环境的保护和改善. ...
抚育间伐对黄龙山辽东栎林木形质的影响
1
2017
... 植物种群的径级结构是指不同径级组的个体在种群内所占比例和配置情况(
田润炜等, 2016 ), 是种群的生殖特性与外界环境相互作用的结果, 在一定程度上体现了种群的径级结构特征和生长发育状况, 进而可以预测种群和群落的动态发展趋势(
涂洪润等, 2019 ).本研究发现, 样地内青海云杉种群的幼苗和小树个体储备较为丰富, 其种群径级结构呈倒“J”型分布, 这说明该种群属增长型种群, 种群天然更新能力良好, 只要未来不遭受强烈干扰, 种群数量会保持逐渐增长.这与
李效雄等(2012) 和
张华等(2016) 分别对祁连山大野口流域和天老池小流域的青海云杉种群结构的研究结果一致.这可能是由青海云杉种群的生物学特性与环境因素共同作用的结果.已有研究表明, 森林更新过程中最重要和最敏感的阶段是幼苗阶段(
李晓笑等, 2011 ;
赵阳等, 2018 ), 林内有限的资源和空间往往成为种群更新的重要限制因素(
项小燕等, 2016 ).青海云杉种群的幼苗和小树个体数量多主要是由其生殖特性决定, 其种群可以产生大量高质量种子, 自然萌发形成的幼苗大都集中分布在林窗下, 形成大量小径级个体, 占据有利资源, 这是其种群在群落中持续处于优势地位的主要原因.随着小径级个体的生长, 它们对光照、水分、土壤养分等环境资源和空间需求随之增加, 由于环境资源和空间有限, 种内和种间竞争加剧, 出现强烈的自疏和他疏现象, 竞争能力弱的幼苗和小树在竞争中会大量死亡, 只有少量小径级个体能够成功转化为大径级个体, 造成样地内仅存在少量的中树和大树, 这成为该青海云杉种群更新和发展的“瓶颈” (
张婕等, 2014 ;
白卫国等, 2017 ;
于世川等, 2017 ).因此, 要实现种群数量的稳定增长, 必须加强对种群结构的保护, 尤其要重视对幼苗和小树及其生存环境的保护和改善. ...
青海云杉不同龄级立木的点格局分析
6
2016
... 祁连山是中国西部的主要山脉之一, 位于青藏高原、蒙古高原、黄土高原三大高原交汇带, 是石羊河、黑河、疏勒河等6大内陆河和黄河流域的重要水源产流地, 维系着河西乃至西部地区生存与发展的命脉, 在国家生态建设中具有十分重要的战略地位(
Gao et al ., 2013 ;
Zhu et al ., 2017 ).祁连山山地森林的主要建群种——青海云杉(
Picea crassifolia ), 为松科云杉属(
Picea )乔木, 是中国青藏高原东北边缘特有树种, 多以纯林形式存在, 呈带状或斑块状分布于海拔2 300-3 300 m的阴坡和半阴坡, 分布面积占祁连山水源涵养林面积的24.74%, 占乔木林面积的75.72%, 对于维系祁连山的生物多样性、水源涵养功能乃至区域生态平衡都具有至关重要的作用(
田风霞等, 2012 ;
何芳兰等, 2016 ;
张华等, 2016 ).部分****(
赵丽琼等, 2010 ;
张华等, 2016 )虽然已对青海云杉种群空间格局及其关联性进行了研究, 但是已有研究的空间尺度相对较小, 仅能揭示小尺度下的种群空间分布格局, 难以客观地反映青海云杉种群空间分布的特点和种内关系.鉴于此, 本研究基于祁连山大野口流域10.2 hm
2 青海云杉动态监测样地数据, 采用空间点格局分析方法中的成对相关函数对不同径级青海云杉种群的空间分布格局及其关联性进行分析, 旨在阐明: (1)祁连山大野口流域青海云杉种群不同径级个体的空间分布格局及其关联性如何随空间尺度进行变化; (2)不同径级个体的空间分布格局及其关联性形成的潜在生态学过程与影响机制.上述科学问题的合理解答, 能从空间格局的角度客观地揭示青海云杉种群发展规律, 可以为制定该种群的配置、种植与保护管理措施提供指导, 并对祁连山地区的植被恢复与重建、生物多样性保护等具有重要的意义和价值. ...
... ;
张华等, 2016 )虽然已对青海云杉种群空间格局及其关联性进行了研究, 但是已有研究的空间尺度相对较小, 仅能揭示小尺度下的种群空间分布格局, 难以客观地反映青海云杉种群空间分布的特点和种内关系.鉴于此, 本研究基于祁连山大野口流域10.2 hm
2 青海云杉动态监测样地数据, 采用空间点格局分析方法中的成对相关函数对不同径级青海云杉种群的空间分布格局及其关联性进行分析, 旨在阐明: (1)祁连山大野口流域青海云杉种群不同径级个体的空间分布格局及其关联性如何随空间尺度进行变化; (2)不同径级个体的空间分布格局及其关联性形成的潜在生态学过程与影响机制.上述科学问题的合理解答, 能从空间格局的角度客观地揭示青海云杉种群发展规律, 可以为制定该种群的配置、种植与保护管理措施提供指导, 并对祁连山地区的植被恢复与重建、生物多样性保护等具有重要的意义和价值. ...
... 植物种群的径级结构是指不同径级组的个体在种群内所占比例和配置情况(
田润炜等, 2016 ), 是种群的生殖特性与外界环境相互作用的结果, 在一定程度上体现了种群的径级结构特征和生长发育状况, 进而可以预测种群和群落的动态发展趋势(
涂洪润等, 2019 ).本研究发现, 样地内青海云杉种群的幼苗和小树个体储备较为丰富, 其种群径级结构呈倒“J”型分布, 这说明该种群属增长型种群, 种群天然更新能力良好, 只要未来不遭受强烈干扰, 种群数量会保持逐渐增长.这与
李效雄等(2012) 和
张华等(2016) 分别对祁连山大野口流域和天老池小流域的青海云杉种群结构的研究结果一致.这可能是由青海云杉种群的生物学特性与环境因素共同作用的结果.已有研究表明, 森林更新过程中最重要和最敏感的阶段是幼苗阶段(
李晓笑等, 2011 ;
赵阳等, 2018 ), 林内有限的资源和空间往往成为种群更新的重要限制因素(
项小燕等, 2016 ).青海云杉种群的幼苗和小树个体数量多主要是由其生殖特性决定, 其种群可以产生大量高质量种子, 自然萌发形成的幼苗大都集中分布在林窗下, 形成大量小径级个体, 占据有利资源, 这是其种群在群落中持续处于优势地位的主要原因.随着小径级个体的生长, 它们对光照、水分、土壤养分等环境资源和空间需求随之增加, 由于环境资源和空间有限, 种内和种间竞争加剧, 出现强烈的自疏和他疏现象, 竞争能力弱的幼苗和小树在竞争中会大量死亡, 只有少量小径级个体能够成功转化为大径级个体, 造成样地内仅存在少量的中树和大树, 这成为该青海云杉种群更新和发展的“瓶颈” (
张婕等, 2014 ;
白卫国等, 2017 ;
于世川等, 2017 ).因此, 要实现种群数量的稳定增长, 必须加强对种群结构的保护, 尤其要重视对幼苗和小树及其生存环境的保护和改善. ...
... 森林的空间格局与空间尺度密切相关, 物种的空间分布格局随着尺度的变化而变化(
Condit et al ., 2000 ;
杨华等, 2014 ).一般来说, 小尺度上的物种分布格局主要受种群自身生物学特性(如种内种间竞争、种子扩散限制和幼苗补充过程等)影响, 而大尺度上物种的空间分布则主要受生境异质性(如光照、土壤养分、水分和温度等)影响(
Shen et al ., 2013 ).本研究表明, 青海云杉种群不同径级的青海云杉个体在较小尺度内均表现为聚集分布, 随着空间尺度的增加, 呈现由聚集分布到随机分布的变化趋势.这一结果与国内外大多数有关种群空间格局的研究结果一致(
刘建泉, 2004 ;
Guo et al ., 2013 ;
张觅等, 2014 ;
张华等, 2016 ).这是因为同一种群的不同个体对生境条件的需求基本上是一致的(
张金屯和孟东平, 2004 ), 且聚集分布有利于种群个体相互庇护, 从而抵御外界不利环境, 以提高存活率(
Diggle, 1981 ).此外, 生境的异质性容易引起种群聚集生长, 同时由于植株对光照、水分、养分等资源的竞争较大(
李海东等, 2011 ), 导致聚集强度随着空间尺度的增大而呈现下降趋势, 表现为在较小尺度上植株倾向于聚集分布逐渐过渡为在较大尺度上的随机分布. ...
... 通常认为, 由于物种的生态学和生物学特性、种子扩散限制和生境异质性等机制的影响, 较小径级的物种常呈聚集分布, 尤其在小尺度上更加明显(
祝燕等, 2011 ).而随着径级增大, 种内和种间为争夺有限资源和空间的竞争也大大增强, 常呈随机或规则分布, 聚集强度也随径级的增加而降低(
Condit et al ., 2000 ).本研究结果也符合这一规律.样地内的青海云杉种群的幼树、小树和中树这类低龄级个体多呈聚集分布, 这一方面可能是由于青海云杉的种子具有种翅, 主要依靠风力传播, 使得大多数种子散落在母树周围一定范围内且随着距离的增大其种子传播的数量逐渐下降, 这些生物学特性使其形成聚集分布的空间分布特征.实地调查过程中也发现, 青海云杉幼苗大都集中分布于林窗下, 在阴蔽的林冠下, 出现幼苗和小树死亡现象, 在地被层特别是苔藓层较厚的地方, 幼苗较少.另一方面, 低龄级种群个体较小, 处在群落的下层, 对光、热、水和土壤养分等生态因子的竞争能力较弱, 聚集可以提高其竞争能力以及抵御外界不良环境, 提高成活率.此外, 小生境的异质作用导致幼树与小树分布不均匀, 如局部地形、土壤和植被等环境因子的差异, 以及采伐、自然灾害等形成的林窗, 都为低龄级个体的大量繁殖和生长创造了良好的条件(
张华等, 2016 ).随着种群年龄的增长, 个体对光照、水分、营养等资源和空间的需求不断增加, 种内和种间竞争加剧而出现强烈的自疏和他疏现象, 导致大量林木死亡, 种群密度下降, 聚集强度逐渐减弱, 种群由聚集分布向随机分布或规则分布转变.此外, 大树主要呈随机分布, 这可能是因为大树位于林冠层, 高度较高, 树冠基本定型, 根系也伸展到一定程度, 种内竞争加剧, 自疏效应使得种群的聚集分布格局被打破, 由聚集分布向随机分布转变. ...
... 种群内不同径级个体的空间关联性描述了某一特定的时间内植物种群组成个体之间的空间分布关系及功能关系(
Shields et al ., 2014 ;
王慧杰等, 2016 ).这种关系是种群内不同径级个体间、种群与环境长期相互作用的结果, 通过它可以揭示种群的现状并预测种群发展趋势(
涂洪润等, 2019 ).空间正关联反映了种群内部正向(互惠)的生态关系, 空间负关联意味着种群内部负向(互斥)的生态关系, 空间无关联则反映了种群内部无明确的生态关系(
张金屯, 1998 ).该研究发现小尺度(<4 m)下大树个体与其他径级个体都呈负关联, 随着尺度的增大, 大树与幼树表现出正关联且关联性随尺度的增大而减弱, 而与小树、中树则表现为无关联或者微弱的负关联.小尺度下大树与其他径级个体都呈负关联, 表明在一定范围内其他径级个体都受到大树的排斥, 这可能是因为大树个体对土壤水分、光等资源的需求量更大, 竞争能力更强, 从而使得其周围其他径级个体的分布密度非常小(
张华等, 2016 ).也有研究表明青海云杉凋落物对种子的萌发和幼苗的生长有自毒作用(
王丽红等, 2017 ), 而大树周围凋落物一般也较为丰富, 导致大树周围种子和幼苗的生长受到抑制.实际调查中也发现在大树周围尤其是林冠下其他径级个体数量很少.大树个体和幼树个体在较大尺度上表现出正关联且随着尺度的增大, 关联性逐渐减弱, 这很可能是由种子扩散限制所引起的(
梁爽等, 2014 ), 大树产生的种子数量较多, 幼树的分布在一定程度上能反映种子的分布, 大树和幼树的关联性变化特点与青海云杉种子主要通过风力传播的特点具有一致性.此外, 在较大尺度下, 大树个体与小树、中树个体都表现为无关联或者微弱的负关联, 一方面表明随着尺度的增大, 大树对小树和中树的影响逐渐降低, 竞争趋于和缓, 另一方面随着径级的增大, 大树与其他径级个体关联性也发生变化, 说明不同径级青海云杉空间关联性随着发育阶段而变化, 这可能是由种群自身的生物生态学特性、种内种间关系以及环境条件共同作用的结果. ...
青海云杉不同龄级立木的点格局分析
6
2016
... 祁连山是中国西部的主要山脉之一, 位于青藏高原、蒙古高原、黄土高原三大高原交汇带, 是石羊河、黑河、疏勒河等6大内陆河和黄河流域的重要水源产流地, 维系着河西乃至西部地区生存与发展的命脉, 在国家生态建设中具有十分重要的战略地位(
Gao et al ., 2013 ;
Zhu et al ., 2017 ).祁连山山地森林的主要建群种——青海云杉(
Picea crassifolia ), 为松科云杉属(
Picea )乔木, 是中国青藏高原东北边缘特有树种, 多以纯林形式存在, 呈带状或斑块状分布于海拔2 300-3 300 m的阴坡和半阴坡, 分布面积占祁连山水源涵养林面积的24.74%, 占乔木林面积的75.72%, 对于维系祁连山的生物多样性、水源涵养功能乃至区域生态平衡都具有至关重要的作用(
田风霞等, 2012 ;
何芳兰等, 2016 ;
张华等, 2016 ).部分****(
赵丽琼等, 2010 ;
张华等, 2016 )虽然已对青海云杉种群空间格局及其关联性进行了研究, 但是已有研究的空间尺度相对较小, 仅能揭示小尺度下的种群空间分布格局, 难以客观地反映青海云杉种群空间分布的特点和种内关系.鉴于此, 本研究基于祁连山大野口流域10.2 hm
2 青海云杉动态监测样地数据, 采用空间点格局分析方法中的成对相关函数对不同径级青海云杉种群的空间分布格局及其关联性进行分析, 旨在阐明: (1)祁连山大野口流域青海云杉种群不同径级个体的空间分布格局及其关联性如何随空间尺度进行变化; (2)不同径级个体的空间分布格局及其关联性形成的潜在生态学过程与影响机制.上述科学问题的合理解答, 能从空间格局的角度客观地揭示青海云杉种群发展规律, 可以为制定该种群的配置、种植与保护管理措施提供指导, 并对祁连山地区的植被恢复与重建、生物多样性保护等具有重要的意义和价值. ...
... ;
张华等, 2016 )虽然已对青海云杉种群空间格局及其关联性进行了研究, 但是已有研究的空间尺度相对较小, 仅能揭示小尺度下的种群空间分布格局, 难以客观地反映青海云杉种群空间分布的特点和种内关系.鉴于此, 本研究基于祁连山大野口流域10.2 hm
2 青海云杉动态监测样地数据, 采用空间点格局分析方法中的成对相关函数对不同径级青海云杉种群的空间分布格局及其关联性进行分析, 旨在阐明: (1)祁连山大野口流域青海云杉种群不同径级个体的空间分布格局及其关联性如何随空间尺度进行变化; (2)不同径级个体的空间分布格局及其关联性形成的潜在生态学过程与影响机制.上述科学问题的合理解答, 能从空间格局的角度客观地揭示青海云杉种群发展规律, 可以为制定该种群的配置、种植与保护管理措施提供指导, 并对祁连山地区的植被恢复与重建、生物多样性保护等具有重要的意义和价值. ...
... 植物种群的径级结构是指不同径级组的个体在种群内所占比例和配置情况(
田润炜等, 2016 ), 是种群的生殖特性与外界环境相互作用的结果, 在一定程度上体现了种群的径级结构特征和生长发育状况, 进而可以预测种群和群落的动态发展趋势(
涂洪润等, 2019 ).本研究发现, 样地内青海云杉种群的幼苗和小树个体储备较为丰富, 其种群径级结构呈倒“J”型分布, 这说明该种群属增长型种群, 种群天然更新能力良好, 只要未来不遭受强烈干扰, 种群数量会保持逐渐增长.这与
李效雄等(2012) 和
张华等(2016) 分别对祁连山大野口流域和天老池小流域的青海云杉种群结构的研究结果一致.这可能是由青海云杉种群的生物学特性与环境因素共同作用的结果.已有研究表明, 森林更新过程中最重要和最敏感的阶段是幼苗阶段(
李晓笑等, 2011 ;
赵阳等, 2018 ), 林内有限的资源和空间往往成为种群更新的重要限制因素(
项小燕等, 2016 ).青海云杉种群的幼苗和小树个体数量多主要是由其生殖特性决定, 其种群可以产生大量高质量种子, 自然萌发形成的幼苗大都集中分布在林窗下, 形成大量小径级个体, 占据有利资源, 这是其种群在群落中持续处于优势地位的主要原因.随着小径级个体的生长, 它们对光照、水分、土壤养分等环境资源和空间需求随之增加, 由于环境资源和空间有限, 种内和种间竞争加剧, 出现强烈的自疏和他疏现象, 竞争能力弱的幼苗和小树在竞争中会大量死亡, 只有少量小径级个体能够成功转化为大径级个体, 造成样地内仅存在少量的中树和大树, 这成为该青海云杉种群更新和发展的“瓶颈” (
张婕等, 2014 ;
白卫国等, 2017 ;
于世川等, 2017 ).因此, 要实现种群数量的稳定增长, 必须加强对种群结构的保护, 尤其要重视对幼苗和小树及其生存环境的保护和改善. ...
... 森林的空间格局与空间尺度密切相关, 物种的空间分布格局随着尺度的变化而变化(
Condit et al ., 2000 ;
杨华等, 2014 ).一般来说, 小尺度上的物种分布格局主要受种群自身生物学特性(如种内种间竞争、种子扩散限制和幼苗补充过程等)影响, 而大尺度上物种的空间分布则主要受生境异质性(如光照、土壤养分、水分和温度等)影响(
Shen et al ., 2013 ).本研究表明, 青海云杉种群不同径级的青海云杉个体在较小尺度内均表现为聚集分布, 随着空间尺度的增加, 呈现由聚集分布到随机分布的变化趋势.这一结果与国内外大多数有关种群空间格局的研究结果一致(
刘建泉, 2004 ;
Guo et al ., 2013 ;
张觅等, 2014 ;
张华等, 2016 ).这是因为同一种群的不同个体对生境条件的需求基本上是一致的(
张金屯和孟东平, 2004 ), 且聚集分布有利于种群个体相互庇护, 从而抵御外界不利环境, 以提高存活率(
Diggle, 1981 ).此外, 生境的异质性容易引起种群聚集生长, 同时由于植株对光照、水分、养分等资源的竞争较大(
李海东等, 2011 ), 导致聚集强度随着空间尺度的增大而呈现下降趋势, 表现为在较小尺度上植株倾向于聚集分布逐渐过渡为在较大尺度上的随机分布. ...
... 通常认为, 由于物种的生态学和生物学特性、种子扩散限制和生境异质性等机制的影响, 较小径级的物种常呈聚集分布, 尤其在小尺度上更加明显(
祝燕等, 2011 ).而随着径级增大, 种内和种间为争夺有限资源和空间的竞争也大大增强, 常呈随机或规则分布, 聚集强度也随径级的增加而降低(
Condit et al ., 2000 ).本研究结果也符合这一规律.样地内的青海云杉种群的幼树、小树和中树这类低龄级个体多呈聚集分布, 这一方面可能是由于青海云杉的种子具有种翅, 主要依靠风力传播, 使得大多数种子散落在母树周围一定范围内且随着距离的增大其种子传播的数量逐渐下降, 这些生物学特性使其形成聚集分布的空间分布特征.实地调查过程中也发现, 青海云杉幼苗大都集中分布于林窗下, 在阴蔽的林冠下, 出现幼苗和小树死亡现象, 在地被层特别是苔藓层较厚的地方, 幼苗较少.另一方面, 低龄级种群个体较小, 处在群落的下层, 对光、热、水和土壤养分等生态因子的竞争能力较弱, 聚集可以提高其竞争能力以及抵御外界不良环境, 提高成活率.此外, 小生境的异质作用导致幼树与小树分布不均匀, 如局部地形、土壤和植被等环境因子的差异, 以及采伐、自然灾害等形成的林窗, 都为低龄级个体的大量繁殖和生长创造了良好的条件(
张华等, 2016 ).随着种群年龄的增长, 个体对光照、水分、营养等资源和空间的需求不断增加, 种内和种间竞争加剧而出现强烈的自疏和他疏现象, 导致大量林木死亡, 种群密度下降, 聚集强度逐渐减弱, 种群由聚集分布向随机分布或规则分布转变.此外, 大树主要呈随机分布, 这可能是因为大树位于林冠层, 高度较高, 树冠基本定型, 根系也伸展到一定程度, 种内竞争加剧, 自疏效应使得种群的聚集分布格局被打破, 由聚集分布向随机分布转变. ...
... 种群内不同径级个体的空间关联性描述了某一特定的时间内植物种群组成个体之间的空间分布关系及功能关系(
Shields et al ., 2014 ;
王慧杰等, 2016 ).这种关系是种群内不同径级个体间、种群与环境长期相互作用的结果, 通过它可以揭示种群的现状并预测种群发展趋势(
涂洪润等, 2019 ).空间正关联反映了种群内部正向(互惠)的生态关系, 空间负关联意味着种群内部负向(互斥)的生态关系, 空间无关联则反映了种群内部无明确的生态关系(
张金屯, 1998 ).该研究发现小尺度(<4 m)下大树个体与其他径级个体都呈负关联, 随着尺度的增大, 大树与幼树表现出正关联且关联性随尺度的增大而减弱, 而与小树、中树则表现为无关联或者微弱的负关联.小尺度下大树与其他径级个体都呈负关联, 表明在一定范围内其他径级个体都受到大树的排斥, 这可能是因为大树个体对土壤水分、光等资源的需求量更大, 竞争能力更强, 从而使得其周围其他径级个体的分布密度非常小(
张华等, 2016 ).也有研究表明青海云杉凋落物对种子的萌发和幼苗的生长有自毒作用(
王丽红等, 2017 ), 而大树周围凋落物一般也较为丰富, 导致大树周围种子和幼苗的生长受到抑制.实际调查中也发现在大树周围尤其是林冠下其他径级个体数量很少.大树个体和幼树个体在较大尺度上表现出正关联且随着尺度的增大, 关联性逐渐减弱, 这很可能是由种子扩散限制所引起的(
梁爽等, 2014 ), 大树产生的种子数量较多, 幼树的分布在一定程度上能反映种子的分布, 大树和幼树的关联性变化特点与青海云杉种子主要通过风力传播的特点具有一致性.此外, 在较大尺度下, 大树个体与小树、中树个体都表现为无关联或者微弱的负关联, 一方面表明随着尺度的增大, 大树对小树和中树的影响逐渐降低, 竞争趋于和缓, 另一方面随着径级的增大, 大树与其他径级个体关联性也发生变化, 说明不同径级青海云杉空间关联性随着发育阶段而变化, 这可能是由种群自身的生物生态学特性、种内种间关系以及环境条件共同作用的结果. ...
灵空山辽东栎种群年龄结构与动态
1
2014
... 植物种群的径级结构是指不同径级组的个体在种群内所占比例和配置情况(
田润炜等, 2016 ), 是种群的生殖特性与外界环境相互作用的结果, 在一定程度上体现了种群的径级结构特征和生长发育状况, 进而可以预测种群和群落的动态发展趋势(
涂洪润等, 2019 ).本研究发现, 样地内青海云杉种群的幼苗和小树个体储备较为丰富, 其种群径级结构呈倒“J”型分布, 这说明该种群属增长型种群, 种群天然更新能力良好, 只要未来不遭受强烈干扰, 种群数量会保持逐渐增长.这与
李效雄等(2012) 和
张华等(2016) 分别对祁连山大野口流域和天老池小流域的青海云杉种群结构的研究结果一致.这可能是由青海云杉种群的生物学特性与环境因素共同作用的结果.已有研究表明, 森林更新过程中最重要和最敏感的阶段是幼苗阶段(
李晓笑等, 2011 ;
赵阳等, 2018 ), 林内有限的资源和空间往往成为种群更新的重要限制因素(
项小燕等, 2016 ).青海云杉种群的幼苗和小树个体数量多主要是由其生殖特性决定, 其种群可以产生大量高质量种子, 自然萌发形成的幼苗大都集中分布在林窗下, 形成大量小径级个体, 占据有利资源, 这是其种群在群落中持续处于优势地位的主要原因.随着小径级个体的生长, 它们对光照、水分、土壤养分等环境资源和空间需求随之增加, 由于环境资源和空间有限, 种内和种间竞争加剧, 出现强烈的自疏和他疏现象, 竞争能力弱的幼苗和小树在竞争中会大量死亡, 只有少量小径级个体能够成功转化为大径级个体, 造成样地内仅存在少量的中树和大树, 这成为该青海云杉种群更新和发展的“瓶颈” (
张婕等, 2014 ;
白卫国等, 2017 ;
于世川等, 2017 ).因此, 要实现种群数量的稳定增长, 必须加强对种群结构的保护, 尤其要重视对幼苗和小树及其生存环境的保护和改善. ...
灵空山辽东栎种群年龄结构与动态
1
2014
... 植物种群的径级结构是指不同径级组的个体在种群内所占比例和配置情况(
田润炜等, 2016 ), 是种群的生殖特性与外界环境相互作用的结果, 在一定程度上体现了种群的径级结构特征和生长发育状况, 进而可以预测种群和群落的动态发展趋势(
涂洪润等, 2019 ).本研究发现, 样地内青海云杉种群的幼苗和小树个体储备较为丰富, 其种群径级结构呈倒“J”型分布, 这说明该种群属增长型种群, 种群天然更新能力良好, 只要未来不遭受强烈干扰, 种群数量会保持逐渐增长.这与
李效雄等(2012) 和
张华等(2016) 分别对祁连山大野口流域和天老池小流域的青海云杉种群结构的研究结果一致.这可能是由青海云杉种群的生物学特性与环境因素共同作用的结果.已有研究表明, 森林更新过程中最重要和最敏感的阶段是幼苗阶段(
李晓笑等, 2011 ;
赵阳等, 2018 ), 林内有限的资源和空间往往成为种群更新的重要限制因素(
项小燕等, 2016 ).青海云杉种群的幼苗和小树个体数量多主要是由其生殖特性决定, 其种群可以产生大量高质量种子, 自然萌发形成的幼苗大都集中分布在林窗下, 形成大量小径级个体, 占据有利资源, 这是其种群在群落中持续处于优势地位的主要原因.随着小径级个体的生长, 它们对光照、水分、土壤养分等环境资源和空间需求随之增加, 由于环境资源和空间有限, 种内和种间竞争加剧, 出现强烈的自疏和他疏现象, 竞争能力弱的幼苗和小树在竞争中会大量死亡, 只有少量小径级个体能够成功转化为大径级个体, 造成样地内仅存在少量的中树和大树, 这成为该青海云杉种群更新和发展的“瓶颈” (
张婕等, 2014 ;
白卫国等, 2017 ;
于世川等, 2017 ).因此, 要实现种群数量的稳定增长, 必须加强对种群结构的保护, 尤其要重视对幼苗和小树及其生存环境的保护和改善. ...
植物种群空间分布的点格局分析
2
1998
... 空间分布格局的研究方法众多, 早期的研究方法主要有: 以频度/密度为基础的样方法、以距离为基础的无样地法、方差区组分析法、双向轨迹方差分析、
χ 2 检验和分形理论等(
张金屯, 1998 ;
王本洋和余世孝, 2005 ).传统的样方取样和分析方法常基于单一尺度(固定大小样方)的若干样方, 常采用
χ 2 分析法和方差/均值比率法等统计分析方法, 然而种群空间分布格局及其关联性对种群密度和空间尺度均有很强的依赖性, 在不同的尺度上往往表现出不同的类型, 故早期的取样和分析方法只能研究特定尺度下种群的空间分布格局特征, 难以全面地反映森林内物种的空间分布特点和种间关系(
He et al ., 1997 ;
李立等, 2010 ).鉴于上述研究方法的局限性,
Ripley (1977) 和
Diggle (1983) 分别提出和完善了点格局分析法, 以分析各种尺度上的空间分布格局.该方法以数字化的植物个体位置信息为基本数据, 将每个个体都视为二维空间中的一个点, 所有个体组成了植物种的空间分布图, 然后在空间分布图的基础上进行种群空间格局分析.其优点是能最大限度地利用点与点之间的距离, 不仅能够提供全面的空间格局信息, 而且具有较强的检验能力, 克服了传统研究方法中尺度单一等不足, 可以分析任意尺度下种群的空间格局特征及种内、种间关系(
Wiegand & Moloney, 2004 ;
Schleicher et al ., 2011 ;
沈志强等, 2016 ;
涂洪润等, 2019 ).因此, 空间点格局分析法在种群空间格局分布特征及潜在生态过程假说的研究方面得到了广泛应用. ...
... 种群内不同径级个体的空间关联性描述了某一特定的时间内植物种群组成个体之间的空间分布关系及功能关系(
Shields et al ., 2014 ;
王慧杰等, 2016 ).这种关系是种群内不同径级个体间、种群与环境长期相互作用的结果, 通过它可以揭示种群的现状并预测种群发展趋势(
涂洪润等, 2019 ).空间正关联反映了种群内部正向(互惠)的生态关系, 空间负关联意味着种群内部负向(互斥)的生态关系, 空间无关联则反映了种群内部无明确的生态关系(
张金屯, 1998 ).该研究发现小尺度(<4 m)下大树个体与其他径级个体都呈负关联, 随着尺度的增大, 大树与幼树表现出正关联且关联性随尺度的增大而减弱, 而与小树、中树则表现为无关联或者微弱的负关联.小尺度下大树与其他径级个体都呈负关联, 表明在一定范围内其他径级个体都受到大树的排斥, 这可能是因为大树个体对土壤水分、光等资源的需求量更大, 竞争能力更强, 从而使得其周围其他径级个体的分布密度非常小(
张华等, 2016 ).也有研究表明青海云杉凋落物对种子的萌发和幼苗的生长有自毒作用(
王丽红等, 2017 ), 而大树周围凋落物一般也较为丰富, 导致大树周围种子和幼苗的生长受到抑制.实际调查中也发现在大树周围尤其是林冠下其他径级个体数量很少.大树个体和幼树个体在较大尺度上表现出正关联且随着尺度的增大, 关联性逐渐减弱, 这很可能是由种子扩散限制所引起的(
梁爽等, 2014 ), 大树产生的种子数量较多, 幼树的分布在一定程度上能反映种子的分布, 大树和幼树的关联性变化特点与青海云杉种子主要通过风力传播的特点具有一致性.此外, 在较大尺度下, 大树个体与小树、中树个体都表现为无关联或者微弱的负关联, 一方面表明随着尺度的增大, 大树对小树和中树的影响逐渐降低, 竞争趋于和缓, 另一方面随着径级的增大, 大树与其他径级个体关联性也发生变化, 说明不同径级青海云杉空间关联性随着发育阶段而变化, 这可能是由种群自身的生物生态学特性、种内种间关系以及环境条件共同作用的结果. ...
植物种群空间分布的点格局分析
2
1998
... 空间分布格局的研究方法众多, 早期的研究方法主要有: 以频度/密度为基础的样方法、以距离为基础的无样地法、方差区组分析法、双向轨迹方差分析、
χ 2 检验和分形理论等(
张金屯, 1998 ;
王本洋和余世孝, 2005 ).传统的样方取样和分析方法常基于单一尺度(固定大小样方)的若干样方, 常采用
χ 2 分析法和方差/均值比率法等统计分析方法, 然而种群空间分布格局及其关联性对种群密度和空间尺度均有很强的依赖性, 在不同的尺度上往往表现出不同的类型, 故早期的取样和分析方法只能研究特定尺度下种群的空间分布格局特征, 难以全面地反映森林内物种的空间分布特点和种间关系(
He et al ., 1997 ;
李立等, 2010 ).鉴于上述研究方法的局限性,
Ripley (1977) 和
Diggle (1983) 分别提出和完善了点格局分析法, 以分析各种尺度上的空间分布格局.该方法以数字化的植物个体位置信息为基本数据, 将每个个体都视为二维空间中的一个点, 所有个体组成了植物种的空间分布图, 然后在空间分布图的基础上进行种群空间格局分析.其优点是能最大限度地利用点与点之间的距离, 不仅能够提供全面的空间格局信息, 而且具有较强的检验能力, 克服了传统研究方法中尺度单一等不足, 可以分析任意尺度下种群的空间格局特征及种内、种间关系(
Wiegand & Moloney, 2004 ;
Schleicher et al ., 2011 ;
沈志强等, 2016 ;
涂洪润等, 2019 ).因此, 空间点格局分析法在种群空间格局分布特征及潜在生态过程假说的研究方面得到了广泛应用. ...
... 种群内不同径级个体的空间关联性描述了某一特定的时间内植物种群组成个体之间的空间分布关系及功能关系(
Shields et al ., 2014 ;
王慧杰等, 2016 ).这种关系是种群内不同径级个体间、种群与环境长期相互作用的结果, 通过它可以揭示种群的现状并预测种群发展趋势(
涂洪润等, 2019 ).空间正关联反映了种群内部正向(互惠)的生态关系, 空间负关联意味着种群内部负向(互斥)的生态关系, 空间无关联则反映了种群内部无明确的生态关系(
张金屯, 1998 ).该研究发现小尺度(<4 m)下大树个体与其他径级个体都呈负关联, 随着尺度的增大, 大树与幼树表现出正关联且关联性随尺度的增大而减弱, 而与小树、中树则表现为无关联或者微弱的负关联.小尺度下大树与其他径级个体都呈负关联, 表明在一定范围内其他径级个体都受到大树的排斥, 这可能是因为大树个体对土壤水分、光等资源的需求量更大, 竞争能力更强, 从而使得其周围其他径级个体的分布密度非常小(
张华等, 2016 ).也有研究表明青海云杉凋落物对种子的萌发和幼苗的生长有自毒作用(
王丽红等, 2017 ), 而大树周围凋落物一般也较为丰富, 导致大树周围种子和幼苗的生长受到抑制.实际调查中也发现在大树周围尤其是林冠下其他径级个体数量很少.大树个体和幼树个体在较大尺度上表现出正关联且随着尺度的增大, 关联性逐渐减弱, 这很可能是由种子扩散限制所引起的(
梁爽等, 2014 ), 大树产生的种子数量较多, 幼树的分布在一定程度上能反映种子的分布, 大树和幼树的关联性变化特点与青海云杉种子主要通过风力传播的特点具有一致性.此外, 在较大尺度下, 大树个体与小树、中树个体都表现为无关联或者微弱的负关联, 一方面表明随着尺度的增大, 大树对小树和中树的影响逐渐降低, 竞争趋于和缓, 另一方面随着径级的增大, 大树与其他径级个体关联性也发生变化, 说明不同径级青海云杉空间关联性随着发育阶段而变化, 这可能是由种群自身的生物生态学特性、种内种间关系以及环境条件共同作用的结果. ...
芦芽山华北落叶松林不同龄级立木的点格局分析
3
2004
... 种群的空间格局是指特定时间内, 种群所有个体在其水平生活空间内的分布状况及其属性在空间上的配置特征, 往往是由种群自身生物学特性、种内种间竞争、环境条件等因素综合作用的结果(
林玥, 2008 ;
Liu et al ., 2020 ).种群的空间分布格局和空间关联性是种群空间格局研究的2个主要内容, 也是种群生态关系在空间格局上的两种表现形式(
张金屯和孟东平, 2004 ).大量研究表明种群的空间分布格局与空间关联性关系密切, 聚集分布和空间正关联表明种群内部正向(互惠)的生态关系, 规则分布和空间负关联则表明种群内部负向(互斥)的生态关系, 随机分布和空间无关联则代表着种群内部不存在明确的生态关系(
杨洪晓, 2006 ).因此, 研究植物种群的空间分布格局及其关联性不仅能够了解种群的空间水平结构与生物、生态学习性(如生活史策略、是否喜光或耐阴等)、种群间相互作用、种群与环境因子的相互作用规律(如植物与生长环境间的适合度、生境异质性等)以及种群在群落中的地位和生态学功能, 更重要的是可以揭示这种分布格局及其关联性的潜在成因, 进而推演种群动态变化的生态学过程(如种内和种间竞争、种子扩散和干扰等), 深化对群落结构的认识, 揭示植物群落的生物多样性形成和维持机制(
McDonald et al ., 2003 ;
Hou et al , 2004 ;
Wiegand & Moloney, 2004 ;
Nathan, 2006 ;
Wang et al ., 2010 ;
Guo et al ., 2013 ). ...
... 两个径级之间的空间关联性分析, 实际上是2个径级之间的多变量点格局分析, 也叫多元点格局分析(
张金屯, 2004 ).该研究采用双变量成对相关函数(
g 12 (
r )), 分析不同径级青海云杉个体的空间关联性.其计算公式如下所示: ...
... 森林的空间格局与空间尺度密切相关, 物种的空间分布格局随着尺度的变化而变化(
Condit et al ., 2000 ;
杨华等, 2014 ).一般来说, 小尺度上的物种分布格局主要受种群自身生物学特性(如种内种间竞争、种子扩散限制和幼苗补充过程等)影响, 而大尺度上物种的空间分布则主要受生境异质性(如光照、土壤养分、水分和温度等)影响(
Shen et al ., 2013 ).本研究表明, 青海云杉种群不同径级的青海云杉个体在较小尺度内均表现为聚集分布, 随着空间尺度的增加, 呈现由聚集分布到随机分布的变化趋势.这一结果与国内外大多数有关种群空间格局的研究结果一致(
刘建泉, 2004 ;
Guo et al ., 2013 ;
张觅等, 2014 ;
张华等, 2016 ).这是因为同一种群的不同个体对生境条件的需求基本上是一致的(
张金屯和孟东平, 2004 ), 且聚集分布有利于种群个体相互庇护, 从而抵御外界不利环境, 以提高存活率(
Diggle, 1981 ).此外, 生境的异质性容易引起种群聚集生长, 同时由于植株对光照、水分、养分等资源的竞争较大(
李海东等, 2011 ), 导致聚集强度随着空间尺度的增大而呈现下降趋势, 表现为在较小尺度上植株倾向于聚集分布逐渐过渡为在较大尺度上的随机分布. ...
芦芽山华北落叶松林不同龄级立木的点格局分析
3
2004
... 种群的空间格局是指特定时间内, 种群所有个体在其水平生活空间内的分布状况及其属性在空间上的配置特征, 往往是由种群自身生物学特性、种内种间竞争、环境条件等因素综合作用的结果(
林玥, 2008 ;
Liu et al ., 2020 ).种群的空间分布格局和空间关联性是种群空间格局研究的2个主要内容, 也是种群生态关系在空间格局上的两种表现形式(
张金屯和孟东平, 2004 ).大量研究表明种群的空间分布格局与空间关联性关系密切, 聚集分布和空间正关联表明种群内部正向(互惠)的生态关系, 规则分布和空间负关联则表明种群内部负向(互斥)的生态关系, 随机分布和空间无关联则代表着种群内部不存在明确的生态关系(
杨洪晓, 2006 ).因此, 研究植物种群的空间分布格局及其关联性不仅能够了解种群的空间水平结构与生物、生态学习性(如生活史策略、是否喜光或耐阴等)、种群间相互作用、种群与环境因子的相互作用规律(如植物与生长环境间的适合度、生境异质性等)以及种群在群落中的地位和生态学功能, 更重要的是可以揭示这种分布格局及其关联性的潜在成因, 进而推演种群动态变化的生态学过程(如种内和种间竞争、种子扩散和干扰等), 深化对群落结构的认识, 揭示植物群落的生物多样性形成和维持机制(
McDonald et al ., 2003 ;
Hou et al , 2004 ;
Wiegand & Moloney, 2004 ;
Nathan, 2006 ;
Wang et al ., 2010 ;
Guo et al ., 2013 ). ...
... 两个径级之间的空间关联性分析, 实际上是2个径级之间的多变量点格局分析, 也叫多元点格局分析(
张金屯, 2004 ).该研究采用双变量成对相关函数(
g 12 (
r )), 分析不同径级青海云杉个体的空间关联性.其计算公式如下所示: ...
... 森林的空间格局与空间尺度密切相关, 物种的空间分布格局随着尺度的变化而变化(
Condit et al ., 2000 ;
杨华等, 2014 ).一般来说, 小尺度上的物种分布格局主要受种群自身生物学特性(如种内种间竞争、种子扩散限制和幼苗补充过程等)影响, 而大尺度上物种的空间分布则主要受生境异质性(如光照、土壤养分、水分和温度等)影响(
Shen et al ., 2013 ).本研究表明, 青海云杉种群不同径级的青海云杉个体在较小尺度内均表现为聚集分布, 随着空间尺度的增加, 呈现由聚集分布到随机分布的变化趋势.这一结果与国内外大多数有关种群空间格局的研究结果一致(
刘建泉, 2004 ;
Guo et al ., 2013 ;
张觅等, 2014 ;
张华等, 2016 ).这是因为同一种群的不同个体对生境条件的需求基本上是一致的(
张金屯和孟东平, 2004 ), 且聚集分布有利于种群个体相互庇护, 从而抵御外界不利环境, 以提高存活率(
Diggle, 1981 ).此外, 生境的异质性容易引起种群聚集生长, 同时由于植株对光照、水分、养分等资源的竞争较大(
李海东等, 2011 ), 导致聚集强度随着空间尺度的增大而呈现下降趋势, 表现为在较小尺度上植株倾向于聚集分布逐渐过渡为在较大尺度上的随机分布. ...
小兴安岭凉水谷地云冷杉林群落组成与空间格局
2
2014
... 由于
g (
r )的非聚集属性, 利用它进行点格局分析时需利用零模型来观察实际过程和理论过程之间的差异(
Stoyan & Penttinen, 2000 ).祁连山大野口流域青海云杉林样地地势相对平坦, 地形因子对于种群空间格局及其关联性影响较弱, 因此采用完全空间随机零模型作为零模型, 即假设物种的空间分布完全随机, 没有潜在的生物学过程发挥作用, 观察青海云杉种群不同径级个体的空间分布格局及其关联性与匀质泊松分布是否存在差异(
张觅等, 2014 ).空间尺度设定为0-60 m, 步长为1 m.采用蒙特卡罗(Monte-Carlo)随机模拟999次产生置信度为99%的包迹线, 以检验物种分布格局及关联性是否显著.在单变量分析中, 若实际分布的
g (
r )值落在包迹线之内, 则符合零模型, 为随机分布; 若在上包迹线以上, 则呈聚集分布; 若在下包迹线以下, 则为规则分布.在双变量分析中, 若实际分布的
g 12 (
r )值落在包迹线之间, 表示接受零假设, 说明不同径级间个体相互独立, 无空间相关性; 若在上包迹线以上, 则两径级间为正相关; 若落在下包迹线以下, 则两径级间为负相关关系(
Hao et al ., 2017 ).该研究所有统计与分析均在R 4.0.1软件中“spatstat”程序包(
Baddeley & Turner, 2005 )的相关函数中完成. ...
... 森林的空间格局与空间尺度密切相关, 物种的空间分布格局随着尺度的变化而变化(
Condit et al ., 2000 ;
杨华等, 2014 ).一般来说, 小尺度上的物种分布格局主要受种群自身生物学特性(如种内种间竞争、种子扩散限制和幼苗补充过程等)影响, 而大尺度上物种的空间分布则主要受生境异质性(如光照、土壤养分、水分和温度等)影响(
Shen et al ., 2013 ).本研究表明, 青海云杉种群不同径级的青海云杉个体在较小尺度内均表现为聚集分布, 随着空间尺度的增加, 呈现由聚集分布到随机分布的变化趋势.这一结果与国内外大多数有关种群空间格局的研究结果一致(
刘建泉, 2004 ;
Guo et al ., 2013 ;
张觅等, 2014 ;
张华等, 2016 ).这是因为同一种群的不同个体对生境条件的需求基本上是一致的(
张金屯和孟东平, 2004 ), 且聚集分布有利于种群个体相互庇护, 从而抵御外界不利环境, 以提高存活率(
Diggle, 1981 ).此外, 生境的异质性容易引起种群聚集生长, 同时由于植株对光照、水分、养分等资源的竞争较大(
李海东等, 2011 ), 导致聚集强度随着空间尺度的增大而呈现下降趋势, 表现为在较小尺度上植株倾向于聚集分布逐渐过渡为在较大尺度上的随机分布. ...
小兴安岭凉水谷地云冷杉林群落组成与空间格局
2
2014
... 由于
g (
r )的非聚集属性, 利用它进行点格局分析时需利用零模型来观察实际过程和理论过程之间的差异(
Stoyan & Penttinen, 2000 ).祁连山大野口流域青海云杉林样地地势相对平坦, 地形因子对于种群空间格局及其关联性影响较弱, 因此采用完全空间随机零模型作为零模型, 即假设物种的空间分布完全随机, 没有潜在的生物学过程发挥作用, 观察青海云杉种群不同径级个体的空间分布格局及其关联性与匀质泊松分布是否存在差异(
张觅等, 2014 ).空间尺度设定为0-60 m, 步长为1 m.采用蒙特卡罗(Monte-Carlo)随机模拟999次产生置信度为99%的包迹线, 以检验物种分布格局及关联性是否显著.在单变量分析中, 若实际分布的
g (
r )值落在包迹线之内, 则符合零模型, 为随机分布; 若在上包迹线以上, 则呈聚集分布; 若在下包迹线以下, 则为规则分布.在双变量分析中, 若实际分布的
g 12 (
r )值落在包迹线之间, 表示接受零假设, 说明不同径级间个体相互独立, 无空间相关性; 若在上包迹线以上, 则两径级间为正相关; 若落在下包迹线以下, 则两径级间为负相关关系(
Hao et al ., 2017 ).该研究所有统计与分析均在R 4.0.1软件中“spatstat”程序包(
Baddeley & Turner, 2005 )的相关函数中完成. ...
... 森林的空间格局与空间尺度密切相关, 物种的空间分布格局随着尺度的变化而变化(
Condit et al ., 2000 ;
杨华等, 2014 ).一般来说, 小尺度上的物种分布格局主要受种群自身生物学特性(如种内种间竞争、种子扩散限制和幼苗补充过程等)影响, 而大尺度上物种的空间分布则主要受生境异质性(如光照、土壤养分、水分和温度等)影响(
Shen et al ., 2013 ).本研究表明, 青海云杉种群不同径级的青海云杉个体在较小尺度内均表现为聚集分布, 随着空间尺度的增加, 呈现由聚集分布到随机分布的变化趋势.这一结果与国内外大多数有关种群空间格局的研究结果一致(
刘建泉, 2004 ;
Guo et al ., 2013 ;
张觅等, 2014 ;
张华等, 2016 ).这是因为同一种群的不同个体对生境条件的需求基本上是一致的(
张金屯和孟东平, 2004 ), 且聚集分布有利于种群个体相互庇护, 从而抵御外界不利环境, 以提高存活率(
Diggle, 1981 ).此外, 生境的异质性容易引起种群聚集生长, 同时由于植株对光照、水分、养分等资源的竞争较大(
李海东等, 2011 ), 导致聚集强度随着空间尺度的增大而呈现下降趋势, 表现为在较小尺度上植株倾向于聚集分布逐渐过渡为在较大尺度上的随机分布. ...
甘肃大野口青海云杉种群的空间分布格局
2
2010
... 祁连山是中国西部的主要山脉之一, 位于青藏高原、蒙古高原、黄土高原三大高原交汇带, 是石羊河、黑河、疏勒河等6大内陆河和黄河流域的重要水源产流地, 维系着河西乃至西部地区生存与发展的命脉, 在国家生态建设中具有十分重要的战略地位(
Gao et al ., 2013 ;
Zhu et al ., 2017 ).祁连山山地森林的主要建群种——青海云杉(
Picea crassifolia ), 为松科云杉属(
Picea )乔木, 是中国青藏高原东北边缘特有树种, 多以纯林形式存在, 呈带状或斑块状分布于海拔2 300-3 300 m的阴坡和半阴坡, 分布面积占祁连山水源涵养林面积的24.74%, 占乔木林面积的75.72%, 对于维系祁连山的生物多样性、水源涵养功能乃至区域生态平衡都具有至关重要的作用(
田风霞等, 2012 ;
何芳兰等, 2016 ;
张华等, 2016 ).部分****(
赵丽琼等, 2010 ;
张华等, 2016 )虽然已对青海云杉种群空间格局及其关联性进行了研究, 但是已有研究的空间尺度相对较小, 仅能揭示小尺度下的种群空间分布格局, 难以客观地反映青海云杉种群空间分布的特点和种内关系.鉴于此, 本研究基于祁连山大野口流域10.2 hm
2 青海云杉动态监测样地数据, 采用空间点格局分析方法中的成对相关函数对不同径级青海云杉种群的空间分布格局及其关联性进行分析, 旨在阐明: (1)祁连山大野口流域青海云杉种群不同径级个体的空间分布格局及其关联性如何随空间尺度进行变化; (2)不同径级个体的空间分布格局及其关联性形成的潜在生态学过程与影响机制.上述科学问题的合理解答, 能从空间格局的角度客观地揭示青海云杉种群发展规律, 可以为制定该种群的配置、种植与保护管理措施提供指导, 并对祁连山地区的植被恢复与重建、生物多样性保护等具有重要的意义和价值. ...
... 研究样地位于祁连山中段北坡大野口流域青海云杉林内, 地理位置为100.25° E, 38.53° N.样地最低海拔2 856 m, 最高海拔2 962 m, 最大高差106 m, 坡向西北, 坡度13°.根据祁连山水源涵养林研究院的资料, 样地所在的大野口流域气候属于温带大陆性高寒半湿润山地气候, 雨热同期, 冬季寒冷漫长, 夏季短暂, 年平均气温5.4 ℃, 最冷月(1月)平均气温为-12.5 ℃, 最热月(7月)平均气温为19.6 ℃, 年降水量为300-500 mm, 主要集中在6-9月, 年平均相对湿度60% (1994-2004年), 年蒸发量为1 488 mm.样地的成土母质为钙质岩, 土壤类型为山地森林灰褐土, 土壤pH 6.3-7.9, 厚度约1 m.林分起源为天然次生林, 为青海云杉纯林, 林分结构简单, 郁闭度较高, 林下光照不足.青海云杉群落成层现象明显, 可明显划分为乔木层、灌木层、草本层和苔藓层4个层次(
赵丽琼等, 2010 ).其中, 乔木树种组成单一, 除青海云杉外, 只有少量祁连圆柏(
Juniperus przewalskii ); 灌木生长发育不良, 物种较少, 主要有金露梅(
Potentilla fruticosa )、银露梅(
Potentilla glabra )、鬼箭锦鸡儿(
Caragana jubata )、吉拉柳(
Salix gilashanica )、鲜黄小檗(
Berberis diaphana )和高山绣线菊(
Spiraea alpina ); 草本植物物种较丰富, 主要有黑穗薹草(
Carex atrata )、珠芽蓼(
Polygonum viviparum )、针茅(
Stipa capillata )和马蔺(
Iris lactea )等典型高山草甸植物.苔藓层发育良好, 盖度达80%以上, 厚度约10 cm, 主要物种有山羽藓(
Abietinella abietina )、欧灰藓(
Hypnum cupressiforme )和匐灯藓(
Plagiomnium cuspidatum )等(
李效雄等, 2013 ). ...
甘肃大野口青海云杉种群的空间分布格局
2
2010
... 祁连山是中国西部的主要山脉之一, 位于青藏高原、蒙古高原、黄土高原三大高原交汇带, 是石羊河、黑河、疏勒河等6大内陆河和黄河流域的重要水源产流地, 维系着河西乃至西部地区生存与发展的命脉, 在国家生态建设中具有十分重要的战略地位(
Gao et al ., 2013 ;
Zhu et al ., 2017 ).祁连山山地森林的主要建群种——青海云杉(
Picea crassifolia ), 为松科云杉属(
Picea )乔木, 是中国青藏高原东北边缘特有树种, 多以纯林形式存在, 呈带状或斑块状分布于海拔2 300-3 300 m的阴坡和半阴坡, 分布面积占祁连山水源涵养林面积的24.74%, 占乔木林面积的75.72%, 对于维系祁连山的生物多样性、水源涵养功能乃至区域生态平衡都具有至关重要的作用(
田风霞等, 2012 ;
何芳兰等, 2016 ;
张华等, 2016 ).部分****(
赵丽琼等, 2010 ;
张华等, 2016 )虽然已对青海云杉种群空间格局及其关联性进行了研究, 但是已有研究的空间尺度相对较小, 仅能揭示小尺度下的种群空间分布格局, 难以客观地反映青海云杉种群空间分布的特点和种内关系.鉴于此, 本研究基于祁连山大野口流域10.2 hm
2 青海云杉动态监测样地数据, 采用空间点格局分析方法中的成对相关函数对不同径级青海云杉种群的空间分布格局及其关联性进行分析, 旨在阐明: (1)祁连山大野口流域青海云杉种群不同径级个体的空间分布格局及其关联性如何随空间尺度进行变化; (2)不同径级个体的空间分布格局及其关联性形成的潜在生态学过程与影响机制.上述科学问题的合理解答, 能从空间格局的角度客观地揭示青海云杉种群发展规律, 可以为制定该种群的配置、种植与保护管理措施提供指导, 并对祁连山地区的植被恢复与重建、生物多样性保护等具有重要的意义和价值. ...
... 研究样地位于祁连山中段北坡大野口流域青海云杉林内, 地理位置为100.25° E, 38.53° N.样地最低海拔2 856 m, 最高海拔2 962 m, 最大高差106 m, 坡向西北, 坡度13°.根据祁连山水源涵养林研究院的资料, 样地所在的大野口流域气候属于温带大陆性高寒半湿润山地气候, 雨热同期, 冬季寒冷漫长, 夏季短暂, 年平均气温5.4 ℃, 最冷月(1月)平均气温为-12.5 ℃, 最热月(7月)平均气温为19.6 ℃, 年降水量为300-500 mm, 主要集中在6-9月, 年平均相对湿度60% (1994-2004年), 年蒸发量为1 488 mm.样地的成土母质为钙质岩, 土壤类型为山地森林灰褐土, 土壤pH 6.3-7.9, 厚度约1 m.林分起源为天然次生林, 为青海云杉纯林, 林分结构简单, 郁闭度较高, 林下光照不足.青海云杉群落成层现象明显, 可明显划分为乔木层、灌木层、草本层和苔藓层4个层次(
赵丽琼等, 2010 ).其中, 乔木树种组成单一, 除青海云杉外, 只有少量祁连圆柏(
Juniperus przewalskii ); 灌木生长发育不良, 物种较少, 主要有金露梅(
Potentilla fruticosa )、银露梅(
Potentilla glabra )、鬼箭锦鸡儿(
Caragana jubata )、吉拉柳(
Salix gilashanica )、鲜黄小檗(
Berberis diaphana )和高山绣线菊(
Spiraea alpina ); 草本植物物种较丰富, 主要有黑穗薹草(
Carex atrata )、珠芽蓼(
Polygonum viviparum )、针茅(
Stipa capillata )和马蔺(
Iris lactea )等典型高山草甸植物.苔藓层发育良好, 盖度达80%以上, 厚度约10 cm, 主要物种有山羽藓(
Abietinella abietina )、欧灰藓(
Hypnum cupressiforme )和匐灯藓(
Plagiomnium cuspidatum )等(
李效雄等, 2013 ). ...
尕海-则岔地区紫果云杉种群结构与动态特征
1
2018
... 植物种群的径级结构是指不同径级组的个体在种群内所占比例和配置情况(
田润炜等, 2016 ), 是种群的生殖特性与外界环境相互作用的结果, 在一定程度上体现了种群的径级结构特征和生长发育状况, 进而可以预测种群和群落的动态发展趋势(
涂洪润等, 2019 ).本研究发现, 样地内青海云杉种群的幼苗和小树个体储备较为丰富, 其种群径级结构呈倒“J”型分布, 这说明该种群属增长型种群, 种群天然更新能力良好, 只要未来不遭受强烈干扰, 种群数量会保持逐渐增长.这与
李效雄等(2012) 和
张华等(2016) 分别对祁连山大野口流域和天老池小流域的青海云杉种群结构的研究结果一致.这可能是由青海云杉种群的生物学特性与环境因素共同作用的结果.已有研究表明, 森林更新过程中最重要和最敏感的阶段是幼苗阶段(
李晓笑等, 2011 ;
赵阳等, 2018 ), 林内有限的资源和空间往往成为种群更新的重要限制因素(
项小燕等, 2016 ).青海云杉种群的幼苗和小树个体数量多主要是由其生殖特性决定, 其种群可以产生大量高质量种子, 自然萌发形成的幼苗大都集中分布在林窗下, 形成大量小径级个体, 占据有利资源, 这是其种群在群落中持续处于优势地位的主要原因.随着小径级个体的生长, 它们对光照、水分、土壤养分等环境资源和空间需求随之增加, 由于环境资源和空间有限, 种内和种间竞争加剧, 出现强烈的自疏和他疏现象, 竞争能力弱的幼苗和小树在竞争中会大量死亡, 只有少量小径级个体能够成功转化为大径级个体, 造成样地内仅存在少量的中树和大树, 这成为该青海云杉种群更新和发展的“瓶颈” (
张婕等, 2014 ;
白卫国等, 2017 ;
于世川等, 2017 ).因此, 要实现种群数量的稳定增长, 必须加强对种群结构的保护, 尤其要重视对幼苗和小树及其生存环境的保护和改善. ...
尕海-则岔地区紫果云杉种群结构与动态特征
1
2018
... 植物种群的径级结构是指不同径级组的个体在种群内所占比例和配置情况(
田润炜等, 2016 ), 是种群的生殖特性与外界环境相互作用的结果, 在一定程度上体现了种群的径级结构特征和生长发育状况, 进而可以预测种群和群落的动态发展趋势(
涂洪润等, 2019 ).本研究发现, 样地内青海云杉种群的幼苗和小树个体储备较为丰富, 其种群径级结构呈倒“J”型分布, 这说明该种群属增长型种群, 种群天然更新能力良好, 只要未来不遭受强烈干扰, 种群数量会保持逐渐增长.这与
李效雄等(2012) 和
张华等(2016) 分别对祁连山大野口流域和天老池小流域的青海云杉种群结构的研究结果一致.这可能是由青海云杉种群的生物学特性与环境因素共同作用的结果.已有研究表明, 森林更新过程中最重要和最敏感的阶段是幼苗阶段(
李晓笑等, 2011 ;
赵阳等, 2018 ), 林内有限的资源和空间往往成为种群更新的重要限制因素(
项小燕等, 2016 ).青海云杉种群的幼苗和小树个体数量多主要是由其生殖特性决定, 其种群可以产生大量高质量种子, 自然萌发形成的幼苗大都集中分布在林窗下, 形成大量小径级个体, 占据有利资源, 这是其种群在群落中持续处于优势地位的主要原因.随着小径级个体的生长, 它们对光照、水分、土壤养分等环境资源和空间需求随之增加, 由于环境资源和空间有限, 种内和种间竞争加剧, 出现强烈的自疏和他疏现象, 竞争能力弱的幼苗和小树在竞争中会大量死亡, 只有少量小径级个体能够成功转化为大径级个体, 造成样地内仅存在少量的中树和大树, 这成为该青海云杉种群更新和发展的“瓶颈” (
张婕等, 2014 ;
白卫国等, 2017 ;
于世川等, 2017 ).因此, 要实现种群数量的稳定增长, 必须加强对种群结构的保护, 尤其要重视对幼苗和小树及其生存环境的保护和改善. ...
Temporal variability in soil moisture after thinning in semi-arid
Picea crassifolia plantations in northwestern China
1
2017
... 祁连山是中国西部的主要山脉之一, 位于青藏高原、蒙古高原、黄土高原三大高原交汇带, 是石羊河、黑河、疏勒河等6大内陆河和黄河流域的重要水源产流地, 维系着河西乃至西部地区生存与发展的命脉, 在国家生态建设中具有十分重要的战略地位(
Gao et al ., 2013 ;
Zhu et al ., 2017 ).祁连山山地森林的主要建群种——青海云杉(
Picea crassifolia ), 为松科云杉属(
Picea )乔木, 是中国青藏高原东北边缘特有树种, 多以纯林形式存在, 呈带状或斑块状分布于海拔2 300-3 300 m的阴坡和半阴坡, 分布面积占祁连山水源涵养林面积的24.74%, 占乔木林面积的75.72%, 对于维系祁连山的生物多样性、水源涵养功能乃至区域生态平衡都具有至关重要的作用(
田风霞等, 2012 ;
何芳兰等, 2016 ;
张华等, 2016 ).部分****(
赵丽琼等, 2010 ;
张华等, 2016 )虽然已对青海云杉种群空间格局及其关联性进行了研究, 但是已有研究的空间尺度相对较小, 仅能揭示小尺度下的种群空间分布格局, 难以客观地反映青海云杉种群空间分布的特点和种内关系.鉴于此, 本研究基于祁连山大野口流域10.2 hm
2 青海云杉动态监测样地数据, 采用空间点格局分析方法中的成对相关函数对不同径级青海云杉种群的空间分布格局及其关联性进行分析, 旨在阐明: (1)祁连山大野口流域青海云杉种群不同径级个体的空间分布格局及其关联性如何随空间尺度进行变化; (2)不同径级个体的空间分布格局及其关联性形成的潜在生态学过程与影响机制.上述科学问题的合理解答, 能从空间格局的角度客观地揭示青海云杉种群发展规律, 可以为制定该种群的配置、种植与保护管理措施提供指导, 并对祁连山地区的植被恢复与重建、生物多样性保护等具有重要的意义和价值. ...
北京暖温带次生林种群分布格局与种间空间关联性
1
2011
... 通常认为, 由于物种的生态学和生物学特性、种子扩散限制和生境异质性等机制的影响, 较小径级的物种常呈聚集分布, 尤其在小尺度上更加明显(
祝燕等, 2011 ).而随着径级增大, 种内和种间为争夺有限资源和空间的竞争也大大增强, 常呈随机或规则分布, 聚集强度也随径级的增加而降低(
Condit et al ., 2000 ).本研究结果也符合这一规律.样地内的青海云杉种群的幼树、小树和中树这类低龄级个体多呈聚集分布, 这一方面可能是由于青海云杉的种子具有种翅, 主要依靠风力传播, 使得大多数种子散落在母树周围一定范围内且随着距离的增大其种子传播的数量逐渐下降, 这些生物学特性使其形成聚集分布的空间分布特征.实地调查过程中也发现, 青海云杉幼苗大都集中分布于林窗下, 在阴蔽的林冠下, 出现幼苗和小树死亡现象, 在地被层特别是苔藓层较厚的地方, 幼苗较少.另一方面, 低龄级种群个体较小, 处在群落的下层, 对光、热、水和土壤养分等生态因子的竞争能力较弱, 聚集可以提高其竞争能力以及抵御外界不良环境, 提高成活率.此外, 小生境的异质作用导致幼树与小树分布不均匀, 如局部地形、土壤和植被等环境因子的差异, 以及采伐、自然灾害等形成的林窗, 都为低龄级个体的大量繁殖和生长创造了良好的条件(
张华等, 2016 ).随着种群年龄的增长, 个体对光照、水分、营养等资源和空间的需求不断增加, 种内和种间竞争加剧而出现强烈的自疏和他疏现象, 导致大量林木死亡, 种群密度下降, 聚集强度逐渐减弱, 种群由聚集分布向随机分布或规则分布转变.此外, 大树主要呈随机分布, 这可能是因为大树位于林冠层, 高度较高, 树冠基本定型, 根系也伸展到一定程度, 种内竞争加剧, 自疏效应使得种群的聚集分布格局被打破, 由聚集分布向随机分布转变. ...
北京暖温带次生林种群分布格局与种间空间关联性
1
2011
... 通常认为, 由于物种的生态学和生物学特性、种子扩散限制和生境异质性等机制的影响, 较小径级的物种常呈聚集分布, 尤其在小尺度上更加明显(
祝燕等, 2011 ).而随着径级增大, 种内和种间为争夺有限资源和空间的竞争也大大增强, 常呈随机或规则分布, 聚集强度也随径级的增加而降低(
Condit et al ., 2000 ).本研究结果也符合这一规律.样地内的青海云杉种群的幼树、小树和中树这类低龄级个体多呈聚集分布, 这一方面可能是由于青海云杉的种子具有种翅, 主要依靠风力传播, 使得大多数种子散落在母树周围一定范围内且随着距离的增大其种子传播的数量逐渐下降, 这些生物学特性使其形成聚集分布的空间分布特征.实地调查过程中也发现, 青海云杉幼苗大都集中分布于林窗下, 在阴蔽的林冠下, 出现幼苗和小树死亡现象, 在地被层特别是苔藓层较厚的地方, 幼苗较少.另一方面, 低龄级种群个体较小, 处在群落的下层, 对光、热、水和土壤养分等生态因子的竞争能力较弱, 聚集可以提高其竞争能力以及抵御外界不良环境, 提高成活率.此外, 小生境的异质作用导致幼树与小树分布不均匀, 如局部地形、土壤和植被等环境因子的差异, 以及采伐、自然灾害等形成的林窗, 都为低龄级个体的大量繁殖和生长创造了良好的条件(
张华等, 2016 ).随着种群年龄的增长, 个体对光照、水分、营养等资源和空间的需求不断增加, 种内和种间竞争加剧而出现强烈的自疏和他疏现象, 导致大量林木死亡, 种群密度下降, 聚集强度逐渐减弱, 种群由聚集分布向随机分布或规则分布转变.此外, 大树主要呈随机分布, 这可能是因为大树位于林冠层, 高度较高, 树冠基本定型, 根系也伸展到一定程度, 种内竞争加剧, 自疏效应使得种群的聚集分布格局被打破, 由聚集分布向随机分布转变. ...
 ,1,2,*, 刘润红3, 赵维俊2, 敬文茂2, 马剑2, 武秀荣2, 赵晶忠2, 马雪娥2
,1,2,*, 刘润红3, 赵维俊2, 敬文茂2, 马剑2, 武秀荣2, 赵晶忠2, 马雪娥2
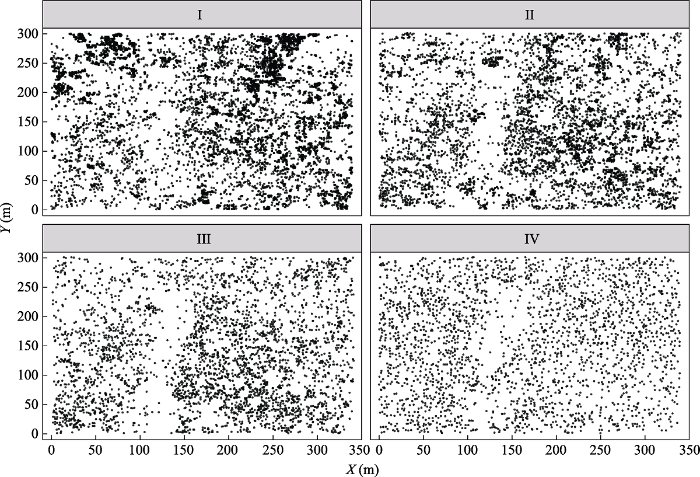 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT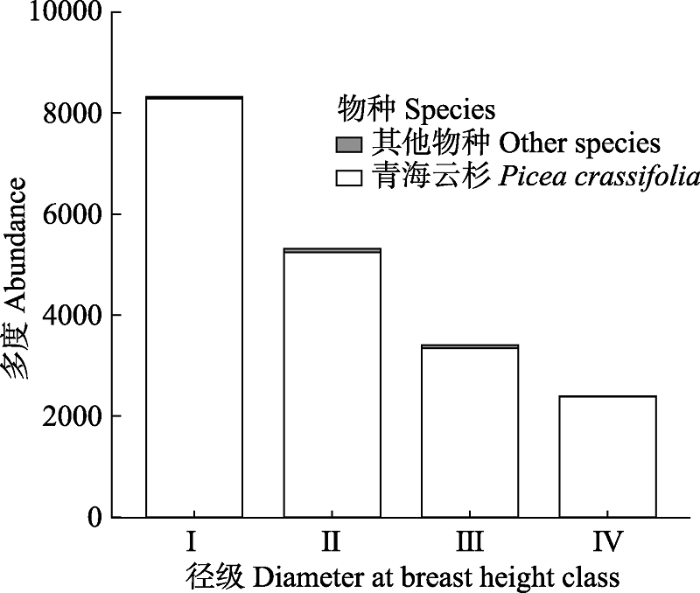 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT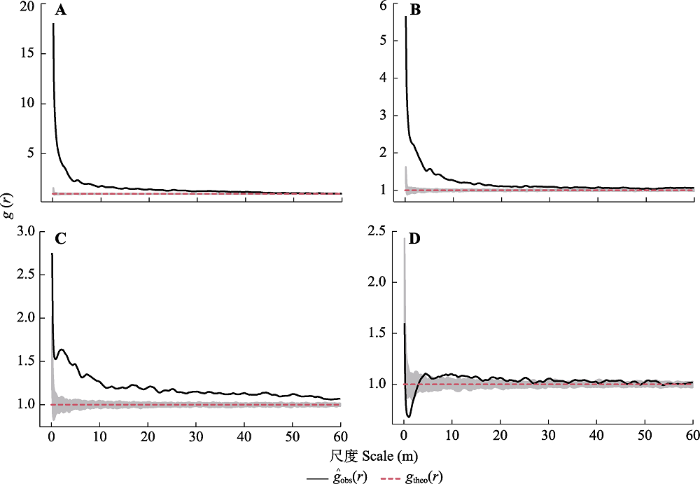 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT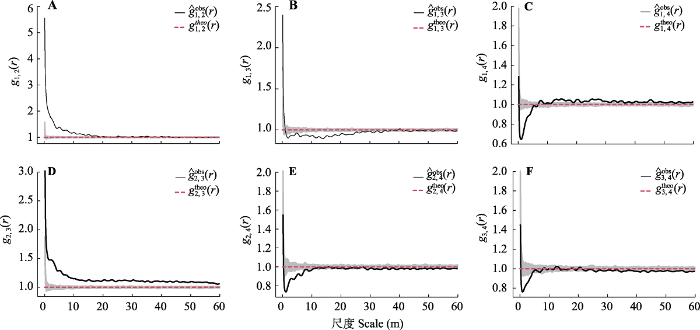 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}