Effect of altered litter input and nitrogen addition on ecosystem aboveground primary productivity and plant functional group composition in a semiarid grassland
GAMADAERJI1,2, Ze YANG1,2, Xing-Ru TAN1,2, Shan-Shan WANG1,2, Wei-Jing LI1,2, Cui-Hai YOU1,2, Yan-Bing WANG1,2, Bing-Wei ZHANG1,3, Ting-Ting REN1,2, Shi-Ping CHEN,1,2,*1State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 2University of Chinese Academy of Sciences, Beijing 100049, China 3State Key Laboratory of Biocontrol, School of Life Sciences, Sun Yat-sen University, Guangzhou 510275, China
National Natural Science Foundation of China(41773084) National Key R&D Program of China(2017YFA0604801) National Key R&D Program of China(2016YFC0500103)
Abstract Aims Litter is the major input source of soil organic carbon and nutrients in natural ecosystems and considered as a key link between above- and belowground carbon cycles. Changes in litter input amount have been proven to exert significant impacts on plant productivity, community structure, and therefore ecosystem function. In Nei Mongol semiarid grasslands, different grassland management practices such as grazing, clipping, and fencing have caused dramatic changes in litter production and input. In addition, as a nitrogen-limited ecosystem, Nei Mongol semiarid grasslands also experienced increasing nitrogen deposition. However, how do changes in litter input and nitrogen addition impact the community productivity and composition of plant functional groups are still unclear in the semiarid grasslands. In this study, our objectives are: 1) to investigate the effects of altered litter input and nitrogen addition on community productivity; 2) to study the changes in aboveground biomass of different plant functional groups and their contribution to community productivity under different litter input and N addition treatments. Methods We established a manipulative experiment with altered litter input and nitrogen addition treatments in a semiarid typical grassland in West Ujimqin Banner, Nei Mongol. A randomized block split-plot design was applied with five blocks. Three litter input treatments, including litter removal (C0), control (C1) and litter addition (C2), were assigned randomly in each block. Each plot (6 m × 7 m) of litter input treatment was separated into two subplots. One of the subplots was assigned as the N addition treatment (N1) and another subplot was considered as the control treatment without N addition (N0). In N addition treatment, 15 g N·m-2·a-1 N fertilizer (as NH4NO3) was applied every year since 2013. Aboveground net primary productivity (ANPP) in community and plant functional group levels of each treatment were determined during the peak season from 2013 to 2018. Important findings Based on 6-year measurements, we found the following results. 1) Litter input increase and nitrogen addition increased community ANPP. Compared with the control, litter removal treatment significantly decreased ANPP by 8.4% and 7.6% in plots without and with N addition, respectively. Litter addition increased ANPP by 10.7% and 6.3% in plots without and with N addition, respectively. 2) Different responses of plant functional groups to altered litter input and N addition led to a change in plant functional group composition. The biomass of perennial bunch grass (PB) and perennial rhizome grass (PR) increased significantly with the increment of litter and nitrogen, which enhanced their dominant status in the community. 3) Improved soil water condition by litter input and nutrient supply by N addition are the major pathways that enhanced ANPP and changed the functional group composition. These results show that proper management, such as grazing exclusion and reducing grazing intensity, can promote productivity by increasing inputs of litter in semiarid grasslands, which leads to the maintenance of ecosystem stability. Suitable nutrients management, like nitrogen addition, is also helpful for productivity improvement and the recovery of degraded grasslands. Keywords:aboveground net primary productivity (ANPP);plant functional group;altered litter input;nitrogen addition;semiarid grassland
PDF (1702KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 嘎玛达尔基, 杨泽, 谭星儒, 王珊珊, 李伟晶, 游翠海, 王彦兵, 张兵伟, 任婷婷, 陈世苹. 凋落物输入变化和氮添加对半干旱草原群落生产力及功能群组成的影响. 植物生态学报, 2020, 44(8): 791-806. DOI: 10.17521/cjpe.2020.0126 GAMADAERJI , YANG Ze, TAN Xing-Ru, WANG Shan-Shan, LI Wei-Jing, YOU Cui-Hai, WANG Yan-Bing, ZHANG Bing-Wei, REN Ting-Ting, CHEN Shi-Ping. Effect of altered litter input and nitrogen addition on ecosystem aboveground primary productivity and plant functional group composition in a semiarid grassland. Chinese Journal of Plant Ecology, 2020, 44(8): 791-806. DOI: 10.17521/cjpe.2020.0126
除了实验处理开始当年(2012年)降水量达到406.3 mm, 为长期平均降水量的140%以上, 其余年份(2013-2018年)均接近或低于长期降水量平均值, 特别是2016和2017年, 其生长季降水量仅为224.1和226.1 mm, 相当于多年平均值的77%和78%, 是较为干旱的年份(图1)。
Fig. 1Changes in growing season precipitation and daily mean air temperature during the studied period (2012-2018) and the longterm mean growing season precipitation during 1955-2011.
新窗口打开|下载原图ZIP|生成PPT 图22013-2018年不同凋落物输入变化和氮添加处理下, 表层(0-10 cm)土壤温度(Ts)、土壤含水量(SWC)和土壤可溶性有机碳氮(DOC和DON)含量的年际变异(A、C、E、G)和6年平均值(B、D、F、H)(平均值±标准误差)。图中给出了凋落物输入变化(C)和氮添加(N)对生产力影响的差异显著性分析结果(ns, p > 0.1; #, p < 0.1; *, p < 0.05; **, p < 0.01; ***, p < 0.001)。柱状图中的字母表示不同凋落物输入变化和氮添加处理间各指标的多重比较结果, 不同小写字母表示不同凋落物输入变化处理间的差异显著(p < 0.05), 不同大写字母代表不同氮添加处理间的差异显著(p < 0.05)。C0, 凋落物去除; C1, 凋落物对照; C2, 凋落物添加; N0, 无氮添加; N1, 氮添加。
Fig. 2Inter-annual variations (A, C, E, G) and the 6-year average (B, D, F, H) of soil temperature (Ts), soil water content (SWC), dissolved organic carbon (DOC) and nitrogen (DON) content in surface soil layer (0-10 cm) in different litter input and nitrogen addition treatments during 2013-2018 (mean ± SE). Significance levels were presented to show the effects of altered litter input (C) and nitrogen addition (N) treatments and their interaction (C × N) on these parameters (ns, p > 0. 1; #, p < 0.1; *, p < 0.05; **, p < 0.01; ***, p < 0.001). The letters in the bar graph indicate the results of multiple comparisons among different litter input and nitrogen addition treatments. Different lowercase letters indicate significant difference among three altered litter input treatments (p < 0.05), and uppercase letters indicate the significant difference between two nitrogen treatments (p < 0.05). C0, litter removal; C1, control; C2, litter addition; N0, without nitrogen addition; N1, nitrogen addition.
新窗口打开|下载原图ZIP|生成PPT 图32013-2018年不同凋落物输入变化和氮添加处理下, 群落地上净初级生产力(ANPP)的年际变异(A)和6年平均值(B)。图中数据均为平均值±标准误差。图中给出了凋落物输入变化(C)和氮添加(N)对生产力影响的差异显著性分析结果(ns, p > 0.1; #, p < 0.1; *, p < 0.05; **, p < 0.01; ***, p < 0.001)。图中不同大写字母表示氮添加对生产力影响的差异显著(p < 0.05)。C0, 凋落物去除; C1, 凋落物对照; C2, 凋落物添加; N0, 无氮添加; N1, 氮添加。
Fig. 3Inter-annual variation (A) and the 6-year average (B) of aboveground net primary productivity (ANPP) in different litter input and nitrogen addition treatments during 2013-2018. The data in the figure are mean ± SE. Significance levels were presented to show the effects of altered litter input (C) and nitrogen addition (N) treatments and their interaction (C × N) on ANPP (ns, p > 0.1; #, p < 0.1; *, p < 0.05; **, p < 0.01; ***, p < 0.001). Different uppercase letters in the bar graph indicate significant difference results of multiple comparisons between two nitrogen treatments (p < 0.05). C0, litter removal; C1, control; C2, litter addition; N0, without nitrogen addition; N1, nitrogen addition.
Table 3 表3 表3凋落物输入变化(C)和氮添加(N)对群落地上净初级生产力(ANPP)及各功能群绝对和相对生物量影响的重复测量方差分析(RANOVA)结果 Table 3Effects of altered litter input (C) and nitrogen addition (N) on community above-ground net primary productivity (ANPP) and absolute and relative biomass of each functional group evaluated using repeated measures analysis of variance (RANOVA)
df
地上净初级生产力 ANPP
绝对生物量 Absolute biomass
相对生物量 Relative biomass
PB
PR
PF
SS
AB
PB%
PR%
PF%
SS%
AB%
区组Block
4
1.72
1.60
0.66
1.17
1.39
1.16
0.87
0.31
1.11
1.94
0.42
C
2
3.20#
0.77
3.30#
0.01
0.66
3.13#
0.18
1.65
0.36
0.70
3.70*
N
1
62.87***
31.22***
7.60*
1.45
0.17
42.97***
9.04**
0.24
12.50**
2.81
37.54***
年Year (Y)
5
57.28***
15.66***
22.04***
17.54***
1.51
4.32**
4.87***
8.73***
6.41***
1.50
4.32**
C × N
2
0.02
0.02
0.02
0.08
1.33
2.65#
0.08
0.18
0.23
1.21
2.96#
C × Y
10
0.59
1.31
6.36***
0.76
0.68
1.85#
2.78**
5.03***
0.86
0.44
1.78#
N × Y
5
7.64***
8.05***
2.63*
5.27***
2.46*
3.91**
8.59***
0.30
10.38***
2.72*
3.67**
C × N × Y
10
0.84
0.95
1.63
1.22
0.65
1.81#
0.39
2.26*
1.46
0.84
1.70#
#, p < 0.1, *, p < 0.05, **, p < 0.01, ***, p < 0.001。AB, 一二年生植物; PB, 多年生丛生禾草; PF, 多年生杂类草; PR, 多年生根茎禾草; SS, 灌木与半灌木。 #, p < 0.1, *, p < 0.05, **, p < 0.01, ***, p < 0.001. AB, perennial plants; PB, perennial bunchgrass; PF, perennial forbs; PR, perennial rhizome grass; SS, shrubs, and semi-shrubs.
新窗口打开|下载原图ZIP|生成PPT 图42013-2018年不同凋落物输入变化和氮添加处理下, 各功能群地上净初级生产力(ANPP)年际变异(A、C、E、G、I)和6年平均值(B、D、F、H、J)。图中数据均为平均值±标准误差。图中给出了每个年份以及多年平均凋落物输入变化(C)氮添加(N)及其交互作用(C × N)对各功能群地上净初级生产力影响的差异显著性分析结果(ns, p > 0.1; #, p < 0.1; *, p < 0.05; **, p < 0.01; ***, p < 0.001)。不同大写字母表示不同氮添加处理(N)对各功能群地上净初级生产力影响的多重比较结果差异显著(p < 0.05)。C0, 凋落物去除; C1, 凋落物对照; C2, 凋落物添加; N0, 无氮添加; N1, 氮添加。AB, 一二年生植物; PB, 多年生丛生禾草; PF, 多年生杂类草; PR, 多年生根茎禾草; SS, 灌木与半灌木。
Fig. 4Inter-annual variations (A, C, E, G, I) and 6-year average (B, D, F, H, J) of aboveground net primary productivity (ANPP) of five functional groups in different litter input and nitrogen addition treatments during 2013-2018. The data in the figure are mean ± SE. Significance levels were presented to show the effects of altered litter input (C) and nitrogen addition (N) treatments and their interaction (C × N) on ANPP of each functional group (ns, p > 0.1; #, p < 0.1; *, p < 0.05; **, p < 0.01; ***, p < 0.001). C0, litter removal; C1, control; C2, litter addition; N0, without nitrogen addition; N1, nitrogen addition. Different uppercase letters in the bar graph indicate significant difference of multiple comparisons between two nitrogen addition treatments. AB, annual and biannual plants; PB, perennial bunch grasses; PF, perennial forbs; PR, perennial rhizome grasses; SS, shrubs and semi-shrubs.
Fig. 5Variations in the relative biomass of the five functional groups in different litter input and nitrogen addition treatments. The data in the figure are mean ± SE. Asterisk indicated significant difference between different nitrogen treatments (**, p < 0.01; ***, p < 0.001). C0, litter removal; C1, control; and C2, litter addition; N0, nitrogen-free control; N1, nitrogen addition. AB, annual and biannual plants; PB, perennial bunch grasses; PF, perennial forbs; PR, perennial rhizome grasses; SS, shrubs, and semi-shrubs.
新窗口打开|下载原图ZIP|生成PPT 图6不同氮处理条件下, 地上净初级生产力(ANPP)、土壤温度(Ts)、土壤含水量(SWC)及其季节变异(CVSWC)与上一年添加的凋落物质量间的相关关系。图中标出相关系数(R2)和差异显著性分析结果(*, p < 0.05; **, p < 0.01; ***, p < 0.001)。N0, 无氮添加; N1, 氮添加。
Fig. 6Correlation analysis of aboveground net primary productivity (ANPP), soil temperature (Ts), soil water content (SWC) and its variation (CVSWC) with litter biomass added in the previous year under each nitrogen treatment. The correlation coefficient (R2) and significance levels are presented in the figure (*, p < 0.05; **, p < 0.01; ***, p < 0.001). N0, nitrogen-free control; N1, nitrogen addition.
新窗口打开|下载原图ZIP|生成PPT 图7群落地上净初级生产力与生长季降水量(A)、土壤温度(B)、土壤含水量(C)和土壤含水量季节变异(D)之间的关系。图中标出相关系数(R2)和差异显著性分析结果(ns, p > 0.05; *, p < 0.05; **, p < 0.01; ***, p < 0.001)。C0, 凋落物去除; C1, 凋落物对照; C2, 凋落物添加; N0, 无氮添加; N1, 氮添加。
Fig. 7Correlation analysis between aboveground net primary productivity (ANPP) and precipitation (A), soil temperature (Ts)(B), soil water content (SWC)(C), and seasonal variation of soil water content (CVSWC)(D) during the growing season. The correlation coefficient (R2) and the significance levels are presented in the figure (ns, p > 0.05; *, p < 0.05; **, p < 0.01; ***, p < 0.001). C0, litter removal; C1, control; C2, litter addition; N0, without nitrogen addition; N1, nitrogen addition.
Fig. 8Structural equation model (SEM) on how litter manipulation and nitrogen addition affect the community functional group composition and aboveground net primary productivity (ANPP) through soil environment. CVSWC, variance of soil water content; DOM, soil dissolved organic matter (the first principal component of DOC and DON, which explained 0.88 of them); PB, perennial bunch grasses; PF, perennial forbs; PR, perennial rhizome grasses; SWC, soil water content; Ts, soil temperature. AIC, Akaike Information Criterion; BIC, Bayesian Information Criterion; Fisher’s Chi, result of Fisher exact test.
3 讨论
3.1 凋落物输入变化和氮添加对ANPP的影响
连续6年的实验处理结果表明, 凋落物输入量的增加显著提高了群落ANPP, 而凋落物去除则降低了ANPP (图3B)。相关分析表明随着凋落物添加量的增加, 群落ANPP显著提高(图6A)。已有研究表明凋落物对ANPP的影响主要分为两个方面, 一方面是通过凋落物自身分解直接增加生态系统碳输入, 提高土壤养分可利用性来促进ANPP (Brearley et al., 2003; Bansal et al., 2014); 另一方面是通过对微环境条件(如土壤温度、水分状况等)的调控来间接影响ANPP (Booth et al., 2005; Deutsch et al., 2010a)。在本研究中, 凋落物输入变化并没有显著改变DOC和DON等土壤养分含量, 而是显著改善了土壤水分和温度等土壤环境条件间接影响植被生产力。ANPP与降水量以及土壤含水量的关系表明, 水分是限制群落生产力的主要限制因子, 凋落物添加通过降低土温, 减少水分蒸发, 提高表层土壤水分可利用性, 降低土壤水分的季节变异, 从而降低了植物遭受干旱胁迫风险, 进而促进植物生长, 提高群落ANPP。Xu等(2013a)在关于生态系统中凋落物作用的meta分析中发现, 在草原生态系统的研究中, 凋落物添加会增加土壤微生物碳以及总碳含量, 并降低土壤温度、增加土壤含水量。我们的研究虽然在统计上尚未检测到凋落物输入变化对土壤DOC和DON含量的显著影响, 但还是发现随着凋落物的增加, DOC含量呈现出增加的趋势。这表明凋落物输入变化对土壤DOC等养分可利用性的影响可能需要更长期的实验研究结果来检验(Wang et al., 2017a)。尤其在干旱半干旱生态系统中, 因为受到水分条件的限制, 凋落物分解速率相对更慢(Zhang & Wang, 2015)。
?grenGI, WetterstedtJ?M, BillbergerMFK (2012). Nutrient limitation on terrestrial plant growth-modeling the interaction between nitrogen and phosphorus New Phytologist, 194, 953-960. DOI:10.1111/j.1469-8137.2012.04116.xURL [本文引用: 1] We use a simple model of plant growth and nutrient uptake to explore the consequences for the plants relative growth rate of letting plants invest differentially in N and P uptake.We find a smooth transition between limiting elements, in contrast to the strict transition in Liebigs law of the minimum. At N : P supply ratios where the two elements simultaneously limit growth, an increase in either of the nutrients will increase the growth rate because more resources can be allocated towards the limiting element, as suggested by the multiple limitation hypothesis. However, the further the supply ratio deviates from these supply rates, the more the plants will follow the law of the minimum.Liebigs law of the minimum will in many cases be a useful first-order approximation.]]>
AmatangeloKL, DukesJS, FieldCB (2008). Responses of a California annual grassland to litter manipulation Journal of Vegetation Science, 19, 605-612. DOI:10.3170/2008-8-18415URL [本文引用: 2]
AustinAT, VivancoL (2006). Plant litter decomposition in a semi-arid ecosystem controlled by photodegradation Nature, 442, 555-558. DOI:10.1038/nature05038URLPMID:16885982 [本文引用: 1] The carbon balance in terrestrial ecosystems is determined by the difference between inputs from primary production and the return of carbon to the atmosphere through decomposition of organic matter. Our understanding of the factors that control carbon turnover in water-limited ecosystems is limited, however, as studies of litter decomposition have shown contradictory results and only a modest correlation with precipitation. Here we evaluate the influence of solar radiation, soil biotic activity and soil resource availability on litter decomposition in the semi-arid Patagonian steppe using the results of manipulative experiments carried out under ambient conditions of rainfall and temperature. We show that intercepted solar radiation was the only factor that had a significant effect on the decomposition of organic matter, with attenuation of ultraviolet-B and total radiation causing a 33 and 60 per cent reduction in decomposition, respectively. We conclude that photodegradation is a dominant control on above-ground litter decomposition in this semi-arid ecosystem. Losses through photochemical mineralization may represent a short-circuit in the carbon cycle, with a substantial fraction of carbon fixed in plant biomass being lost directly to the atmosphere without cycling through soil organic matter pools. Furthermore, future changes in radiation interception due to decreased cloudiness, increased stratospheric ozone depletion, or reduced vegetative cover may have a more significant effect on the carbon balance in these water-limited ecosystems than changes in temperature or precipitation.
BaiYF, HanXG, WuJG, ChenZZ, LiLH (2004). Ecosystem stability and compensatory effects in the Inner Mongolia grassland Nature, 431, 181-184. DOI:10.1038/nature02850URLPMID:15356630 [本文引用: 2] Numerous studies have suggested that biodiversity reduces variability in ecosystem productivity through compensatory effects; that is, a species increases in its abundance in response to the reduction of another in a fluctuating environment. But this view has been challenged on several grounds. Because most studies have been based on artificially constructed grasslands with short duration, long-term studies of natural ecosystems are needed. On the basis of a 24-year study of the Inner Mongolia grassland, here we present three key findings. First, that January-July precipitation is the primary climatic factor causing fluctuations in community biomass production; second, that ecosystem stability (conversely related to variability in community biomass production) increases progressively along the hierarchy of organizational levels (that is, from species to functional group to whole community); and finally, that the community-level stability seems to arise from compensatory interactions among major components at both species and functional group levels. From a hierarchical perspective, our results corroborate some previous findings of compensatory effects. Undisturbed mature steppe ecosystems seem to culminate with high biodiversity, productivity and ecosystem stability concurrently. Because these relationships are correlational, further studies are necessary to verify the causation among these factors. Our study provides new insights for better management and restoration of the rapidly degrading Inner Mongolia grassland.
BaiYF, PanQM, XingQ (2016). Fundamental theories and technologies for optimizing the production functions and ecological functions in grassland ecosystems Chinese Science Bulletin, 61, 201-212. [本文引用: 1]
BansalS, SheleyRL, BlankB, VasquezEA (2014). Plant litter effects on soil nutrient availability and vegetation dynamics: changes that occur when annual grasses invade shrub- steppe communities Plant Ecology, 215, 367-378. DOI:10.1007/s11258-014-0307-1URL [本文引用: 2] Changes in the quantity and quality of plant litter occur in many ecosystems as they are invaded by exotic species, which impact soil nutrient cycling and plant community composition. Such changes in sagebrush-steppe communities are occurring with invasion of annual grasses (AG) into a perennial grass (PG) dominated system. We conducted a 5-year litter manipulation study located in the northern Great Basin, USA. Springtime litter was partially or completely removed in three communities with differing levels of invasion (invaded, mixed, and native) to determine how litter removal and litter biomass affected plant-available soil N and plant community composition. Litter biomass (prior to the removal treatment) was negatively correlated with plant-available N in the invaded community, but was positively correlated in the native community. Plant-available N had greater intra- and inter-annual fluctuations in the invaded compared to the mixed or native communities, but was not generally affected by removal treatments. Litter removal had negative effects on AG cover during a warm/dry year and negative effects on PG cover during a cool/wet year in the mixed community. Overall, the effectiveness of springtime litter manipulations on plant-available N were limited and weather dependent, and only removal treatments > 75 % had effects on the plant community. Our study demonstrates how communities invaded by AGs have significantly increased temporal variability in nutrient cycling, which may decrease ecosystem stability. Further, we found that the ecological impacts from litter manipulation on sagebrush communities were dependent on the extent of AG invasion, the timing of removal, and seasonal precipitation.
BoothMS, StarkJM, RastetterE (2005). Controls on nitrogen cycling in terrestrial ecosystems: a synthetic analysis of literature data Ecological Monographs, 75, 139-157. DOI:10.1890/04-0988URL [本文引用: 1]
BosyJL, ReaderRJ (1995). Mechanisms underlying the suppression of forb seedling emergence by grass (Poa pratensis) litter Functional Ecology, 9, 635-639. DOI:10.2307/2390155URL [本文引用: 1]
BrearleyFQ, PressMC, ScholesJD (2003). Nutrients obtained from leaf litter can improve the growth of dipterocarp seedlings New Phytologist, 160, 101-110. [本文引用: 2]
CarlyleCN, FraserLH, TurkingtonR (2014). Response of grassland biomass production to simulated climate change and clipping along an elevation gradient Oecologia, 174, 1065-1073. DOI:10.1007/s00442-013-2833-2URL [本文引用: 1] Changes in rainfall and temperature regimes are altering plant productivity in grasslands worldwide, and these climate change factors are likely to interact with grassland disturbances, particularly grazing. Understanding how plant production responds to both climate change and defoliation, and how this response varies among grassland types, is important for the long-term sustainability of grasslands. For 4 years, we manipulated temperature [ambient and increased using open-top chambers (OTC)], water (ambient, reduced using rainout shelters and increased using hand watering) and defoliation (clipped, and unclipped) in three grassland types along an elevation gradient. We monitored plant cover and biomass and found that OTC reduced biomass by 15 %, but clipping and water treatments interacted with each other and their effects varied in different grassland types. For example, total biomass did not decline in the higher elevation grasslands due to clipping, and water addition mitigated the effects of clipping on subordinate grasses in the lower grasslands. The response of total biomass was driven by dominant plant species while subordinate grasses and forbs showed more variable responses. Overall, our results demonstrate that biomass in the highest elevation grassland was least effected by the treatments and the response of biomass tended to be dependent on interactions between climate change treatments and defoliation. Together, the results suggest that ecosystem function of these grasslands under altered climate patterns will be dependent on site-specific management.
ClarkCM, TilmanD (2008). Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands Nature, 451, 712-715. DOI:10.1038/nature06503URLPMID:18256670 [本文引用: 1] Rates of atmospheric deposition of biologically active nitrogen (N) are two to seven times the pre-industrial rates in many developed nations because of combustion of fossil fuels and agricultural fertilization. They are expected to increase similarly over the next 50 years in industrializing nations of Asia and South America. Although the environmental impacts of high rates of nitrogen addition have been well studied, this is not so for the lower, chronic rates that characterize much of the globe. Here we present results of the first multi-decadal experiment to examine the impacts of chronic, experimental nitrogen addition as low as 10 kg N ha(-1) yr(-1) above ambient atmospheric nitrogen deposition (6 kg N ha(-1) yr(-1) at our site). This total input rate is comparable to terrestrial nitrogen deposition in many industrialized nations. We found that this chronic low-level nitrogen addition rate reduced plant species numbers by 17% relative to controls receiving ambient N deposition. Moreover, species numbers were reduced more per unit of added nitrogen at lower addition rates, suggesting that chronic but low-level nitrogen deposition may have a greater impact on diversity than previously thought. A second experiment showed that a decade after cessation of nitrogen addition, relative plant species number, although not species abundances, had recovered, demonstrating that some effects of nitrogen addition are reversible.
ClarkCM, TilmanD (2010). Recovery of plant diversity following N cessation: effects of recruitment, litter, and elevated N cycling Ecology, 91, 3620-3630. DOI:10.1890/09-1268.1URLPMID:21302833 [本文引用: 1] Plant species richness has declined and composition shifted in response to elevated atmospheric deposition of biologically active nitrogen over much of the industrialized world. Litter thickness, litter nitrogen (N) content, and soil N mineralization rates often remain elevated long after inputs cease, clouding the prospects that plant community diversity and composition would recover should N inputs be reduced. Here we determined how N cycling, litter accumulation, and recruitment limitation influenced community recovery following cessation of long-term N inputs to prairie-like grasslands. We alleviated each of these potential inhibitors through a two-year full-factorial experiment involving organic carbon addition, litter removal, and seed addition. Seed addition had the largest effect on increasing seedling and species numbers and may be necessary to overcome long-term burial of seeds of target perennial grassland species. Litter removal increased light availability and bare sites for colonization, though it had little effect on reducing the biomass of competing neighbors or altering extractable soil N. Nonetheless, these positive influences were enough to lead to small increases in species richness within one year. We found that, although C addition quickly altered many factors assumed favorable for the target community (decreased N availability and biomass of nearby competitors, increased light and site availability), these changes were insufficient to positively impact species richness or seedling numbers over the experimental duration. However, only carbon addition had species-specific effects on the existing plant community, suggesting that its apparent limited utility may be more a result of slow recovery under ambient recruitment rather than from a lack of a restorative effect. There were dramatic interactions among treatments, with the positive effects of litter removal largely negated by carbon addition, and the positive effects of seed addition generally amplified by litter removal. It remains unclear whether each mechanism explored here will induce community recovery, but over different temporal scales. Long-term monitoring will help resolve these remaining questions. Regardless, our results suggest that reversal of species loss and compositional shifts from N deposition in prairies may be more inhibited by habitat fragmentation, recruitment limitation, and long-term suppression of fire than from continued effects of elevated N.
DengMF, LiuLL, JiangL, LiuWX, WangX, LiSP, YangS, WangB (2018). Ecosystem scale trade-off in nitrogen acquisition pathways Nature Ecology & Evolution, 2, 1724-1734. DOI:10.1038/s41559-018-0677-1URLPMID:30250156 [本文引用: 1] The nitrogen (N) cycle in terrestrial ecosystems is strongly influenced by resorption before litter fall and by mineralization after litter fall. Although both resorption and mineralization make N available to plants and are influenced by climate, their linkage in a changing environment remains largely unknown. Here, our synthesis study shows that, at the global scale, increasing N-resorption efficiency negatively affects the N-mineralization rate. As temperature and precipitation increase, the increasing rates of N cycling closely correspond to a shift from the more conservative resorption pathway to the mineralization pathway. Furthermore, ecosystems with faster N-cycle rates support plant species that have higher foliar N:P ratios and microbial communities with lower fungi:bacteria ratios. Our study shows an ecosystem scale trade-off in N-acquisition pathways. We propose that incorporating the dynamic interaction between N resorption and N mineralization into Earth system models will improve the simulation of nutrient constraints on ecosystem productivity.
DeutschES, BorkEW, WillmsWD (2010a). Separation of grassland litter and ecosite influences on seasonal soil moisture and plant growth dynamics Plant Ecology, 209, 135-145. DOI:10.1007/s11258-010-9729-6URL [本文引用: 1] Festuca hallii) grassland. We also examined the relationship between litter and plant biomass throughout the growing season, including linkages between litter, plant growth, and the effects of litter on microclimate. During May, only ecosite was found to be associated with soil moisture, with a similar finding for plant biomass. Litter became important in maintaining greater soil moisture in June and July, however, likely through its corresponding negative impact on soil temperature and associated evaporation. In general, litter had a stronger and more consistent influence on soil moisture than ecosite. Finally, litter had a positive relationship with above-ground biomass, but only during June and July, the same months when litter exhibited the strongest relationship with soil moisture. Litter therefore appears to promote mid-season plant growth in these temperate grasslands, presumably through its ability to reduce evaporation and maintain greater soil water during seasonal moisture limitations.]]>
DeutschES, BorkEW, WillmsWD (2010b). Soil moisture and plant growth responses to litter and defoliation impacts in Parkland grasslands Agriculture Ecosystems & Environment, 135, 1-9. [本文引用: 2]
FacelliJM, PickettSTA (1991a). Plant litter: light interception and effects on an old-field plant community Ecology, 72, 1024-1031. DOI:10.2307/1940602URL [本文引用: 2]
FacelliJM, PickettSTA (1991b). Plant litter: its dynamics and effects on plant community structure The Botanical Review, 57, 1-32. DOI:10.1007/BF02858763URL [本文引用: 2]
FangJY, YangYH, MaWH, MohammatA, ShenHH (2010). Ecosystem carbon stocks and their changes in China?s grasslands Science China Life Sciences, 53, 757-765. DOI:10.1007/s11427-010-4029-xURLPMID:20697865 [本文引用: 1] The knowledge of carbon (C) stock and its dynamics is crucial for understanding the role of grassland ecosystems in China's terrestrial C cycle. To date, a comprehensive assessment on C balance in China's grasslands is still lacking. By reviewing published literature, this study aims to evaluate ecosystem C stocks (both vegetation biomass and soil organic C) and their changes in China's grasslands. Our results are summarized as follows: (1) biomass C density (C stock per area) of China's grasslands differed greatly among previous studies, ranging from 215.8 to 348.1 g C m(-2) with an average of 300.2 g C m(-2). Likewise, soil C density also varied greatly between 8.5 and 15.1 kg C m(-2). In total, ecosystem C stock in China's grasslands was estimated at 29.1 Pg C. (2) Both the magnitude and direction of ecosystem C changes in China's grasslands differed greatly among previous studies. According to recent reports, neither biomass nor soil C stock in China's grasslands showed a significant change during the past 20 years, indicating that grassland ecosystems are C neutral. (3) Spatial patterns and temporal dynamics of grassland biomass were closely correlated with precipitation, while changes in soil C stocks exhibited close associations with soil moisture and soil texture. Human activities, such as livestock grazing and fencing could also affect ecosystem C dynamics in China's grasslands.
FangJY, YuGR, LiuLL, HuSJ, ChapinIII FS (2018). Climate change, human impacts, and carbon sequestration in China Proceedings of the National Academy of Sciences of the United States of America, 115, 4015-4020. [本文引用: 1]
Heisler-WhiteJL, KnappAK, KellyEF (2008). Increasing precipitation event size increases aboveground net primary productivity in a semi-arid grassland Oecologia, 158, 129-140. DOI:10.1007/s00442-008-1116-9URLPMID:18670792 [本文引用: 1] Water availability is the primary constraint to aboveground net primary productivity (ANPP) in many terrestrial biomes, and it is an ecosystem driver that will be strongly altered by future climate change. Global circulation models predict a shift in precipitation patterns to growing season rainfall events that are larger in size but fewer in number. This
IsbellF, ReichPB, TilmanD, HobbieSE, PolaskyS, BinderS (2013). Nutrient enrichment, biodiversity loss, and consequent declines in ecosystem productivity Proceedings of the National Academy of Sciences of the United States of America, 110, 11911-11916. [本文引用: 1]
JensenK, GutekunstK (2003). Effects of litter on establishment of grassland plant species: the role of seed size and successional status Basic and Applied Ecology, 4, 579-587. DOI:10.1078/1439-1791-00179URL [本文引用: 1] AbstractLitter layers have a negative effect on seedling recruitment in many ecosystems. Successional fen grasslands are characterized by the built-up of litter layers which might be a key-factor influencing species composition and diversity during succession. Seed size is thought to be an important feature for the regeneration niche of species. Large seed size has been shown to be of advantage for germination and establishment under shaded conditions and beneath litter layers and thus for late-successional species.We investigated the effects of litter (control, litter layer of 3 cm and 8 cm) on seedling recruitment in 35 grassland species varying in seed size and successional status and on the abiotic conditions temperature, light quantity and light quality. Furthermore, we analyzed the species' light requirement for germination and its relation to seed size, successional status and establishment beneath litter.Light quantity was significantly lower beneath litter (reduction of 78% and 91% beneath 3 cm and 8 cm litter, respectively). The R/FR-ratio decreased from 1.2 without litter to 0.8 and 0.6 in presence of 3 cm and 8 cm litter. Seedling recruitment of 33 species was hampered by a litter layer. Establishment in presence of litter was positively related to seed size (r = 0.62, p < 0.001 and r = 0.46, p < 0.01 for 3 cm and 8 cm litter, respectively). In addition, establishment of early-successional species was hampered by litter to a significantly greater extent than recruitment of late-successional species independent of seed mass (ANCOVA: F = 7.6, p < 0.01). We found a positive relation between seed mass and the proportion of germination in darkness (r = 0.57, p < 0.001) indicating that small-seeded species had a higher light requirement for germination. We also found positive relationships between the proportion of germination in darkness and establishment in the 3 cm and the 8 cm litter treatment (r = 0.4, p < 0.05 and r = 0.3, p < 0.05 for 3 cm and 8 cm litter, respectively).We conclude that species-specific effects on seedling establishment are mainly due to differences in seed mass, successional status, and light requirement for germination and that these factors thus influence species composition and diversity during abandoned fen grassland succession.Nach der Nutzungsaufgabe ist die Sukzession auf Feuchtgrünland-Brachen durch die Bildung mächtiger Streuauflagen gekennzeichnet. Durch Effekte auf Keimung und Etablierung beeinflusst die Streuauflage die Artenzusammensetzung und -vielfalt und wird deshalb als ein Schlüsselfaktor für die Vegetationsentwicklung angesehen.In einem Gewächshaus-Experiment wurde der Einfluss unterschiedlicher Streumächtigkeiten (Kontrolle, 3 cm und 8 cm) auf die abiotischen Faktoren Temperatur, Lichtintensität und Lichtqualität sowie auf die Keimlingsetablierung von 35 Grünlandarten untersucht. Die ausgewählten Arten unterscheiden sich hinsichtlich ihres Samengewichts (von 0,1 bis 4,8 mg) sowie ihres Verhaltens im Sukzessionsverlauf (Arten früher sowie später Sukzessionstadien). Weiterhin wurden die Zusammenhänge zwischen der Lichtbedürftigkeit der Keimung und dem Samengewicht, dem Verhalten im Sukzessionsverlauf und der Keimlingsetablierung unter Streuauflagen analysiert.Unter den Streuvarianten war die Lichtintensität signifikant niedriger als in der Kontrolle (Reduktion um 78% und 91% unter 3 und 8 cm Streuauflage). Das Hellrot/Dunkelrot-Verhältnis sank von 1,2 ohne Streuauflage auf 0,8 und 0,6 unter 3 bzw. 8 cm Streumächtigkeit. Die Keimlingsetablierung von 33 Arten wurde durch die Streu signifikant verringert. Die Etablierung von Arten mit einem hohen Samengewicht wurde weniger durch die Streuauflagen beeinflusst als diejenige kleinsamiger Arten (r = 0,62, p < 0,001 und r = 0,46, p < 0,01 für 3 cm und 8 cm Streumächtigkeit). Zusätzlich wurde (auch bei Berücksichtigung des Samengewichts als Kovariable) die Keimlingsetablierung von Arten früher Sukzessionsstadien stärker verringert als die von Arten später Sukzessionsstadien (ANCOVA: F = 7,6, p < 0,01). Weiterhin wurde ein positiver Zusammenhang zwischen dem Samengewicht und dem Keimungsprozentsatz im Dunkeln gefunden (r = 0,57, p < 0,001), was zeigt, dass kleinsamige Arten eine höhere Lichtbedürftigkeit für die Keimung aufweisen. Auch zwischen dem Anteil der Keimung im Dunkeln sowie der Keimlingsetablierung unter Streuauflagen wurden positive Zusammenhänge nachgewiesen (r = 0,4, p < 0,05 und r = 0,3, p < 0,05 für 3 cm bzw. 8 cm Streumächtigkeit).Die Ergebnisse zeigen, dass die artenspezifischen Unterschiede hinsichtlich der Keimlingsetablierung unter Streuauflagen im Wesentlichen durch die Faktoren Samengewicht, Lichtbedürftigkeit für die Keimung sowie das Sukzessionsverhalten erklärt werden können und dass diese Eigenschaften somit die Artenzusammensetzung und -vielfalt während der Sukzession auf Feuchtgrünland-Brachen beeinflussen.]]>
JiangHZ (2018). Effects of Litter Removal and Litter Addition on Community Structure and Ecosystem Function of Stepper Ecosystem. Master degree dissertation, University of Chinese Academy of Sciences, Beijing. [本文引用: 1]
JingZB, ChengJM, ChenA (2013). Assessment of vegetative ecological characteristics and the succession process during three decades of grazing exclusion in a continental steppe grassland Ecological Engineering, 57, 162-169. DOI:10.1016/j.ecoleng.2013.04.035URL [本文引用: 1] Stipa bungeana steppe widely distributes in western China and Loess Plateau region, and is mainly used for soil and water conservation. Because of serious degradation, a 30 years (1982-2011) experiment has been carried out involving long-term grazing exclusion. What impacts will climate change have on vegetative ecological characteristics, ecosystem productivity and the succession of semiarid grasslands? The temperature, rainfall, soil moisture, vegetation characteristics, biomass and succession process were analyzed using 5 reference years in the period of grazing exclusion. Our results showed that the temperature increased gradually, but annual rainfall decreased gradually during the years 1982-2011. For soil moisture in the 0-60 cm soil layer, the main changes were an initial increase in the early stages of restoration (from 0 to 15 years), followed by a gradual decrease (from 16 to 30 years). The coverage, numbers of plants, plant density, and the Shannon-Wiener index showed similar variation tendencies as they increased during the plant community restoration process and all reached peak values in the 20th year. Above-and below-ground biomass also initially increased and then decreased with time of restoration, with highest values observed in the 20th year. The coverage, numbers of plants, plant density, Shannon-Wiener index, and above-and below-ground biomass all significantly increased with increasing annual mean temperature, growing season temperature and soil moisture in each sampled soil layer, but were not related to rainfall. The plant communities were composed of Gramineae, Leguminosae, and Compositae. Perennial species, C-3 plants and xerophytes were the main indicators of the succession process and vegetation restoration. Long-term grazing exclusion could exert a negative influence on the species renewal and productivity. Therefore, time is a very important factor for ecological restoration. These results provide reliable reference information for the similar natural world grasslands. (C) 2013 Elsevier B.V.
JonesDL, WillettVB (2006). Experimental evaluation of methods to quantify dissolved organic nitrogen (DON) and dissolved organic carbon (DOC) in soil Soil Biology & Biochemistry, 38, 991-999. DOI:10.1016/j.soilbio.2005.08.012URL [本文引用: 1]
KelemenA, T?r?kP, ValkóO, MigléczT, TóthmérészB (2013). Mechanisms shaping plant biomass and species richness: plant strategies and litter effect in alkali and loess grasslands Journal of Vegetation Science, 24, 1195-1203. DOI:10.1111/jvs.2013.24.issue-6URL [本文引用: 1]
LambEG (2008). Direct and indirect control of grassland community structure by litter, resources, and biomass Ecology, 89, 216-225. DOI:10.1890/07-0393.1URLPMID:18376563 [本文引用: 1] Multiple factors linked through complex networks of interaction including fertilization, aboveground biomass, and litter control the diversity of plant communities. The challenge of explaining plant diversity is to determine not only how each individual mechanism directly influences diversity, but how those mechanisms indirectly influence diversity through interactions with other mechanisms. This approach is well established in the study of plant species richness, but surprisingly little effort has been dedicated toward understanding the controls of community evenness, despite the recognition that this aspect of diversity can influence a variety of critical ecosystem functions. Similarly, studies of diversity have predominantly focused on the influence of shoot, rather than root, biomass, despite the fact that the majority of plant biomass is belowground in many natural communities. In this study, I examine the roles of belowground biomass, live aboveground biomass, litter, and light availability in controlling the species richness and evenness of a rough fescue grassland community using structural equation modeling. Litter was the primary mechanism structuring grassland diversity, with both richness and evenness declining with increasing litter cover. There were few relationships between shoot biomass, shading, and diversity, and more importantly, no relationship between root biomass and diversity. The lack of relationship between root biomass and species richness and evenness suggests that, even though root competition in grasslands is intense, belowground interactions may not play an important role in structuring community diversity or composition.
LeBauerDS, TresederKK (2008). Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed Ecology, 89, 371-379. DOI:10.1890/06-2057.1URLPMID:18409427 [本文引用: 1] Our meta-analysis of 126 nitrogen addition experiments evaluated nitrogen (N) limitation of net primary production (NPP) in terrestrial ecosystems. We tested the hypothesis that N limitation is widespread among biomes and influenced by geography and climate. We used the response ratio (R approximately equal ANPP(N)/ANPP(ctrl)) of aboveground plant growth in fertilized to control plots and found that most ecosystems are nitrogen limited with an average 29% growth response to nitrogen (i.e., R = 1.29). The response ratio was significant within temperate forests (R = 1.19), tropical forests (R = 1.60), temperate grasslands (R = 1.53), tropical grasslands (R = 1.26), wetlands (R = 1.16), and tundra (R = 1.35), but not deserts. Eight tropical forest studies had been conducted on very young volcanic soils in Hawaii, and this subgroup was strongly N limited (R = 2.13), which resulted in a negative correlation between forest R and latitude. The degree of N limitation in the remainder of the tropical forest studies (R = 1.20) was comparable to that of temperate forests, and when the young Hawaiian subgroup was excluded, forest R did not vary with latitude. Grassland response increased with latitude, but was independent of temperature and precipitation. These results suggest that the global N and C cycles interact strongly and that geography can mediate ecosystem response to N within certain biome types.
LettsB, LambEG, MischkolzJM, RomoJT (2015). Litter accumulation drives grassland plant community composition and functional diversity via leaf traits Plant Ecology, 216, 357-370. DOI:10.1007/s11258-014-0436-6URL [本文引用: 2]
LiJZ, LinS, TaubeF, PanQM, DittertK (2011). Above and belowground net primary productivity of grassland influenced by supplemental water and nitrogen in Inner Mongolia Plant and Soil, 340, 253-264. DOI:10.1007/s11104-010-0612-yURL [本文引用: 2] While water availability determines grassland productivity in semiarid regions, nutrient availability is the main limiting factor under wet conditions. An experiment was conducted in 2008 at two sites in Inner Mongolia with histories of heavy grazing (HG) and moderate grazing (MG) to study the interactive effects of water and nitrogen on above- and belowground net primary productivity (ANPP and BNPP), biomass partitioning, and plant species composition. The study comprises two water treatments (no irrigation and irrigated when soil water content was below 70% of the field capacity), and two nitrogen (N) levels (0 and 100 kg N ha(-1)). Mean values of ANPP at the peak biomass time reached 1,028 +/- 95 SD g m(-2) at the HG site and 568 +/- 32 SD g m(-2) at the MG site in irrigated and fertilized treatment. Nitrogen use efficiency (NUE) was significantly higher at irrigated plots compared to rain-fed plots at both HG and MG sites. Water use efficiency (WUEt) based on total water input and ANPP decreased with irrigation at the HG site. Meanwhile, N application significantly increased WUEt, WUEp (based on precipitation), and WUEi (based on irrigation water) at both sites. BNPP was significantly higher at irrigated plots compared to rain-fed plots at both HG and MG sites, and it tended to decrease with N addition. However, the fraction of belowground to total biomass (f(BNPP) = BNPP/(ANPP+BNPP) decreased with the addition of supplemental resources and exhibited a negative correlation with ANPP. Species diversity remained lower at the HG site compared to the MG site; it decreased with the addition of supplemental resources at the latter site. The annual Salsola collina contributed the most to the total biomass under irrigation. Based on global climate models, more frequent extreme climates are predicted in the future, which can result in changes in resource availabilities. Therefore, our research results have important implications for predicting the production and other properties of grassland ecosystems.
LiuJS, CuiY, LiXF, WilseyBJ, IsbellF, WanSQ, WangL, WangDL (2018). Reversal of nitrogen-induced species diversity declines mediated by change in dominant grass and litter Oecologia, 188, 921-929. DOI:10.1007/s00442-018-4252-xURLPMID:30143873 [本文引用: 1] Atmospheric nitrogen (N) deposition reduces plant diversity. However, it often remains unclear how dominant species and litter accumulation feedbacks mediate N-induced plant diversity declines. We tested mechanisms of N-induced diversity change through dominant grasses and litter in a 7-year field experiment. Nitrogen addition reduced species richness, Shannon-Wiener diversity (H') and evenness from the second to the fourth year, however, surprisingly, increased them in the sixth and seventh year. The reversal in the response of diversity to N addition was explained by changes in grass dominance and standing litter accumulation. The diversity recovery during later years in fertilized plots was attributed to a decrease in the dominant grass and an increase in standing litter: standing litter reduced bud numbers of the dominant grass by decreasing light availability. The decreased light availability by standing litter reduced completion from the dominant species, which resulted in diversity increase. The negative feedback between dominant grasses and standing litter led to transient N-induced diversity loss in the short-term, but recovery of plant diversity in the long-term. Grassland management that affects litter accumulation, such as firing, grazing and mowing, can therefore, have substantial effects on the long-term response of plant diversity to N deposition.
LuM, ZhouXH, LuoYQ, YangYH, FangCM, ChenJK, LiB (2011). Minor stimulation of soil carbon storage by nitrogen addition: a meta-analysis. Agriculture, Ecosystems & Environment, 140, 234-244. [本文引用: 1]
ManningP, SaundersM, BardgettRD, BonkowskiM, BradfordMA, EllisRJ, KandelerE, MarhanS, TscherkoD (2008). Direct and indirect effects of nitrogen deposition on litter decomposition Soil Biology & Biochemistry, 40, 688-698. DOI:10.1016/j.soilbio.2007.08.023URL [本文引用: 1]
PanQM, BaiYF, HanXG, YangJC (2005). Effects of nitrogen additions on a Leymus chinensis population in typical steppe of Inner Mongolia Acta Phytoecologica Sinica, 29, 311-317. [本文引用: 2]
PatrickLB, FraserLH, KershnerMW (2008). Large-scale manipulation of plant litter and fertilizer in a managed successional temperate grassland Plant Ecology, 197, 183-195. DOI:10.1007/s11258-007-9369-7URL [本文引用: 2] Plant litter may play an important role in herbaceous plant communities by limiting primary production and influencing plant species richness. However, it is not known how the effect of litter interacts with fertilization. We tested for the role of litter and fertilization in a large-scale experiment to investigate effects on diversity and biomass of plant species, growth forms, native vs. non-native groups, and abiotic ecosystem components (e.g., soil moisture, PAR). We manipulated plant litter (removed vs. left insitu) and nutrient availability (NPK-fertilized vs. unfertilized) for 4years in 314-m2 plots, replicated six times, in an old-field grassland. While many of our species-level results supported previously published studies and theory, our plant group results generally did not. Specifically, grass species richness and forb biomass was not affected by either fertilization or plant litter. Moreover, plant litter removal significantly increased non-native plant species richness. Relative to native plant species, all of our experimental manipulations significantly increased both the biomass and the species richness of non-native plant species. Thus, this grassland system was sensitive to management treatments through the facilitation of non-native plant species. We coupled biotic and abiotic components within a nonmetric multidimensional scaling (NMS) analysis to investigate treatment effects, which revealed that specific treatments altered ecosystem development. These results suggest that fertilization and plant litter may have larger impacts on plant communities and on ecosystem properties than previously understood, underscoring the need for larger-scale and longer-term experiments.]]>
PiaoS, FangJ, CiaisP, PeylinP, HuangY, SitchS, WangT (2009). The carbon balance of terrestrial ecosystems in China Nature, 458, 1009-1013. DOI:10.1038/nature07944URLPMID:19396142 [本文引用: 1] Global terrestrial ecosystems absorbed carbon at a rate of 1-4 Pg yr(-1) during the 1980s and 1990s, offsetting 10-60 per cent of the fossil-fuel emissions. The regional patterns and causes of terrestrial carbon sources and sinks, however, remain uncertain. With increasing scientific and political interest in regional aspects of the global carbon cycle, there is a strong impetus to better understand the carbon balance of China. This is not only because China is the world's most populous country and the largest emitter of fossil-fuel CO(2) into the atmosphere, but also because it has experienced regionally distinct land-use histories and climate trends, which together control the carbon budget of its ecosystems. Here we analyse the current terrestrial carbon balance of China and its driving mechanisms during the 1980s and 1990s using three different methods: biomass and soil carbon inventories extrapolated by satellite greenness measurements, ecosystem models and atmospheric inversions. The three methods produce similar estimates of a net carbon sink in the range of 0.19-0.26 Pg carbon (PgC) per year, which is smaller than that in the conterminous United States but comparable to that in geographic Europe. We find that northeast China is a net source of CO(2) to the atmosphere owing to overharvesting and degradation of forests. By contrast, southern China accounts for more than 65 per cent of the carbon sink, which can be attributed to regional climate change, large-scale plantation programmes active since the 1980s and shrub recovery. Shrub recovery is identified as the most uncertain factor contributing to the carbon sink. Our data and model results together indicate that China's terrestrial ecosystems absorbed 28-37 per cent of its cumulated fossil carbon emissions during the 1980s and 1990s.
RiceEL, ParentiRL (1978). Causes of decreases in productivity in undisturbed tall grass prairie American Journal of Botany, 65, 1091-1097. DOI:10.1002/ajb2.1978.65.issue-10URL [本文引用: 1]
RuprechtE, EnyediMZ, EcksteinRL, DonathTW (2010). Restorative removal of plant litter and vegetation 40 years after abandonment enhances re-emergence of steppe grassland vegetation Biological Conservation, 143, 449-456. DOI:10.1016/j.biocon.2009.11.012URL [本文引用: 1]
SasakiT, LuX, HirotaM, BaiY (2019). Species asynchrony and response diversity determine multifunctional stability of natural grasslands Journal of Ecology, 107, 1862-1875. DOI:10.1111/jec.2019.107.issue-4URL [本文引用: 1]
ShenY (2016). Effects of Water and Nitrogen and Litter Addition on Leymus chinensis Garssland. PhD dissertation, China Agricultural University, Beijing. [本文引用: 3]
ShenY, ChenWQ, YangGW, YangX, LiuN, SunX, ChenJS, ZhangYJ (2016). Can litter addition mediate plant productivity responses to increased precipitation and nitrogen deposition in a typical steppe? Ecological Research, 31, 579-587. DOI:10.1007/s11284-016-1368-5URL [本文引用: 1]
StevensCJ, DiseNB, MountfordJO, GowingDJ (2004). Impact of nitrogen deposition on the species richness of grasslands Science, 303, 1876-1879. DOI:10.1126/science.1094678URLPMID:15031507 [本文引用: 3] A transect of 68 acid grasslands across Great Britain, covering the lower range of ambient annual nitrogen deposition in the industrialized world (5 to 35 kg Nha-1 year-1), indicates that long-term, chronic nitrogen deposition has significantly reduced plant species richness. Species richness declines as a linear function of the rate of inorganic nitrogen deposition, with a reduction of one species per 4-m2 quadrat for every 2.5 kg Nha-1 year-1 of chronic nitrogen deposition. Species adapted to infertile conditions are systematically reduced at high nitrogen deposition. At the mean chronic nitrogen deposition rate of central Europe (17 kg Nha-1 year-1), there is a 23% species reduction compared with grasslands receiving the lowest levels of nitrogen deposition.
SudingKN, CollinsSL, GoughL, ClarkC, ClelandEE, GrossKL, MilchunasDG, PenningsS (2005). Functional-and abundance-based mechanisms explain diversity loss due to N fertilization Proceedings of the National Academy of Sciences of the United States of America, 102, 4387-4392. [本文引用: 1]
TianQY, LiuNN, BaiWM, LiLH, ChenJQ, ReichPB, YuQ, GuoDL, SmithMD, KnappAK, ChengWX, LuP, GaoY, YangA, WangTZ, LiX, WangZW, MaYB, HanXG, ZhangWH (2016). A novel soil manganese mechanism drives plant species loss with increased nitrogen deposition in a temperate steppe Ecology, 97, 65-74. DOI:10.1890/15-0917.1URLPMID:27008776 [本文引用: 1] Loss of plant diversity with increased anthropogenic nitrogen (N) deposition in grasslands has occurred globally. In most cases, competitive exclusion driven by preemption of light or space is invoked as a key mechanism. Here, we provide evidence from a 9-yr N-addition experiment for an alternative mechanism: differential sensitivity of forbs and grasses to increased soil manganese (Mn) levels. In Inner Mongolia steppes, increasing the N supply shifted plant community composition from grass-forb codominance (primarily Stipa krylovii and Artemisia frigida, respectively) to exclusive dominance by grass, with associated declines in overall species richness. Reduced abundance of forbs was linked to soil acidification that increased mobilization of soil Mn, with a 10-fold greater accumulation of Mn in forbs than in grasses. The enhanced accumulation of Mn in forbs was correlated with reduced photosynthetic rates and growth, and is consistent with the loss of forb species. Differential accumulation of Mn between forbs and grasses can be linked to fundamental differences between dicots and monocots in the biochemical pathways regulating metal transport. These findings provide a mechanistic explanation for N-induced species loss in temperate grasslands by linking metal mobilization in soil to differential metal acquisition and impacts on key functional groups in these ecosystems.
VargaC, FeketeI, KotroczóZ, KrakompergerZ, VinczeG (2008). The effect of litter on soil organic matter (SOM) turnover in Síkf?kút site Cereal Research Communications, 36, 547-550. [本文引用: 1]
VitousekPM, HowarthRW (1991). Nitrogen limitation on land and in the sea: How can it occur? Biogeochemistry, 13, 87-115. [本文引用: 1]
WanHW, YangY, BaiSQ, XuYH, BaiYH (2008). Variation in leaf functional traits of six species along a nitrogen addition gradient in Leymus chinensis steppe in Inner Mongolia Journal of Plant Ecology (Chinese Version), 32, 611-321. [本文引用: 1]
WangJ (2011). The Effect of Litter on Vegetation and Soil Moisture in Typical Grassland. PhD dissertation, Inner Mongolia Agricultural University, Hohhot. [本文引用: 1]
WangJ, GaoYZ, ZhangYH, YangJJ, SmithMD, KnappAK, EissenstatDM, HanXG (2019). Asymmetry in above- and belowground productivity responses to N addition in a semi-arid temperate steppe Global Change Biology, 25, 2958-2969. DOI:10.1111/gcb.14719URLPMID:31152626 [本文引用: 1]
WangJ, ZhaoML, WillmsWD, HanGD, WangZW, BaiYF (2011). Can plant litter affect net primary production of a typical steppe in Inner Mongolia? Journal of Vegetation Science, 22, 367-376. DOI:10.1111/j.1654-1103.2011.01257.xURL [本文引用: 2] Methods;We examined the role of plant litter in two experiments where litter was either removed or added in a protected or heavily grazed site, respectively, in autumn and in spring in a split plot design. The treatments (control, moderate and heavy litter application) were applied once in five replications but repeated at new locations in each of 3 years. This was done to examine only the direct effect of litter on annual net primary production and selected plant characteristics and not potential secondary effects. We also measured soil moisture and soil temperature.Results;Removing litter caused a reduction in the amount of grass (Leymus chinensis) that was produced, but litter addition caused an inconsistent effect among years, with moderate applications producing the most positive effects. Litter removal resulted in shorter and less dense plants of L. chinensis and Carex duriuscula, while heavy litter addition in autumn reduced plant height of both Cleistogenes squarrosa and C. duriuscula.Conclusions;Litter was effective for enhancing soil moisture status and reducing soil heat units in the typical steppe of Inner Mongolia. Therefore, litter mass may serve as an index of grassland health in such environments.]]>
WangJ-J, PisaniO, LinLH, LunOOY, BowdenRD, LajthaK, SimpsonAJ, SimpsonMJ (2017a). Long-term litter manipulation alters soil organic matter turnover in a temperate deciduous forest Science of the Total Environment, 608, 865-875. [本文引用: 2]
WangSS (2016). Effects of Altered Litter Input on Productivity and Carbon Fluxes of a Semiarid Grassland, Inner Mongolia. Master degree dissertation, University of Chinese Academy of Sciences, Beijing. [本文引用: 1]
WangX, XuZ, LüX, WangR, CaiJ, YangS, LiM-H, JiangY (2017b). Responses of litter decomposition and nutrient release rate to water and nitrogen addition differed among three plant species dominated in a semi-arid grassland Plant and Soil, 418, 241-253. DOI:10.1007/s11104-017-3288-8URL [本文引用: 1]
WardleDA, BardgettRD, KlironomosJN, Set?l?H, vander Putten WH, WallDH (2004). Ecological linkages between aboveground and belowground biota Science, 304, 1629-1633. DOI:10.1126/science.1094875URLPMID:15192218 [本文引用: 1] All terrestrial ecosystems consist of aboveground and belowground components that interact to influence community- and ecosystem-level processes and properties. Here we show how these components are closely interlinked at the community level, reinforced by a greater degree of specificity between plants and soil organisms than has been previously supposed. As such, aboveground and belowground communities can be powerful mutual drivers, with both positive and negative feedbacks. A combined aboveground-belowground approach to community and ecosystem ecology is enhancing our understanding of the regulation and functional significance of biodiversity and of the environmental impacts of human-induced global change phenomena.
WikeemBM, NewmanRF, RyswykAL (1989). Effect of fertilization date and litter removal on grassland forage production Journal of Range Management, 42, 412-415. DOI:10.2307/3899551URL [本文引用: 1]
WilcoxKR, TredennickAT, KoernerSE, GrmanE, HallettLM, AvolioML, LaPierre KJ, HousemanGR, IsbellF, JohnsonDS, AlataloJM, BaldwinAH, BorkEW, BoughtonEH, BowmanWD, BrittonAJ, CahillJr JF, CollinsSL, DuGZ, EskelinenA, GoughL, JentschA, KernC, KlanderudK, KnappAK, KreylingJ, LuoYQ, McLarenJR, MegonigalP, OnipchenkoV, PrevéyJ, PriceJN, RobinsonCH, SalaOE, SmithMD, SoudzilovskaiaNA, SouzaL, TilmanD, WhiteSR, XuZW, YahdjianL, YuQ, ZhangPF, ZhangYH (2017). Asynchrony among local communities stabilises ecosystem function of metacommunities Ecology Letters, 20, 1534-1545. DOI:10.1111/ele.12861URLPMID:29067791 [本文引用: 1] Temporal stability of ecosystem functioning increases the predictability and reliability of ecosystem services, and understanding the drivers of stability across spatial scales is important for land management and policy decisions. We used species-level abundance data from 62 plant communities across five continents to assess mechanisms of temporal stability across spatial scales. We assessed how asynchrony (i.e. different units responding dissimilarly through time) of species and local communities stabilised metacommunity ecosystem function. Asynchrony of species increased stability of local communities, and asynchrony among local communities enhanced metacommunity stability by a wide range of magnitudes (1-315%); this range was positively correlated with the size of the metacommunity. Additionally, asynchronous responses among local communities were linked with species' populations fluctuating asynchronously across space, perhaps stemming from physical and/or competitive differences among local communities. Accordingly, we suggest spatial heterogeneity should be a major focus for maintaining the stability of ecosystem services at larger spatial scales.
WuGL, DuGZ, LiuZH, ThirgoodS (2009). Effect of fencing and grazing on a Kobresia-dominated meadow in the Qinghai-Tibetan Plateau Plant and Soil, 319, 115-126. DOI:10.1007/s11104-008-9854-3URL [本文引用: 2] Grazing is one of the most important factors influencing community structure and productivity in natural grasslands. Fencing to exclude grazers is one of the main management practices used to protect grasslands. Can fencing improve grassland community status by restraining grazing? We conducted a field community study and indoor soil analyses to determine the long-term effects of fencing and grazing on the above-ground community and soil in a Kobresia-dominated meadow in the Qinghai-Tibetan Plateau, NW China. Our results showed that fencing significantly improved above-ground vegetation productivity but reduced plant density and species diversity. Long-term fencing favored the improvement of forage grass functional groups and restrained the development noxious weed functional groups. There were significant positive effects of fencing on below-ground organic matter, total nitrogen, available nitrogen, total phosphorus and available phosphorus. The productivity of grazed meadow showed a weak decrease over time. There were long-term decreasing trends for plant density both in fenced and grazed meadows. Our study suggests that grazing can be considered as a useful management practice to improve species diversity and plant density in long-term fenced grasslands and that periodic grazing and fencing is beneficial in grassland management.]]>
WuJB, WangXD (2019). Temporal stability of aboveground net primary production in northern Tibet alpine steppe in response to nitrogen addition Journal of Mountain Science, 16, 2679-2686. DOI:10.1007/s11629-018-5135-7URL
XiaJY, WanSQ (2008). Global response patterns of terrestrial plant species to nitrogen addition New Phytologist, 179, 428-439. DOI:10.1111/nph.2008.179.issue-2URL [本文引用: 1]
XuS, LiuL, SayerEJ (2013a). Variability of above-ground litter inputs alters soil physicochemical and biological processes: a meta-analysis of litterfall-manipulation experiments Biogeosciences, 10, 7423-7433. DOI:10.5194/bg-10-7423-2013URL [本文引用: 2] Global change has been shown to alter the amount of above-ground litter inputs to soil greatly, which could cause substantial cascading effects on below-ground biogeochemical cycling. Despite extensive study, there is uncertainty about how changes in above-ground litter inputs affect soil carbon and nutrient turnover and transformation. Here, we conducted a meta-analysis on 70 litter-manipulation experiments in order to assess how changes in above-ground litter inputs alter soil physicochemical properties, carbon dynamics and nutrient cycles. Our results demonstrated that litter removal decreased soil respiration by 34%, microbial biomass carbon in the mineral soil by 39% and total carbon in the mineral soil by 10%, whereas litter addition increased them by 31, 26 and 10%, respectively. This suggests that greater litter inputs increase the soil carbon sink despite higher rates of carbon release and transformation. Total nitrogen and extractable inorganic nitrogen in the mineral soil decreased by 17 and 30%, respectively, under litter removal, but were not altered by litter addition. Overall, litter manipulation had a significant impact upon soil temperature and moisture, but not soil pH; litter inputs were more crucial in buffering soil temperature and moisture fluctuations in grassland than in forest. Compared to other ecosystems, tropical and subtropical forests were more sensitive to variation in litter inputs, as altered litter inputs affected the turnover and accumulation of soil carbon and nutrients more substantially over a shorter time period. Our study demonstrates that although the magnitude of responses differed greatly among ecosystems, the direction of the responses was very similar across different ecosystems. Interactions between plant productivity and below-ground biogeochemical cycling need to be taken into account to predict ecosystem responses to environmental change.
XuX, SherryRA, NiuS, LiD, LuoY (2013b). Net primary productivity and rain-use efficiency as affected by warming, altered precipitation, and clipping in a mixed-grass prairie Global Change Biology, 19, 2753-2764. DOI:10.1111/gcb.12248URL [本文引用: 1] Grassland productivity in response to climate change and land use is a global concern. In order to explore the effects of climate change and land use on net primary productivity (NPP), NPP partitioning [f(BNPP), defined as the fraction of belowground NPP (BNPP) to NPP], and rain-use efficiency (RUE) of NPP, we conducted a field experiment with warming (+3 degrees C), altered precipitation (double and half), and annual clipping in a mixed-grass prairie in Oklahoma, USA since July, 2009. Across the years, warming significantly increased BNPP, f(BNPP), and RUEBNPP by an average of 11.6%, 2.8%, and 6.6%, respectively. This indicates that BNPP was more sensitive to warming than aboveground NPP (ANPP) since warming did not change ANPP and RUEANPP much. Double precipitation stimulated ANPP, BNPP, and NPP but suppressed RUEANPP, RUEBNPP, and RUENPP while half precipitation decreased ANPP, BNPP, and NPP but increased RUEANPP, RUEBNPP, and RUENPP. Clipping interacted with altered precipitation in impacting RUEANPP, RUEBNPP, and RUENPP, suggesting land use could confound the effects of precipitation changes on ecosystem processes. Soil moisture was found to be a main factor in regulating variation in ANPP, BNPP, and NPP while soil temperature was the dominant factor influencing f(BNPP). These findings suggest that BNPP is critical point to future research. Additionally, results from single-factor manipulative experiments should be treated with caution due to the non-additive interactive effects of warming with altered precipitation and land use (clipping).
XuZW, RenHY, LiMH, vanRuijven J, HanXG, WanSQ, LiH, YuQ, JiangY, JiangL (2015). Environmental changes drive the temporal stability of semi-arid natural grasslands through altering species asynchrony Journal of Ecology, 103, 1308-1316. DOI:10.1111/1365-2745.12441URL [本文引用: 1]
YanLM, ChenSP, HuangJH, LinGH (2010). Differential responses of auto- and heterotrophic soil respiration to water and nitrogen addition in a semiarid temperate steppe Global Change Biology, 16, 2345-2357. DOI:10.1111/gcb.2010.16.issue-8URL [本文引用: 1]
YangH, JiangL, LiL, LiA, WuMY, WanSQ (2012). Diversity-dependent stability under mowing and nutrient addition: evidence from a 7-year grassland experiment Ecology Letters, 15, 619-626. DOI:10.1111/j.1461-0248.2012.01778.xURL [本文引用: 1] Ecology Letters (2012) Abstract Anthropogenic perturbations may affect biodiversity and ecological stability as well as their relationships. However, diversitystability patterns and associated mechanisms under human disturbances have rarely been explored. We conducted a 7-year field experiment examining the effects of mowing and nutrient addition on the diversity and temporal stability of herbaceous plant communities in a temperate steppe in northern China. Mowing increased population and community stability, whereas nutrient addition had the opposite effects. Stability exhibited positive relationships with species richness at population, functional group and community levels. Treatments did not alter these positive diversitystability relationships, which were associated with the stabilising effect of species richness on component populations, species asynchrony and portfolio effects. Despite the difficulty of pinpointing causal mechanisms of diversitystability patterns observed in nature, our results suggest that diversity may still be a useful predictor of the stability of ecosystems confronted with anthropogenic disturbances.
YangH, LiY, WuMY, ZhangZ, LiL, WanSQ (2011). Plant community responses to nitrogen addition and increased precipitation: the importance of water availability and species traits Global Change Biology, 17, 2936-2944. DOI:10.1111/j.1365-2486.2011.02423.xURL [本文引用: 1] Global nitrogen (N) enrichment and changing precipitation regimes are likely to alter plant community structure and composition, with consequent influences on biodiversity and ecosystem functioning. Responses of plant community structure and composition to N addition and increased precipitation were examined in a temperate steppe in northern China. Increased precipitation and N addition stimulated and suppressed community species richness, respectively, across 6 years (2005-2010) of the manipulative experiment. N addition and increased precipitation significantly altered plant community structure and composition at functional groups levels. The significant relationship between species richness and soil moisture (SM) suggests that plant community structure is mediated by water under changing environmental conditions. In addition, plant height played an important role in affecting the responses of plant communities to N addition, and the effects of increased precipitation on plant community were dependent on species rooting depth. Our results highlight the importance and complexity of both abiotic (SM) and biotic factors (species traits) in structuring plant community under changing environmental scenarios. These findings indicate that knowledge of species traits can contribute to mechanistic understanding and projection of vegetation dynamics in response to future environmental change.
YarwoodS, BrewerE, YarwoodR, LajthaK, MyroldD (2013). Soil microbe active community composition and capability of responding to litter addition after 12 years of no inputs Applied and Environmental Microbiology, 79, 1385-1392. DOI:10.1128/AEM.03181-12URL [本文引用: 1] One explanation given for the high microbial diversity found in soils is that they contain a large inactive biomass that is able to persist in soils for long periods of time. This persistent microbial fraction may help to buffer the functionality of the soil community during times of low nutrients by providing a reservoir of specialized functions that can be reactivated when conditions improve. A study was designed to test the hypothesis: in soils lacking fresh root or detrital inputs, microbial community composition may persist relatively unchanged. Upon addition of new inputs, this community will be stimulated to grow and break down litter similarly to control soils. Soils from two of the Detrital Input and Removal Treatments (DIRT) at the H. J. Andrews Experimental Forest, the no-input and control treatment plots, were used in a microcosm experiment where Douglas-fir needles were added to soils. After 3 and 151 days of incubation, soil microbial DNA and RNA was extracted and characterized using quantitative PCR (qPCR) and 454 pyrosequencing. The abundance of 16S and 28S gene copies and RNA copies did not vary with soil type or amendment; however, treatment differences were observed in the abundance of archaeal ammonia-oxidizing amoA gene abundance. Analysis of similar to 110,000 bacterial sequences showed a significant change in the active (RNA-based) community between day 3 and day 151, but microbial composition was similar between soil types. These results show that even after 12 years of plant litter exclusion, the legacy of community composition was well buffered against a dramatic disturbance.
YueK, PengY, PengCH, YangWQ, PengX, WuFZ (2016). Stimulation of terrestrial ecosystem carbon storage by nitrogen addition: a meta-analysis Scientific Reports, 6, 19895. DOI: 10.1038/srep19895. DOI:10.1038/srep19895URLPMID:26813078 [本文引用: 2] Elevated nitrogen (N) deposition alters the terrestrial carbon (C) cycle, which is likely to feed back to further climate change. However, how the overall terrestrial ecosystem C pools and fluxes respond to N addition remains unclear. By synthesizing data from multiple terrestrial ecosystems, we quantified the response of C pools and fluxes to experimental N addition using a comprehensive meta-analysis method. Our results showed that N addition significantly stimulated soil total C storage by 5.82% ([2.47%, 9.27%], 95% CI, the same below) and increased the C contents of the above- and below-ground parts of plants by 25.65% [11.07%, 42.12%] and 15.93% [6.80%, 25.85%], respectively. Furthermore, N addition significantly increased aboveground net primary production by 52.38% [40.58%, 65.19%] and litterfall by 14.67% [9.24%, 20.38%] at a global scale. However, the C influx from the plant litter to the soil through litter decomposition and the efflux from the soil due to microbial respiration and soil respiration showed insignificant responses to N addition. Overall, our meta-analysis suggested that N addition will increase soil C storage and plant C in both above- and below-ground parts, indicating that terrestrial ecosystems might act to strengthen as a C sink under increasing N deposition.
ZhangBW, TanXR, WangSS, ChenML, ChenSP, RenTT, XiaJY, BaiYF, HuangJH, HanXG (2017). Asymmetric sensitivity of ecosystem carbon and water processes in response to precipitation change in a semi-arid steppe Functional Ecology, 31, 1301-1311. DOI:10.1111/fec.2017.31.issue-6URL [本文引用: 1]
ZhangTA, LuoY, ChenHYH, RuanH (2018a). Responses of litter decomposition and nutrient release to N addition: a meta-analysis of terrestrial ecosystems Applied Soil Ecology, 128, 35-42. DOI:10.1016/j.apsoil.2018.04.004URL [本文引用: 2]
ZhangXY, WangW (2015). Control of climate and litter quality on leaf litter decomposition in different climatic zones Journal of Plant Research, 128, 791-802. DOI:10.1007/s10265-015-0743-6URLPMID:26135888 [本文引用: 1] Climate and initial litter quality are the major factors influencing decomposition rates on large scales. We established a comprehensive database of terrestrial leaf litter decomposition, including 785 datasets, to examine the relationship between climate and litter quality and evaluate the factors controlling decomposition on a global scale, the arid and semi-arid (AS) zone, the humid middle and humid low (HL) latitude zones. Initial litter nitrogen (N) and phosphorus (P) concentration only increased with mean annual temperature (MAT) in the AS zone and decreased with mean annual precipitation (MAP) in the HL zone. Compared with nutrient content, MAT imposed less effect on initial litter lignin content than MAP. MAT were the most important decomposition driving factors on a global scale as well as in different climatic zones. MAP only significantly affected decomposition constants in AS zone. Although litter quality parameters also showed significant influence on decomposition, their importance was less than the climatic factors. Besides, different litter quality parameters exerted significant influence on decomposition in different climatic zones. Our results emphasized that climate consistently exerted important effects on decomposition constants across different climatic zones.
ZhangY, LoreauM, LüX, HeN, ZhangG, HanX (2016). Nitrogen enrichment weakens ecosystem stability through decreased species asynchrony and population stability in a temperate grassland Global Change Biology, 22, 1445-1455. DOI:10.1111/gcb.13140URLPMID:26511538 [本文引用: 2] Biodiversity generally promotes ecosystem stability. To assess whether the diversity-stability relationship observed under ambient nitrogen (N) conditions still holds under N enriched conditions, we designed a 6-year field experiment to test whether the magnitude and frequency of N enrichment affects ecosystem stability and its relationship with species diversity in a temperate grassland. Results of this experiment showed that the frequency of N addition had no effect on either the temporal stability of ecosystem and population or the relationship between diversity and stability. Nitrogen addition decreased ecosystem stability significantly through decreases in species asynchrony and population stability. Species richness was positively associated with ecosystem stability, but no significant relationship between diversity and the residuals of ecosystem stability was detected after controlling for the effects of the magnitude of N addition, suggesting collinearity between the effects of N addition and species richness on ecosystem stability, with the former prevailing over the latter. Both population stability and the residuals of population stability after controlling for the effects of the magnitude of N addition were positively associated with ecosystem stability, indicating that the stabilizing effects of component populations were still present after N enrichment. Our study supports the theory predicting that the effects of environmental factors on ecosystem functioning are stronger than those of biodiversity. Understanding such mechanisms is important and urgent to protect biodiversity in mediating ecosystem functioning and services in the face of global changes.
ZhangYH, HeNP, LoreauM, PanQM, HanXG (2018b). Scale dependence of the diversity-stability relationship in a temperate grassland Journal of Ecology, 106, 1277-1285. DOI:10.1111/jec.2018.106.issue-3URL [本文引用: 1]
ZhaoYN, YangB, LiMX, XiaoRQ, RaoKY, WangJQ, ZhangT, GuoJX (2019). Community composition, structure and productivity in response to nitrogen and phosphorus additions in a temperate meadow Science of the Total Environment, 654, 863-871. DOI:10.1016/j.scitotenv.2018.11.155URL [本文引用: 2]
ZongN, ZhaoGS, ShiPL (2019). Different sensitivity and threshold in response to nitrogen addition in four alpine grasslands along a precipitation transect on the Northern Tibetan Plateau Ecology and Evolution, 9, 9782-9793. DOI:10.1002/ece3.5514URLPMID:31534693 [本文引用: 1] The increase in atmospheric nitrogen (N) deposition has resulted in some terrestrial ecological changes. In order to identify the response of sensitive indicators to N input and estimate the sensitivity and saturation thresholds in alpine grasslands, we set up a series of multilevel N addition experiments in four types of alpine grasslands (alpine meadow [AM], alpine meadow-steppe [AMS], alpine steppe [AS], and alpine desert-steppe [ADS]) along with a decreasing precipitation gradient from east to west on the Northern Tibetan Plateau. N addition only had significant effects on species diversity in AMS, while had no effects on the other three alpine grasslands. Aboveground biomass of grasses and overall community in ADS were enhanced with increasing N addition, but such effects did not occur in AS. Legume biomass in ADS and AS showed similar unimodal patterns and exhibited a decreasing tend in AM. Regression fitting showed that the most sensitive functional groups were grasses, and the N saturation thresholds were 103, 115, 136, and 156 kg N hm(-2) year(-1) in AM, AMS, AS, and ADS, respectively. This suggests that alpine grasslands become more and more insensitive to N input with precipitation decrease. N saturation thresholds also negatively correlated with soil N availability. N sensitivity differences caused by precipitation and nutrient availability suggest that alpine grasslands along the precipitation gradient will respond differently to atmospheric N deposition in the future global change scenario. This different sensitivity should also be taken into consideration when using N fertilization to restore degraded grasslands. Nutrient limitation on terrestrial plant growth-modeling the interaction between nitrogen and phosphorus 1 2012
... 在大多数陆地生态系统中, 氮是植物生长的主要限制因子(Vitousek & Howarth, 1991; LeBauer & Treseder, 2008; Xia & Wan, 2008; Lu et al., 2011; ?gren et al., 2012; Gao et al., 2019).大量研究表明氮添加会提高植物群落生产力, 但群落植物物种多样性却显著降低(Stevens et al., 2004; Suding et al., 2005; Clark & Tilman, 2008; Isbell et al., 2013).由于植物对氮、磷等养分元素的重吸收作用, 凋落物中的碳氮比相对较高(Deng et al., 2018).日益加剧的大气氮沉降, 会提高环境中活性氮的含量(Gruber & Galloway, 2008).这一方面降低了凋落物中碳氮比, 另一方面缓解了微生物的营养限制, 促进微生物活性, 进而加速凋落物的分解(Austin & Vivanco, 2006; Yue et al., 2016; Wang et al., 2017b), 改善了环境中的养分状况(Manning et al., 2008; Zhang et al., 2018a).那么, 凋落物和氮两者共同作用会对生态系统生产力和群落结构产生怎样的影响呢? 目前还缺乏长期的实验研究. ...
Responses of a California annual grassland to litter manipulation 2 2008
... 近年来, 凋落物输入变化对生态系统结构功能的影响更多聚焦在植被生产力、土壤微生物介导的养分循环过程和土壤碳库稳定性的变化上(Xu et al., 2013a), 而对于凋落物输入变化对群落结构影响的报道比较有限(王静, 2011; 蒋红志, 2018).凋落物对群落结构的影响则主要通过其对不同植物功能群的影响差异来实现, 如Facelli和Pickett (1991b)发现凋落物输入增加会抑制草本植物的生长, 其中既发现对外来种生长建群的抑制(Patrick et al., 2008), 也有对原生种生长的抑制, 这主要与凋落物类型有关(Facelli & Pickett, 1991b; Foster & Gross, 1998; Amatangelo et al., 2008; Letts et al., 2015).此外, 凋落物的累积能通过防止种子进入土壤层而对种子补充过程形成抑制, 也会通过化感作用抑制种子萌发, 累积的凋落物也能通过降低物种丰富度和均匀度来改变群落的物种组成(Foster & Gross, 1998; Amatangelo et al., 2008; Letts et al., 2015). ...
... ; Amatangelo et al., 2008; Letts et al., 2015). ...
Plant litter decomposition in a semi-arid ecosystem controlled by photodegradation 1 2006
... 在大多数陆地生态系统中, 氮是植物生长的主要限制因子(Vitousek & Howarth, 1991; LeBauer & Treseder, 2008; Xia & Wan, 2008; Lu et al., 2011; ?gren et al., 2012; Gao et al., 2019).大量研究表明氮添加会提高植物群落生产力, 但群落植物物种多样性却显著降低(Stevens et al., 2004; Suding et al., 2005; Clark & Tilman, 2008; Isbell et al., 2013).由于植物对氮、磷等养分元素的重吸收作用, 凋落物中的碳氮比相对较高(Deng et al., 2018).日益加剧的大气氮沉降, 会提高环境中活性氮的含量(Gruber & Galloway, 2008).这一方面降低了凋落物中碳氮比, 另一方面缓解了微生物的营养限制, 促进微生物活性, 进而加速凋落物的分解(Austin & Vivanco, 2006; Yue et al., 2016; Wang et al., 2017b), 改善了环境中的养分状况(Manning et al., 2008; Zhang et al., 2018a).那么, 凋落物和氮两者共同作用会对生态系统生产力和群落结构产生怎样的影响呢? 目前还缺乏长期的实验研究. ...
Ecosystem stability and compensatory effects in the Inner Mongolia grassland 2 2004
... 为了更好地研究物种组成的变化, 本研究按照物种的生活型将植物划分为5个不同功能群, 包括多年生丛生禾草(PB)、多年生根茎禾草(PR)、多年生杂类草(PF)、灌木与半灌木(SS)和一二年生植物(AB)(Bai et al., 2004). ...
... 在我国半干旱草原, 持续大量的氮添加会降低群落生物多样性是已经被大量的实验证实的结论(Stevens et al., 2004; Clark & Tilman, 2010; Zhang et al., 2016; Liu et al., 2018), 但与以往研究不同的是, 本研究经过6年的氮添加处理, 并没有发现物种和功能群多样性发生显著变化, 这可能是与本实验的氮添加量有关.在本研究区域已开展的长期氮添加梯度实验中, 草原生产力和多样性均与氮添加量表现出非线性关系, 呈现先增加后降低的趋势, 而拐点为10-15 g·m-2·a-1, 过量的氮添加会因为土壤酸化和有害离子的富集而导致群落生产力和多样性的降低(Stevens et al., 2004; Tian et al., 2016; Gao et al., 2019).本研究的施氮量为15 g·m-2·a-1, 对该区域草原生态系统是一个比较适宜的氮添加水平, 因此本实验中的氮添加处理对物种和功能群多样性并没有显著的影响.我们发现, 氮添加提高土壤养分含量, 显著促进多年生丛生禾草、多年生根茎禾草和一二年生植物的生长, 增大其对群落ANPP的贡献.而另一个优势功能群多年生杂类草的生物量主要与多年生根茎禾草此消彼长, 由于多年生根茎禾草的增加, 最终造成多年生杂类草在群落ANPP相对贡献的显著下降; 并且还发现灌木与半灌木的生物量因多年生禾草的增加而减少.Bai等(2004)通过分析本区域草原生产力的长期动态, 提出多年生丛生禾草和多年生根茎禾草两个主要功能群之间的补偿作用是维持半干旱草原生产力稳定性的重要机制.与多年生根茎禾草相比, 多年生丛生禾草(以大针茅、冰草(Agropyron cristatum)等丛生禾草为主)更能忍耐干旱和贫瘠的生境, 又因其生长速率较慢, 因此在较为干旱的年份可以保持相对稳定的生长; 而在较为湿润的年份, 多年生根茎禾草则可以迅速利用水分和养分资源, 实现快速生长.在本研究中, 我们也发现了类似的补偿现象, 实验前3年(2013-2015)雨水较为充足, 特别是在氮添加处理下, 多年生根茎禾草的生物量持续增加; 但在2016年后, 由于降水减少, 多年生根茎禾草的生物量迅速下降, 而多年生丛生禾草仍然维持了较高的生物量.近年来的研究也证实物种间的非同步性是维持草原群落生产力稳定性的重要途径(Yang et al., 2012; Xu et al., 2015; Zhang et al., 2016; Wilcox et al., 2017; Zhang et al., 2018b; Sasaki et al., 2019; Wu & Wang, 2019). ...
草地生产与生态功能合理配置的理论基础与关键技术 1 2016
... 我国天然草地面积约4亿hm2, 是我国面积最大的陆地植被类型, 其中北方温带草原所占比例最大(陈佐忠和汪诗平, 2000; Piao et al., 2009; Fang et al., 2010).由于不合理的开发利用和气候变化, 全国90%的天然草地存在不同程度的退化(白永飞等, 2016).为了治理草原退化, 国家从2003年开始通过一系列管理措施, 改善了草原生态环境(张英俊等, 2013).与放牧和打草等传统草原利用方式相比, 围封禁牧可以使退化草原植被得到迅速恢复, 因而作为最经济有效的草地恢复方式得到推广(Wu et al., 2009; Jing et al., 2013).但也有研究发现长期围封所造成的凋落物大量累积反而会抑制生产力(Rice & Parenti, 1978; Knapp & Seastedt, 1986). ...
草地生产与生态功能合理配置的理论基础与关键技术 1 2016
... 我国天然草地面积约4亿hm2, 是我国面积最大的陆地植被类型, 其中北方温带草原所占比例最大(陈佐忠和汪诗平, 2000; Piao et al., 2009; Fang et al., 2010).由于不合理的开发利用和气候变化, 全国90%的天然草地存在不同程度的退化(白永飞等, 2016).为了治理草原退化, 国家从2003年开始通过一系列管理措施, 改善了草原生态环境(张英俊等, 2013).与放牧和打草等传统草原利用方式相比, 围封禁牧可以使退化草原植被得到迅速恢复, 因而作为最经济有效的草地恢复方式得到推广(Wu et al., 2009; Jing et al., 2013).但也有研究发现长期围封所造成的凋落物大量累积反而会抑制生产力(Rice & Parenti, 1978; Knapp & Seastedt, 1986). ...
Plant litter effects on soil nutrient availability and vegetation dynamics: changes that occur when annual grasses invade shrub- steppe communities 2 2014
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
... 连续6年的实验处理结果表明, 凋落物输入量的增加显著提高了群落ANPP, 而凋落物去除则降低了ANPP (图3B).相关分析表明随着凋落物添加量的增加, 群落ANPP显著提高(图6A).已有研究表明凋落物对ANPP的影响主要分为两个方面, 一方面是通过凋落物自身分解直接增加生态系统碳输入, 提高土壤养分可利用性来促进ANPP (Brearley et al., 2003; Bansal et al., 2014); 另一方面是通过对微环境条件(如土壤温度、水分状况等)的调控来间接影响ANPP (Booth et al., 2005; Deutsch et al., 2010a).在本研究中, 凋落物输入变化并没有显著改变DOC和DON等土壤养分含量, 而是显著改善了土壤水分和温度等土壤环境条件间接影响植被生产力.ANPP与降水量以及土壤含水量的关系表明, 水分是限制群落生产力的主要限制因子, 凋落物添加通过降低土温, 减少水分蒸发, 提高表层土壤水分可利用性, 降低土壤水分的季节变异, 从而降低了植物遭受干旱胁迫风险, 进而促进植物生长, 提高群落ANPP.Xu等(2013a)在关于生态系统中凋落物作用的meta分析中发现, 在草原生态系统的研究中, 凋落物添加会增加土壤微生物碳以及总碳含量, 并降低土壤温度、增加土壤含水量.我们的研究虽然在统计上尚未检测到凋落物输入变化对土壤DOC和DON含量的显著影响, 但还是发现随着凋落物的增加, DOC含量呈现出增加的趋势.这表明凋落物输入变化对土壤DOC等养分可利用性的影响可能需要更长期的实验研究结果来检验(Wang et al., 2017a).尤其在干旱半干旱生态系统中, 因为受到水分条件的限制, 凋落物分解速率相对更慢(Zhang & Wang, 2015). ...
Controls on nitrogen cycling in terrestrial ecosystems: a synthetic analysis of literature data 1 2005
... 连续6年的实验处理结果表明, 凋落物输入量的增加显著提高了群落ANPP, 而凋落物去除则降低了ANPP (图3B).相关分析表明随着凋落物添加量的增加, 群落ANPP显著提高(图6A).已有研究表明凋落物对ANPP的影响主要分为两个方面, 一方面是通过凋落物自身分解直接增加生态系统碳输入, 提高土壤养分可利用性来促进ANPP (Brearley et al., 2003; Bansal et al., 2014); 另一方面是通过对微环境条件(如土壤温度、水分状况等)的调控来间接影响ANPP (Booth et al., 2005; Deutsch et al., 2010a).在本研究中, 凋落物输入变化并没有显著改变DOC和DON等土壤养分含量, 而是显著改善了土壤水分和温度等土壤环境条件间接影响植被生产力.ANPP与降水量以及土壤含水量的关系表明, 水分是限制群落生产力的主要限制因子, 凋落物添加通过降低土温, 减少水分蒸发, 提高表层土壤水分可利用性, 降低土壤水分的季节变异, 从而降低了植物遭受干旱胁迫风险, 进而促进植物生长, 提高群落ANPP.Xu等(2013a)在关于生态系统中凋落物作用的meta分析中发现, 在草原生态系统的研究中, 凋落物添加会增加土壤微生物碳以及总碳含量, 并降低土壤温度、增加土壤含水量.我们的研究虽然在统计上尚未检测到凋落物输入变化对土壤DOC和DON含量的显著影响, 但还是发现随着凋落物的增加, DOC含量呈现出增加的趋势.这表明凋落物输入变化对土壤DOC等养分可利用性的影响可能需要更长期的实验研究结果来检验(Wang et al., 2017a).尤其在干旱半干旱生态系统中, 因为受到水分条件的限制, 凋落物分解速率相对更慢(Zhang & Wang, 2015). ...
Mechanisms underlying the suppression of forb seedling emergence by grass (Poa pratensis) litter 1 1995
Nutrients obtained from leaf litter can improve the growth of dipterocarp seedlings 2 2003
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
... 连续6年的实验处理结果表明, 凋落物输入量的增加显著提高了群落ANPP, 而凋落物去除则降低了ANPP (图3B).相关分析表明随着凋落物添加量的增加, 群落ANPP显著提高(图6A).已有研究表明凋落物对ANPP的影响主要分为两个方面, 一方面是通过凋落物自身分解直接增加生态系统碳输入, 提高土壤养分可利用性来促进ANPP (Brearley et al., 2003; Bansal et al., 2014); 另一方面是通过对微环境条件(如土壤温度、水分状况等)的调控来间接影响ANPP (Booth et al., 2005; Deutsch et al., 2010a).在本研究中, 凋落物输入变化并没有显著改变DOC和DON等土壤养分含量, 而是显著改善了土壤水分和温度等土壤环境条件间接影响植被生产力.ANPP与降水量以及土壤含水量的关系表明, 水分是限制群落生产力的主要限制因子, 凋落物添加通过降低土温, 减少水分蒸发, 提高表层土壤水分可利用性, 降低土壤水分的季节变异, 从而降低了植物遭受干旱胁迫风险, 进而促进植物生长, 提高群落ANPP.Xu等(2013a)在关于生态系统中凋落物作用的meta分析中发现, 在草原生态系统的研究中, 凋落物添加会增加土壤微生物碳以及总碳含量, 并降低土壤温度、增加土壤含水量.我们的研究虽然在统计上尚未检测到凋落物输入变化对土壤DOC和DON含量的显著影响, 但还是发现随着凋落物的增加, DOC含量呈现出增加的趋势.这表明凋落物输入变化对土壤DOC等养分可利用性的影响可能需要更长期的实验研究结果来检验(Wang et al., 2017a).尤其在干旱半干旱生态系统中, 因为受到水分条件的限制, 凋落物分解速率相对更慢(Zhang & Wang, 2015). ...
Response of grassland biomass production to simulated climate change and clipping along an elevation gradient 1 2014
... 我国天然草地面积约4亿hm2, 是我国面积最大的陆地植被类型, 其中北方温带草原所占比例最大(陈佐忠和汪诗平, 2000; Piao et al., 2009; Fang et al., 2010).由于不合理的开发利用和气候变化, 全国90%的天然草地存在不同程度的退化(白永飞等, 2016).为了治理草原退化, 国家从2003年开始通过一系列管理措施, 改善了草原生态环境(张英俊等, 2013).与放牧和打草等传统草原利用方式相比, 围封禁牧可以使退化草原植被得到迅速恢复, 因而作为最经济有效的草地恢复方式得到推广(Wu et al., 2009; Jing et al., 2013).但也有研究发现长期围封所造成的凋落物大量累积反而会抑制生产力(Rice & Parenti, 1978; Knapp & Seastedt, 1986). ...
... 本研究区域位于内蒙古自治区锡林郭勒盟西乌珠穆沁旗, 117.58° E, 44.37° N, 海拔1 148 m, 属典型温带半干旱大陆性气候(陈佐忠和汪诗平, 2000).根据当地气象站长期气象数据(1955-2011年), 本区域年平均气温为1.5 ℃, 年降水量333 mm, 大部分降水(超过87%)发生在生长季(5-9月).降水量年际间波动剧烈, 年降水量最多和最少分别能达到508和118 mm.本区域典型植被类型为温带半干旱草原, 以多年生禾草和多年生杂类草为优势植物, 放牧和打草是本区域最普遍的草原利用方式.土壤类型为栗钙土, 表层土壤(0-10 cm)有机碳含量(14.2 ± 0.4) g·kg-1, 氮含量(1.8 ± 0.1) g·kg-1, pH为7.7 ± 0.5 (王珊珊, 2016; Zhang et al., 2017). ...
中国典型草原生态系统 2 2000
... 我国天然草地面积约4亿hm2, 是我国面积最大的陆地植被类型, 其中北方温带草原所占比例最大(陈佐忠和汪诗平, 2000; Piao et al., 2009; Fang et al., 2010).由于不合理的开发利用和气候变化, 全国90%的天然草地存在不同程度的退化(白永飞等, 2016).为了治理草原退化, 国家从2003年开始通过一系列管理措施, 改善了草原生态环境(张英俊等, 2013).与放牧和打草等传统草原利用方式相比, 围封禁牧可以使退化草原植被得到迅速恢复, 因而作为最经济有效的草地恢复方式得到推广(Wu et al., 2009; Jing et al., 2013).但也有研究发现长期围封所造成的凋落物大量累积反而会抑制生产力(Rice & Parenti, 1978; Knapp & Seastedt, 1986). ...
... 本研究区域位于内蒙古自治区锡林郭勒盟西乌珠穆沁旗, 117.58° E, 44.37° N, 海拔1 148 m, 属典型温带半干旱大陆性气候(陈佐忠和汪诗平, 2000).根据当地气象站长期气象数据(1955-2011年), 本区域年平均气温为1.5 ℃, 年降水量333 mm, 大部分降水(超过87%)发生在生长季(5-9月).降水量年际间波动剧烈, 年降水量最多和最少分别能达到508和118 mm.本区域典型植被类型为温带半干旱草原, 以多年生禾草和多年生杂类草为优势植物, 放牧和打草是本区域最普遍的草原利用方式.土壤类型为栗钙土, 表层土壤(0-10 cm)有机碳含量(14.2 ± 0.4) g·kg-1, 氮含量(1.8 ± 0.1) g·kg-1, pH为7.7 ± 0.5 (王珊珊, 2016; Zhang et al., 2017). ...
Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands 1 2008
... 在大多数陆地生态系统中, 氮是植物生长的主要限制因子(Vitousek & Howarth, 1991; LeBauer & Treseder, 2008; Xia & Wan, 2008; Lu et al., 2011; ?gren et al., 2012; Gao et al., 2019).大量研究表明氮添加会提高植物群落生产力, 但群落植物物种多样性却显著降低(Stevens et al., 2004; Suding et al., 2005; Clark & Tilman, 2008; Isbell et al., 2013).由于植物对氮、磷等养分元素的重吸收作用, 凋落物中的碳氮比相对较高(Deng et al., 2018).日益加剧的大气氮沉降, 会提高环境中活性氮的含量(Gruber & Galloway, 2008).这一方面降低了凋落物中碳氮比, 另一方面缓解了微生物的营养限制, 促进微生物活性, 进而加速凋落物的分解(Austin & Vivanco, 2006; Yue et al., 2016; Wang et al., 2017b), 改善了环境中的养分状况(Manning et al., 2008; Zhang et al., 2018a).那么, 凋落物和氮两者共同作用会对生态系统生产力和群落结构产生怎样的影响呢? 目前还缺乏长期的实验研究. ...
Recovery of plant diversity following N cessation: effects of recruitment, litter, and elevated N cycling 1 2010
... 在我国半干旱草原, 持续大量的氮添加会降低群落生物多样性是已经被大量的实验证实的结论(Stevens et al., 2004; Clark & Tilman, 2010; Zhang et al., 2016; Liu et al., 2018), 但与以往研究不同的是, 本研究经过6年的氮添加处理, 并没有发现物种和功能群多样性发生显著变化, 这可能是与本实验的氮添加量有关.在本研究区域已开展的长期氮添加梯度实验中, 草原生产力和多样性均与氮添加量表现出非线性关系, 呈现先增加后降低的趋势, 而拐点为10-15 g·m-2·a-1, 过量的氮添加会因为土壤酸化和有害离子的富集而导致群落生产力和多样性的降低(Stevens et al., 2004; Tian et al., 2016; Gao et al., 2019).本研究的施氮量为15 g·m-2·a-1, 对该区域草原生态系统是一个比较适宜的氮添加水平, 因此本实验中的氮添加处理对物种和功能群多样性并没有显著的影响.我们发现, 氮添加提高土壤养分含量, 显著促进多年生丛生禾草、多年生根茎禾草和一二年生植物的生长, 增大其对群落ANPP的贡献.而另一个优势功能群多年生杂类草的生物量主要与多年生根茎禾草此消彼长, 由于多年生根茎禾草的增加, 最终造成多年生杂类草在群落ANPP相对贡献的显著下降; 并且还发现灌木与半灌木的生物量因多年生禾草的增加而减少.Bai等(2004)通过分析本区域草原生产力的长期动态, 提出多年生丛生禾草和多年生根茎禾草两个主要功能群之间的补偿作用是维持半干旱草原生产力稳定性的重要机制.与多年生根茎禾草相比, 多年生丛生禾草(以大针茅、冰草(Agropyron cristatum)等丛生禾草为主)更能忍耐干旱和贫瘠的生境, 又因其生长速率较慢, 因此在较为干旱的年份可以保持相对稳定的生长; 而在较为湿润的年份, 多年生根茎禾草则可以迅速利用水分和养分资源, 实现快速生长.在本研究中, 我们也发现了类似的补偿现象, 实验前3年(2013-2015)雨水较为充足, 特别是在氮添加处理下, 多年生根茎禾草的生物量持续增加; 但在2016年后, 由于降水减少, 多年生根茎禾草的生物量迅速下降, 而多年生丛生禾草仍然维持了较高的生物量.近年来的研究也证实物种间的非同步性是维持草原群落生产力稳定性的重要途径(Yang et al., 2012; Xu et al., 2015; Zhang et al., 2016; Wilcox et al., 2017; Zhang et al., 2018b; Sasaki et al., 2019; Wu & Wang, 2019). ...
Ecosystem scale trade-off in nitrogen acquisition pathways 1 2018
... 在大多数陆地生态系统中, 氮是植物生长的主要限制因子(Vitousek & Howarth, 1991; LeBauer & Treseder, 2008; Xia & Wan, 2008; Lu et al., 2011; ?gren et al., 2012; Gao et al., 2019).大量研究表明氮添加会提高植物群落生产力, 但群落植物物种多样性却显著降低(Stevens et al., 2004; Suding et al., 2005; Clark & Tilman, 2008; Isbell et al., 2013).由于植物对氮、磷等养分元素的重吸收作用, 凋落物中的碳氮比相对较高(Deng et al., 2018).日益加剧的大气氮沉降, 会提高环境中活性氮的含量(Gruber & Galloway, 2008).这一方面降低了凋落物中碳氮比, 另一方面缓解了微生物的营养限制, 促进微生物活性, 进而加速凋落物的分解(Austin & Vivanco, 2006; Yue et al., 2016; Wang et al., 2017b), 改善了环境中的养分状况(Manning et al., 2008; Zhang et al., 2018a).那么, 凋落物和氮两者共同作用会对生态系统生产力和群落结构产生怎样的影响呢? 目前还缺乏长期的实验研究. ...
Separation of grassland litter and ecosite influences on seasonal soil moisture and plant growth dynamics 1 2010
... 连续6年的实验处理结果表明, 凋落物输入量的增加显著提高了群落ANPP, 而凋落物去除则降低了ANPP (图3B).相关分析表明随着凋落物添加量的增加, 群落ANPP显著提高(图6A).已有研究表明凋落物对ANPP的影响主要分为两个方面, 一方面是通过凋落物自身分解直接增加生态系统碳输入, 提高土壤养分可利用性来促进ANPP (Brearley et al., 2003; Bansal et al., 2014); 另一方面是通过对微环境条件(如土壤温度、水分状况等)的调控来间接影响ANPP (Booth et al., 2005; Deutsch et al., 2010a).在本研究中, 凋落物输入变化并没有显著改变DOC和DON等土壤养分含量, 而是显著改善了土壤水分和温度等土壤环境条件间接影响植被生产力.ANPP与降水量以及土壤含水量的关系表明, 水分是限制群落生产力的主要限制因子, 凋落物添加通过降低土温, 减少水分蒸发, 提高表层土壤水分可利用性, 降低土壤水分的季节变异, 从而降低了植物遭受干旱胁迫风险, 进而促进植物生长, 提高群落ANPP.Xu等(2013a)在关于生态系统中凋落物作用的meta分析中发现, 在草原生态系统的研究中, 凋落物添加会增加土壤微生物碳以及总碳含量, 并降低土壤温度、增加土壤含水量.我们的研究虽然在统计上尚未检测到凋落物输入变化对土壤DOC和DON含量的显著影响, 但还是发现随着凋落物的增加, DOC含量呈现出增加的趋势.这表明凋落物输入变化对土壤DOC等养分可利用性的影响可能需要更长期的实验研究结果来检验(Wang et al., 2017a).尤其在干旱半干旱生态系统中, 因为受到水分条件的限制, 凋落物分解速率相对更慢(Zhang & Wang, 2015). ...
Soil moisture and plant growth responses to litter and defoliation impacts in Parkland grasslands 2 2010
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
Plant litter: light interception and effects on an old-field plant community 2 1991
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
Plant litter: its dynamics and effects on plant community structure 2 1991
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
... 近年来, 凋落物输入变化对生态系统结构功能的影响更多聚焦在植被生产力、土壤微生物介导的养分循环过程和土壤碳库稳定性的变化上(Xu et al., 2013a), 而对于凋落物输入变化对群落结构影响的报道比较有限(王静, 2011; 蒋红志, 2018).凋落物对群落结构的影响则主要通过其对不同植物功能群的影响差异来实现, 如Facelli和Pickett (1991b)发现凋落物输入增加会抑制草本植物的生长, 其中既发现对外来种生长建群的抑制(Patrick et al., 2008), 也有对原生种生长的抑制, 这主要与凋落物类型有关(Facelli & Pickett, 1991b; Foster & Gross, 1998; Amatangelo et al., 2008; Letts et al., 2015).此外, 凋落物的累积能通过防止种子进入土壤层而对种子补充过程形成抑制, 也会通过化感作用抑制种子萌发, 累积的凋落物也能通过降低物种丰富度和均匀度来改变群落的物种组成(Foster & Gross, 1998; Amatangelo et al., 2008; Letts et al., 2015). ...
Ecosystem carbon stocks and their changes in China?s grasslands 1 2010
... 我国天然草地面积约4亿hm2, 是我国面积最大的陆地植被类型, 其中北方温带草原所占比例最大(陈佐忠和汪诗平, 2000; Piao et al., 2009; Fang et al., 2010).由于不合理的开发利用和气候变化, 全国90%的天然草地存在不同程度的退化(白永飞等, 2016).为了治理草原退化, 国家从2003年开始通过一系列管理措施, 改善了草原生态环境(张英俊等, 2013).与放牧和打草等传统草原利用方式相比, 围封禁牧可以使退化草原植被得到迅速恢复, 因而作为最经济有效的草地恢复方式得到推广(Wu et al., 2009; Jing et al., 2013).但也有研究发现长期围封所造成的凋落物大量累积反而会抑制生产力(Rice & Parenti, 1978; Knapp & Seastedt, 1986). ...
Climate change, human impacts, and carbon sequestration in China 1 2018
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
Species richness in a successional grassland: effects of nitrogen enrichment and plant litter 2 1998
... 近年来, 凋落物输入变化对生态系统结构功能的影响更多聚焦在植被生产力、土壤微生物介导的养分循环过程和土壤碳库稳定性的变化上(Xu et al., 2013a), 而对于凋落物输入变化对群落结构影响的报道比较有限(王静, 2011; 蒋红志, 2018).凋落物对群落结构的影响则主要通过其对不同植物功能群的影响差异来实现, 如Facelli和Pickett (1991b)发现凋落物输入增加会抑制草本植物的生长, 其中既发现对外来种生长建群的抑制(Patrick et al., 2008), 也有对原生种生长的抑制, 这主要与凋落物类型有关(Facelli & Pickett, 1991b; Foster & Gross, 1998; Amatangelo et al., 2008; Letts et al., 2015).此外, 凋落物的累积能通过防止种子进入土壤层而对种子补充过程形成抑制, 也会通过化感作用抑制种子萌发, 累积的凋落物也能通过降低物种丰富度和均匀度来改变群落的物种组成(Foster & Gross, 1998; Amatangelo et al., 2008; Letts et al., 2015). ...
... ).此外, 凋落物的累积能通过防止种子进入土壤层而对种子补充过程形成抑制, 也会通过化感作用抑制种子萌发, 累积的凋落物也能通过降低物种丰富度和均匀度来改变群落的物种组成(Foster & Gross, 1998; Amatangelo et al., 2008; Letts et al., 2015). ...
Nitrogen addition interacted with salinity-alkalinity to modify plant diversity, microbial PLFAs and soil coupled elements: a 5-year experiment 2 2019
... 在大多数陆地生态系统中, 氮是植物生长的主要限制因子(Vitousek & Howarth, 1991; LeBauer & Treseder, 2008; Xia & Wan, 2008; Lu et al., 2011; ?gren et al., 2012; Gao et al., 2019).大量研究表明氮添加会提高植物群落生产力, 但群落植物物种多样性却显著降低(Stevens et al., 2004; Suding et al., 2005; Clark & Tilman, 2008; Isbell et al., 2013).由于植物对氮、磷等养分元素的重吸收作用, 凋落物中的碳氮比相对较高(Deng et al., 2018).日益加剧的大气氮沉降, 会提高环境中活性氮的含量(Gruber & Galloway, 2008).这一方面降低了凋落物中碳氮比, 另一方面缓解了微生物的营养限制, 促进微生物活性, 进而加速凋落物的分解(Austin & Vivanco, 2006; Yue et al., 2016; Wang et al., 2017b), 改善了环境中的养分状况(Manning et al., 2008; Zhang et al., 2018a).那么, 凋落物和氮两者共同作用会对生态系统生产力和群落结构产生怎样的影响呢? 目前还缺乏长期的实验研究. ...
... 在我国半干旱草原, 持续大量的氮添加会降低群落生物多样性是已经被大量的实验证实的结论(Stevens et al., 2004; Clark & Tilman, 2010; Zhang et al., 2016; Liu et al., 2018), 但与以往研究不同的是, 本研究经过6年的氮添加处理, 并没有发现物种和功能群多样性发生显著变化, 这可能是与本实验的氮添加量有关.在本研究区域已开展的长期氮添加梯度实验中, 草原生产力和多样性均与氮添加量表现出非线性关系, 呈现先增加后降低的趋势, 而拐点为10-15 g·m-2·a-1, 过量的氮添加会因为土壤酸化和有害离子的富集而导致群落生产力和多样性的降低(Stevens et al., 2004; Tian et al., 2016; Gao et al., 2019).本研究的施氮量为15 g·m-2·a-1, 对该区域草原生态系统是一个比较适宜的氮添加水平, 因此本实验中的氮添加处理对物种和功能群多样性并没有显著的影响.我们发现, 氮添加提高土壤养分含量, 显著促进多年生丛生禾草、多年生根茎禾草和一二年生植物的生长, 增大其对群落ANPP的贡献.而另一个优势功能群多年生杂类草的生物量主要与多年生根茎禾草此消彼长, 由于多年生根茎禾草的增加, 最终造成多年生杂类草在群落ANPP相对贡献的显著下降; 并且还发现灌木与半灌木的生物量因多年生禾草的增加而减少.Bai等(2004)通过分析本区域草原生产力的长期动态, 提出多年生丛生禾草和多年生根茎禾草两个主要功能群之间的补偿作用是维持半干旱草原生产力稳定性的重要机制.与多年生根茎禾草相比, 多年生丛生禾草(以大针茅、冰草(Agropyron cristatum)等丛生禾草为主)更能忍耐干旱和贫瘠的生境, 又因其生长速率较慢, 因此在较为干旱的年份可以保持相对稳定的生长; 而在较为湿润的年份, 多年生根茎禾草则可以迅速利用水分和养分资源, 实现快速生长.在本研究中, 我们也发现了类似的补偿现象, 实验前3年(2013-2015)雨水较为充足, 特别是在氮添加处理下, 多年生根茎禾草的生物量持续增加; 但在2016年后, 由于降水减少, 多年生根茎禾草的生物量迅速下降, 而多年生丛生禾草仍然维持了较高的生物量.近年来的研究也证实物种间的非同步性是维持草原群落生产力稳定性的重要途径(Yang et al., 2012; Xu et al., 2015; Zhang et al., 2016; Wilcox et al., 2017; Zhang et al., 2018b; Sasaki et al., 2019; Wu & Wang, 2019). ...
An Earth-system perspective of the global nitrogen cycle 1 2008
... 在大多数陆地生态系统中, 氮是植物生长的主要限制因子(Vitousek & Howarth, 1991; LeBauer & Treseder, 2008; Xia & Wan, 2008; Lu et al., 2011; ?gren et al., 2012; Gao et al., 2019).大量研究表明氮添加会提高植物群落生产力, 但群落植物物种多样性却显著降低(Stevens et al., 2004; Suding et al., 2005; Clark & Tilman, 2008; Isbell et al., 2013).由于植物对氮、磷等养分元素的重吸收作用, 凋落物中的碳氮比相对较高(Deng et al., 2018).日益加剧的大气氮沉降, 会提高环境中活性氮的含量(Gruber & Galloway, 2008).这一方面降低了凋落物中碳氮比, 另一方面缓解了微生物的营养限制, 促进微生物活性, 进而加速凋落物的分解(Austin & Vivanco, 2006; Yue et al., 2016; Wang et al., 2017b), 改善了环境中的养分状况(Manning et al., 2008; Zhang et al., 2018a).那么, 凋落物和氮两者共同作用会对生态系统生产力和群落结构产生怎样的影响呢? 目前还缺乏长期的实验研究. ...
Increasing precipitation event size increases aboveground net primary productivity in a semi-arid grassland 1 2008
... 以往的研究表明, 氮添加会显著提高半干旱草原生态系统生产力(潘庆民等, 2005; 沈月, 2016; Yue et al., 2016; Wang et al., 2019; Zhao et al., 2019).本研究也发现, 氮添加显著提高了土壤DOC和DON含量, 促进植物生长, 显著提高了群落生产力.内蒙古半干旱草原作为一个水氮共同限制的生态系统(Heisler-White et al., 2008; Li et al., 2011; Ren et al., 2017).氮对草原生产力的影响强度甚至方向都是依赖于生态系统的水分状况的(Yan et al., 2010; Shen et al., 2016).在本研究中, 氮添加对生产力的促进作用在降水量较高的湿润年份表现得更为明显, 而在较为干旱的年份(2016和2017年), 氮添加对ANPP的促进作用明显减弱(图3A).ANPP与降水量之间的关系也表明, 氮添加显著提高了ANPP-降水量关系的阈值, 随着降水量增加, ANPP增加的趋势更加明显, 这也进一步证实了氮添加处理对水分条件的依赖性(Yang et al., 2011; Zong et al., 2019).此外, 氮添加显著降低了土壤温度, 这主要是由于氮添加促进植物生长, 提高植被盖度, 同时导致第二年凋落物输入量的进一步增多, 对土壤温度调节表现出显著的负反馈作用, 这也有助于缓解干旱年份的水分胁迫作用(Li et al., 2011; Zhao et al., 2019).我们的研究还发现氮添加并未改变凋落物输入变化对群落ANPP的影响, 二者没有显著的交互作用.与Shen等(2016)在本区域进行氮和凋落物添加实验的结果相同.这可能与半干旱草原降水量和生产力剧烈的年际变异有关, 要想进一步探讨这两者之间的交互作用是否会对系统生产力形成影响, 还需要更长期的实验数据积累减弱或去除数据分析过程中年份等其他因素的影响. ...
Nutrient enrichment, biodiversity loss, and consequent declines in ecosystem productivity 1 2013
... 在大多数陆地生态系统中, 氮是植物生长的主要限制因子(Vitousek & Howarth, 1991; LeBauer & Treseder, 2008; Xia & Wan, 2008; Lu et al., 2011; ?gren et al., 2012; Gao et al., 2019).大量研究表明氮添加会提高植物群落生产力, 但群落植物物种多样性却显著降低(Stevens et al., 2004; Suding et al., 2005; Clark & Tilman, 2008; Isbell et al., 2013).由于植物对氮、磷等养分元素的重吸收作用, 凋落物中的碳氮比相对较高(Deng et al., 2018).日益加剧的大气氮沉降, 会提高环境中活性氮的含量(Gruber & Galloway, 2008).这一方面降低了凋落物中碳氮比, 另一方面缓解了微生物的营养限制, 促进微生物活性, 进而加速凋落物的分解(Austin & Vivanco, 2006; Yue et al., 2016; Wang et al., 2017b), 改善了环境中的养分状况(Manning et al., 2008; Zhang et al., 2018a).那么, 凋落物和氮两者共同作用会对生态系统生产力和群落结构产生怎样的影响呢? 目前还缺乏长期的实验研究. ...
Effects of litter on establishment of grassland plant species: the role of seed size and successional status 1 2003
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
1 2018
... 近年来, 凋落物输入变化对生态系统结构功能的影响更多聚焦在植被生产力、土壤微生物介导的养分循环过程和土壤碳库稳定性的变化上(Xu et al., 2013a), 而对于凋落物输入变化对群落结构影响的报道比较有限(王静, 2011; 蒋红志, 2018).凋落物对群落结构的影响则主要通过其对不同植物功能群的影响差异来实现, 如Facelli和Pickett (1991b)发现凋落物输入增加会抑制草本植物的生长, 其中既发现对外来种生长建群的抑制(Patrick et al., 2008), 也有对原生种生长的抑制, 这主要与凋落物类型有关(Facelli & Pickett, 1991b; Foster & Gross, 1998; Amatangelo et al., 2008; Letts et al., 2015).此外, 凋落物的累积能通过防止种子进入土壤层而对种子补充过程形成抑制, 也会通过化感作用抑制种子萌发, 累积的凋落物也能通过降低物种丰富度和均匀度来改变群落的物种组成(Foster & Gross, 1998; Amatangelo et al., 2008; Letts et al., 2015). ...
1 2018
... 近年来, 凋落物输入变化对生态系统结构功能的影响更多聚焦在植被生产力、土壤微生物介导的养分循环过程和土壤碳库稳定性的变化上(Xu et al., 2013a), 而对于凋落物输入变化对群落结构影响的报道比较有限(王静, 2011; 蒋红志, 2018).凋落物对群落结构的影响则主要通过其对不同植物功能群的影响差异来实现, 如Facelli和Pickett (1991b)发现凋落物输入增加会抑制草本植物的生长, 其中既发现对外来种生长建群的抑制(Patrick et al., 2008), 也有对原生种生长的抑制, 这主要与凋落物类型有关(Facelli & Pickett, 1991b; Foster & Gross, 1998; Amatangelo et al., 2008; Letts et al., 2015).此外, 凋落物的累积能通过防止种子进入土壤层而对种子补充过程形成抑制, 也会通过化感作用抑制种子萌发, 累积的凋落物也能通过降低物种丰富度和均匀度来改变群落的物种组成(Foster & Gross, 1998; Amatangelo et al., 2008; Letts et al., 2015). ...
Assessment of vegetative ecological characteristics and the succession process during three decades of grazing exclusion in a continental steppe grassland 1 2013
... 我国天然草地面积约4亿hm2, 是我国面积最大的陆地植被类型, 其中北方温带草原所占比例最大(陈佐忠和汪诗平, 2000; Piao et al., 2009; Fang et al., 2010).由于不合理的开发利用和气候变化, 全国90%的天然草地存在不同程度的退化(白永飞等, 2016).为了治理草原退化, 国家从2003年开始通过一系列管理措施, 改善了草原生态环境(张英俊等, 2013).与放牧和打草等传统草原利用方式相比, 围封禁牧可以使退化草原植被得到迅速恢复, 因而作为最经济有效的草地恢复方式得到推广(Wu et al., 2009; Jing et al., 2013).但也有研究发现长期围封所造成的凋落物大量累积反而会抑制生产力(Rice & Parenti, 1978; Knapp & Seastedt, 1986). ...
Experimental evaluation of methods to quantify dissolved organic nitrogen (DON) and dissolved organic carbon (DOC) in soil 1 2006
Mechanisms shaping plant biomass and species richness: plant strategies and litter effect in alkali and loess grasslands 1 2013
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
Detritus accumulation limits productivity of tallgrass prairie 1 1986
... 我国天然草地面积约4亿hm2, 是我国面积最大的陆地植被类型, 其中北方温带草原所占比例最大(陈佐忠和汪诗平, 2000; Piao et al., 2009; Fang et al., 2010).由于不合理的开发利用和气候变化, 全国90%的天然草地存在不同程度的退化(白永飞等, 2016).为了治理草原退化, 国家从2003年开始通过一系列管理措施, 改善了草原生态环境(张英俊等, 2013).与放牧和打草等传统草原利用方式相比, 围封禁牧可以使退化草原植被得到迅速恢复, 因而作为最经济有效的草地恢复方式得到推广(Wu et al., 2009; Jing et al., 2013).但也有研究发现长期围封所造成的凋落物大量累积反而会抑制生产力(Rice & Parenti, 1978; Knapp & Seastedt, 1986). ...
Legume proportion in grassland litter affects decomposition dynamics and nutrient mineralization 1 2019
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
Direct and indirect control of grassland community structure by litter, resources, and biomass 1 2008
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed 1 2008
... 在大多数陆地生态系统中, 氮是植物生长的主要限制因子(Vitousek & Howarth, 1991; LeBauer & Treseder, 2008; Xia & Wan, 2008; Lu et al., 2011; ?gren et al., 2012; Gao et al., 2019).大量研究表明氮添加会提高植物群落生产力, 但群落植物物种多样性却显著降低(Stevens et al., 2004; Suding et al., 2005; Clark & Tilman, 2008; Isbell et al., 2013).由于植物对氮、磷等养分元素的重吸收作用, 凋落物中的碳氮比相对较高(Deng et al., 2018).日益加剧的大气氮沉降, 会提高环境中活性氮的含量(Gruber & Galloway, 2008).这一方面降低了凋落物中碳氮比, 另一方面缓解了微生物的营养限制, 促进微生物活性, 进而加速凋落物的分解(Austin & Vivanco, 2006; Yue et al., 2016; Wang et al., 2017b), 改善了环境中的养分状况(Manning et al., 2008; Zhang et al., 2018a).那么, 凋落物和氮两者共同作用会对生态系统生产力和群落结构产生怎样的影响呢? 目前还缺乏长期的实验研究. ...
Litter accumulation drives grassland plant community composition and functional diversity via leaf traits 2 2015
... 近年来, 凋落物输入变化对生态系统结构功能的影响更多聚焦在植被生产力、土壤微生物介导的养分循环过程和土壤碳库稳定性的变化上(Xu et al., 2013a), 而对于凋落物输入变化对群落结构影响的报道比较有限(王静, 2011; 蒋红志, 2018).凋落物对群落结构的影响则主要通过其对不同植物功能群的影响差异来实现, 如Facelli和Pickett (1991b)发现凋落物输入增加会抑制草本植物的生长, 其中既发现对外来种生长建群的抑制(Patrick et al., 2008), 也有对原生种生长的抑制, 这主要与凋落物类型有关(Facelli & Pickett, 1991b; Foster & Gross, 1998; Amatangelo et al., 2008; Letts et al., 2015).此外, 凋落物的累积能通过防止种子进入土壤层而对种子补充过程形成抑制, 也会通过化感作用抑制种子萌发, 累积的凋落物也能通过降低物种丰富度和均匀度来改变群落的物种组成(Foster & Gross, 1998; Amatangelo et al., 2008; Letts et al., 2015). ...
... ; Letts et al., 2015). ...
Above and belowground net primary productivity of grassland influenced by supplemental water and nitrogen in Inner Mongolia 2 2011
... 以往的研究表明, 氮添加会显著提高半干旱草原生态系统生产力(潘庆民等, 2005; 沈月, 2016; Yue et al., 2016; Wang et al., 2019; Zhao et al., 2019).本研究也发现, 氮添加显著提高了土壤DOC和DON含量, 促进植物生长, 显著提高了群落生产力.内蒙古半干旱草原作为一个水氮共同限制的生态系统(Heisler-White et al., 2008; Li et al., 2011; Ren et al., 2017).氮对草原生产力的影响强度甚至方向都是依赖于生态系统的水分状况的(Yan et al., 2010; Shen et al., 2016).在本研究中, 氮添加对生产力的促进作用在降水量较高的湿润年份表现得更为明显, 而在较为干旱的年份(2016和2017年), 氮添加对ANPP的促进作用明显减弱(图3A).ANPP与降水量之间的关系也表明, 氮添加显著提高了ANPP-降水量关系的阈值, 随着降水量增加, ANPP增加的趋势更加明显, 这也进一步证实了氮添加处理对水分条件的依赖性(Yang et al., 2011; Zong et al., 2019).此外, 氮添加显著降低了土壤温度, 这主要是由于氮添加促进植物生长, 提高植被盖度, 同时导致第二年凋落物输入量的进一步增多, 对土壤温度调节表现出显著的负反馈作用, 这也有助于缓解干旱年份的水分胁迫作用(Li et al., 2011; Zhao et al., 2019).我们的研究还发现氮添加并未改变凋落物输入变化对群落ANPP的影响, 二者没有显著的交互作用.与Shen等(2016)在本区域进行氮和凋落物添加实验的结果相同.这可能与半干旱草原降水量和生产力剧烈的年际变异有关, 要想进一步探讨这两者之间的交互作用是否会对系统生产力形成影响, 还需要更长期的实验数据积累减弱或去除数据分析过程中年份等其他因素的影响. ...
... ).此外, 氮添加显著降低了土壤温度, 这主要是由于氮添加促进植物生长, 提高植被盖度, 同时导致第二年凋落物输入量的进一步增多, 对土壤温度调节表现出显著的负反馈作用, 这也有助于缓解干旱年份的水分胁迫作用(Li et al., 2011; Zhao et al., 2019).我们的研究还发现氮添加并未改变凋落物输入变化对群落ANPP的影响, 二者没有显著的交互作用.与Shen等(2016)在本区域进行氮和凋落物添加实验的结果相同.这可能与半干旱草原降水量和生产力剧烈的年际变异有关, 要想进一步探讨这两者之间的交互作用是否会对系统生产力形成影响, 还需要更长期的实验数据积累减弱或去除数据分析过程中年份等其他因素的影响. ...
Reversal of nitrogen-induced species diversity declines mediated by change in dominant grass and litter 1 2018
... 在我国半干旱草原, 持续大量的氮添加会降低群落生物多样性是已经被大量的实验证实的结论(Stevens et al., 2004; Clark & Tilman, 2010; Zhang et al., 2016; Liu et al., 2018), 但与以往研究不同的是, 本研究经过6年的氮添加处理, 并没有发现物种和功能群多样性发生显著变化, 这可能是与本实验的氮添加量有关.在本研究区域已开展的长期氮添加梯度实验中, 草原生产力和多样性均与氮添加量表现出非线性关系, 呈现先增加后降低的趋势, 而拐点为10-15 g·m-2·a-1, 过量的氮添加会因为土壤酸化和有害离子的富集而导致群落生产力和多样性的降低(Stevens et al., 2004; Tian et al., 2016; Gao et al., 2019).本研究的施氮量为15 g·m-2·a-1, 对该区域草原生态系统是一个比较适宜的氮添加水平, 因此本实验中的氮添加处理对物种和功能群多样性并没有显著的影响.我们发现, 氮添加提高土壤养分含量, 显著促进多年生丛生禾草、多年生根茎禾草和一二年生植物的生长, 增大其对群落ANPP的贡献.而另一个优势功能群多年生杂类草的生物量主要与多年生根茎禾草此消彼长, 由于多年生根茎禾草的增加, 最终造成多年生杂类草在群落ANPP相对贡献的显著下降; 并且还发现灌木与半灌木的生物量因多年生禾草的增加而减少.Bai等(2004)通过分析本区域草原生产力的长期动态, 提出多年生丛生禾草和多年生根茎禾草两个主要功能群之间的补偿作用是维持半干旱草原生产力稳定性的重要机制.与多年生根茎禾草相比, 多年生丛生禾草(以大针茅、冰草(Agropyron cristatum)等丛生禾草为主)更能忍耐干旱和贫瘠的生境, 又因其生长速率较慢, 因此在较为干旱的年份可以保持相对稳定的生长; 而在较为湿润的年份, 多年生根茎禾草则可以迅速利用水分和养分资源, 实现快速生长.在本研究中, 我们也发现了类似的补偿现象, 实验前3年(2013-2015)雨水较为充足, 特别是在氮添加处理下, 多年生根茎禾草的生物量持续增加; 但在2016年后, 由于降水减少, 多年生根茎禾草的生物量迅速下降, 而多年生丛生禾草仍然维持了较高的生物量.近年来的研究也证实物种间的非同步性是维持草原群落生产力稳定性的重要途径(Yang et al., 2012; Xu et al., 2015; Zhang et al., 2016; Wilcox et al., 2017; Zhang et al., 2018b; Sasaki et al., 2019; Wu & Wang, 2019). ...
Minor stimulation of soil carbon storage by nitrogen addition: a meta-analysis. Agriculture, 1 2011
... 在大多数陆地生态系统中, 氮是植物生长的主要限制因子(Vitousek & Howarth, 1991; LeBauer & Treseder, 2008; Xia & Wan, 2008; Lu et al., 2011; ?gren et al., 2012; Gao et al., 2019).大量研究表明氮添加会提高植物群落生产力, 但群落植物物种多样性却显著降低(Stevens et al., 2004; Suding et al., 2005; Clark & Tilman, 2008; Isbell et al., 2013).由于植物对氮、磷等养分元素的重吸收作用, 凋落物中的碳氮比相对较高(Deng et al., 2018).日益加剧的大气氮沉降, 会提高环境中活性氮的含量(Gruber & Galloway, 2008).这一方面降低了凋落物中碳氮比, 另一方面缓解了微生物的营养限制, 促进微生物活性, 进而加速凋落物的分解(Austin & Vivanco, 2006; Yue et al., 2016; Wang et al., 2017b), 改善了环境中的养分状况(Manning et al., 2008; Zhang et al., 2018a).那么, 凋落物和氮两者共同作用会对生态系统生产力和群落结构产生怎样的影响呢? 目前还缺乏长期的实验研究. ...
Direct and indirect effects of nitrogen deposition on litter decomposition 1 2008
... 在大多数陆地生态系统中, 氮是植物生长的主要限制因子(Vitousek & Howarth, 1991; LeBauer & Treseder, 2008; Xia & Wan, 2008; Lu et al., 2011; ?gren et al., 2012; Gao et al., 2019).大量研究表明氮添加会提高植物群落生产力, 但群落植物物种多样性却显著降低(Stevens et al., 2004; Suding et al., 2005; Clark & Tilman, 2008; Isbell et al., 2013).由于植物对氮、磷等养分元素的重吸收作用, 凋落物中的碳氮比相对较高(Deng et al., 2018).日益加剧的大气氮沉降, 会提高环境中活性氮的含量(Gruber & Galloway, 2008).这一方面降低了凋落物中碳氮比, 另一方面缓解了微生物的营养限制, 促进微生物活性, 进而加速凋落物的分解(Austin & Vivanco, 2006; Yue et al., 2016; Wang et al., 2017b), 改善了环境中的养分状况(Manning et al., 2008; Zhang et al., 2018a).那么, 凋落物和氮两者共同作用会对生态系统生产力和群落结构产生怎样的影响呢? 目前还缺乏长期的实验研究. ...
氮素对内蒙古典型草原羊草种群的影响 2 2005
... 以往的研究表明, 氮添加会显著提高半干旱草原生态系统生产力(潘庆民等, 2005; 沈月, 2016; Yue et al., 2016; Wang et al., 2019; Zhao et al., 2019).本研究也发现, 氮添加显著提高了土壤DOC和DON含量, 促进植物生长, 显著提高了群落生产力.内蒙古半干旱草原作为一个水氮共同限制的生态系统(Heisler-White et al., 2008; Li et al., 2011; Ren et al., 2017).氮对草原生产力的影响强度甚至方向都是依赖于生态系统的水分状况的(Yan et al., 2010; Shen et al., 2016).在本研究中, 氮添加对生产力的促进作用在降水量较高的湿润年份表现得更为明显, 而在较为干旱的年份(2016和2017年), 氮添加对ANPP的促进作用明显减弱(图3A).ANPP与降水量之间的关系也表明, 氮添加显著提高了ANPP-降水量关系的阈值, 随着降水量增加, ANPP增加的趋势更加明显, 这也进一步证实了氮添加处理对水分条件的依赖性(Yang et al., 2011; Zong et al., 2019).此外, 氮添加显著降低了土壤温度, 这主要是由于氮添加促进植物生长, 提高植被盖度, 同时导致第二年凋落物输入量的进一步增多, 对土壤温度调节表现出显著的负反馈作用, 这也有助于缓解干旱年份的水分胁迫作用(Li et al., 2011; Zhao et al., 2019).我们的研究还发现氮添加并未改变凋落物输入变化对群落ANPP的影响, 二者没有显著的交互作用.与Shen等(2016)在本区域进行氮和凋落物添加实验的结果相同.这可能与半干旱草原降水量和生产力剧烈的年际变异有关, 要想进一步探讨这两者之间的交互作用是否会对系统生产力形成影响, 还需要更长期的实验数据积累减弱或去除数据分析过程中年份等其他因素的影响. ...
... 以往的研究表明, 氮添加会显著提高半干旱草原生态系统生产力(潘庆民等, 2005; 沈月, 2016; Yue et al., 2016; Wang et al., 2019; Zhao et al., 2019).本研究也发现, 氮添加显著提高了土壤DOC和DON含量, 促进植物生长, 显著提高了群落生产力.内蒙古半干旱草原作为一个水氮共同限制的生态系统(Heisler-White et al., 2008; Li et al., 2011; Ren et al., 2017).氮对草原生产力的影响强度甚至方向都是依赖于生态系统的水分状况的(Yan et al., 2010; Shen et al., 2016).在本研究中, 氮添加对生产力的促进作用在降水量较高的湿润年份表现得更为明显, 而在较为干旱的年份(2016和2017年), 氮添加对ANPP的促进作用明显减弱(图3A).ANPP与降水量之间的关系也表明, 氮添加显著提高了ANPP-降水量关系的阈值, 随着降水量增加, ANPP增加的趋势更加明显, 这也进一步证实了氮添加处理对水分条件的依赖性(Yang et al., 2011; Zong et al., 2019).此外, 氮添加显著降低了土壤温度, 这主要是由于氮添加促进植物生长, 提高植被盖度, 同时导致第二年凋落物输入量的进一步增多, 对土壤温度调节表现出显著的负反馈作用, 这也有助于缓解干旱年份的水分胁迫作用(Li et al., 2011; Zhao et al., 2019).我们的研究还发现氮添加并未改变凋落物输入变化对群落ANPP的影响, 二者没有显著的交互作用.与Shen等(2016)在本区域进行氮和凋落物添加实验的结果相同.这可能与半干旱草原降水量和生产力剧烈的年际变异有关, 要想进一步探讨这两者之间的交互作用是否会对系统生产力形成影响, 还需要更长期的实验数据积累减弱或去除数据分析过程中年份等其他因素的影响. ...
Large-scale manipulation of plant litter and fertilizer in a managed successional temperate grassland 2 2008
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
... 近年来, 凋落物输入变化对生态系统结构功能的影响更多聚焦在植被生产力、土壤微生物介导的养分循环过程和土壤碳库稳定性的变化上(Xu et al., 2013a), 而对于凋落物输入变化对群落结构影响的报道比较有限(王静, 2011; 蒋红志, 2018).凋落物对群落结构的影响则主要通过其对不同植物功能群的影响差异来实现, 如Facelli和Pickett (1991b)发现凋落物输入增加会抑制草本植物的生长, 其中既发现对外来种生长建群的抑制(Patrick et al., 2008), 也有对原生种生长的抑制, 这主要与凋落物类型有关(Facelli & Pickett, 1991b; Foster & Gross, 1998; Amatangelo et al., 2008; Letts et al., 2015).此外, 凋落物的累积能通过防止种子进入土壤层而对种子补充过程形成抑制, 也会通过化感作用抑制种子萌发, 累积的凋落物也能通过降低物种丰富度和均匀度来改变群落的物种组成(Foster & Gross, 1998; Amatangelo et al., 2008; Letts et al., 2015). ...
The carbon balance of terrestrial ecosystems in China 1 2009
... 我国天然草地面积约4亿hm2, 是我国面积最大的陆地植被类型, 其中北方温带草原所占比例最大(陈佐忠和汪诗平, 2000; Piao et al., 2009; Fang et al., 2010).由于不合理的开发利用和气候变化, 全国90%的天然草地存在不同程度的退化(白永飞等, 2016).为了治理草原退化, 国家从2003年开始通过一系列管理措施, 改善了草原生态环境(张英俊等, 2013).与放牧和打草等传统草原利用方式相比, 围封禁牧可以使退化草原植被得到迅速恢复, 因而作为最经济有效的草地恢复方式得到推广(Wu et al., 2009; Jing et al., 2013).但也有研究发现长期围封所造成的凋落物大量累积反而会抑制生产力(Rice & Parenti, 1978; Knapp & Seastedt, 1986). ...
Exacerbated nitrogen limitation ends transient stimulation of grassland productivity by increased precipitation 1 2017
... 以往的研究表明, 氮添加会显著提高半干旱草原生态系统生产力(潘庆民等, 2005; 沈月, 2016; Yue et al., 2016; Wang et al., 2019; Zhao et al., 2019).本研究也发现, 氮添加显著提高了土壤DOC和DON含量, 促进植物生长, 显著提高了群落生产力.内蒙古半干旱草原作为一个水氮共同限制的生态系统(Heisler-White et al., 2008; Li et al., 2011; Ren et al., 2017).氮对草原生产力的影响强度甚至方向都是依赖于生态系统的水分状况的(Yan et al., 2010; Shen et al., 2016).在本研究中, 氮添加对生产力的促进作用在降水量较高的湿润年份表现得更为明显, 而在较为干旱的年份(2016和2017年), 氮添加对ANPP的促进作用明显减弱(图3A).ANPP与降水量之间的关系也表明, 氮添加显著提高了ANPP-降水量关系的阈值, 随着降水量增加, ANPP增加的趋势更加明显, 这也进一步证实了氮添加处理对水分条件的依赖性(Yang et al., 2011; Zong et al., 2019).此外, 氮添加显著降低了土壤温度, 这主要是由于氮添加促进植物生长, 提高植被盖度, 同时导致第二年凋落物输入量的进一步增多, 对土壤温度调节表现出显著的负反馈作用, 这也有助于缓解干旱年份的水分胁迫作用(Li et al., 2011; Zhao et al., 2019).我们的研究还发现氮添加并未改变凋落物输入变化对群落ANPP的影响, 二者没有显著的交互作用.与Shen等(2016)在本区域进行氮和凋落物添加实验的结果相同.这可能与半干旱草原降水量和生产力剧烈的年际变异有关, 要想进一步探讨这两者之间的交互作用是否会对系统生产力形成影响, 还需要更长期的实验数据积累减弱或去除数据分析过程中年份等其他因素的影响. ...
Causes of decreases in productivity in undisturbed tall grass prairie 1 1978
... 我国天然草地面积约4亿hm2, 是我国面积最大的陆地植被类型, 其中北方温带草原所占比例最大(陈佐忠和汪诗平, 2000; Piao et al., 2009; Fang et al., 2010).由于不合理的开发利用和气候变化, 全国90%的天然草地存在不同程度的退化(白永飞等, 2016).为了治理草原退化, 国家从2003年开始通过一系列管理措施, 改善了草原生态环境(张英俊等, 2013).与放牧和打草等传统草原利用方式相比, 围封禁牧可以使退化草原植被得到迅速恢复, 因而作为最经济有效的草地恢复方式得到推广(Wu et al., 2009; Jing et al., 2013).但也有研究发现长期围封所造成的凋落物大量累积反而会抑制生产力(Rice & Parenti, 1978; Knapp & Seastedt, 1986). ...
Restorative removal of plant litter and vegetation 40 years after abandonment enhances re-emergence of steppe grassland vegetation 1 2010
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
Species asynchrony and response diversity determine multifunctional stability of natural grasslands 1 2019
... 在我国半干旱草原, 持续大量的氮添加会降低群落生物多样性是已经被大量的实验证实的结论(Stevens et al., 2004; Clark & Tilman, 2010; Zhang et al., 2016; Liu et al., 2018), 但与以往研究不同的是, 本研究经过6年的氮添加处理, 并没有发现物种和功能群多样性发生显著变化, 这可能是与本实验的氮添加量有关.在本研究区域已开展的长期氮添加梯度实验中, 草原生产力和多样性均与氮添加量表现出非线性关系, 呈现先增加后降低的趋势, 而拐点为10-15 g·m-2·a-1, 过量的氮添加会因为土壤酸化和有害离子的富集而导致群落生产力和多样性的降低(Stevens et al., 2004; Tian et al., 2016; Gao et al., 2019).本研究的施氮量为15 g·m-2·a-1, 对该区域草原生态系统是一个比较适宜的氮添加水平, 因此本实验中的氮添加处理对物种和功能群多样性并没有显著的影响.我们发现, 氮添加提高土壤养分含量, 显著促进多年生丛生禾草、多年生根茎禾草和一二年生植物的生长, 增大其对群落ANPP的贡献.而另一个优势功能群多年生杂类草的生物量主要与多年生根茎禾草此消彼长, 由于多年生根茎禾草的增加, 最终造成多年生杂类草在群落ANPP相对贡献的显著下降; 并且还发现灌木与半灌木的生物量因多年生禾草的增加而减少.Bai等(2004)通过分析本区域草原生产力的长期动态, 提出多年生丛生禾草和多年生根茎禾草两个主要功能群之间的补偿作用是维持半干旱草原生产力稳定性的重要机制.与多年生根茎禾草相比, 多年生丛生禾草(以大针茅、冰草(Agropyron cristatum)等丛生禾草为主)更能忍耐干旱和贫瘠的生境, 又因其生长速率较慢, 因此在较为干旱的年份可以保持相对稳定的生长; 而在较为湿润的年份, 多年生根茎禾草则可以迅速利用水分和养分资源, 实现快速生长.在本研究中, 我们也发现了类似的补偿现象, 实验前3年(2013-2015)雨水较为充足, 特别是在氮添加处理下, 多年生根茎禾草的生物量持续增加; 但在2016年后, 由于降水减少, 多年生根茎禾草的生物量迅速下降, 而多年生丛生禾草仍然维持了较高的生物量.近年来的研究也证实物种间的非同步性是维持草原群落生产力稳定性的重要途径(Yang et al., 2012; Xu et al., 2015; Zhang et al., 2016; Wilcox et al., 2017; Zhang et al., 2018b; Sasaki et al., 2019; Wu & Wang, 2019). ...
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
3 2016
... 以往的研究表明, 氮添加会显著提高半干旱草原生态系统生产力(潘庆民等, 2005; 沈月, 2016; Yue et al., 2016; Wang et al., 2019; Zhao et al., 2019).本研究也发现, 氮添加显著提高了土壤DOC和DON含量, 促进植物生长, 显著提高了群落生产力.内蒙古半干旱草原作为一个水氮共同限制的生态系统(Heisler-White et al., 2008; Li et al., 2011; Ren et al., 2017).氮对草原生产力的影响强度甚至方向都是依赖于生态系统的水分状况的(Yan et al., 2010; Shen et al., 2016).在本研究中, 氮添加对生产力的促进作用在降水量较高的湿润年份表现得更为明显, 而在较为干旱的年份(2016和2017年), 氮添加对ANPP的促进作用明显减弱(图3A).ANPP与降水量之间的关系也表明, 氮添加显著提高了ANPP-降水量关系的阈值, 随着降水量增加, ANPP增加的趋势更加明显, 这也进一步证实了氮添加处理对水分条件的依赖性(Yang et al., 2011; Zong et al., 2019).此外, 氮添加显著降低了土壤温度, 这主要是由于氮添加促进植物生长, 提高植被盖度, 同时导致第二年凋落物输入量的进一步增多, 对土壤温度调节表现出显著的负反馈作用, 这也有助于缓解干旱年份的水分胁迫作用(Li et al., 2011; Zhao et al., 2019).我们的研究还发现氮添加并未改变凋落物输入变化对群落ANPP的影响, 二者没有显著的交互作用.与Shen等(2016)在本区域进行氮和凋落物添加实验的结果相同.这可能与半干旱草原降水量和生产力剧烈的年际变异有关, 要想进一步探讨这两者之间的交互作用是否会对系统生产力形成影响, 还需要更长期的实验数据积累减弱或去除数据分析过程中年份等其他因素的影响. ...
... 以往的研究表明, 氮添加会显著提高半干旱草原生态系统生产力(潘庆民等, 2005; 沈月, 2016; Yue et al., 2016; Wang et al., 2019; Zhao et al., 2019).本研究也发现, 氮添加显著提高了土壤DOC和DON含量, 促进植物生长, 显著提高了群落生产力.内蒙古半干旱草原作为一个水氮共同限制的生态系统(Heisler-White et al., 2008; Li et al., 2011; Ren et al., 2017).氮对草原生产力的影响强度甚至方向都是依赖于生态系统的水分状况的(Yan et al., 2010; Shen et al., 2016).在本研究中, 氮添加对生产力的促进作用在降水量较高的湿润年份表现得更为明显, 而在较为干旱的年份(2016和2017年), 氮添加对ANPP的促进作用明显减弱(图3A).ANPP与降水量之间的关系也表明, 氮添加显著提高了ANPP-降水量关系的阈值, 随着降水量增加, ANPP增加的趋势更加明显, 这也进一步证实了氮添加处理对水分条件的依赖性(Yang et al., 2011; Zong et al., 2019).此外, 氮添加显著降低了土壤温度, 这主要是由于氮添加促进植物生长, 提高植被盖度, 同时导致第二年凋落物输入量的进一步增多, 对土壤温度调节表现出显著的负反馈作用, 这也有助于缓解干旱年份的水分胁迫作用(Li et al., 2011; Zhao et al., 2019).我们的研究还发现氮添加并未改变凋落物输入变化对群落ANPP的影响, 二者没有显著的交互作用.与Shen等(2016)在本区域进行氮和凋落物添加实验的结果相同.这可能与半干旱草原降水量和生产力剧烈的年际变异有关, 要想进一步探讨这两者之间的交互作用是否会对系统生产力形成影响, 还需要更长期的实验数据积累减弱或去除数据分析过程中年份等其他因素的影响. ...
Can litter addition mediate plant productivity responses to increased precipitation and nitrogen deposition in a typical steppe? 1 2016
... 以往的研究表明, 氮添加会显著提高半干旱草原生态系统生产力(潘庆民等, 2005; 沈月, 2016; Yue et al., 2016; Wang et al., 2019; Zhao et al., 2019).本研究也发现, 氮添加显著提高了土壤DOC和DON含量, 促进植物生长, 显著提高了群落生产力.内蒙古半干旱草原作为一个水氮共同限制的生态系统(Heisler-White et al., 2008; Li et al., 2011; Ren et al., 2017).氮对草原生产力的影响强度甚至方向都是依赖于生态系统的水分状况的(Yan et al., 2010; Shen et al., 2016).在本研究中, 氮添加对生产力的促进作用在降水量较高的湿润年份表现得更为明显, 而在较为干旱的年份(2016和2017年), 氮添加对ANPP的促进作用明显减弱(图3A).ANPP与降水量之间的关系也表明, 氮添加显著提高了ANPP-降水量关系的阈值, 随着降水量增加, ANPP增加的趋势更加明显, 这也进一步证实了氮添加处理对水分条件的依赖性(Yang et al., 2011; Zong et al., 2019).此外, 氮添加显著降低了土壤温度, 这主要是由于氮添加促进植物生长, 提高植被盖度, 同时导致第二年凋落物输入量的进一步增多, 对土壤温度调节表现出显著的负反馈作用, 这也有助于缓解干旱年份的水分胁迫作用(Li et al., 2011; Zhao et al., 2019).我们的研究还发现氮添加并未改变凋落物输入变化对群落ANPP的影响, 二者没有显著的交互作用.与Shen等(2016)在本区域进行氮和凋落物添加实验的结果相同.这可能与半干旱草原降水量和生产力剧烈的年际变异有关, 要想进一步探讨这两者之间的交互作用是否会对系统生产力形成影响, 还需要更长期的实验数据积累减弱或去除数据分析过程中年份等其他因素的影响. ...
Impact of nitrogen deposition on the species richness of grasslands 3 2004
... 在大多数陆地生态系统中, 氮是植物生长的主要限制因子(Vitousek & Howarth, 1991; LeBauer & Treseder, 2008; Xia & Wan, 2008; Lu et al., 2011; ?gren et al., 2012; Gao et al., 2019).大量研究表明氮添加会提高植物群落生产力, 但群落植物物种多样性却显著降低(Stevens et al., 2004; Suding et al., 2005; Clark & Tilman, 2008; Isbell et al., 2013).由于植物对氮、磷等养分元素的重吸收作用, 凋落物中的碳氮比相对较高(Deng et al., 2018).日益加剧的大气氮沉降, 会提高环境中活性氮的含量(Gruber & Galloway, 2008).这一方面降低了凋落物中碳氮比, 另一方面缓解了微生物的营养限制, 促进微生物活性, 进而加速凋落物的分解(Austin & Vivanco, 2006; Yue et al., 2016; Wang et al., 2017b), 改善了环境中的养分状况(Manning et al., 2008; Zhang et al., 2018a).那么, 凋落物和氮两者共同作用会对生态系统生产力和群落结构产生怎样的影响呢? 目前还缺乏长期的实验研究. ...
... 在我国半干旱草原, 持续大量的氮添加会降低群落生物多样性是已经被大量的实验证实的结论(Stevens et al., 2004; Clark & Tilman, 2010; Zhang et al., 2016; Liu et al., 2018), 但与以往研究不同的是, 本研究经过6年的氮添加处理, 并没有发现物种和功能群多样性发生显著变化, 这可能是与本实验的氮添加量有关.在本研究区域已开展的长期氮添加梯度实验中, 草原生产力和多样性均与氮添加量表现出非线性关系, 呈现先增加后降低的趋势, 而拐点为10-15 g·m-2·a-1, 过量的氮添加会因为土壤酸化和有害离子的富集而导致群落生产力和多样性的降低(Stevens et al., 2004; Tian et al., 2016; Gao et al., 2019).本研究的施氮量为15 g·m-2·a-1, 对该区域草原生态系统是一个比较适宜的氮添加水平, 因此本实验中的氮添加处理对物种和功能群多样性并没有显著的影响.我们发现, 氮添加提高土壤养分含量, 显著促进多年生丛生禾草、多年生根茎禾草和一二年生植物的生长, 增大其对群落ANPP的贡献.而另一个优势功能群多年生杂类草的生物量主要与多年生根茎禾草此消彼长, 由于多年生根茎禾草的增加, 最终造成多年生杂类草在群落ANPP相对贡献的显著下降; 并且还发现灌木与半灌木的生物量因多年生禾草的增加而减少.Bai等(2004)通过分析本区域草原生产力的长期动态, 提出多年生丛生禾草和多年生根茎禾草两个主要功能群之间的补偿作用是维持半干旱草原生产力稳定性的重要机制.与多年生根茎禾草相比, 多年生丛生禾草(以大针茅、冰草(Agropyron cristatum)等丛生禾草为主)更能忍耐干旱和贫瘠的生境, 又因其生长速率较慢, 因此在较为干旱的年份可以保持相对稳定的生长; 而在较为湿润的年份, 多年生根茎禾草则可以迅速利用水分和养分资源, 实现快速生长.在本研究中, 我们也发现了类似的补偿现象, 实验前3年(2013-2015)雨水较为充足, 特别是在氮添加处理下, 多年生根茎禾草的生物量持续增加; 但在2016年后, 由于降水减少, 多年生根茎禾草的生物量迅速下降, 而多年生丛生禾草仍然维持了较高的生物量.近年来的研究也证实物种间的非同步性是维持草原群落生产力稳定性的重要途径(Yang et al., 2012; Xu et al., 2015; Zhang et al., 2016; Wilcox et al., 2017; Zhang et al., 2018b; Sasaki et al., 2019; Wu & Wang, 2019). ...
... , 过量的氮添加会因为土壤酸化和有害离子的富集而导致群落生产力和多样性的降低(Stevens et al., 2004; Tian et al., 2016; Gao et al., 2019).本研究的施氮量为15 g·m-2·a-1, 对该区域草原生态系统是一个比较适宜的氮添加水平, 因此本实验中的氮添加处理对物种和功能群多样性并没有显著的影响.我们发现, 氮添加提高土壤养分含量, 显著促进多年生丛生禾草、多年生根茎禾草和一二年生植物的生长, 增大其对群落ANPP的贡献.而另一个优势功能群多年生杂类草的生物量主要与多年生根茎禾草此消彼长, 由于多年生根茎禾草的增加, 最终造成多年生杂类草在群落ANPP相对贡献的显著下降; 并且还发现灌木与半灌木的生物量因多年生禾草的增加而减少.Bai等(2004)通过分析本区域草原生产力的长期动态, 提出多年生丛生禾草和多年生根茎禾草两个主要功能群之间的补偿作用是维持半干旱草原生产力稳定性的重要机制.与多年生根茎禾草相比, 多年生丛生禾草(以大针茅、冰草(Agropyron cristatum)等丛生禾草为主)更能忍耐干旱和贫瘠的生境, 又因其生长速率较慢, 因此在较为干旱的年份可以保持相对稳定的生长; 而在较为湿润的年份, 多年生根茎禾草则可以迅速利用水分和养分资源, 实现快速生长.在本研究中, 我们也发现了类似的补偿现象, 实验前3年(2013-2015)雨水较为充足, 特别是在氮添加处理下, 多年生根茎禾草的生物量持续增加; 但在2016年后, 由于降水减少, 多年生根茎禾草的生物量迅速下降, 而多年生丛生禾草仍然维持了较高的生物量.近年来的研究也证实物种间的非同步性是维持草原群落生产力稳定性的重要途径(Yang et al., 2012; Xu et al., 2015; Zhang et al., 2016; Wilcox et al., 2017; Zhang et al., 2018b; Sasaki et al., 2019; Wu & Wang, 2019). ...
Functional-and abundance-based mechanisms explain diversity loss due to N fertilization 1 2005
... 在大多数陆地生态系统中, 氮是植物生长的主要限制因子(Vitousek & Howarth, 1991; LeBauer & Treseder, 2008; Xia & Wan, 2008; Lu et al., 2011; ?gren et al., 2012; Gao et al., 2019).大量研究表明氮添加会提高植物群落生产力, 但群落植物物种多样性却显著降低(Stevens et al., 2004; Suding et al., 2005; Clark & Tilman, 2008; Isbell et al., 2013).由于植物对氮、磷等养分元素的重吸收作用, 凋落物中的碳氮比相对较高(Deng et al., 2018).日益加剧的大气氮沉降, 会提高环境中活性氮的含量(Gruber & Galloway, 2008).这一方面降低了凋落物中碳氮比, 另一方面缓解了微生物的营养限制, 促进微生物活性, 进而加速凋落物的分解(Austin & Vivanco, 2006; Yue et al., 2016; Wang et al., 2017b), 改善了环境中的养分状况(Manning et al., 2008; Zhang et al., 2018a).那么, 凋落物和氮两者共同作用会对生态系统生产力和群落结构产生怎样的影响呢? 目前还缺乏长期的实验研究. ...
A novel soil manganese mechanism drives plant species loss with increased nitrogen deposition in a temperate steppe 1 2016
... 在我国半干旱草原, 持续大量的氮添加会降低群落生物多样性是已经被大量的实验证实的结论(Stevens et al., 2004; Clark & Tilman, 2010; Zhang et al., 2016; Liu et al., 2018), 但与以往研究不同的是, 本研究经过6年的氮添加处理, 并没有发现物种和功能群多样性发生显著变化, 这可能是与本实验的氮添加量有关.在本研究区域已开展的长期氮添加梯度实验中, 草原生产力和多样性均与氮添加量表现出非线性关系, 呈现先增加后降低的趋势, 而拐点为10-15 g·m-2·a-1, 过量的氮添加会因为土壤酸化和有害离子的富集而导致群落生产力和多样性的降低(Stevens et al., 2004; Tian et al., 2016; Gao et al., 2019).本研究的施氮量为15 g·m-2·a-1, 对该区域草原生态系统是一个比较适宜的氮添加水平, 因此本实验中的氮添加处理对物种和功能群多样性并没有显著的影响.我们发现, 氮添加提高土壤养分含量, 显著促进多年生丛生禾草、多年生根茎禾草和一二年生植物的生长, 增大其对群落ANPP的贡献.而另一个优势功能群多年生杂类草的生物量主要与多年生根茎禾草此消彼长, 由于多年生根茎禾草的增加, 最终造成多年生杂类草在群落ANPP相对贡献的显著下降; 并且还发现灌木与半灌木的生物量因多年生禾草的增加而减少.Bai等(2004)通过分析本区域草原生产力的长期动态, 提出多年生丛生禾草和多年生根茎禾草两个主要功能群之间的补偿作用是维持半干旱草原生产力稳定性的重要机制.与多年生根茎禾草相比, 多年生丛生禾草(以大针茅、冰草(Agropyron cristatum)等丛生禾草为主)更能忍耐干旱和贫瘠的生境, 又因其生长速率较慢, 因此在较为干旱的年份可以保持相对稳定的生长; 而在较为湿润的年份, 多年生根茎禾草则可以迅速利用水分和养分资源, 实现快速生长.在本研究中, 我们也发现了类似的补偿现象, 实验前3年(2013-2015)雨水较为充足, 特别是在氮添加处理下, 多年生根茎禾草的生物量持续增加; 但在2016年后, 由于降水减少, 多年生根茎禾草的生物量迅速下降, 而多年生丛生禾草仍然维持了较高的生物量.近年来的研究也证实物种间的非同步性是维持草原群落生产力稳定性的重要途径(Yang et al., 2012; Xu et al., 2015; Zhang et al., 2016; Wilcox et al., 2017; Zhang et al., 2018b; Sasaki et al., 2019; Wu & Wang, 2019). ...
The effect of litter on soil organic matter (SOM) turnover in Síkf?kút site 1 2008
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
Nitrogen limitation on land and in the sea: How can it occur? 1 1991
... 在大多数陆地生态系统中, 氮是植物生长的主要限制因子(Vitousek & Howarth, 1991; LeBauer & Treseder, 2008; Xia & Wan, 2008; Lu et al., 2011; ?gren et al., 2012; Gao et al., 2019).大量研究表明氮添加会提高植物群落生产力, 但群落植物物种多样性却显著降低(Stevens et al., 2004; Suding et al., 2005; Clark & Tilman, 2008; Isbell et al., 2013).由于植物对氮、磷等养分元素的重吸收作用, 凋落物中的碳氮比相对较高(Deng et al., 2018).日益加剧的大气氮沉降, 会提高环境中活性氮的含量(Gruber & Galloway, 2008).这一方面降低了凋落物中碳氮比, 另一方面缓解了微生物的营养限制, 促进微生物活性, 进而加速凋落物的分解(Austin & Vivanco, 2006; Yue et al., 2016; Wang et al., 2017b), 改善了环境中的养分状况(Manning et al., 2008; Zhang et al., 2018a).那么, 凋落物和氮两者共同作用会对生态系统生产力和群落结构产生怎样的影响呢? 目前还缺乏长期的实验研究. ...
... 近年来, 凋落物输入变化对生态系统结构功能的影响更多聚焦在植被生产力、土壤微生物介导的养分循环过程和土壤碳库稳定性的变化上(Xu et al., 2013a), 而对于凋落物输入变化对群落结构影响的报道比较有限(王静, 2011; 蒋红志, 2018).凋落物对群落结构的影响则主要通过其对不同植物功能群的影响差异来实现, 如Facelli和Pickett (1991b)发现凋落物输入增加会抑制草本植物的生长, 其中既发现对外来种生长建群的抑制(Patrick et al., 2008), 也有对原生种生长的抑制, 这主要与凋落物类型有关(Facelli & Pickett, 1991b; Foster & Gross, 1998; Amatangelo et al., 2008; Letts et al., 2015).此外, 凋落物的累积能通过防止种子进入土壤层而对种子补充过程形成抑制, 也会通过化感作用抑制种子萌发, 累积的凋落物也能通过降低物种丰富度和均匀度来改变群落的物种组成(Foster & Gross, 1998; Amatangelo et al., 2008; Letts et al., 2015). ...
1 2011
... 近年来, 凋落物输入变化对生态系统结构功能的影响更多聚焦在植被生产力、土壤微生物介导的养分循环过程和土壤碳库稳定性的变化上(Xu et al., 2013a), 而对于凋落物输入变化对群落结构影响的报道比较有限(王静, 2011; 蒋红志, 2018).凋落物对群落结构的影响则主要通过其对不同植物功能群的影响差异来实现, 如Facelli和Pickett (1991b)发现凋落物输入增加会抑制草本植物的生长, 其中既发现对外来种生长建群的抑制(Patrick et al., 2008), 也有对原生种生长的抑制, 这主要与凋落物类型有关(Facelli & Pickett, 1991b; Foster & Gross, 1998; Amatangelo et al., 2008; Letts et al., 2015).此外, 凋落物的累积能通过防止种子进入土壤层而对种子补充过程形成抑制, 也会通过化感作用抑制种子萌发, 累积的凋落物也能通过降低物种丰富度和均匀度来改变群落的物种组成(Foster & Gross, 1998; Amatangelo et al., 2008; Letts et al., 2015). ...
Asymmetry in above- and belowground productivity responses to N addition in a semi-arid temperate steppe 1 2019
... 以往的研究表明, 氮添加会显著提高半干旱草原生态系统生产力(潘庆民等, 2005; 沈月, 2016; Yue et al., 2016; Wang et al., 2019; Zhao et al., 2019).本研究也发现, 氮添加显著提高了土壤DOC和DON含量, 促进植物生长, 显著提高了群落生产力.内蒙古半干旱草原作为一个水氮共同限制的生态系统(Heisler-White et al., 2008; Li et al., 2011; Ren et al., 2017).氮对草原生产力的影响强度甚至方向都是依赖于生态系统的水分状况的(Yan et al., 2010; Shen et al., 2016).在本研究中, 氮添加对生产力的促进作用在降水量较高的湿润年份表现得更为明显, 而在较为干旱的年份(2016和2017年), 氮添加对ANPP的促进作用明显减弱(图3A).ANPP与降水量之间的关系也表明, 氮添加显著提高了ANPP-降水量关系的阈值, 随着降水量增加, ANPP增加的趋势更加明显, 这也进一步证实了氮添加处理对水分条件的依赖性(Yang et al., 2011; Zong et al., 2019).此外, 氮添加显著降低了土壤温度, 这主要是由于氮添加促进植物生长, 提高植被盖度, 同时导致第二年凋落物输入量的进一步增多, 对土壤温度调节表现出显著的负反馈作用, 这也有助于缓解干旱年份的水分胁迫作用(Li et al., 2011; Zhao et al., 2019).我们的研究还发现氮添加并未改变凋落物输入变化对群落ANPP的影响, 二者没有显著的交互作用.与Shen等(2016)在本区域进行氮和凋落物添加实验的结果相同.这可能与半干旱草原降水量和生产力剧烈的年际变异有关, 要想进一步探讨这两者之间的交互作用是否会对系统生产力形成影响, 还需要更长期的实验数据积累减弱或去除数据分析过程中年份等其他因素的影响. ...
Can plant litter affect net primary production of a typical steppe in Inner Mongolia? 2 2011
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
... ; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
Long-term litter manipulation alters soil organic matter turnover in a temperate deciduous forest 2 2017
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
... 连续6年的实验处理结果表明, 凋落物输入量的增加显著提高了群落ANPP, 而凋落物去除则降低了ANPP (图3B).相关分析表明随着凋落物添加量的增加, 群落ANPP显著提高(图6A).已有研究表明凋落物对ANPP的影响主要分为两个方面, 一方面是通过凋落物自身分解直接增加生态系统碳输入, 提高土壤养分可利用性来促进ANPP (Brearley et al., 2003; Bansal et al., 2014); 另一方面是通过对微环境条件(如土壤温度、水分状况等)的调控来间接影响ANPP (Booth et al., 2005; Deutsch et al., 2010a).在本研究中, 凋落物输入变化并没有显著改变DOC和DON等土壤养分含量, 而是显著改善了土壤水分和温度等土壤环境条件间接影响植被生产力.ANPP与降水量以及土壤含水量的关系表明, 水分是限制群落生产力的主要限制因子, 凋落物添加通过降低土温, 减少水分蒸发, 提高表层土壤水分可利用性, 降低土壤水分的季节变异, 从而降低了植物遭受干旱胁迫风险, 进而促进植物生长, 提高群落ANPP.Xu等(2013a)在关于生态系统中凋落物作用的meta分析中发现, 在草原生态系统的研究中, 凋落物添加会增加土壤微生物碳以及总碳含量, 并降低土壤温度、增加土壤含水量.我们的研究虽然在统计上尚未检测到凋落物输入变化对土壤DOC和DON含量的显著影响, 但还是发现随着凋落物的增加, DOC含量呈现出增加的趋势.这表明凋落物输入变化对土壤DOC等养分可利用性的影响可能需要更长期的实验研究结果来检验(Wang et al., 2017a).尤其在干旱半干旱生态系统中, 因为受到水分条件的限制, 凋落物分解速率相对更慢(Zhang & Wang, 2015). ...
1 2016
... 本研究区域位于内蒙古自治区锡林郭勒盟西乌珠穆沁旗, 117.58° E, 44.37° N, 海拔1 148 m, 属典型温带半干旱大陆性气候(陈佐忠和汪诗平, 2000).根据当地气象站长期气象数据(1955-2011年), 本区域年平均气温为1.5 ℃, 年降水量333 mm, 大部分降水(超过87%)发生在生长季(5-9月).降水量年际间波动剧烈, 年降水量最多和最少分别能达到508和118 mm.本区域典型植被类型为温带半干旱草原, 以多年生禾草和多年生杂类草为优势植物, 放牧和打草是本区域最普遍的草原利用方式.土壤类型为栗钙土, 表层土壤(0-10 cm)有机碳含量(14.2 ± 0.4) g·kg-1, 氮含量(1.8 ± 0.1) g·kg-1, pH为7.7 ± 0.5 (王珊珊, 2016; Zhang et al., 2017). ...
1 2016
... 本研究区域位于内蒙古自治区锡林郭勒盟西乌珠穆沁旗, 117.58° E, 44.37° N, 海拔1 148 m, 属典型温带半干旱大陆性气候(陈佐忠和汪诗平, 2000).根据当地气象站长期气象数据(1955-2011年), 本区域年平均气温为1.5 ℃, 年降水量333 mm, 大部分降水(超过87%)发生在生长季(5-9月).降水量年际间波动剧烈, 年降水量最多和最少分别能达到508和118 mm.本区域典型植被类型为温带半干旱草原, 以多年生禾草和多年生杂类草为优势植物, 放牧和打草是本区域最普遍的草原利用方式.土壤类型为栗钙土, 表层土壤(0-10 cm)有机碳含量(14.2 ± 0.4) g·kg-1, 氮含量(1.8 ± 0.1) g·kg-1, pH为7.7 ± 0.5 (王珊珊, 2016; Zhang et al., 2017). ...
Responses of litter decomposition and nutrient release rate to water and nitrogen addition differed among three plant species dominated in a semi-arid grassland 1 2017
... 在大多数陆地生态系统中, 氮是植物生长的主要限制因子(Vitousek & Howarth, 1991; LeBauer & Treseder, 2008; Xia & Wan, 2008; Lu et al., 2011; ?gren et al., 2012; Gao et al., 2019).大量研究表明氮添加会提高植物群落生产力, 但群落植物物种多样性却显著降低(Stevens et al., 2004; Suding et al., 2005; Clark & Tilman, 2008; Isbell et al., 2013).由于植物对氮、磷等养分元素的重吸收作用, 凋落物中的碳氮比相对较高(Deng et al., 2018).日益加剧的大气氮沉降, 会提高环境中活性氮的含量(Gruber & Galloway, 2008).这一方面降低了凋落物中碳氮比, 另一方面缓解了微生物的营养限制, 促进微生物活性, 进而加速凋落物的分解(Austin & Vivanco, 2006; Yue et al., 2016; Wang et al., 2017b), 改善了环境中的养分状况(Manning et al., 2008; Zhang et al., 2018a).那么, 凋落物和氮两者共同作用会对生态系统生产力和群落结构产生怎样的影响呢? 目前还缺乏长期的实验研究. ...
Ecological linkages between aboveground and belowground biota 1 2004
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
Effect of fertilization date and litter removal on grassland forage production 1 1989
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
Asynchrony among local communities stabilises ecosystem function of metacommunities 1 2017
... 在我国半干旱草原, 持续大量的氮添加会降低群落生物多样性是已经被大量的实验证实的结论(Stevens et al., 2004; Clark & Tilman, 2010; Zhang et al., 2016; Liu et al., 2018), 但与以往研究不同的是, 本研究经过6年的氮添加处理, 并没有发现物种和功能群多样性发生显著变化, 这可能是与本实验的氮添加量有关.在本研究区域已开展的长期氮添加梯度实验中, 草原生产力和多样性均与氮添加量表现出非线性关系, 呈现先增加后降低的趋势, 而拐点为10-15 g·m-2·a-1, 过量的氮添加会因为土壤酸化和有害离子的富集而导致群落生产力和多样性的降低(Stevens et al., 2004; Tian et al., 2016; Gao et al., 2019).本研究的施氮量为15 g·m-2·a-1, 对该区域草原生态系统是一个比较适宜的氮添加水平, 因此本实验中的氮添加处理对物种和功能群多样性并没有显著的影响.我们发现, 氮添加提高土壤养分含量, 显著促进多年生丛生禾草、多年生根茎禾草和一二年生植物的生长, 增大其对群落ANPP的贡献.而另一个优势功能群多年生杂类草的生物量主要与多年生根茎禾草此消彼长, 由于多年生根茎禾草的增加, 最终造成多年生杂类草在群落ANPP相对贡献的显著下降; 并且还发现灌木与半灌木的生物量因多年生禾草的增加而减少.Bai等(2004)通过分析本区域草原生产力的长期动态, 提出多年生丛生禾草和多年生根茎禾草两个主要功能群之间的补偿作用是维持半干旱草原生产力稳定性的重要机制.与多年生根茎禾草相比, 多年生丛生禾草(以大针茅、冰草(Agropyron cristatum)等丛生禾草为主)更能忍耐干旱和贫瘠的生境, 又因其生长速率较慢, 因此在较为干旱的年份可以保持相对稳定的生长; 而在较为湿润的年份, 多年生根茎禾草则可以迅速利用水分和养分资源, 实现快速生长.在本研究中, 我们也发现了类似的补偿现象, 实验前3年(2013-2015)雨水较为充足, 特别是在氮添加处理下, 多年生根茎禾草的生物量持续增加; 但在2016年后, 由于降水减少, 多年生根茎禾草的生物量迅速下降, 而多年生丛生禾草仍然维持了较高的生物量.近年来的研究也证实物种间的非同步性是维持草原群落生产力稳定性的重要途径(Yang et al., 2012; Xu et al., 2015; Zhang et al., 2016; Wilcox et al., 2017; Zhang et al., 2018b; Sasaki et al., 2019; Wu & Wang, 2019). ...
Effect of fencing and grazing on a Kobresia-dominated meadow in the Qinghai-Tibetan Plateau 2 2009
... 我国天然草地面积约4亿hm2, 是我国面积最大的陆地植被类型, 其中北方温带草原所占比例最大(陈佐忠和汪诗平, 2000; Piao et al., 2009; Fang et al., 2010).由于不合理的开发利用和气候变化, 全国90%的天然草地存在不同程度的退化(白永飞等, 2016).为了治理草原退化, 国家从2003年开始通过一系列管理措施, 改善了草原生态环境(张英俊等, 2013).与放牧和打草等传统草原利用方式相比, 围封禁牧可以使退化草原植被得到迅速恢复, 因而作为最经济有效的草地恢复方式得到推广(Wu et al., 2009; Jing et al., 2013).但也有研究发现长期围封所造成的凋落物大量累积反而会抑制生产力(Rice & Parenti, 1978; Knapp & Seastedt, 1986). ...
... 在我国半干旱草原, 持续大量的氮添加会降低群落生物多样性是已经被大量的实验证实的结论(Stevens et al., 2004; Clark & Tilman, 2010; Zhang et al., 2016; Liu et al., 2018), 但与以往研究不同的是, 本研究经过6年的氮添加处理, 并没有发现物种和功能群多样性发生显著变化, 这可能是与本实验的氮添加量有关.在本研究区域已开展的长期氮添加梯度实验中, 草原生产力和多样性均与氮添加量表现出非线性关系, 呈现先增加后降低的趋势, 而拐点为10-15 g·m-2·a-1, 过量的氮添加会因为土壤酸化和有害离子的富集而导致群落生产力和多样性的降低(Stevens et al., 2004; Tian et al., 2016; Gao et al., 2019).本研究的施氮量为15 g·m-2·a-1, 对该区域草原生态系统是一个比较适宜的氮添加水平, 因此本实验中的氮添加处理对物种和功能群多样性并没有显著的影响.我们发现, 氮添加提高土壤养分含量, 显著促进多年生丛生禾草、多年生根茎禾草和一二年生植物的生长, 增大其对群落ANPP的贡献.而另一个优势功能群多年生杂类草的生物量主要与多年生根茎禾草此消彼长, 由于多年生根茎禾草的增加, 最终造成多年生杂类草在群落ANPP相对贡献的显著下降; 并且还发现灌木与半灌木的生物量因多年生禾草的增加而减少.Bai等(2004)通过分析本区域草原生产力的长期动态, 提出多年生丛生禾草和多年生根茎禾草两个主要功能群之间的补偿作用是维持半干旱草原生产力稳定性的重要机制.与多年生根茎禾草相比, 多年生丛生禾草(以大针茅、冰草(Agropyron cristatum)等丛生禾草为主)更能忍耐干旱和贫瘠的生境, 又因其生长速率较慢, 因此在较为干旱的年份可以保持相对稳定的生长; 而在较为湿润的年份, 多年生根茎禾草则可以迅速利用水分和养分资源, 实现快速生长.在本研究中, 我们也发现了类似的补偿现象, 实验前3年(2013-2015)雨水较为充足, 特别是在氮添加处理下, 多年生根茎禾草的生物量持续增加; 但在2016年后, 由于降水减少, 多年生根茎禾草的生物量迅速下降, 而多年生丛生禾草仍然维持了较高的生物量.近年来的研究也证实物种间的非同步性是维持草原群落生产力稳定性的重要途径(Yang et al., 2012; Xu et al., 2015; Zhang et al., 2016; Wilcox et al., 2017; Zhang et al., 2018b; Sasaki et al., 2019; Wu & Wang, 2019). ...
Temporal stability of aboveground net primary production in northern Tibet alpine steppe in response to nitrogen addition 2019
Global response patterns of terrestrial plant species to nitrogen addition 1 2008
... 在大多数陆地生态系统中, 氮是植物生长的主要限制因子(Vitousek & Howarth, 1991; LeBauer & Treseder, 2008; Xia & Wan, 2008; Lu et al., 2011; ?gren et al., 2012; Gao et al., 2019).大量研究表明氮添加会提高植物群落生产力, 但群落植物物种多样性却显著降低(Stevens et al., 2004; Suding et al., 2005; Clark & Tilman, 2008; Isbell et al., 2013).由于植物对氮、磷等养分元素的重吸收作用, 凋落物中的碳氮比相对较高(Deng et al., 2018).日益加剧的大气氮沉降, 会提高环境中活性氮的含量(Gruber & Galloway, 2008).这一方面降低了凋落物中碳氮比, 另一方面缓解了微生物的营养限制, 促进微生物活性, 进而加速凋落物的分解(Austin & Vivanco, 2006; Yue et al., 2016; Wang et al., 2017b), 改善了环境中的养分状况(Manning et al., 2008; Zhang et al., 2018a).那么, 凋落物和氮两者共同作用会对生态系统生产力和群落结构产生怎样的影响呢? 目前还缺乏长期的实验研究. ...
Variability of above-ground litter inputs alters soil physicochemical and biological processes: a meta-analysis of litterfall-manipulation experiments 2 2013
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
... 近年来, 凋落物输入变化对生态系统结构功能的影响更多聚焦在植被生产力、土壤微生物介导的养分循环过程和土壤碳库稳定性的变化上(Xu et al., 2013a), 而对于凋落物输入变化对群落结构影响的报道比较有限(王静, 2011; 蒋红志, 2018).凋落物对群落结构的影响则主要通过其对不同植物功能群的影响差异来实现, 如Facelli和Pickett (1991b)发现凋落物输入增加会抑制草本植物的生长, 其中既发现对外来种生长建群的抑制(Patrick et al., 2008), 也有对原生种生长的抑制, 这主要与凋落物类型有关(Facelli & Pickett, 1991b; Foster & Gross, 1998; Amatangelo et al., 2008; Letts et al., 2015).此外, 凋落物的累积能通过防止种子进入土壤层而对种子补充过程形成抑制, 也会通过化感作用抑制种子萌发, 累积的凋落物也能通过降低物种丰富度和均匀度来改变群落的物种组成(Foster & Gross, 1998; Amatangelo et al., 2008; Letts et al., 2015). ...
Net primary productivity and rain-use efficiency as affected by warming, altered precipitation, and clipping in a mixed-grass prairie 1 2013
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
Environmental changes drive the temporal stability of semi-arid natural grasslands through altering species asynchrony 1 2015
... 在我国半干旱草原, 持续大量的氮添加会降低群落生物多样性是已经被大量的实验证实的结论(Stevens et al., 2004; Clark & Tilman, 2010; Zhang et al., 2016; Liu et al., 2018), 但与以往研究不同的是, 本研究经过6年的氮添加处理, 并没有发现物种和功能群多样性发生显著变化, 这可能是与本实验的氮添加量有关.在本研究区域已开展的长期氮添加梯度实验中, 草原生产力和多样性均与氮添加量表现出非线性关系, 呈现先增加后降低的趋势, 而拐点为10-15 g·m-2·a-1, 过量的氮添加会因为土壤酸化和有害离子的富集而导致群落生产力和多样性的降低(Stevens et al., 2004; Tian et al., 2016; Gao et al., 2019).本研究的施氮量为15 g·m-2·a-1, 对该区域草原生态系统是一个比较适宜的氮添加水平, 因此本实验中的氮添加处理对物种和功能群多样性并没有显著的影响.我们发现, 氮添加提高土壤养分含量, 显著促进多年生丛生禾草、多年生根茎禾草和一二年生植物的生长, 增大其对群落ANPP的贡献.而另一个优势功能群多年生杂类草的生物量主要与多年生根茎禾草此消彼长, 由于多年生根茎禾草的增加, 最终造成多年生杂类草在群落ANPP相对贡献的显著下降; 并且还发现灌木与半灌木的生物量因多年生禾草的增加而减少.Bai等(2004)通过分析本区域草原生产力的长期动态, 提出多年生丛生禾草和多年生根茎禾草两个主要功能群之间的补偿作用是维持半干旱草原生产力稳定性的重要机制.与多年生根茎禾草相比, 多年生丛生禾草(以大针茅、冰草(Agropyron cristatum)等丛生禾草为主)更能忍耐干旱和贫瘠的生境, 又因其生长速率较慢, 因此在较为干旱的年份可以保持相对稳定的生长; 而在较为湿润的年份, 多年生根茎禾草则可以迅速利用水分和养分资源, 实现快速生长.在本研究中, 我们也发现了类似的补偿现象, 实验前3年(2013-2015)雨水较为充足, 特别是在氮添加处理下, 多年生根茎禾草的生物量持续增加; 但在2016年后, 由于降水减少, 多年生根茎禾草的生物量迅速下降, 而多年生丛生禾草仍然维持了较高的生物量.近年来的研究也证实物种间的非同步性是维持草原群落生产力稳定性的重要途径(Yang et al., 2012; Xu et al., 2015; Zhang et al., 2016; Wilcox et al., 2017; Zhang et al., 2018b; Sasaki et al., 2019; Wu & Wang, 2019). ...
Differential responses of auto- and heterotrophic soil respiration to water and nitrogen addition in a semiarid temperate steppe 1 2010
... 以往的研究表明, 氮添加会显著提高半干旱草原生态系统生产力(潘庆民等, 2005; 沈月, 2016; Yue et al., 2016; Wang et al., 2019; Zhao et al., 2019).本研究也发现, 氮添加显著提高了土壤DOC和DON含量, 促进植物生长, 显著提高了群落生产力.内蒙古半干旱草原作为一个水氮共同限制的生态系统(Heisler-White et al., 2008; Li et al., 2011; Ren et al., 2017).氮对草原生产力的影响强度甚至方向都是依赖于生态系统的水分状况的(Yan et al., 2010; Shen et al., 2016).在本研究中, 氮添加对生产力的促进作用在降水量较高的湿润年份表现得更为明显, 而在较为干旱的年份(2016和2017年), 氮添加对ANPP的促进作用明显减弱(图3A).ANPP与降水量之间的关系也表明, 氮添加显著提高了ANPP-降水量关系的阈值, 随着降水量增加, ANPP增加的趋势更加明显, 这也进一步证实了氮添加处理对水分条件的依赖性(Yang et al., 2011; Zong et al., 2019).此外, 氮添加显著降低了土壤温度, 这主要是由于氮添加促进植物生长, 提高植被盖度, 同时导致第二年凋落物输入量的进一步增多, 对土壤温度调节表现出显著的负反馈作用, 这也有助于缓解干旱年份的水分胁迫作用(Li et al., 2011; Zhao et al., 2019).我们的研究还发现氮添加并未改变凋落物输入变化对群落ANPP的影响, 二者没有显著的交互作用.与Shen等(2016)在本区域进行氮和凋落物添加实验的结果相同.这可能与半干旱草原降水量和生产力剧烈的年际变异有关, 要想进一步探讨这两者之间的交互作用是否会对系统生产力形成影响, 还需要更长期的实验数据积累减弱或去除数据分析过程中年份等其他因素的影响. ...
Diversity-dependent stability under mowing and nutrient addition: evidence from a 7-year grassland experiment 1 2012
... 在我国半干旱草原, 持续大量的氮添加会降低群落生物多样性是已经被大量的实验证实的结论(Stevens et al., 2004; Clark & Tilman, 2010; Zhang et al., 2016; Liu et al., 2018), 但与以往研究不同的是, 本研究经过6年的氮添加处理, 并没有发现物种和功能群多样性发生显著变化, 这可能是与本实验的氮添加量有关.在本研究区域已开展的长期氮添加梯度实验中, 草原生产力和多样性均与氮添加量表现出非线性关系, 呈现先增加后降低的趋势, 而拐点为10-15 g·m-2·a-1, 过量的氮添加会因为土壤酸化和有害离子的富集而导致群落生产力和多样性的降低(Stevens et al., 2004; Tian et al., 2016; Gao et al., 2019).本研究的施氮量为15 g·m-2·a-1, 对该区域草原生态系统是一个比较适宜的氮添加水平, 因此本实验中的氮添加处理对物种和功能群多样性并没有显著的影响.我们发现, 氮添加提高土壤养分含量, 显著促进多年生丛生禾草、多年生根茎禾草和一二年生植物的生长, 增大其对群落ANPP的贡献.而另一个优势功能群多年生杂类草的生物量主要与多年生根茎禾草此消彼长, 由于多年生根茎禾草的增加, 最终造成多年生杂类草在群落ANPP相对贡献的显著下降; 并且还发现灌木与半灌木的生物量因多年生禾草的增加而减少.Bai等(2004)通过分析本区域草原生产力的长期动态, 提出多年生丛生禾草和多年生根茎禾草两个主要功能群之间的补偿作用是维持半干旱草原生产力稳定性的重要机制.与多年生根茎禾草相比, 多年生丛生禾草(以大针茅、冰草(Agropyron cristatum)等丛生禾草为主)更能忍耐干旱和贫瘠的生境, 又因其生长速率较慢, 因此在较为干旱的年份可以保持相对稳定的生长; 而在较为湿润的年份, 多年生根茎禾草则可以迅速利用水分和养分资源, 实现快速生长.在本研究中, 我们也发现了类似的补偿现象, 实验前3年(2013-2015)雨水较为充足, 特别是在氮添加处理下, 多年生根茎禾草的生物量持续增加; 但在2016年后, 由于降水减少, 多年生根茎禾草的生物量迅速下降, 而多年生丛生禾草仍然维持了较高的生物量.近年来的研究也证实物种间的非同步性是维持草原群落生产力稳定性的重要途径(Yang et al., 2012; Xu et al., 2015; Zhang et al., 2016; Wilcox et al., 2017; Zhang et al., 2018b; Sasaki et al., 2019; Wu & Wang, 2019). ...
Plant community responses to nitrogen addition and increased precipitation: the importance of water availability and species traits 1 2011
... 以往的研究表明, 氮添加会显著提高半干旱草原生态系统生产力(潘庆民等, 2005; 沈月, 2016; Yue et al., 2016; Wang et al., 2019; Zhao et al., 2019).本研究也发现, 氮添加显著提高了土壤DOC和DON含量, 促进植物生长, 显著提高了群落生产力.内蒙古半干旱草原作为一个水氮共同限制的生态系统(Heisler-White et al., 2008; Li et al., 2011; Ren et al., 2017).氮对草原生产力的影响强度甚至方向都是依赖于生态系统的水分状况的(Yan et al., 2010; Shen et al., 2016).在本研究中, 氮添加对生产力的促进作用在降水量较高的湿润年份表现得更为明显, 而在较为干旱的年份(2016和2017年), 氮添加对ANPP的促进作用明显减弱(图3A).ANPP与降水量之间的关系也表明, 氮添加显著提高了ANPP-降水量关系的阈值, 随着降水量增加, ANPP增加的趋势更加明显, 这也进一步证实了氮添加处理对水分条件的依赖性(Yang et al., 2011; Zong et al., 2019).此外, 氮添加显著降低了土壤温度, 这主要是由于氮添加促进植物生长, 提高植被盖度, 同时导致第二年凋落物输入量的进一步增多, 对土壤温度调节表现出显著的负反馈作用, 这也有助于缓解干旱年份的水分胁迫作用(Li et al., 2011; Zhao et al., 2019).我们的研究还发现氮添加并未改变凋落物输入变化对群落ANPP的影响, 二者没有显著的交互作用.与Shen等(2016)在本区域进行氮和凋落物添加实验的结果相同.这可能与半干旱草原降水量和生产力剧烈的年际变异有关, 要想进一步探讨这两者之间的交互作用是否会对系统生产力形成影响, 还需要更长期的实验数据积累减弱或去除数据分析过程中年份等其他因素的影响. ...
Soil microbe active community composition and capability of responding to litter addition after 12 years of no inputs 1 2013
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
Stimulation of terrestrial ecosystem carbon storage by nitrogen addition: a meta-analysis 2 2016
... 在大多数陆地生态系统中, 氮是植物生长的主要限制因子(Vitousek & Howarth, 1991; LeBauer & Treseder, 2008; Xia & Wan, 2008; Lu et al., 2011; ?gren et al., 2012; Gao et al., 2019).大量研究表明氮添加会提高植物群落生产力, 但群落植物物种多样性却显著降低(Stevens et al., 2004; Suding et al., 2005; Clark & Tilman, 2008; Isbell et al., 2013).由于植物对氮、磷等养分元素的重吸收作用, 凋落物中的碳氮比相对较高(Deng et al., 2018).日益加剧的大气氮沉降, 会提高环境中活性氮的含量(Gruber & Galloway, 2008).这一方面降低了凋落物中碳氮比, 另一方面缓解了微生物的营养限制, 促进微生物活性, 进而加速凋落物的分解(Austin & Vivanco, 2006; Yue et al., 2016; Wang et al., 2017b), 改善了环境中的养分状况(Manning et al., 2008; Zhang et al., 2018a).那么, 凋落物和氮两者共同作用会对生态系统生产力和群落结构产生怎样的影响呢? 目前还缺乏长期的实验研究. ...
... 以往的研究表明, 氮添加会显著提高半干旱草原生态系统生产力(潘庆民等, 2005; 沈月, 2016; Yue et al., 2016; Wang et al., 2019; Zhao et al., 2019).本研究也发现, 氮添加显著提高了土壤DOC和DON含量, 促进植物生长, 显著提高了群落生产力.内蒙古半干旱草原作为一个水氮共同限制的生态系统(Heisler-White et al., 2008; Li et al., 2011; Ren et al., 2017).氮对草原生产力的影响强度甚至方向都是依赖于生态系统的水分状况的(Yan et al., 2010; Shen et al., 2016).在本研究中, 氮添加对生产力的促进作用在降水量较高的湿润年份表现得更为明显, 而在较为干旱的年份(2016和2017年), 氮添加对ANPP的促进作用明显减弱(图3A).ANPP与降水量之间的关系也表明, 氮添加显著提高了ANPP-降水量关系的阈值, 随着降水量增加, ANPP增加的趋势更加明显, 这也进一步证实了氮添加处理对水分条件的依赖性(Yang et al., 2011; Zong et al., 2019).此外, 氮添加显著降低了土壤温度, 这主要是由于氮添加促进植物生长, 提高植被盖度, 同时导致第二年凋落物输入量的进一步增多, 对土壤温度调节表现出显著的负反馈作用, 这也有助于缓解干旱年份的水分胁迫作用(Li et al., 2011; Zhao et al., 2019).我们的研究还发现氮添加并未改变凋落物输入变化对群落ANPP的影响, 二者没有显著的交互作用.与Shen等(2016)在本区域进行氮和凋落物添加实验的结果相同.这可能与半干旱草原降水量和生产力剧烈的年际变异有关, 要想进一步探讨这两者之间的交互作用是否会对系统生产力形成影响, 还需要更长期的实验数据积累减弱或去除数据分析过程中年份等其他因素的影响. ...
Asymmetric sensitivity of ecosystem carbon and water processes in response to precipitation change in a semi-arid steppe 1 2017
... 本研究区域位于内蒙古自治区锡林郭勒盟西乌珠穆沁旗, 117.58° E, 44.37° N, 海拔1 148 m, 属典型温带半干旱大陆性气候(陈佐忠和汪诗平, 2000).根据当地气象站长期气象数据(1955-2011年), 本区域年平均气温为1.5 ℃, 年降水量333 mm, 大部分降水(超过87%)发生在生长季(5-9月).降水量年际间波动剧烈, 年降水量最多和最少分别能达到508和118 mm.本区域典型植被类型为温带半干旱草原, 以多年生禾草和多年生杂类草为优势植物, 放牧和打草是本区域最普遍的草原利用方式.土壤类型为栗钙土, 表层土壤(0-10 cm)有机碳含量(14.2 ± 0.4) g·kg-1, 氮含量(1.8 ± 0.1) g·kg-1, pH为7.7 ± 0.5 (王珊珊, 2016; Zhang et al., 2017). ...
Responses of litter decomposition and nutrient release to N addition: a meta-analysis of terrestrial ecosystems 2 2018
... 凋落物是陆地生态系统重要碳库之一, 其分解过程作为重要养分释放的途径, 连接着地上地下碳循环过程(Facelli & Pickett, 1991b; Sayer et al., 2011; Wang et al., 2011).人类活动和土地利用方式的改变正在显著影响着凋落物的产生及其向土壤的输入(Xu et al., 2013a; Fang et al., 2018).凋落物数量及其分解速度会直接影响植物和土壤微生物吸收利用养分的过程, 并进一步调控生态系统结构和功能(Wardle et al., 2004; Zhang et al., 2018a).目前有关凋落物输入变化对群落生产力影响的研究已有很多(Lamb, 2008; Ruprecht et al., 2010), 并得到了普遍的共识, 即凋落物输入量增加能够提高植被生产力(Patrick et al., 2008; Deutsch et al., 2010b; Wang et al., 2011; Kelemen et al., 2013), 而去除凋落物会导致群落生产力的下降(Wikeem et al., 1989; Xu et al., 2013b).凋落物输入量变化主要通过以下途径影响群落生产力: 首先, 作为土壤养分的重要来源, 凋落物输入量的变化会改变土壤养分的可利用性, 并直接影响植物的生长.如凋落物输入量增加会显著提高土壤可溶性碳氮含量(Brearley et al., 2003; Bansal et al., 2014; Wang et al., 2017a), 但对土壤有机碳库的影响通常是不显著的(Varga et al., 2008).第二, 凋落物输入量变化通过改变底物供给状况影响土壤微生物活性和群落组成, 从而影响凋落物分解和养分释放速率(Yarwood et al., 2013; Kohmann et al., 2019).第三, 作为植被和大气间的缓冲层, 覆盖地表的凋落物层会造成地表光、热和水等资源的重新分配, 影响土壤和植物群落微环境, 如凋落物层通过减少地表日间接收到的光辐射和夜间的热量散失来维持土壤相对稳定的温度状态(Facelli & Pickett, 1991a), 也可以通过减少蒸发来提高土壤含水量(Deutsch et al., 2010b), 进而间接影响植物生长和群落结构(Facelli & Pickett, 1991a, 1991b; Jensen & Gutekunst, 2003). ...
... 在大多数陆地生态系统中, 氮是植物生长的主要限制因子(Vitousek & Howarth, 1991; LeBauer & Treseder, 2008; Xia & Wan, 2008; Lu et al., 2011; ?gren et al., 2012; Gao et al., 2019).大量研究表明氮添加会提高植物群落生产力, 但群落植物物种多样性却显著降低(Stevens et al., 2004; Suding et al., 2005; Clark & Tilman, 2008; Isbell et al., 2013).由于植物对氮、磷等养分元素的重吸收作用, 凋落物中的碳氮比相对较高(Deng et al., 2018).日益加剧的大气氮沉降, 会提高环境中活性氮的含量(Gruber & Galloway, 2008).这一方面降低了凋落物中碳氮比, 另一方面缓解了微生物的营养限制, 促进微生物活性, 进而加速凋落物的分解(Austin & Vivanco, 2006; Yue et al., 2016; Wang et al., 2017b), 改善了环境中的养分状况(Manning et al., 2008; Zhang et al., 2018a).那么, 凋落物和氮两者共同作用会对生态系统生产力和群落结构产生怎样的影响呢? 目前还缺乏长期的实验研究. ...
Control of climate and litter quality on leaf litter decomposition in different climatic zones 1 2015
... 连续6年的实验处理结果表明, 凋落物输入量的增加显著提高了群落ANPP, 而凋落物去除则降低了ANPP (图3B).相关分析表明随着凋落物添加量的增加, 群落ANPP显著提高(图6A).已有研究表明凋落物对ANPP的影响主要分为两个方面, 一方面是通过凋落物自身分解直接增加生态系统碳输入, 提高土壤养分可利用性来促进ANPP (Brearley et al., 2003; Bansal et al., 2014); 另一方面是通过对微环境条件(如土壤温度、水分状况等)的调控来间接影响ANPP (Booth et al., 2005; Deutsch et al., 2010a).在本研究中, 凋落物输入变化并没有显著改变DOC和DON等土壤养分含量, 而是显著改善了土壤水分和温度等土壤环境条件间接影响植被生产力.ANPP与降水量以及土壤含水量的关系表明, 水分是限制群落生产力的主要限制因子, 凋落物添加通过降低土温, 减少水分蒸发, 提高表层土壤水分可利用性, 降低土壤水分的季节变异, 从而降低了植物遭受干旱胁迫风险, 进而促进植物生长, 提高群落ANPP.Xu等(2013a)在关于生态系统中凋落物作用的meta分析中发现, 在草原生态系统的研究中, 凋落物添加会增加土壤微生物碳以及总碳含量, 并降低土壤温度、增加土壤含水量.我们的研究虽然在统计上尚未检测到凋落物输入变化对土壤DOC和DON含量的显著影响, 但还是发现随着凋落物的增加, DOC含量呈现出增加的趋势.这表明凋落物输入变化对土壤DOC等养分可利用性的影响可能需要更长期的实验研究结果来检验(Wang et al., 2017a).尤其在干旱半干旱生态系统中, 因为受到水分条件的限制, 凋落物分解速率相对更慢(Zhang & Wang, 2015). ...
Nitrogen enrichment weakens ecosystem stability through decreased species asynchrony and population stability in a temperate grassland 2 2016
... 在我国半干旱草原, 持续大量的氮添加会降低群落生物多样性是已经被大量的实验证实的结论(Stevens et al., 2004; Clark & Tilman, 2010; Zhang et al., 2016; Liu et al., 2018), 但与以往研究不同的是, 本研究经过6年的氮添加处理, 并没有发现物种和功能群多样性发生显著变化, 这可能是与本实验的氮添加量有关.在本研究区域已开展的长期氮添加梯度实验中, 草原生产力和多样性均与氮添加量表现出非线性关系, 呈现先增加后降低的趋势, 而拐点为10-15 g·m-2·a-1, 过量的氮添加会因为土壤酸化和有害离子的富集而导致群落生产力和多样性的降低(Stevens et al., 2004; Tian et al., 2016; Gao et al., 2019).本研究的施氮量为15 g·m-2·a-1, 对该区域草原生态系统是一个比较适宜的氮添加水平, 因此本实验中的氮添加处理对物种和功能群多样性并没有显著的影响.我们发现, 氮添加提高土壤养分含量, 显著促进多年生丛生禾草、多年生根茎禾草和一二年生植物的生长, 增大其对群落ANPP的贡献.而另一个优势功能群多年生杂类草的生物量主要与多年生根茎禾草此消彼长, 由于多年生根茎禾草的增加, 最终造成多年生杂类草在群落ANPP相对贡献的显著下降; 并且还发现灌木与半灌木的生物量因多年生禾草的增加而减少.Bai等(2004)通过分析本区域草原生产力的长期动态, 提出多年生丛生禾草和多年生根茎禾草两个主要功能群之间的补偿作用是维持半干旱草原生产力稳定性的重要机制.与多年生根茎禾草相比, 多年生丛生禾草(以大针茅、冰草(Agropyron cristatum)等丛生禾草为主)更能忍耐干旱和贫瘠的生境, 又因其生长速率较慢, 因此在较为干旱的年份可以保持相对稳定的生长; 而在较为湿润的年份, 多年生根茎禾草则可以迅速利用水分和养分资源, 实现快速生长.在本研究中, 我们也发现了类似的补偿现象, 实验前3年(2013-2015)雨水较为充足, 特别是在氮添加处理下, 多年生根茎禾草的生物量持续增加; 但在2016年后, 由于降水减少, 多年生根茎禾草的生物量迅速下降, 而多年生丛生禾草仍然维持了较高的生物量.近年来的研究也证实物种间的非同步性是维持草原群落生产力稳定性的重要途径(Yang et al., 2012; Xu et al., 2015; Zhang et al., 2016; Wilcox et al., 2017; Zhang et al., 2018b; Sasaki et al., 2019; Wu & Wang, 2019). ...
... ; Zhang et al., 2016; Wilcox et al., 2017; Zhang et al., 2018b; Sasaki et al., 2019; Wu & Wang, 2019). ...
Scale dependence of the diversity-stability relationship in a temperate grassland 1 2018
... 在我国半干旱草原, 持续大量的氮添加会降低群落生物多样性是已经被大量的实验证实的结论(Stevens et al., 2004; Clark & Tilman, 2010; Zhang et al., 2016; Liu et al., 2018), 但与以往研究不同的是, 本研究经过6年的氮添加处理, 并没有发现物种和功能群多样性发生显著变化, 这可能是与本实验的氮添加量有关.在本研究区域已开展的长期氮添加梯度实验中, 草原生产力和多样性均与氮添加量表现出非线性关系, 呈现先增加后降低的趋势, 而拐点为10-15 g·m-2·a-1, 过量的氮添加会因为土壤酸化和有害离子的富集而导致群落生产力和多样性的降低(Stevens et al., 2004; Tian et al., 2016; Gao et al., 2019).本研究的施氮量为15 g·m-2·a-1, 对该区域草原生态系统是一个比较适宜的氮添加水平, 因此本实验中的氮添加处理对物种和功能群多样性并没有显著的影响.我们发现, 氮添加提高土壤养分含量, 显著促进多年生丛生禾草、多年生根茎禾草和一二年生植物的生长, 增大其对群落ANPP的贡献.而另一个优势功能群多年生杂类草的生物量主要与多年生根茎禾草此消彼长, 由于多年生根茎禾草的增加, 最终造成多年生杂类草在群落ANPP相对贡献的显著下降; 并且还发现灌木与半灌木的生物量因多年生禾草的增加而减少.Bai等(2004)通过分析本区域草原生产力的长期动态, 提出多年生丛生禾草和多年生根茎禾草两个主要功能群之间的补偿作用是维持半干旱草原生产力稳定性的重要机制.与多年生根茎禾草相比, 多年生丛生禾草(以大针茅、冰草(Agropyron cristatum)等丛生禾草为主)更能忍耐干旱和贫瘠的生境, 又因其生长速率较慢, 因此在较为干旱的年份可以保持相对稳定的生长; 而在较为湿润的年份, 多年生根茎禾草则可以迅速利用水分和养分资源, 实现快速生长.在本研究中, 我们也发现了类似的补偿现象, 实验前3年(2013-2015)雨水较为充足, 特别是在氮添加处理下, 多年生根茎禾草的生物量持续增加; 但在2016年后, 由于降水减少, 多年生根茎禾草的生物量迅速下降, 而多年生丛生禾草仍然维持了较高的生物量.近年来的研究也证实物种间的非同步性是维持草原群落生产力稳定性的重要途径(Yang et al., 2012; Xu et al., 2015; Zhang et al., 2016; Wilcox et al., 2017; Zhang et al., 2018b; Sasaki et al., 2019; Wu & Wang, 2019). ...
草原碳汇管理对策 1 2013
... 我国天然草地面积约4亿hm2, 是我国面积最大的陆地植被类型, 其中北方温带草原所占比例最大(陈佐忠和汪诗平, 2000; Piao et al., 2009; Fang et al., 2010).由于不合理的开发利用和气候变化, 全国90%的天然草地存在不同程度的退化(白永飞等, 2016).为了治理草原退化, 国家从2003年开始通过一系列管理措施, 改善了草原生态环境(张英俊等, 2013).与放牧和打草等传统草原利用方式相比, 围封禁牧可以使退化草原植被得到迅速恢复, 因而作为最经济有效的草地恢复方式得到推广(Wu et al., 2009; Jing et al., 2013).但也有研究发现长期围封所造成的凋落物大量累积反而会抑制生产力(Rice & Parenti, 1978; Knapp & Seastedt, 1986). ...
草原碳汇管理对策 1 2013
... 我国天然草地面积约4亿hm2, 是我国面积最大的陆地植被类型, 其中北方温带草原所占比例最大(陈佐忠和汪诗平, 2000; Piao et al., 2009; Fang et al., 2010).由于不合理的开发利用和气候变化, 全国90%的天然草地存在不同程度的退化(白永飞等, 2016).为了治理草原退化, 国家从2003年开始通过一系列管理措施, 改善了草原生态环境(张英俊等, 2013).与放牧和打草等传统草原利用方式相比, 围封禁牧可以使退化草原植被得到迅速恢复, 因而作为最经济有效的草地恢复方式得到推广(Wu et al., 2009; Jing et al., 2013).但也有研究发现长期围封所造成的凋落物大量累积反而会抑制生产力(Rice & Parenti, 1978; Knapp & Seastedt, 1986). ...
Community composition, structure and productivity in response to nitrogen and phosphorus additions in a temperate meadow 2 2019
... 以往的研究表明, 氮添加会显著提高半干旱草原生态系统生产力(潘庆民等, 2005; 沈月, 2016; Yue et al., 2016; Wang et al., 2019; Zhao et al., 2019).本研究也发现, 氮添加显著提高了土壤DOC和DON含量, 促进植物生长, 显著提高了群落生产力.内蒙古半干旱草原作为一个水氮共同限制的生态系统(Heisler-White et al., 2008; Li et al., 2011; Ren et al., 2017).氮对草原生产力的影响强度甚至方向都是依赖于生态系统的水分状况的(Yan et al., 2010; Shen et al., 2016).在本研究中, 氮添加对生产力的促进作用在降水量较高的湿润年份表现得更为明显, 而在较为干旱的年份(2016和2017年), 氮添加对ANPP的促进作用明显减弱(图3A).ANPP与降水量之间的关系也表明, 氮添加显著提高了ANPP-降水量关系的阈值, 随着降水量增加, ANPP增加的趋势更加明显, 这也进一步证实了氮添加处理对水分条件的依赖性(Yang et al., 2011; Zong et al., 2019).此外, 氮添加显著降低了土壤温度, 这主要是由于氮添加促进植物生长, 提高植被盖度, 同时导致第二年凋落物输入量的进一步增多, 对土壤温度调节表现出显著的负反馈作用, 这也有助于缓解干旱年份的水分胁迫作用(Li et al., 2011; Zhao et al., 2019).我们的研究还发现氮添加并未改变凋落物输入变化对群落ANPP的影响, 二者没有显著的交互作用.与Shen等(2016)在本区域进行氮和凋落物添加实验的结果相同.这可能与半干旱草原降水量和生产力剧烈的年际变异有关, 要想进一步探讨这两者之间的交互作用是否会对系统生产力形成影响, 还需要更长期的实验数据积累减弱或去除数据分析过程中年份等其他因素的影响. ...
... ; Zhao et al., 2019).我们的研究还发现氮添加并未改变凋落物输入变化对群落ANPP的影响, 二者没有显著的交互作用.与Shen等(2016)在本区域进行氮和凋落物添加实验的结果相同.这可能与半干旱草原降水量和生产力剧烈的年际变异有关, 要想进一步探讨这两者之间的交互作用是否会对系统生产力形成影响, 还需要更长期的实验数据积累减弱或去除数据分析过程中年份等其他因素的影响. ...
Different sensitivity and threshold in response to nitrogen addition in four alpine grasslands along a precipitation transect on the Northern Tibetan Plateau 1 2019
... 以往的研究表明, 氮添加会显著提高半干旱草原生态系统生产力(潘庆民等, 2005; 沈月, 2016; Yue et al., 2016; Wang et al., 2019; Zhao et al., 2019).本研究也发现, 氮添加显著提高了土壤DOC和DON含量, 促进植物生长, 显著提高了群落生产力.内蒙古半干旱草原作为一个水氮共同限制的生态系统(Heisler-White et al., 2008; Li et al., 2011; Ren et al., 2017).氮对草原生产力的影响强度甚至方向都是依赖于生态系统的水分状况的(Yan et al., 2010; Shen et al., 2016).在本研究中, 氮添加对生产力的促进作用在降水量较高的湿润年份表现得更为明显, 而在较为干旱的年份(2016和2017年), 氮添加对ANPP的促进作用明显减弱(图3A).ANPP与降水量之间的关系也表明, 氮添加显著提高了ANPP-降水量关系的阈值, 随着降水量增加, ANPP增加的趋势更加明显, 这也进一步证实了氮添加处理对水分条件的依赖性(Yang et al., 2011; Zong et al., 2019).此外, 氮添加显著降低了土壤温度, 这主要是由于氮添加促进植物生长, 提高植被盖度, 同时导致第二年凋落物输入量的进一步增多, 对土壤温度调节表现出显著的负反馈作用, 这也有助于缓解干旱年份的水分胁迫作用(Li et al., 2011; Zhao et al., 2019).我们的研究还发现氮添加并未改变凋落物输入变化对群落ANPP的影响, 二者没有显著的交互作用.与Shen等(2016)在本区域进行氮和凋落物添加实验的结果相同.这可能与半干旱草原降水量和生产力剧烈的年际变异有关, 要想进一步探讨这两者之间的交互作用是否会对系统生产力形成影响, 还需要更长期的实验数据积累减弱或去除数据分析过程中年份等其他因素的影响. ...
 ,1,2,*
,1,2,*
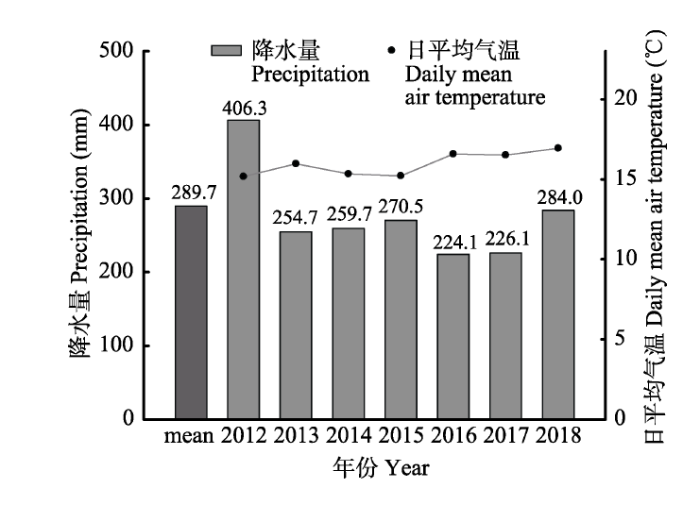 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT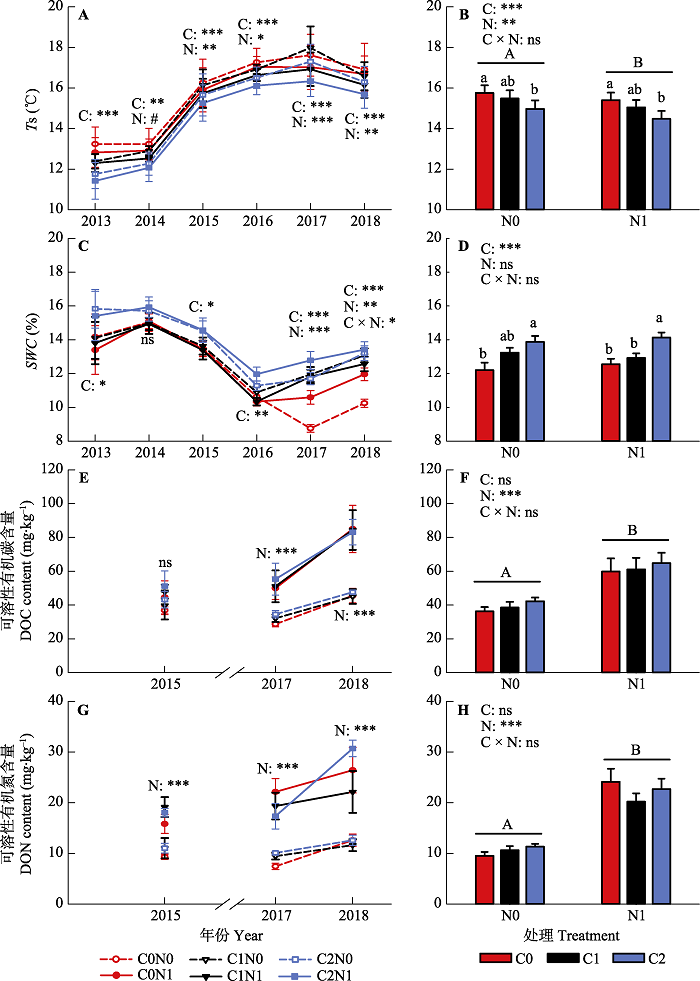 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT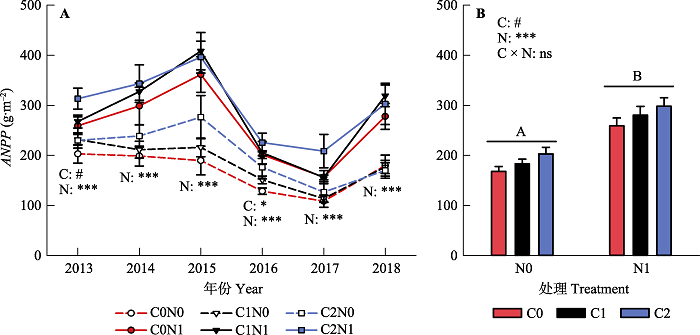 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT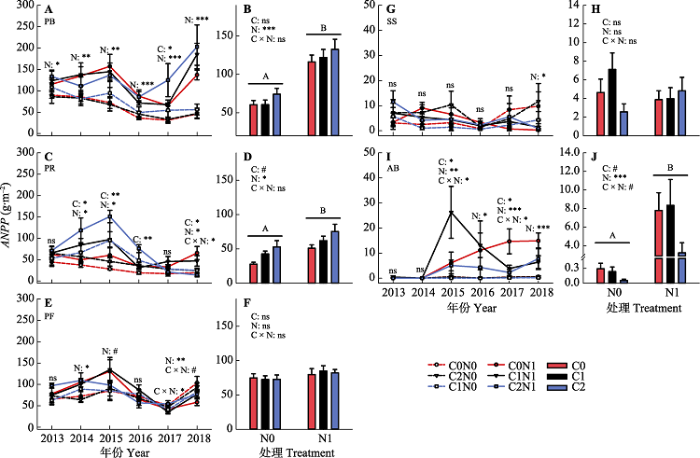 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT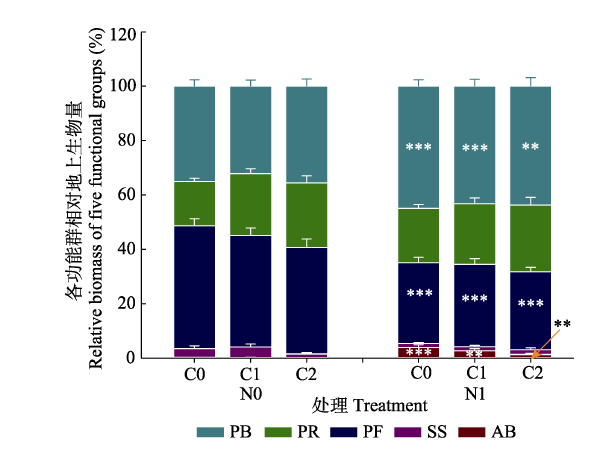 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT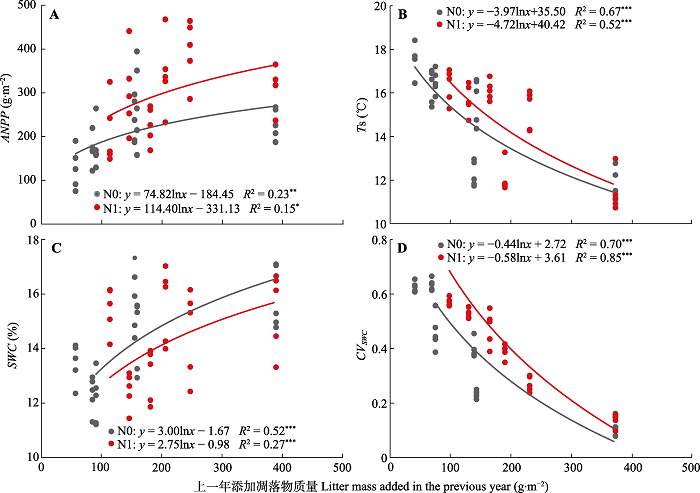 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT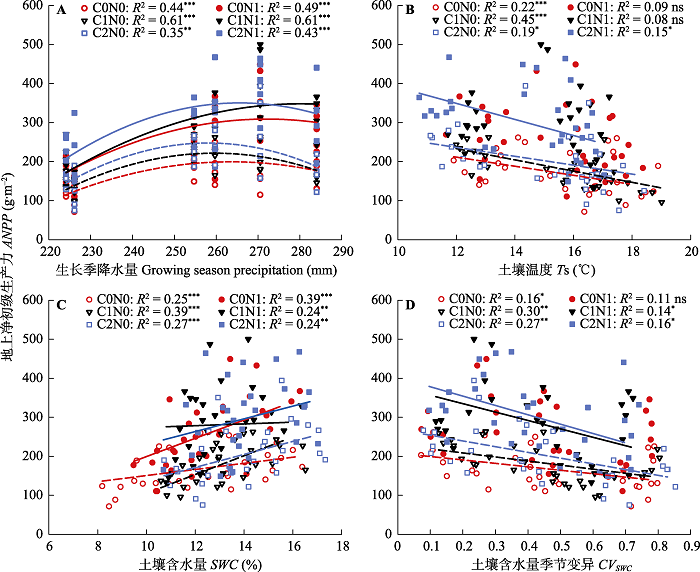 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT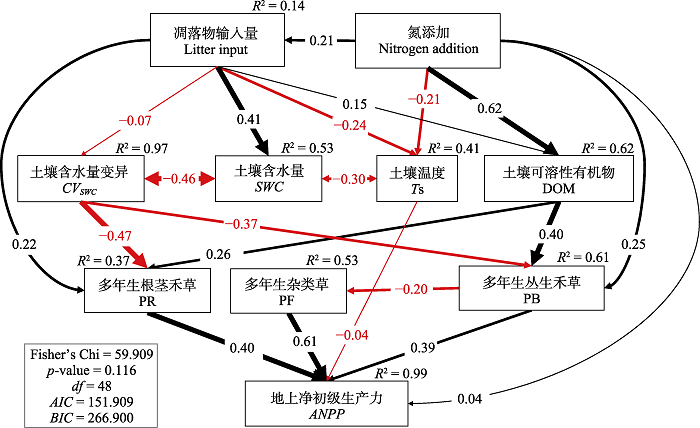 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}