 ,, 马克明
,, 马克明 ,*中国科学院生态环境研究中心城市与区域生态国家重点实验室, 北京 100085;中国科学院大学, 北京 100049
,*中国科学院生态环境研究中心城市与区域生态国家重点实验室, 北京 100085;中国科学院大学, 北京 100049Selection of null models in nestedness pattern detection of highly asymmetric mycorrhizal networks
Li-Tao LIN,, Ke-Ming MA,*State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085,China;and University of Chinese Academy of Sciences, Beijing 100049, China通讯作者: *mkm@rcees.ac.cn
编委: 周淑荣
责任编辑: 李敏, 赵航(实习)
收稿日期:2019-03-25接受日期:2019-06-05网络出版日期:2019-07-20
| 基金资助: |
Received:2019-03-25Accepted:2019-06-05Online:2019-07-20
| Fund supported: |

摘要
零模型是判定网络嵌套性的重要依据, 菌根共生关系网络经常出现高度非对称性, 该文通过探究矩阵非对称变化对基于不同零模型构建方法的网络嵌套性的影响, 试图为非对称网络零模型的选择提供依据。结果表明: 不同零模型保守性不同, 增加限定条件减少零模型构建过程中的自由空间, 高度限定条件易导致第II类错误。高度非对称网络会增加基于完全随机(r00)零模型的矩阵温度(NT)偏离、降低配对重叠度(NODF)偏离, 标准化指数z-score值显示网络非对称增加后有助于NT和NODF显著性判定。行或列限定对非对称网络嵌套性判定的影响存在差异, 列限定(c0)的网络嵌套性判定对网络非对称性变化的响应规律与r00零模型的响应趋势基本一致, 具有更低的嵌套性偏离和标准差值。行限定(r0, 包括行列限定(backtrack))零模型NT值和NT偏移随矩阵非对称性的变化保持稳定, 较之c0零模型在高度非对称网络中呈现更低的NODF偏离值。选用完全随机和限定零模型相结合的方法, 有助于更加准确判断非对称网络是否具有嵌套结构。高度非对称网络嵌套性判定中对行属性特征比较敏感, 不同非对称性网络间嵌套性水平相比较时选用r0零模型要优于r00和c0零模型。
关键词:
Abstract
Aims Null model is an important basis for nesting judgment. Highly asymmetric structures often appear in plant symbolic fungal networks. This study aims to explore the influence of matrix asymmetric changes on network nesting judgment.
Methods The study was conducted based on various null model construction methods.
Important findings Constraints vary with changing null models, with reducing null space when additional qualifications were added during null model establishment. Highly constrained nulls are prone to causing type II errors. Highly asymmetric networks increase matrix temperature (NT) deviation based on random (Equiprobable- equiprobable, r00) null model while reducing overlap and decreasing fill (NODF) deviation. Values of z-score show that highly asymmetric networks contribute to the significant determination level of NT and NODF. The impacts on the judgment of nestedness of asymmetric networks differ between row and column fixed null models. The effects of network asymmetry change on nesting detection based on column constrained (c0) nulls are similar to that of random null model, but with smaller nesting deviation and standard deviations. No significant differences in both NT and NT deviations were observed among different asymmetry networks based on the row fixed (r0) nulls, with a lower NODF deviation in highly asymmetric network based on c0 nulls. To more accurately determine whether the asymmetric networks would have nested structures, we recommend using a combination of random and constrained null models. Our results also demonstrate that the r0 null model performs better than either the r00 null model or the c0 null model when comparing nesting level of different asymmetric networks.
Keywords:
PDF (2747KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
林力涛, 马克明. 菌根共生网络嵌套性判定的零模型选择. 植物生态学报, 2019, 43(7): 611-623. DOI: 10.17521/cjpe.2019.0065
LIN Li-Tao, MA Ke-Ming.
群落中物种之间通过相互作用(如捕食、竞争、寄生、互惠等)彼此关联构成网络(Bascompte, 2010; Thebault & Fontaine, 2010)。将网络分析方法应用于生态学研究由来已久(陈水华和王玉军, 2004; Ulrich et al., 2009; 李中才等, 2011), 其中嵌套性是互惠共生关系网络(传粉、种子传播、菌根共生等)的典型特征(Ulrich et al., 2009; 方强和黄双全, 2012), 即相互作用关系中相对特化种(specialist)相互作用的物种是相对泛化种(generalist)相互作用的物种子集的作用模式。为对相互作用关系类型进行甄别, 需要通过建立零模型(null model)来对网络嵌套结构进行显著性检验(Bascompte et al., 2003; Ulrich & Gotelli, 2007)。当前, 不同零模型构建方法均在传粉网络和种子传播网络(具有较低的非对称性)嵌套性判定中应用较为成熟, 而基于不同零模型的网络非对称性变化对嵌套性判定影响的研究相对较少。近年来, 高通量技术的进步使得微生物信息获取量呈指数式增长(Liu et al., 2012; Poisot et al., 2013), 在构建菌根共生关系网络时, 宿主植物物种数量与共生真菌操作分类单元(OTUs)数量之间可能出现数量级的差异(Bahram et al., 2014), 导致网络呈现高度的非对称性。相较于传统近方形的二分网络, 高度非对称网络对零模型构建自由空间具有较强的限定作用(Chagnon et al., 2016)。因此, 明确矩阵非对称变化对基于不同零模型嵌套性判定的影响规律有助于高度非对称网嵌套性结构判定中选取适宜零模型, 提高网络嵌套结构判定的精准度。
不同的零模型检验反映的生态学意义存在区别(Atmar & Patterson, 1993; Wright et al., 1997; Ulrich et al., 2009; Kennedy et al., 2014), 选取适当的零模型有助于准确揭示类群间相互作用关系的维持机制。目前, 基于被动取样的嵌套性判定的零模型选择已经进行了较多的研究。如完全随机零模型认为同一类群内的各元素以相同的概率与另一类群中任意元素建立共生关系(Atmar & Patterson, 1993); 在传粉网络研究中, 数量多且(或)分布广泛的物种更容易被调查观测到, 需要通过增加限定零模型构建条件(行列限定)抵消由于被动取样对共生关系嵌套性判定的影响(Bascompte et al., 2003); 在探究海岛距离对鸟类栖息地嵌套性的影响时, 将海岛承载能力作为零模型构建限定因子(Vázquez et al., 2009; 罗芳等, 2013; 张雪梅等, 2016)等。然而, 关于非对称矩阵嵌套性判定中零模型的选择缺乏深入探究(Blüthgen et al., 2007), Almeida-Neto等(2008)指出矩阵形状、大小、填充度等因素均是影响矩阵嵌套性度量值的重要因素, 矩阵非对称变化后对不同限定零模型的构建自由度的影响程度存在差异(Chagnon et al., 2016)。因此, 选取适宜的零模型对于非对称菌根共生关系网络嵌套性判定具有重要价值。当前, 在非对称网络属性特征方面已经进行了一些基础研究, 如Blüthgen等(2007)对51个动植物间互惠网络分析显示随矩阵非对称性增加网络特异性水平升高, Almeida-Neto等(2008)指出零模型构建方法是影响不同形状随机矩阵嵌套性度量值保守性的重要因素, 但仍缺乏基于多种零模型选择的网络非对称变化对嵌套性判定影响的研究。
目前, 已有相当多的研究显示宿主植物可与大量不同真菌建立共生关系(Bahram et al., 2011; Gao et al., 2015), 即基于高通量测序结果的植物共生真菌关系网络呈现高度非对称性(Bahram et al., 2014)。同时, 进行关系网络构建时也会面临相互作用物种数量不保守情形, 如Toju等(2014)在构建外生菌根(EM)、丛枝菌根(AM)等关系子网络时面临共生真菌物种数存在差异的问题。共生关系网络中, 物种连接点数是直接反映物种间相互作用关系强度的重要指标, 然而针对不同形状、大小网络, 该指标可能存在上限不保守的问题; 灰度填充度是表征Atmar和Patterson (1995)研究中287个网络分析显示, 行数相近的各组网络其灰度填充度与非对称性之间相关性未达到显著水平(附录I), 即行数相近时网络内相对作用强度随矩阵非对称性变化差异不显著。为此, 本文以灰度填充度保守为前提进行非对称网络的阐述, 假定行元素表征宿主植物物种, 列元素表征共生真菌, 通过物种总数保守和宿主植物物种总数保守两种方式构建不同非对称性网络, 探究矩阵非对称变化对网络嵌套性判定的影响及其在不同限定零模型中的表现, 为高度非对称共生关系网络嵌套性判定零模型选择提供借鉴。
1 研究方法
1.1 关系矩阵的构建
本研究中所有运算均在统计分析软件R 3.5.2 (https://cran.r-project.org)中实现。随机矩阵由vegan 2.5-4包(Oksanen et al., 2019)中nullmodel命令创 建, 用method参数选用完全随机(equiprobable row-equiprobable column, r00)、行限定(fixed row-equiprobable column, r0)、列限定(equiprobable row-fixed column, c0)和行列限定(fixed row-fixed column, backtrack)等算法构建定性矩阵(附录II)。嵌套性指数用bipartite 2.11包(Dormann et al., 2018)中nested命令实现, method参数确定嵌套性指标类型。本文中, 所有选用和构建的矩阵均假定行元素表征植物、列元素表征菌根真菌, 通过改变列数/行数比值来模拟关系网络中共生真菌OTUs分类单元数远高于宿主植物物种数量的情形。以填充度50%、行列数10 × 10完全嵌套和均匀分布的矩阵为初始矩阵(IM), 初步探析不同零模型构建方法对矩阵嵌套性和反嵌套性判定的影响(附录II)。在矩阵非对称变化对嵌套性判定影响中, 为保证矩阵非对称变化后具有相近的嵌套性水平, 使其更具可对比性, 本文选用了物种总数保守和宿主植物物种总数保守两种网络构建方式:
(1)物种总数保守: 以物种总数60, 行列数30/30、25/35、20/40、15/35、10/50、5/55, 填充度50%左右的600个(每水平100个)完全随机r00矩阵作为初始矩阵, 用以表征不同非对称性同时具有相同嵌套水平的网络。
(2)宿主植物物种总数保守: 本研究选取bipartite包中5个最接近方形互惠关系网络作为初始矩阵(附录III), 假定行元素表征植物、列元素表征菌根真菌, 基于初始矩阵通过列合并方式产生1倍、5倍、10倍的初始矩阵, 探究矩阵非对称性变化对网络嵌套性判定的影响, 及零模型选择对其的影响。通过列合并的方式产生矩阵, 虽然物种连接强度发生变化, 但其具有相同水平的相对连接强度, 即网络具有相同的灰度填充度水平。列合并非对称网络中共生真菌和宿主植物在选择合作对象时与近方形网络具有相同的作用格局, 因而, 用其来表征具有相同嵌套性水平但不同非对称性水平的关系矩阵。
1.2 嵌套性指标选择
矩阵温度(NT)和配对重叠度(NODF)是两种较为常用的计算网络嵌套性的方法(表1)。T由Atmar和Patterson (1993)提出, 是针对矩阵与相同大小且填充度完全嵌套矩阵之间偏离的计算指标, 由于T值随嵌套性增加而下降, 我们用NT = 100 - T转换来表征基于矩阵温度的嵌套性指标(Bascompte et al., 2003; Almeida-Neto et al., 2008)。调整矩阵行列顺序使灰度单元格尽可能地排在矩阵的左上角, 辨识矩阵中与其相应嵌套矩阵不同的“缺失”和“多余”, 计算与其相应最大程度嵌套矩阵的偏离:式中, uij为矩阵中位于i行j列的意外值(“缺失”和“多余”); Dij为贯穿ij点对角线距离; dij为ij点沿这条线的特定距离。
Table 1
表1
表1嵌套性度量指标特征
Table 1
| 指标 Index | 反映特征 Underlying feature | 下限保守 Bottom boundary conservative | 上限保守 Top boundary conservative | 零模型保守 Null model conservative | 存在上限 Top limit |
|---|---|---|---|---|---|
| 矩阵温度 matrix temperature (NT) | 矩阵中异常点数量、位置 Relative abundance and position of unexpected absences or presences in a matrix | 是 Yes | 是 Yes | 否 No | 是 Yes |
| 配对重叠度 nestedness metric based on overlap and decreasing fill (NODF) | 矩阵配对重叠中非子集行列的数量及位置 Percentage and position of columns (rows) overlaps with other columns (rows) | 是 Yes | 是 Yes | 是 Yes | 是 Yes |
| 嵌套性偏移 Nestedness deviation | 网络与零模型的距离 Distance deviates from null models | 是 Yes | 否 No | 是 Yes | 是 Yes |
| 标准化指数 Standardized effect size | 网络达到显著水平的难易程度 Significance of a network deviates from null models | 是 Yes | 否 No | 是 Yes | 否 No |
新窗口打开|下载CSV
式中, f为行数, P为列数, U为总意外值, 为两次标准化后的指标, 不受矩阵大小和性状约束; 矩阵完全嵌套时U值等于0, 矩阵完全无序化Um ≈ 0.04145。
式中, k = 100/Umax, T值的变化范围是从0° (矩阵完全嵌套)到100° (矩阵完全无序), 且不受矩阵大小约束。
配对重叠度由Almeida-Neto等(2008)基于矩阵两个基本性质, 填充逐渐下降(DF)和配对重叠(PO), 针对矩阵中物种相互配对重叠比例提出的计算指标。调整矩阵物种填充(1’s)尽可能地排在矩阵的左上角, 矩阵中行总和、列总和逐渐递减; 假设调整后的m × n矩阵, i行在j行上方, k列在l列左方, MT为行或列的填充之和; 对任意两行, 如果MTj < MTi, 则DFij等于100, 否则DFij为0; 同样, 矩阵中任意两列MTl < MTk, 则DFkl等于100, 否则DFkl为0。列配对重叠POkl表示l列中1’s在k列中具有相同1’s所占的比例, 行配对重叠POij表示j行中1’s在i行中具有相同1’s所占的比例。配对嵌套程度Npaired定义为: 若DFpaired = 0, 则Npaired = 0; 若DFpaired = 100, 则Npaired = PO。
式中, NODF值的变化范围是从0 (矩阵完全无序)到100 (矩阵完全嵌套), 且不受矩阵大小约束。
为方便不同矩阵嵌套性对比, Ulrich和Gotelli (2007)提出标准化指数(SES)记为z-score = (x - μ)/σ,其中x为实际观测值, μ为100个模拟值矩阵嵌套性平均值, σ为100个模拟矩阵嵌套性标准差。当z-score高于或低于±1.96时达到统计学双尾检验0.05显著水平, 认为该网络具有显著(反)嵌套结构。除直接的显著水平比较之外, 这里将嵌套性偏移定义为矩阵真实值与模拟矩阵平均值的差值, 能够量化显示矩阵嵌套性值与零模型值数值差距(Montesinos-?Navarro et al., 2012)。
2 结果
2.1 不同零模型嵌套性度量值变化特征
不同零模型相比较, 高嵌套性和均匀分布(高模块性)矩阵均表明行列频度(关系矩阵中非0单元格的个数)限定零模型(backtrack)能够较大程度上保留初始矩阵特征属性; 同时, 高度规整的矩阵具有较小的零模型构建自由空间, 其标准差接近于0 (图1)。完全随机零模型(r00)在嵌套性或非嵌套性判定中更易达到显著水平, 其嵌套性水平仅与初始矩阵内部元素分布格局相关, 以高嵌套或均匀分布矩阵为初始矩阵构建r00零模型具有相同的NT值及NODF值。行(r0)或列(c0)频度限定零模型较r00保守性高, 表明矩阵需要更高的嵌套性(反嵌套性)特征时与零模型对比才能达到显著水平, 如图1B中行(列)频率总和限制零模型显著高于(低于)完全随机零模型。图1
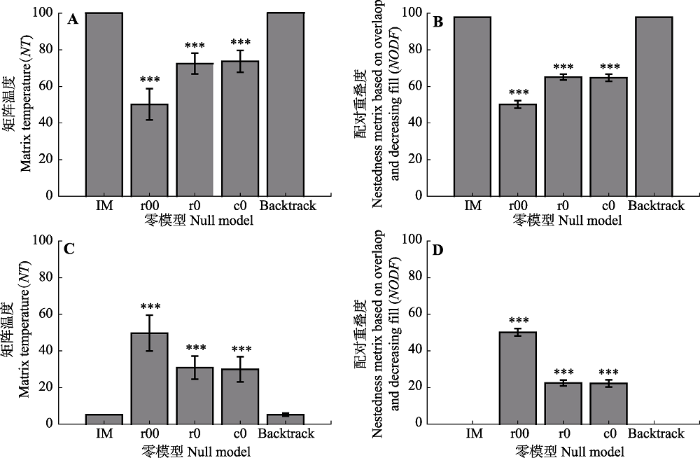 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1基于完全嵌套(A, B)和均匀分布(C, D)矩阵构建的不同零模型嵌套性特征(平均值±标准偏差)。Backtrack, 行列限定; c0, 列限定; IM, 初始矩阵; r0, 行限定; r00, 完全随机。***, 0.001水平显著。
Fig. 1Absolute values of nestedness from null models constructed based on fully nested (A, B) and uniformly distributed matrices (C, D) (mean ± SD). Backtrack, fixed row-fixed column; c0, equiprobable row-fixed column; IM, initial matrix; r0, fixed row- equiprobable column; r00, equiprobable row-equiprobable column. ***, significant at 0.001 level.
2.2 矩阵行列属性对完全随机矩阵嵌套值的影响
根据嵌套性的定义, 认为不同非对称性各组随机矩阵具有相同的嵌套性显著水平。由图2可知, 不同嵌套性度量指标对矩阵非对称性变化相应存在区 别, NT随矩阵非对称性增大呈显著增加的趋势, 对矩阵形状具有显著的依赖性; 而矩阵非对称性变化对NODF指标无显著影响, 二者相关性未达到显著水平。在嵌套显著水平判定中, 基于r00、r0、c0零模型的标准化两种度量指标z-score指数随矩阵非对称变化均能保持稳定, 然而基于高度限定的bactrack零模型的z-score指数随矩阵非对称性增加呈下降的趋势。图2
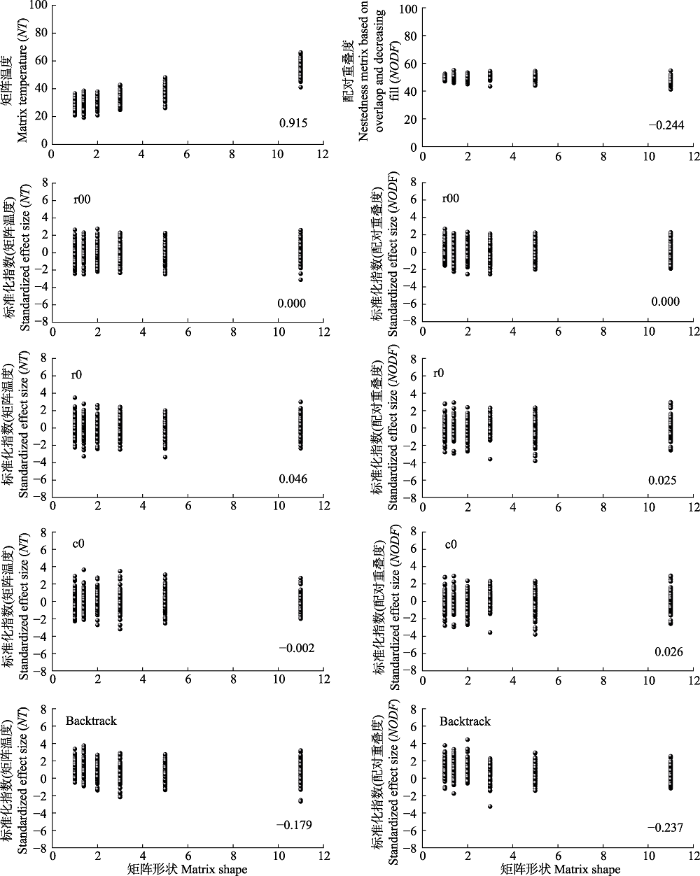 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2矩阵形状变动对矩阵嵌套性指数影响。矩阵形状, 矩阵列数/矩阵行数。Backtrack, 行列限定; c0, 列限定; r0, 行限定; r00, 完全随机。
Fig. 2Effects of changes in matrix shape on nestedness metrics. Matrix shape, number of columns/number of rows. Backtrack, fixed row-fixed column; c0, equiprobable row-fixed column; r0, fixed row-equiprobable column; r00, equiprobable row-equiprobable column.
2.3 非对称性对不同零模型矩阵温度的影响
行元素总数限定的网络中, 基于不同零模型构建方法NT值对矩阵非对称性变化的响应规律存在差异(图3)。r0和backtrack零模型NT值对矩阵非对称变化响应不敏感; 而r00和c0零模型NT值随矩阵非对称性增加而降低, 为IM > 5倍>10倍, 这可能是与矩阵属性特征改变相关(图1)。嵌套性偏移显示, r0和backtrack构建矩阵的NT偏移在非对称性发生变化后保持稳定; 而r00和c0构建矩阵的NT偏移值随矩阵非对称性增加而逐渐增加, 与NT值下降趋势相对应。此外, c0零模型对不同对称性矩阵NT偏移影响存在差异, 与完全随机r00方法相比, IM矩阵NT降低(34.37 ± 17.40)%, 而5倍和10倍矩阵NT偏移较之r00矩阵仅降低(13.40 ± 16.34)%和(13.15 ± 15.55)%。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3非对称性变化对构建二分零模型矩阵温度(NT)和NT偏移的影响(平均值±标准偏差)。Backtrack, 行列限定; c0, 列限定; r0, 行限定; r00, 完全随机。IM, 初始矩阵; 5 times, 5倍列合并网络; 10 times, 10倍列合并网络。
Fig. 3Effects of matrix asymmetric variations on matrix temperature (NT) and NT deviation based on different binary matrix construction methods (mean ± SD). Backtrack, fixed row-fixed column; c0, equiprobable row-fixed column; r0, fixed row-equiprobable column; r00, equiprobable row-equiprobable column. IM, Initial matrix; 5 times, 5 times column merge network; 10 times, 10 times column merge network.
标准化指数(z-score)显示, 不同零模型构建方法对矩阵嵌套性显著性判定具有较大的影响(图4)。如基于r00零模型显著的olesen2002flores和mosquin1967网络在对比r0零模型时其z-score接近0, 即行属性特征是导致网络矩阵嵌套性的因素, 对行属性限定后其嵌套性未达到显著水平。参照backtrack零模型的bezerra2009网络在与其他零模型相比较时其嵌套显著水平发生转变, 较之r00较高的z-score指数以及r0、c0达到显著嵌套水平, backtrack具有更高的矩阵嵌套判定保守性。随初始矩阵非对称性增加, 基于r00和c0零模型z-score呈增加趋势; 而基于r0零模型, 嵌套性中性的网络在非对称性发生变化后z-score指数基本稳定, 显著嵌套的网络在非对称变化后z-score指数呈增加趋势; 基于backtrack构建方法z-score指数变化与r0具有相近的变化趋势, 但其z-score绝对值低于r0。
图4
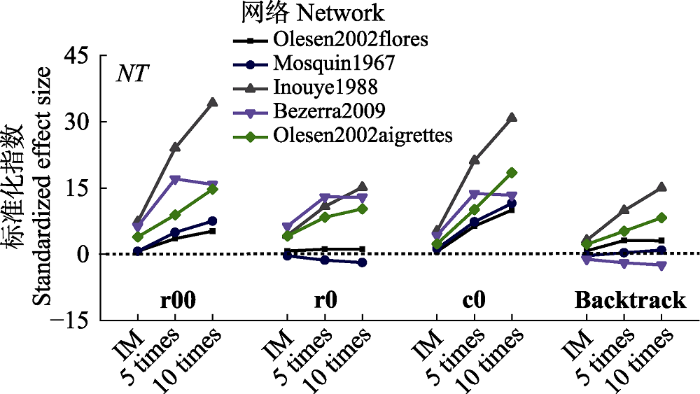 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4非对称性变化对矩阵温度标准化指数的影响。Backtrack, 行列限定; c0, 列限定; r0, 行限定; r00, 完全随机。IM, 初始矩阵; 5 times, 5倍列合并网络; 10 times, 10倍列合并网络。
Fig. 4Effects of matrix asymmetric variations on the standardized effect size (z-score) of matrix temperature (NT). Backtrack, fixed row-fixed column; c0, equiprobable row-fixed column; r0, fixed row-equiprobable column; r00, equiprobable row-equiprobable column. IM, initial matrix; 5 times, 5 times column merge network; 10 times, 10 times column merge network.
2.4 非对称性对不同零模型配对重叠度的影响
NODF是基于配对重叠的嵌套性度量指标, 不同零模型构建方法NODF值对矩阵非对称性变化响应亦存在差异(图5)。矩阵非对称性对r00零模型NODF值无显著影响, 而r0、c0和backtrack零模型NODF值随矩阵非对称变化表现不同: 网络在r0零模型随对称性增大NODF值表现为增加趋势; 在c0零模型中表现为下降趋势; backtrack零模型NODF值变化特征介于二者之间, 如r0增幅较大的olesen2002flores网络在backtrack零模型中随非对称增加呈增大的趋势, c0降幅较大的bezerra2009和olesen2002aigrettes网络表现为下降趋势。结果显示, 基于c0零模型的NODF偏移随网络非对称性变化基本保持稳定, 而基于r00、r0和backtrack零模型的NODF偏移随网络非对称性变化呈下降的趋势。相较于r0构建方法NT偏移基本稳定, NODF偏移随网络非对称性增加呈现较大幅度的下降。基于backtrack零模型的NODF偏移随网络非对称增加逐渐降低至负值, 即呈现反嵌套模式。图5
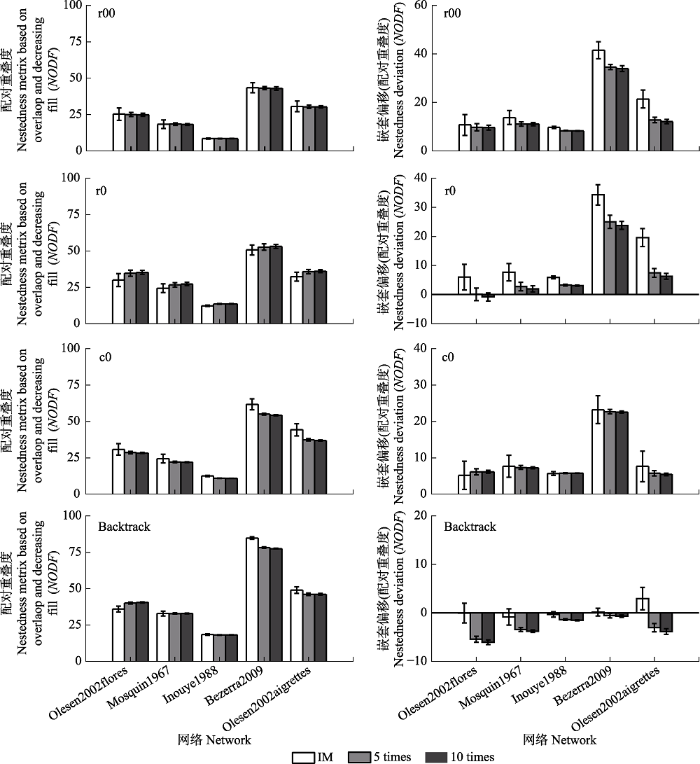 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5非对称性变化对构建二分零模型配对重叠度(NODF)和NODF偏移影响(平均值±标准偏差)。Backtrack, 行列限定; c0, 列限定; r0, 行限定; r00, 完全随机。IM, 初始矩阵; 5 times, 5倍列合并网络; 10 times, 10倍列合并网络。
Fig. 5Effects of matrix asymmetric variations on nestedness metric based on overlap and decreasing fill (NODF) and NODF deviation based on different binary matrix construction methods (mean ± SD). Backtrack, fixed row-fixed column; c0, equiprobable row-fixed column; r0, fixed row-equiprobable column; r00, equiprobable row-equiprobable column. IM, Initial matrix; 5 times, 5 times column merge network; 10 times, 10 times column merge network.
标准化指数(z-score)显示, 不同零模型构建方法的矩阵嵌套性显著水平可能发生改变, 如inouye1988网络z-score指数在r00、r0和c0零模型比对中达到嵌套性显著嵌套水平, 而在backtrack零模型比对中呈现随机或显著反嵌套结构。如图6所示, 随矩阵非对称性增加基于r00和c0零模型z-score呈增加趋势, 而基于r0方法网络矩阵在非对称性发生变化后z-score基本保持稳定。与NT值z-score不同, 基于backtrack构建方法的NODF值z-score在矩阵非对称性增加后对矩阵嵌套性判定中更为保守, 而对反嵌套性判定更为敏感, 如backtrack方法olesen2002aigrettes网络嵌套性的显著性发生转变, NODF值z-score低于NT值z-score。
图6
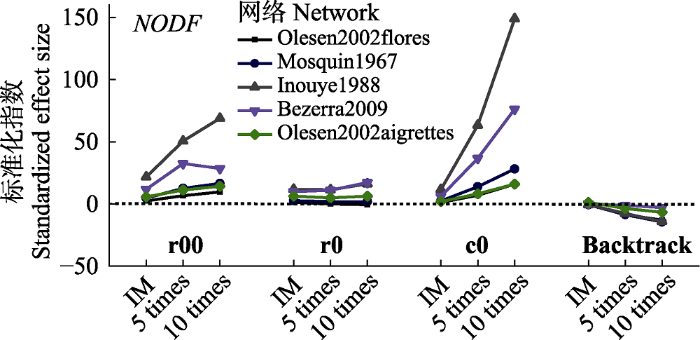 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6非对称性变化对矩阵配对重叠度标准化指数的影响。Backtrack, 行列限定; c0, 列限定; r0, 行限定; r00, 完全随机。IM, 初始矩阵; 5 times, 5倍列合并网络; 10 times, 10倍列合并网络。
Fig. 6Effects of matrix asymmetric variations o z-scores of on the standardized effect size (z-score) of nestedness metric based on overlap and decreasing fill (NODF). Backtrack, fixed row-fixed column; c0, equiprobable row-fixed column; r0, fixed row-equiprobable column; r00, equiprobable row- equiprobable column. IM, Initial matrix; 5 times, 5 times column merge network; 10 times, 10 times column merge network.
3 讨论
3.1 不同零模型在嵌套性判定中保守性的差异
零模型是进行嵌套性判定的重要依据(Ulrich & Gotelli, 2007; Ulrich et al., 2009), 通过零模型比较可以看出: r00零模型具有较高的构建自由空间, 受初始矩阵内单元格嵌套性或反嵌套性分布格局影响较小(图2), 仅与矩阵自身填充度相关(Almeida-Neto et al., 2008)。在探究菌丝网络中共生真菌在网络中作用差别时, 通过与r00对比显示部分共生真菌在整个菌丝网络中可能起关键节点作用(Southworth et al., 2005)。通常认为嵌套性检验中最好的零假设可能是所有的物种具有相同的出现概率(Cook & Quinn, 1998), 然而, 受取样因素的影响, 稀有物种之间的相互作用往往难以被直接观测到(Vázquez et al., 2009)。因此, 需要通过增加零模型行(列)限定条件来抵消被动取样对嵌套性判定的影响(Gotelli, 2000)。近方形的传粉网络或种子传播网络中, 植物限定的r0和传粉昆虫或食果动物限定的c0零模型较完全随机r00零模型保守性高, 应用该方法生成的随机矩阵能够保留初始矩阵中植物(传粉昆虫或食果动物)的部分特征, 其嵌套性度量值较之r00零模型更为接近初始矩阵真实值(图1, 图3, 图5)。限定零模型应用较为广泛, 鸟类岛屿生物地理学研究中常利用该种方法将海岛承载能力作为构建零模型的限定因素用于探究距离因子对鸟类栖息地嵌套性的影响(Vázquez et al., 2009; 罗芳等, 2013; 张雪梅等, 2016)。此外, 当二分网络相互作用两大类群其中一个类群的各物种为均等取样时, 即该类群在构建共生关系网络时不受种群数量差异的影响, 可使用r0或c0零模型去除被动取样因素影响并进行网络显著性的检验(Ulrich & Gotelli, 2013)。高度限定的backtrack零模型较之r0和c0方法又具有更高的保守性, 适用于相互作用两大类群在构建关系网络时均受被动取样影响的情况, 如传粉网络调查取样过程中数量多且(或)分布广泛的植物或传粉昆虫种更容易被调查观测到(Bascompte et al., 2003), 但该方法对于高度规整网络嵌套性检验中具有一定局限性(Chagnon et al., 2016), 易引发第II类错误(图1), 即显著嵌套的关系网络在统计分析过程中判定为未达到嵌套性显著水平, 过低估计网络嵌套结构的显著性水平。嵌套性是互惠关系网络的典型特征, 基于不同零模型构建方法网络的嵌套性水平可能会发生改变, 如Bascompte等(2003)对52个食果、传粉网络嵌套性分析发现当参照零模型由r00变为backtrack时有6个网络嵌套性水平发生改变; Kennedy等(2014)的研究也表明backtrack零模型在嵌套性判断中具有较高的保守性, 基于backtrack方法未达到显著的网络在与r0零模型对比后达到嵌套性显著水平。本研究亦显示出类似规律, 如bezerra2009网络嵌套性NT在r00、r0、c0构建方法对比中达到显著水平, 而与backtrack零模型对比后未达到显著水平; 甚者, NODF嵌套性衡量方法中bezerra2009网络在backtrack零模型对比后呈现显著的反嵌套水平。
3.2 非对称网络与传统近方形网络在嵌套性判定中的不同
矩阵嵌套性度量指标对矩阵属性变化的响应存在差异(Ulrich & Gotelli, 2007), 完全零模型大小及形状改变后NODF值无显著变化, 而NT值与矩阵形状呈显著正相关关系(图2), 与Almeida-Neto等(2008)的研究结果相一致。宿主植物总数保持不变时, r00零模型NT值随矩阵非对称性增加呈降低的趋势, 而NODF值基本保持稳定(图3, 图5)。两种嵌套性方法相比较, 随矩阵有序程度降低至完全随机分布, NT值在测定高度有序水平矩阵时发挥出色, 但其在完全随机分布时NT度量值不保守; 而NODF值在低有序水平度量时发挥出色, 矩阵随机分布时具有相同的NODF度量值(图7, 图8)。虽然NT和NODF对矩阵非对称性变化相应存在差异, 但在与r00零模型进行显著性检验时, 网络非对称变化对各组矩阵NT和NODF显著性检验均无显著的影响(图2), 二者的z-score指数均能呈现相等的下限, 但随矩阵有序程度增加, z-score指数上限不保守, 近方形网络z-score指数要高于非对称性网络(图9; 表1), 即基于r00的z-score指数可在显著性判定中进行应用, 在不同非对称性的高度有序网络间进行比较时仍具有一定局限性。图7
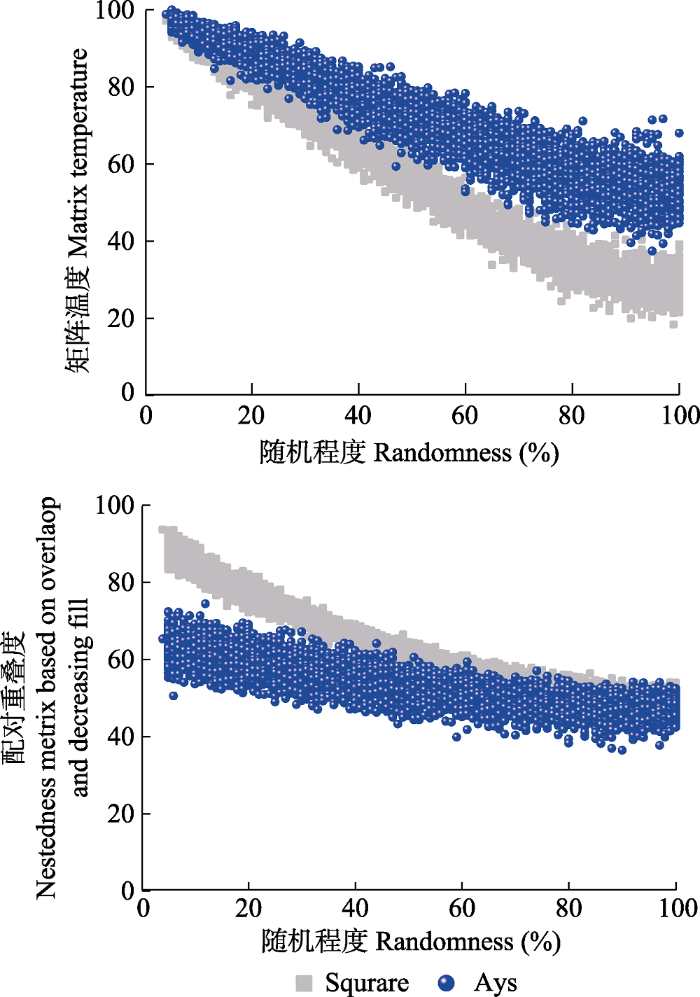 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7矩阵随机化程度对嵌套性度量指标的影响。Square, 对称网络, 行列数30/30; Asy, 非对称网络, 行列数5/55。随机化程度, 网络中无序单元格的数量/单元格总数量。随机程度包含4:100 (%)共计97水平, 每水平50个重复, 共计4 850矩阵。
Fig. 7Effects of the level of matrix randomness on nestedness metrics. Square, symmetric networks with rows and columns 30/30; Asy, asymmetric networks with rows and columns 5/55. Randomness, percentage of disordered cells deviates from fully nested matrix. Data contains a total of 4 850 matrices, with 4:100 (%) randomness degree 97 levels of, 50 repetitions per level.
图8
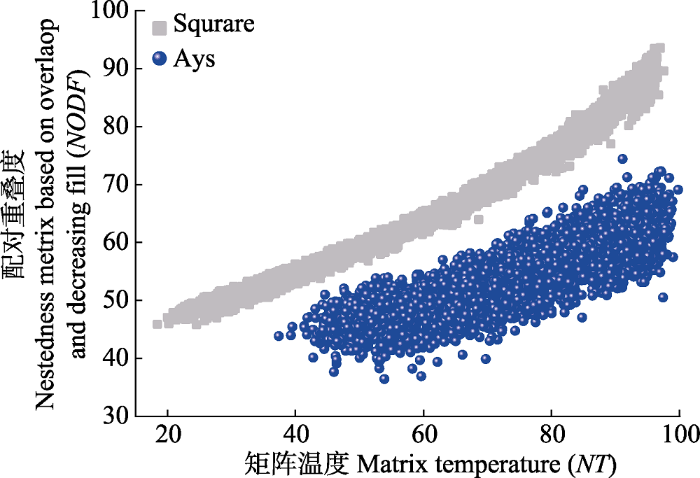 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8矩阵温度(NT)与配对重叠度(NODF)间的相关性。Square, 对称网络, 行列数30/30; Asy, 非对称网络, 行列数5/55。随机化程度, 网络中无序单元格的数量/单元格总数量。随机程度包含4:100 (%)共计97水平, 每水平50个重复, 共计4 850矩阵。
Fig. 8Correlation between matrix temperature (NT) and nestedness metric based on overlap and decreasing fill (NODF). Square, symmetric networks with rows and columns 30/30; Asy, asymmetric networks with rows and columns 5/55. Randomness, percentage of disordered cells deviates from fully nested matrix. Data contains a total of 4 850 matrices, with 4:100 (%) randomness degree 97 levels of, 50 repetitions per level.
图9
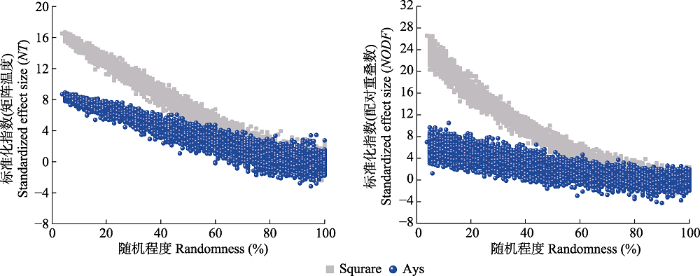 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9矩阵随机化程度对嵌套标准化指标影响。Square, 对称网络, 行列数30/30; Asy, 非对称网络, 行列数5/55。随机化程度, 网络中无序单元格的数量/单元格总数量。随机程度包含4:100 (%)共计97水平, 每水平50个重复, 共计4 850矩阵。NT, 矩阵温度; NODF, 配对重叠度。
Fig. 9Effects of the level of matrix randomness on the standardized effect size of nestedness. Square, symmetric networks with rows and columns 30/30; Asy, asymmetric networks with rows and columns 5/55. Randomness, percentage of disordered cells deviates from fully nested matrix. Data contains a total of 4 850 matrices, with 4:100 (%) random-ness degree 97 levels of, 50 repetitions per level. NT, matrix temperature; NODF, nestedness metric based on overlap and decreasing fill.
增加限定条件的零模型构建方法能够去除取样环节中被动取样因素的影响, 有助于更加保守地判定物种间相互作用的本质(Ulrich et al., 2009)。同时, 增加限定条件的同时将会极大减少零模型构建过程中的自由空间(Chagnon et al., 2016), 对于低填充度、高非对称网络选用限定零模型不能有效地随机化共生关系矩阵。物种总数一定的随机网络中, 基于backtrack零模型NT和NODF值的z-score指数在提高判定保守性的同时, 容易引发第II类错误; 从backtrack零模型构建自由度来看, NT和NODF值的z-score指数与矩阵非对称性负相关, 表明高度非对称网络较之近方形网络保留更多原始矩阵的信息(图2)。本研究显示, 利用backtrack零模型对高度规整网络进行显著性验证时亦存在较大的局限性(图1, 图5), 完全随机的零模型是较好的替代选择(Ulrich & Gotelli, 2013), 利用r00方法生成矩阵NODF值随矩阵非对称性变化基本稳定, NT值随矩阵非对称性增大而减小,NT偏移和z-score随非对称性增大而增大, 均未对网络嵌套性判定产生负效应。
行(列)限定对矩阵非对称变化后嵌套性判定影响不同, 本研究中, 行限定的r0和backtrack零模型NT值和NT偏移随网络非对称性增加无显著变化, 而列限定c0零模型NT值和NT偏移变化与r00零模型结果相近(图3)。基于矩阵温度的嵌套性判定方法在高度非对称矩阵嵌套性判定中对行属性特征更敏感, 即宿主植物行属性特征是影响非对称网络零模型构建自由度的主导因素。NODF值对矩阵大小及形状不敏感, 但基于r00方法生成矩阵NODF偏移随矩阵非对称性增加呈下降趋势, 与NT指标趋势相反。这主要是由于二者对待调整后矩阵中互为子集列(行)处理方式不同(表1), NT强调异常点出现点数量和位置, 而NODF将相同列(行)结构视为妨碍矩阵嵌套的结构(Ulrich et al., 2009)。随非对称性增加基于backtrack零模型的NODF偏移表现出与r00方法相反的结果, 如基于backtrack方法的bezerra2009网络嵌套偏移显示由嵌套向反嵌套方向发生转变(图5)。此外, 矩阵非对称性变化对基于不同零模型的z-score指数影响存在差异, 行限定的r0方法网络矩阵非对称性变化z-score基本稳定, 列限定z-score指数随矩阵非对称增加而呈现升高趋势。这可能与不同零模型生成矩阵的标准差存在差异相关联, c0零模型构建自由度低于r00零模型导致c0零模型生成随机矩阵的标准差相对较小, 当标准差的变化超过嵌套性偏移的变化时z-score指数发生相应改变。
3.3 单网络嵌套性的判定及多网络间的比较
菌根非对称网络嵌套性判定中, 兼顾保守型的同时也需要警惕第II类错误的发生, 可以采用多个零模型相结合的方法进行验证。同时, 集合种群嵌套性研究指出, 对于可能影响物种分布的因素(如生境大小、隔离程度及其质量等)都是当嵌套性在统计上被证明以后才考虑的问题(刘灿然等, 2002)。完全随机零模型(r00), 假定同一类群内所有物种具有相等概率与另一类群内任一物种建立联系, 具有较高的零模型构建自由度(图2), 可以先将菌根非对称网络通过与r00零模型比较判定网络的嵌套性情况, 然后对嵌套性的成因进行解析。经由r00检验后达到嵌套性显著的网络仍无法确定相互作用的双方是否存在彼此选择的倾向性(Ulrich et al., 2009), 被动取样亦是导致网络嵌套性的可能因素。后续嵌套性检验中, 可通过构建限定零模型来去除物种的出现频率差异影响(Bahram et al., 2011), 探究导致网络出现嵌套性结构的原因。在检验过程中, 行或列限定零模型对高度非对称网络嵌套性判定的影响存在差异(图3, 图5), 这主要与矩阵本身宿主植物行或共生真菌列自由度空间差异有关。高度非对称矩阵嵌套性判定中对宿主植物行属性特征更敏感(Beiler et al., 2015), 与r00方法相比呈现更低的嵌套性偏离值(图3, 图5)。不同于昆虫传粉网络、动物种子传播网络, 在植物-共生真菌网络研究中由宿主植物频度差异部分产生的影响能够在取样过程中予以控制(Ulrich & Gotelli, 2013), 因此, 被动取样影响主要来自于共生真菌出现频率的差异, 可选用c0零模型对网络中共生真菌物种出现频率进行限定, c0零模型较大程度地保留了r00方法的特征, 具有较高的零模型构建自由度(图3)。如果不同零模型嵌套性判定结果存在差异且限定零模型方差值很小时, 可以谨慎选用r00零模型结果进行解释, 同时辅以物种累计曲线来阐述被动取样在网络构建中的影响。当物种累计曲线达到饱和时, 则认为取样量能够涵盖该研究区内所有的物种, 不存在由于数量少而未被监测到的物种, 即被动取样效应对共生关系网络结构无影响。通常, NT和NODF值均能够直观地反映相互作用关系网络的有序水平, 且NODF对于关系矩阵大小、性状变化不敏感, 能够较好地指示不同网络的有序水平差异。然而, 网络间相互作用关系对网络结构的贡献很难判断比较, 相关指示指标也相对较少。在与零模型比较中容易确定网络是否具有显著嵌套结构, 判定过程中嵌套性偏移和z-score指数均能定量化反映当前网络结构与随机情况的差异。本研究中5组网络结果均显示, 基于行限定零模型的NT偏移值随网络非对称性变化保持稳定(图3), 而NT标准化的z-score指数由于随机矩阵方差差异随非对称性增加呈现增大的趋势(附录I; 图4)。同时, 基于行限定零模型NODF标准化z-score指数亦显示随矩阵非对称性变化维持不变(图5)。因此, 非对称性差异较大的网络间进行嵌套性水平比较时, 鉴于非对称性网络中行属性因素较强的限定作用, 选择r0零模型要优于r00和c0零模型。
4 结论
高度非对称关系网络嵌套性判定, 选择零模型强调保守性同时要注意零模型构建的自由空间, r00和c0方法均是不错的选择。选用r00和限定零模型相结合的方法, 有助于更加准确地判断非对称关系网络是否嵌套, 揭示类群水平物种间相互作用的本质。行或列限定零模型对高度非对称网络嵌套性判定的影响存在差异, 矩阵嵌套性判定中对行属性特征更敏感, 不同非对称性网络间嵌套性水平相比较时选用r0零模型要优于r00和c0零模型。附录? 行数相近矩阵形状变化对填充度的影响
Supplement ? Effects of matrix shape variations on connectance附录II 不同零模型构建方法及其特征
Supplement II Null model construction methods and characteristics附录III 5个互惠共生关系网络及基本特征
Supplement III Five mutually-symbiotic relationship networks and their basic properties
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1111/oik.2008.117.issue-8URL [本文引用: 6]
DOI:10.1007/BF00317508URLPMID:11513889 [本文引用: 3]

The measure of order and disorder in the distribution of species in fragmented habitats proposed by Atmar and Patterson (Oecologia, 96 (1993) 373-82) was applied to investigate nested patterns of Dactylogyrus species parasitising the gills of roach. Organisation in dactylogyrid assemblages was investigated at three levels: (1) host populations between localities; (2) local host populations over seasons; and (3) individual hosts over one season within a local host population. Dactylogyrid assemblages showed nested patterns when analyses were conducted at the level of localities (among host populations) and at the level of seasons (among host populations within localities). The analysis at the level of hosts (infracommunities of parasites) revealed that nested pattern is not common. We suggest that nestedness may have a variety of causes and does not necessarily imply competition.
[本文引用: 1]
DOI:10.1016/j.funeco.2013.10.003URL [本文引用: 2]
DOI:10.1111/j.1574-6941.2010.01000.xURLPMID:21114502 [本文引用: 2]
Ectomycorrhizal fungi (EcMF) form diverse communities and link different host plants into mycorrhizal networks, yet little is known about the magnitude of mycobiont diversity of a single tree individual. This study addresses species richness and spatial structure of EcMF in the root system of a single European aspen (Populus tremula) individual in an old-growth boreal mixed forest ecosystem in Estonia. Combining morphological and molecular identification methods for both plant and fungi, 122 species of EcMF were recovered from 103 root samples of the single tree. Richness estimators predicted the total EcMF richness to range from 182 to 207 species, reflecting the observation of 62.3% singletons and doubletons within the community. Fine-scale genetic diversity in Cenococcum geophilum indicates the presence of 23 internal transcribed spacer genotypes. EcMF community was significantly spatially autocorrelated only at the lineage level up to 3 m distance, but not at the species level. Proximity of other hosts had a significant effect on the spatial distribution of EcMF lineages. This study demonstrates that a single tree may host as many EcMF species and individuals as recovered on multiple hosts in diverse communities over larger areas.
DOI:10.1126/science.1194255URLPMID:20705836 [本文引用: 1]
DOI:10.1073/pnas.1633576100URLPMID:12881488 [本文引用: 5]
Most studies of plant-animal mutualisms involve a small number of species. There is almost no information on the structural organization of species-rich mutualistic networks despite its potential importance for the maintenance of diversity. Here we analyze 52 mutualistic networks and show that they are highly nested; that is, the more specialist species interact only with proper subsets of those species interacting with the more generalists. This assembly pattern generates highly asymmetrical interactions and organizes the community cohesively around a central core of interactions. Thus, mutualistic networks are neither randomly assembled nor organized in compartments arising from tight, parallel specialization. Furthermore, nestedness increases with the complexity (number of interactions) of the network: for a given number of species, communities with more interactions are significantly more nested. Our results indicate a nonrandom pattern of community organization that may be relevant for our understanding of the organization and persistence of biodiversity.
DOI:10.1111/jec.2015.103.issue-3URL [本文引用: 1]
DOI:10.1111/j.1365-2656.2009.01567.xURLPMID:19515098
1. In the Neotropics, most plants depend on animals for pollination. Solitary bees are the most important vectors, and among them members of the tribe Centridini depend on oil from flowers (mainly Malpighiaceae) to feed their larvae. This specialized relationship within 'the smallest of all worlds' (a whole pollination network) could result in a 'tiny world' different from the whole system. This 'tiny world' would have higher nestedness, shorter path lengths, lower modularity and higher resilience if compared with the whole pollination network. 2. In the present study, we contrasted a network of oil-flowers and their visitors from a Brazilian steppe ('caatinga') to whole pollination networks from all over the world. 3. A network approach was used to measure network structure and, finally, to test fragility. The oil-flower network studied was more nested (NODF = 0.84, N = 0.96) than all of the whole pollination networks studied. Average path lengths in the two-mode network were shorter (one node, both for bee and plant one-mode network projections) and modularity was lower (M = 0.22 and four modules) than in all of the whole pollination networks. Extinctions had no or small effects on the network structure, with an average change in nestedness smaller than 2% in most of the cases studied; and only two species caused coextinctions. The higher the degree of the removed species, the stronger the effect and the higher the probability of a decrease in nestedness. 4. We conclude that the oil-flower subweb is more cohesive and resilient than whole pollination networks. Therefore, the Malpighiaceae have a robust pollination service in the Neotropics. Our findings reinforce the hypothesis that each ecological service is in fact a mosaic of different subservices with a hierarchical structure ('webs within webs').
DOI:10.1016/j.cub.2006.12.039URLPMID:17275300 [本文引用: 2]
The topology of ecological interaction webs holds important information for theories of coevolution, biodiversity, and ecosystem stability . However, most previous network analyses solely counted the number of links and ignored variation in link strength. Because of this crude resolution, results vary with scale and sampling intensity, thus hampering a comparison of network patterns at different levels . We applied a recently developed quantitative and scale-independent analysis based on information theory to 51 mutualistic plant-animal networks, with interaction frequency as measure of link strength. Most networks were highly structured, deviating significantly from random associations. The degree of specialization was independent of network size. Pollination webs were significantly more specialized than seed-dispersal webs, and obligate symbiotic ant-plant mutualisms were more specialized than nectar-mediated facultative ones. Across networks, the average specialization of animal and plants was correlated, but is constrained by the ratio of plant to animal species involved. In pollination webs, rarely visited plants were on average more specialized than frequently attended ones, whereas specialization of pollinators was positively correlated with their interaction frequency. We conclude that quantitative specialization in ecological communities mirrors evolutionary trade-offs and constraints of web architecture. This approach can be easily expanded to other types of biological interactions.
DOI:10.1007/s00442-015-3457-5URLPMID:27106850 [本文引用: 4]
Understanding the factors that shape community assembly remains one of the most enduring and important questions in modern ecology. Network theory can reveal rules of community assembly within and across study systems and suggest novel hypotheses regarding the formation and stability of communities. However, such studies generally face the challenge of disentangling the relative influence of factors such as interaction type and environmental conditions on shaping communities and associated networks. Endophytic and endolichenic symbioses, characterized by microbial species that occur within healthy plants and lichen thalli, represent some of the most ubiquitous interactions in nature. Fungi that engage in these symbioses are hyperdiverse, often horizontally transmitted, and functionally beneficial in many cases, and they represent the diversification of multiple phylogenetic groups. We evaluated six measures of ecological network structure for &gt;4100 isolates of endophytic and endolichenic fungi collected systematically from five sites across North America. Our comparison of these co-occurring interactions in biomes ranging from tundra to subtropical forest showed that the type of interactions (i.e., endophytic vs. endolichenic) had a much more pronounced influence on network structure than did environmental conditions. In particular, endophytic networks were less nested, less connected, and more modular than endolichenic networks in all sites. The consistency of the network structure within each interaction type, independent of site, is encouraging for current efforts devoted to gathering metadata on ecological network structure at a global scale. We discuss several mechanisms potentially responsible for such patterns and draw attention to knowledge gaps in our understanding of networks for diverse interaction types.
URLPMID:22489483 [本文引用: 1]
A pot experiment was conducted to study the effects of combined application of organic and inorganic fertilizers on the nitrogen uptake by rice and the nitrogen supply by soil in a wheat-rice rotation system, and approach the mechanisms for the increased fertilizer nitrogen use efficiency of rice under the combined fertilization from the viewpoint of microbiology. Comparing with applying inorganic fertilizers, combined application of organic and inorganic fertilizers decreased the soil microbial biomass carbon and nitrogen and soil mineral nitrogen contents before tillering stage, but increased them significantly from heading to filling stage. Under the combined fertilization, the dynamics of soil nitrogen supply matched best the dynamics of rice nitrogen uptake and utilization, which promoted the nitrogen accumulation in rice plant and the increase of rice yield and biomass, and increased the fertilizer nitrogen use efficiency of rice significantly. Combined application of inorganic and organic fertilizers also promoted the propagation of soil microbes, and consequently, more mineral nitrogen in soil was immobilized by the microbes at rice early growth stage, and the immobilized nitrogen was gradually released at the mid and late growth stages of rice, being able to better satisfy the nitrogen demand of rice in its various growth and development stages.
URLPMID:22489483 [本文引用: 1]
A pot experiment was conducted to study the effects of combined application of organic and inorganic fertilizers on the nitrogen uptake by rice and the nitrogen supply by soil in a wheat-rice rotation system, and approach the mechanisms for the increased fertilizer nitrogen use efficiency of rice under the combined fertilization from the viewpoint of microbiology. Comparing with applying inorganic fertilizers, combined application of organic and inorganic fertilizers decreased the soil microbial biomass carbon and nitrogen and soil mineral nitrogen contents before tillering stage, but increased them significantly from heading to filling stage. Under the combined fertilization, the dynamics of soil nitrogen supply matched best the dynamics of rice nitrogen uptake and utilization, which promoted the nitrogen accumulation in rice plant and the increase of rice yield and biomass, and increased the fertilizer nitrogen use efficiency of rice significantly. Combined application of inorganic and organic fertilizers also promoted the propagation of soil microbes, and consequently, more mineral nitrogen in soil was immobilized by the microbes at rice early growth stage, and the immobilized nitrogen was gradually released at the mid and late growth stages of rice, being able to better satisfy the nitrogen demand of rice in its various growth and development stages.
DOI:10.1007/s004420050412URLPMID:28308039 [本文引用: 1]
Randomization models, often termed &quot;null&quot; models, have been widely used since the 1970s in studies of species community and biogeographic patterns. More recently they have been used to test for nested species subset patterns (or nestedness) among assemblages of species occupying spatially subdivided habitats, such as island archipelagoes and terrestrial habitat patches. Nestedness occurs when the species occupying small or species-poor sites have a strong tendency to form proper subsets of richer species assemblages. In this paper, we examine the ability of several published simulation models to detect, in an unbiased way, nested subset patterns from a simple matrix of site-by-species presence-absence data. Each approach attempts to build in biological realism by following the assumption that the ecological processes that generated the patterns observed in nature would, if they could be repeated many times over using the same species and landscape configuration, produce islands with the same number of species and species present on the same number of islands as observed. In mathematical terms, the mean marginal totals (column and row sums) of many simulated matrices would match those of the observed matrix. Results of model simulations suggest that the true probability of a species occupying any given site cannot be estimated unambiguously. Nearly all of the models tested were shown to bias simulation matrices toward low levels of nestedness, increasing the probability of a Type I statistical error. Further, desired marginal totals could be obtained only through ad-hoc manipulation of the calculated probabilities. Paradoxically, when such results are achieved, the model is shown to have little statistical power to detect nestedness. This is because nestedness is determined largely by the marginal totals of the matrix themselves, as suggested earlier by Wright and Reeves. We conclude that at the present time, the best null model for nested subset patterns may be one based on equal probabilities of occurrence for all species. Examples of such models are readily available in the literature.
URL [本文引用: 1]
DOI:10.3724/SP.J.1003.2012.08026URL [本文引用: 1]
DOI:10.3724/SP.J.1003.2012.08026URL [本文引用: 1]
DOI:10.1111/nph.13068URLPMID:25303438 [本文引用: 1]
Environmental selection and dispersal limitation are two of the primary processes structuring biotic communities in ecosystems, but little is known about these processes in shaping soil microbial communities during secondary forest succession. We examined the communities of ectomycorrhizal (EM) fungi in young, intermediate and old forests in a Chinese subtropical ecosystem, using 454 pyrosequencing. The EM fungal community consisted of 393 operational taxonomic units (OTUs), belonging to 21 EM fungal lineages, in which three EM fungal lineages and 11 EM fungal OTUs showed significantly biased occurrence among the young, intermediate and old forests. The EM fungal community was structured by environmental selection and dispersal limitation in old forest, but only by environmental selection in young, intermediate, and whole forests. Furthermore, the EM fungal community was affected by different factors in the different forest successional stages, and the importance of these factors in structuring EM fungal community dramatically decreased along the secondary forest succession series. This study suggests that different assembly mechanisms operate on the EM fungal community at different stages in secondary subtropical forest succession.
DOI:10.1002/ece3.5372URLPMID:31410260 [本文引用: 1]
Patterns of species associations have been commonly used to infer interactions among species. If species positively co-occur, they may form predominantly neutral assemblages, and such patterns suggest a relatively weak role for compensatory dynamics. The main objective of this study was to test this prediction on temporal samples of bird assemblages (n?=?19, 10-57?years) by the presence/absence and quantitative null models on assemblage and guild levels. These null model outcomes were further analyzed to evaluate the effects of various data set characteristics on the outcomes of the null models. The analysis of two binary null models in combination with three association indices revealed 20% with significant aggregations, 61% with random associations, and only 19% with significant segregations (n?=?95 simulations). The results of the quantitative null model simulations detected more none-random associations: 61% aggregations, 6% random associations, and 33% segregations (n?=?114 simulations). Similarly, quantitative analyses on guild levels showed 58% aggregations, 20% segregations, and 22% random associations (n?=?450 simulations). Bayesian GLMs detected that the outcomes of the binary and quantitative null models applied to the assemblage analyses were significantly related to census plot size, whereas the outcomes of the quantitative analyses were also related to the mean population densities of species in the data matrices. In guild-level analyses, only 9% of the GLMs showed a significant influence of matrix properties (plot size, matrix size, species richness, and mean species population densities) on the null model outcomes. The results did not show the prevalence of negative associations that would have supported compensatory dynamics. Instead, we assume that a similar response of the majority of species to climate-driven and stochastic factors may be responsible for the revealed predominance of positive associations.
DOI:10.7717/peerj.686URLPMID:25548729 [本文引用: 2]
A number of recent studies suggest that interspecific competition plays a key role in determining the structure of ectomycorrhizal (ECM) fungal communities. Despite this growing consensus, there has been limited study of ECM fungal community dynamics in abiotically stressful environments, which are often dominated by positive rather than antagonistic interactions. In this study, we examined the ECM fungal communities associated with the host genus Alnus, which live in soils high in both nitrate and acidity. The nature of ECM fungal species interactions (i.e., antagonistic, neutral, or positive) was assessed using taxon co-occurrence and DNA sequence abundance correlational analyses. ECM fungal communities were sampled from root tips or mesh in-growth bags in three monodominant A. rubra plots at a site in Oregon, USA and identified using Illumina-based amplification of the ITS1 gene region. We found a total of 175 ECM fungal taxa; 16 of which were closely related to known Alnus-associated ECM fungi. Contrary to previous studies of ECM fungal communities, taxon co-occurrence analyses on both the total and Alnus-associated ECM datasets indicated that the ECM fungal communities in this system were not structured by interspecific competition. Instead, the co-occurrence patterns were consistent with either random assembly or significant positive interactions. Pair-wise correlational analyses were also more consistent with neutral or positive interactions. Taken together, our results suggest that interspecific competition does not appear to determine the structure of all ECM fungal communities and that abiotic conditions may be important in determining the specific type of interaction occurring among ECM fungi.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1128/AEM.06826-11URLPMID:22194300 [本文引用: 1]
Taxonomic and phylogenetic fingerprinting based on sequence analysis of gene fragments from the large-subunit rRNA (LSU) gene or the internal transcribed spacer (ITS) region is becoming an integral part of fungal classification. The lack of an accurate and robust classification tool trained by a validated sequence database for taxonomic placement of fungal LSU genes is a severe limitation in taxonomic analysis of fungal isolates or large data sets obtained from environmental surveys. Using a hand-curated set of 8,506 fungal LSU gene fragments, we determined the performance characteristics of a na?ve Bayesian classifier across multiple taxonomic levels and compared the classifier performance to that of a sequence similarity-based (BLASTN) approach. The na?ve Bayesian classifier was computationally more rapid (&gt;460-fold with our system) than the BLASTN approach, and it provided equal or superior classification accuracy. Classifier accuracies were compared using sequence fragments of 100 bp and 400 bp and two different PCR primer anchor points to mimic sequence read lengths commonly obtained using current high-throughput sequencing technologies. Accuracy was higher with 400-bp sequence reads than with 100-bp reads. It was also significantly affected by sequence location across the 1,400-bp test region. The highest accuracy was obtained across either the D1 or D2 variable region. The na?ve Bayesian classifier provides an effective and rapid means to classify fungal LSU sequences from large environmental surveys. The training set and tool are publicly available through the Ribosomal Database Project.
[本文引用: 2]
[本文引用: 2]
DOI:10.1111/j.1469-8137.2011.04045.xURLPMID:22269207 [本文引用: 1]
Ecological network theory predicts that in mutualistic systems specialists tend to interact with a subset of species with which generalists interact (i.e. nestedness). Approaching plant-arbuscular mycorrhizal fungi (AMF) association using network analyses will allow the generality of this pattern to be expanded to the ubiquitous plant-AMF mutualism. Based on certain plant-AMF specificity recently suggested, networks are expected to be nested as a result of their mutualistic nature, and modular, with certain species interacting more tightly than others. Network analyses were used to test for nestedness and modularity and to compare the different contribution of plant and AMF to the overall nestedness. Plant-AMF networks share general network properties with other mutualisms. Plant species with few AMFs in their roots tend to associate with those AMFs recorded in most plant species. AMFs present in a few plant species occur in plant species sheltering most AMF (i.e. nestedness). This plant-AMF network presents weakly interlinked subsets of species, strongly connected internally (i.e. modularity). Both plants and AMF show a nested structure, although AMFs have lower nestedness than plants. The plant-AMF interaction pattern is interpreted in the context of how plant-AMF associations can be underlying mechanisms shaping plant community assemblages.
DOI:10.3389/fmicb.2019.02371URLPMID:31708882 [本文引用: 1]
Despite their potential importance with regard to tobacco-related health outcomes, as well as their hypothesized role in the production of tobacco-specific N-nitrosamines, bacterial constituents of tobacco products lack characterization. Specifically, to our knowledge, there has been no comprehensive characterization of the effects of storage conditions on the bacterial communities associated with little cigars and cigarillos. To address this knowledge gap, we characterized the bacterial community composition of the tobacco and wrapper components of the following four products: Swisher Sweets Original; Swisher Sweets, Sweet Cherry; Cheyenne Cigars Full Flavor 100's; and Cheyenne Menthol Box. Each product was stored under three different conditions of temperature and relative humidity to mimic different user storage conditions: room (20°C 50% RH), refrigerator (5°C 18% RH) and pocket (25°C 30% RH). On days 0, 5, 9 and 14, subsamples were collected, the wrapper and tobacco were separated, and their total DNA was extracted separately and purified. Resulting DNA was then used in PCR assays targeting the V3 V4 region of the bacterial 16S rRNA gene, followed by sequencing using Illumina HiSeq 300bp PE. Resulting sequences were processed using the Quantitative Insights Into Microbial Ecology (QIIME) software package, followed by analyses in R using the Phyloseq and Vegan packages. A single bacterial phylum, Firmicutes, dominated in the wrapper subsamples whereas the tobacco subsamples were dominated by Proteobacteria. Cheyenne Menthol Box (CMB) samples were characterized by significant differential abundances for 23 bacterial operational taxonomic units (OTUs) in tobacco subsamples and 27 OTUs in the wrapper subsamples between day 0 and day 14 under all conditions. OTUs from the genera Acinetobacter and Bacillus significantly increased in the CMB tobacco subsamples, and OTUs from Bacillus, Streptococcus, Lactobacillus, and Enterococcus significantly increased in the CMB wrapper subsamples over time. These initial results suggest that the bacterial communities of little cigars and cigarillos are dynamic over time and varying storage conditions.
DOI:10.1046/j.1472-4642.2002.00148.xURL
DOI:10.1002/ece3.508URLPMID:23610649 [本文引用: 1]
High-throughput sequencing is becoming increasingly important in microbial ecology, yet it is surprisingly under-used to generate or test biogeographic hypotheses. In this contribution, we highlight how adding these methods to the ecologist toolbox will allow the detection of new patterns, and will help our understanding of the structure and dynamics of diversity. Starting with a review of ecological questions that can be addressed, we move on to the technical and analytical issues that will benefit from an increased collaboration between different disciplines.
DOI:10.1007/s00572-005-0368-zURLPMID:15997390 [本文引用: 1]
The concept of a common mycorrhizal network implies that the arrangement of plants and mycorrhizal fungi in a community shares properties with other networks. A network is a system of nodes connected by links. Here we apply network theory to mycorrhizas to determine whether the architecture of a potential common mycorrhizal network is random or scale-free. We analyzed mycorrhizal data from an oak woodland from two perspectives: the phytocentric view using trees as nodes and fungi as links and the mycocentric view using fungi as nodes and trees as links. From the phytocentric perspective, the distribution of potential mycorrhizal links, as measured by the number of ectomycorrhizal morphotypes on trees of Quercus garryana, was random with a short tail, implying that all the individuals of this species are more or less equal in linking to fungi in a potential network. From the mycocentric perspective, however, the distribution of plant links to fungi was scale-free, suggesting that certain fungus species may act as hubs with frequent connections to the network. Parallels exist between social networks and mycorrhizas that suggest future lines of study on mycorrhizal networks.
DOI:10.1126/science.1188321URLPMID:20705861 [本文引用: 1]
Research on the relationship between the architecture of ecological networks and community stability has mainly focused on one type of interaction at a time, making difficult any comparison between different network types. We used a theoretical approach to show that the network architecture favoring stability fundamentally differs between trophic and mutualistic networks. A highly connected and nested architecture promotes community stability in mutualistic networks, whereas the stability of trophic networks is enhanced in compartmented and weakly connected architectures. These theoretical predictions are supported by a meta-analysis on the architecture of a large series of real pollination (mutualistic) and herbivory (trophic) networks. We conclude that strong variations in the stability of architectural patterns constrain ecological networks toward different architectures, depending on the type of interaction.
DOI:10.1038/ncomms6273URLPMID:25327887 [本文引用: 1]
Species in ecological communities build complex webs of interaction. Although revealing the architecture of these networks is fundamental to understanding ecological and evolutionary dynamics in nature, it has been difficult to characterize the structure of most species-rich ecological systems. By overcoming this limitation through next-generation sequencing technology, we herein uncover the network architecture of below-ground plant-fungus symbioses, which are ubiquitous to terrestrial ecosystems. The examined symbiotic network of a temperate forest in Japan includes 33 plant species and 387 functionally and phylogenetically diverse fungal taxa, and the overall network architecture differs fundamentally from that of other ecological networks. In contrast to results for other ecological networks and theoretical predictions for symbiotic networks, the plant-fungus network shows moderate or relatively low levels of interaction specialization and modularity and an unusual pattern of 'nested' network architecture. These results suggest that species-rich ecological networks are more architecturally diverse than previously recognized.
DOI:10.1111/oik.2009.118.issue-1URL [本文引用: 7]
DOI:10.1002/ecy.2945URLPMID:31834622 [本文引用: 4]
Identifying species interactions and detecting when ecological communities are structured by them is an important problem in ecology and biogeography.?Ecologists have developed specialized statistical hypothesis tests to detect patterns indicative of community-wide processes in their field data. In this respect, null model approaches have proved particularly popular. The freedom allowed in choosing the null model and statistic to construct a hypothesis test leads to a proliferation of possible hypothesis tests from which ecologists can choose to detect these processes. Here, we point out some serious shortcomings of a popular approach to choosing the best hypothesis for the ecological problem at hand that involves benchmarking different hypothesis tests by assessing their performance on artificially constructed datasets. Terminological errors concerning the use of Type-I and Type-II errors that underlie these approaches are discussed. We argue that the key benchmarking methods proposed in the literature are not a sound guide for selecting null hypothesis tests, and further, that there is no simple way to benchmark null hypothesis tests. Surprisingly, the basic problems identified here do not appear to have been addressed previously, and these methods are still being used to develop and test new null models and summary statistics, from quantifying community structure (e.g., nestedness and modularity) to analyzing ecological networks.
DOI:10.1111/1365-2745.13080URLPMID:31007275 [本文引用: 3]
Uncovering the roles of biotic interactions in assembling and maintaining species-rich communities remains a major challenge in ecology. In plant communities, interactions between individuals of different species are expected to generate positive or negative spatial interspecific associations over short distances. Recent studies using individual-based point pattern datasets have concluded that (a) detectable interspecific interactions are generally rare, but (b) are most common in communities with fewer species; and (c) the most abundant species tend to have the highest frequency of interactions. However, it is unclear how the detection of spatial interactions may change with the abundances of each species, or the scale and intensity of interactions. We ask if statistical power is sufficient to explain all three key results.We use a simple two-species model, assuming no habitat associations, and where the abundances, scale and intensity of interactions are controlled to simulate point pattern data. In combination with an approximation to the variance of the spatial summary statistics that we sample, we investigate the power of current spatial point pattern methods to correctly reject the null model of pairwise species independence.We show the power to detect interactions is positively related to both the abundances of the species tested, and the intensity and scale of interactions, but negatively related to imbalance in abundances. Differences in detection power in combination with the abundance distributions found in natural communities are sufficient to explain all the three key empirical results, even if all pairwise interactions are identical. Critically, many hundreds of individuals of both species may be required to detect even intense interactions, implying current abundance thresholds for including species in the analyses are too low. Synthesis. The widespread failure to reject the null model of spatial interspecific independence could be due to low power of the tests rather than any key biological process. Since we do not model habitat associations, our results represent a first step in quantifying sample sizes required to make strong statements about the role of biotic interactions in diverse plant communities. However, power should be factored into analyses and considered when designing empirical studies.
DOI:10.1093/aob/mcp057URLPMID:19304996 [本文引用: 3]
Ecologists and evolutionary biologists are becoming increasingly interested in networks as a framework to study plant-animal mutualisms within their ecological context. Although such focus on networks has brought about important insights into the structure of these interactions, relatively little is still known about the mechanisms behind these patterns.
DOI:10.1007/s004420050348URLPMID:11027779 [本文引用: 1]
Nested species subset patterns consist in a hierarchical structure of species composition in related assemblages, with the species found in depauperate assemblages representing non-random subsets of progressively richer ones. This pattern has been found at the infracommunity level in about a third of the fish ectoparasite assemblages studied to date. Here we present evidence for another non-random structural pattern in assemblages of fish ectoparasites, anti-nestedness, which corresponds to situations in which parasite species are always absent from infracommunities richer than the most depauperate one in which they occur. We show that this pattern is exactly as common as nestedness, and that anti-nested assemblages are characterised by significantly lower prevalence and mean intensities of parasites than nested assemblages. In addition, we found a positive relationship between the prevalence and the mean intensity of parasites across the different assemblages. We propose a link between the nestedness/anti-nestedness continuum and the prevalence-intensity relationship that may involve colonisation-extinction processes. The results presented here suggest that, although nestedness may not be common in parasite communities, other departures from random species assembly are possible, and that some form of structure may be present in many communities. The continuum between nestedness and anti-nestedness also has implications for recent models of species coexistence in communities.
DOI:10.17520/biods.2015315URL [本文引用: 2]
DOI:10.17520/biods.2015315URL [本文引用: 2]
A consistent metric for nestedness analysis in ecological systems: Reconciling concept and measurement
6
2008
... 不同的零模型检验反映的生态学意义存在区别(
... 对51个动植物间互惠网络分析显示随矩阵非对称性增加网络特异性水平升高,
... 矩阵温度(NT)和配对重叠度(NODF)是两种较为常用的计算网络嵌套性的方法(
... 配对重叠度由
... 零模型是进行嵌套性判定的重要依据(
... 矩阵嵌套性度量指标对矩阵属性变化的响应存在差异(
The measure of order and disorder in the distribution of species in fragmented habitat
3
1993
... 不同的零模型检验反映的生态学意义存在区别(
... ), 选取适当的零模型有助于准确揭示类群间相互作用关系的维持机制.目前, 基于被动取样的嵌套性判定的零模型选择已经进行了较多的研究.如完全随机零模型认为同一类群内的各元素以相同的概率与另一类群中任意元素建立共生关系(
... 矩阵温度(NT)和配对重叠度(NODF)是两种较为常用的计算网络嵌套性的方法(
The Nestedness Temperature Calculator: A Visual Basic Program, Including 294 Presence-absence Matrices
1
1995
... 目前, 已有相当多的研究显示宿主植物可与大量不同真菌建立共生关系(
Network perspectives of ectomycorrhizal associations
2
2014
... 群落中物种之间通过相互作用(如捕食、竞争、寄生、互惠等)彼此关联构成网络(
... 目前, 已有相当多的研究显示宿主植物可与大量不同真菌建立共生关系(
A single European aspen ( Populus tremula) tree individual may potentially harbour dozens of Cenococcum geophilum ITS genotypes and hundreds of species of ectomycorrhizal fungi
2
2011
... 目前, 已有相当多的研究显示宿主植物可与大量不同真菌建立共生关系(
... 菌根非对称网络嵌套性判定中, 兼顾保守型的同时也需要警惕第II类错误的发生, 可以采用多个零模型相结合的方法进行验证.同时, 集合种群嵌套性研究指出, 对于可能影响物种分布的因素(如生境大小、隔离程度及其质量等)都是当嵌套性在统计上被证明以后才考虑的问题(
Structure and dynamics of ecological networks
1
2010
... 群落中物种之间通过相互作用(如捕食、竞争、寄生、互惠等)彼此关联构成网络(
The nested assembly of plant-animal mutualistic networks
5
2003
... 群落中物种之间通过相互作用(如捕食、竞争、寄生、互惠等)彼此关联构成网络(
... 不同的零模型检验反映的生态学意义存在区别(
... 矩阵温度(NT)和配对重叠度(NODF)是两种较为常用的计算网络嵌套性的方法(
... 高度限定的backtrack零模型较之r0和c0方法又具有更高的保守性, 适用于相互作用两大类群在构建关系网络时均受被动取样影响的情况, 如传粉网络调查取样过程中数量多且(或)分布广泛的植物或传粉昆虫种更容易被调查观测到(
... ), 即显著嵌套的关系网络在统计分析过程中判定为未达到嵌套性显著水平, 过低估计网络嵌套结构的显著性水平.嵌套性是互惠关系网络的典型特征, 基于不同零模型构建方法网络的嵌套性水平可能会发生改变, 如
Topology of tree-mycorrhizal fungus interaction networks in xeric and mesic Douglas-fir forests
1
2015
... 菌根非对称网络嵌套性判定中, 兼顾保守型的同时也需要警惕第II类错误的发生, 可以采用多个零模型相结合的方法进行验证.同时, 集合种群嵌套性研究指出, 对于可能影响物种分布的因素(如生境大小、隔离程度及其质量等)都是当嵌套性在统计上被证明以后才考虑的问题(
Pollination networks of oil-flowers: A tiny world within the smallest of all worlds
2009
Specialization, constraints, and conflicting interests in mutualistic networks
2
2007
... 不同的零模型检验反映的生态学意义存在区别(
... ).因此, 选取适宜的零模型对于非对称菌根共生关系网络嵌套性判定具有重要价值.当前, 在非对称网络属性特征方面已经进行了一些基础研究, 如
Interaction type influences ecological network structure more than local abiotic conditions: Evidence from endophytic and endolichenic fungi at a continental scale
4
2016
... 群落中物种之间通过相互作用(如捕食、竞争、寄生、互惠等)彼此关联构成网络(
... 不同的零模型检验反映的生态学意义存在区别(
... 高度限定的backtrack零模型较之r0和c0方法又具有更高的保守性, 适用于相互作用两大类群在构建关系网络时均受被动取样影响的情况, 如传粉网络调查取样过程中数量多且(或)分布广泛的植物或传粉昆虫种更容易被调查观测到(
... 增加限定条件的零模型构建方法能够去除取样环节中被动取样因素的影响, 有助于更加保守地判定物种间相互作用的本质(
岛屿群落组成的嵌套格局及其应用
1
2004
... 群落中物种之间通过相互作用(如捕食、竞争、寄生、互惠等)彼此关联构成网络(
岛屿群落组成的嵌套格局及其应用
1
2004
... 群落中物种之间通过相互作用(如捕食、竞争、寄生、互惠等)彼此关联构成网络(
An evaluation of randomization models for nested species subsets analysis
1
1998
... 零模型是进行嵌套性判定的重要依据(
Bipartite: Analysis of bipartite ecological webs
1
2018
... 本研究中所有运算均在统计分析软件R 3.5.2 (https://cran.r-project.org)中实现.随机矩阵由vegan 2.5-4包(
传粉网络的研究进展: 网络的结构和动态
1
2012
... 群落中物种之间通过相互作用(如捕食、竞争、寄生、互惠等)彼此关联构成网络(
传粉网络的研究进展: 网络的结构和动态
1
2012
... 群落中物种之间通过相互作用(如捕食、竞争、寄生、互惠等)彼此关联构成网络(
Community assembly of ectomycorrhizal fungi along a subtropical secondary forest succession
1
2015
... 目前, 已有相当多的研究显示宿主植物可与大量不同真菌建立共生关系(
Null model analysis of species co-occurrence patterns
1
2000
... 零模型是进行嵌套性判定的重要依据(
Pollination biology in the Snowy Mountains of Australia: Comparisons with montane Colorado, USA
1988
Missing checkerboards? An absence of competitive signal in Alnus-??associated ectomycorrhizal fungal communities
2
2014
... 不同的零模型检验反映的生态学意义存在区别(
... 高度限定的backtrack零模型较之r0和c0方法又具有更高的保守性, 适用于相互作用两大类群在构建关系网络时均受被动取样影响的情况, 如传粉网络调查取样过程中数量多且(或)分布广泛的植物或传粉昆虫种更容易被调查观测到(
生态网络分析方法研究综述
1
2011
... 群落中物种之间通过相互作用(如捕食、竞争、寄生、互惠等)彼此关联构成网络(
生态网络分析方法研究综述
1
2011
... 群落中物种之间通过相互作用(如捕食、竞争、寄生、互惠等)彼此关联构成网络(
嵌套性: 研究方法、形成机制及其对生物保护的意义
1
2002
... 菌根非对称网络嵌套性判定中, 兼顾保守型的同时也需要警惕第II类错误的发生, 可以采用多个零模型相结合的方法进行验证.同时, 集合种群嵌套性研究指出, 对于可能影响物种分布的因素(如生境大小、隔离程度及其质量等)都是当嵌套性在统计上被证明以后才考虑的问题(
嵌套性: 研究方法、形成机制及其对生物保护的意义
1
2002
... 菌根非对称网络嵌套性判定中, 兼顾保守型的同时也需要警惕第II类错误的发生, 可以采用多个零模型相结合的方法进行验证.同时, 集合种群嵌套性研究指出, 对于可能影响物种分布的因素(如生境大小、隔离程度及其质量等)都是当嵌套性在统计上被证明以后才考虑的问题(
Accurate, rapid taxonomic classification of fungal large-subunit rRNA genes
1
2012
... 群落中物种之间通过相互作用(如捕食、竞争、寄生、互惠等)彼此关联构成网络(
动植物互惠网络结构及影响因素
2
2013
... 不同的零模型检验反映的生态学意义存在区别(
... 零模型是进行嵌套性判定的重要依据(
动植物互惠网络结构及影响因素
2
2013
... 不同的零模型检验反映的生态学意义存在区别(
... 零模型是进行嵌套性判定的重要依据(
The network structure of plant- arbuscular mycorrhizal fungi
1
2012
... 为方便不同矩阵嵌套性对比,
Observations on the pollination biology of plants on Melville Island, N.W.T, Canada
1967
Vegan: Community ecology package
1
2019
... 本研究中所有运算均在统计分析软件R 3.5.2 (https://cran.r-project.org)中实现.随机矩阵由vegan 2.5-4包(
Invasion of pollination networks on oceanic islands: Importance of invader complexes and endemic super generalists
2002
High-throughput sequencing: A roadmap toward community ecology
1
2013
... 群落中物种之间通过相互作用(如捕食、竞争、寄生、互惠等)彼此关联构成网络(
Application of network theory to potential mycorrhizal networks
1
2005
... 零模型是进行嵌套性判定的重要依据(
Stability of ecological communities and the architecture of mutualistic and trophic networks
1
2010
... 群落中物种之间通过相互作用(如捕食、竞争、寄生、互惠等)彼此关联构成网络(
Assembly of complex plant-fungus networks
1
2014
... 目前, 已有相当多的研究显示宿主植物可与大量不同真菌建立共生关系(
A consumer’s guide to nestedness analysis
7
2009
... 群落中物种之间通过相互作用(如捕食、竞争、寄生、互惠等)彼此关联构成网络(
... ), 其中嵌套性是互惠共生关系网络(传粉、种子传播、菌根共生等)的典型特征(
... 不同的零模型检验反映的生态学意义存在区别(
... 零模型是进行嵌套性判定的重要依据(
... 增加限定条件的零模型构建方法能够去除取样环节中被动取样因素的影响, 有助于更加保守地判定物种间相互作用的本质(
... 行(列)限定对矩阵非对称变化后嵌套性判定影响不同, 本研究中, 行限定的r0和backtrack零模型NT值和NT偏移随网络非对称性增加无显著变化, 而列限定c0零模型NT值和NT偏移变化与r00零模型结果相近(
... 菌根非对称网络嵌套性判定中, 兼顾保守型的同时也需要警惕第II类错误的发生, 可以采用多个零模型相结合的方法进行验证.同时, 集合种群嵌套性研究指出, 对于可能影响物种分布的因素(如生境大小、隔离程度及其质量等)都是当嵌套性在统计上被证明以后才考虑的问题(
Null model analysis of species nestedness patterns
4
2007
... 群落中物种之间通过相互作用(如捕食、竞争、寄生、互惠等)彼此关联构成网络(
... 为方便不同矩阵嵌套性对比,
... 零模型是进行嵌套性判定的重要依据(
... 矩阵嵌套性度量指标对矩阵属性变化的响应存在差异(
Pattern detection in null model analysis
3
2013
... 零模型是进行嵌套性判定的重要依据(
... 增加限定条件的零模型构建方法能够去除取样环节中被动取样因素的影响, 有助于更加保守地判定物种间相互作用的本质(
... 菌根非对称网络嵌套性判定中, 兼顾保守型的同时也需要警惕第II类错误的发生, 可以采用多个零模型相结合的方法进行验证.同时, 集合种群嵌套性研究指出, 对于可能影响物种分布的因素(如生境大小、隔离程度及其质量等)都是当嵌套性在统计上被证明以后才考虑的问题(
Uniting pattern and process in plant-animal mutualistic networks: A review
3
2009
... 不同的零模型检验反映的生态学意义存在区别(
... 零模型是进行嵌套性判定的重要依据(
... ).限定零模型应用较为广泛, 鸟类岛屿生物地理学研究中常利用该种方法将海岛承载能力作为构建零模型的限定因素用于探究距离因子对鸟类栖息地嵌套性的影响(
A comparative analysis of nested subset patterns of species composition
1
1997
... 不同的零模型检验反映的生态学意义存在区别(
舟山群岛蝶类群落嵌套分布格局及其影响因素
2
2016
... 不同的零模型检验反映的生态学意义存在区别(
... 零模型是进行嵌套性判定的重要依据(
舟山群岛蝶类群落嵌套分布格局及其影响因素
2
2016
... 不同的零模型检验反映的生态学意义存在区别(
... 零模型是进行嵌套性判定的重要依据(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}