Effects of long-term vegetation cover changes on the organic carbon fractions in soil aggregates of mollisols
Na LI, Yi-He ZHANG, Xiao-Zeng HAN,*, Meng-Yang YOU, Xiang-Xiang HAONortheast Institute of Geography and Agroecology, Chinese Academy of Sciences, Harbin 150081, China
Supported by the National Key R&D Program of China(2016YFD0300802-01) the Key Research Program of Frontier Sciences, Chinese Academy of Sciences(QYZDB-SSW-SYS022) the Youth Innovation Promotion Association of Chinese Academy of Sciences(2016211)
Abstract Aims Soil aggregate is the main habitat for decomposition and transformation of soil organic carbon (SOC) and is important to regulate SOC sequestration. The mechanisms of the stability of SOC fractions may vary among different aggregate sizes. The aims of this study were to explore the characteristics of SOC “fractionation” in soil aggregates, and to reveal the mechanisms of carbon (C) sequestration in soil aggregates of mollisols after 31-year changes in vegetation cover. Methods A long-term field experiment with different vegetation cover (grassland, farmland and bareland) was established in National Observation Station of Hailun Agro-ecosystem System. Soil aggregate fractionation, the density and humus fractionation within different aggregate sizes were further carried out. Important findings The results showed that after 31 years of land cover change, the surface SOC and total nitrogen (TN) contents in grassland with higher C inputs increased significantly with time, while the SOC and TN contents decreased significantly in bareland, but with no statistical significance in farmland. The 2-0.25 mm (include 2 mm, the same below) aggregates was the excellent fraction for SOC sequestration under all three land cover. The stability of soil aggregate was in the order of: grassland > farmland > bareland. The mass proportion of soil aggregate and its associated content were highest in grassland, while the proportion of microaggregate and its carbon allocation rate were lowest in grassland. However, due to the lower C inputs in farmland and bareland, the distribution of aggregates was in the order of microaggregate > macroaggregate > silt-clay fraction under these two types of land cover, and organic carbon (OC) content was highest in microaggregates. Different vegetation cover changed the C “fractionation” of density and humus fractions in aggregates. Compared with farmland and bareland soils, OC contents in light fractions in >2 mm and 2-0.25 mm aggregates were higher in grassland, and the OC contents in furic acid, humic acid and humin were highest in 2-0.25 mm aggregates in grassland, while the humus OC accumulated in microaggregates in farmland and bareland. Our results indicated that the plant-derived C entered macroaggregates first, and long-term grass cover enhanced free and light C fractions in macroaggregate, which consequently improved the stability of soil aggregates and enhanced the “fractionation” effects of large aggregates on the humus fractions. Our results revealed the characteristics of carbon sequestration in soil aggregates under different vegetation cover. Keywords:vegetation cover;water-stable aggregates;soil organic carbon;density fraction;humus
PDF (1295KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 李娜, 张一鹤, 韩晓增, 尤孟阳, 郝翔翔. 长期不同植被覆盖对黑土团聚体内有机碳组分的影响. 植物生态学报, 2019, 43(7): 624-634. DOI: 10.17521/cjpe.2019.0028 LI Na, ZHANG Yi-He, HAN Xiao-Zeng, YOU Meng-Yang, HAO Xiang-Xiang. Effects of long-term vegetation cover changes on the organic carbon fractions in soil aggregates of mollisols. Chinese Journal of Plant Ecology, 2019, 43(7): 624-634. DOI: 10.17521/cjpe.2019.0028
土壤团聚体是土壤结构的基本单元, 是土壤的重要组成部分, 具有保证和协调土壤水肥气热、影响土壤微生物活性、维持和稳定土壤适宜三相比等作用(李娜等, 2013)。近年来, 土壤团聚体的固碳机制一直是全球碳源汇研究的热点。土壤有机碳(SOC)是土壤团聚结构的重要构成物质, 经微生物的分解转化, 被分解成密度、大小和稳定性不同的组分(Six et al., 2012), 这些组分在胶结物质胶结、矿物吸附和电性吸附等作用下形成不同粒级大小的团聚体(Chenu & Stotzky, 2002), 导致不同粒径大小的团聚体中, 有机碳的数量和稳定程度不尽相同。团聚体内有机碳的生物、物理和化学结合方式是有机碳固定及保护机制研究的重要内容(Six et al., 1998, 2012; Chenu & Stotzky, 2002)。土壤团聚体对有机碳物理保护过程已进行了较多研究(Six et al., 2000, 2004; Lugato et al., 2010), 但对于各粒级团聚体内有机碳的碳固存机制尚无定论。有****认为, 较大粒级团聚体内的有机碳大部分为半腐解态植物残体或松散结合态有机碳和多糖类物质(Beare et al., 1994), 而微团聚体内的有机碳主要来源于微生物代谢产物等较难分解的、腐殖化程度较高的有机物(Chaney & Swift, 1986)。外源物质进入土壤后, 需经一系列的生物化学分解转化过程, 才能转变为土壤有机碳, 土壤团聚体为这一系列过程提供了合适的“场所”, 进而引起有机碳组分在不同粒径团聚体中的“分馏”, 即有机碳被重新分配。分馏所形成的不同活性和密度组分的有机碳只有在合适的“场所”或者与矿质相结合, 形成稳定性有机碳, 才能被长时间保存下来(Bossuyt et al., 2001; 李凯等, 2010)。所以土壤团聚体是土壤固碳的核心。但是, 到目前为止, 关于团聚体的固碳机制还未完全清楚, 团聚体的形成对有机碳的“分馏”特征如何?特别是团聚体内部有机碳密度组分和化学组分分异方面的研究还相对缺乏。
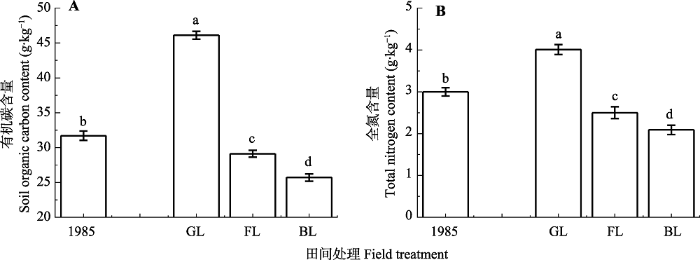
Fig. 1Contents of soil organic carbon and total nitrogen in initial soil and soils under different vegetation covers (mean ± SD). Different lowercase letters above the bar differ at 0.05 levels among different vegetation covers. BL, bareland; FL, farmland; GL, grassland.
Fig. 2Distribution of soil aggregates and mean weight diameter (MWD) of aggregates under different vegetation covers (mean ± SD). Different lowercase letters above the bar differ at 0.05 levels among different vegetation covers.
Fig. 3Contents of organic carbon in soil aggregates fractions under different vegetation covers (mean ± SD). Different lowercase letters above the bar differ at 0.05 levels among different vegetation covers.
Fig. 4Organic carbon contents in different density fractions of aggregates under different vegetation covers (mean ± SD). Different lowercase letters above the bar differ at 0.05 levels among different vegetation covers.
新窗口打开|下载原图ZIP|生成PPT 图5不同植被覆盖下表层全土腐殖质组分的碳含量和腐殖化指标(平均值±标准偏差)。FA, 富里酸; HA, 胡敏酸; HU, 胡敏素。不同小写字母表示处理间差异显著(p < 0.05)。
Fig. 5Contents of organic carbon in humic substances and humification index in bulk soil under different vegetation covers (mean ± SD). FA, fulvic acid; HA, humic acid; HU, humin. Different lowercase letters above the bar differ at 0.05 levels among different vegetation covers.
Fig. 6Contents of organic carbon in soil aggregates under different vegetation covers (mean ± SD). Different lowercase letters above the bar differ at 0.05 levels among different vegetation covers.
BeareMH, HendrixPF, CabreraML, ColemanDC ( 1994). Aggregate-protected and unprotected organic matter pools in conventional- and no-tillage soils Soil Science Society of America Journal, 58, 787-795. DOI:10.2136/sssaj1994.03615995005800030021xURL [本文引用: 1]
BossuytH, DenefK, SixJ, FreySD, MerckxR, PaustianK ( 2001). Influence of microbial populations and residue quality on aggregate stability Applied Soil Ecology, 16, 195-208. DOI:10.1016/S0929-1393(00)00116-5URL [本文引用: 2]
BronickCJ, LalR ( 2005). Soil structure and management: A review Geoderma, 124, 3-22. DOI:10.1016/j.jhazmat.2019.121139URLPMID:31520935 [本文引用: 1] Hydroxyapatite (Ca10(PO4)6(OH)2), a calcium phosphate biomaterial, is a very promising candidate for the treatment of air, water and soil pollution. Indeed, hydroxyapatite (Hap) can be extremely useful in the field of environmental management, due in one part to its particular structure and attractive properties, such as its great adsorption capacities, its acid-base adjustability, its ion-exchange capability and its good thermal stability. Moreover, Hap is able to constitute a valuable resource recovery route. The first part of this review will be dedicated towards presenting Hap's structure and defining properties that result in its viability as an environmental remediation material. The second will focus on its use as adsorbent for wastewater and soil treatment, while indicating the mechanisms involved in this remediation process. Finally, the last part will impart all findings on Hap's applications in the field of catalysis, whether it be as catalyst, as photocatalyst, or as active phase support. Hence, all of the above will have served in showcasing the benefits gained by employing hydroxyapatite in air, water and soil clean-up.
ChaneyK, SwiftRS ( 1986). Studies on aggregate stability. II. The effect of humic substances on the stability of reformed soil aggregates Journal of Soil Science, 37, 337-343. DOI:10.1111/ejs.1986.37.issue-2URL [本文引用: 1]
ChenuC, StotzkyG ( 2002). Interactions between microorganisms and soil particles: An overview In: Interactions between Soil Particles and Microorganisms: Impact on Terrestrial Ecosystem. John Wiley & Sons, Chichester, UK. 3-40. [本文引用: 2]
EynardA, SchumacherTE, LindstromMJ, MaloDD ( 2004). Aggregate sizes and stability in cultivated South Dakota prairie ustolls and usterts Soil Science Society of America Journal, 68, 1360-1365. DOI:10.2136/sssaj2004.1360URL [本文引用: 1]
GolchinA, OadesJM, SkjemstadJO, ClarkeP ( 1994). Soil structure and carbon cycling Australian Journal of Soil Research, 32, 1043-1068. DOI:10.1038/s41598-019-55089-8URLPMID:31811223 [本文引用: 1] Anthropogenic disturbance, such as agricultural and architectural activities, can greatly influence belowground soil microbes, and thus soil formation and nutrient cycling. The objective of this study was to investigate microbial community variation in deep soils affected by strong disturbances. In present study, twelve soil samples were collected from different depths (0-300?cm) and placed onto the surface. We investigated the structure variation of the microbial community down through the soil profiles in response to disturbance originated by legume plants (robinia and clover) cultivation vs. plant-free controls. The high-throughput sequencing of 16S rRNA genes showed that microbial α-diversity decreased with depth, and that growing both plants significantly impacted the diversity in the topsoil. The soil profile was clustered into three layers: I (0-40?cm), II (40-120?cm), and III (120-300?cm); with significantly different taxa found among them. Soil properties explained a large amount of the variation (23.5%) in the microbial community, and distinct factors affected microbial assembly in the different layers, e.g., available potassium in layer I, pH and total nitrogen in layer II, pH and organic matter in layer III. The prediction of metabolic functions and oxygen requirements indicated that the number of aerobic bacteria increased with more air exposure, which may further accelerate the transformation of nitrogen, sulfur, carbon, and pesticides in the soil. The diversity of soil microorganisms followed a depth-decay pattern, but became higher following legume growth and air exposure, with notable abundance variation of several important bacterial species, mainly belonging to Nitrospira, Verrucomicrobia, and Planctomycetes, and soil properties occurring across the soil profiles.
HanXZ, WangSY, VenemanPLM, XingBS ( 2006). Change of organic carbon content and its fractions in black soil under long-term application of chemical fertilizers and recycled organic manure Communications in Soil Science and Plant Analysis, 37, 1127-1137. DOI:10.1080/00103620600588553URL [本文引用: 1]
HaoXX ( 2017). Change Characteristic of Soil Organic Matter in Mollisol Profile Under Different Ecosystems PhD dissertation, University of Chinese Academy of Sciences (Northeast Institute of Geography and Agro-ecology, Chinese Academy of Sciences), Changchun. [本文引用: 1]
HaoXX, DouS, AnFH, LiMM ( 2010). Humus composition and structural characteristics of humic acid in soil aggregates under different utilization of land Journal of Soil and Water Conservation, 24(5), 248-252. [本文引用: 3]
HaoXX, DouS, HanXZ, LiMM, AnFH ( 2014). Structure of humic acid in soil aggregates under different ecosystems in typical black soil region of northeast China Acta Pedologica Sinica, 51, 824-833. [本文引用: 1]
JohnB, YamashitaT, LudwigB, FlessaH ( 2005). Storage of organic carbon in aggregate and density fractions of silty soils under different types of land use Geoderma, 128, 63-79. DOI:10.1016/j.geoderma.2004.12.013URL [本文引用: 1]
LalR ( 2008). Carbon sequestration Philosophical Transactions of the Royal Society B: Biological Sciences, 363, 815-830. DOI:10.1016/j.scitotenv.2019.135806URLPMID:31838420 [本文引用: 1] Seagrass biomass represents an important source of organic carbon that can contribute to long-term sediment carbon stocks in coastal ecosystems. There is little empirical data on the long-term microbial decomposition of seagrass detritus, despite this process being one of the key drivers of carbon-cycling in coastal ecosystems, that is, it influences the amount and quality of carbon available for sequestration. Here, our goal was to investigate how litter quality (leaf vs. rhizome/root) and the microbial communities involved in organic matter remineralisation shift over a 2-year field decomposition study north of Sydney, Australia using the temperate seagrass Zostera muelleri. The sites varied in bulk sediment characteristics and the sediment-associated microbial communities, but these variables overall had little influence on long-term seagrass decomposition rates or seagrass-associated microbiomes. The results showed a clear succession of bacterial and archaeal communities for both tissues types from r-strategists such as α- and γ-proteobacteria to K-strategies, including δ-proteobacteria, Bacteroidia and Spirochaetes. We used a new mathematical model to capture how decay rates varied over time and found that two decomposition events occurred for some seagrass leaf samples, possibly due to exudate input from living seagrass roots growing into the litter bag. The new model also indicated that conventional single exponential models overestimate long-term decay rates, and we detected for the first time the refractory, or stable, phase of decomposition for rhizome/root biomass. The stable phase began at approximately 20% mass remaining and after 600?days, and the persistence of rhizome/root biomass was attributed to the anoxic conditions and the preservation of refractory organic matter. While we predict that rhizome/root biomass will contribute more to the long-term sediment carbon stocks, the preservation of leaf carbon may be enhanced at locations were sedimentation is high and burial in anoxic conditions is rapid and constant.
LiHB, HanXZ, XuYL, HouXY ( 2008). Aggregate stability of rhizosphere soil as affected by land management in black soil Journal of Soil and Water Conservation, 22(3), 110-115. [本文引用: 2]
LugatoE, SimonettiG, MorariF, NardiS, BertiA, GiardiniL ( 2010). Distribution of organic and humic carbon in wet-sieved aggregates of different soils under long-term fertilization experiment Geoderma, 157(3-4), 80-85. DOI:10.1016/j.geoderma.2010.03.017URL [本文引用: 1]
OadesJM ( 1984). Soil organic matter and structural stability: Mechanisms and implications for management Plant and Soil, 76, 319-337. DOI:10.1007/BF02205590URL [本文引用: 1]
PérezMG, Martin-NetoL, SaabSC, NovotnyEH, MiloriDMBP, BagnatoVS, ColnagoLA, MeloWJ, KnickerH ( 2004). Characterization of humic acids from a Brazilian Oxisol under different tillage systems by EPR, 13C NMR, FTIR and fluorescence spectroscopy Geoderma, 118(3-4), 181-190. DOI:10.1007/bf01401306URLPMID:1456104 [本文引用: 1] A highly ossified teratoma was diagnosed and surgically treated in a 2-year old girl. More than 150 teeth were macroscopically identified during the operation. Pathological study established the diagnosis of mature teratoma with teeth formation. Only six analogous cases have been reported previously and only two patients survived the operation.
PugetP, AngersDA, ChenuC ( 1998). Nature of carbohydrates associated with water-stable aggregates of two cultivated soils Soil Biology & Biochemistry, 31, 55-63. DOI:10.1111/gcb.14962URLPMID:31838767 [本文引用: 1] Numerous studies have demonstrated that fertilization with nutrients such as nitrogen, phosphorus, and potassium increase plant productivity in both natural and managed ecosystems, demonstrating that primary productivity is nutrient limited in most terrestrial ecosystems. In contrast, it has been demonstrated that heterotrophic microbial communities in soil are primarily limited by organic carbon or energy. While this concept of contrasting limitations, i.e., microbial carbon and plant nutrient limitation, is based on strong evidence that we review in this paper, it is often ignored in discussions of ecosystem response to global environment changes. The plant-centric perspective has equated plant-nutrient limitations with those of whole ecosystems, thereby ignoring the important role of the heterotrophs responsible for soil decomposition in driving ecosystem carbon storage. In order to truly integrate carbon and nutrient cycles in ecosystem science, we must account for the fact that while plant productivity may be nutrient- limited, the secondary productivity by heterotrophic communities is inherently carbon-limited. Ecosystem carbon cycling integrates the independent physiological responses of its individual components, as well as tightly coupled exchanges between autotrophs and heterotrophs. To the extent that the interacting autotrophic and heterotrophic processes are controlled by organisms that are limited by nutrient versus carbon accessibility, respectively, we propose that ecosystems by definition cannot be 'limited' by nutrients or carbon alone. Here, we outline how models aimed at predicting non-steady state ecosystem responses over time can benefit from dissecting ecosystems into the organismal components and their inherent limitations to better represent plant-microbe interactions in coupled carbon and nutrient models.
SeddaiuG, PorcuG, LeddaL, RoggeroPP, AgnelliA, CortiG ( 2013). Soil organic matter content and composition as influenced by soil management in a semi-arid Mediterranean agro-silvo-pastoral system Agriculture, Ecosystems & Environment, 167, 1-11. DOI:10.1007/s11356-019-06538-4URLPMID:31838703 [本文引用: 1] Valuation of environmental goods and services are frequently spatially heterogeneous, the significance of this heterogeneity for policy analysis is gradually recognized. For the valuation of environmental goods and services in the context of attaining a better environmental status across Heihe River basin (HRB), Northwest of China by 2020, a survey was conducted employing choice experiment approach across the entire river basin. In this research we estimate the impact of distance from inland rivers' origin (spatial attribute) on the inhabitants' willingness to pay for restoration of ecological attributes. A total of 11 ecological attributes were selected including 6 land related attributes and 5 water related attributes. The present study displays the outcomes of the research planned to examine differences in willingness to pay across different locations/distances. A total of five cities and 33 surround villages/townships were included to examine for location effect, while four ad hoc base distances split samples were recognized for distance effect i.e. ≤100?km, ≤200?km, ≤300 and?&gt;?300?km from rivers' origin. The results of mixed logit model recognized that people living at different locations value the attributes differently. For instance the people of Zhangye region are willing to pay RMB 95.66 annually for improvements in biodiversity while in Gaotai the individuals' willingness to pay (WTP) for the same attributes was RMB 45.68. Similarly, the respondents' living nearer to the origin of river were willing to pay relatively higher amount for the upgradation in the degraded ecosystem services than the remainders. The results of willingness to pay obtained by Krinsky Robb method confirmed that the attributes quality of agricultural products and water quality were the most preferred attributes with the willingness to pay RMB 91.09 and 122.89 respectively. The significant results of willingness to pay may serve as a reference for sustainable improvements and uplifting of the degraded ecological attributes.
SixJ, BossuytH, DegryzeS, DenefK ( 2004). A history of research on the link between (micro) aggregates, soil biota, and soil organic matter dynamics Soil & Tillage Research, 79, 7-31. DOI:10.4014/jmb.1911.11003URLPMID:31838828 [本文引用: 1] Deinococcus actinosclerus BM2T (GenBank: KT448814) is a radio-resistant bacterium that is newly isolated from the soil of a rocky hillside in Seoul. As an extremophile, D. actinosclerus BM2T may possess anti-inflammatory properties that may be beneficial to human health. In this study, we evaluated the anti-inflammatory effects of BM2U, an aqueous extract of D. actinosclerus BM2T, on lipopolysaccharide (LPS)-mediated inflammatory responses in RAW264.7 macrophage cells. BM2U showed antioxidant capacity, as determined by the DPPH radical scavenging (IC50 = 349.3 μg/mL) and ORAC (IC50 = 50.24 μg/mL) assays. At 20 μg/mL, BM2U induced a significant increase in heme oxygenase-1 (HO-1) expression (p <0.05). BM2U treatment (0.2 - 20 μg/mL) significantly suppressed LPS-induced increase in the mRNA expression of proinflammatory cytokines tumor necrosis factor-α (TNF-α), interleukin (IL)-1β, and IL-6 (p <0.05). BM2U treatment also suppressed the expression of inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2), which are involved in the production of inflammatory mediators. BM2U treatment also inhibited the activation of nuclear factor-κB (NF-κB) and mitogen-activated protein kinases (MAPKs): JNK, ERK, and p-38 (p <0.05). Collectively, BM2U exhibited anti-inflammatory potential that can be exploited in attenuating inflammatory responses.
SixJ, ElliottET, PaustianK, DoranJW ( 1998). Aggregation and soil organic matter accumulation in cultivated and native grassland soils Soil Science Society of America Journal, 62, 1367-1377. DOI:10.2136/sssaj1998.03615995006200050032xURL [本文引用: 2]
SixJ, ElliottET, PaustianK ( 2000). Soil macroaggregate turnover and microaggregate formation: A mechanism for C sequestration under no-tillage agriculture Soil Biology & Biochemistry, 32, 2099-2103. DOI:10.1111/gcb.14962URLPMID:31838767 [本文引用: 1] Numerous studies have demonstrated that fertilization with nutrients such as nitrogen, phosphorus, and potassium increase plant productivity in both natural and managed ecosystems, demonstrating that primary productivity is nutrient limited in most terrestrial ecosystems. In contrast, it has been demonstrated that heterotrophic microbial communities in soil are primarily limited by organic carbon or energy. While this concept of contrasting limitations, i.e., microbial carbon and plant nutrient limitation, is based on strong evidence that we review in this paper, it is often ignored in discussions of ecosystem response to global environment changes. The plant-centric perspective has equated plant-nutrient limitations with those of whole ecosystems, thereby ignoring the important role of the heterotrophs responsible for soil decomposition in driving ecosystem carbon storage. In order to truly integrate carbon and nutrient cycles in ecosystem science, we must account for the fact that while plant productivity may be nutrient- limited, the secondary productivity by heterotrophic communities is inherently carbon-limited. Ecosystem carbon cycling integrates the independent physiological responses of its individual components, as well as tightly coupled exchanges between autotrophs and heterotrophs. To the extent that the interacting autotrophic and heterotrophic processes are controlled by organisms that are limited by nutrient versus carbon accessibility, respectively, we propose that ecosystems by definition cannot be 'limited' by nutrients or carbon alone. Here, we outline how models aimed at predicting non-steady state ecosystem responses over time can benefit from dissecting ecosystems into the organismal components and their inherent limitations to better represent plant-microbe interactions in coupled carbon and nutrient models.
SixJ, GregorichEG, K?gel-KnabnerI ( 2012). Commentary on the impact of Tisdall & Oades (1982) European Journal of Soil Science, 63, 1-21. DOI:10.1111/j.1365-2389.2011.01408.xURL [本文引用: 2]
SpacciniR, MbagwuJSC, ConteP, PiccoloA ( 2006). Changes of humic substances characteristics from forested to cultivated soils in Ethiopia Geoderma, 132(1-2), 9-19. DOI:10.1016/j.geoderma.2005.04.015URL [本文引用: 1]
SpacciniR, PiccoloA, HaberhauerGF, GerzabekMH ( 2000). Transformation of organic matter from maize residues into labile and humic fractions of three European soils as revealed by 13C distribution and CPMAS-NMR Spectra European Journal of Soil Science, 51, 583-594. DOI:10.1111/ejs.2000.51.issue-4URL [本文引用: 1]
StumpfL, PaulettoEA, PintoLFS ( 2016). Soil aggregation and root growth of perennial grasses in a constructed clay minesoil Soil & Tillage Research, 161, 71-78. DOI:10.4014/jmb.1911.11003URLPMID:31838828 [本文引用: 1] Deinococcus actinosclerus BM2T (GenBank: KT448814) is a radio-resistant bacterium that is newly isolated from the soil of a rocky hillside in Seoul. As an extremophile, D. actinosclerus BM2T may possess anti-inflammatory properties that may be beneficial to human health. In this study, we evaluated the anti-inflammatory effects of BM2U, an aqueous extract of D. actinosclerus BM2T, on lipopolysaccharide (LPS)-mediated inflammatory responses in RAW264.7 macrophage cells. BM2U showed antioxidant capacity, as determined by the DPPH radical scavenging (IC50 = 349.3 μg/mL) and ORAC (IC50 = 50.24 μg/mL) assays. At 20 μg/mL, BM2U induced a significant increase in heme oxygenase-1 (HO-1) expression (p <0.05). BM2U treatment (0.2 - 20 μg/mL) significantly suppressed LPS-induced increase in the mRNA expression of proinflammatory cytokines tumor necrosis factor-α (TNF-α), interleukin (IL)-1β, and IL-6 (p <0.05). BM2U treatment also suppressed the expression of inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2), which are involved in the production of inflammatory mediators. BM2U treatment also inhibited the activation of nuclear factor-κB (NF-κB) and mitogen-activated protein kinases (MAPKs): JNK, ERK, and p-38 (p <0.05). Collectively, BM2U exhibited anti-inflammatory potential that can be exploited in attenuating inflammatory responses.
TisdallJM, OadesJM ( 1982). Organic matter and water-stable aggregates in soils Journal of Soil Science, 33, 141-163. DOI:10.1016/j.scitotenv.2019.135736URLPMID:31791773 [本文引用: 1] Soil compaction can be a major impediment to tree growth as it damages soil physical and biological properties and reduces plant available water. This may result in trees that are more vulnerable to seasonal water stress. Improving soil physical and biological properties by increasing soil organic matter content may lead to improved tree establishment. Organic matter (OM), in the form of municipal green waste compost (MGWC) or biochar was incorporated into compacted urban soils at two sites. We established six soil treatments: 1) unamended, 2) tillage only, 3) tillage with MGWC (20% v/v), 4) tillage with biochar (10% v/v), 5) tillage with MGWC + biochar (10% &amp; 5% v/v - low), and 6) tillage with MGWC + biochar (20% &amp; 10% v/v - high) (one site only). The treatments were established to a depth of 0.5?m in 2?×?2?m plots. One Corymbia maculata sapling was planted into each plot. Bulk density, hydraulic conductivity, stem diameter growth and tree water status were measured during tree establishment. At the end of the 30-month experiment, development of water stable aggregates, the rate of microbiological decomposition of OM, and tree size (diameter at breast height; DBH, and canopy growth index) were measured. All OM amended treatments improved soil physical and biological properties. There were no significant differences among the OM treatments. At the end of the experiment, tree DBH and canopy growth index were greater in the OM treatments than tillage only and unamended. As such, we recommend using local and sustainable forms of OM to improve soils and assist tree establishment in challenging sites where soil water is limited, or evapotranspiration demand is high.
WangT, XuS, ZhaoMY, LiH, KouD, FangJY, HuHF ( 2017). Allocation of mass and stability of soil aggregate in different types of Nei Mongol grasslands Chinese Journal of Plant Ecology, 41, 1168-1176. DOI:10.17521/cjpe.2017.0220URL [本文引用: 1]
YuanYR, LiN, ZouWX, YouMY, HanXZ, MaDL ( 2018). Distribution characteristics of organic carbon in aggregates of soils of three ecosystems in typical Mollisols of Northeast China Acta Ecologica Sinica, 38, 6025-6032. [本文引用: 1]
ZhuGY, ShangguanZP, DengL ( 2017). Soil aggregate stability and aggregate-associated carbon and nitrogen in natural restoration grassland and Chinese red pine plantation on the Loess Plateau Catena, 149, 253-260. DOI:10.1016/j.catena.2016.10.004URL [本文引用: 2]
Aggregate-protected and unprotected organic matter pools in conventional- and no-tillage soils 1 1994
... 土壤团聚体是土壤结构的基本单元, 是土壤的重要组成部分, 具有保证和协调土壤水肥气热、影响土壤微生物活性、维持和稳定土壤适宜三相比等作用(李娜等, 2013).近年来, 土壤团聚体的固碳机制一直是全球碳源汇研究的热点.土壤有机碳(SOC)是土壤团聚结构的重要构成物质, 经微生物的分解转化, 被分解成密度、大小和稳定性不同的组分(Six et al., 2012), 这些组分在胶结物质胶结、矿物吸附和电性吸附等作用下形成不同粒级大小的团聚体(Chenu & Stotzky, 2002), 导致不同粒径大小的团聚体中, 有机碳的数量和稳定程度不尽相同.团聚体内有机碳的生物、物理和化学结合方式是有机碳固定及保护机制研究的重要内容(Six et al., 1998, 2012; Chenu & Stotzky, 2002).土壤团聚体对有机碳物理保护过程已进行了较多研究(Six et al., 2000, 2004; Lugato et al., 2010), 但对于各粒级团聚体内有机碳的碳固存机制尚无定论.有****认为, 较大粒级团聚体内的有机碳大部分为半腐解态植物残体或松散结合态有机碳和多糖类物质(Beare et al., 1994), 而微团聚体内的有机碳主要来源于微生物代谢产物等较难分解的、腐殖化程度较高的有机物(Chaney & Swift, 1986).外源物质进入土壤后, 需经一系列的生物化学分解转化过程, 才能转变为土壤有机碳, 土壤团聚体为这一系列过程提供了合适的“场所”, 进而引起有机碳组分在不同粒径团聚体中的“分馏”, 即有机碳被重新分配.分馏所形成的不同活性和密度组分的有机碳只有在合适的“场所”或者与矿质相结合, 形成稳定性有机碳, 才能被长时间保存下来(Bossuyt et al., 2001; 李凯等, 2010).所以土壤团聚体是土壤固碳的核心.但是, 到目前为止, 关于团聚体的固碳机制还未完全清楚, 团聚体的形成对有机碳的“分馏”特征如何?特别是团聚体内部有机碳密度组分和化学组分分异方面的研究还相对缺乏. ...
Influence of microbial populations and residue quality on aggregate stability 2 2001
... 土壤团聚体是土壤结构的基本单元, 是土壤的重要组成部分, 具有保证和协调土壤水肥气热、影响土壤微生物活性、维持和稳定土壤适宜三相比等作用(李娜等, 2013).近年来, 土壤团聚体的固碳机制一直是全球碳源汇研究的热点.土壤有机碳(SOC)是土壤团聚结构的重要构成物质, 经微生物的分解转化, 被分解成密度、大小和稳定性不同的组分(Six et al., 2012), 这些组分在胶结物质胶结、矿物吸附和电性吸附等作用下形成不同粒级大小的团聚体(Chenu & Stotzky, 2002), 导致不同粒径大小的团聚体中, 有机碳的数量和稳定程度不尽相同.团聚体内有机碳的生物、物理和化学结合方式是有机碳固定及保护机制研究的重要内容(Six et al., 1998, 2012; Chenu & Stotzky, 2002).土壤团聚体对有机碳物理保护过程已进行了较多研究(Six et al., 2000, 2004; Lugato et al., 2010), 但对于各粒级团聚体内有机碳的碳固存机制尚无定论.有****认为, 较大粒级团聚体内的有机碳大部分为半腐解态植物残体或松散结合态有机碳和多糖类物质(Beare et al., 1994), 而微团聚体内的有机碳主要来源于微生物代谢产物等较难分解的、腐殖化程度较高的有机物(Chaney & Swift, 1986).外源物质进入土壤后, 需经一系列的生物化学分解转化过程, 才能转变为土壤有机碳, 土壤团聚体为这一系列过程提供了合适的“场所”, 进而引起有机碳组分在不同粒径团聚体中的“分馏”, 即有机碳被重新分配.分馏所形成的不同活性和密度组分的有机碳只有在合适的“场所”或者与矿质相结合, 形成稳定性有机碳, 才能被长时间保存下来(Bossuyt et al., 2001; 李凯等, 2010).所以土壤团聚体是土壤固碳的核心.但是, 到目前为止, 关于团聚体的固碳机制还未完全清楚, 团聚体的形成对有机碳的“分馏”特征如何?特别是团聚体内部有机碳密度组分和化学组分分异方面的研究还相对缺乏. ...
Change of organic carbon content and its fractions in black soil under long-term application of chemical fertilizers and recycled organic manure 1 2006
Transformation of organic matter from maize residues into labile and humic fractions of three European soils as revealed by 13C distribution and CPMAS-NMR Spectra 1 2000
Soil aggregate stability and aggregate-associated carbon and nitrogen in natural restoration grassland and Chinese red pine plantation on the Loess Plateau 2 2017
 ,*, 尤孟阳, 郝翔翔中国科学院东北地理与农业生态研究所, 哈尔滨 150081
,*, 尤孟阳, 郝翔翔中国科学院东北地理与农业生态研究所, 哈尔滨 150081
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT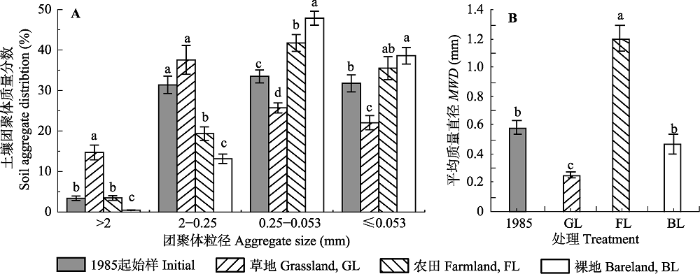 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT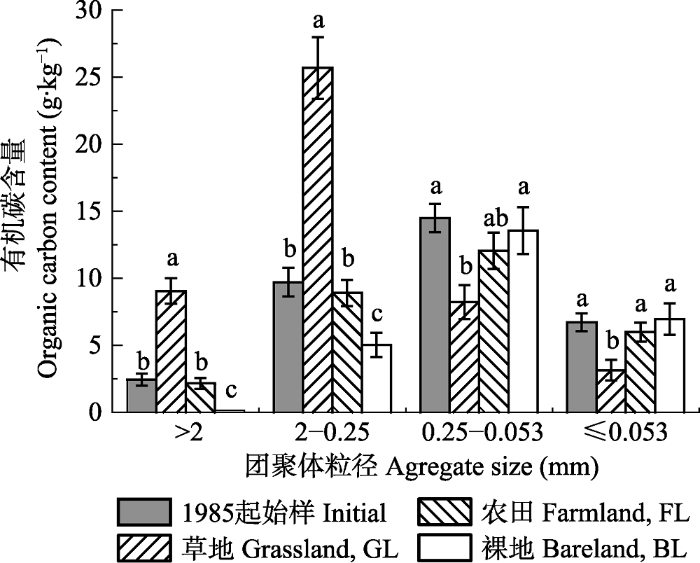 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT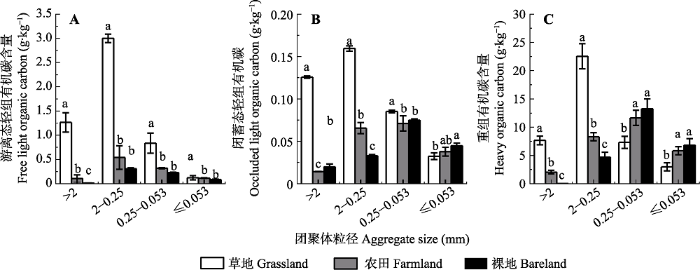 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT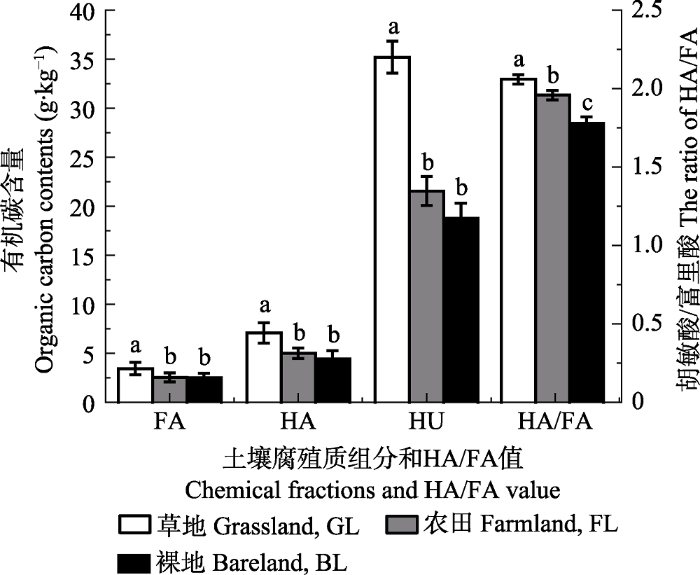 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT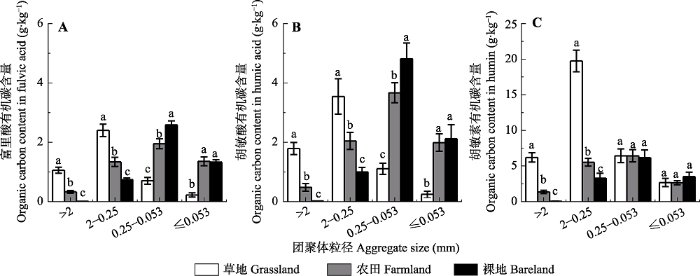 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}