吕中诚
1 , 4 , 康文星
, 1 , 2 , 3 , * , 黄志宏
1 , 2 , 3 , 赵仲辉
1 , 2 , 3 , 邓湘雯
1 , 2 , 3 1.
中南林业科技大学, 长沙 410004 2.
南方林业生态应用技术国家工程实验室, 长沙 410004 3.
湖南会同杉木林国家重点野外科学观测研究站, 湖南会同 418307 4.
湖南警察学院, 长沙 410138 Reuse of retranslocated nutrients in tissues of Chinese fir in plantations of different ages Zhong-Cheng Lü
1 , 4 , Wen-Xing KANG
, 1 , 2 , 3 , * , Zhi-Hong HUANG
1 , 2 , 3 , Zhong-Hui ZHAO
1 , 2 , 3 , Xiang-Wen DENG
1 , 2 , 3 1.
Central South University of Forestry and Technology, Changsha 410004, China 2.
National Engineering Laboratory for Applied Technology of Forestry & Ecology in South China, Changsha 410004, China; 3.
Huitong National Field Station for Scientific Observation and Research of Chinese Fir Plantation Ecosystem, Huitong, Hunan 418307, China 4.
Hunan Police College, Changsha 410138, China 通讯作者: *(
kwx1218@126.com )
编委: 黄建国
责任编辑: 李敏, 实习编辑: 赵航
基金资助: 国家林业公益性行业科研专项 (201104009 )科技部公益性研究项目 (2007-04-15 )国家野外科学观测研究站项目 (20080615 )
Editorial board: HUANG Jian-guo Editor: LI Min, ZHAO Hang Online: 2019-05-20
Fund supported: Supported by the National Forestry Public Welfare Industry Research Project (201104009 )The Ministry of Science and Technology Public Welfare Research Project (2007-04-15 )The National Field Science Observation and Research Station Project (20080615 )
摘要 为了清楚了解不同林龄杉木(
Cunninghamia lanceolata )组织迁移养分(氮、磷、钾、钙、镁)再利用特征, 为人工林丰产的经营管理提供科学依据, 该文利用湖南会同杉木林不同林龄段的活的枝叶与枯死枝叶养分浓度及其差异和枝叶枯死量, 估算枝叶枯死前的养分迁移量。用某林龄段开始和结束时的杉木器官的养分浓度差异, 估算那些在某林龄段开始前林分生产的, 在林龄段结束时仍存活的器官(干、皮、枝、叶、根), 即仍存活物质, 在该林龄段的养分迁移量。将枝叶枯死前迁移的养分和这些仍存活物质中转移的养分与从土壤中吸收的养分相结合, 根据森林生产的生物量, 综合分析森林物质生产的养分利用特征。研究表明: 1-7年生林分, 利用枝叶枯死前迁移的养分生产的生物量及占总生产生物量的比例, 分别为217 kg·hm
-2 ·a
-1 和3.52%; 20-25年生林分则分别上升到2 540 kg·hm
-2 ·a
-1 和17.50%。枝叶枯死前迁移的养分生产的生物量及占总生产生物量的比例都随林龄增加而增大。林分在12-16、17-20、21-25年林龄段, 由这些仍存活物质中迁移出的养分生产的生物量分别为385、561和450 kg·hm
-2 ·a
-1 , 分别占总生产生物量的3.40%、3.40%和3.11%。这些仍存活物质中迁移出的养分量随林龄增加呈现先上升后下降的变化, 由这些养分生产的物质量占总生产物质量的比例随林龄增加呈下降趋势。结果显示, 只要有枝叶枯死发生, 就有枝叶枯死前迁移出养分用于物质再生产。林分郁闭后, 才会发生这些仍存活物质中迁移出的养分再利用。杉木体内养分再分配及贮备机制、杉木生长规律和不同生长发育阶段对养分的需求和利用效率等, 共同调节控制着枝叶枯死前迁移的和这些仍存活物质中迁移出的养分再利用的年变化。
关键词: 杉木 ;
人工林 ;
养分迁移 ;
养分再利用 ;
物质生产 Abstract Aims The purpose of this study is to investigate the reuse characteristics of nutrients (Nitrogen, Phosphorus, Potassium, Calcium, Magnesium) in Cunninghamia lanceolata with different ages, and therefore provide scientific support for better management of the high-yield plantations in China.
Methods We estimated the nutrient retranslocation from dying branches and leaves according to nutrient concentration differences between living and dead, and the amount of dead branches and leaves in Chinese fir forests of different age classes in Hunan Huitong. Based on the difference in the nutrient concentrations of Chinese fir organs at the beginning and end of each age class, the nutrient transport amount from “biomass produced before the beginning of a given age class, which is still alive at the end of the age class” (referred as “biomass-still-alive” hereafter) was estimated for the age class. The nutrient utilization characteristics in forest production were comprehensively analyzed by combining the nutrients transferred from dying branches and leaves with those transferred from the biomass still alive and absorbed from the soil.
Important findings The results showed that the annual biomass produced by the retranslocated nutrients of dying branches and leaves and their proportion in the total biomass were 217 kg·hm
-2 ·a
-1 and 3.52%, respectively, for forests 1-7 years old, and increased to 2 540 kg·hm
-2 ·a
-1 and 17.50%, respectively, when forests reached 20-25 years old. The biomass produced by the retranslocated nutrients of dying branches and leaves and its proportion in the total biomass increased with the increase of forest age. The average annual biomass produced by the biomass-still-alive in stands of 12-16, 17-20 and 21-25 years old was 385, 561 and 450 kg·hm
-2 ·a
-1 , accounting for 3.40%, 3.40% and 3.11% of the total biomass production, respectively. The nutrient retranslocation from the biomass-still-alive first increased and then decreased with the increase of forest age, and the proportion of the biomass produced by these nutrients to the total biomass showed a decreasing trend with the increase of forest age. The results show that the death of branches and leaves always results in reuse of retranslocated nutrients. On the other hand, the reuse of retranslocated nutrients from the biomass-still-alive occurs only after canopy closure. The mechanism of nutrient redistribution and storage, the growth pattern and the nutrient demand and utilization efficiency at different growth stages of Chinese fir jointly regulate and control the annual changes of nutrient retranslocation before branches and leaves die and the reuse of retranslocated nutrients from the biomass-still-alive.
Keywords: Cunninghamia lanceolata ;
plantation ;
retranslocated nutrients ;
reuse of nutrients ;
biomass production PDF (1090KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 吕中诚, 康文星, 黄志宏, 赵仲辉, 邓湘雯. 不同林龄杉木组织迁移养分的再利用. 植物生态学报, 2019, 43(5): 458-470. DOI:
10.17521/cjpe.2018.0212 Lü Zhong-Cheng, KANG Wen-Xing, HUANG Zhi-Hong, ZHAO Zhong-Hui, DENG Xiang-Wen.
Reuse of retranslocated nutrients in tissues of Chinese fir in plantations of different ages .
Chinese Journal of Plant Ecology , 2019, 43(5): 458-470. DOI:
10.17521/cjpe.2018.0212 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节。20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义。以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 )。植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义。然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用。虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道。如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理。本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量。将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据。
1 材料和方法 1.1 研究区域概况 本研究所选林分位于湖南会同杉木林生态系统国家野外科学观测研究站第3集水区试验林内。1987年冬采伐了该集水区第1代杉木林。1988年春在采伐迹地炼山、全垦挖穴营造第2代杉木林, 造林密度2 200株·hm
-2 。前3年(包括造林当年)的春、秋季各抚育1次, 以后任其自然生长。研究林分的地理位置为26.83° N, 109.75° E, 坡向西北, 坡度15°-25°, 海拔270- 350 m。研究区年降水量1 100-1 400 mm, 年平均气温16.9 ℃, 属于中亚热带季风湿润气候区。土壤为板页岩发育的黄壤, 风化程度较深, 土层厚度超过80 cm。
1.2 林分观测样地设置 湖南会同杉木林生态系统国家野外科学观测研究站为了研究杉木林生态系统的结构功能, 1994年在第3集水区山坡中上部和中下部的7年生第2代杉木林内各设2块(共4块) 20 m × 33.33 m长方形样地进行各项观测试验。
1.3 生物量测定 在林龄为7、11、16、20、25年时(与第1代林测定生物量的林龄相同), 在4块观测样地测定林分生物量。生物量测定采用克拉夫特的林木生长状况分级法(
林业部科技司, 1994 ), 把样地内所有林木分成优势木(I级木)、亚优势木(II级木)、中级木(III级木)、被压木(IV级木)、濒死木(V级木) 5个生长等级, 测定样地内每株树木的树高、胸径和冠幅。根据样地所有树木平均树高和胸径选取样地标准木1株, 根据各生长级树木的平均树高和胸径选取各生长级标准木1株(样地中总计6株样木), 将它们连根挖出。在现场将树干按2 m分区段, 测定各区段树干、皮、枝和叶的生物量鲜质量, 并测定根的鲜质量。选取杉木各器官样品, 称量后放在80 ℃的烘箱内烘干至恒质量。求出样品的含水率, 将各器官鲜质量换算成干质量。根据相对生长法则建立生物量估算模型, 再用经过检验的模型估算单位面积林分生物量。
1.4 枝叶枯死量估算 根据杉木枝、叶生长周期(寿命)估算不同林龄时林分枝叶枯死量。杉木叶片生长周期定为5年(
刘爱琴等, 2005 ), 杉木树枝生长周期定为10年(根据在会同长期观测的结果)。用下式估算单位面积林分不同林龄时的枝、叶枯死量。
(1) W 1 t W 1( t - 5) (2) W 2 t W 2( t - 10) 式中,
W 1 t 表示单位面积林分
t 年生时的叶枯死量;
W 2 t 表示单位面积林分
t 年生时的枝枯死量;
W 1( t - 5) 表示单位面积林分(
t - 5)年生时生产的叶量;
W 2( t - 10) 表示单位面积林分(
t - 10)年生时生产的枝量。
1.5 分析样品采集 每次测定生物量的同时采集分析样品。为了保证样品具有代表性, 将每次生物量测定的伐倒木分成树干基部、中下、中、中上和上部5个区段, 根据每一区段树干质量的权重比分区段采集树干样品, 再把各区段釆集的树干样品混合在一起组合成树干样品。树皮取样采用与树干相同的方法。枝和叶都分成当年生、一年生、二年生、多年生分别取样。将根分成根头、大根(直径≥1 cm)、粗根(直径0.2- 1.0 cm)、细根(直径<0.2 cm)分别取样。同时从活树上收集刚枯死枝叶样品。
1.6 养分测定 取适量待测的样品置85 ℃恒温干燥箱内烘干至恒质量, 称质量并磨碎, 过60目筛, 按《森林生态系统研究方法》(
林业部科技司, 1994 )中的养分测定方法配置待测溶液。全钾(TK)、全钙(TCa)、全镁(TMg)含量用原子吸收分光光度计测定, 全磷(TP)含量用分光光度计测定, 全氮(TN)含量用半微量凯氏法测定。每个样品重复测定3次, 用3次重复测定的平均值作为计量标准。
1.7 不同林龄段林分生产的生物量估算 单位面积林分某林龄段生产的树干、树皮生物量, 根据生长过程中树干、树皮凋落甚少的规律, 用某林龄段开始和结束时单位面积林分测定的这些器官现存量之差求得。根系存在更新代谢过程, 但要准确测定根系(尤其是细根)的生长周期, 需采用微根管技术或同位素追踪技术。这些技术方法成本高且操作有一定的难度, 因而大多数研究者在估算某林龄段根生产量时, 将根与干材和树皮同等对待(
梅莉等, 2004 ;
王端丽等, 2012 )。本研究也是如此(这样估算出的根生产量要低于它的实际生产量)。杉木是常绿树种, 枝、叶都有一定的生长周期, 根据
陈日升等(2018) 的研究方法, 估算出某林龄段这些还存活的枝叶量。然后用下式估算某林龄段单位面积林分生产的生物量(
W )。
(3) W = (W 2 k W 1 k W 3 k W 2 i W 1 i 式中,
W 2 k 表示某林龄段结束时的单位面积林分枝叶现存量;
W 1 k 表示某林龄段“这些还存活的枝叶量”;
W 3 k 表示单位面积林分在某林龄段生长的枝叶在该林龄段的枯死量;
W 2 i 、
W 1 i 分别表示某林龄段结束和开始时单位面积林分的树干、树皮和根现存量;
k = 1, 2 (枝、叶);
i = 1, 2, 3 (干、皮、根)。
1.8 不同林龄段杉木活组织的养分转移量计算 根据某林龄段结束时的杉木各器官养分浓度(
L 2 p )和开始时的浓度(
L 1 p ), 以及这林龄段“仍存活物质” (
Wmp ), 估算单位面积“仍存活物质”在这林龄段的养分转移量。若
L 2 p <
L 1 p , 表示这林龄段“仍存活物质”有部分养分被转移出来, 转移量(
M )为:
(4) M = Wmp × (L 2 p L 1 p 若
L 2 p >
L 1 p , 表示着这林龄段“仍存活物质”的养分增加了, 有新的养分补充到“仍存活物质”内, 转移到“仍存活物质”的养分量是:
(5) M = Wmp × (L 1 p L 2 p 式中,
p = 1, 2, 3, 4, 5 (干、皮、枝、叶、根)。
1.9 不同林龄段枝、叶枯死前养分转移量估算 采用某林龄段枝叶枯死前养分浓度(
Lkm )与枯死后养分浓度(
Lkn )之差, 以及在某林龄段单位面积林分的枝叶枯死量(
W 3 k ), 用下式计算某林龄段单位面积林分枝叶枯死前的养分转移量(
M 3 )。
(6) M 3 = W 3 k Lkm - Lkn ) 1.10 不同林龄段土壤养分吸收量估算 单位面积林分不同林龄段土壤养分吸收量根据
陈日升等(2018) 的方法估算。该方法考虑了枝叶枯死前迁移的养分, 以及“仍存活物质”迁移养分的再利用。在估算出单位面积林分某林龄段生产的生物量(公式(3))和某林龄段“仍存活物质”迁移出(或又吸收)养分量(公式(4)和公式(5)), 以及单位面积林分枝叶枯死前养分转移量(公式(6))后, 如果某林龄段结束时杉木各器官养分浓度低于开始时, 则某林龄段单位面积林分“仍存活物质”有部分养分迁移出来, 可用于林分物质生产, 从而减少了土壤养分的吸收量(
M 4 )。其土壤养分吸收量用下面公式求得:
(7) M 4 = Lkm (W 2 k W 1 k W 3 k × Lkn Lp (W 2 i W 1 i M 1 - M 3 式中,
Lp 为某林龄段树干、皮和根的养分浓度;
M 1 为某林龄段单位面积林分“仍存活物质”迁移出的养分量;
p = 1, 2, 3 (干、皮、根)。
当某林龄段结束时杉木各器官养分浓度高于开始时, 表示某林龄段“仍存活物质”的养分增加了, 因此, 该林龄段林分从土壤吸收的养分不仅用于本林龄段林分的物质生产, 还有一些要输送到“仍存活物质”内, 这就增加了土壤养分的吸收量。其计算公式为:
(8) M 4 = Lkm (W 2 k W 1 k W 3 k × Lkn Lp (W 2i -W 1 i M 2 - M 3 式中,
M 2 为某林龄段单位面积林分补充到“仍存活物质”的养分量。
1.11 不同林龄段林分养分利用效率估算 不同林龄段林分的养分利用效率根据下面公式计算:
(9) N = (M 4 + M 3 + M 2 )/W 式中,
N 表示养分的利用效率;
M 4 表示某林龄段单位面积林分用于物质生产的土壤养分量;
M 3 表示某林龄单位面积林分枝叶枯死前迁移养分的再利用量;
M 2 表示某林龄段单位面积林分“仍存活物质”迁移养分的再利用量;
W 表示某林龄段单位面积林分生产的生物量。
1.12 数据分析处理 所有数据经Excel软件处理, 用SPSS 20.0软件进行统计分析。釆用最小显著差异(
LSD )法对林分吸收的土壤养分量, 从活立木组织迁移的养分量, 枝叶凋落前迁移的养分量进行显著性检验, 显著水平为
α = 0.05。
2 结果和分析 2.1 不同林龄段林分的生产力 从
图1 看出, 同一林龄段杉木各器官生产力不同, 其中树干生产力最大。杉木是用材(干材利用)树种, 这种特性决定了树干生产力是构成林分生产力的主要部分。从
图1 中不同林龄段的杉木各器官生产力的对比分析看出, 同一林龄段不同器官生产力不同, 不同林龄段同一器官生产力也不一样, 这是杉木自身的生长规律引起的。如杉木在快速生长阶段(8-11年), 各器官生长速率都较大, 因而各器官生产力之间的差别较其他林龄段更为均衡; 又如干材积累阶段(17-20年), 杉木树干长粗, 干材积累增大, 因此此林龄段树干生产力增大速率大于其他器官。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1不同林龄段杉木林的生产力。同一林龄时, 不同小写字母表示器官间差异显著(p < 0.05)。 Fig. 1Productivity of Chinese fir forests in different age classes. In the same age, the different lowercase letters indicate significant differences among organs (p < 0.05). 林分在1-20年内其生产力持续增加, 20年后生产力下降。各器官生产力随林龄的变化是: 干、叶的变化与林分生产力一样; 枝的生产力在1-11年时上升、12-16年时下降、17-20年时上升、21-25年时下降; 皮和根的生产力在1-11年时上升, 12-16年时下降, 17-25年时又上升。
2.2 不同林龄段杉木活组织内的养分迁移量 因为造林的苗木为一年生苗木, 其苗木生物量在7年生林分生物量中所占比例非常少, 可认为林分7年生时测定的生物量, 即是林分从造林那年到第7年林分生产的生物量。那么7年生的林分就不存在这个林龄阶段的“仍存活物质”。所以
表1 和
表2 中8年后的各林龄段才有“仍存活物质”养分迁移数据。
Table 1 表1 表1 不同林龄杉木体内的养分元素迁移量
Table 1
Average annual nutrient element transfer within the body of Chinese fir in different ages 林龄 养分元素 Nutrient elements (kg·hm-2 ·a-1 ) 合计 -2 ·a-1 ) 氮 Nitrogen 磷 Phosphorus 钾 Potassium 钙 Calcium 镁 Magnesium 8-11 +3.11aM +0.37bM +1.36cM +1.96dM +1.03eM +7.83M 12-16 -1.41aN -0.43bN -1.27cN -1.90dM -0.82eN -5.83N 17-20 -1.66aR -0.53bR -1.40cM -2.26dR -0.90eR -6.75R 21-25 -1.30aS -0.40bN -1.06cS -1.77dS -0.65eS -5.18S
“-”, the nutrient was transported from individuals at the previous stage to the new growth biomass at this stage; “+”, the newly absorbed nutrients in this growth stage flew to the nutrients of the existing organisms in the previous stage. The same row in the same lowercase letters that the difference is not significant (p > 0.05). A certain age in the same column in the same uppercase letter that the difference is not significant (p > 0.05), “-”, 以前生育阶段生长的植物体内养分中输送到这一生育阶段新生长生物量中的养分; “+”, 这一生育阶段新吸收的养分中流向以前生育阶段现存生物体内的养分。同一行相同小写字母表示差异不显著(p > 0.05); 同一列相同大写字母表示差异不显著(p > 0.05)。 新窗口打开 |
下载CSV Table 2 表2 表2 不同林龄杉木器官的养分迁移量
Table 2
Average annual nutrient transfer in organs of Chinese fir in different ages 林龄 干 Stem wood (kg·hm-2 ·a-1 ) 皮 Stem bark-2 ·a-1 ) 枝 Twig-2 ·a-1 ) 叶 Needle-2 ·a-1 ) 根 Root-2 ·a-1 ) 合计 Total-2 ·a-1 ) 8- 11 +1.01aM +1.21bM +2.10cM +2.92dM +0.59eM +7.83M 12- 16 -1.09aN -0.64bN -1.46cN -1.57dN -1.07aN -5.83N 17- 20 -1.79aR -0.87bR -1.63aR -1.18dR -1.28dR -6.75R 21- 25 -1.65aR -0.71bS -1.08cS -0.86dS -0.88dS -5.18S
“-”, the nutrient transferred from individuals at the previous stage to the new growth biomass at this stage; “+”, the newly absorbed nutrients in this growth stage that flow to the existing organisms in the previous stage. Different lowercase letters in the same row indicate significant differences (p < 0.05), different uppercase letters in the same column indicate significant differences (p < 0.05). “-”, 以前生育阶段生长的植物体内养分中输送到这一生育阶段新生长生物量中的养分; “+”, 这一生育阶段新吸收的养分中流向以前生育阶段现存生物体内的养分。同一行不同小写字母表示差异显著(p < 0.05); 同一列不同大写字母表示差异显著(p < 0.05)。 新窗口打开 |
下载CSV 从
表1 看出, 8-11年生的林分, “仍存活物质”新补充的养分为7.83 kg·hm
-2 ·a
-1 , 其中, N、P、K、Ca、Mg的平均补充量分别为3.11、0.37、1.36、1.96和1.03 kg·hm
-2 ·a
-1 。12年以后各林龄段“仍存活物质”平均每年迁移出的养分为5.18-6.75 kg·hm
-2 , 其中, N、P、K、Ca、Mg迁移量分别为1.30-1.66、0.40-0.53、1.06-1.40、1.77-2.26、0.65-0.90 kg·hm
-2 ·?a
-1 。从“仍存活物质”迁移出的各养分量在12-20年内都呈上升趋势, 20-25年内都呈下降趋势。
由
表2 可知, 8-11年生的林分补充到“仍存活物质”(干、皮、枝、叶和根)的平均养分量, 分别为1.01、1.21、2.10、0.92和0.59 kg·hm
-2 ·a
-1 。可见, 在快速生长阶段林分吸收的养分不仅要满足本林龄段林分生产物质的需要, 而且还要运送一些到“仍存活物质”。这是杉木在快速生长阶段为了壮大自己的个体所采取的策略。从运送到“仍存活物质”的各器官养分量看出, 生理功能越强的器官如叶补充的养分越多。
林分自第12年起, 从“仍存活物质” (干、皮、枝、叶和根)中迁移出的养分量, 依次是1.09-1.79、0.64-?0.87、1.08-1.63、0.86-1.57、0.88-1.28 kg·hm
-2 ·?a
-1 。各器官迁移出来的养分随林龄变化的规律是: 除从叶中迁移出的养分随林龄增加而减少外, 干、皮、枝和根中迁移出的养分均随林龄增加呈现先上升后下降的现象。从
表2 还看出, 当杉木进入干材积累阶段后, 生理功能越强的器官, 如叶, 转移出来的养分越少; 生理功能越弱的器官, 如树干、树枝中迁移出来的养分相对较多。可见, 各器官迁移出来的养分量也与器官的功能有关。
2.3 不同林龄段枝、叶枯死前的养分迁移量 表3 表明, 1-25年生的林分, 不同林龄段枝叶枯死前迁移出的养分量为3.23-29.28 kg·hm
-2 ·a
-1 。其中, 枝的迁移量0.19-8.40 kg·hm
-2 ·a
-1 , 占养分总迁移量的5.59%-28.68%; 叶的迁移量3.04-20.88 kg·hm
-2 ·?a
-1 , 占养分总迁移量71.32%-94.41%。由此可见, 杉木枝叶枯死前的养分迁移量绝大部分由叶贡献。枝的养分迁移量随林龄增加而增加; 叶的养分迁移量在20年以前随林龄增加而增加, 20年以后随林龄增加而下降。
Table 3 表3 表3 杉木枝叶枯死前的养分元素迁移量
Table 3
Average annual nutrient element transferred before death of branches and leaves of Chinese fir 林龄 器官 养分元素 Nutrient elements (kg·hm-2 ·a-1 ) 合计 Total-2 ·a-1 ) 磷 Phosphorus 氮 Nitrogen 钾 Potassium 钙 Calcium 镁 Magnesium 1- 7 枝 Twig 0.01bA 0.05aA 0.11cA 0.01bA 0.01bA 0.19A 叶 Needle 0.18bB 1.25aB 1.30cB 0.19bB 0.12cB 3.04B 合 Total 0.19bB 1.30aQ 1.41cQ 0.20dB 0.13eQ 3.23Q 8- 11 枝 Twig 0.04bC 0.24aC 0.42cC 0.03dD 0.03dC 0.77C 叶 Needle 0.62bD 5.24aD 4.62cD 0.86dF 0.38eD 11.72D 合 Total 0.66bE 5.48aE 5.04cE 0.89dE 0.41eE 12.48E 12- 16 枝 Twig 0.15bM 1.04aM 1.49cM 0.16dM 0.09eM 2.93M 叶 Needle 1.03bN 8.47aN 6.76cN 1.44dN 0.43eN 18.13N 合 Total 1.18bT 9.51aT 8.25cT 1.60dT 0.52eT 21.06T 17- 20 枝 Twig 0.39bK 2.87aK 3.47cK 0.45dK 0.18eK 7.36K 叶 Needle 1.52bR 11.98aR 8.51cR 2.11dR 0.41eR 24.53R 合 Total 1.91bS 14.85aS 11.98cS 2.56dS 0.59eS 31.89S 21- 25 枝 Twig 0.41bK 3.57aW 3.71cW 0.57dW 0.14eW 8.40W 叶 Needle 1.34bG 10.30aG 6.94cG 2.02dR 0.28eG 20.88G 合 Total 1.75bP 13.87aP 10.65cP 2.59dP 0.42eP 29.28P
The same row in the same lowercase letters indicates that the difference is not significant (p > 0.05). A certain age in the same column in the same uppercase letter indicates that the difference is not significant (p > 0.05). 同一行小写字母相同表示差异不显著(p > 0.05); 同一列大写字母相同表示差异不显著(p > 0.05)。 新窗口打开 |
下载CSV 从
表3 还可看出, 不同林龄段枝叶枯死前的N、P、K、Ca、Mg迁移量分别是1.30-14.85、0.19-1.91、1.41-11.98、0.20-2.59、0.13-0.59 kg·hm
-2 ·a
-1 , 分别占总迁移量的40.37%-47.38%、5.29%-5.97%、36.37%-43.79%、6.09%-8.85%、1.43%-3.97%。从中看出, 枝叶枯死前迁移的养分主要是N和K, 占总迁移量的83.75%-84.25%, P、Ca、Mg的迁移量只占总迁移量的15.75%-16.25%。不同林龄段枝叶枯死前迁移的N中, 枝占3.85%-25.74%, 叶占74.26%-?96.15%; 迁移的P中, 枝占3.23%-23.43%, 叶占76.57%-96.77%; 迁移的K中, 枝占7.80%-?34.84%, 叶占65.16%-92.20%; 迁移的Ca中, 枝占3.06%-?22.01%, 叶占77.99%-96.94%; 迁移的Mg中, 枝占6.25%-33.33%, 叶占66.67%-93.75%。无论哪一种养分, 叶的迁移量均占其中绝大部分。
2.4 不同林龄段林分吸收的土壤养分量 根据“吸收=存留+归还”的方法(传统方法)计算的林分的土壤养分吸收量, 以及按公式(7)或公式(8) 计算的林分的土壤养分吸收量都列于
表4 中。
1-7年生的林分, 传统方法计算的林分吸收的土壤养分为90.79 kg·hm
-2 ·a
-1 。由于该林龄段不存在“仍存活物质”, 因此也就没有“仍存活物质”的养分迁移量。但该林龄段内枝叶枯死前迁移的3.22 kg·hm
-2 ·a
-1 养分可再利用, 这就减少了林分对土壤养分的吸收量, 因此, 按公式(8)计算得到林分吸收的土壤养分为87.57 kg·hm
-2 ·a
-1 。
8-11年生的林分, 传统方法计算的林分吸收土壤养分为164.08 kg·hm
-2 ·a
-1 。在该林龄段补充到“这些仍存活的物质”的养分为7.83 kg·hm
-2 ·a
-1 , 这增加了林分对土壤养分的吸收量。但此林龄段枝叶枯死前迁移的12.48 kg·hm
-2 ·a
-1 养分的再利用, 又可减少林分对土壤养分的吸收量。按公式(8)计算得到林分吸收土壤养分159.43 kg·hm
-2 ·a
-1 。
12-16年生的林分, 从“仍存活物质”迁移出5.83 kg·hm
-2 ·a
-1 养分和该林龄段枯死枝叶中迁移的21.06 kg·hm
-2 ·a
-1 养分可再利用。按公式(7)计算得到林分平均吸收了146.21 kg·hm
-2 ·a
-1 土壤中的养分, 比传统方法计算的173.10 kg·hm
-2 ·a
-1 少26.89 kg·hm
-2 ·a
-1 。
17-20年和21-25年两个林龄段与12-16年林龄段相同, 都发生了“仍存活物质”迁移出的养分和某林龄段的枝叶枯死前迁移的养分的再利用。因此, 按公式(7)计算得到这两个林龄段吸收的土壤养分依次为160.72和132.73 kg·hm
-2 ·a
-1 , 分别比传统方法计算的199.36和167.19 kg·hm
-2 ·a
-1 少38.64和34.46 kg·hm
-2 ·a
-1 。
从
表4 中可看出, 任一个林龄时, 林分吸收土壤营养元素的量都是N > Ca > K > Mg > P。1-7年平均吸收土壤养分87.57 kg·hm
-2 ·a
-1 , 8-11年上升到159.43 kg·hm
-2 ·a
-1 , 12-16年下降到146.21 kg·hm
-2 ·?a
-1 , 17-20年又上升到160.72 kg·hm
-2 ·?a
-1 , 21-25年再降到132.73 kg·hm
-2 ·a
-1 , 在1-25年生的林分内, 吸收的土壤养分随林龄增加呈现上升、下降、再上升、再下降的变化。
2.5 不同林龄段杉木体内转移的养分再利用 1-7年生的林分, 从土壤吸收养分87.57 kg·?hm
-2 ·a
-1 (
表4 ), 从枝叶枯死前迁移的可再利用养分3.22 kg·hm
-2 ·a
-1 (
表3 ), 用于物质生产的养分共90.79 kg·hm
-2 ·a
-1 。该林龄段每年生产干物质6.17 t·hm
-2 (
表4 ), 即生产1 t干物质消耗养分14.77 kg, 其中土壤提供14.25 kg养分, 枝叶枯死前转移提供0.52 kg (
表5 )。每生产1 t干物质中, 利用土壤养分生产了964.8 kg, 利用枝叶枯死前迁移的养分生产了35.20 kg, 枝叶枯死前迁移的养分生产的物质占生产物质总量的3.52%。
Table 4 表4 表4 杉木林在不同林龄阶段年吸收的土壤养分
Table 4
Annual nutrients taken up from soils by Chinese fir plantations at different ages 林龄 生产力-2 ·a-1 ) 计算公式 养分元素 Nutrient elements (kg·hm-2 ·a-1 ) 合计 Total-2 ·a-1 ) 磷 氮 钾 钙 镁 1- 7 6.17 ARR 2.92 30.43 21.44 28.66 7.54 90.79 公式(8) Formula (8) 2.74 29.18 20.14 28.47 7.42 87.57 8- 11 10.63 ARR 5.60 57.30 36.45 50.26 14.47 164.08 公式(8) Formula (8) 5.31 54.93 32.77 51.33 15.09 159.43 12- 16 11.31 ARR 5.79 61.09 37.79 53.10 15.35 173.10 公式(7) Formula (7) 4.18 50.17 28.27 49.60 13.99 146.21 17- 20 16.51 ARR 6.72 73.30 44.69 58.24 16.41 199.36 公式(7) Formula (7) 4.28 56.79 31.31 53.42 14.92 160.72 21- 25 14.45 ARR 5.61 60.77 38.46 48.66 13.70 167.19 公式(7) Formula (7) 3.45 45.60 26.75 44.30 12.63 132.73
ARR, means “absorption = retention + return”. ARR, 表示“吸收=存留+归还”。 新窗口打开 |
下载CSV 8-11年生的林分, 从土壤中吸收159.43 kg·?hm
-2 ·a
-1 养分(包括输送到“仍存活物质”中7.83 kg·hm
-2 ·a
-1 养分)(
表4 ), 加上每年枝叶枯死前迁移出12.48 kg·hm
-2 ·a
-1 养分再利用(
表3 ), 因而用于生产物质的养分共171.91 kg·hm
-2 ·a
-1 。该林龄段每年生产干物质10.63 t·hm
-2 (
表4 ), 每生产1 t干物质所需的16.17 kg养分中, 从土壤中吸收和枝叶枯死前迁移的养分分别为15.00和1.17 kg (
表5 )。每生产1 t干物质中, 有927.6 kg干物质是利用土壤养分生产的, 有72.4 kg干物质是利用枝叶枯死前迁移的养分生产的, 分别占生产干物质总量的92.76%和7.24%。
Table 5 表5 表5 杉木林生产1 t整树干物质所需养分及来源
Table 5
Nutrients and sources needed for producing 1 t dry biomass in Chinese fir plantations 林龄 养分来源 养分元素 Nutrient elements (kg·hm-2 ·a-1 ) 合计 Total-2 ·a-1 ) 磷 氮 钾 钙 镁 1- 7 土壤中吸收 Absorbed from the soil 0.44 4.73 3.26 4.62 1.20 14.25 衰老枝叶回流 Return from senescent branches and leaves 0.03 0.21 0.23 0.03 0.02 0.52 TPGM 0 0 0 0 0 0 合计 Total 0.47 4.94 3.49 4.65 1.22 14.77 8 - 11 ASNAPGM 0.50 5.17 3.08 4.83 1.42 15.00 衰老枝叶回流 Return from senescent branches and leaves 0.06 0.52 0.47 0.08 0.04 1.17 TPGM 0 0 0 0 0 0 合计 Total 0.56 5.69 3.55 4.91 1.46 16.17 12- 16 土壤中吸收 Absorbed from the soil 0.37 4.44 2.50 4.39 1.23 12.93 衰老枝叶回流 Return from senescent branches and leaves 0.10 0.84 0.73 0.14 0.05 1.86 TPGM 0.05 0.10 0.13 0.14 0.10 0.52 合计 Total 0.52 5.38 3.36 4.67 1.38 15.31 17- 20 土壤中吸收 Absorbed from the soil 0.26 3.44 1.90 3.23 0.90 9.73 衰老枝叶回流 Return from senescent branches and leaves 0.12 0.89 0.72 0.16 0.04 1.93 TPGM 0.05 0.11 0.10 0.07 0.08 0.41 合计 Total 0.43 4.44 2.72 3.46 1.02 12.07 21-2 5 从土壤中吸收 Absorbed from the soil 0.24 3.16 1.85 3.06 0.87 9.18 衰老枝叶回流 Return from senescent branches and leaves 0.12 0.96 0.74 0.18 0.03 2.03 TPGM 0.03 0.09 0.07 0.12 0.05 0.36 合计 Total 0.39 4.21 2.66 3.36 0.95 11.57
ASNAPGM, which is taken from the soil (including nutrients added to substances that survived 8 years ago and survived in this forest age); TPGM, which refers to nutrients transferred from previously grown materials that are still alive in this forest age. ASNAPGM, 表示从土壤中吸收(包括补充到8年以前生长的, 在此林龄段内仍存活的物质中)的养分; TPGM, 表示从以前生长的在该林龄段还活着的物质中迁移出的养分。 新窗口打开 |
下载CSV 12-16年生的林分, 用于物质生产的养分来自3个方面, 一是林分吸收的土壤养分146.21 kg·hm
-2 ·a
-1 (
表4 ), 二是“仍存活物质”迁移出养分5.83 kg·hm
-2 ·a
-1 (
表2 ), 再是枝叶枯死前迁移出养分21.06 kg·hm
-2 ·a
-1 (
表3 )。该林龄段林分每年生产干物质11.31 t·hm
-2 (
表4 )。每生产1 t干物质需要15.31 kg养分, 其中, 土壤吸收的、枝叶枯死前迁移的、“仍存活物质”迁移出的养分分别为12.93、1.86和0.52 kg (
表5 )。该林龄段林分每生产1 t干物质, 从“仍存活物质”迁移出的养分再利用生产了34 kg, 从枝叶凋落前迁移的养分再利用生产了121.5 kg, 分别占生产物质总量的3.40%和12.15%。
17-20年生的林分, 用于物质生产的养分有“仍存活物质”中迁移出的养分6.75 kg·hm
-2 ·a
-1 (
表2 ), 枝叶枯死前迁移的养分31.89 kg·hm
-2 ·a
-1 (
表3 ), 从土壤吸收的养分160.72 kg·hm
-2 ·a
-1 (
表4 ), 共计199.36 kg·hm
-2 ·a
-1 。该林龄段每年生产16.51 t·hm
-2 干物质(
表4 )。从
表5 可知每生产1 t干物质消耗土壤养分9.73 kg, 消耗枝叶枯死前迁移的养分1.93 kg, 消耗“仍存活物质”迁移出的养分0.41 kg。林分每年生产的16.51 t·hm
-2 干物质中, 利用枝叶枯死前迁移的养分生产了2.32 t·hm
-2 ·a
-1 , 利用“仍存活物质”迁移出的养分生产了0.56 t·hm
-2 ·a
-1 , 分别占林分生产物质总量的15.99%和3.40%。
21-25年生的林分, 生产干物质14.45 t·hm
-2 ·a
-1 (
表4 ), 共消耗养分168.69 kg·hm
-2 ·a
-1 。其中, 消耗土壤中的养分132.73 kg·hm
-2 ·a
-1 (
表4 ), 枝叶枯死前迁移的养分26.28 kg·hm
-2 ·a
-1 (
表3 ), 从“仍存活物质”迁移出的养分5.18 kg·hm
-2 ·a
-1 (
表2 )。那么每生产1 t干物质需利用养分11.57 kg, 其中, 利用土壤中的养分9.18 kg, 利用“仍存活物质”迁移出的养分0.36 kg, 利用枝叶凋落前迁移的养分2.03 kg (
表5 )。林分生产的14.45 t·hm
-2 ·a
-1 干物质中, 利用枝叶枯死前迁移的和“仍存活物质”迁移出的养分分别生产了2.54和0.45 t·hm
-2 ·a
-1 , 分别占生产物质总量的17.55%和3.11%。
从上面分析看出, 林分物质生产过程中, 利用的枝叶枯死前迁移的养分生产的物质占生产物质总量的比例, 从1-7年生的林分的3.52%持续上升到21-25年生的林分的17.55%。只有林龄达到12年后, 才发生从“仍存活物质”中迁移出的养分的再利用。利用“仍存活物质”迁移出的养分生产的物质, 占林分生产物质总量的3.11%-3.40%。
3 讨论 会同杉木枝叶枯死前N、P、K迁移率大于Ca、Mg。各养分元素迁移率与福建杉木(
周丽丽, 2014 )相比, 除叶的Mg相近外, 其余枝叶中各元素转移率均低于福建杉木。与林宝平等(2017)研究的杉木相比, P、Ca转移率偏高, N、K、Mg则偏低。与其他研究结果存在差别可能是如下原因造成的: (1)研究林分的立地环境条件不相同, 同一树种在不同的立地环境条件下可能造成枝叶枯死前后养分浓度的差异, 进而导致枝叶枯死前养分迁移率的差异。(2)研究方法不同, 本研究是在同一林分内连续定位测定不同林龄时的数据, 其他同类研究大都用“空间换时间”法, 空间的异质性可能导致其研究结果偏移。(3)本研究把1-25年生的林分分成几个连续的林龄段, 依据每林龄段前后测定数据分段逐一分析, 同类研究则是在林分某一林龄时测定数据, 把林分1年生到这个林龄时作为一个时间序列处理, 所取的时间尺度不一样, 导致研究结果也不一样。
从本研究结果看出, 会同1-25年生杉木林生产的物质中, 有3.52%-17.55%是利用枝叶枯死前迁移的养分生产的, 有3.11%-3.40%是利用“仍存活物质”迁移出的养分生产的。可见, 用于林分物质生产的养分并不都来自土壤。如果忽略林分物质生产中对植物组织衰老时迁移的和“仍存活物质”迁移出的养分的再利用, 就不能真实地反映林分养分利用特征, 而且也可能会低估林分对养分的利用效率。
枝叶枯死前迁移的养分, 以及这些养分生产的物质占生产物质总量的比例都随林龄增加而增大。这是由不同林龄林分的枝叶枯死量决定的。杉木是常绿树种, 枝叶都有一定的生长周期, 当年生长的枝叶不会在当年枯死, 要到它生长周期的后期才会死亡。从
图1 可以看出, 杉木从幼树到干材积累阶段(1-20年生), 叶的生产量逐年增加。这些不同年份生长的叶完成它的生长期后逐渐死亡, 所以随林龄增加叶枯死量逐渐增多, 叶枯死前迁移的养分也逐渐增多, 再用于物质生产的养分也增多。可见, 树木生长发育特征和枝叶的生理特性控制着枝叶枯死前的养分迁移和再利用的年变化规律。
会同杉木林生长12年时, 才发生某林龄段“仍存活物质”迁移出养分用于物质生产。这主要由杉木不同生长阶段对养分的不同需求引起的。杉木从幼树到快速生长阶段为了壮大自己的个体, 为树木未来生长打好基础, 已生产的物质(器官)中需要不断补充养分。因此, 这个生育阶段的“仍存活物质”中不仅没有养分转移出来再利用, 而且还需要新的养分补充。如本研究中8-11年生的林分吸收的土壤养分中, 每年还需输送7.83 kg·hm
-2 到该林龄段“仍存活物质”内(
表2 )。此外,
Miller (1986) 的研究表明, 林分郁闭后, 多年生乔木林生产单位物质所需的养分量减少。树木体内养分再分配机制控制着将积累在“仍存活物质”内的养分迁移出来重新用于物质生产。从本研究结果看, 林分生长12年后(杉木林已郁闭), 虽然从“仍存活物质”内迁移出的养分并不很多, 但“仍存活物质”迁移出的养分生产的物质量仍占生产物质总量的3.11%-3.40%。这是因为, 树木体内需要一定的养分量才能正常生长, 只有超过生长需要的那部分积累的养分才能迁移出来再利用(
Miller, 1986 ;
Marschner et al ., 1997 ;
Chen & Brassan, 2013 )。从本研究还可看出, 会同杉木林郁闭后, “仍存活物质”有一个较为稳定的速率迁移出养分重新用于物质生产。
植物组织的养分迁移是植物对养分贫瘠环境的一种适应机制, 也是植物保存养分并维持体内养分平衡的一种重要的养分利用策略(
Kobe et al ., 2005 ;
Milla et al ., 2005 ;
Huang & Wang, 2007 ;
Hosseini et al ., 2008 )。然而, 植物组织迁移养分的机制是什么? 这些迁移的养分是如何被活化重新用于物质生产的? 对于植物组织迁移养分的机制, 有研究提出, 土壤贫瘠生境中植物养分再吸收效率高于土壤肥沃生境中的植物, 土壤贫瘠的胁迫作用可能是植物组织养分迁移的机理(
Moghaddas & Stephens, 2007 )。也有研究指出, 植物组织的养分再吸收与土壤养分供给性相关不大, 贫瘠土壤并不一定是植物体养分迁移的驱动力(
Del Arco et al ., 1991 ;
Eckstein & Karlsson, 1997 ;
Aerts & Chapin, 1999 ;
王希华等, 2004 )。一些研究者用植物生理中“源-汇”理论分析植物组织内养分迁移机制, 认为树木器官在某些生理条件下是养分的“源”, 在另一些生理条件下可能是“汇”。正这种“源-汇”作用调控着树木体内的养分迁移(
Marschner et al ., 1997 )。如本研究中杉木快速增长阶段, 由于树木要壮大自己的个体, 因而将部分从土壤吸收的养分输送到“仍存活物质”内。这时杉木各器官成了养分的“汇”。当林分郁闭后, 生产单位生物量所需养分量下降(
Miller, 1986 ), 积累在“仍存活物质”内的养分被迁移出来用于新的物质生产, 这时杉木各器官成了养分的“源”。也有****认为, 树木体内养分迁移的机理是植物本身具有这种自我调控的反馈机制, 这种调控机制是植物在自然界长期的生存竞争中形成的(
Killingbeck, 1996 ;
Harrington et al ., 2001 )。可见, 对于植物组织养分迁移的机制还没有一个被广泛认可的解析。目前, 迁移的养分如何被活化且重新用于物质生产的机理研究, 更是一个非常薄弱的环节。树木体内的养分迁移及养分再利用是一个非常复杂的生理生化过程, 人们对此还缺乏基本的了解。因此, 揭示树木体内养分迁移和再利用的调控机制是生理生态学亟待解决的重要问题。
杉木林某林龄时凋落在林地的枝叶量与某林龄时的枯死枝叶量是不相等的。这是因为杉木枯死枝叶的宿存性, 枯死枝叶不会立即从树上掉落, 有些还残存在树上。如果用某林龄时凋落在林地的枝叶量估算某林龄时的枯死枝叶量, 就有可能忽略了那些存留在树上的枯死枝叶。此外, 那些存留在树上的枯死枝叶由于雨水的风化作用, 掉落后其养分含量有可能低于刚枯死的枝叶。因此, 本研究不同林龄时杉木枝叶的枯死量是根据杉木枝叶生长周期估算的, 而不是用某林龄时收集的林地凋落枝叶量估算的。而且测定枯死枝叶的养分样品不是从林地凋落枝叶中收集, 而是在活树上采集的, 这样就不用考虑枯死的枝叶是残存在树上还是掉落, 克服了枯死枝叶在树上宿存的影响, 同时也弥补了刚枯死枝叶和凋落枝叶的养分浓度差异的缺陷。由此推算出的不同林龄时枝叶枯死前迁移的养分及其再利用量, 所得的结果更接近实际。
本文虽然分析了“仍存活物质”中根的迁移养分的再利用, 也探讨了枝叶枯死前的养分迁移。但是, 植物细根在枯死前也存在养分转移, 而且这种养分循环对提高植物养分利用效率意义重大(
Meier et al ., 1985 ;
Ratnam & Sankara, 2008 )。由于很难准确估测细根的枯死量, 本文没有研究细根在枯死前转移的养分, 因此, 本研究得出的杉木枯死组织迁移的养分量要低于它的实际迁移量。
4 结论 (1)会同杉木林只要有枝叶枯死发生, 枝叶枯死前就有养分迁移出来用于物质再生产。枝叶枯死前迁移的再利用的养分, 以及这些养分生产的物质占总生产生物量的比例随林龄增加而增大。枝叶枯死前迁移的养分再利用的年变化受树木生长发育特征和枝叶的生理特性控制。
(2)会同杉木林郁闭后, 才发生“仍存活物质”迁移出的养分的再利用。这与不同生长发育阶段树木器官对养分的需求不同, 以及与林木体内的养分调节机制采取的策略有关。林分郁闭后, “仍存活物质”内有一个较为稳定的速率迁移出养分重新用于物质生产。
(3)会同1-25年生的杉木林内, 每年生产的物质中有3.52%-17.55%是利用枝叶枯死前转移的养分生产的; 12-25年生的林分内, 有3.11%-3.40%是利用某林龄段“仍存活物质”迁移出的养分生产的。枝叶枯死前迁移的养分和“仍存活物质”迁移出的养分也参与了林分物质生产过程。忽略了枝叶枯死前迁移的养分和“仍存活物质”迁移出的养分再利用, 就不能真实地反映林分养分利用特征和林分对养分的吸收和利用策略。
[16] Killingbeck KT 1996 ). Nutrient in senesced leaves: Keys to the search for potential resorption and resorption proficiencyEcology, 77, 1716 -1727 . [本文引用: 1] [17] Kobe RK Lepcryk CA Iyer M 2005 ). Resorption efficiency decreases with increasing green leaf nutrients in a global data setEcology, 86, 2780 -2792 . [本文引用: 1] [18] Li QL Mo QF Wang FM Li YW Xu X Zou B Li XB Chen Y Li ZA 2015 ). Nutrient utilization by Casuarina equisetifolia plantation of different ages in the tropical coastal area of South China Chinese Journal of Applied and Environmental Biology 139 -146 . [本文引用: 1] [ 李钦禄 , 莫其锋 , 王法明 , 李应文 , 徐馨 , 邹碧 , 李晓波 , 陈瑶 , 李志安 (2015 ). 华南热带沿海不同林龄木麻黄人工林养分利用特征应用与环境生物学报 , 21, 139 -146 .] [本文引用: 1] [19] Lim MT Cousens JF 1986 ). The internal transfer of nutrients in Scot pine stand 2. The patterns of transfer and the effects of nitrogen availabilityForestry, 59, 17 -21 . [本文引用: 1] [20] Lin BP He ZM Lin SZ Hu HT Qiu LJ Liu ZM 2017 ). Needles macronutrient concentrations and retrains location characteristics in Chinese fir plantations of different agesJournal of Forest and Environment, 37(1 ), 34 -39 . [本文引用: 1] [ 林宝平 , 何宗明 , 林思祖 , 胡欢甜 , 邱岭军 , 刘桌明 (2017 ). 不同林龄杉木针叶大量元素转移特征森林与环境学报 , 37(1 ), 34 -39 .] [本文引用: 1] [21] Lin DX Liu KH Luo SF 2002 ). Dynamics and cycling analysis of nutrient elements in Eucalyptus urophylla Chinese Journal of Applied and Environmental Biology, 148 -153 . [本文引用: 1] [ 林德喜 , 刘开汉 , 罗水发 (2002 ). 尾叶桉营养元素动态和循环分析应用与环境生物学报 , 8, 148 -153 .] [本文引用: 1] [22] Liu AQ Fan SH Lin KM Ma XQ Sheng WT 2005 ). Comparison on nutrient cycling in different generation plantations of Chinese firPlant Nutrition and Fertilizer Science, 11, 273 -278 . [本文引用: 2] [ 刘爱琴 , 范少辉 , 林开敏 , 马祥庆 , 盛炜彤 (2005 ). 不同栽植代数杉木林养分循环的比较研究植物营养与肥料学报 , 11, 273 -278 .] [本文引用: 2] [23] Marschner H Kirkby EA Engels C 1997 ). Importance of cycling and recycling of mineral nutrients within plants for growth and developmentBotanica Acta, 110, 265 -273 . [本文引用: 2] [24] Mayor JR Wright SJ Turner BL 2013 ). Species-specific responses of foliar nutrients to long-term nitrogen and phosphorus additions in a lowland tropical forestJournal of Ecology, 102, 36 -44 . [本文引用: 1] [25] Mei L Wang ZQ Cheng YH Guo DL 2004 ). A review: Factors influencing fine root longevity in forest ecosystemsActa Phytoecologica Sinica, 28, 704 -710 . [本文引用: 1] [ 梅莉 , 王政权 , 程云环 , 郭大立 (2004 ). 林木细根寿命及其影响因子研究进展植物生态学报 , 28, 704 -710 .] [本文引用: 1] [26] Meier CE Grier CC Cole DW 1985 ). Below and ground N and P use by Abies amabilis standsEcology, 1928 -1942 . [本文引用: 1] [27] Milla R Castro-Diez P Maestro-Martinez M 2005 ). Does the gradualness of leaf shedding govern nutrient resorption from senescing leaves in Mediterranean woody plants?Plant and Soil, 278, 303 -313 . [本文引用: 1] [28] Miller HG 1984 ). Dynamics of nutrient cycling in plantation ecosystems. In: Bowen GD, Nambiar EKS eds. Nutrition of Plantation Forests . Academic Press, London. 53 -78 . [本文引用: 1] [29] Miller HG 1986 ). Carbon × nutrient interaction—The limitations to productivityTree Physiology, 2, 373 -385 . [本文引用: 4] [30] Moghaddas EEY Stephens SL 2007 ). Thinning burning and thin-burn fuel treatment effects on soil properties in a Sierra Nevada mixed conifer forestsForest Ecology and Management, 250, 156 -166 . [本文引用: 1] [1] Aerts R 1990 ). Nutrient use efficiency in evergreen and deciduous species from heath landsOecologia, 84, 391 -397 . [本文引用: 1] [2] Aerts R Chapin III FS 1999 ). The mineral nutrition of wild plants revisited: A reevaluation of processes and patternsAdvances in Ecological Research, 37, 1 -67 . [本文引用: 1] [31] Ratnam J Sankaran M 2008 ). Hanan in a tropical savanna: Variation N, P nutrient resorption patterns of plant functional groups and functional significanceOecologia , 157, 141 -151 . [本文引用: 1] [32] Shaver GR Melillo JM 1984 ). Nutrient budgets of marsh plants: Efficiency concepts and relation to availabilityEcology , 65, 1491 -1510 . [本文引用: 1] [3] An Z Niu DC Wen HY Yang Y Zhang HR Fu H 2011 ). Effects of N addition on nutrient resorption efficiency and C:N:P stoichiometric characteristics in Stipa bungeana of steppe grass lands in the Loess Plateau, ChinaChinese Journal of Plant Ecology, 801 -807 . [本文引用: 1] [ 安卓 , 牛得草 , 文海燕 , 杨益 , 张洪荣 , 傅华 (2011 ). 氮素添加对黄土高原典型草原长芒草氮磷重吸收率及C:N:P化学计量特征的影响植物生态学报 , 35, 801 -807 .] [本文引用: 1] [33] Tian DL Shen Y Kang WX Xiang WH Yan WD Deng XW 2011 ). Characteristics of nutrient cycling in first and second rotations of Chinese fir plantationsActa Ecologica Sinica, 31, 5025 -5032 . [本文引用: 1] [ 田大伦 , 沈燕 , 康文星 , 项文化 , 闫文德 , 邓湘雯 (2011 ). 连栽第1和第2代杉木人工林养分循环的比较生态学报 , 31, 5025 -5032 .] [本文引用: 1] [4] Cao JH Tao ZL Jiang JS Xie GS Zhao CM 2010 ). Nutrient use efficiency of clone PR107 at various ageChinese Journal of Tropical Crops, 31, 2091 -097 . [本文引用: 1] [ 曹建华 , 陶忠良 , 蒋菊生 , 谢贵水 , 赵春梅 (2010 ). 不同年龄橡胶树PR107养分利用效率研究热带作物学报 , 31, 2091 -2097 .] [本文引用: 1] [34] Wang RL Cheng RM Xiao WF Feng XH Liu ZB Wang XR 2012 ). Influencing factors of fine root production and turnover in forest ecosystemWorld Forestry Research, 21(1 ), 19 -24 . [ 王瑞丽 , 程瑞梅 , 肖文发 , 封晓辉 , 刘泽彬 , 王晓荣 (2012 ). 森林细根生产和周转的影响因素世界林业研究 , 21(1 ), 19 -24 .] [35] Wang XH Huang JJ Yan ER 2004 ). A study on leaf nutrient resorption of some trees in Tiantong National Forest ParkChinese Journal of Ecology, 33(4 ), 13 -16 . [本文引用: 1] [5] Chen HYH Brassard BW 2013 ). Intrinsic and extrinsic controls of fine root life spanCritical Reviews in Plant Sciences, 32, 151 -161 . [本文引用: 2] [6] Chen RS Kang WX Zhou YQ Tian DL Xiang WH 2018 ). Changes in nutrient cycling with age in a Cunninghamia lanceolata plantation forestChinese Journal of Plant Ecology, 173 -184 . [本文引用: 2] [35] [ 王希华 , 黄建军 , 闫恩荣 (2004 ). 天童国家森林公园若干树种叶水平上养分利用效率的研究生态学杂志 , 33(4 ), 13 -16 .] [本文引用: 1] [36] Williams RF 1955 ). Redistribution of mineral elements during developmentAnnual Review of Plant Physiology, 6, 25 -42 . [本文引用: 1] [6] [ 陈日升 , 康文星 , 周玉泉 , 田大伦 , 项文化 (2018 ). 杉木人工林养分循环随林龄变化的特征植物生态学报 , 42, 173 -184 .] [本文引用: 2] [7] Del Arco JM Esucdero A Garrido MV 1991 ). Effects of site characteristics on nitrogen retrains location from senescing leavesEcology, 58, 701 -708 . [本文引用: 1] [37] Wu PF Ma XQ 2009 ). Research advances in the mechanisms of high nutrient use efficiency in plantsActa Ecologica Sinica, 29, 427 -437 . [本文引用: 1] [ 吴鹏飞 , 马祥庆 (2009 ). 植物养分高效利用机制研究进展生态学报 , 29, 427 -437 .] [本文引用: 1] [8] Deng HJ Chen AM Yan SW Lin YM Zhang GS Du K Wu CZ Hong W 2015 ). Nutrient resorption efficiency and C:N:P stoichiometry in different ages of Leucaena leucocephala Chinese Journal of Applied and Environmental Biology, 522 -527 . [本文引用: 1] [ 邓浩俊 , 陈爱民 , 严思维 , 林勇明 , 张广帅 , 杜锟 , 吴承祯 , 洪伟 (2015 ). 不同林龄新银合欢重吸收率及其C:N:P化学计量特征应用与环境生物学报 , 21, 522 -527 .] [本文引用: 1] [38] Xiang WH Tian DL 2002 ). Nutrient cycling in Pinus massoniana stands of different age classesActa Phytoecologica Sinica, 89 -95 . [本文引用: 1] [ 项文化 , 田大伦 (2002 ). 不同年龄阶段马尾松人工林养分循环的研究植物生态学报 , 26, 89 -95 .] [本文引用: 1] [9] Division of Science and Technology of Ministry of Forestry Compiles (1994 ). Forest Ecosystem Research Methods . China Science and Technology Press, Beijing. 156 -158 . [本文引用: 2] [ 林业部科技司 (1994 ). 森林生态系统研究方法 . 中国科学技术出版社, 北京. 156 -158 .] [本文引用: 2] [39] Xiao XC Li ZH Tang ZJ Zeng Q Wang HF 2013 ). Effects of stand density on nutrient cycling rate and use efficiency of Pinus elliottii plantationChinese Journal of Ecology 2871 -2880 . [本文引用: 1] [ 肖兴翠 , 李志辉 , 唐作钧 , 曾琴 , 王海风 (2013 ). 林分密度对湿地松人工林养分循环速率和利用效率的影响生态学杂志 , 32, 2871 -2880 .] [本文引用: 1] [10] Eckstein RL Karlsson PS 1977 ). Above-ground growth and nitrogen use by plant in a subarctic environment: Effects of habitat life-form and speciesOikos 311 -324 . [本文引用: 1] [11] Harrington RA Fownes JH Vitousek PM 2001 ). Production and resource use efficiencies in N- and P-limited tropical forests: A comparison of responses to long-term fertilizationEcosystems , 4, 646 -657 . [本文引用: 1] [40] Ye GF 2012 ). Age-related changes in nutrient resorption patterns and tannin concentration of Casuarina equisetifolia plantationsJournal of Tropical Forest Science 546 -556 . [本文引用: 1] [41] Zeng DH Chen GS Chen FS Zhao Q Ji XY 2005 ). Foliar nutrients and their resorption efficiencies in four Pinus sylvestris var. mongolica plantations of different ages on sandy soilScientia Silvae Sinicae 5 ), 21 -27 . DOI:
10.11707/j.1001-7488.20050504 Magsci [本文引用: 1] <p>树木叶片的养分再吸收效率能够反映树木对养分保存、利用以及对养分贫瘠环境的适应能力。以科尔沁沙地东南缘章古台地区樟子松人工林为研究对象,分析了11、20、29、45年生树木叶片的基本特征、养分含量及其再吸收效率。结果表明:叶片衰老后其质量和面积明显减少;叶片凋落前的平均养分含量没有表现出随樟子松年龄增加而出现有规律的变化;凋落叶片中的N、P、K、Mg含量表现出随年龄增加而增加的趋势,而Ca的趋势与之相反;11年生和20年生的樟子松叶片N、P、K的再吸收效率相似,都显著高于29年生和45年生樟子松(
P<0.05),而樟子松叶片对Mg的再吸收效率表现出随年龄增大而显著降低,Ca随叶片的衰老而不断累积,再吸收效率表现为负值,20年生的樟子松叶片Ca再吸收效率最大,11年生和45年生最低。樟子松叶片的N、P、K、Mg养分再吸收效率随年龄增加而降低的趋势表明,随年龄增加樟子松对贫瘠养分生境的适应能力逐渐降低,反映了樟子松养分保存方面的衰退特征。 </p>
[12] He B Qin WM Yu HG Liu YH Qin L Qin YH 2007 ). Biological cycling of nutrients in different ages classes of Acacia mangium plantationActa Ecologica Sinica, 5158 -5167 . [本文引用: 1] [ 何斌 , 秦武明 , 余浩光 , 刘运华 , 覃林 , 覃永华 (2007 ). 不同年龄阶段马占相思(Acacia mangium )人工林营养元素的生物循环生态学报 , 27, 5158 -5167 .] [本文引用: 1] [41] [ 曾德慧 , 陈广生 , 陈伏生 , 赵琼 , 冀小燕 (2005 ). 不同林龄樟子松叶片养分含量及其再吸收效率林业科学 , 41(5 ), 21 -27 .] DOI:
10.11707/j.1001-7488.20050504 Magsci [本文引用: 1] <p>树木叶片的养分再吸收效率能够反映树木对养分保存、利用以及对养分贫瘠环境的适应能力。以科尔沁沙地东南缘章古台地区樟子松人工林为研究对象,分析了11、20、29、45年生树木叶片的基本特征、养分含量及其再吸收效率。结果表明:叶片衰老后其质量和面积明显减少;叶片凋落前的平均养分含量没有表现出随樟子松年龄增加而出现有规律的变化;凋落叶片中的N、P、K、Mg含量表现出随年龄增加而增加的趋势,而Ca的趋势与之相反;11年生和20年生的樟子松叶片N、P、K的再吸收效率相似,都显著高于29年生和45年生樟子松(
P<0.05),而樟子松叶片对Mg的再吸收效率表现出随年龄增大而显著降低,Ca随叶片的衰老而不断累积,再吸收效率表现为负值,20年生的樟子松叶片Ca再吸收效率最大,11年生和45年生最低。樟子松叶片的N、P、K、Mg养分再吸收效率随年龄增加而降低的趋势表明,随年龄增加樟子松对贫瘠养分生境的适应能力逐渐降低,反映了樟子松养分保存方面的衰退特征。 </p>
[42] Zeng Q Gao GW Lin YM Fan HQ 2008 ). Resorption efficiencies of nitrogen and phosphorus of leaves during senescence for two growth forms of Avicennia marina Journal of Xiamen University (Natural Science )181 -185 . [本文引用: 1] [13] Hosseini SM Rouhi-Moghaddam E Ebrahimi E 2008 ). Comparison of growth, nutrition and soil properties of pure stands of Quercus castaneifolia and mixed with Zelkova carpinifolia in the Hyrcanian forests of IranForest Ecology and Management 1149 -1160 . [本文引用: 1] [14] Huang JJ Wang XH 2007 ). Leaf nutrient concentration, nutrient resorption and litter decomposition in an evergreen broad-leaved forest in eastern ChinaForest Ecology and Management, 239, 150 -158 . [本文引用: 1] [42] [ 曾琦 , 高国伟 , 林益明 , 范航清 (2008 ). 红树植物白骨壤叶片衰老过程的氮磷内吸收变化研究厦门大学学报(自然科学版) , 47, 181 -185 .] [本文引用: 1] [43] Zhang LH Lin YM Ye GF Yin L Zhou HC 2009 ). Nitrogen and phosphorus concentrations, N:P ratio and resorption efficiency of leaves in different forest typesJournal of Beijing Forestry University, 31(5 ), 67 -72 . [本文引用: 1] [ 张立华 , 林益明 , 叶功富 , 殷亮 , 周海超 (2009 ). 不同林分类型叶片氮磷含量, 氮磷比及其内吸收率北京林业大学学报 , 31(5 ), 67 -72 .] [本文引用: 1] [44] Zhao Q Liu XY Hu YL Zeng DH 2010 ). Effects of nitrogen addition on nutrient allocation and nutrient resorption efficiency in Larix gmelinii Scientia Silvae Sinicae, 5 ), 14 -19 . DOI:
10.11707/j.1001-7488.20100503 Magsci [本文引用: 1] <p><font face="Verdana">以东北林业大学帽儿山实验林场18年生兴安落叶松人工林为对象,研究连续5年N添加(NH
4 NO
3 ,10 g·m
-2 a
-1 )对落叶松各器官N,P,K,Ca和Mg含量及叶片养分再吸收效率的影响。结果表明:N添加对叶片5种元素含量均无显著影响,但可显著提高树根、树枝和树干的N和P含量; N添加能显著提高凋落叶片的N含量,降低凋落叶片的C/N比和叶片N再吸收效率。研究结果反映出研究区兴安落叶松人工林土壤N并不缺乏。N添加导致兴安落叶松对N和P的奢侈吸收,可提高凋落物的分解速率,加快兴安落叶松人工林生态系统的N循环。</font></p>
[ 赵琼 , 刘兴宇 , 胡亚林 , 曾德慧 (2010 ). 氮添加对兴安落叶松养分分配和再吸收效率的影响林业科学 , 46(5 ), 14 -19 .] DOI:
10.11707/j.1001-7488.20100503 Magsci [本文引用: 1] <p><font face="Verdana">以东北林业大学帽儿山实验林场18年生兴安落叶松人工林为对象,研究连续5年N添加(NH
4 NO
3 ,10 g·m
-2 a
-1 )对落叶松各器官N,P,K,Ca和Mg含量及叶片养分再吸收效率的影响。结果表明:N添加对叶片5种元素含量均无显著影响,但可显著提高树根、树枝和树干的N和P含量; N添加能显著提高凋落叶片的N含量,降低凋落叶片的C/N比和叶片N再吸收效率。研究结果反映出研究区兴安落叶松人工林土壤N并不缺乏。N添加导致兴安落叶松对N和P的奢侈吸收,可提高凋落物的分解速率,加快兴安落叶松人工林生态系统的N循环。</font></p>
[45] Zheng Y Guo YR Wang MT Li M Fan RR Sun J Yang FC Zhong QL Cheng DL 2017 ). Foliar nutrients and their resorption efficiencies of Pinus hwangshanensis along an elevation gradient of Wuyi Mountains in JiangxiJournal of Anhui Agricultural University 415 -421 . [本文引用: 1] [ 郑媛 , 郭英荣 , 王满堂 , 李曼 , 范瑞瑞 , 孙俊 , 杨福春 , 钟全林 , 程栋梁 (2017 ). 武夷山不同海拔梯度黄山松叶片养分含量及其再吸收效率安徽农业大学学报 , 44, 415 -421 .] [本文引用: 1] [46] Zhou LL 2014 ). Study on Nutrient Retranslocation and Nutrient Use Efficiency in Different Developmental-Staged Chinese Fir Plantations. PhD dissertation, Fujian Agriculture and Forestry University, Fuzhou . 80 -82 . [本文引用: 1] [ 周丽丽 (2014 ). 不同发育阶段杉木人工林养分内循环与周转利用效率的研究博士学位论文, 福建农林大学, 福州 . 80 -82 .] [本文引用: 1] [47] Zong N Shi PL Geng SB Ma WL 2017 ). Nitrogen and phosphorus resorption efficiency of forests in North ChinaChinese Journal of Eco-Agriculture, 25, 520 -529 . [本文引用: 1] [15] Killingbeck KT 1986 ). The terminological jungle revisited: Making a case for use of the term resorptionOikos, 46, 263 -264 . [本文引用: 1] [47] [ 宗宁 , 石培礼 , 耿守保 , 马维玲 (2017 ). 北方山区主要森林类型树木叶片氮、磷回收效率研究中国生态农业学报 , 25, 520 -529 .] [本文引用: 1] Nutrient in senesced leaves: Keys to the search for potential resorption and resorption proficiency
1
1996
... 植物组织的养分迁移是植物对养分贫瘠环境的一种适应机制, 也是植物保存养分并维持体内养分平衡的一种重要的养分利用策略(
Kobe et al ., 2005 ;
Milla et al ., 2005 ;
Huang & Wang, 2007 ;
Hosseini et al ., 2008 ).然而, 植物组织迁移养分的机制是什么? 这些迁移的养分是如何被活化重新用于物质生产的? 对于植物组织迁移养分的机制, 有研究提出, 土壤贫瘠生境中植物养分再吸收效率高于土壤肥沃生境中的植物, 土壤贫瘠的胁迫作用可能是植物组织养分迁移的机理(
Moghaddas & Stephens, 2007 ).也有研究指出, 植物组织的养分再吸收与土壤养分供给性相关不大, 贫瘠土壤并不一定是植物体养分迁移的驱动力(
Del Arco et al ., 1991 ;
Eckstein & Karlsson, 1997 ;
Aerts & Chapin, 1999 ;
王希华等, 2004 ).一些研究者用植物生理中“源-汇”理论分析植物组织内养分迁移机制, 认为树木器官在某些生理条件下是养分的“源”, 在另一些生理条件下可能是“汇”.正这种“源-汇”作用调控着树木体内的养分迁移(
Marschner et al ., 1997 ).如本研究中杉木快速增长阶段, 由于树木要壮大自己的个体, 因而将部分从土壤吸收的养分输送到“仍存活物质”内.这时杉木各器官成了养分的“汇”.当林分郁闭后, 生产单位生物量所需养分量下降(
Miller, 1986 ), 积累在“仍存活物质”内的养分被迁移出来用于新的物质生产, 这时杉木各器官成了养分的“源”.也有****认为, 树木体内养分迁移的机理是植物本身具有这种自我调控的反馈机制, 这种调控机制是植物在自然界长期的生存竞争中形成的(
Killingbeck, 1996 ;
Harrington et al ., 2001 ).可见, 对于植物组织养分迁移的机制还没有一个被广泛认可的解析.目前, 迁移的养分如何被活化且重新用于物质生产的机理研究, 更是一个非常薄弱的环节.树木体内的养分迁移及养分再利用是一个非常复杂的生理生化过程, 人们对此还缺乏基本的了解.因此, 揭示树木体内养分迁移和再利用的调控机制是生理生态学亟待解决的重要问题. ...
Resorption efficiency decreases with increasing green leaf nutrients in a global data set
1
2005
... 植物组织的养分迁移是植物对养分贫瘠环境的一种适应机制, 也是植物保存养分并维持体内养分平衡的一种重要的养分利用策略(
Kobe et al ., 2005 ;
Milla et al ., 2005 ;
Huang & Wang, 2007 ;
Hosseini et al ., 2008 ).然而, 植物组织迁移养分的机制是什么? 这些迁移的养分是如何被活化重新用于物质生产的? 对于植物组织迁移养分的机制, 有研究提出, 土壤贫瘠生境中植物养分再吸收效率高于土壤肥沃生境中的植物, 土壤贫瘠的胁迫作用可能是植物组织养分迁移的机理(
Moghaddas & Stephens, 2007 ).也有研究指出, 植物组织的养分再吸收与土壤养分供给性相关不大, 贫瘠土壤并不一定是植物体养分迁移的驱动力(
Del Arco et al ., 1991 ;
Eckstein & Karlsson, 1997 ;
Aerts & Chapin, 1999 ;
王希华等, 2004 ).一些研究者用植物生理中“源-汇”理论分析植物组织内养分迁移机制, 认为树木器官在某些生理条件下是养分的“源”, 在另一些生理条件下可能是“汇”.正这种“源-汇”作用调控着树木体内的养分迁移(
Marschner et al ., 1997 ).如本研究中杉木快速增长阶段, 由于树木要壮大自己的个体, 因而将部分从土壤吸收的养分输送到“仍存活物质”内.这时杉木各器官成了养分的“汇”.当林分郁闭后, 生产单位生物量所需养分量下降(
Miller, 1986 ), 积累在“仍存活物质”内的养分被迁移出来用于新的物质生产, 这时杉木各器官成了养分的“源”.也有****认为, 树木体内养分迁移的机理是植物本身具有这种自我调控的反馈机制, 这种调控机制是植物在自然界长期的生存竞争中形成的(
Killingbeck, 1996 ;
Harrington et al ., 2001 ).可见, 对于植物组织养分迁移的机制还没有一个被广泛认可的解析.目前, 迁移的养分如何被活化且重新用于物质生产的机理研究, 更是一个非常薄弱的环节.树木体内的养分迁移及养分再利用是一个非常复杂的生理生化过程, 人们对此还缺乏基本的了解.因此, 揭示树木体内养分迁移和再利用的调控机制是生理生态学亟待解决的重要问题. ...
华南热带沿海不同林龄木麻黄人工林养分利用特征
1
2015
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
华南热带沿海不同林龄木麻黄人工林养分利用特征
1
2015
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
The internal transfer of nutrients in Scot pine stand 2. The patterns of transfer and the effects of nitrogen availability
1
1986
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
不同林龄杉木针叶大量元素转移特征
1
2017
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
不同林龄杉木针叶大量元素转移特征
1
2017
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
尾叶桉营养元素动态和循环分析
1
2002
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
尾叶桉营养元素动态和循环分析
1
2002
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
不同栽植代数杉木林养分循环的比较研究
2
2005
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
... 根据杉木枝、叶生长周期(寿命)估算不同林龄时林分枝叶枯死量.杉木叶片生长周期定为5年(
刘爱琴等, 2005 ), 杉木树枝生长周期定为10年(根据在会同长期观测的结果).用下式估算单位面积林分不同林龄时的枝、叶枯死量. ...
不同栽植代数杉木林养分循环的比较研究
2
2005
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
... 根据杉木枝、叶生长周期(寿命)估算不同林龄时林分枝叶枯死量.杉木叶片生长周期定为5年(
刘爱琴等, 2005 ), 杉木树枝生长周期定为10年(根据在会同长期观测的结果).用下式估算单位面积林分不同林龄时的枝、叶枯死量. ...
Importance of cycling and recycling of mineral nutrients within plants for growth and development
2
1997
... 会同杉木林生长12年时, 才发生某林龄段“仍存活物质”迁移出养分用于物质生产.这主要由杉木不同生长阶段对养分的不同需求引起的.杉木从幼树到快速生长阶段为了壮大自己的个体, 为树木未来生长打好基础, 已生产的物质(器官)中需要不断补充养分.因此, 这个生育阶段的“仍存活物质”中不仅没有养分转移出来再利用, 而且还需要新的养分补充.如本研究中8-11年生的林分吸收的土壤养分中, 每年还需输送7.83 kg·hm
-2 到该林龄段“仍存活物质”内(
表2 ).此外,
Miller (1986) 的研究表明, 林分郁闭后, 多年生乔木林生产单位物质所需的养分量减少.树木体内养分再分配机制控制着将积累在“仍存活物质”内的养分迁移出来重新用于物质生产.从本研究结果看, 林分生长12年后(杉木林已郁闭), 虽然从“仍存活物质”内迁移出的养分并不很多, 但“仍存活物质”迁移出的养分生产的物质量仍占生产物质总量的3.11%-3.40%.这是因为, 树木体内需要一定的养分量才能正常生长, 只有超过生长需要的那部分积累的养分才能迁移出来再利用(
Miller, 1986 ;
Marschner et al ., 1997 ;
Chen & Brassan, 2013 ).从本研究还可看出, 会同杉木林郁闭后, “仍存活物质”有一个较为稳定的速率迁移出养分重新用于物质生产. ...
... 植物组织的养分迁移是植物对养分贫瘠环境的一种适应机制, 也是植物保存养分并维持体内养分平衡的一种重要的养分利用策略(
Kobe et al ., 2005 ;
Milla et al ., 2005 ;
Huang & Wang, 2007 ;
Hosseini et al ., 2008 ).然而, 植物组织迁移养分的机制是什么? 这些迁移的养分是如何被活化重新用于物质生产的? 对于植物组织迁移养分的机制, 有研究提出, 土壤贫瘠生境中植物养分再吸收效率高于土壤肥沃生境中的植物, 土壤贫瘠的胁迫作用可能是植物组织养分迁移的机理(
Moghaddas & Stephens, 2007 ).也有研究指出, 植物组织的养分再吸收与土壤养分供给性相关不大, 贫瘠土壤并不一定是植物体养分迁移的驱动力(
Del Arco et al ., 1991 ;
Eckstein & Karlsson, 1997 ;
Aerts & Chapin, 1999 ;
王希华等, 2004 ).一些研究者用植物生理中“源-汇”理论分析植物组织内养分迁移机制, 认为树木器官在某些生理条件下是养分的“源”, 在另一些生理条件下可能是“汇”.正这种“源-汇”作用调控着树木体内的养分迁移(
Marschner et al ., 1997 ).如本研究中杉木快速增长阶段, 由于树木要壮大自己的个体, 因而将部分从土壤吸收的养分输送到“仍存活物质”内.这时杉木各器官成了养分的“汇”.当林分郁闭后, 生产单位生物量所需养分量下降(
Miller, 1986 ), 积累在“仍存活物质”内的养分被迁移出来用于新的物质生产, 这时杉木各器官成了养分的“源”.也有****认为, 树木体内养分迁移的机理是植物本身具有这种自我调控的反馈机制, 这种调控机制是植物在自然界长期的生存竞争中形成的(
Killingbeck, 1996 ;
Harrington et al ., 2001 ).可见, 对于植物组织养分迁移的机制还没有一个被广泛认可的解析.目前, 迁移的养分如何被活化且重新用于物质生产的机理研究, 更是一个非常薄弱的环节.树木体内的养分迁移及养分再利用是一个非常复杂的生理生化过程, 人们对此还缺乏基本的了解.因此, 揭示树木体内养分迁移和再利用的调控机制是生理生态学亟待解决的重要问题. ...
Species-specific responses of foliar nutrients to long-term nitrogen and phosphorus additions in a lowland tropical forest
1
2013
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
林木细根寿命及其影响因子研究进展
1
2004
... 单位面积林分某林龄段生产的树干、树皮生物量, 根据生长过程中树干、树皮凋落甚少的规律, 用某林龄段开始和结束时单位面积林分测定的这些器官现存量之差求得.根系存在更新代谢过程, 但要准确测定根系(尤其是细根)的生长周期, 需采用微根管技术或同位素追踪技术.这些技术方法成本高且操作有一定的难度, 因而大多数研究者在估算某林龄段根生产量时, 将根与干材和树皮同等对待(
梅莉等, 2004 ;
王端丽等, 2012 ).本研究也是如此(这样估算出的根生产量要低于它的实际生产量).杉木是常绿树种, 枝、叶都有一定的生长周期, 根据
陈日升等(2018) 的研究方法, 估算出某林龄段这些还存活的枝叶量.然后用下式估算某林龄段单位面积林分生产的生物量(
W ). ...
林木细根寿命及其影响因子研究进展
1
2004
... 单位面积林分某林龄段生产的树干、树皮生物量, 根据生长过程中树干、树皮凋落甚少的规律, 用某林龄段开始和结束时单位面积林分测定的这些器官现存量之差求得.根系存在更新代谢过程, 但要准确测定根系(尤其是细根)的生长周期, 需采用微根管技术或同位素追踪技术.这些技术方法成本高且操作有一定的难度, 因而大多数研究者在估算某林龄段根生产量时, 将根与干材和树皮同等对待(
梅莉等, 2004 ;
王端丽等, 2012 ).本研究也是如此(这样估算出的根生产量要低于它的实际生产量).杉木是常绿树种, 枝、叶都有一定的生长周期, 根据
陈日升等(2018) 的研究方法, 估算出某林龄段这些还存活的枝叶量.然后用下式估算某林龄段单位面积林分生产的生物量(
W ). ...
Below and ground N and P use by
Abies amabilis stands
1
1985
... 本文虽然分析了“仍存活物质”中根的迁移养分的再利用, 也探讨了枝叶枯死前的养分迁移.但是, 植物细根在枯死前也存在养分转移, 而且这种养分循环对提高植物养分利用效率意义重大(
Meier et al ., 1985 ;
Ratnam & Sankara, 2008 ).由于很难准确估测细根的枯死量, 本文没有研究细根在枯死前转移的养分, 因此, 本研究得出的杉木枯死组织迁移的养分量要低于它的实际迁移量. ...
Does the gradualness of leaf shedding govern nutrient resorption from senescing leaves in Mediterranean woody plants?
1
2005
... 植物组织的养分迁移是植物对养分贫瘠环境的一种适应机制, 也是植物保存养分并维持体内养分平衡的一种重要的养分利用策略(
Kobe et al ., 2005 ;
Milla et al ., 2005 ;
Huang & Wang, 2007 ;
Hosseini et al ., 2008 ).然而, 植物组织迁移养分的机制是什么? 这些迁移的养分是如何被活化重新用于物质生产的? 对于植物组织迁移养分的机制, 有研究提出, 土壤贫瘠生境中植物养分再吸收效率高于土壤肥沃生境中的植物, 土壤贫瘠的胁迫作用可能是植物组织养分迁移的机理(
Moghaddas & Stephens, 2007 ).也有研究指出, 植物组织的养分再吸收与土壤养分供给性相关不大, 贫瘠土壤并不一定是植物体养分迁移的驱动力(
Del Arco et al ., 1991 ;
Eckstein & Karlsson, 1997 ;
Aerts & Chapin, 1999 ;
王希华等, 2004 ).一些研究者用植物生理中“源-汇”理论分析植物组织内养分迁移机制, 认为树木器官在某些生理条件下是养分的“源”, 在另一些生理条件下可能是“汇”.正这种“源-汇”作用调控着树木体内的养分迁移(
Marschner et al ., 1997 ).如本研究中杉木快速增长阶段, 由于树木要壮大自己的个体, 因而将部分从土壤吸收的养分输送到“仍存活物质”内.这时杉木各器官成了养分的“汇”.当林分郁闭后, 生产单位生物量所需养分量下降(
Miller, 1986 ), 积累在“仍存活物质”内的养分被迁移出来用于新的物质生产, 这时杉木各器官成了养分的“源”.也有****认为, 树木体内养分迁移的机理是植物本身具有这种自我调控的反馈机制, 这种调控机制是植物在自然界长期的生存竞争中形成的(
Killingbeck, 1996 ;
Harrington et al ., 2001 ).可见, 对于植物组织养分迁移的机制还没有一个被广泛认可的解析.目前, 迁移的养分如何被活化且重新用于物质生产的机理研究, 更是一个非常薄弱的环节.树木体内的养分迁移及养分再利用是一个非常复杂的生理生化过程, 人们对此还缺乏基本的了解.因此, 揭示树木体内养分迁移和再利用的调控机制是生理生态学亟待解决的重要问题. ...
1
1984
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
Carbon × nutrient interaction—The limitations to productivity
4
1986
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
... 会同杉木林生长12年时, 才发生某林龄段“仍存活物质”迁移出养分用于物质生产.这主要由杉木不同生长阶段对养分的不同需求引起的.杉木从幼树到快速生长阶段为了壮大自己的个体, 为树木未来生长打好基础, 已生产的物质(器官)中需要不断补充养分.因此, 这个生育阶段的“仍存活物质”中不仅没有养分转移出来再利用, 而且还需要新的养分补充.如本研究中8-11年生的林分吸收的土壤养分中, 每年还需输送7.83 kg·hm
-2 到该林龄段“仍存活物质”内(
表2 ).此外,
Miller (1986) 的研究表明, 林分郁闭后, 多年生乔木林生产单位物质所需的养分量减少.树木体内养分再分配机制控制着将积累在“仍存活物质”内的养分迁移出来重新用于物质生产.从本研究结果看, 林分生长12年后(杉木林已郁闭), 虽然从“仍存活物质”内迁移出的养分并不很多, 但“仍存活物质”迁移出的养分生产的物质量仍占生产物质总量的3.11%-3.40%.这是因为, 树木体内需要一定的养分量才能正常生长, 只有超过生长需要的那部分积累的养分才能迁移出来再利用(
Miller, 1986 ;
Marschner et al ., 1997 ;
Chen & Brassan, 2013 ).从本研究还可看出, 会同杉木林郁闭后, “仍存活物质”有一个较为稳定的速率迁移出养分重新用于物质生产. ...
... 的研究表明, 林分郁闭后, 多年生乔木林生产单位物质所需的养分量减少.树木体内养分再分配机制控制着将积累在“仍存活物质”内的养分迁移出来重新用于物质生产.从本研究结果看, 林分生长12年后(杉木林已郁闭), 虽然从“仍存活物质”内迁移出的养分并不很多, 但“仍存活物质”迁移出的养分生产的物质量仍占生产物质总量的3.11%-3.40%.这是因为, 树木体内需要一定的养分量才能正常生长, 只有超过生长需要的那部分积累的养分才能迁移出来再利用(
Miller, 1986 ;
Marschner et al ., 1997 ;
Chen & Brassan, 2013 ).从本研究还可看出, 会同杉木林郁闭后, “仍存活物质”有一个较为稳定的速率迁移出养分重新用于物质生产. ...
... 植物组织的养分迁移是植物对养分贫瘠环境的一种适应机制, 也是植物保存养分并维持体内养分平衡的一种重要的养分利用策略(
Kobe et al ., 2005 ;
Milla et al ., 2005 ;
Huang & Wang, 2007 ;
Hosseini et al ., 2008 ).然而, 植物组织迁移养分的机制是什么? 这些迁移的养分是如何被活化重新用于物质生产的? 对于植物组织迁移养分的机制, 有研究提出, 土壤贫瘠生境中植物养分再吸收效率高于土壤肥沃生境中的植物, 土壤贫瘠的胁迫作用可能是植物组织养分迁移的机理(
Moghaddas & Stephens, 2007 ).也有研究指出, 植物组织的养分再吸收与土壤养分供给性相关不大, 贫瘠土壤并不一定是植物体养分迁移的驱动力(
Del Arco et al ., 1991 ;
Eckstein & Karlsson, 1997 ;
Aerts & Chapin, 1999 ;
王希华等, 2004 ).一些研究者用植物生理中“源-汇”理论分析植物组织内养分迁移机制, 认为树木器官在某些生理条件下是养分的“源”, 在另一些生理条件下可能是“汇”.正这种“源-汇”作用调控着树木体内的养分迁移(
Marschner et al ., 1997 ).如本研究中杉木快速增长阶段, 由于树木要壮大自己的个体, 因而将部分从土壤吸收的养分输送到“仍存活物质”内.这时杉木各器官成了养分的“汇”.当林分郁闭后, 生产单位生物量所需养分量下降(
Miller, 1986 ), 积累在“仍存活物质”内的养分被迁移出来用于新的物质生产, 这时杉木各器官成了养分的“源”.也有****认为, 树木体内养分迁移的机理是植物本身具有这种自我调控的反馈机制, 这种调控机制是植物在自然界长期的生存竞争中形成的(
Killingbeck, 1996 ;
Harrington et al ., 2001 ).可见, 对于植物组织养分迁移的机制还没有一个被广泛认可的解析.目前, 迁移的养分如何被活化且重新用于物质生产的机理研究, 更是一个非常薄弱的环节.树木体内的养分迁移及养分再利用是一个非常复杂的生理生化过程, 人们对此还缺乏基本的了解.因此, 揭示树木体内养分迁移和再利用的调控机制是生理生态学亟待解决的重要问题. ...
Thinning burning and thin-burn fuel treatment effects on soil properties in a Sierra Nevada mixed conifer forests
1
2007
... 植物组织的养分迁移是植物对养分贫瘠环境的一种适应机制, 也是植物保存养分并维持体内养分平衡的一种重要的养分利用策略(
Kobe et al ., 2005 ;
Milla et al ., 2005 ;
Huang & Wang, 2007 ;
Hosseini et al ., 2008 ).然而, 植物组织迁移养分的机制是什么? 这些迁移的养分是如何被活化重新用于物质生产的? 对于植物组织迁移养分的机制, 有研究提出, 土壤贫瘠生境中植物养分再吸收效率高于土壤肥沃生境中的植物, 土壤贫瘠的胁迫作用可能是植物组织养分迁移的机理(
Moghaddas & Stephens, 2007 ).也有研究指出, 植物组织的养分再吸收与土壤养分供给性相关不大, 贫瘠土壤并不一定是植物体养分迁移的驱动力(
Del Arco et al ., 1991 ;
Eckstein & Karlsson, 1997 ;
Aerts & Chapin, 1999 ;
王希华等, 2004 ).一些研究者用植物生理中“源-汇”理论分析植物组织内养分迁移机制, 认为树木器官在某些生理条件下是养分的“源”, 在另一些生理条件下可能是“汇”.正这种“源-汇”作用调控着树木体内的养分迁移(
Marschner et al ., 1997 ).如本研究中杉木快速增长阶段, 由于树木要壮大自己的个体, 因而将部分从土壤吸收的养分输送到“仍存活物质”内.这时杉木各器官成了养分的“汇”.当林分郁闭后, 生产单位生物量所需养分量下降(
Miller, 1986 ), 积累在“仍存活物质”内的养分被迁移出来用于新的物质生产, 这时杉木各器官成了养分的“源”.也有****认为, 树木体内养分迁移的机理是植物本身具有这种自我调控的反馈机制, 这种调控机制是植物在自然界长期的生存竞争中形成的(
Killingbeck, 1996 ;
Harrington et al ., 2001 ).可见, 对于植物组织养分迁移的机制还没有一个被广泛认可的解析.目前, 迁移的养分如何被活化且重新用于物质生产的机理研究, 更是一个非常薄弱的环节.树木体内的养分迁移及养分再利用是一个非常复杂的生理生化过程, 人们对此还缺乏基本的了解.因此, 揭示树木体内养分迁移和再利用的调控机制是生理生态学亟待解决的重要问题. ...
Nutrient use efficiency in evergreen and deciduous species from heath lands
1
1990
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
The mineral nutrition of wild plants revisited: A reevaluation of processes and patterns
1
1999
... 植物组织的养分迁移是植物对养分贫瘠环境的一种适应机制, 也是植物保存养分并维持体内养分平衡的一种重要的养分利用策略(
Kobe et al ., 2005 ;
Milla et al ., 2005 ;
Huang & Wang, 2007 ;
Hosseini et al ., 2008 ).然而, 植物组织迁移养分的机制是什么? 这些迁移的养分是如何被活化重新用于物质生产的? 对于植物组织迁移养分的机制, 有研究提出, 土壤贫瘠生境中植物养分再吸收效率高于土壤肥沃生境中的植物, 土壤贫瘠的胁迫作用可能是植物组织养分迁移的机理(
Moghaddas & Stephens, 2007 ).也有研究指出, 植物组织的养分再吸收与土壤养分供给性相关不大, 贫瘠土壤并不一定是植物体养分迁移的驱动力(
Del Arco et al ., 1991 ;
Eckstein & Karlsson, 1997 ;
Aerts & Chapin, 1999 ;
王希华等, 2004 ).一些研究者用植物生理中“源-汇”理论分析植物组织内养分迁移机制, 认为树木器官在某些生理条件下是养分的“源”, 在另一些生理条件下可能是“汇”.正这种“源-汇”作用调控着树木体内的养分迁移(
Marschner et al ., 1997 ).如本研究中杉木快速增长阶段, 由于树木要壮大自己的个体, 因而将部分从土壤吸收的养分输送到“仍存活物质”内.这时杉木各器官成了养分的“汇”.当林分郁闭后, 生产单位生物量所需养分量下降(
Miller, 1986 ), 积累在“仍存活物质”内的养分被迁移出来用于新的物质生产, 这时杉木各器官成了养分的“源”.也有****认为, 树木体内养分迁移的机理是植物本身具有这种自我调控的反馈机制, 这种调控机制是植物在自然界长期的生存竞争中形成的(
Killingbeck, 1996 ;
Harrington et al ., 2001 ).可见, 对于植物组织养分迁移的机制还没有一个被广泛认可的解析.目前, 迁移的养分如何被活化且重新用于物质生产的机理研究, 更是一个非常薄弱的环节.树木体内的养分迁移及养分再利用是一个非常复杂的生理生化过程, 人们对此还缺乏基本的了解.因此, 揭示树木体内养分迁移和再利用的调控机制是生理生态学亟待解决的重要问题. ...
Hanan in a tropical savanna: Variation N, P nutrient resorption patterns of plant functional groups and functional significance
1
2008
... 本文虽然分析了“仍存活物质”中根的迁移养分的再利用, 也探讨了枝叶枯死前的养分迁移.但是, 植物细根在枯死前也存在养分转移, 而且这种养分循环对提高植物养分利用效率意义重大(
Meier et al ., 1985 ;
Ratnam & Sankara, 2008 ).由于很难准确估测细根的枯死量, 本文没有研究细根在枯死前转移的养分, 因此, 本研究得出的杉木枯死组织迁移的养分量要低于它的实际迁移量. ...
Nutrient budgets of marsh plants: Efficiency concepts and relation to availability
1
1984
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
氮素添加对黄土高原典型草原长芒草氮磷重吸收率及C:N:P化学计量特征的影响
1
2011
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
氮素添加对黄土高原典型草原长芒草氮磷重吸收率及C:N:P化学计量特征的影响
1
2011
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
连栽第1和第2代杉木人工林养分循环的比较
1
2011
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
连栽第1和第2代杉木人工林养分循环的比较
1
2011
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
不同年龄橡胶树PR107养分利用效率研究
1
2010
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
不同年龄橡胶树PR107养分利用效率研究
1
2010
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
森林细根生产和周转的影响因素
2012
森林细根生产和周转的影响因素
2012
天童国家森林公园若干树种叶水平上养分利用效率的研究
1
2004
... 植物组织的养分迁移是植物对养分贫瘠环境的一种适应机制, 也是植物保存养分并维持体内养分平衡的一种重要的养分利用策略(
Kobe et al ., 2005 ;
Milla et al ., 2005 ;
Huang & Wang, 2007 ;
Hosseini et al ., 2008 ).然而, 植物组织迁移养分的机制是什么? 这些迁移的养分是如何被活化重新用于物质生产的? 对于植物组织迁移养分的机制, 有研究提出, 土壤贫瘠生境中植物养分再吸收效率高于土壤肥沃生境中的植物, 土壤贫瘠的胁迫作用可能是植物组织养分迁移的机理(
Moghaddas & Stephens, 2007 ).也有研究指出, 植物组织的养分再吸收与土壤养分供给性相关不大, 贫瘠土壤并不一定是植物体养分迁移的驱动力(
Del Arco et al ., 1991 ;
Eckstein & Karlsson, 1997 ;
Aerts & Chapin, 1999 ;
王希华等, 2004 ).一些研究者用植物生理中“源-汇”理论分析植物组织内养分迁移机制, 认为树木器官在某些生理条件下是养分的“源”, 在另一些生理条件下可能是“汇”.正这种“源-汇”作用调控着树木体内的养分迁移(
Marschner et al ., 1997 ).如本研究中杉木快速增长阶段, 由于树木要壮大自己的个体, 因而将部分从土壤吸收的养分输送到“仍存活物质”内.这时杉木各器官成了养分的“汇”.当林分郁闭后, 生产单位生物量所需养分量下降(
Miller, 1986 ), 积累在“仍存活物质”内的养分被迁移出来用于新的物质生产, 这时杉木各器官成了养分的“源”.也有****认为, 树木体内养分迁移的机理是植物本身具有这种自我调控的反馈机制, 这种调控机制是植物在自然界长期的生存竞争中形成的(
Killingbeck, 1996 ;
Harrington et al ., 2001 ).可见, 对于植物组织养分迁移的机制还没有一个被广泛认可的解析.目前, 迁移的养分如何被活化且重新用于物质生产的机理研究, 更是一个非常薄弱的环节.树木体内的养分迁移及养分再利用是一个非常复杂的生理生化过程, 人们对此还缺乏基本的了解.因此, 揭示树木体内养分迁移和再利用的调控机制是生理生态学亟待解决的重要问题. ...
Intrinsic and extrinsic controls of fine root life span
2
2013
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
... 会同杉木林生长12年时, 才发生某林龄段“仍存活物质”迁移出养分用于物质生产.这主要由杉木不同生长阶段对养分的不同需求引起的.杉木从幼树到快速生长阶段为了壮大自己的个体, 为树木未来生长打好基础, 已生产的物质(器官)中需要不断补充养分.因此, 这个生育阶段的“仍存活物质”中不仅没有养分转移出来再利用, 而且还需要新的养分补充.如本研究中8-11年生的林分吸收的土壤养分中, 每年还需输送7.83 kg·hm
-2 到该林龄段“仍存活物质”内(
表2 ).此外,
Miller (1986) 的研究表明, 林分郁闭后, 多年生乔木林生产单位物质所需的养分量减少.树木体内养分再分配机制控制着将积累在“仍存活物质”内的养分迁移出来重新用于物质生产.从本研究结果看, 林分生长12年后(杉木林已郁闭), 虽然从“仍存活物质”内迁移出的养分并不很多, 但“仍存活物质”迁移出的养分生产的物质量仍占生产物质总量的3.11%-3.40%.这是因为, 树木体内需要一定的养分量才能正常生长, 只有超过生长需要的那部分积累的养分才能迁移出来再利用(
Miller, 1986 ;
Marschner et al ., 1997 ;
Chen & Brassan, 2013 ).从本研究还可看出, 会同杉木林郁闭后, “仍存活物质”有一个较为稳定的速率迁移出养分重新用于物质生产. ...
杉木人工林养分循环随林龄变化的特征
2
2018
... 单位面积林分某林龄段生产的树干、树皮生物量, 根据生长过程中树干、树皮凋落甚少的规律, 用某林龄段开始和结束时单位面积林分测定的这些器官现存量之差求得.根系存在更新代谢过程, 但要准确测定根系(尤其是细根)的生长周期, 需采用微根管技术或同位素追踪技术.这些技术方法成本高且操作有一定的难度, 因而大多数研究者在估算某林龄段根生产量时, 将根与干材和树皮同等对待(
梅莉等, 2004 ;
王端丽等, 2012 ).本研究也是如此(这样估算出的根生产量要低于它的实际生产量).杉木是常绿树种, 枝、叶都有一定的生长周期, 根据
陈日升等(2018) 的研究方法, 估算出某林龄段这些还存活的枝叶量.然后用下式估算某林龄段单位面积林分生产的生物量(
W ). ...
... 单位面积林分不同林龄段土壤养分吸收量根据
陈日升等(2018) 的方法估算.该方法考虑了枝叶枯死前迁移的养分, 以及“仍存活物质”迁移养分的再利用.在估算出单位面积林分某林龄段生产的生物量(公式(3))和某林龄段“仍存活物质”迁移出(或又吸收)养分量(公式(4)和公式(5)), 以及单位面积林分枝叶枯死前养分转移量(公式(6))后, 如果某林龄段结束时杉木各器官养分浓度低于开始时, 则某林龄段单位面积林分“仍存活物质”有部分养分迁移出来, 可用于林分物质生产, 从而减少了土壤养分的吸收量(
M 4 ).其土壤养分吸收量用下面公式求得: ...
天童国家森林公园若干树种叶水平上养分利用效率的研究
1
2004
... 植物组织的养分迁移是植物对养分贫瘠环境的一种适应机制, 也是植物保存养分并维持体内养分平衡的一种重要的养分利用策略(
Kobe et al ., 2005 ;
Milla et al ., 2005 ;
Huang & Wang, 2007 ;
Hosseini et al ., 2008 ).然而, 植物组织迁移养分的机制是什么? 这些迁移的养分是如何被活化重新用于物质生产的? 对于植物组织迁移养分的机制, 有研究提出, 土壤贫瘠生境中植物养分再吸收效率高于土壤肥沃生境中的植物, 土壤贫瘠的胁迫作用可能是植物组织养分迁移的机理(
Moghaddas & Stephens, 2007 ).也有研究指出, 植物组织的养分再吸收与土壤养分供给性相关不大, 贫瘠土壤并不一定是植物体养分迁移的驱动力(
Del Arco et al ., 1991 ;
Eckstein & Karlsson, 1997 ;
Aerts & Chapin, 1999 ;
王希华等, 2004 ).一些研究者用植物生理中“源-汇”理论分析植物组织内养分迁移机制, 认为树木器官在某些生理条件下是养分的“源”, 在另一些生理条件下可能是“汇”.正这种“源-汇”作用调控着树木体内的养分迁移(
Marschner et al ., 1997 ).如本研究中杉木快速增长阶段, 由于树木要壮大自己的个体, 因而将部分从土壤吸收的养分输送到“仍存活物质”内.这时杉木各器官成了养分的“汇”.当林分郁闭后, 生产单位生物量所需养分量下降(
Miller, 1986 ), 积累在“仍存活物质”内的养分被迁移出来用于新的物质生产, 这时杉木各器官成了养分的“源”.也有****认为, 树木体内养分迁移的机理是植物本身具有这种自我调控的反馈机制, 这种调控机制是植物在自然界长期的生存竞争中形成的(
Killingbeck, 1996 ;
Harrington et al ., 2001 ).可见, 对于植物组织养分迁移的机制还没有一个被广泛认可的解析.目前, 迁移的养分如何被活化且重新用于物质生产的机理研究, 更是一个非常薄弱的环节.树木体内的养分迁移及养分再利用是一个非常复杂的生理生化过程, 人们对此还缺乏基本的了解.因此, 揭示树木体内养分迁移和再利用的调控机制是生理生态学亟待解决的重要问题. ...
Redistribution of mineral elements during development
1
1955
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
杉木人工林养分循环随林龄变化的特征
2
2018
... 单位面积林分某林龄段生产的树干、树皮生物量, 根据生长过程中树干、树皮凋落甚少的规律, 用某林龄段开始和结束时单位面积林分测定的这些器官现存量之差求得.根系存在更新代谢过程, 但要准确测定根系(尤其是细根)的生长周期, 需采用微根管技术或同位素追踪技术.这些技术方法成本高且操作有一定的难度, 因而大多数研究者在估算某林龄段根生产量时, 将根与干材和树皮同等对待(
梅莉等, 2004 ;
王端丽等, 2012 ).本研究也是如此(这样估算出的根生产量要低于它的实际生产量).杉木是常绿树种, 枝、叶都有一定的生长周期, 根据
陈日升等(2018) 的研究方法, 估算出某林龄段这些还存活的枝叶量.然后用下式估算某林龄段单位面积林分生产的生物量(
W ). ...
... 单位面积林分不同林龄段土壤养分吸收量根据
陈日升等(2018) 的方法估算.该方法考虑了枝叶枯死前迁移的养分, 以及“仍存活物质”迁移养分的再利用.在估算出单位面积林分某林龄段生产的生物量(公式(3))和某林龄段“仍存活物质”迁移出(或又吸收)养分量(公式(4)和公式(5)), 以及单位面积林分枝叶枯死前养分转移量(公式(6))后, 如果某林龄段结束时杉木各器官养分浓度低于开始时, 则某林龄段单位面积林分“仍存活物质”有部分养分迁移出来, 可用于林分物质生产, 从而减少了土壤养分的吸收量(
M 4 ).其土壤养分吸收量用下面公式求得: ...
Effects of site characteristics on nitrogen retrains location from senescing leaves
1
1991
... 植物组织的养分迁移是植物对养分贫瘠环境的一种适应机制, 也是植物保存养分并维持体内养分平衡的一种重要的养分利用策略(
Kobe et al ., 2005 ;
Milla et al ., 2005 ;
Huang & Wang, 2007 ;
Hosseini et al ., 2008 ).然而, 植物组织迁移养分的机制是什么? 这些迁移的养分是如何被活化重新用于物质生产的? 对于植物组织迁移养分的机制, 有研究提出, 土壤贫瘠生境中植物养分再吸收效率高于土壤肥沃生境中的植物, 土壤贫瘠的胁迫作用可能是植物组织养分迁移的机理(
Moghaddas & Stephens, 2007 ).也有研究指出, 植物组织的养分再吸收与土壤养分供给性相关不大, 贫瘠土壤并不一定是植物体养分迁移的驱动力(
Del Arco et al ., 1991 ;
Eckstein & Karlsson, 1997 ;
Aerts & Chapin, 1999 ;
王希华等, 2004 ).一些研究者用植物生理中“源-汇”理论分析植物组织内养分迁移机制, 认为树木器官在某些生理条件下是养分的“源”, 在另一些生理条件下可能是“汇”.正这种“源-汇”作用调控着树木体内的养分迁移(
Marschner et al ., 1997 ).如本研究中杉木快速增长阶段, 由于树木要壮大自己的个体, 因而将部分从土壤吸收的养分输送到“仍存活物质”内.这时杉木各器官成了养分的“汇”.当林分郁闭后, 生产单位生物量所需养分量下降(
Miller, 1986 ), 积累在“仍存活物质”内的养分被迁移出来用于新的物质生产, 这时杉木各器官成了养分的“源”.也有****认为, 树木体内养分迁移的机理是植物本身具有这种自我调控的反馈机制, 这种调控机制是植物在自然界长期的生存竞争中形成的(
Killingbeck, 1996 ;
Harrington et al ., 2001 ).可见, 对于植物组织养分迁移的机制还没有一个被广泛认可的解析.目前, 迁移的养分如何被活化且重新用于物质生产的机理研究, 更是一个非常薄弱的环节.树木体内的养分迁移及养分再利用是一个非常复杂的生理生化过程, 人们对此还缺乏基本的了解.因此, 揭示树木体内养分迁移和再利用的调控机制是生理生态学亟待解决的重要问题. ...
植物养分高效利用机制研究进展
1
2009
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
植物养分高效利用机制研究进展
1
2009
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
不同林龄新银合欢重吸收率及其C:N:P化学计量特征
1
2015
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
不同林龄新银合欢重吸收率及其C:N:P化学计量特征
1
2015
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
不同年龄阶段马尾松人工林养分循环的研究
1
2002
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
不同年龄阶段马尾松人工林养分循环的研究
1
2002
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
2
1994
... 在林龄为7、11、16、20、25年时(与第1代林测定生物量的林龄相同), 在4块观测样地测定林分生物量.生物量测定采用克拉夫特的林木生长状况分级法(
林业部科技司, 1994 ), 把样地内所有林木分成优势木(I级木)、亚优势木(II级木)、中级木(III级木)、被压木(IV级木)、濒死木(V级木) 5个生长等级, 测定样地内每株树木的树高、胸径和冠幅.根据样地所有树木平均树高和胸径选取样地标准木1株, 根据各生长级树木的平均树高和胸径选取各生长级标准木1株(样地中总计6株样木), 将它们连根挖出.在现场将树干按2 m分区段, 测定各区段树干、皮、枝和叶的生物量鲜质量, 并测定根的鲜质量.选取杉木各器官样品, 称量后放在80 ℃的烘箱内烘干至恒质量.求出样品的含水率, 将各器官鲜质量换算成干质量.根据相对生长法则建立生物量估算模型, 再用经过检验的模型估算单位面积林分生物量. ...
... 取适量待测的样品置85 ℃恒温干燥箱内烘干至恒质量, 称质量并磨碎, 过60目筛, 按《森林生态系统研究方法》(
林业部科技司, 1994 )中的养分测定方法配置待测溶液.全钾(TK)、全钙(TCa)、全镁(TMg)含量用原子吸收分光光度计测定, 全磷(TP)含量用分光光度计测定, 全氮(TN)含量用半微量凯氏法测定.每个样品重复测定3次, 用3次重复测定的平均值作为计量标准. ...
2
1994
... 在林龄为7、11、16、20、25年时(与第1代林测定生物量的林龄相同), 在4块观测样地测定林分生物量.生物量测定采用克拉夫特的林木生长状况分级法(
林业部科技司, 1994 ), 把样地内所有林木分成优势木(I级木)、亚优势木(II级木)、中级木(III级木)、被压木(IV级木)、濒死木(V级木) 5个生长等级, 测定样地内每株树木的树高、胸径和冠幅.根据样地所有树木平均树高和胸径选取样地标准木1株, 根据各生长级树木的平均树高和胸径选取各生长级标准木1株(样地中总计6株样木), 将它们连根挖出.在现场将树干按2 m分区段, 测定各区段树干、皮、枝和叶的生物量鲜质量, 并测定根的鲜质量.选取杉木各器官样品, 称量后放在80 ℃的烘箱内烘干至恒质量.求出样品的含水率, 将各器官鲜质量换算成干质量.根据相对生长法则建立生物量估算模型, 再用经过检验的模型估算单位面积林分生物量. ...
... 取适量待测的样品置85 ℃恒温干燥箱内烘干至恒质量, 称质量并磨碎, 过60目筛, 按《森林生态系统研究方法》(
林业部科技司, 1994 )中的养分测定方法配置待测溶液.全钾(TK)、全钙(TCa)、全镁(TMg)含量用原子吸收分光光度计测定, 全磷(TP)含量用分光光度计测定, 全氮(TN)含量用半微量凯氏法测定.每个样品重复测定3次, 用3次重复测定的平均值作为计量标准. ...
林分密度对湿地松人工林养分循环速率和利用效率的影响
1
2013
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
林分密度对湿地松人工林养分循环速率和利用效率的影响
1
2013
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
Above-ground growth and nitrogen use by plant in a subarctic environment: Effects of habitat life-form and species
1
1977
... 植物组织的养分迁移是植物对养分贫瘠环境的一种适应机制, 也是植物保存养分并维持体内养分平衡的一种重要的养分利用策略(
Kobe et al ., 2005 ;
Milla et al ., 2005 ;
Huang & Wang, 2007 ;
Hosseini et al ., 2008 ).然而, 植物组织迁移养分的机制是什么? 这些迁移的养分是如何被活化重新用于物质生产的? 对于植物组织迁移养分的机制, 有研究提出, 土壤贫瘠生境中植物养分再吸收效率高于土壤肥沃生境中的植物, 土壤贫瘠的胁迫作用可能是植物组织养分迁移的机理(
Moghaddas & Stephens, 2007 ).也有研究指出, 植物组织的养分再吸收与土壤养分供给性相关不大, 贫瘠土壤并不一定是植物体养分迁移的驱动力(
Del Arco et al ., 1991 ;
Eckstein & Karlsson, 1997 ;
Aerts & Chapin, 1999 ;
王希华等, 2004 ).一些研究者用植物生理中“源-汇”理论分析植物组织内养分迁移机制, 认为树木器官在某些生理条件下是养分的“源”, 在另一些生理条件下可能是“汇”.正这种“源-汇”作用调控着树木体内的养分迁移(
Marschner et al ., 1997 ).如本研究中杉木快速增长阶段, 由于树木要壮大自己的个体, 因而将部分从土壤吸收的养分输送到“仍存活物质”内.这时杉木各器官成了养分的“汇”.当林分郁闭后, 生产单位生物量所需养分量下降(
Miller, 1986 ), 积累在“仍存活物质”内的养分被迁移出来用于新的物质生产, 这时杉木各器官成了养分的“源”.也有****认为, 树木体内养分迁移的机理是植物本身具有这种自我调控的反馈机制, 这种调控机制是植物在自然界长期的生存竞争中形成的(
Killingbeck, 1996 ;
Harrington et al ., 2001 ).可见, 对于植物组织养分迁移的机制还没有一个被广泛认可的解析.目前, 迁移的养分如何被活化且重新用于物质生产的机理研究, 更是一个非常薄弱的环节.树木体内的养分迁移及养分再利用是一个非常复杂的生理生化过程, 人们对此还缺乏基本的了解.因此, 揭示树木体内养分迁移和再利用的调控机制是生理生态学亟待解决的重要问题. ...
Production and resource use efficiencies in N- and P-limited tropical forests: A comparison of responses to long-term fertilization
1
2001
... 植物组织的养分迁移是植物对养分贫瘠环境的一种适应机制, 也是植物保存养分并维持体内养分平衡的一种重要的养分利用策略(
Kobe et al ., 2005 ;
Milla et al ., 2005 ;
Huang & Wang, 2007 ;
Hosseini et al ., 2008 ).然而, 植物组织迁移养分的机制是什么? 这些迁移的养分是如何被活化重新用于物质生产的? 对于植物组织迁移养分的机制, 有研究提出, 土壤贫瘠生境中植物养分再吸收效率高于土壤肥沃生境中的植物, 土壤贫瘠的胁迫作用可能是植物组织养分迁移的机理(
Moghaddas & Stephens, 2007 ).也有研究指出, 植物组织的养分再吸收与土壤养分供给性相关不大, 贫瘠土壤并不一定是植物体养分迁移的驱动力(
Del Arco et al ., 1991 ;
Eckstein & Karlsson, 1997 ;
Aerts & Chapin, 1999 ;
王希华等, 2004 ).一些研究者用植物生理中“源-汇”理论分析植物组织内养分迁移机制, 认为树木器官在某些生理条件下是养分的“源”, 在另一些生理条件下可能是“汇”.正这种“源-汇”作用调控着树木体内的养分迁移(
Marschner et al ., 1997 ).如本研究中杉木快速增长阶段, 由于树木要壮大自己的个体, 因而将部分从土壤吸收的养分输送到“仍存活物质”内.这时杉木各器官成了养分的“汇”.当林分郁闭后, 生产单位生物量所需养分量下降(
Miller, 1986 ), 积累在“仍存活物质”内的养分被迁移出来用于新的物质生产, 这时杉木各器官成了养分的“源”.也有****认为, 树木体内养分迁移的机理是植物本身具有这种自我调控的反馈机制, 这种调控机制是植物在自然界长期的生存竞争中形成的(
Killingbeck, 1996 ;
Harrington et al ., 2001 ).可见, 对于植物组织养分迁移的机制还没有一个被广泛认可的解析.目前, 迁移的养分如何被活化且重新用于物质生产的机理研究, 更是一个非常薄弱的环节.树木体内的养分迁移及养分再利用是一个非常复杂的生理生化过程, 人们对此还缺乏基本的了解.因此, 揭示树木体内养分迁移和再利用的调控机制是生理生态学亟待解决的重要问题. ...
Age-related changes in nutrient resorption patterns and tannin concentration of
Casuarina equisetifolia plantations
1
2012
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
不同林龄樟子松叶片养分含量及其再吸收效率
1
2005
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
不同年龄阶段马占相思(
Acacia mangium )人工林营养元素的生物循环
1
2007
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
不同年龄阶段马占相思(
Acacia mangium )人工林营养元素的生物循环
1
2007
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
不同林龄樟子松叶片养分含量及其再吸收效率
1
2005
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
红树植物白骨壤叶片衰老过程的氮磷内吸收变化研究
1
2008
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
Comparison of growth, nutrition and soil properties of pure stands of
Quercus castaneifolia and mixed with
Zelkova carpinifolia in the Hyrcanian forests of Iran
1
2008
... 植物组织的养分迁移是植物对养分贫瘠环境的一种适应机制, 也是植物保存养分并维持体内养分平衡的一种重要的养分利用策略(
Kobe et al ., 2005 ;
Milla et al ., 2005 ;
Huang & Wang, 2007 ;
Hosseini et al ., 2008 ).然而, 植物组织迁移养分的机制是什么? 这些迁移的养分是如何被活化重新用于物质生产的? 对于植物组织迁移养分的机制, 有研究提出, 土壤贫瘠生境中植物养分再吸收效率高于土壤肥沃生境中的植物, 土壤贫瘠的胁迫作用可能是植物组织养分迁移的机理(
Moghaddas & Stephens, 2007 ).也有研究指出, 植物组织的养分再吸收与土壤养分供给性相关不大, 贫瘠土壤并不一定是植物体养分迁移的驱动力(
Del Arco et al ., 1991 ;
Eckstein & Karlsson, 1997 ;
Aerts & Chapin, 1999 ;
王希华等, 2004 ).一些研究者用植物生理中“源-汇”理论分析植物组织内养分迁移机制, 认为树木器官在某些生理条件下是养分的“源”, 在另一些生理条件下可能是“汇”.正这种“源-汇”作用调控着树木体内的养分迁移(
Marschner et al ., 1997 ).如本研究中杉木快速增长阶段, 由于树木要壮大自己的个体, 因而将部分从土壤吸收的养分输送到“仍存活物质”内.这时杉木各器官成了养分的“汇”.当林分郁闭后, 生产单位生物量所需养分量下降(
Miller, 1986 ), 积累在“仍存活物质”内的养分被迁移出来用于新的物质生产, 这时杉木各器官成了养分的“源”.也有****认为, 树木体内养分迁移的机理是植物本身具有这种自我调控的反馈机制, 这种调控机制是植物在自然界长期的生存竞争中形成的(
Killingbeck, 1996 ;
Harrington et al ., 2001 ).可见, 对于植物组织养分迁移的机制还没有一个被广泛认可的解析.目前, 迁移的养分如何被活化且重新用于物质生产的机理研究, 更是一个非常薄弱的环节.树木体内的养分迁移及养分再利用是一个非常复杂的生理生化过程, 人们对此还缺乏基本的了解.因此, 揭示树木体内养分迁移和再利用的调控机制是生理生态学亟待解决的重要问题. ...
Leaf nutrient concentration, nutrient resorption and litter decomposition in an evergreen broad-leaved forest in eastern China
1
2007
... 植物组织的养分迁移是植物对养分贫瘠环境的一种适应机制, 也是植物保存养分并维持体内养分平衡的一种重要的养分利用策略(
Kobe et al ., 2005 ;
Milla et al ., 2005 ;
Huang & Wang, 2007 ;
Hosseini et al ., 2008 ).然而, 植物组织迁移养分的机制是什么? 这些迁移的养分是如何被活化重新用于物质生产的? 对于植物组织迁移养分的机制, 有研究提出, 土壤贫瘠生境中植物养分再吸收效率高于土壤肥沃生境中的植物, 土壤贫瘠的胁迫作用可能是植物组织养分迁移的机理(
Moghaddas & Stephens, 2007 ).也有研究指出, 植物组织的养分再吸收与土壤养分供给性相关不大, 贫瘠土壤并不一定是植物体养分迁移的驱动力(
Del Arco et al ., 1991 ;
Eckstein & Karlsson, 1997 ;
Aerts & Chapin, 1999 ;
王希华等, 2004 ).一些研究者用植物生理中“源-汇”理论分析植物组织内养分迁移机制, 认为树木器官在某些生理条件下是养分的“源”, 在另一些生理条件下可能是“汇”.正这种“源-汇”作用调控着树木体内的养分迁移(
Marschner et al ., 1997 ).如本研究中杉木快速增长阶段, 由于树木要壮大自己的个体, 因而将部分从土壤吸收的养分输送到“仍存活物质”内.这时杉木各器官成了养分的“汇”.当林分郁闭后, 生产单位生物量所需养分量下降(
Miller, 1986 ), 积累在“仍存活物质”内的养分被迁移出来用于新的物质生产, 这时杉木各器官成了养分的“源”.也有****认为, 树木体内养分迁移的机理是植物本身具有这种自我调控的反馈机制, 这种调控机制是植物在自然界长期的生存竞争中形成的(
Killingbeck, 1996 ;
Harrington et al ., 2001 ).可见, 对于植物组织养分迁移的机制还没有一个被广泛认可的解析.目前, 迁移的养分如何被活化且重新用于物质生产的机理研究, 更是一个非常薄弱的环节.树木体内的养分迁移及养分再利用是一个非常复杂的生理生化过程, 人们对此还缺乏基本的了解.因此, 揭示树木体内养分迁移和再利用的调控机制是生理生态学亟待解决的重要问题. ...
红树植物白骨壤叶片衰老过程的氮磷内吸收变化研究
1
2008
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
不同林分类型叶片氮磷含量, 氮磷比及其内吸收率
1
2009
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
不同林分类型叶片氮磷含量, 氮磷比及其内吸收率
1
2009
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
氮添加对兴安落叶松养分分配和再吸收效率的影响
1
2010
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
氮添加对兴安落叶松养分分配和再吸收效率的影响
1
2010
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
武夷山不同海拔梯度黄山松叶片养分含量及其再吸收效率
1
2017
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
武夷山不同海拔梯度黄山松叶片养分含量及其再吸收效率
1
2017
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
不同发育阶段杉木人工林养分内循环与周转利用效率的研究
1
2014
... 会同杉木枝叶枯死前N、P、K迁移率大于Ca、Mg.各养分元素迁移率与福建杉木(
周丽丽, 2014 )相比, 除叶的Mg相近外, 其余枝叶中各元素转移率均低于福建杉木.与林宝平等(2017)研究的杉木相比, P、Ca转移率偏高, N、K、Mg则偏低.与其他研究结果存在差别可能是如下原因造成的: (1)研究林分的立地环境条件不相同, 同一树种在不同的立地环境条件下可能造成枝叶枯死前后养分浓度的差异, 进而导致枝叶枯死前养分迁移率的差异.(2)研究方法不同, 本研究是在同一林分内连续定位测定不同林龄时的数据, 其他同类研究大都用“空间换时间”法, 空间的异质性可能导致其研究结果偏移.(3)本研究把1-25年生的林分分成几个连续的林龄段, 依据每林龄段前后测定数据分段逐一分析, 同类研究则是在林分某一林龄时测定数据, 把林分1年生到这个林龄时作为一个时间序列处理, 所取的时间尺度不一样, 导致研究结果也不一样. ...
不同发育阶段杉木人工林养分内循环与周转利用效率的研究
1
2014
... 会同杉木枝叶枯死前N、P、K迁移率大于Ca、Mg.各养分元素迁移率与福建杉木(
周丽丽, 2014 )相比, 除叶的Mg相近外, 其余枝叶中各元素转移率均低于福建杉木.与林宝平等(2017)研究的杉木相比, P、Ca转移率偏高, N、K、Mg则偏低.与其他研究结果存在差别可能是如下原因造成的: (1)研究林分的立地环境条件不相同, 同一树种在不同的立地环境条件下可能造成枝叶枯死前后养分浓度的差异, 进而导致枝叶枯死前养分迁移率的差异.(2)研究方法不同, 本研究是在同一林分内连续定位测定不同林龄时的数据, 其他同类研究大都用“空间换时间”法, 空间的异质性可能导致其研究结果偏移.(3)本研究把1-25年生的林分分成几个连续的林龄段, 依据每林龄段前后测定数据分段逐一分析, 同类研究则是在林分某一林龄时测定数据, 把林分1年生到这个林龄时作为一个时间序列处理, 所取的时间尺度不一样, 导致研究结果也不一样. ...
北方山区主要森林类型树木叶片氮、磷回收效率研究
1
2017
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
The terminological jungle revisited: Making a case for use of the term resorption
1
1986
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
北方山区主要森林类型树木叶片氮、磷回收效率研究
1
2017
... 林分的养分主要被用于物质生产(
吴鹏飞和马祥庆, 2009 ;
李钦禄等, 2015 ), 因而养分利用成为森林生态系统生产力和养分循环研究的重要环节.20世纪80年代以来, 我国****将林分养分动态与生物量相结合开展了杉木(
Cunninghamia lanceolata )(
刘爱琴等, 2005 ;
田大伦等, 2011 )、湿地松(
Pinus elliottii )(
肖兴翠等, 2013 )、马占相思(
Acacia mangium )(
何斌等, 2007 )、尾叶桉(
Eucalyptus grandis × E. urophylla )(
林德喜等, 2002 )、马尾松(
Pinus massoniana )(
项文化和田大伦, 2002 )、橡胶树(
Hevea brasiliensis )(
曹建华等, 2010 )等人工林的养分利用研究, 这些研究成果具有重要的理论和实践意义.以往研究林分生产物质利用的养分都用“吸收=存留+归还”方法求算, 这种估算忽略了植物组织迁移养分的再利用(
Killingbeck, 1986 ;
Aerts, 1990 ;
Chen & Brassard, 2013 ).植物组织迁移的养分在新的物质生产中再利用, 不仅提高了养分的利用效率, 促进了林分养分的循环(
Mayor et al ., 2013 ), 而且减少了植物对土壤养分的吸收, 维护了林地地力, 降低了植物对环境的依赖性(
Shaver & Melillo, 1984 ;
Lim & Cousens, 1986 ), 对森林生态系统的养分循环、植物种群和群落稳定性都具有重要的生态学意义.然而目前对植物组织内的养分迁移与利用研究, 几乎全部集中在不同生活型植物(
曾琦等, 2008 ;
张立华等, 2009 ;
宗宁等, 2017 ), 不同林龄植物(
曾德慧等, 2005 ;
Ye, 2012 ;
邓浩俊等, 2015 ;
林宝平等, 2017 )衰老组织养分转移率的差异; 以及海拔(
郑媛等, 2017 )、土壤肥力(
赵琼等, 2010 ;
安卓等, 2011 )对植物组织衰老时养分转移的影响上, 很少探讨植物衰老组织转移的养分在林分新的物质生产中的再利用.虽然在20世纪50年代已发现草本植物活组织内养分的循环和再分配现象(
Williams, 1955 ), 发现植物活组织迁移出的养分可活化用以补充(至少部分地补充)物质生产中所需养分(
Miller, 1984 ,
1986 ), 但由于研究方法(同位素追踪技术)成本高且不安全, 因此, 鲜见对乔木活组织的养分迁移与再利用的报道.如果仅用从土壤中吸收的养分, 忽略植物组织迁移养分的再利用, 就不能真实地反映植物养分利用特征, 也不能完全揭示植物养分利用的机理.本研究利用湖南会同杉木林生态系统国家野外科学观测研究站连续25年定位测定的生物量和林木养分含量数据, 定量估算了不同林龄段杉木枝叶枯死前向活体组织转移的养分量, 并估算了“那些在某林龄段开始前林分生产的, 在这林龄段结束时仍存活的器官(干、皮、枝、叶、根)” (“ ”内文字后文中用“仍存活物质”表示)中, 在这林龄段内的养分迁移量.将这些养分与从土壤吸收的养分结合一起, 综合分析林分物质生产的养分利用特征, 旨在揭示树木组织的迁移养分再利用策略, 为人工林的经营管理提供科学依据. ...
 ,1,2,3,*, 黄志宏1,2,3, 赵仲辉1,2,3, 邓湘雯1,2,31.
,1,2,3,*, 黄志宏1,2,3, 赵仲辉1,2,3, 邓湘雯1,2,31. 
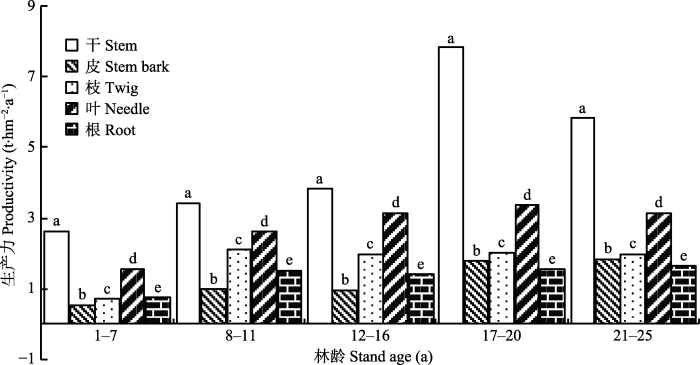 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}