0 引言
【研究意义】细菌性溃疡病(bacterial canker disease,BCD)是严重危害柑橘的一种病害,在世界范围内对柑橘产业造成了大的损失,被列入检疫性病害,其病原为柑橘黄单胞菌柑橘亚种(Xanthomonas citri subsp. citri,Xcc)[1]。Xcc可侵染柑橘叶片、皮刺、枝条及果实,造成落叶、梢枯、削弱树势等危害,严重时还引起落果。由于细菌性溃疡病对于柑橘树体正常生长和果实产量及品质均有重要的影响,培育抗溃疡病的柑橘品种或采取有效的防病措施对于柑橘产业持续、健康发展具有重要意义。【前人研究进展】目前,从寄主角度针对柑橘溃疡病开展的工作主要包括以下几个方面:分析不同柑橘品种或资源对Xcc的抗性差异[2]、克隆和鉴定抗病基因[3,4,5]、解析柑橘对Xcc应答的生理或分子机制[6]、柑橘遗传转化和转基因植株抗性评价[7,8,9]、外源药剂(如水杨酸)防控强溃疡病等[10],对揭示柑橘抗病性差异的生理和分子机制、发掘和创制抗性资源等起到了重要的作用。柑橘基因型是影响溃疡病发生程度的一个重要因素,大多数柑橘栽培品种均易发生溃疡病,而金柑和枸橼C-05(Citrus medica)则较少发病,后者更是被认为对柑橘溃疡病具有免疫抗性[2,6]。对抗病和感病的柑橘资源进行分析,发现叶片结构、气孔大小与密度、叶片分泌物质、抗氧化酶活性及基因表达谱等与溃疡病抗性强弱有关[2,6,10]。如有报道表明气孔小可能是金柑抗病的一个的结构机制,能够限制进入叶片的细菌数量[6];枸橼C-05叶片分泌特殊的物质抑制溃疡病菌生长,使其表现出较好的抗性[2]。植物在遭受病原入侵时,会主动合成一些物质去增强抗病性。研究表明,在抗病中起作用的次生代谢物较多,多胺是其中一种。多胺是广泛存在于活体生物中的一类低分子量含氮脂肪碱,具有多聚阳离子特性,在生理pH下带正电荷,能够共价结合带负电荷的大分子物质(如DNA、RNA、染色质和蛋白质等),因而参与了许多植物生理和生物学过程[11,12]。高等植物中常见的多胺为腐胺(putrescine,Put)、亚精胺(spermidine,Spd)和精胺(spermine,Spm)。此外,多胺还被认为具有清除活性氧(reactive oxygen species,ROS)、调节渗透压的作用[11]。基于多胺这些特性,大量研究认为,多胺是植物应答非生物胁迫的一类重要代谢物[12]。研究还表明,多胺在植物应答生物胁迫中也发挥着重要的作用[13,14],这主要是基于以下研究结果:(1)植物在病原菌入侵时,多胺合成酶活性上升,使体内积累多胺[15]。如大麦在接种白粉病菌(Blumeria graminis)后,鸟胺酸脱羧酶活性显著增加,叶片中自由态Spd、共轭态Spd和Put含量升高[16]。对烟草接种活体营养型细菌后发现,植株体内Spm显著增加[17]。这些结果说明多胺参与寄主植物的病原应答过程;(2)外源施用多胺能增强植物的抗病能力,如外源添加Spm增强了烟草对烟草花叶病毒(Tobacco mosaic virus,TMV)的抗性[18];(3)超表达多胺合成基因,使转基因植株中的多胺含量增加,其抗病性也得以增强[9]。【本研究切入点】无论是开展抗病育种还是研发溃疡病防控技术,理论上均需要对柑橘应答Xcc的侵染进行深入地研究。柑橘溃疡病在高温环境下容易发生,但这一现象的机制却知之甚少。此外,利用外源多胺防控溃疡病发生的研究尚未见报道。【拟解决的关键问题】以冰糖橙(Citrus sinensis)为供试材料,比较不同温度下植株接种溃疡病菌后的发病情况、多胺含量及抗病基因的表达,同时分析外源多胺处理能否增强抗病能力。阐明高温下柑橘易发溃疡病的可能机制,同时为研发和应用基于外源物质处理增强抗病性的技术提供理论指导。1 材料与方法
试验于2014年在华中农业大学园艺林学学院和广西特色作物研究院完成。1.1 植物材料和菌株
植物材料为三年生冰糖橙植株,接种用的柑橘溃疡病菌株由华中农业大学植物病理实验室洪霓教授提供。病菌在SPA培养基(蔗糖20 g,蛋白胨5 g,K2HPO4 0.5 g,MgSO4·7H2O 0.25 g,pH 7.2—7.4)中悬浮培养,接种试验在广西特色作物研究院进行。1.2 处理
选取叶龄为30—45 d、部位相同且长势良好的夏稍枝条,田间取回后迅速将枝条下端插入清水中,分为两组,分别放置于21℃和30℃光照培养箱中,72 h后用于接种,在温度处理结束后取叶片用于多胺含量测定和抗病基因表达分析。此外,为研究多胺是否有利于增强溃疡病抗性,将冰糖橙植株分别用清水或0.4 mmol·L-1亚精胺(Spd)溶液每24 h喷施叶面一次,连续处理72 h。Spd处理结束后取叶片测定内源多胺含量(所取叶片先经清水洗净擦干),同时将处理的叶片接种Xcc。为分析抗病基因表达,将Spd喷施叶片3 d和7 d,取处理前和处理后的叶片用于抗病基因表达分析。1.3 Xcc接种
参考WANG等[6]针刺接种法,将需要接种的叶片用清水洗净擦干,在叶脉两侧相同部位用接种针各刺5个伤口,在每个伤口周围分别再刺5个小伤口。随后用移液枪吸取配置好的菌液10 μL(浓度约为108 cfu/mL)打在伤口上。将接种后的叶片套上塑料袋,放置于28℃条件下培养,在接种后不同时间(温度处理材料在接种后第3、7、10天;多胺处理材料在接种后第7、10、14天)统计发病率与病情指数。每个处理组各统计50个病斑,分析软件为ImageJ(NIH)。1.4 多胺提取与测定
参考ZHANG等[19]方法,将叶片在液氮中研磨成粉末,取0.1 g加入1 mL浓度为5%的高氯酸(perchloric acid,PCA),涡旋混匀后冰上放置30 min,4℃、12 000 r/min离心15 min,将上清液转入一个新的离心管。沉淀中加入1 mL PCA,重复前面的步骤,离心后将两次上清液合并(即为多胺粗提液)。取1 mL粗提液至10 mL离心管,加入50 μL己二胺(1 mmol·L-1),加入1 mL的2 mol·L-1 NaOH,涡旋混匀;加入10 μL苯甲酰氯,涡旋20 s,37℃水浴20 min;加入2 mL饱和NaCl和2 mL乙醚,混匀后于8 000 r/min离心5 min;取1 mL乙醚相,真空浓缩干燥25 min;加200—500 μL色谱级甲醇(Fisher,美国)溶解,0.22 μm有机滤膜过滤后取20 μL在安装紫外检测器高效液相色谱系统(HPLC,Agilent 1200)中上样。多胺检测波长为230 nm,采用安捷伦C18反向色谱柱(4.6 mm×150 mm,孔径5 μm)。洗脱液为色谱级甲醇∕水(A相/B相),采用梯度洗脱,流速0.7 mL·min-1,柱温25℃。梯度洗脱程序见表1[20]。Table 1
表1
表1自由态多胺分离所用HPLC洗脱程序
Table 1HPLC elution procedures for separation of free polyamines
| 时间 Time (min) | A相 A phase (%) | B相 B phase (%) |
|---|---|---|
| 0 | 45 | 55 |
| 15 | 95 | 5 |
| 17 | 100 | 0 |
| 18 | 45 | 55 |
| 25 | 45 | 55 |
新窗口打开
1.5 RNA提取及RT-PCR
利用RNAiso Plus RNA试剂盒(TaKaRa公司)提取总RNA,总RNA使用DnaseI(TaKaRa)在37℃下去除DNA污染,然后采用ReverTra Ace-a-TM试剂盒(Toyobo,Japan)合成cDNA。使用Nanodrop 1000紫外分光光度计(Thermo Scientific)测定cDNA浓度,稀释到200 ng·μL-1后用于RT-PCR。RT-PCR反应体系(25 μL)包括5×缓冲液2.5 μL,dNTP(2.5 mmol·L-1 each)0.5 μL,MgCl2(1.5 mmol·L-1)1.5 μL,Taq聚合酶1 μL,正向和反向引物各1 μL(终浓度为0.25 μmol·L-1,引物见表2),cDNA模板1 μL,加去离子水至25 μL。反应程序:94℃预变性5 min;94℃变性40 s,58℃退火40 s,72℃延伸40 s,共28个循环,72℃保温10 min。用于表达分析的抗病相关基因有CHI(chitinase)、AOS(allene oxide synthase)、GPX(glutathione peroxidase)和PR4A(pathogenesis-related protein 4A),以Actin作为内参基因。Table 2
表2
表2本研究中RT-PCR分析所用引物序列
Table 2Primer sequences used for RT-PCR analysis in this study
| 基因 Gene | 引物序列Primer sequence (5′-3′) | |
|---|---|---|
| 正向引物Forward primer | 反向引物Reverse primer | |
| CHI | TCTTGCCCTAGCTTTTCCCAC | GCAATCTCACGCTTC GAAACTT |
| AOS | ATTCCACCTACACGGAGGCAT | TAAC GGAGCGAGCTGAAACAG |
| GPX | CAGTGTGGCTTGACCAATTCAA | CCCCTCCTTATCCACCAAGAAC |
| PR4A | ACATAACTGTAGTGCCCATGAGC | GGAGGCTTAGATTTGGACGAAGG |
| Actin | ATCCCTCAGCACCTTCC | CCAACCTTAGCACTTCTCC |
新窗口打开
1.6 数据统计与分析
每个处理设置3个重复,数据采用Excel 2010作图,通过Excel自带软件ANOVA(Analysis of Variance)进行差异显著性分析(P<0.05)。2 结果
2.1 不同温度培养植株叶片发病情况
冰糖橙植株在21℃和30℃培养3 d后用于溃疡病菌接种,在接种后第3、7和10天观察两个处理组植株的发病率。两个处理组植株叶片在第3天均出现病斑,但二者的发病率却存在差异。21℃条件下培养植株叶片在第3天的发病率为10%,而30℃条件下培养植株叶片的发病率接近40%。在第7天时两组材料的发病率均有所增加,但30℃条件下培养植株的发病率更高。然而,在第10天时,两个处理组植株的发病率接近(图1)。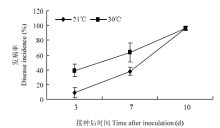 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图121℃和30℃条件下预培养3 d的植株接种Xcc后的发病率
-->Fig. 1Canker disease incidence of Xcc-inoculated plants that have been pre-cultured at 21℃ or 30℃ for three days
-->
2.2 不同温度处理后叶片多胺含量和抗病基因表达比较
多胺与植物的抗病能力有关[13,14],因此,分析了21℃和30℃下培养3 d的冰糖橙植株自由态多胺含量。由图2-A可见,两组处理材料中均能成功地检测到常见的3种自由态多胺,即腐胺(Put)、亚精胺(Spd)和精胺(Spm)。但21℃条件下预培养植株叶片自由态Put、Spd和Spm含量均高于30℃下培养植株,表明高温条件下植株多胺合成减少。植物抗病性差异很大程度上与抗病基因表达有关[21,22],因此,对21℃和30℃培养植株4个抗病基因CHI、GPX、AOS和PR4A的表达进行分析。由图2-B可以看出,除AOS表达水平在两组材料中差异不明显外,其余3个基因的表达水平在30℃培养植株中均要低于21℃培养植株,表明高温可能抑制了抗病基因的表达。
2.3 外源亚精胺处理后植株内源多胺含量及抗病基因表达变化
由图2中可以看出,冰糖橙中Spd含量高于Put和Spm。因此,对冰糖橙植株外源施用Spd,以分析是否有利于增强抗病性。外源Spd处理3 d后,采集叶片分析游离态多胺含量(图3-A)。可以看出,与对照(清水)处理相比,Spd处理材料中Put和Spd含量均明显增加,但Spm含量差异不大。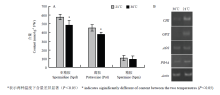 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图221℃和30℃预培养3 d的植株自由态多胺含量(A)和抗病基因表达量(B)
-->Fig. 2Free polyamine contents (A) and expression levels of defense-related genes (B) in plants pre-cultured at 21℃ or 30℃ for three days
-->
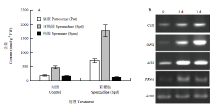 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3外源亚精胺和清水处理后叶片自由态多胺含量(A)和抗病基因表达量(B)
-->Fig. 3Free polyamine levels (A) and expression levels of defense-related genes (B) in plants treated with exogenous spermidine or water
-->
Spd处理3 d后,AOS、CHI、GPX和PR4A 4个抗病基因的表达上调,其中GPX和AOS诱导最为强烈;在处理7 d后CHI和GPX表达水平达到最大值,而AOS和PR4A的表达水平维持稳定(图3-B)。
2.4 外源亚精胺处理对溃疡病抗性的影响
将清水或Spd处理叶片针刺接种Xcc,接种后观察发病情况。由表型可见,接种后第7天时两个处理组的叶片可以看到白色病斑,但Spd处理材料出现病斑的伤口明显比对照组少或小;在第14天时对照发病程度重于Spd处理材料(图4-A)。通过发病率比较也可以看出,在接种后14 d时Spd处理材料的发病率约为40%,而对照组在接种后14 d时的发病率为85%(图4-B)。Spd处理组14 d时病情指数为4.2,但清水对照组在接种后第14天病情指数为9.0(图4-C)。结果表明,外源Spd处理能够增强冰糖橙叶片对柑橘溃疡病的抗性。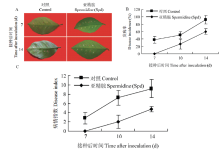 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4外源Spd处理对溃疡病抗性的影响
-->Fig. 4Effects of exogenous spermidine treatment on canker disease resistance
-->
3 讨论
植物对病原入侵的应答受多个因素影响,其中一个重要因素是温度[23]。通常认为,适合Xcc生长的最佳温度范围为20—30℃,在此范围里温度增加则会加重溃疡病发生[24]。本研究发现,在较高温度下培养一段时间的植株对Xcc更敏感,表明高温能促进寄主的发病或降低其抗病性,研究结果在一定程度上解释了柑橘在高温季节更容易感染溃疡病的原因;另一方面也暗示,在高温季节做好柑橘溃疡病防治可能对于控制该病的发生和蔓延具有关键作用。当病原入侵时,植物会启动非病原特异性抗病反应,这是一种先天免疫特征,在感病或抗病植株、合适或不合适生长条件下均存在[25]。此过程中,植物在生理、代谢、细胞和分子水平上发生一系列的改变,包括代谢物的积累和基因表达的变化[21,26]。本研究发现,30℃条件下植株体内积累的多胺含量低于21℃条件下生长的植株;此外,还发现3个抗病基因表达水平在高温下受到抑制。高温下生长的植株易感染溃疡病可能与其体内积累的代谢物(如本研究中的多胺)减少或抗病基因(如CHI、AOS和PR4A等)表达受抑制有关。高温下多胺含量减少可能由于合成受阻所致,前期有研究表明,多胺合成关键基因ADC(精氨酸脱羧酶)在高温下表达下调,而多胺含量一定程度上取决于其合成基因在转录水平上的表达[11,27]。因此,高温下多胺合成可能受到抑制,从而减少体内多胺含量。高温下CHI、AOS和PR4A表达受抑制的原因尚不清楚,一个可能的机制是高温抑制了调控上述3个抗病基因的转录激活因子或激活了某些转录抑制因子,使抗病基因的表达受到抑制,这一推论是否正确需要进一步证实。
本研究中,高温下植株多胺含量减少,其溃疡病抗性降低;相反,外源Spd处理提高了内源Spd含量,并增强了所处理植株的溃疡病抗性,表明柑橘植株积累较高水平的多胺有利于提高其溃疡病抗性。事实上,前期已有研究发现,超表达多胺合成基因MdSPDS1提高了甜橙转基因植株内源多胺含量,显著增强了转基因植株对溃疡病的抗性[8]。多胺在抗病中的作用机制可能体现在如下两个方面:(1)多胺作为信号分子参与抗病信号的传递[28,29],但这一机制是否在溃疡病抗性中发挥作用尚待证实;(2)多胺降解产生H2O2诱发抗病反应。研究表明,多胺合成后被转运到质外体,进而由多胺氧化酶分解产生H2O2[30,31],而H2O2是调节气孔运动和引起过敏性反应(HR)的重要信号分子[32]。气孔是溃疡病菌进入植株体内的主要通道,当H2O2产生后能够促进气孔关闭,阻止溃疡病菌通过气孔进入体内[7];此外,H2O2可以促进HR引起的细胞死亡从而将病原菌局限在接种部位。已有较多研究表明多胺分解形成的H2O2在植物防御病原入侵方面发挥着重要的作用[14,16,32-36]。因此,多胺更可能是通过产生H2O2来促进气孔关闭或触发局部HR来增强溃疡病抗性。
病原入侵会产生相应的胁迫信号,植物感知和传递该信号并启动相应的防御网络,最终通过改变大量抗病基因的表达水平来应答病原入侵,暗示抗病基因的表达变化是植物-病原互作过程中一个极为重要的防御反应[22,26,37]。研究表明,PR蛋白是植物防御病原侵染最为重要的一类基因,通常被用作植物抗病反应的标记基因。CHI与PR4A分别是PR3和PR4类PR蛋白,在植物系统获得性抗性中发挥着重要作用[38]。GPX参与细胞生化反应的多个过程,在保护寄主植物被病原入侵中也发挥关键作用[39],AOS是茉莉酸合成的一种重要酶,而茉莉酸在植物防御反应中具有重要作用[40]。本研究中,发现两种温度下培养的植株虽然AOS表达水平变化不大,但30℃下生长的植株CHI、GPX和PR4A表达水平明显低于21℃下生长的植株。此外,外源Spd增强了溃疡病抗性,与之一致的是4个基因的表达水平得以升高。可以清楚地看出,上述基因的表达水平与溃疡病抗性呈正相关,表明它们在柑橘抗病反应中可能有重要的作用,是今后开展分子育种增强溃疡病抗性的重要候选基因。
4 结论
高温下甜橙更容易受柑橘溃疡病菌危害,其可能的机制是高温抑制抗病基因的表达和降低多胺的合成。外源多胺处理能够增强甜橙溃疡病抗性,可能原因是多胺处理后增强了抗病基因的表达,从而诱发植株的抗病反应最终表现出抗病。高温是影响溃疡病发生的一个关键环境因素,多胺有助于提高对柑橘溃疡病的抗性。The authors have declared that no competing interests exist.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | . |
| [2] | . 【目的】采用新的接种方式,比较冰糖橙与枸橼 C-05叶片对溃疡病菌生长特性的的影响,探讨冰糖橙与枸橼 C-05对溃疡病菌敏感度的差异性。【方法】将完全展开、颜色淡绿的冰糖橙与枸橼 C-05叶片进行75%酒精和1% NaClO 消毒并切割后,分别与溃疡病菌在离体条件下共培养,观察叶片在 MT 培养基(对照)、MT 中间嵌入NYGA 培养基(MA)中对溃疡病菌生长的影响;分别提取冰糖橙和枸橼 C-05叶片的粗提液,并以25%、50%和75%的比例添加至 MT 培养基中,观察冰糖橙和枸橼 C-05叶片提取液分别对溃疡病菌生长的影响;同时以不同比例的叶片粗提液添加至 NYGB 培养基中,观察冰糖橙与枸橼 C-05叶片提取液分别对溃疡病菌胞外多糖含量的影响。【结果】Xac 在培养冰糖橙叶片的 MT 培养基中生长迅速,接种1周后即出现 Xac 的增殖现象,菌斑直径均值比对照大0.5 mm;接种2周后菌斑直径均值为5.27 mm,几乎是对照的3倍;而2—3周为 Xac 的急速生长期,菌斑直径由5.27 mm 快速增长至13.41 mm,是对照菌斑直径的5倍多;3周后菌斑生长缓慢趋于平稳;各个时间点的菌斑直径与对照相比均达到极显著差异水平。在培养冰糖橙叶片的 MA 培养基中,Xac 生长1 d 后菌斑直径均值为4.58 mm,小于对照的6.19 mm;接种3 d 后 Xac 继续生长,菌斑直径几乎与对照组相同,差异不显著;但是在接种5 d 后,菌斑直径快速增长至21.31 mm,大于对照组的16.33 mm,3个菌斑长势连在一起,两者差异达到极显著水平。冰糖橙的叶片提取液对溃疡病菌的生长及胞外多糖含量均有促进作用,随着其添加浓度的增加,促进效果 越明显,与对照相比均可达到极显著水平。Xac 在培养枸橼 C-05叶片的 MT 培养基中生长较为缓慢,在接种1周后菌斑直径与对照相比差异不显著;2周后菌斑直径约为对照的一半;3周时,菌斑直径均值为2.06 mm,小于对照的2.62 mm;2周和3周时的菌斑直径与对照相比均达到显著水平。在培养枸橼 C-05叶片的 MA 培养基中,Xac 生长较为缓慢,菌斑直径均小于对照组,在1、3和5 d 时与对照相比均达到极显著水平。枸橼 C-05的叶片提取液对溃疡病菌生长和胞外多糖含量的影响不大,与对照相比均差异不显著。【结论】冰糖橙叶片中存在可促进溃疡病菌的生长、增加胞外多糖含 量的物质,这种物质可能突破了其自身免疫系统,导致其对 Xac 高度敏感而极易感病。枸橼 C-05叶片与溃疡病菌共培养时,分泌抑制溃疡病菌生长的物质,使得枸橼 C-05对 Xac 具有较低敏感度而表现抗性。 【目的】采用新的接种方式,比较冰糖橙与枸橼 C-05叶片对溃疡病菌生长特性的的影响,探讨冰糖橙与枸橼 C-05对溃疡病菌敏感度的差异性。【方法】将完全展开、颜色淡绿的冰糖橙与枸橼 C-05叶片进行75%酒精和1% NaClO 消毒并切割后,分别与溃疡病菌在离体条件下共培养,观察叶片在 MT 培养基(对照)、MT 中间嵌入NYGA 培养基(MA)中对溃疡病菌生长的影响;分别提取冰糖橙和枸橼 C-05叶片的粗提液,并以25%、50%和75%的比例添加至 MT 培养基中,观察冰糖橙和枸橼 C-05叶片提取液分别对溃疡病菌生长的影响;同时以不同比例的叶片粗提液添加至 NYGB 培养基中,观察冰糖橙与枸橼 C-05叶片提取液分别对溃疡病菌胞外多糖含量的影响。【结果】Xac 在培养冰糖橙叶片的 MT 培养基中生长迅速,接种1周后即出现 Xac 的增殖现象,菌斑直径均值比对照大0.5 mm;接种2周后菌斑直径均值为5.27 mm,几乎是对照的3倍;而2—3周为 Xac 的急速生长期,菌斑直径由5.27 mm 快速增长至13.41 mm,是对照菌斑直径的5倍多;3周后菌斑生长缓慢趋于平稳;各个时间点的菌斑直径与对照相比均达到极显著差异水平。在培养冰糖橙叶片的 MA 培养基中,Xac 生长1 d 后菌斑直径均值为4.58 mm,小于对照的6.19 mm;接种3 d 后 Xac 继续生长,菌斑直径几乎与对照组相同,差异不显著;但是在接种5 d 后,菌斑直径快速增长至21.31 mm,大于对照组的16.33 mm,3个菌斑长势连在一起,两者差异达到极显著水平。冰糖橙的叶片提取液对溃疡病菌的生长及胞外多糖含量均有促进作用,随着其添加浓度的增加,促进效果 越明显,与对照相比均可达到极显著水平。Xac 在培养枸橼 C-05叶片的 MT 培养基中生长较为缓慢,在接种1周后菌斑直径与对照相比差异不显著;2周后菌斑直径约为对照的一半;3周时,菌斑直径均值为2.06 mm,小于对照的2.62 mm;2周和3周时的菌斑直径与对照相比均达到显著水平。在培养枸橼 C-05叶片的 MA 培养基中,Xac 生长较为缓慢,菌斑直径均小于对照组,在1、3和5 d 时与对照相比均达到极显著水平。枸橼 C-05的叶片提取液对溃疡病菌生长和胞外多糖含量的影响不大,与对照相比均差异不显著。【结论】冰糖橙叶片中存在可促进溃疡病菌的生长、增加胞外多糖含 量的物质,这种物质可能突破了其自身免疫系统,导致其对 Xac 高度敏感而极易感病。枸橼 C-05叶片与溃疡病菌共培养时,分泌抑制溃疡病菌生长的物质,使得枸橼 C-05对 Xac 具有较低敏感度而表现抗性。 |
| [3] | . 以纽荷尔脐橙和四季橘为试验材料,基于溃疡病菌诱导的柑橘转录组数据库,利用PCR方法克隆获得4个WRKY家族基因Cs WRKY22、Cs WRKY50、CsWRKY72-1和CsWRKY72-2的cDNA全长序列。序列分析结果表明,这4个基因的cDNA全长分别为1 123、1 312、1 809和2 208 bp,开放阅读框长度分别为921、480、1 809和1 767 bp,各编码306、159、602和588个氨基酸。氨基酸序列和结构分析显示,这4个基因属于第Ⅱ类WRKY蛋白。进化树分析显示,所克隆的4个柑橘WRKY蛋白与可可、葡萄等WRKY蛋白亲缘关系较近。亚细胞定位显示,所克隆的4个WRKY基因定位于细胞核,与亚细胞定位预测相符。对纽荷尔脐橙和四季橘中4个基因受溃疡病菌诱导的表达分析表明,CsWRKY22参与寄主的感病反应,而CsWRKY50参与抗溃疡病的免疫应答反应。柑橘溃疡病菌能抑制CsWRKY72-1和CsWRKY72-2介导的寄主基础免疫。水杨酸(SA)和茉莉酸甲酯(MeJA)处理表明,4个Cs WRKY基因均不参与SA和MeJA介导的寄主抗病和抗逆反应。 以纽荷尔脐橙和四季橘为试验材料,基于溃疡病菌诱导的柑橘转录组数据库,利用PCR方法克隆获得4个WRKY家族基因Cs WRKY22、Cs WRKY50、CsWRKY72-1和CsWRKY72-2的cDNA全长序列。序列分析结果表明,这4个基因的cDNA全长分别为1 123、1 312、1 809和2 208 bp,开放阅读框长度分别为921、480、1 809和1 767 bp,各编码306、159、602和588个氨基酸。氨基酸序列和结构分析显示,这4个基因属于第Ⅱ类WRKY蛋白。进化树分析显示,所克隆的4个柑橘WRKY蛋白与可可、葡萄等WRKY蛋白亲缘关系较近。亚细胞定位显示,所克隆的4个WRKY基因定位于细胞核,与亚细胞定位预测相符。对纽荷尔脐橙和四季橘中4个基因受溃疡病菌诱导的表达分析表明,CsWRKY22参与寄主的感病反应,而CsWRKY50参与抗溃疡病的免疫应答反应。柑橘溃疡病菌能抑制CsWRKY72-1和CsWRKY72-2介导的寄主基础免疫。水杨酸(SA)和茉莉酸甲酯(MeJA)处理表明,4个Cs WRKY基因均不参与SA和MeJA介导的寄主抗病和抗逆反应。 |
| [4] | . . |
| [5] | . Abstract The lateral organ boundary domain (LBD) genes encode a group of plant-specific proteins that function as transcription factors in the regulation of plant growth and development. Citrus sinensis lateral organ boundary 1 (CsLOB1) is a member of the LBD family and functions as a disease susceptibility gene in citrus bacterial canker (CBC). Thirty-four LBD members have been identified from the Citrus sinensis genome. We assessed the potential for additional members of LBD genes in citrus to function as surrogates for CsLOB1 in CBC, and compared host gene expression on induction of different LBD genes. Using custom-designed transcription activator-like (TAL) effectors, two members of the same clade as CsLOB1, named CsLOB2 and CsLOB3, were found to be capable of functioning similarly to CsLOB1 in CBC. RNA sequencing and quantitative reverse transcription-polymerase chain reaction analyses revealed a set of cell wall metabolic genes that are associated with CsLOB1, CsLOB2 and CsLOB3 expression and may represent downstream genes involved in CBC. 脗漏 2016 BSPP AND JOHN WILEY & SONS LTD. |
| [6] | . |
| [7] | Abstract Citrus is a highly valued tree crop worldwide, while, at the same time, citrus production faces many biotic challenges, including bacterial canker and Huanglongbing (HLB). Breeding for disease???resistant varieties is the most efficient and sustainable approach to control plant diseases. Traditional breeding of citrus varieties is challenging due to multiple limitations, including polyploidy, polyembryony, extended juvenility and long crossing cycles. Targeted genome editing technology has the potential to shorten varietal development for some traits, including disease resistance. Here, we used CRISPR/Cas9/sgRNA technology to modify the canker susceptibility gene CsLOB1 in Duncan grapefruit. Six independent lines, DLOB2, DLOB3, DLOB9, DLOB10, DLOB11 and DLOB12, were generated. Targeted next???generation sequencing of the six lines showed the mutation rate was 31.58%, 23.80%, 89.36%, 88.79%, 46.91% and 51.12% for DLOB2, DLOB3, DLOB9, DLOB10, DLOB11 and DLOB12, respectively, of the cells in each line. DLOB2 and DLOB3 showed canker symptoms similar to wild???type grapefruit, when inoculated with the pathogen Xanthomonas citri subsp. citri (Xcc). No canker symptoms were observed on DLOB9, DLOB10, DLOB11 and DLOB12 at 4??days postinoculation (DPI) with Xcc. Pustules caused by Xcc were observed on DLOB9, DLOB10, DLOB11 and DLOB12 in later stages, which were much reduced compared to that on wild???type grapefruit. The pustules on DLOB9 and DLOB10 did not develop into typical canker symptoms. No side effects and off???target mutations were detected in the mutated plants. This study indicates that genome editing using CRISPR technology will provide a promising pathway to generate disease???resistant citrus varieties. |
| [8] | pAbstract/p pBackground/p pEnormous work has shown that polyamines are involved in a variety of physiological processes, but information is scarce on the potential of modifying disease response through genetic transformation of a polyamine biosynthetic gene./p pResults/p pIn the present work, an apple itspermidine synthase /itgene (itMdSPDS1/it) was introduced into sweet orange (itCitrus sinensis /itOsbeck Anliucheng) via itAgrobacterium/it-mediated transformation of embryogenic calluses. Two transgenic lines (TG4 and TG9) varied in the transgene expression and cellular endogenous polyamine contents. Pinprick inoculation demonstrated that the transgenic lines were less susceptible to itXanthomonas axonopodis /itpv. itcitri /it(Xac), the causal agent of citrus canker, than the wild type plants (WT). In addition, our data showed that upon Xac attack TG9 had significantly higher free spermine (Spm) and polyamine oxidase (PAO) activity when compared with the WT, concurrent with an apparent hypersensitive response and the accumulation of more Hsub2/subOsub2/sub. Pretreatment of TG9 leaves with guazatine acetate, an inhibitor of PAO, repressed PAO activity and reduced Hsub2/subOsub2 /subaccumulation, leading to more conspicuous disease symptoms than the controls when both were challenged with Xac. Moreover, mRNA levels of most of the defense-related genes involved in synthesis of pathogenesis-related protein and jasmonic acid were upregulated in TG9 than in the WT regardless of Xac infection./p pConclusion/p pOur results demonstrated that overexpression of the itMdSPDS1 /itgene prominently lowered the sensitivity of the transgenic plants to canker. This may be, at least partially, correlated with the generation of more Hsub2/subOsub2 /subdue to increased production of polyamines and enhanced PAO-mediated catabolism, triggering hypersensitive response or activation of defense-related genes./p |
| [9] | |
| [10] | Citrus canker caused by Xanthomonas axonopodis pv. citri (Xac) is a devastating bacterial disease threatening the citrus industry. Salicylic acid (SA) plays a key role in plant defense response to biotic stress, but information is scarce concerning the application of SA to enhancing Xac resistance. In the present research attempts were made to investigate how exogenous application of SA influenced canker disease outbreak in navel orange (Citrus sinensis). Exogenously applied SA at 0.25mM significantly enhanced the endogenous free and bound SA, particularly the latter. Upon exposure to Xac, lower disease incidence rate and smaller lesion sites were observed in the samples pre-treated with SA, accompanied by repression of bacterial growth at the lesion sites. Concurrent with the augmented disease resistance, SA-treated leaves had higher H2O2 level and smaller stomata apertures before or after Xac infection when compared with their counterparts pre-treated with water (control). SA treatment elevated the activities of phenylalanine ammonia-lyase and 尾-1,3-glucanase, but only the latter was higher in the SA-treated samples after Xac infection. In addition, mRNA levels of two pathogenesis-related genes, CsCHI and CsPR4A, were higher in the SA-treated samples relative to the control. Taken together, our results strongly suggest that the exogenously applied SA has evoked a cascade of physiological and molecular events that function singly or in concert to confer resistance to Xac invasion. |
| [11] | . Plants are challenged by a variety of biotic or abiotic stresses, which can affect their growth and development, productivity, and geographic distribution. In order to survive adverse environmental conditions, plants have evolved various adaptive strategies, among which is the accumulation of metabolites that play protective roles. A well-established example of the metabolites that are involved in stress responses, or stress tolerance, is the low-molecular-weight aliphatic polyamines, including putrescine, spermidine, and spermine. The critical role of polyamines in stress tolerance is suggested by several lines of evidence: firstly, the transcript levels of polyamine biosynthetic genes, as well as the activities of the corresponding enzymes, are induced by stresses; secondly, elevation of endogenous polyamine levels by exogenous supply of polyamines, or overexpression of polyamine biosynthetic genes, results in enhanced stress tolerance; and thirdly, a reduction of endogenous polyamines is accompanied by compromised stress tolerance. A number of studies have demonstrated that polyamines function in stress tolerance largely by modulating the homeostasis of reactive oxygen species (ROS) due to their direct, or indirect, roles in regulating antioxidant systems or suppressing ROS production. The transcriptional regulation of polyamine synthesis by transcription factors is also reviewed here. Meanwhile, future perspectives on polyamine research are also suggested. |
| [12] | . |
| [13] | . |
| [14] | . Although most work on polyamines in incompatible interactions between plants and pathogens has focussed on polyamines conjugated to phenolic compounds (hydroxycinnamic acid amides), changes in free polyamines and their catabolism have been shown to occur in such interactions. A common feature of these interactions is an increase in diamine oxidase (DAO) activity and, in some interactions, of polyamine oxidase (PAO). The activities of these two enzymes produces hydrogen peroxide (H 2 O 2 ), which may act in structural defense, as a signal molecule, or as an antimicrobial compound in host resistance. There are several possible roles for polyamines and polyamine catabolism in plant resistance to pathogen infection; H 2 O 2 produced might trigger the hypersensitive response (HR), thought to be a form of programmed cell death (PCD), the polyamine spermine might act as an inducer of PR proteins, and as a trigger for caspase activity and hence HR. There is, however, a need for more precise information on the timing and location of changes in polyamine metabolism in the development of resistance. Only with this information can a case be made for the involvement of polyamines and polyamine catabolism in plant defense. |
| [15] | . Abstract 090004 Change is reported in the biosynthetic and oxidative activity of hypersensitive (NN) and susceptible (nn) tobacco ( Nicotiana tabacum ) plants in response to tobacco mosaic virus (TMV). 090004 Mature leaves of nn and NN tobacco were collected over 009000972 h as uninoculated controls or after inoculation with TMV or phosphate buffer (mock-inoculation). The polyamine response to inoculation was analysed by measuring activity and gene expression of S-adenosylmethionine decarboxylase (SAMDC), arginine-(ADC) and ornithine decarboxylases (ODC); incorporation of labelled putrescine; and activity of diamine oxidase (DAO). 090004 In NN leaves SAMDC activity and transcript levels, and DAO activity increased in the TMV-inoculated plants but not in the other treatments; a two-fold increase in DAO activity was seen after 72 h. Both ADC and ODC activity increased in NN leaves at 72 h in TMV-inoculated plants; ADC mRNA increased with activity. The increase in SAMDC mRNA (24 h) preceded the rise in activity (72 h). [ 3 H]putrescine added to NN leaves led to enhanced label recovery and incorporation into spermidine and spermine in TMV-inoculated plants. No significant changes in biosynthetic or oxidative activity occurred in nn plants. 090004 After TMV inoculation, NN, unlike nn, tobacco plants upgrade polyamine synthesis and oxidation; this leads to changes in cellular components which might induce programmed cell death. |
| [16] | Polyamine levels and activities of enzymes of polyamine biosynthesis and catabolism were examined in the barley cultivar Delibes ( Ml1al + Ml(Ab) ) reacting hypersensitively to the powdery mildew fungus, Blumeria graminis f. sp. hordei (race CC220). Levels of free putrescine and spermine and of conjugated forms of putrescine, spermidine and spermine were greatly increased 1鈥4 d following inoculation of barley with the powdery mildew. These changes in polyamine levels were accompanied by elevated activities of the polyamine biosynthetic enzymes ornithine decarboxylase (ODC), arginine decarboxylase (ADC) and S -adenosylmethionine decarboxylase (AdoMetDC) and the polyamine catabolic enzymes diamine oxidase (DAO) and polyamine oxidase (PAO). Activities of two enzymes involved in conjugating polyamines to hydroxycinnamic acids, putrescine hydroxycinnamoyl transferase (PHT) and tyramine feruloyl-CoA transferase (TFT) were also examined and were found to increase significantly 1鈥4 d after inoculation. The possibility that the increased levels of free spermine, increased polyamine conjugates, and increased DAO and PAO activities are involved in development of the hypersensitive response of Delibes to powdery mildew infection is discussed. |
| [17] | The role of polyamine (PA) metabolism in tobacco (Nicotiana tabacum) defense against pathogens with contrasting pathogenic strategies was evaluated. Infection by the necrotrophic fungus Sclerotinia sclerotiorum resulted in increased arginine decarboxylase expression and activity in host tissues, as well as putrescine and spermine accumulation in leaf apoplast. Enhancement of leaf PA levels, either by using transgenic plants or infiltration with exogenous PAs, led to increased necrosis due to infection by S. sclerotiorum. Specific inhibition of diamine and PA oxidases attenuated the PA-induced enhancement of leaf necrosis during fungal infection. When tobacco responses to infection by the biotrophic bacterium Pseudomonas viridiflava were investigated, an increase of apoplastic spermine levels was detected. Enhancement of host PA levels by the above-described experimental approaches strongly decreased in planta bacterial growth, an effect that was blocked by a PA oxidase inhibitor. It can be concluded that accumulation and further oxidation of free PAs in the leaf apoplast of tobacco plants occurs in a similar, although not identical way during tobacco defense against infection by microorganisms with contrasting pathogenesis strategies. This response affects the pathogen's ability to colonize host tissues and results are detrimental for plant defense against necrotrophic pathogens that feed on necrotic tissue; on the contrary, this response plays a beneficial role in defense against biotrophic pathogens that depend on living tissue for successful host colonization. Thus, apoplastic PAs play important roles in plant-pathogen interactions, and modulation of host PA levels, particularly in the leaf apoplast, may lead to significant changes in host susceptibility to different kinds of pathogens. |
| [18] | . |
| [19] | Abscisic acid-responsive element (ABRE)-binding factors (ABFs) play important roles in abiotic stress responses; however, the underlying mechanisms are poorly understood. In this study, it is reported that overexpression of Poncirus trifoliata PtrABF significantly enhanced dehydration tolerance. The transgenic lines displayed smaller stomatal apertures, reduced stomatal density/index, and lower expression levels of genes associated with stomatal development. PtrABF was found to interact with PtrICE1, a homologue of ICE1 (Inducer of CBF Expression 1) that has been shown to be critical for stomatal development. Microarray analysis revealed that a total of 70 genes were differentially expressed in the transgenic line, 42 induced and 28 repressed. At least two units of ABREs and coupling elements were present in the promoters of most of the induced genes, among which peroxidase and arginine decarboxylase were verified as bona fide targets of PtrABF. Transgenic plants exhibited higher antioxidant enzyme activities and free polyamine levels, but lower levels of reactive oxygen species (ROS) and malondialdehyde. Polyamines were revealed to be associated with ROS scavenging in the transgenic plants due to a modulation of antioxidant enzymes triggered by signalling mediated by H2O2 derived from polyamine oxidase (PAO)-mediated catabolism. Taken together, the results indicate that PtrABF functions positively in dehydration tolerance by limiting water loss through its influence on stomatal movement or formation and maintaining ROS homeostasis via modulation of antioxidant enzymes and polyamines through transcriptional regulation of relevant target genes. |
| [20] | Polyamine oxidase (PAO) is a key enzyme catalyzing polyamine catabolism leading to H2O2production. We previously demonstrated thatCitrus sinensiscontains six putativePAOgenes, but their functions are not well understood. In this work, we reported functional elucidation ofCsPAO4in polyamine catabolism and salt stress response. CsPAO4 was localized to the apoplast and used both spermidine (Spd) and spermine (Spm) as substrates for terminal catabolism. Transgenic plants overexpressingCsPAO4displayed prominent increase in PAO activity, concurrent with marked decrease of Spm and Spd and elevation of H2O2. Seeds of transgenic lines displayed better germination when compared with wild type (WT) under salt stress. However, both vegetative growth and root elongation of the transgenic lines were prominently inhibited under salt stress, accompanied by higher level of H2O2and more conspicuous programmed cell death (PCD). Exogenous supply of catalase (CAT), a H2O2scavenger, partially recovered the vegetative growth and root elongation. In addition, spermine inhibited root growth of transgenic plants. Taken together, these data demonstrated thatCsPAO4accounts for production of H2O2causing oxidative damages under salt stress and that down-regulation of aPAOgene involved in polyamine terminal catabolism may be an alternative approach for improving salt stress tolerance. |
| [21] | . Induced defence responses are elicited when plants are exposed to biotic stresses such as attack by herbivores or pathogens. In nature, plants are often subjected to attack by more than one organism, and defence responses elicited by one organism can thereby be modified by the presence of another. Below-ground attack can influence responses to above-ground attack and vice versa, due to systemic induction of defence metabolism. In some interactions defence is enhanced through prior attack by another organism, whereas in others there are conflicting signals. Recent research has shown how plants integrate these signals to coordinate defence by regulation of key metabolic pathways, although there is still much to be learnt. |
| [22] | . Abstract The hormone jasmonate (JA), which functions in plant immunity, regulates resistance to pathogen infection and insect attack through triggering genome-wide transcriptional reprogramming in plants. We show that the basic helix-loop-helix transcription factor (TF) MYC2 in tomato (Solanum lycopersicum) acts downstream of the JA receptor to orchestrate JA-mediated activation of both the wounding and pathogen responses. Using chromatin immunoprecipitation sequencing (ChIP-seq) coupled with RNA sequencing (RNA-seq) assays, we identified 655 MYC2-targeted JA-responsive genes. These genes are highly enriched in Gene Ontology categories related to TFs and the early response to JA, indicating that MYC2 functions at a high hierarchical level to regulate JA-mediated gene transcription. We also identified a group of MYC2-targeted TFs (MTFs) that may directly regulate the JA-induced transcription of late defense genes. Our findings suggest that MYC2 and its downstream MTFs form a hierarchical transcriptional cascade during JA-mediated plant immunity that initiates and amplifies transcriptional output. As proof of concept, we showed that during plant resistance to the necrotrophic pathogen Botrytis cinerea, MYC2 and the MTF JA2-Like (JA2L) form a transcription module that preferentially regulates wounding-responsive genes, whereas MYC2 and the MTF ETHYLENE RESPONSE FACTOR.C3 (ERF.C3) form a transcription module that preferentially regulates pathogen-responsive genes. 漏 2017 American Society of Plant Biologists. All rights reserved. |
| [23] | . Plants perceive and integrate intrinsic and extrinsic signals to execute appropriate responses for maximal survival and reproductive success. Plant immune responses are tightly controlled to ensure effective defenses against pathogens while minimizing their adverse effects on plant growth and development. Plant defenses induced in response to pathogen infection are modulated by abiotic signals such as light, circadian rhythm, and temperature. The modulation occurs on specific key components of plant immunity, indicating an intricate integration of biotic and abiotic signals. This review will summarize very recent studies revealing the intersection of plant defenses with light, circadian rhythm and temperature. In addition, it will discuss the adaptive value and evolutionary constraints of abiotic regulation of plant immunity. |
| [24] | . 多年田间系统观察表明 ,柑橘溃疡病的消长 ,在春梢期 ,主要影响因子为气温。 4月下旬平均气温 2 0℃持续 1 0~ 1 5d ,田间始病 ;平均气温 2 5℃连续 3d ,田间进入病情激增期。在夏、秋梢期 ,主要影响因子为湿度和降雨。相对湿度低于 70 % ,不利于发病 ;持续阴雨或短时大雨极有利于流行。溃疡病的药剂防治应掌握在春梢展叶后 7d用药 ,夏、秋梢则萌芽后即用药。药剂品种以可杀得和百菌清最理想 ,喷药次数以 3次为好。 . 多年田间系统观察表明 ,柑橘溃疡病的消长 ,在春梢期 ,主要影响因子为气温。 4月下旬平均气温 2 0℃持续 1 0~ 1 5d ,田间始病 ;平均气温 2 5℃连续 3d ,田间进入病情激增期。在夏、秋梢期 ,主要影响因子为湿度和降雨。相对湿度低于 70 % ,不利于发病 ;持续阴雨或短时大雨极有利于流行。溃疡病的药剂防治应掌握在春梢展叶后 7d用药 ,夏、秋梢则萌芽后即用药。药剂品种以可杀得和百菌清最理想 ,喷药次数以 3次为好。 |
| [25] | . Salicylic acid (SA) has been reported to induce plant defense responses. The transcriptions of defense genes that are responsible for a given plant鈥檚 resistance to an array of plant pathogens are activated in a process called non-host resistance. Biotic signals capable of carrying out the activation of pathogenesis-related (PR) genes in pea tissue include fungal DNase and chitosan, two components released fromFusarium solanispores that are known to target host DNA. Recent reports indicate that SA also has a physical affinity for DNA. Here, we report that SA-induced reactive oxygen species release results in fragment alterations in pea nuclear DNA and cytologically detectable diameter and structural changes in the pea host nuclei. Additionally, we examine the subsequent SA-related increase of resistance to the true pea pathogenF. solanif.sp.pisiand the accumulation of the phytoalexin pisatin. This is the first report showing that SA-induced PR gene activation may be attributed to the host pea genomic DNA damage and that at certain concentrations, SA can be temporally associated with subsequent increases in the defense response of this legume. |
| [26] | . |
| [27] | Arginine decarboxylase (ADC), one of the enzymes responsible for putrescine (Put) biosynthesis, has been shown to be implicated in stress response. In the current paper attempts were made to clone and characterize a gene encoding ADC from peach ( Prunus persica (L.) Batsch, ‘Akatsuki’). Rapid amplification of cDNA ends (RACE) gave rise to a full-length ADC cDNA ( PpADC) with a complete open reading frame of 217802bp, encoding a 725 amino acid polypeptide. Homology search and sequence multi-alignment demonstrated that the deduced PpADC protein sequence shared a high identity with ADCs from other plants, including several highly conservative motifs and amino acids. Southern blotting indicated that PpADC existed in peach genome as a single gene. Expression levels of PpADC in different tissues of peach ( P. persica ‘Akatsuki’) were spatially and developmentally regulated. Treatment of peach shoots from ‘Mochizuki’ with exogenous 502mM Put, an indirect product of ADC, remarkably induced accumulation of PpADC mRNA. Transcripts of PpADC in peach leaves from ‘Mochizuki’ were quickly induced, either transiently or continuously, in response to dehydration, high salinity (20002mM NaCl), low temperature (402°C) and heavy metal (1500208M CdCl 2), but repressed by high temperature 3702°C) during a 2-day treatment, which changed in an opposite direction when the stresses were otherwise removed with the exception of CdCl 2 treatment. In addition, steady-state of PpADC mRNA could be also transiently up-regulated by abscisic acid (ABA) in ‘Mochizuki’ leaves. All of these, taken together, suggest that PpADC is a stress-responsive gene and can be considered as a potential target that is genetically manipulated so as to create novel germplasms with enhanced stress tolerance in the future. |
| [28] | Polyamines play pivotal roles in plant defense to environmental stresses. However, stress tolerance of genetically engineered plants for polyamine biosynthesis has been little examined so far. We cloned spermidine synthase cDNA from Cucurbita ficifolia and the gene was introduced to Arabidopsis thaliana under the control of the cauliflower mosaic virus 35S promoter. The transgene was stably integrated and actively transcribed in the transgenic plants. As compared with the wild-type plants, the T2 and T3 transgenic plants exhibited a significant increase in spermidine synthase activity and spermidine content in leaves together with enhanced tolerance to various stresses including chilling, freezing, salinity, hyperosmosis, drought, and paraquat toxicity. During exposure to chilling stress (5脗掳C), the transgenics displayed a remarkable increase in arginine decarboxylase activity and conjugated spermidine contents in leaves compared to the wild type. A cDNA microarray analysis revealed that several genes were more abundantly transcribed in the transgenics than in the wild type under chilling stress. These genes included those for stress-responsive transcription factors such as DREB and stress-protective proteins like rd29A. These results strongly suggest an important role for spermidine as a signaling regulator in stress signaling pathways, leading to build-up of stress tolerance mechanisms in plants under stress conditions. |
| [29] | We have proposed that the polyamine spermine functions as a signaling molecule to evoke defense reactions/cell death in N-gene bearing Nicotiana tabacuminfected with tobacco mosaic virus (TMV). In this work we examined whether this spermine-based signal transduction pathway is present in other plants such as Arabidopsis thaliana. For this purpose, we used the experimental system of cucumber mosaic virus-yellow strain (CMV-Y) and A. thaliana ecotype C24 carrying the RCY-1 resistance gene, in which a hypersensitive response is triggered. Almost all spermine-responsive genes in Arabidopsis were similarly regulated as in the TMV-Nicotiana tabacum system. In addition to the formerly identified spermine-signal component genes such as alternative oxidase- and mitogen-activated protein kinase 3 (WIPK orthologue)-genes, AtbZIP60, encoding an endoplasmic reticulum (ER)-localized transcription factor, was identified as a spermine-responsive gene, indicating that the ER may be involved in a spermine-signaling pathway. Treatment with polyamine oxidase inhibitors prior to CMV inoculation suppressed the expression of the genes mentioned above and compromised the host defense against CMV. Furthermore, exogenously added spermine suppressed the multiplication of CMV in Arabidopsis as did spermidine but less efficiently, while putrescine did not show such effects. Collectively, we conclude that the spermine-signaling pathway plays an important defensive role inA. thalianaas well as in N. tabacum when those plants are attacked by incompatible viral pathogens. |
| [30] | . |
| [31] | In contrast to animals, where polyamine (PA) catabolism efficiently converts spermine (Spm) to putrescine (Put), plants have been considered to possess a PA catabolic pathway producing 1,3-diaminopropane, 螖鹿-pyrroline, the corresponding aldehyde, and hydrogen peroxide but unable to back-convert Spm to Put. Arabidopsis (Arabidopsis thaliana) genome contains at least five putative PA oxidase (PAO) members with yet-unknown localization and physiological role(s). AtPAO1 was recently identified as an enzyme similar to the mammalian Spm oxidase, which converts Spm to spermidine (Spd). In this work, we have performed in silico analysis of the five Arabidopsis genes and have identified PAO3 (AtPAO3) as a nontypical PAO, in terms of homology, compared to other known PAOs. We have expressed the gene AtPAO3 and have purified a protein corresponding to it using the inducible heterologous expression system of Escherichia coli. AtPAO3 catalyzed the sequential conversion/oxidation of Spm to Spd, and of Spd to Put, thus exhibiting functional homology to the mammalian PAOs. The best substrate for this pathway was Spd, whereas the N鹿-acetyl-derivatives of Spm and Spd were oxidized less efficiently. On the other hand, no activity was detected when diamines (agmatine, cadaverine, and Put) were used as substrates. Moreover, although AtPAO3 does not exhibit significant similarity to the other known PAOs, it is efficiently inhibited by guazatine, a potent PAO inhibitor. AtPAO3 contains a peroxisomal targeting motif at the C terminus, and it targets green fluorescence protein to peroxisomes when fused at the N terminus but not at the C terminus. These results reveal that AtPAO3 is a peroxisomal protein and that the C terminus of the protein contains the sorting information. The overall data reinforce the view that plants and mammals possess a similar PA oxidation system, concerning both the subcellular localization and the mode of its action. |
| [32] | . Programmed cell death plays a critical role during the hypersensitive response in the plant defense system. One of components that triggers it is hydrogen peroxide, which is generated through multiple pathways. One example is proposed to be polyamine oxidation, but direct evidence for this has been limited. In this article, we investigated relationships among polyamine oxidase, hydrogen peroxide, and programmed cell death using a model system constituted of tobacco (Nicotiana tabacum) cultured cell and its elicitor, cryptogein. When cultured cells were treated with cryptogein, programmed cell death occurred with a distinct pattern of DNA degradation. The level of hydrogen peroxide was simultaneously increased, along with polyamine oxidase activity in apoplast. With the same treatment in the presence of 伪-difluoromethyl-Orn, an inhibitor of polyamine biosynthesis, production of hydrogen peroxide was suppressed and programmed cell death did not occur. A gene encoding a tobacco polyamine oxidase that resides in the apoplast was isolated and used to construct RNAi transgenic cell lines. When these lines were treated with cryptogein, polyamines were not degraded but secreted into culture medium and hydrogen peroxide was scarcely produced, with a concomitant suppression of cell death. Activities of mitogen-activated protein kinases (wound- and salicylic acid-induced protein kinases) were also suppressed, indicating that phosphorylation cascade is involved in polyamine oxidation-derived cell death. These results suggest that polyamine oxidase is a key element for the oxidative burst, which is essential for induction of programmed cell death, and that mitogen-activated protein kinase is one of the factors that mediate this pathway. |
| [33] | In the incompatible interaction between the barley cultivar Hart ( mlo allele) and the powdery mildew fungus Blumeria graminis DC. Speer f. sp. hordei Marchal, free spermidine and conjugated forms of putrescine and spermidine were found to accumulate 1鈥3 days following inoculation. The increase in free spermidine could be accounted for by increased activities of the biosynthetic enzymes ornithine decarboxylase (EC 4.1.1.17) and S-adenosylmethionine decarboxylase (EC 4.1.1.50). Activities of the transferase enzymes putrescine hydroxycinnamoyl transferase (EC 2.3.1.-) and tyramine feruloyl-CoA transferase (EC 2.3.1.-) were also increased in leaves of Hart inoculated with powdery mildew, as were activities of the polyamine catabolic enzymes diamine oxidase (DAO, EC 1.4.3.6) and polyamine oxidase (PAO, EC 1.4.3.4). The possible role of conjugated polyamines, and increases in DAO and PAO, in penetration based resistance to powdery mildew infection is discussed. |
| [34] | Wounding chickpea (Cicer arietinum) internodes or cotyledons resulted in an increase in the steady-state level of copper amine oxidase (CuAO) expression both locally and systemically. Dissection of the molecular mechanisms controlling CuAO expression indicated that jasmonic acid worked as a potent inducer of the basal and wound-inducible CuAO expression, whereas salicylic acid and abscisic acid caused a strong reduction of the wound-induced CuAO expression, without having any effect on the basal levels. Epicotyl treatment with the CuAO mechanism-based inhibitor 2-bromoethylamine decreased hydrogen peroxide (H2O2) levels in all the internodes, as evidenced in vivo by 3,3鈥-diaminobenzidine oxidation. Moreover, inhibitor pretreatment of wounded epicotyls resulted in a lower accumulation of H2O2both at the wound site and in distal organs. In vivo CuAO inhibition by 2-bromoethylamine after inoculation of resistant chickpea cv Sultano with Ascochyta rabiei resulted in the development of extended necrotic lesions, with extensive cell damage occurring in sclerenchyma and cortical parenchyma tissues. These results, besides stressing the fine-tuning by key signaling molecules in wound-induced CuAO regulation, demonstrate that local and systemic CuAO induction is essential for H2O2production in response to wounding and indicate the relevance of these enzymes in protection against pathogens. |
| [35] | . Screening immediate-early responding genes during the hypersensitive response (HR) against tobacco mosaic virus infection in tobacco (Nicotiana tabacum) plants, we identified a gene encoding ornithine decarboxylase. Subsequent analyses showed that other genes involved in polyamine biosynthesis were also up-regulated, resulting in the accumulation of polyamines in apoplasts of tobacco mosaic virus-infected leaves. Inhibitors of polyamine biosynthesis, 伪-difluoromethyl-ornithine, however, suppressed accumulation of polyamines, and the rate of HR was reduced. In contrast, polyamine infiltration into a healthy leaf induced the generation of hydrogen peroxide and simultaneously caused HR-like cell death. Polyamine oxidase activity in the apoplast increased up to 3-fold that of the basal level during the HR, and its suppression with a specific inhibitor, guazatine, resulted in reduced HR. Because it is established that hydrogen peroxide is one of the degradation products of polyamines, these results indicate that one of the biochemical events in the HR is production of polyamines, whose degradation induces hydrogen peroxide, eventually resulting in hypersensitive cell death. |
| [36] | . The hypersensitive response (HR) is a powerful resistance system that plants have developed against pathogen attack. There are two major pathways for HR induction; one is through recognition of the pathogen by a specific host protein, and is known as the host HR. The other is through common biochemical changes upon infection鈥攖he nonhost HR. We previously demonstrated that hydrogen peroxide derived from polyamine degradation by polyamine oxidase triggers the typical host HR in tobacco plants upon infection with tobacco mosaic virus. However, it remains to be determined whether or not polyamines are involved in the nonhost HR in tobacco, and in the host HR in other plant species. When tobacco plants were infected with Pseudomonas cichorii , a representative nonhost pathogen, transcripts for six genes encoding enzymes for polyamine metabolism were simultaneously induced, and polyamines were accumulated in apoplasts. Hydrogen peroxide was concomitantly produced and hypersensitive cell death occurred at infected sites. Silencing of polyamine oxidase by the virus-induced gene silencing method resulted in suppression of hydrogen peroxide production and in disappearance of visible hypersensitive cell death with an increase in bacterial growth. Our results indicated that polyamines served as the source of hydrogen peroxide during the nonhost HR in tobacco plants. Further analysis revealed that polyamines were accumulated in apoplasts of Arabidopsis thaliana infected with Pseudomonas syringae , and of rice infected with Magnaporthe grisea , both causing the typical host HR. As in tobacco, it is conceivable that the same mechanism operates for nonhost HR in these plants. Our present observations thus suggested that polyamines are commonly utilized as the source of hydrogen peroxide during host- and nonhost HRs in higher plants. |
| [37] | . Plant diseases can drastically abate the crop yields as the degree of disease outbreak is getting severe around the world. Therefore, plant disease management has always been one of the main objectives of any crop improvement program. Plant disease resistance (R) genes have the ability to detect a pathogen attack and facilitate a counter attack against the pathogen. Numerous plant R-genes have been used with varying degree of success in crop improvement programs in the past and many of them are being continuously exploited. With the onset of recent genomic, bioinformatics and molecular biology techniques, it is quite possible to tame the R-genes for efficiently controlling the plant diseases caused by pathogens. This review summarizes the recent applications and future potential of R-genes in crop disease management. |
| [38] | . |
| [39] | . The ubiquitous antioxidant thiol tripeptide glutathione is present in millimolar concentrations in plant tissues and is regarded as one of the major determinants of cellular redox homeostasis. Recent research has highlighted a regulatory role for glutathione in influencing the expression of many genes important in plants' responses to both abiotic and biotic stress. Therefore, it becomes important to consider how glutathione levels and its redox state are influenced by environmental factors, how glutathione is integrated into primary metabolism and precisely how it can influence the functioning of signal transduction pathways by modulating cellular redox state. This review draws on a number of recent important observations and papers to present a unified view of how the responsiveness of glutathione to changes in photosynthesis may be one means of linking changes in nuclear gene expression to changes in the plant's external environment. |
| [40] | . The octadecanoid pathway metabolite jasmonic acid (JA) plays a vital role in rice ( Oryza sativa L. cv. Nipponbare) defense/stress response(s). However, genes involved in its biosynthesis remain unidentified. Here, we cloned a novel rice cDNA highly homologous to the allene oxide synthase (EC 4.2.1.92) AOS gene, the first committed step in JA biosynthesis, showing significant similarity at the amino acid level with a related monocotyledoneous barley AOS. OsAOS is a novel member of the cytochrome P450 CYP74A subfamily and exists as a single copy gene in the rice genome. An examination of its steady state mRNA level in two-week-old seedling leaves revealed that OsAOS does not express constitutively in healthy leaves, and shows a weak responsiveness to cut. Signaling components of defense/stress pathways, in particular JA itself, strongly up-regulated the OsAOS transcript, whereas salicylate, ethylene, abscisic acid, and hydrogen peroxide were not so effective. Copper, a heavy metal also significantly enhanced the OsAOS expression. Protein phosphatase inhibitors proved to be the most potent in up-regulating the OsAOS mRNA level, suggesting the involvement of phosphorylation/dephosphorylation events in its regulation. Moreover, the inducible nature of OsAOS was influenced by light signal(s). Blast pathogen ( Magnaporthe grisea) specifically elicited the accumulation of OsAOS mRNA in leaves in an incompatible versus compatible interaction, a first demonstration of pathogen responsiveness for any AOS gene to date. These results strongly suggest the importance of OsAOS in rice defense/stress response pathway(s). |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}