0 引言
【研究意义】单倍体育种是现代育种的一项重要内容,单倍体植株经诱变后再进行染色体加倍形成双单倍体纯系,可以缩短育种周期,丰富种质资源[1]。花药培养是人工诱导产生单倍体的有效途径,目前已在小麦、辣椒、甜橙等多种植物上广泛应用,并成功与杂交育种、转基因相结合形成一套较为完整的育种技术体系[2,3,4]。苹果为基因组高度杂合木本植物,童期长,加上自交不亲和,导致通过多代自交获得纯合植株的方法难以实现[5]。苹果花药培养获得具有配子体来源的纯合基因型植株,该植株目标性状稳定,便于遗传分析,提高了选择效率[6]。同时,苹果花药培养获得的每个株系均来自不同的花粉粒,因此具有独一性、唯一性,是进行杂交育种、遗传分析的重要材料[7]。来自不同花粉粒的株系,其植物学特征、生物学特性,以及诱导培养、生根的条件等都有差异,因此对每个株系试管苗的倍性、生长特性、特征、生根培养条件及试管苗生根移植到田间后的性状进行观察分析,对苹果新种质的创新具有重要意义。【前人研究进展】苹果花药培养开始于20世纪70年代末。1990年中国农业科学院果树研究所相继获得10个品种的胚状体,以及‘元帅’‘赤阳’‘祝光’‘国光’‘富士’‘红玉’‘新红星’‘金冠’8个花药培养的再生植株,经染色体观察和同工酶分析证明来源于花粉[8]。ZHANG等[9]以‘红星’苹果花药为试材,发现在苹果花药培养诱导胚状体形成的过程中,花药中的小孢子在经过一定时间的低温诱导后才能获得胚性潜能。温鑫等[10]用SSR标记对30个‘嘎啦’苹果花药培养再生株系进行基因型鉴定,经检测全部株系为纯合基因型。H?FER等[11,12,13]经多年研究,获得单倍体来源的苹果植株共100多株,并证明通过苹果花药培养获得的再生植株,与供体相比,其结实率低,形态特征发生了变化。【本研究切入点】苹果营养丰富、口感爽脆,是中国果实产量最高的经济树种,中国的苹果主栽品种多数由外国引进[14]。近年来,国内外关于苹果花药培养的研究不断深入,但关于苹果花药培养获得的纯合基因型株系表型方面的鉴定、倍性分析,以及生根条件的筛选尚未得到大量、有效的重复验证,更没有后续相关研究报道。【拟解决的关键问题】本研究对‘嘎啦’‘富士’‘红星’的纯合基因型株系进行倍性分析,筛选其最适的生根条件,并对32个株系进行表型多样性分析,进一步选育优良的苹果新种质。1 材料与方法
1.1 植物材料
2013—2016年每年4月初,采集山西省农业科学院果树所苹果基地的‘红星’‘富士’‘嘎啦’苹果(Malus domestica Borkh.)的健壮花蕾,经过苹果花药培养获得再生植株。本研究以花药培养再生植株为材料[15,16],经SSR鉴定均为单倍体来源[10],其中包括‘红星’7株、‘富士’5株、‘嘎啦’20株(表1中DH0:‘红星’;DH1:‘富士’;DH2:‘嘎啦’)。1.2 再生植株倍性分析
取2—3片0.5 cm2左右再生植株嫩叶置于预冷的干净培养皿中,加入1 mL WBP细胞裂解液,用刀片快速切碎后静置1—3 min,经直径50 μm滤网过滤于EP管中,滤液中加入核糖核酸酶和荧光染料(propidium iodide,PI),浓度为50 μm?mL-1,4℃静置20 min。利用流式细胞仪(BD C6)进行倍性分析。以杂合二倍体‘嘎啦’试管苗叶片为对照。1.3 生根培养基的筛选
取生长旺盛的继代组培苗为材料,当再生植株不定芽长度为3 cm左右时,对其剪切并转移至添加不同IBA浓度的培养基中进行不定根诱导处理,培养基为1/2 MS+2 %蔗糖+0.6 %琼脂,IBA浓度设3个处理,分别为1 mg·L-1(T1)、2 mg·L-1(T2)、3 mg·L-1(T3)。置于25℃温室光照培养(光照强度3 000 lx,光照时间14 h/d)。以上共3个处理,每个处理15株。40 d后开始统计每个处理的生根率(生根株数/总株数),并从每个处理中选取根系较发达的3株测量植株主根数和平均根长,以筛选适宜苹果纯合基因型株系生根的激素浓度。1.4 生根试管苗的性状观察
将苹果纯合基因型株系接入生根培养基后从生根培养的3个处理中选取叶片充分展开,根系长势较好的3株,在40 d后观察统计试管苗生长状态、长势、叶色、叶片形状等。测量试管苗叶片的叶长度、叶宽度、叶柄长度,计算叶形指数(叶长度/叶宽度)。测量植株主根数、根长度和株高。观察生长情况并拍照,分析不同株系的性状差异。观察记载方法参照《果树种质资源描述符—记载项目及评价标准》[17]。1.5 生根苗驯化、移栽
生根培养40 d后每个株系选取10株进行驯化移栽。置于室内自然光照下培养5—7 d后,去掉瓶盖加入自来水10 mL左右,炼苗3 d。将组培苗的根部残留培养基用清水冲洗干净,移栽到含有草炭、蛭石、珍珠盐的花盆中(比例为1﹕1﹕1),在花盆上盖一层保鲜膜后放入光照培养箱(光照培养:12 h、25℃,暗培养:12 h、21℃),3周后转移至温室条件进行培养。1.6 数据统计分析
采用 Microsoft Excel 2010软件进行数据整理和统计;用SPSS 20.0 对数据进行单因素方差分析,并使用Duncan法对不同处理进行多重分析,P≤0.05认为具有显著性差异,图表中所有数据均为3个重复的平均数±标准差(Mean±SE)[18]。2 结果
2.1 苹果纯合基因型株系倍性分析
对‘嘎啦’‘富士’‘红星’的32个苹果纯合基因型株系进行倍性分析。结果表明,有单倍体、二倍体、三倍体和四倍体4种不同的倍性(表1、图1)。其中单倍体(n)1株、二倍体(2n)27株、三倍体(3n)1株、四倍体(4n)3株,二倍体占84.38%,说明苹果纯合基因型株系的主要倍性为二倍体。‘嘎啦’‘富士’‘红星’苹果纯合基因型株系的倍性分化率具有差异性,其中‘红星’苹果纯合基因型株系的倍性分化率最大,为28.57%,‘富士’‘嘎拉’苹果纯合基因型株系的倍性分化率分别为20%和10%。Table 1
表1
表1‘红星’‘富士’‘嘎啦’苹果纯合基因型株系倍性
Table 1‘Red Star’, ‘Fuji’, ‘Gala’ apple homozygous genotype lines ploidy analysis results
| 红星株系 Red star strain | 倍性 Ploidy | 富士株系 Fuji strain | 倍性 Ploidy | 嘎啦株系 Gala strains | 倍性 Ploidy | 嘎啦株系 Gala strains | 倍性 Ploidy | 嘎啦株系 Gala strains | 倍性 Ploidy | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DH0-1 | 2n | DH1-1 | 2n | DH2-1 | 2n | DH2-8 | 2n | DH2-15 | 4n | ||||
| DH0-2 | 2n | DH1-2 | 4n | DH2-2 | 2n | DH2-9 | 2n | DH2-16 | 2n | ||||
| DH0-3 | 2n | DH1-3 | 2n | DH2-3 | 1n | DH2-10 | 2n | DH2-17 | 2n | ||||
| DH0-4 | 2n | DH1-4 | 2n | DH2-4 | 2n | DH2-11 | 2n | DH2-18 | 2n | ||||
| DH0-5 | 4n | DH1-5 | 2n | DH2-5 | 2n | DH2-12 | 2n | DH2-19 | 2n | ||||
| DH0-6 | 2n | DH2-6 | 2n | DH2-13 | 2n | DH2-20 | 2n | ||||||
| DH0-7 | 3n | DH2-7 | 2n | DH2-14 | 2n |
新窗口打开
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1流式细胞仪对再生植株倍性分析
-->Fig. 1Flow cytometric (FCM) analysis of ploidy level of regenerated plantlets
-->
2.2 苹果纯合基因型株系生根培养
2.2.1 ‘红星’苹果纯合基因型株系生根培养 由表2可知,与T1培养基相比,T2、T3培养基的生根率较高,T1培养基生根率为6.67%—70%,而T2、T3培养基生根率均为53.33%—100%。从根长来看,IBA浓度对大多数‘红星’纯合基因型株系的根长影响不明显,DH0-3株系在T2培养基根长最长,为20.83 cm,在T3和T1培养基的最大根长分别为20.13 cm和18.13 cm(图2-A)。分析7个株系的生根数,生根数最多的培养基主要为T2或T3培养基,DH0-7株系在T3培养基中生根数为22.33条,而在T1培养基中仅为1条。说明IBA浓度不足显著降低了生根率和根数,根长也有所减小,但影响不显著。综上所述,从生根率、生根数、根的长度来看,‘红星’苹果纯合基因型株系以1/2 MS培养基中添加2—3 mg L-1IBA(T2、T3培养基)为宜。其中,T2培养基中生根率达到最高的有4个株系,略好于T3培养基。Table 2
表2
表2不同IBA浓度对‘红星’苹果纯合基因型株系根系形态的影响
Table 2Effects of different IBA concentrations on root morphology of 'Red Star' apple homozygous lines
| 根系形态 Root morphology | 培养基 Medium | 株系 Line | ||||||
|---|---|---|---|---|---|---|---|---|
| DH0-1 | DH0-2 | DH0-3 | DH0-4 | DH0-5 | DH0-6 | DH0-7 | ||
| 生根率 Root rate (%) | T1 | 26.67 | 6.67 | 70 | 46.67 | 33.33 | 46.67 | 33.33 |
| T2 | 53.33 | 86.67 | 100 | 93.33 | 86.67 | 66.67 | 100 | |
| T3 | 73.33 | 73.33 | 93.33 | 93.33 | 100 | 53.33 | 66.67 | |
| 根长 Root length (cm/plant) | T1 | 5.70±0.81a | 4.80b | 18.13±1.53a | 3.70±1.16a | 3.50±0.58b | 3.50±0.79a | 3.90±0.56a |
| T2 | 3.87±1.13a | 6.63±1.19a | 20.83±1.66a | 3.73±0.77a | 4.63±0.91b | 3.17±0.77a | 3.93±0.67a | |
| T3 | 4.60±0.71a | 4.50±0.67b | 20.13±1.51a | 4.07±1.13a | 8.33±1.17a | 3.13±0.90a | 4.27±0.95a | |
| 根数 Root number | T1 | 7.00±1.26b | 3.00b | 5.00±1.00b | 2.33±1.05c | 3.33±0.83b | 3.33±1.32b | 1.00c |
| T2 | 4.67±1.32b | 5.00±1.00a | 10.00±1.26a | 15.33±1.28a | 5.67±1.05a | 9.00±1.00a | 22.33±1.59a | |
| T3 | 17.67±1.56a | 1.33±0.83c | 7.00±1.26ab | 9.33±0.83b | 6.33±1.15a | 8.67±0.83a | 12.33±1.45b | |
新窗口打开
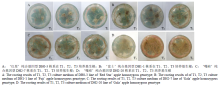 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2‘红星’‘富士’‘嘎啦’苹果纯合基因型株系生根情况
-->Fig. 2The root of ‘Red Star’, ‘Fuji’, ‘Gala’ apple homozygous genotype lines
-->
2.2.2 ‘富士’苹果纯合基因型株系生根培养 如表3所示,从生根率来看,‘富士’纯合基因型株系在T2培养基生根率最高,T1培养基生根率为0—53.33%,T2培养基生根率为26.67%—100%,T3培养基生根率为6.67%—86.67%。分析所有株系的根长和根数,在T2、T3培养基的根长和根数显著高于T1培养基(P<0.05),DH1-3株系在T2、T3培养基的根长分别为21.00 cm和18.37 cm,而在T1培养基的根长仅为14.00 cm;DH1-1株系在T1、T2、T3培养基的根数分别为3.00、18.33、17.00条(图2-B)。表明激素浓度明显影响‘富士’纯合基因型株系的生根率、根长和根数,‘富士’纯合基因型株系在添加2—3 mg L-1 IBA的培养基中(T2、T3培养基),根的长势最好,其中,T2培养基好于T3培养基。
Table 3
表3
表3不同IBA浓度对‘富士’苹果纯合基因型株系根系形态的影响
Table 3Effects of different IBA concentrations on root morphology of 'Fuji' apple homozygous
| 根系形态 Root morphology | 培养基 Medium | 株系 Line | ||||
|---|---|---|---|---|---|---|
| DH1-1 | DH1-2 | DH1-3 | DH1-4 | DH1-5 | ||
| 生根率 Root rate(%) | T1 | 53.33 | 13.33 | 26.67 | _ | _ |
| T2 | 100 | 60 | 100 | 40 | 26.67 | |
| T3 | 86.67 | 73.33 | 66.67 | 6.67 | 53.33 | |
| 根长 Root length (cm/plant) | T1 | 9.53±1.21b | 8.40±1.37b | 14.00±1.20b | _ | _ |
| T2 | 17.43±1.00a | 19.67±1.41a | 21.00±1.15a | 4.47±1.31a | 4.17±1.21a | |
| T3 | 8.57±1.26b | 22.27±1.63a | 18.37±1.21a | 4.5a | 7.30±1.48a | |
| 根数 Root number | T1 | 3.00b | 5.00±1.00b | 2.67±1.28c | _ | _ |
| T2 | 18.33±1.48a | 2.00±1.00b | 26.67±1.65a | 4.00±1.38a | 3.00±1.20a | |
| T3 | 17.00±1.26a | 12.33±1.28a | 18.33±1.15b | 3a | 1.33±0.83a | |
新窗口打开
2.2.3 ‘嘎啦’苹果纯合基因型株系生根培养基分析 表4表明,‘嘎啦’纯合基因型株系的生根率高达100%,其中DH2-7株系在3个培养基均为100%(图2-C),DH2-5、DH2-13、DH2-18在T2、T3培养基中生根率一样,为100%;DH2-2株系在T1、T2培养基中的生根率最高,为100%。除此之外,其余‘嘎啦’纯合基因型株系在T1培养基的生根率低于T2、T3培养基,T2培养基的生根率略好于T3培养基。DH2-1、DH2-8、DH2-9、DH2-15、DH2-19、DH2-20株系在T1、T2、T3培养基的根长无显著差别(P>0.05),而其他株系在T1培养基的根长显著低于T2或T3培养基(P<0.05),且T2、T3培养基的根长基本无差别,其中,DH2-11株系在T2、T3培养基的根长为20.70 cm,而在T1培养基中仅为14.50 cm。分析20个株系在3个培养基中的生根数,生根数最多的培养基主要为T2或T3培养基,T2培养基的生根数略好于T3培养基。DH2-20株系在T2培养基中生根数为86.67条,而在T1培养基中仅为27条(图2-D)。综上所述,激素浓度是‘嘎啦’纯合基因型株系生根率、生根数及生根长度的限制因子之一,在生根诱导培养基中添加2—3 mg·L-1的生根激素(T2、T3培养基),生根状态最好。
Table 4
表4
表4不同IBA浓度对‘嘎啦’苹果纯合基因型株系根系形态的影响
Table 4Effects of different IBA concentrations on root morphology of ‘Gala’ apple homozygous lines
| 根系形态 Root morphology | 培养基 Medium | 株系 Line | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| DH2-1 | DH2-2 | DH2-3 | DH2-4 | DH2-5 | DH2-6 | DH2-7 | DH2-8 | DH2-9 | DH2-10 | ||
| 生根率 Root rate (%) | T1 | 6.67 | 100 | 20 | _ | 6.67 | 80 | 100 | 53.33 | 66.67 | 20 |
| T2 | 60 | 100 | 66.67 | 100 | _ | 100 | 100 | 73.33 | 86.67 | 60 | |
| T3 | 13.33 | 90 | 80 | 86.67 | 13.33 | 100 | 100 | 73.33 | 86.67 | 86.67 | |
| 根长 Root length (cm.plant-1) | T1 | 4.65±0.97a | 1.70±0.67b | 2.93±0.49b | _ | 5.9a | 4.27±0.88b | 0.57±0.39b | 32.60±1.05a | 9.50±1.25a | 7.53±1.31b |
| T2 | 6.43±1.47a | 1.63±0.78b | 1.87±1.19b | 29.43±1.80a | _ | 8.90±1.07a | 2.50±1.02a | 30.00±1.10a | 8.93±1.25a | 8.23±1.76b | |
| T3 | 1.5a | 5.77±1.17a | 13.17±1.20a | 14.83±1.40b | 5.85±1.18a | 9.40±1.15a | 3.67±0.71a | 31.33±1.28a | 10.93±1.54a | 22.10±1.31a | |
| 根数 Root number | T1 | 1.50±0.89b | 24.67±1.65b | 3.67±1.15a | _ | 1a | 9.33±0.83c | 16c | 9.00±1.20b | 5.67±1.45a | 1.33±0.83b |
| T2 | 17.00±1.66a | 39.67±1.67ab | 1.67±0.83a | 4.00±1.38a | _ | 42.33±1.82a | 29.33±1.83b | 21.67±1.61a | 9.67±1.45a | 5.00±1.20b | |
| T3 | 1b | 51.67±2.42a | 2.33±0.83a | 6.00±1.38a | 1a | 34.67±1.28b | 43.67±1.73a | 8.00±1.59b | 12.00±1.74a | 22.33±1.86a | |
| 根系形态 Root morphology | 培养基 Medium | 株系 Line | |||||||||
| DH2-11 | DH2-12 | DH2-13 | DH2-14 | DH2-15 | DH2-16 | DH2-17 | DH2-18 | DH2-19 | DH2-20 | ||
| 生根率 Root rate(%) | T1 | 66.67 | 20 | 80 | 6.67 | 53.33 | _ | 6.67 | 93.33 | 73.33 | 6.67 |
| T2 | 100 | 100 | 100 | 73.33 | 66.67 | 20 | 53.33 | 100 | 73.33 | 86.67 | |
| T3 | 93.33 | 53.33 | 100 | 60 | 53.33 | _ | 13.33 | 100 | 93.33 | 66.67 | |
| 根长 Root length (cm.plant-1) | T1 | 14.5±1.62b | 12.7±1.26b | 1.33±0.69c | 4.8b | 14.27±1.46a | _ | 4.5b | 7.60±1.30c | 13.43±1.54a | 13.8a |
| T2 | 20.7±1.40a | 35.27±1.78a | 3.83±1.18b | 17.33±1.63a | 10.23±1.25a | 3.43±1.02 | 11.77±1.40a | 16.10±1.72b | 14.43±1.38a | 18.40±1.57a | |
| T3 | 20.7±1.40a | 23.2±1.90b | 6.27±0.96a | 6.40±1.36b | 9.80±1.26a | _ | 3.20±1.46b | 23.83±1.42a | 11.07±1.33a | 14.77±1.48a | |
| 根数 Root number | T1 | 3.00±1.20b | 2.33±0.83b | 11.67±0.83b | 1a | 13.67±1.36a | _ | 1b | 5.00±1.00b | 3.33±1.15b | 27c |
| T2 | 10.33±1.36a | 11.33±1.15a | 18.67±1.59ab | 6.33±1.36a | 7.00±1.63b | 1.33±0.83 | 2.67±0.83a | 8.33±1.67ab | 3.67±1.15b | 86.67±2.05a | |
| T3 | 5.33±1.59ab | 3.00±1.20b | 22.33±1.71a | 5.33±1.48a | 5.00±1.26b | _ | 1.50±0.89ab | 14.33±1.61a | 16.00±1.53a | 59.67±2.50b | |
新窗口打开
2.3 苹果纯合基因型株系性状分析
2.3.1 ‘红星’苹果纯合基因型株系性状分析 如表5所示,‘红星’纯合基因型株系DH0-2、DH0-5株系叶数较少,分别为12.33和15.00,DH0-1株系的叶数最多,为27.00;DH0-1、DH0-4、DH0-7株系的根数显著较高(P<0.05),为17.67—22.33条,而其余株系根数仅为4.33—11.00条。从根长来看,DH0-3株系根长最大,为18.17 cm(图3-A),其他株系根长最长为5.73 cm(DH0-2株系,图3-B);‘红星’纯合基因型株系的株高差异不明显;DH0-1、DH0-3、DH0-4、DH0-7株系的叶柄较长,为0.77—0.87 cm。表明‘红星’纯合基因型的大部分株系性状较好,其中DH0-1、DH0-3、DH0-4、DH0-7株系的综合性状明显好于其他株系,且4个株系的叶形指数差异不显著,说明它们的叶片形状无显著差别。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3苹果纯合基因型株系性状
-->Fig. 3Plants traits of apple homozygous genotype lines
-->
Table 5
表5
表5‘红星’苹果纯合基因型株系性状
Table 5Plant traits of ‘Red Star’ apple homozygous genotype
| 株系 Line | 叶数 Leave number | 根数 Root number | 根长 Root length (cm) | 株高 Plant height (cm) | 叶柄长 Petiole length (cm) | 叶形指数 Leaf shape index |
|---|---|---|---|---|---|---|
| DH0-1 | 27.00±1.00a | 17.67±1.56a | 4.27±0.91a | 3.70±0.91ab | 0.83±0.49a | 1.66±0.52b |
| DH0-2 | 12.33±1.45b | 4.33±1.28c | 5.73±1.15b | 2.37±0.80c | 0.5±0.46c | 2.07±0.49a |
| DH0-3 | 22.33±1.28a | 10.67±1.28b | 18.17±1.89a | 4.13±0.80a | 0.80±0.46a | 1.60±0.51b |
| DH0-4 | 26.67±1.71a | 18.67±1.56a | 3.17±0.85b | 3.47±0.71abc | 0.87±0.39a | 1.74±0.54b |
| DH0-5 | 15.00±1.00b | 8.00±1.00bc | 3.53±0.91b | 2.57±0.86bc | 0.63±0.39bc | 1.71±0.37b |
| DH0-6 | 26.00±1.84a | 11.00±1.38b | 2.87±0.49b | 4.57±0.49a | 0.7±0.46b | 1.68±0.59b |
| DH0-7 | 22.67±1.28a | 22.33±1.59a | 2.67±1.08b | 3.67±1.01ab | 0.77±0.39a | 1.88±0.67ab |
新窗口打开
2.3.2 ‘富士’苹果纯合基因型株系性状分析 从表6可以看出,‘富士’苹果纯合基因型株系叶数差异较小,为15.33—22.67片;比较所有株系的根数,DH1-3株系根数显著最多(P<0.05),为26.00条;从根长来看,DH1-2、DH1-3株系根长为19.00—22.27 cm,而其他株系仅为4.43—7.93 cm;‘富士’苹果纯合基因型DH1-5株系的株高最高,为3.33 cm(图3-C),DH1-1株系的株高仅为1.97 cm(图3-D);分析叶柄长度,DH1-2、DH1-3株系的叶柄较长,分别为1.53和1.17 cm。表明‘富士’纯合基因型DH1-3株系的综合性状好,叶形指数较小,叶片形状呈阔圆形。
Table 6
表6
表6‘富士’苹果纯合基因型株系性状
Table 6Plant traits of ‘Fuji’ apple homozygous genotype
| 株系 Line | 叶数 Leave number | 根数 Root number | 根长 Root length(cm) | 株高 Plant height (cm) | 叶柄长 Petiole length (cm) | 叶形指数 Leaf shape index |
|---|---|---|---|---|---|---|
| DH1-1 | 15.33±1.32b | 17.67±1.56b | 7.93±0.87b | 1.97±0.67d | 0.83±0.39bc | 1.90±0.63ab |
| DH1-2 | 16.00±1.38b | 12.33±1.28c | 22.27±1.63a | 2.23±0.63cd | 1.53±0.77a | 1.61±0.41b |
| DH1-3 | 17.67±1.15ab | 26.00±1.53a | 19.00±1.56a | 3.07±0.69ab | 1.17±0.53ba | 1.58±0.73b |
| DH1-4 | 22.67±1.28a | 4.00±1.38d | 4.47±1.32b | 2.57±0.59bc | 0.87±0.39bc | 2.12±0.55a |
| DH1-5 | 22.33±1.65a | 1.33±0.83d | 4.43±1.55b | 3.33±0.72a | 0.70±0.58c | 1.52±0.57b |
新窗口打开
2.3.3 ‘嘎啦’苹果纯合基因型株系性状分析 表7表明,‘嘎啦’苹果纯合基因型DH2-5株系叶数最少,为11.33片,DH2-4株系叶数最多,为36.67片;就根数而言,DH2-20株系根数多达85.67条(图3-E),DH2-2、DH2-7株系根数少于DH2-20株系,但显著多于其他株系(P<0.05),分别为41.67和43.67条;从根长来看,DH2-7株系根长最大,为35.27 cm(图3-F),而其他株系的根长最大仅为22.10 cm(DH2-10株系);比较株高,DH2-2株系株高显著最高,为5.57 cm(图3-G),DH2-8株系株高显著最矮,为1.67 cm(图3-H);‘嘎啦’苹果纯合基因型株系叶柄长度无明显差别。综上所述,‘嘎啦’纯合基因型DH2-2、DH2-4、DH2-7、DH2-20株系的综合性状好(图3-E—G),相较DH2-4、DH2-7株系,DH2-2、DH2-20株系的叶形指数较大,所以叶片形状呈窄长形。
Table 7
表7
表7‘嘎啦’苹果纯合基因型株系性状
Table 7Plant traits of ‘Gala’ apple homozygous genotype
| 株系 Line | 叶数 Leave number | 根数 Root number | 根长 Root length(cm) | 株高 Plant height | 叶柄长 Petiole length (cm) | 叶形指数 Leaf shape index |
|---|---|---|---|---|---|---|
| DH2-1 | 13.67±1.48gh | 17.00±1.66ef | 6.43±1.47ghi | 1.70±0.46g | 0.70±0.46ef | 2.15±0.60bc |
| DH2-2 | 20.00±1.00defg | 41.67±1.52b | 6.33±1.01ghi | 5.57±1.01a | 1.27±0.59ab | 2.29±0.57b |
| DH2-3 | 32.33±1.83abc | 3.67±1.15hij | 13.17±1.20f | 3.17±0.71d | 1.0±0.67bcde | 3.07±0.68a |
| DH2-4 | 36.67±2.30a | 5.00±1.00ghij | 27.17±2.08bc | 2.97±0.86de | 1.33±0.39a | 1.74±0.34defg |
| DH2-5 | 11.33±1.36h | 1j | 5.87±1.05ghi | 1.70±0.70g | 0.57±0.39f | 1.69±0.75efg |
| DH2-6 | 21.33±1.28defg | 35.67±1.15c | 9.97±0.98fgh | 3.20±0.71d | 0.80±0.64cdef | 2.21±0.45b |
| DH2-7 | 18.67±1.28defgh | 43.67±1.72b | 35.27±1.78a | 5.10±0.58ab | 1.10±0.46abc | 2.07±0.51bcd |
| DH2-8 | 20.67±1.93defg | 21.67±1.61de | 3.67±0.71i | 1.67±0.53g | 0.80±0.64cdef | 1.92±0.68bcdef |
| DH2-9 | 19.33±1.45defgh | 8.00±1.26ghi | 11.67±1.40fg | 4.10±0.46c | 0.83±0.53cdef | 1.63±0.56fg |
| DH2-10 | 20.00±1.53defg | 5.00±1.20ghij | 22.10±1.31cd | 2.20±0.64fg | 0.83±0.49cdef | 1.92±0.54bcdef |
| DH2-11 | 17.33±1.05efgh | 10.00±1.44gh | 12.53±1.31f | 2.90±0.64de | 0.90±0.46cde | 1.40±0.45g |
| DH2-12 | 27.00±1.53bcd | 11.33±1.15fg | 19.63±1.36de | 2.90±0.64de | 0.73±0.53def | 1.58±0.47fg |
| DH2-13 | 21.67±1.05defg | 24.67±1.28d | 4.93±1.47hi | 1.77±0.53g | 0.70±0.46ef | 2.21±0.37b |
| DH2-14 | 25.00±1.38cde | 6.33±1.36ghij | 9.17±1.46fghi | 2.53±0.53def | 0.83±0.63cdef | 2.12±0.59bc |
| DH2-15 | 20.67±1.36defg | 4.00±1.00hij | 14.89±1.28ef | 2.47±0.39ef | 0.93±0.39cde | 1.80±0.55cdef |
| DH2-16 | 15.33±1.36fgh | 1.33±0.83j | 3.43±1.02i | 3.17±0.59d | 1.03±0.39bcd | 2.29±0.59b |
| DH2-17 | 21.00±1.26defg | 2.67±0.83ij | 11.77±1.40fg | 2.40±0.56ef | 0.83±0.49cdef | 2.11±0.39bc |
| DH2-18 | 34.33±1.78ab | 11.00±1.20fg | 9.87±1.03fgh | 3.07±0.67de | 0.80±0.58cdef | 2.07±0.49bcd |
| DH2-19 | 25.00±1.38cde | 11.33±1.68fg | 11.17±1.31fg | 3.17±0.71d | 0.90±0.46cde | 2.00±0.44bcde |
| DH2-20 | 23.67±1.61def | 85.67±2.16a | 30.00±1.10b | 4.60±0.67bc | 1.03±0.53bcd | 2.21±0.53b |
新窗口打开
2.4 苹果不同倍性纯合基因型株系叶片形态特征
叶片形态特征可被用作鉴定植物倍性水平的指示标记(如茎的粗度、叶色、叶缘、叶片形状和叶形指数等)[19,20]。本研究中,苹果纯合基因型株系的单倍体、多倍体变异株系和二倍体株系在叶片形态特征上表现出明显差异。苹果纯合基因型单倍体、多倍体株系叶片形态特征表现出了表型变异的多样性。叶形指数在单倍体、二倍体和多倍体之间表现差异显著(表5—7),单倍体DH2-3株系的叶形指数(3.07)显著大于其他倍性株系,二倍体的叶形指数为1.40—2.29,三倍体DH0-7株系、四倍体DH0-5株系的叶形指数分别为1.88和1.71。二倍体为椭圆形(图4-A),单倍体的叶片形状为狭椭圆形(图4-B),多倍体为广椭圆形(图4-C—F)。单倍体DH2-3株系叶片呈窄长形(图4-B),而三倍体、四倍体叶片呈阔圆形(图4-C—F)。与二倍体DH2-20株系相比,单倍体叶基更窄,叶柄较细(图4-B);而多倍体株系的叶基变得更阔,叶柄较粗(图4-C—F)。不同倍性株系叶色、叶缘锯齿的变异也表现出多样性,单倍体的叶色、叶缘锯齿较浅(图4-B),多倍体的叶色、叶缘锯齿一般变深(图4-C—F)。这一结果表明叶色、叶片形状、叶形指数、叶缘变异及叶柄长度都可用做苹果纯合基因型株系不同倍体变异早期选择的有效指示标记。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4苹果纯合基因型株系不同倍性叶片性状
-->Fig. 4Leaf traits of different ploidy in apple homozygous genotype lines
-->
2.5 生根苗驯化、移栽
经移栽后,‘红星’纯合基因型DH0-3、DH0-4株系,‘富士’纯合基因型DH1-3株系,‘嘎啦’纯合基因型DH2-2、DH2-4、DH2-6、DH2-7、DH2-11株系成活(图5)。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5‘红星’、‘富士’、‘嘎啦’苹果纯合基因型株系移栽后性状
-->Fig. 5Transplanted plants traits of ‘Red Star’, ‘Fuji’, ‘Gala’ apple homozygous genotype lines
-->
3 讨论
本研究中的32个纯合基因型株系有4种不同的倍性(单倍体、二倍体、三倍体、四倍体),其中主要倍性为二倍体。H?FER等[21]研究发现花药培养获得的再生植株染色体倍数变化多样,在通过花药培养诱导的植株中,87.5%是二倍体,而单倍体、三倍体和四倍体胚胎的发生率非常低。TESTILLANO[22]、GRIGGS[23]等研究表明,再生植株染色体自然加倍现象是由于单倍体细胞发生了核融合或核内加倍。笔者课题组使用SSR标记对花药来源植株鉴定揭示了再生植株100%的纯合性,32个株系均为单倍体来源的再生植株,可以排除二倍体由体细胞发育而来[10,15]。组培过程中,不同的品种、外植体在培养基中添加不同浓度、种类的生长调节物质(如BA、IBA、KT等)对生根率、根长、根数有一定的影响[24,25]。在诱导试管苗生根时,以MS或1/2 MS为基本培养基[26]。DOBRáNSZKI等[27]报道‘皇家嘎啦’苹果在培养基中添加2 mg?L-1 IBA时,生根状况良好;‘旭’苹果在添加1 mg?L-1 IBA的培养基中生根良好;‘Prima’苹果在3 mg?L-1 IBA水平上生根状态良好。谢璇等[28]研究表明,‘红富士’苹果组培苗在添加IBA 1 mg?L-1、蔗糖20 mg L -1的1/2 MS培养基适宜不定根诱导。有关苹果纯合基因型株系生根培养条件的报道较少,本试验表明IBA浓度对‘红星’‘富士’‘嘎啦’苹果纯合基因型株系的生根有影响,以1/2 MS培养基中添加2—3 mg?L-1 生根诱导激素(IBA)为宜。
陈银全等[29]研究表明,由于花粉来源的株系配子体丰富,因此不同的植株性状表现多样化。苹果花药来源不同株系的树种习性和叶片形态有很大差异[13]。与供体叶相比,苹果花药来源植株的叶长、宽度、果实显著变小,花的形态也发生了改变[30,31]。通过对叶片大小和生长习性的测定,苹果花药培养株系比杂合母本植株的活力更低[32,33]。LI等[34]描述了Populus×beijingensis再生系的不同倍性间叶片形态和植物大小的差异。本研究中‘红星’苹果纯合基因型DH0-1、DH0-3、DH0-4、DH0-7株系,‘富士’苹果纯合基因型DH1-3株系,‘嘎啦’苹果纯合基因型DH2-2、DH2-4、DH2-12、DH2-20株系的性状较好。
苹果纯合基因型单倍体、二倍体、多倍体株系的生长形态具有差异性,本研究中单倍体、二倍体、多倍体株系的叶基、叶片形状、叶柄、叶色、叶缘锯齿具有明显差异。除此之外,还有其他性状的变异,如单倍体的叶形变得更窄长,表现为叶形指数增大。从分子机制方面分析,不同倍性株系生长形态多样性可能是由于染色体加倍后引起染色体结构的改变或表观遗传的修饰[35],也可能是因为基因的变化增加了基因表达的多样性而导致表型变异[36,37]。由于材料有限,本研究只对不同倍性的植株表型变异进行了观察分析,但RIDDLE等[38]表明不同种或不同基因型的单倍体、多倍体其表型变异也不同,所以还有待进一步探究。
4 结论
苹果花药培养获得的32个纯合基因型株系中,‘红星’苹果有1个三倍体、1个四倍体;‘富士’苹果有1个四倍体;‘嘎啦’苹果有1个单倍体、1个四倍体,苹果纯合基因型株系的主要倍性为二倍体,且‘红星’苹果纯合基因型株系的倍性分化率最大,为28.57%。单倍体株系叶基窄、叶柄细、叶色和叶缘锯齿较浅,而多倍体株系与之相反。除此之外,还有其他性状的变异,如单倍体的叶形变得更窄长,表现为叶形指数增大。IBA浓度2—3 mg?L-1为苹果纯合基因型株系组培的最适浓度。‘红星’纯合基因型DH0-1、DH0-3、DH0-4、DH0-7株系,‘富士’纯合基因型DH1-3株系,‘嘎啦’纯合基因型DH2-2、DH2-4、DH2-7、DH2-20株系优于同一品种其他株系。‘红星’纯合基因型株系的移栽成活率最高,为28.57%。The authors have declared that no competing interests exist.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | . |
| [2] | . 单倍体在基础研究和育种实践中具有重要的利用价值,而花药培养是生产小麦单倍体的主要方法之一.本文重点从影响小麦花药培养的基因型依赖性、脱分化培养基及其附加成分等主要因素,小麦花药培养在育种实践中的应用,与辐射诱变、远缘杂交和转基因技术的结合等方面展开综述,较为全面地统计了相关研究者所筛选的具有优良花药培养特性的基因型,分析了目前存在的问题并提出了建议,旨在为相关研究提供参考. . 单倍体在基础研究和育种实践中具有重要的利用价值,而花药培养是生产小麦单倍体的主要方法之一.本文重点从影响小麦花药培养的基因型依赖性、脱分化培养基及其附加成分等主要因素,小麦花药培养在育种实践中的应用,与辐射诱变、远缘杂交和转基因技术的结合等方面展开综述,较为全面地统计了相关研究者所筛选的具有优良花药培养特性的基因型,分析了目前存在的问题并提出了建议,旨在为相关研究提供参考. |
| [3] | |
| [4] | . [Objective]In order to increase anther culture efficiency of pepper.[Method]MS culture media and Bolajiaohong were used in this experiment to study the influences of carbon sources and concentrations on anther callus induction of pepper.Jiayu was taken as a material to study influences of plant growth regulators and concentrations on anther callus induction of pepper according to L16(45) orthogonal design.[Result]The average callus and embryoid induction rates of maltose at all concentrations were higher than these of sucrose but the difference was not significant.Taking maltose or sucrose as a carbon source,3% to 6% concentration was good for increasing induction frequencies of calli and embryoids.However,If the concentration was over 6%,the induction rates were declined dramatically with the increase of sugar concentration.The influences of growth regulators on induction rate of calli were listed as 2,4-D ZT NAA KT 6-BA;the influences on induction rates of embryoids were listed as 2,4-D NAA ZT KT 6-BA.The 2,4-D,ZT,NAA and KT had signficant or extremely significant influences on induction rates of calli and embryoids.2,4-D,ZT at 1.0 mg/L and NNA,KT at 0.5 mg/L had the best effects.The influences of ZT on calli and embryoids were better than those of KT and 6-BA.1.0 mg/L 2,4-D +1.0 mg/L ZT +0.5 mg/L KT +0.5 mg/L 6-BA was the best regulator combination for induction culture of Jiayu anther.[Conclusion]The experiment provided research basis for anther culture of pepper. |
| [5] | . |
| [6] | . . |
| [7] | . |
| [8] | . . |
| [9] | |
| [10] | . [目的]单倍体花药离体培养是农作物包括果树育种中种质资源创新的最有效方法之一,苹果是染色体高度杂合且自交不亲和树种之一.在当前苹果主栽品种中,‘嘎啦’具有早熟、丰产、稳产、多抗的优良性状,是苹果育种的重要种质资源之一.花药单倍体育种也是苹果新品种培育的重要手段.本研究通过‘嘎啦’苹果花药培养诱导胚状体并获得纯合再生植株,为创制新的纯合体种质资源,加速苹果新品种培育进程提供材料.[方法]采集‘嘎啦,苹果单核靠边期到双核早期的花药(未开放的花蕾),低温处理后进行离体培养,经胚状体诱导,分化培养形成再生苗,再经生根培养获得花药再生植株.之后利用FACS流式细胞仪对再生植株进行倍性分析.取再生植株叶片分离DNA,选用80个来源于苹果HIDRAS数据库的SSR标记对所有植株进行PCR扩增,经过凝胶电泳和荧光毛细管电泳鉴定再生植株纯合基因型.移栽成活后,对每个再生株系进行形态学特征观察及统计分析.[结果]过去3年中,共接种的74 200个‘嘎啦’花药,从未被污染的5万多个花药中成功诱导形成386个胚状体(胚状体诱导率0.7%),经分化培养获得64株再生苗(植株再生率16.6%),最终经生根培养、移栽获得30个成活再生株系.其中包括28个二倍体株系,1个单倍体株系和1个四倍体株系.SSR标记用于纯合性鉴定,PAGE结果表明再生株系均为花粉(小孢子)单倍体细胞来源.为了鉴定这些再生植株基因型,从80个SSR中筛选出17个SSR标记(其余SSR标记不具有多态性或带型杂乱)对所获得的30个再生株系进行基因型鉴定.17个SSR标记所对应的PCR扩增物能有效区分鉴定不同再生植株基因型.继代培养60d后的形态学观察显示不同再生株系的株高、叶长、叶宽等特征差异明显.不同二倍体纯合植株的植物学特征也存在差异:Gala 5植株相对较高,叶基变宽,叶尖渐尖;Gala 7叶片变小、变厚,叶柄变短且基部宽大,叶色深且有很强的光泽度;Gala 18叶片较小,叶数较多.纯合二倍体再生植株长势弱于‘嘎啦’杂合供体,但强于单倍体和纯合四倍体.[结论]采用优化花药培养技术,成功获得了一批苹果纯合体再生植株种质并建立了SSR标记鉴定体系.这些新的种质很大程度丰富了苹果育种亲本种质资源,为挖掘‘嘎啦’苹果优良性状基因提供了重要材料,为后期的田间性状筛选,杂交育种奠定了基础. . [目的]单倍体花药离体培养是农作物包括果树育种中种质资源创新的最有效方法之一,苹果是染色体高度杂合且自交不亲和树种之一.在当前苹果主栽品种中,‘嘎啦’具有早熟、丰产、稳产、多抗的优良性状,是苹果育种的重要种质资源之一.花药单倍体育种也是苹果新品种培育的重要手段.本研究通过‘嘎啦’苹果花药培养诱导胚状体并获得纯合再生植株,为创制新的纯合体种质资源,加速苹果新品种培育进程提供材料.[方法]采集‘嘎啦,苹果单核靠边期到双核早期的花药(未开放的花蕾),低温处理后进行离体培养,经胚状体诱导,分化培养形成再生苗,再经生根培养获得花药再生植株.之后利用FACS流式细胞仪对再生植株进行倍性分析.取再生植株叶片分离DNA,选用80个来源于苹果HIDRAS数据库的SSR标记对所有植株进行PCR扩增,经过凝胶电泳和荧光毛细管电泳鉴定再生植株纯合基因型.移栽成活后,对每个再生株系进行形态学特征观察及统计分析.[结果]过去3年中,共接种的74 200个‘嘎啦’花药,从未被污染的5万多个花药中成功诱导形成386个胚状体(胚状体诱导率0.7%),经分化培养获得64株再生苗(植株再生率16.6%),最终经生根培养、移栽获得30个成活再生株系.其中包括28个二倍体株系,1个单倍体株系和1个四倍体株系.SSR标记用于纯合性鉴定,PAGE结果表明再生株系均为花粉(小孢子)单倍体细胞来源.为了鉴定这些再生植株基因型,从80个SSR中筛选出17个SSR标记(其余SSR标记不具有多态性或带型杂乱)对所获得的30个再生株系进行基因型鉴定.17个SSR标记所对应的PCR扩增物能有效区分鉴定不同再生植株基因型.继代培养60d后的形态学观察显示不同再生株系的株高、叶长、叶宽等特征差异明显.不同二倍体纯合植株的植物学特征也存在差异:Gala 5植株相对较高,叶基变宽,叶尖渐尖;Gala 7叶片变小、变厚,叶柄变短且基部宽大,叶色深且有很强的光泽度;Gala 18叶片较小,叶数较多.纯合二倍体再生植株长势弱于‘嘎啦’杂合供体,但强于单倍体和纯合四倍体.[结论]采用优化花药培养技术,成功获得了一批苹果纯合体再生植株种质并建立了SSR标记鉴定体系.这些新的种质很大程度丰富了苹果育种亲本种质资源,为挖掘‘嘎啦’苹果优良性状基因提供了重要材料,为后期的田间性状筛选,杂交育种奠定了基础. |
| [11] | Based on optimized protocols for anther and microspore culture in apple ( Malus x domestica Borkh.), the regeneration phase and the efficiency of the processes in general were compared by using the same androgenic material of two experimental years. Microspore culture resulted in an increase in embryo induction depending on the genotype (H枚fer 2004), however anther culture was superior to microspore culture in the total number of regenerated plants. The regeneration process in anther and microspore culture is similar. Two developmental pathways were observed: 1) secondary embryogenesis followed by adventitious shoot formation and 2) direct adventitious shoot formation from primary embryos. Induction and regeneration processes are delayed in microspore culture as compared with anther culture. The reasons for the reduced regeneration efficiency in microspore culture are discussed. |
| [12] | The production of doubled haploids offers new possibilities for genetic studies and breeding. In apple Malus 脳 domestica Borkh., double haploid and thus homozygous material was obtained by in vitro androgenesis and in situ parthenogenesis followed by embryo or cotyledon culture. This paper describes a comprehensive evaluation of material induced concerning ploidy level, zygosity state using isozyme and SSR analysis, tree morphology, flower, and fruit quality, respectively. The use of homozygous lines in apple will be discussed in view of the efficiency of the induction process and the fertility of the plant material. The application in running breeding programmes is not a realistic approach at present, while single lines with selected characteristics can already be used for pre-breeding experiments and detailed genetic studies. |
| [13] | . In apple Malus聽脳聽domestica Borkh., double haploid and thus homozygous material was obtained by in vitro androgenesis. Grafted trees of nine lines induced by anther culture and one line induced by isolated microspore culture were investigated at the molecular genetics level and further characterized in the orchard. The double haploid lines originated from five different apple cultivars, which were used as donor material for the in vitro androgenesis. This paper describes a comprehensive evaluation of doubled haploids after a long period in the orchard: including ploidy level using flow cytometry, zygosity state using simple sequence repeat (SSR) marker analysis with a total of 28 SSR markers, resistance traits by screening with linked molecular markers, tree and leaf morphology, pollen fertility of the androgenic genotypes and fruit characteristics. Based on the results obtained we conclude that the use of these homozygous lines in apple breeding is currently not a realistic approach. The homozygous lines characterized in this study will be re-grafted and preserved in the fruit genebank of the Julius K眉hn-Institut. They could be used for pre-breeding experiments and detailed basic genetic studies such as whole genome sequencing and transcriptome analysis for avoiding problems during genome assembly and for reducing the number of expressed alleles for one and same gene, respectively. They are intended to be used internally, but also externally by material requests of other interested research institutions. |
| [14] | . . |
| [15] | . 利用花药培养诱导产生胚状体获得纯合基因型株系,对基因型高度杂合的苹果育种及遗传分析有重要意义。通常花药培养后代为染色体倍性水平不同的混合群体,而且可能会有花药壁等体细胞发育而来的再生植株的混入,再生植株倍性以及来源的准确鉴定,是关乎到单倍体育种技术的有效利用及创新种质遗传分析等的重要问题。本研究中对苹果花药培养获得的再生植株利用流式细胞仪进行了倍性分析,同时提取叶片DNA后利用SSR分析鉴定植株来源。5月上旬苹果现蕾后采集不同时期的花簇,用密封袋封好置于冰箱4℃冷藏室进行低温预处理。低温处理后花蕾在超净工作台内用0.1%次氯酸钠灭菌后,用镊子从花蕾内取出花药接种于胚状体诱导培养基,25℃下暗培养,3~5个月后待胚状体长至8~10 mm,转移至再生培养基上进行植株再生。再生植株获得后利用流式细胞仪(Partec,德国)进行倍性分析,同时提取叶片DNA后SSR分析鉴定植株来源。SSR分析所用10个marker为CH02c06、MS14h03、CH04e02、CH03d12、CH05d03、CH01f12、CH03d02、CH03b06、CH03b10和CH04f10。目前获得‘丹霞'、‘富士'、‘嘎拉'、‘红星'、‘千秋'、‘祝光'等品种的再生植株97株,其中60株已成功继代并得到扩增,37株在继代过程中枯死。从两年的流式细胞仪倍性鉴定结果看,花粉植株的倍性在继代培养的过程中不断变化。共检测了41株苹果花药培养再生植株,第一次检测结果显示这些植株中有单倍体(x)2株、二倍体(2x)32株、三倍体(3x)2株、四倍体(4x)3株、单倍体和二倍体的混倍体(x+2x)2株。继代3~4次后再次检测,其中的2株混倍体(x+2x)消失,一株在继代2次后枯死,另一株转变为二倍体。还有一株单倍体表现为继代3~4次后部分植株自然加倍为二倍体,即在后代群体中既有单倍体又有二倍体。由此可以看到能够存活并成功继代扩增的植株最终都表现为整倍体,之后将对获得的花粉植株的倍性继续进行跟踪鉴定,以了解花粉植株在继代培养过程中的倍性变化情况。利用CH02c06、MSl4h03、CH04e02、CH03d12、CH05d03、CH01f12、CH03d02、CH03b06、CH03b10和CH04f10等10个marker对‘红星'和‘千秋'来源的花药培养再生植株进行SSR分析,结果显示所有植株均只扩增出亲本中一个片段,因此可以证实这些植株均是由花粉发育而来。 . 利用花药培养诱导产生胚状体获得纯合基因型株系,对基因型高度杂合的苹果育种及遗传分析有重要意义。通常花药培养后代为染色体倍性水平不同的混合群体,而且可能会有花药壁等体细胞发育而来的再生植株的混入,再生植株倍性以及来源的准确鉴定,是关乎到单倍体育种技术的有效利用及创新种质遗传分析等的重要问题。本研究中对苹果花药培养获得的再生植株利用流式细胞仪进行了倍性分析,同时提取叶片DNA后利用SSR分析鉴定植株来源。5月上旬苹果现蕾后采集不同时期的花簇,用密封袋封好置于冰箱4℃冷藏室进行低温预处理。低温处理后花蕾在超净工作台内用0.1%次氯酸钠灭菌后,用镊子从花蕾内取出花药接种于胚状体诱导培养基,25℃下暗培养,3~5个月后待胚状体长至8~10 mm,转移至再生培养基上进行植株再生。再生植株获得后利用流式细胞仪(Partec,德国)进行倍性分析,同时提取叶片DNA后SSR分析鉴定植株来源。SSR分析所用10个marker为CH02c06、MS14h03、CH04e02、CH03d12、CH05d03、CH01f12、CH03d02、CH03b06、CH03b10和CH04f10。目前获得‘丹霞'、‘富士'、‘嘎拉'、‘红星'、‘千秋'、‘祝光'等品种的再生植株97株,其中60株已成功继代并得到扩增,37株在继代过程中枯死。从两年的流式细胞仪倍性鉴定结果看,花粉植株的倍性在继代培养的过程中不断变化。共检测了41株苹果花药培养再生植株,第一次检测结果显示这些植株中有单倍体(x)2株、二倍体(2x)32株、三倍体(3x)2株、四倍体(4x)3株、单倍体和二倍体的混倍体(x+2x)2株。继代3~4次后再次检测,其中的2株混倍体(x+2x)消失,一株在继代2次后枯死,另一株转变为二倍体。还有一株单倍体表现为继代3~4次后部分植株自然加倍为二倍体,即在后代群体中既有单倍体又有二倍体。由此可以看到能够存活并成功继代扩增的植株最终都表现为整倍体,之后将对获得的花粉植株的倍性继续进行跟踪鉴定,以了解花粉植株在继代培养过程中的倍性变化情况。利用CH02c06、MSl4h03、CH04e02、CH03d12、CH05d03、CH01f12、CH03d02、CH03b06、CH03b10和CH04f10等10个marker对‘红星'和‘千秋'来源的花药培养再生植株进行SSR分析,结果显示所有植株均只扩增出亲本中一个片段,因此可以证实这些植株均是由花粉发育而来。 |
| [16] | . . |
| [17] | |
| [18] | . 为探明高粱养分吸收和根系生长对氮、磷、钾胁迫的响应,通过长期定位试验,在高粱/玉米轮作条件下研究了不同养分配比NPK、PK、NK、NP、CK对高粱根系生长及养分吸收的影响.结果表明:与NPK相比,长期不施氮肥(PK)条件下高粱总根长增加18.29%,总根体积降低26.52%,且根系主要分布在0~10 cm土层,直径小于0.5 mm细根所占比例显著增加.不施磷肥(NK)显著抑制了高粱根系生长,总根长、总根表面积和总根体积分别降低24.03%、27.48%和41.29%.不施钾肥(NP)对细根生长有明显抑制作用.不施氮、磷、钾均降低高粱对相应养分的吸收和累积,不施氮促进了营养器官中氮和钾素向籽粒转运,不施磷或钾肥抑制了氮、磷及钾的转运.高粱对养分的吸收、积累和转运与根系形态有关,不同养分积累与运转与根系形态关系表现不尽相同:氮素、钾素积累和转运与根系形态具有较好的相关性,氮素的积累和转运与植株生物量和产量的相关性大于磷素和钾素.综上,高粱根系形态及养分吸收对氮、磷及钾胁迫响应不同,该研究可为不同养分瘠薄地高粱高效栽培提供理论依据. . 为探明高粱养分吸收和根系生长对氮、磷、钾胁迫的响应,通过长期定位试验,在高粱/玉米轮作条件下研究了不同养分配比NPK、PK、NK、NP、CK对高粱根系生长及养分吸收的影响.结果表明:与NPK相比,长期不施氮肥(PK)条件下高粱总根长增加18.29%,总根体积降低26.52%,且根系主要分布在0~10 cm土层,直径小于0.5 mm细根所占比例显著增加.不施磷肥(NK)显著抑制了高粱根系生长,总根长、总根表面积和总根体积分别降低24.03%、27.48%和41.29%.不施钾肥(NP)对细根生长有明显抑制作用.不施氮、磷、钾均降低高粱对相应养分的吸收和累积,不施氮促进了营养器官中氮和钾素向籽粒转运,不施磷或钾肥抑制了氮、磷及钾的转运.高粱对养分的吸收、积累和转运与根系形态有关,不同养分积累与运转与根系形态关系表现不尽相同:氮素、钾素积累和转运与根系形态具有较好的相关性,氮素的积累和转运与植株生物量和产量的相关性大于磷素和钾素.综上,高粱根系形态及养分吸收对氮、磷及钾胁迫响应不同,该研究可为不同养分瘠薄地高粱高效栽培提供理论依据. |
| [19] | . In the present paper attempts were made to induce chromosome doubling of ‘Meiwa’ kumquat ( Fortunella02crassifolia ) protoplasts and ‘Frost’ navel orange ( Citrus02sinensis 02Osbeck) embryogenic callus via colchicine treatment. Colchicine decreased protoplast viability, delayed protoplast division and inhibited callus growth, indicating presence of toxicity to cells. Cell lines established from ‘Meiwa’ protoplasts treated with 0.01 and 0.1% colchicine for 8, 16 and 2402h at each concentration showed different responses when they were cultured on embryoid-induction medium. Flow cytometry (FCM) demonstrated that tetraploids were detected in cell lines and embryoids from all of the treatments, with the highest frequency being 19.23%. As for ‘Frost’, tetraploid cells were only detected when the callus was treated with 0.1% colchicine for either 4 or 802days, from which plantlets were regenerated. FCM and chromosome counting confirmed them as true tetraploids. The diploid cells were more active in mitotic division during a 12-day culture and smaller in size than their tetraploid counterpart. Potential applications of the novel tetraploid germplasms obtained through in02vitro chromosome doubling to citrus cultivar improvement are discussed. |
| [20] | . |
| [21] | . Based on a protocol for microspore culture in apple ( Malus domestica Borkh.), the embryo induction phase has been improved with regard to pretreatment of microspores for initiation of microspore embryogenesis, the concentration of carbon source in the induction medium and the microspore density in the suspension. Furthermore, the effect of the genotype was studied. To determine the efficiency of in vitro androgenesis, both methods, via anther and microspore culture, were investigated using the same bud material. A comparison of the efficiency of embryo induction in anther and microspore cultures showed that microspore culture resulted in an increase up to 10 times, depending on the genotype. The regeneration route in microspore culture is similar to that of androgenic embryos via anther culture and showed adventitious shoot formation in most cases after a long period of secondary embryogenesis. |
| [22] | . A multidisciplinary study was carried out to analyse the chromosome doubling process during the early stages of in vitro maize microspore embryogenesis. The main stages (microspore derivatives) that were formed in the course of the culture were analysed. Chromosome number was determined from squashed cells, and DNA content was measured by cytometry. In parallel, an ultrastructural analysis of the microspore derivatives demonstrated the occurrence of a nuclear fusion process. It seems likely that nuclear fusion ensures chromosome doubling at early stages of induced microspore embryogenesis. It occurs precisely at the 5/7聽day stage in the embryonic domain and probably leads to polyploidy in the endosperm domain of the microspore derivatives. As a conclusion a scheme summarises the results and proposes an interpretation of the sequence of chromosome doubling events during early maize microspore embryogenesis. Understanding of this process will be important for future efforts to increase the percentage of homozygous plants for crop improvement. |
| [23] | |
| [24] | . 以国内外核果类果树茎尖培养及其应用方面的文献为基础,对茎尖培养在核果类果树上的主要应用,影响核果类果树茎尖培养的主要因素进行了综述,提出目前核果类果树茎尖培养过程中存在的主要问题及有效的解决途径,分析了核果类果树试管苗的驯化移栽问题. . 以国内外核果类果树茎尖培养及其应用方面的文献为基础,对茎尖培养在核果类果树上的主要应用,影响核果类果树茎尖培养的主要因素进行了综述,提出目前核果类果树茎尖培养过程中存在的主要问题及有效的解决途径,分析了核果类果树试管苗的驯化移栽问题. |
| [25] | . . |
| [26] | . In vitro clonal multiplication of apple rootstock MM 111 using axillary buds and shoot apices were carried out. Vegetative axillary buds of the size of 0.2-2.0 cm and shoot apices measuring 4 mm in length were initiated to shoot proliferation on MS medium supplemented with BA (0.5 - 1.0 mgl(-1)), GA3(0.5 mgl(-1)), with or without IBA(0.05 - 0.1 mgl(-1)). Small size explants showed less phenol exudation and less contamination. Following establishment phase, the small shoots emerged from explants were subcultured on MS medium supplemented with different combinations and concentrations of growth regulators. BA (1.0 mgl(-1)) and GA3 (0.5 mgl(-1)) combination showed highest multiplication rate (1:5), andcl also produced longer shoots. Two step rooting was done by transferring microcuttings to auxin free solid medium after root initiation in dark on 1/2 strength MS liquid medium containing IBA (0.5 mgl(-1) ). Rooted plantlets were transferred to peat containing paper cups and resulting plants of MM 111 acclimated successfully for transfer to field. |
| [27] | . <h2 class="secHeading" id="section_abstract">Abstract</h2><p id="">Micropropagation of apple has played an important role in the production of healthy, disease-free plants and in the rapid multiplication of scions and rootstocks with desirable traits. During the last few decades, in apple, many reliable methods have been developed for both rootstocks and scions from a practical, commercial point of view. Successful micropropagation of apple using pre-existing meristems (culture of apical buds or nodal segments) is influenced by several internal and external factors including <em>ex vitro</em> (e.g. genotype and physiological state) and <em>in vitro</em> conditions (e.g., media constituents and light). Specific requirements during stages of micropropagation, such as the establishment of <em>in vitro</em> cultures, shoot multiplication, rooting of microshoots and acclimatization are summarized in this review. New approaches for increasing shoot multiplication and rooting for apple and current use of micropropagated plantlets as tools in basic and applied research are also discussed.</p> |
| [28] | . 为提高苹果苗木繁殖速度及苗木品质,本文以‘红富士’苹果组培苗为试材,探索苹果茎尖启动培养过程中黑暗培养对不定芽再生、生根诱导培养过程中IBA和蔗糖浓度对不定根发生及不同炼苗方式对组培苗移栽成活的影响,优化了苹果茎尖培养的快繁体系。结果表明,茎尖启动培养中,在培养基MS+6-BA 4.0 mg·L~(-1)+NAA 0.5 mg·L~(-1)+山梨醇20 g·L~(-1)+蔗糖10g·L~(-1)上,20 d暗培养后再转入不含NAA的MS培养基(其他成份同上)进行光照培养,茎尖愈伤组织的诱导率和不定芽再生率较高,优于黑暗培养40和60 d。培养基1/2 MS+IBA 1.00 mg·L~(-1)+蔗糖20 g·L~(-1)适宜不定芽的诱导生根,并且在低浓度IBA的生根培养基中,蔗糖浓度的提高有利于苹果组培苗的不定根发生。炼苗的初始2周采用改良的霍格兰德半营养液覆膜水培炼苗,后转入全营养液中敞口培养3周,再移栽至营养钵中,组培苗移栽成活率可达到75%,优于常规的苹果组培苗炼苗处理。 . 为提高苹果苗木繁殖速度及苗木品质,本文以‘红富士’苹果组培苗为试材,探索苹果茎尖启动培养过程中黑暗培养对不定芽再生、生根诱导培养过程中IBA和蔗糖浓度对不定根发生及不同炼苗方式对组培苗移栽成活的影响,优化了苹果茎尖培养的快繁体系。结果表明,茎尖启动培养中,在培养基MS+6-BA 4.0 mg·L~(-1)+NAA 0.5 mg·L~(-1)+山梨醇20 g·L~(-1)+蔗糖10g·L~(-1)上,20 d暗培养后再转入不含NAA的MS培养基(其他成份同上)进行光照培养,茎尖愈伤组织的诱导率和不定芽再生率较高,优于黑暗培养40和60 d。培养基1/2 MS+IBA 1.00 mg·L~(-1)+蔗糖20 g·L~(-1)适宜不定芽的诱导生根,并且在低浓度IBA的生根培养基中,蔗糖浓度的提高有利于苹果组培苗的不定根发生。炼苗的初始2周采用改良的霍格兰德半营养液覆膜水培炼苗,后转入全营养液中敞口培养3周,再移栽至营养钵中,组培苗移栽成活率可达到75%,优于常规的苹果组培苗炼苗处理。 |
| [29] | . 结合作者多年水稻花培育种的实践,论述该育种特点、程序、培养条件及其效率比较。认为花培育种既继承杂浆、诱变育种等经验,又吸收了细胞生物学、胚胎学、遗传学等生物科学的新成熟。是产生“纯系”的捷径,能控制杂种分离,克服杂交育种选择效率低、时间长、品种易退化等问题,是快速、高效培育水稻新品种较好方法之一。 . 结合作者多年水稻花培育种的实践,论述该育种特点、程序、培养条件及其效率比较。认为花培育种既继承杂浆、诱变育种等经验,又吸收了细胞生物学、胚胎学、遗传学等生物科学的新成熟。是产生“纯系”的捷径,能控制杂种分离,克服杂交育种选择效率低、时间长、品种易退化等问题,是快速、高效培育水稻新品种较好方法之一。 |
| [30] | . Apple is a temperate fruit species with several breeding disadvantages such as the highly heterozygous nature of the genome and a long reproductive cycle. It is expected that homozygous apple genotypes can increase breeding efficiency, but inbreeding is prevented by the gametophytic self-incompatibility mechanism. In this paper, the production and characterization of homozygous genotypes, generated from anther culture in Malus 脳 domestica cv. 'Braeburn', is described as well as their potential for use in breeding programmes. After determination of large scale anther culture efficiency, anther culture-derived genotypes were simple sequence repeat (SSR)-fingerprinted to determine their homozygous nature and confirm their androgenic origin. Their value for breeding was estimated based on growth and fertility measurements. The use of homozygous androgenic genotypes from 'Braeburn' in apple breeding programmes is currently not a realistic approach, partly because of the low efficiency of anther culture, but mainly because of the reduced vigour and severe sterility of the androgenic genotypes produced. |
| [31] | . Doubled haploids can improve the efficiency of breeding and genetic study in apple (Malus 脳 domestica Borkh.). Seventeen homozygous genotypes were obtained by in vitro anther culture from 'Senshu' apple. Flow cytometry analysis revealed that the ploidy level of the anther-derived plantlets was diploid. Simple sequence repeat (SSR) analysis determined the origin and homozygous status of the anther-derived plantlets. The results of S-RNase polymerase chain reaction (PCR)-digestion analysis reinforced the homozygous state. The morphological characteristics and reproductive potential of the doubled haploids were investigated. It is especially interesting that one of the doubled haploids derived from anther culture had fertile pollen grains (rate of in vitro germination was 20.6%-33.0%), and several progenies were obtained from the cross between 'Prima' apple and the doubled haploid. The hybridism of these seedlings was confirmed by SSR analysis. Furthermore, the female gametes of the doubled haploid showed slight fertility. Thus, this doubled haploid will be valuable for breeding and genetic studies. |
| [32] | . Homozygous genotypes are valuable for genetic and genomic studies in higher plants. However, obtaining homozygous perennial plants using conventional breeding techniques is currently a challenge because of a long juvenile period, high heterozygosity and the substantial inbreeding depression. In vitro androgenesis has been used to develop haploid and doubled haploid plants. In this study, we report the regeneration of doubled haploid lines of Valencia sweet orange cv. Rohde Red ( Citrus sinensis [L.] Osbeck) via anther culture. Anthers at the uninucleate stage were induced and two embryogenic calli were obtained that further regenerated to embryoids (2/400). Plantlets were obtained after transferring the embryoids to a shoot regeneration medium, but were short-lived. Ploidy analysis via both flow cytometry and chromosome counting verified that these two lines were diploids. Additionally, 43 simple sequence repeat (SSR) markers which showed to be heterozygous in the Valencia sweet orange donor line confirmed homozygosity and doubled haploids in the anther-derived lines. Furthermore, analysis of the doubled haploids via cleaved amplified polymorphic sequence (CAPS) markers and target region sequencing confirmed the allelic state of two genes ( LCYE and LCYB ) involved in the carotenoid biosynthesis of sweet oranges. |
| [33] | . Three gametoclonal plants ofCitrus clementinaHort. ex Tan., cv. Nules, designated ESP, FRA, and ITA (derived from three labs in Spain, France, and Italy, respectively), were selected for cytological and molecular characterization in order to elucidate genomic rearrangements provoked by haploidization. The study included comparisons of their ploidy, homozygosity, genome integrity, and gene dosage, using chromosome counting, flow cytometry, SSR marker genotyping, and array-Comparative Genomic Hybridization (array-CGH). Chromosome counting and flow cytometry revealed that ESP and FRA were haploid, but ITA was tri-haploid. Homozygous patterns, represented by a single peak (allele), were observed among the three plants at almost all SSR loci distributed across the entire diploid donor genome. Those few loci with extra peaks visualized as output from automated sequencing runs, generally low or ambiguous, might result from amplicons of paralogous members at the locus, non-specific sites, or unexpected recombinant alleles. No new alleles were found, suggesting the genomes remained stable and intact during gametogenesis and regeneration. The integrity of the haploid genome also was supported by array-CGH studies, in which genomic profiles were comparable to the diploid control. The presence of few gene hybridization abnormalities, corroborated by gene dosage measurements, were hypothetically due to the segregation of hemizygous alleles and minor genomic rearrangements occurring during the haploidization procedure. In conclusion, these plants that are valuable genetic and breeding materials contain completely homozygous and essentially intact genomes. |
| [34] | |
| [35] | . New Phytol. 2007;174(4):717-20. Congresses; Research Support, N.I.H., Extramural; Research Support, U.S. Gov't, Non-P.H.S. |
| [36] | . |
| [37] | . |
| [38] | Abstract Polyploidization is an important process in the evolutionary history of most eukaryotic species. It oftentimes causes large-scale genomic reorganizations and is accompanied by a wide variety of phenotypic alterations in morphology, niche preference and fitness characteristics. Despite their importance, the morphological effects of alterations in ploidy are not well understood. We investigated these changes in four diverse maize inbred lines, using monoploid, diploid, triploid and tetraploid derivatives, measuring 13 characters in a randomized field study. Employing several analysis of variance approaches, we find that all characters investigated strongly respond to alterations in ploidy. This response appears to have two sources: one source is shared by all inbred lines and constitutes a common response to ploidy change. The other source is genotype specific and results in a response to ploidy change that varies among inbred lines. This finding demonstrates the existence of genetic variation for the morphological response to ploidy change in Zea mays. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}