0 引言
【研究意义】针对甘蓝类蔬菜(Brassica oleracea)中缺乏Ogura细胞质雄性不育(Ogura cytoplasmic male sterility,Ogura CMS)育性恢复材料的问题,利用远缘杂交结合胚挽救将育性恢复基因Rfo(restorer- of-fertility gene)从甘蓝型油菜导入芥蓝中[1]。对加倍后的种间杂种进行回交,已分别获得BC1和BC2代育性恢复材料[2]。但这些材料遗传背景还与芥蓝有较大差异,仍需进一步回交,保留Rfo的同时尽可能消除甘蓝型油菜的遗传背景,这对最终实现Ogura CMS甘蓝类蔬菜的育性恢复、打破国外公司利用Ogura CMS对特异种质的垄断具有重要意义。【前人研究进展】甘蓝类蔬菜是芸薹属中典型的异花授粉植物,杂种优势十分明显,利用雄性不育系进行杂交种生产是甘蓝类蔬菜杂种优势利用的主要途径之一[3]。Ogura细胞质雄性不育系由于其不育性稳定,转育容易,已成为甘蓝类蔬菜可实用的主要不育源[4]。早在20世纪70年代,BANNEROT等[5]利用远缘杂交将萝卜中的Ogura胞质不育转到甘蓝类蔬菜中,并经过原生质体融合将其改良后[6],Ogura CMS已逐渐成为甘蓝类蔬菜杂交制种中广泛应用的细胞质不育类型[7]。但随着Ogura CMS的推广利用,近几年来越来越多的优异品种是胞质不育育成,不能自交分离,无法继续进行材料的创新利用[8,9,10]。而创制Ogura CMS的恢复系是解决这一问题的有效途径,但在甘蓝中不存在天然的Ogura CMS恢复材料。甘蓝型油菜Ogura CMS恢复系的研究历经30余年[11,12],已将Rfo从萝卜属中导入到甘蓝型油菜,并整合到了甘蓝型油菜的C09染色体上[13,14],成功创制出恢复系并应用于甘蓝型油菜Ogura CMS制种,Ogu-CMS/Rfo育种体系开始成为甘蓝型油菜杂交种生产的主要技术之一,这为研究者将甘蓝型油菜中Rfo向甘蓝类蔬菜转移提供了可能。为缩短转育年限,YU等[1,2]选择甘蓝类蔬菜中容易春化、繁殖周期短的变种芥蓝作为转育材料,利用远缘杂交结合胚挽救技术,将Rfo导入Ogura CMS芥蓝中,并对加倍的F1代(种间六倍体杂种)多代回交,结合形态、育性、遗传背景分析,筛选获得育性恢复、结实性好的BC2代Rfo阳性单株。【本研究切入点】选育的BC2代Rfo阳性单株(15Q23)的遗传背景与芥蓝仍有不少差异,需要进一步回交以减少背景的干扰;且15Q23自然结实性仍很低(<0.1粒/种荚),远达不到正常水平,自然授粉结实性有待进一步提升。【拟解决的关键问题】利用芥蓝对BC2代育性较好的单株15Q23进一步回交,分析BC3代和BC4代Rfo的传递效率(transmission rate,TR),分析BC3代单株遗传背景,获得育性恢复、花粉活力好、结实性强的BC3代单株。1 材料与方法
1.1 试验材料
回交母本:Ogura CMS芥蓝15Y102。第3代回交父本:芥蓝与甘蓝型油菜杂交后的BC2代中育性较好的单株15Q23,这是前期试验中利用芥蓝和含Rfo的甘蓝型油菜进行种间杂交,获得的种间杂种加倍后,回交两代后利用Rfo特异分子标记筛选结合形态、遗传背景分析获得的,该单株育性恢复、花粉活力显著优于其余单株,形态更接近于亲本芥蓝且结实性良好[1,2]。第4代回交父本:第3代回交后用标记筛选获得的育性恢复、背景更接近芥蓝母本的优良单株。上述材料均由中国农业科学院蔬菜花卉研究所甘蓝课题组提供,于2015年、2016年秋季定植于本所南圃场,常规栽培管理。
1.2 BC3代单株的获得及Rfo阳性单株的筛选
2016年3月,在盛花期进行人工剥蕾去雄授粉,并挂牌标记杂交组合、授粉花蕾数,计算结实率(seed setting)等,结实率=饱满种子数/授粉花蕾数×100%。将收获的BC3代种子分两批播种,第一批挑选少量种子于2016年7月底播种(编号为16Q),用于初筛,调查Rfo阳性株比例、形态特征和分析遗传背景;第二批在10月初播种,筛选Rfo阳性单株用于大规模的回交转育(编号为16QW)。所有BC3单株成苗后,采用改良CTAB法提取基因组DNA[15]。利用已开发的Rfo特异标记[1](RFO-2F/RFO-NEW-R)对两批BC3群体进行筛选,PCR扩增方法、体系及检测方法参照YU等[1],计算Rfo的传递效率,Rfo传递效率= Rfo阳性单株数目/群体总单株数×100%。
1.3 16Q群体形态学性状调查及遗传背景分析
将16Q群体于9月初定植于温室,对各单株成熟期的形态特征进行观察和调查。共对16个形态特征进行调查,其中10个性状采取分级赋值法(表1),包括株型、基生叶颜色、基生叶形状、基生叶叶缘、基生叶叶面、叶面蜡粉、叶面光泽、基生叶翼、基生叶叶脉、花瓣颜色。另外6个性状利用刻度尺进行测量,包括株高、基生叶长、基生叶宽、叶柄长、花直径、花蕾长,这些测量数据根据王述民等[16]方法进行10级分类转化,1级<M-2σ,10级≥M+2σ,M代表平均值,σ为标准差,每一级差0.5σ。对统计的性状数据进行标准化,利用NTSYSpc2.11a软件,采用非加权类平均法(UPGMA)进行聚类分析。Table 1
表1
表1芥蓝和甘蓝型油菜杂交后回交后代的性状调查分级标准
Table 1The traits investigation grade for the backcross progenies from the hybrids between Chinese kale and rapeseed
| 性状 Trait | 赋值标准 Classification standard | |||||||
|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
| 株型Plant type | 直立 Erect | 半直立 Semi-erect | 半开展 Semi-patulous | 平展 Prostrate | ||||
| 基生叶颜色 Color of radical leaves | 浅绿 Light green | 绿 Green | 深绿 Dark green | 浅灰绿 Light grayish green | 灰绿 Grayish green | 深灰绿 Dark grayish green | 紫绿 Purplish green | |
| 基生叶形状 Shape of radical leaves | 近圆 Nearly circular | 卵圆 Ovate | 倒卵圆 Obovate | 宽卵圆 Broad ovate | 椭圆 Elliptic | |||
| 基生叶叶缘 Margin of radical leaves | 全缘 Entire | 波状 Undulant | 锯齿 Serrate | |||||
| 基生叶叶面 Leaf surface of radical leaves | 平展 Smooth | 微皱 Slightly wrinkling | 皱 Wrinkling | 多皱 Much wrinkling | ||||
| 叶面蜡粉 Wax on leaf surface | 无 Absent | 少 Little | 中 Intermediate | 多 Much | ||||
| 叶面光泽 Luster on leaf surface | 无 Absent | 有 Present | ||||||
| 基生叶翼 Auricle of radical leaf | 无 Absent | 有 Present | ||||||
| 基生叶叶脉 Vein in radical leaf | 不明显 Indistinct | 中 Intermediate | 明显 Distinct | |||||
| 花瓣颜色 Flower color | 白 White | 淡黄 Light yellow | 黄 Yellow | |||||
新窗口打开
利用于海龙等[17]开发的芥蓝与甘蓝型油菜之间的32对SSR多态性背景标记,对16Q群体的遗传背景进行分析。PCR反应体系及PCR扩增产物的检测方法均参照于海龙等[17]。针对引物扩增产生的每个等位变异,不同材料在该等位变异处有带记为1,无带记为0。利用NTSYSpc2.11a软件计算亲本和各世代单株间的遗传相似系数,采用UPGMA进行聚类分析。
1.4 BC3代Rfo阳性单株花粉活力评估和倍性分析
于盛花期对16Q群体中筛选的Rfo阳性单株的育性进行观察,并测定花粉活力。花粉活力测定采用醋酸洋红染色法,取植株新开的3朵花,将其花粉均匀涂于载玻片上,用1%的醋酸洋红染色,观察3个视野(视野内花粉粒数≥300)。为方便数据处理,对花粉活力进行分级,共划分4个等级,1级:≤25%;2级:25%<花粉活力≤50%;3级:50%<花粉活力<75%;4级:花粉活力≥75%。利用流式细胞仪对16Q群体中的Rfo阳性单株采用流式细胞术(flow cytometry,FCM)方法进行倍性鉴定。参照DOLEZEL等[18]关于植物DNA相对含量测定的方法,略有改动,具体方法如下:取待测样品叶片200 mg,分别加入1—1.5 mL细胞裂解液(Galbraith’s buffer),以一次性锋利刀片切碎组织,混合匀浆,利用400目滤网将匀浆过滤到1.5 mL离心管中,滤液体积应>500 μL,弃上清,加入PI染液(工作浓度50 μg·mL-1)对核进行染色,避光处理30 min后,上机(BD FACSCaliburTM,BD Biosciences,San Jose,CA,USA)测量。以芥蓝2C DNA相对含量为参照,将其G0/G1期对应峰值调整到200通道处,根据杂种后代G0/G1期峰值的位置,判断其倍性。
1.5 BC4回交后代的获得、Rfo阳性单株的筛选及倍性分析
结合背景分析和育性调查结果,挑选16Q群体中育性较好、背景近于芥蓝的Rfo阳性单株为重点,与芥蓝15Y102进行第4代回交,统计每单株授粉花蕾数及结籽情况,计算各Rfo阳性单株的结实率。将获得的部分BC4代种子于2017年1月初播种,成苗后利用Rfo特异标记筛选Rfo阳性株,计算Rfo传递效率。选取部分BC4代Rfo阳性单株利用流式细胞仪对其倍性进行鉴定。
1.6 数据统计与分析
数据统计和计算利用Microsoft Office Excel 2003,不同组合间方差分析、相关性分析和显著性检验均使用SPSS 11.0软件(SPSS,Chicago,IL,USA)。2 结果
2.1 BC3代单株的获得及Rfo阳性单株的筛选
以15Q23为父本,利用Ogura CMS芥蓝15Y102为母本进行回交。在整个花期对15Q23的育性观察发现,15Q23在整个花期的不同阶段,该单株育性的差异较大,整体表现为开花后20 d内15Q23育性较好,花粉活力较高,平均花粉活力87%(表2、图1-a);20 d后,该单株花粉活力43%,部分花朵的花药上甚至观察不到花粉(表2、图1-b)。两个时期结实率存在极显著差异(P<0.01),开花20 d内共收获种子265粒,结实率为13%,而开花20 d后,这段时期仅收获种子40粒(表2)。相关性分析表明,结实率和花粉活力间存在极显著正相关(P<0.01)。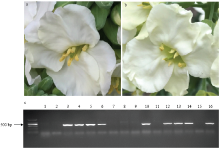 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图115Q23开花前期(a)和开花后期(b)育性表现及16Q群体单株Rfo筛选(c)
图c中1、2:芥蓝Chinese kale;3、4:Rfo甘蓝型油菜Rapeseed with Rfo;5:15Q23;6—16:16Q群体部分单株(6、10、12、13、14、16为Rfo阳性株)Some individuals of 16Q group (6, 10, 12, 13, 14, 16 are Rfo-positive individuals)
-->Fig. 1Fertility performance of 15Q23 on fore-period of flowering (a) and latter-period of flowering (b), and screening of partial 16Q individuals by Rfo specific marker (c)
-->
Table 2
表2
表215Q23单株在开花前期及后期的结实性比较
Table 2The seed setting comparison of 15Q23 between fore-period of flowering and latter-period of flowering
| 花期 Flowering period | 花粉活力 Pollen viability (%) | 授粉花数 Number of pollinated flowers | 种子数 Number of seeds | 结实率 Seed setting rate (%) (Mean±SE) | 整个花期平均结实率 Average seed setting rate in the whole flowering period (%) | 成苗数 Number of viable plants | Rfo阳性植株 Number of Rfo-positive plants | Rfo传递效率 Transmission rate of Rfo (%) |
|---|---|---|---|---|---|---|---|---|
| 开花前期(15Q23开花后20 d内,2.25-3.15) Fore-period of flowering (before the 20th day of flowering) | 87 | 2304 | 265 | 13±6** | 7 | 263 | 126 | 48 |
| 开花后期(15Q23开花20 d后)Latter-period of flowering (after the 20th day of flowering) | 43 | 1915 | 40 | 2±0.8 |
新窗口打开
将收获的305粒种子分两批播种,第一批播种43粒,成苗34株(即16Q群体),利用Rfo特异标记筛选,17株为Rfo阳性(图1-c),Rfo传递效率为50%;第二批播种262粒,成苗229株(即16QW群体),Rfo特异标记检测结果表明,109株为Rfo阳性,Rfo传递效率为48%。两个群体Rfo阳性株与不含Rfo单株比值都近于1﹕1。
2.2 BC3后代的遗传背景、形态和倍性分析
利用背景标记分析16Q群体的34株成苗后代的遗传背景,并观察其形态特征,期望获得形态学特征和遗传背景近于亲本芥蓝的单株。将16Q群体的34株单株于2016年9月初定植于温室,对各单株的形态进行观察比较。34株BC3单株的形态整体上都近于芥蓝,如株型(半直立),叶型(卵圆、椭圆),叶面(微皱),叶色(深绿),叶面蜡粉(中),花瓣颜色(白)等性状基本与芥蓝一致(图2)。较亲本芥蓝,BC3代Rfo阳性单株的生长势较强,最大外叶长、最大外叶宽、茎粗、株高、花朵直径等性状的值均略高于亲本芥蓝。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2不同时期芥蓝亲本和BC3代Rfo阳性单株16Q1-4的形态特征对比
苗期Seedling stage(a:15Y102;d:16Q1-4)、成熟期Mature stage(b:15Y102;e:16Q1-4)、开花期Flowering stage(c:15Y102;f:16Q1-4)
-->Fig. 2Morphological characterization comparison between the Rfo-positive BC3 individual 16Q1-4 and Chinese kale
-->
对34株BC3代单株进行遗传背景聚类,结果表明在遗传相似系数0.74处,34株BC3后代和芥蓝15Y102聚为一类,与芥蓝15Y102遗传相似系数在0.81—0.92,高于其回交用父本15Q23(0.73)。其中,单株16Q1-34与芥蓝间遗传相似系数最高,为0.92,17个Rfo阳性株中,单株16Q1-8、16Q1-13、16Q1-18与亲本芥蓝间的遗传相似系数为0.90(图3-a)。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3SSR背景标记(a)和形态标记(b)用于34株BC3后代的聚类分析
-->Fig. 3Cluster analysis 34 BC3 progenies by SSR marker (a) and morphological markers (b)
-->
利用形态学数据对BC3后代植株的表型进行聚类分析,结果表明34株BC3后代与芥蓝15Y102聚为一个分支,甘蓝型油菜单独聚成一个分支,背景标记和形态学标记都表明所有BC3后代遗传背景近于亲本芥蓝。与背景标记聚类结果相比,利用形态数据聚类时,16Q群体中部分单株无法区分,表明形态学数据聚类分辨率较低(图3-b)。
利用流式细胞术,对34株BC3后代(16Q)的倍性进行鉴定,结果如图4所示,34株BC3后代单株倍性仍为多倍体形式。以芥蓝为参照标准(200),34个BC3后代中,只有单株16Q1-13的荧光峰值位于300通道处,推测倍性接近于三倍体;其余33个单株(包括Rfo阳性单株和不含Rfo的单株)的荧光峰值都位于400通道处,推测倍性接近于四倍体。
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4BC2后代15Q23及BC3后代16Q1-4和16Q1-13的倍性鉴定
-->Fig. 4Ploidy identification of 15Q23 and BC3 individuals 16Q1-4 & 16Q1-13
-->
2.3 BC3代Rfo阳性单株的育性调查
开花后对所有单株育性进行调查,34个单株育性表现与Rfo标记筛选结果完全一致,所有阳性单株均有不同程度的花粉出现(图2-f),其余单株均为不育,标记筛选准确率达到100%。不同单株间育性表现存在差异,对Rfo阳性单株的花粉活力进行测定,单株16Q1-4、16Q1-7、16Q1-10在整个花期育性较好,花粉活力一直在75%以上(表3)。利用芥蓝对育性较好的Rfo阳性单株(16Q1-4、16Q1-7、16Q1-8、16Q1-10、16Q1-13、16Q1-19、16Q1-25、16Q1-27、16Q1-35,花粉活力>2级)进行回交,结实率统计结果表明(表3),单株16Q1-4和单株16Q-10结实性最好,结实率分别为15%和9%。显著性分析结果表明,较回交用父本BC2代阳性单株15Q23,单株16Q1-4和单株16Q-10结实率显著提高(P<0.05)。相关性分析表明,BC3代Rfo阳性单株的结实率与花粉活力显著正相关(P<0.05,r=0.697*)。Table 3
表3
表3BC3代花粉活力>2级的单株的遗传相似系数、结实率及其回交后代Rfo传递效率
Table 3The genetic similarity coefficient with parent Chinese kale, seed setting rate and TR of Rfo of fertility-restored BC3 individuals (pollen viability level >2)
| 单株编号 Individual code | 花粉活力级别 Pollen viability level | 与芥蓝15Y102遗传相似系数 Genetic similarity coefficient with parent Chinese kale 15Y102 | 结实率 Seed setting rate (%) | 回交后Rfo传递效率 TR of Rfo in backcross progenies |
|---|---|---|---|---|
| 15Q23 | 3.3 | 0.74 | 7 | 48% (126/263) |
| 16Q1-4 | 4 | 0.87 | 15 | 34% (24/70) |
| 16Q1-7 | 4 | 0.88 | 4 | 34% (14/41) |
| 16Q1-8 | 3.7 | 0.90 | 7 | 33% (19/57) |
| 16Q1-10 | 4 | 0.88 | 9 | 33% (18/55) |
| 16Q1-13 | 2.6 | 0.90 | 2 | |
| 16Q1-19 | 2.3 | 0.81 | 0 | |
| 16Q1-25 | 2.7 | 0.88 | 0 | |
| 16Q1-27 | 2.3 | 0.87 | 2 | |
| 16Q1-35 | 3.7 | 0.87 | 4 | 45% (14/31) |
新窗口打开
2.4 BC4代的获得及Rfo传递效率
收获16Q群体中Rfo阳性单株16Q1-4、16Q1-7、16Q1-8、16Q1-10、16Q1-35回交获得的种子,播种成苗后利用Rfo特异标记进行检测,结果表明(表3、图5-a),不同单株回交后获得的BC4代中均含有Rfo阳性株,5株BC3代Rfo阳性株中除16Q1-35外(TR=45%),其余4株的Rfo传递效率均接近33%。该结果表明Rfo在BC4代可以稳定传递,但传递效率仍不正常,这可能是由于BC3代Rfo阳性单株的染色体组成仍为多倍体所导致。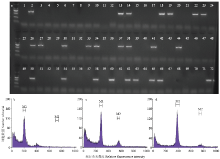 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图516Q1-4回交后代的Rfo筛选(a)和倍性鉴定(b-d)
a:16Q1-4的70个回交后代Rfo筛选 Rfo screening of 70 backcross progenies of 16Q1-4。1:芥蓝Chinese kale 15Y102;2:15Q23;3—72:16Q1-4的回交后代 70 backcross progenies of 16Q1-4。b:单株的荧光峰值位于200 The peaks of G0/G1 peak on 200 channels;c:单株的荧光峰值位于290 The peaks of G0/G1 peak on 290 channels;d:单株的荧光峰值位于390 The peaks of G0/G1 peak on 390 channels
-->Fig. 5Rfo screening (a) and ploidy identification (b-d) in the backcross progenies of 16Q1-4
-->
对16Q1-4的24株阳性株进行倍性检测(图5),不同单株间倍性差异较大,3株位于180—200(图5-b),14株阳性株峰值位于200—300(图5-c),7株位于300—400(图5-d)。上述结果表明,利用芥蓝对种间六倍体阳性株多代回交,其倍性存在分离,可从中获得倍性近于亲本芥蓝的材料,但其遗传组成仍需进一步分析。
3 讨论
3.1 芸薹属远缘杂交中种间三倍体的利用
甘蓝型油菜(AACC)是由甘蓝(CC)和白菜(AA)杂交后自然加倍获得的,三者同属十字花科芸薹属,亲缘关系较近[19](U,1935)。远缘杂交已成为三者间遗传背景拓展和种质资源改良的重要手段,但往往表现出杂交亲和障碍,杂种后代育性较差等问题,需要进行多轮胚挽救,多代回交和大量授粉工作才能克服[20,21,22,23,24]。白菜(AA)与甘蓝型油菜(AACC)间亲和性较强,较易获得F1种间杂种AAC,利用两者间远缘杂交后多代自交是甘蓝型油菜遗传背景拓展的重要手段之一[25,26],且前人研究表明F1后代AAC育性较高,花粉活力>50%,且在减数分裂时期,A染色体间配对稳定,整倍体配子出现频率较高(5%—15%,A/AC,n=10/19)[27,28]。不同于白菜AA和甘蓝型油菜AACC远缘杂交较易获得后代,甘蓝类蔬菜CC和甘蓝型油菜之间进行远缘杂交很难获得后代,前人相关研究较少,且很少进行多代回交转育[20-21,29]。此外,CC×AACC杂交后获得的种间杂种ACC单株育性较差,很难进行回交得到后代,张玉成等[30]利用结球甘蓝做父本与ACC单株进行杂交,结荚正常,但后代结实率仅为0.01%;YANG等[31]对ACC单株的育性和减数分裂行为进行调查,试验结果表明,ACC单株减数分裂时期染色体分离异常,且在A和C染色体组之间常出现异源联会,植株不育,无法获得后代;LI等[32]观察到ACC的花粉母细胞终变期配对形式多为9Ⅱ+10Ⅰ或10Ⅱ+8Ⅰ形式,在减数分裂后期多为不均等分离,配子以13﹕15和14﹕14的分离形式比例最高,有观察到出现9﹕19的分离形式,这表明ACC单株在花粉母细胞减数分裂时期可能会出现整倍体配子(C/AC)。在前期研究中,利用芥蓝对获得的种间F1三倍体育性恢复单株YL2进行了回交[1],获得了形态特征近于母本芥蓝的后代,但经Rfo特异标记检测,没有Rfo阳性株,全为不育类型。目前没有研究报道ACC可以与CC回交获得CC型后代。今后,在甘蓝与甘蓝型油菜的远缘杂交试验中,可尝试对种间杂种F1(ACC)进行大量正反交,期望直接分离出整倍体C型配子,从而缩短异源基因的转育年限。3.2 六倍体AACCCC在Rfo基因转育中的影响
前人研究表明,对CC和AACC获得的ACC杂种进行染色体加倍,获得AACCCC单株,可有效提高与AA或AACC基因型材料杂交的结实性[33,34]。AACCCC多数花粉母细胞在减数分裂时期多是28﹕28(18C+10A﹕18C+10A)分离,分离规律相对稳定[34],钱伟等[35]发明了以甘蓝型油菜与甘蓝杂交,染色体加倍获得的六倍体为桥梁再与白菜杂交,创建新型甘蓝型油菜的方法。利用该桥梁,将野生甘蓝中的菌核病抗性分别转移到白菜型油菜和甘蓝型油菜中,且后代结实正常[33,34];不同于AACCCC与CC或AACC杂交,AACCCC与CC杂交很难正常结实,且回交两代后染色体仍分离异常[2],在前期研究中,为保证Rfo传递,提高种间杂种单株育性,笔者将获得F1代种间Rfo阳性株进行加倍处理,并利用芥蓝对加倍后AACCCC单株进行第二代回交,20 733个种荚中仅获得98个单株,其中5株为Rfo阳性,结合形态、背景和育性比较,筛选出1株遗传背景、形态特征近于芥蓝、育性恢复稳定、可不进行胚挽救自然少量结实(结实率=7%)的BC2代单株15Q23[2]。本研究以该单株15Q23为父本,利用芥蓝对其进一步回交后获得126株Rfo阳性单株,Rfo可稳定传递(表2,TR=48%),BC3后代遗传背景较上一代单株15Q23更近于芥蓝,部分单株育性稳定,花粉活力>75%(表3,16Q1-4、16Q1-7、16Q1-10),单株16Q1-4和16Q1-10结实性较15Q23有显著提高(P<0.05)。后期倍性鉴定结果表明,15Q23为BC1代单株(AnCnCoCo)与芥蓝回交后获得,倍性近于四倍体(图4),是对照芥蓝的2倍,DNA相对含量高于甘蓝型油菜(AnCnAnCn)。根据倍性推测,15Q23组成形式可能为CoCoCnC*A*(C*和A*表示非整套染色体组),其中一套完整的Co来源于回交芥蓝,其余染色体是由于BC1代AnCnCoCo染色体配对异常,导致染色体不均等分离,BC1单株形成了CoCnC*A*型的配子,根据染色体组成形式和15Q23后代Rfo阳性株比例,推测Rfo位点的基因型可能为RfRfrfrf,分别位于4套C染色体组上。而15Q23回交后产生的BC3代Rfo阳性单株的倍性介于三倍体和四倍体之间,Rfo传递效率均接近33%(表3、图4),推测染色体组成形式可能为CoCoC*A*,Rfo位点的基因型可能为Rfrfrf,分别位于3套C染色体组上。因此,对ACC进行染色体加倍有利于提高杂种后代的育性以及保证Rfo的稳定传递,但BC3后代的染色体倍性还比较混杂,杂种后代的染色体要完全恢复到亲本芥蓝的水平仍需多代回交和大量授粉。然而,当利用BC3代育性较好的单株进一步回交获得BC4后代时,发现不仅Rfo可以继续稳定传递(表3),而且还有部分单株的倍性非常接近于亲本芥蓝(图5)。因此,在后续试验中,设想可结合小孢子培养和遗传组成分析,期望在多倍体Rfo阳性后代中分离出正常的CC配子;或对杂种后代中倍性近于二倍体的后代进行自交和正反交,加快甘蓝类蔬菜(2C=18)Ogura CMS恢复材料的获得。4 结论
利用芥蓝对芥蓝-甘蓝型油菜BC2代育性较好单株15Q23进一步回交,获得了BC3和BC4后代。Rfo特异标记筛选结果表明,Rfo在两个世代可稳定传递。倍性鉴定结果表明,BC4代倍性存在分离,部分Rfo阳性单株倍性接近于亲本芥蓝。结合背景分析、形态观察、育性比较,获得了两株育性稳定、形态特征近于芥蓝、遗传背景和结实性较上一代显著提升的BC3单株16Q1-4和16Q1-10。The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}