0 引言
【研究意义】小麦秆锈病由小麦秆锈菌(Puccinia graminis Pers. f. sp. tritici Eriks. and E. Henn)引起,是小麦的重要真菌病害之一。近30多年来,由于全球小麦普遍引入Sr31等抗秆锈病基因,小麦秆锈病得到了很好控制,基本上只在局部地区零星发生[1]。然而,1999年能够克服Sr31抗性的小麦秆锈菌小种Ug99的出现,使得世界小麦生产又重新处于秆锈病的威胁之下。Ug99除能克服Sr31的抗性外,还对其他多数重要抗秆锈病基因具有极罕见的联合致病力,全球90%以上的重要小麦生产品种都表现为高度感病[2]。2006年中国118份小麦品种送到肯尼亚进行Ug99抗性分析鉴定,其中只有2份材料表现抗性[3],由此可见中国小麦品种对Ug99的抗性较差。SINGH等[4]对52个抗秆锈病基因的Ug99抗性鉴定,只有22(42.3%)个基因对Ug99表现一定抗性,因此,发掘新抗源是中国应对Ug99以及其变异小种的最关键的任务之一[5,6,7]。【前人研究进展】迄今为止,小麦抗秆锈病基因已被正式命名至Sr59,并且还有许多未命名的抗病基因。小麦抗秆锈病基因来源广泛,除来自普通小麦种之外,还有来自小麦属的其他种,如一粒小麦、二粒小麦、硬粒小麦、提莫菲维小麦、阿拉拉特小麦以及小麦近缘属。其中,Sr24、Sr25、Sr26和Sr43来自长穗偃麦草(Agropyruon elongatum),Sr34来自于顶芒山羊草(Aegilops comosa),Sr38来自偏凸山羊草(Ae. ventricosa),Sr32、Sr39和Sr47来自拟斯卑尔脱山羊草(Ae. speltoides),Sr27、Sr31、Sr50和Sr59来自栽培黑麦(Secale cereale),Sr44来自中间偃麦草(Thinopyrum intermedium),Sr51来自希尔斯山羊草(Ae. searsii),Sr52来自簇毛麦(Dasypyrum villosum),Sr53来自卵穗山羊草(Ae. geniculata)[1,8]。SINGH等[4]鉴定的抗Ug99的基因中有19个来源于小麦亲缘种属。可见小麦近缘种属是小麦抗秆锈的重要基因源。【本研究切入点】小麦族含有300多个物种,其中小麦近缘物种250个以上[9],前人仅对小麦近缘种属的极少数物种进行了抗秆锈研究[1,8];另外,由于小麦与近缘种属之间的不亲和性等诸多因素,导致杂交成效较低,远缘种属的优异基因无法导入小麦,而小麦–近缘物种染色体系是向小麦转育近缘物种优异基因的良好载体和中间桥梁。因此,对较多的小麦-近缘植物染色体系进行抗秆锈鉴定,有利于筛选小麦秆锈病新抗源。【拟解决的关键问题】本研究利用中国小麦秆锈菌流行小种34MKGQM和21C3CTHSM对165份小麦-近缘植物染色体系进行苗期抗秆锈鉴定,与已报道的秆锈基因染色体定位信息比较,获得小麦秆锈病新抗源;筛选基于水稻EST序列设计的PLUG(PCR-base landmark unique gene)引物,建立抗病基因所在染色体的特异分子标记,为中国应对Ug99增加新的候选抗源,也为染色体工程诱导获得可用于小麦抗病育种的小麦-近缘植物染色体易位系奠定基础。1 材料与方法
1.1 材料
供试材料为165份小麦-近缘植物染色体系(附加系、代换系和易位系)和3份六倍体小麦(对照)(电子附表1)。TA编号材料由美国堪萨斯州立大学植物病理系Jon Raupp博士提供;JIC编号材料、CSDA1N-CSDA4N、6N、7N和XX029由英国约翰英纳斯中心READER S M教授提供;中国春(Chinese Spring,CS)由电子科技大学杨足君教授提供;感病对照小麦小密穗(little club,LC)和中国当前优势秆锈菌生理小种34MKGQM和21C3CTHSM(均采集于四川阿坝州)由沈阳农业大学植物保护学院植物病理教研室保存并提供。其中,34MKGQM的有效抗病基因包括Sr9e、Sr10、Sr11、Sr13、Sr14、Sr17、Sr18、Sr19、Sr20、Sr21、Sr23、Sr25、Sr26、Sr30、Sr31、Sr32、Sr33、Sr34、Sr35、Sr36、Sr37、Sr38、Sr47、Srdp-2、SrTmp、SrTt3和SrWld-1;21C3CTHSM的有效抗病基因包括Sr5、Sr9e、Sr19、Sr20、Sr21、Sr22、Sr23、Sr25、Sr26、Sr27、Sr30、Sr31、Sr32、Sr33、Sr36、Sr37、Sr38、Sr47和SrTmp。作为分子标记筛选对照的济麦19(JM19)、济麦20(JM20)、济麦22(JM22)、济麦23(JM23)、济麦262(JM262)和济麦229(JM229)由山东省农业科学院作物研究所小麦育种团队育成。Table S1
附表1
附表1供试168份种质的小麦感锈病苗期反应型
Table S1Wheat stem rust infection types of seedling stage of 168 germplasm
| 编号 Accession No. | 材料 Material | 侵染型 Infection type | 编号 Accession No. | 材料 Material | 侵染型 Infection type | |
|---|---|---|---|---|---|---|
| 中国春 | 中国春 T. aestivum CV. Chinese Spring (CS) | 4 | TA3601 | 中国春-帝国黑麦1R附加系 CS-Imperial rye 1R addition | 0 | |
| TA4072 | 小麦-顶芒山羊草易位系 Wheat-Ae. comosa translocation | 3 | TA3603 | 中国春-帝国黑麦2R附加系 CS-Imperial rye 2R addition | 4 | |
| ALCD | 小麦品种Alcedo Wheat variety Alcedo | 3 | TA3604 | 中国春-帝国黑麦3R附加系 CS-Imperial rye 3R addition | 4 | |
| TA3558 | ALCD-尾状山羊草2C#1附加系 ALCD-Ae. caudata 2C#1 addition | 4 | TA3605 | 中国春-帝国黑麦4R附加系 CS-Imperial rye 4R addition | 4 | |
| TA3559 | ALCD-尾状山羊草5C#1附加系 ALCD-Ae. caudata 5C#1 addition | 3- | TA3606 | 中国春-帝国黑麦5R附加系 CS-Imperial rye 5R addition | 4 | |
| TA3560 | ALCD-尾状山羊草6C#1附加系 ALCD-Ae. caudata 6C#1 addition | 3- | TA3608 | 中国春-帝国黑麦6R附加系 CS-Imperial rye 6R addition | 1 | |
| TA3562 | ALCD-尾状山羊草7C#1附加系 ALCD-Ae. caudata 7C#1 addition | 1+ | TA3609 | 中国春-帝国黑麦7R附加系 CS-Imperial rye 7R addition | 4 | |
| TA3561 | ALCD-尾状山羊草3C#1附加系 ALCD-Ae. caudata3C#1 addition | 4 | TA3681 | 中国春-中间偃麦草?Ai附加系 CS- Th. intermedium?Ai addition | 0 | |
| TA3563 | ALCD-尾状山羊草4C#1附加系 ALCD-Ae. caudata4C#1 addition | 4 | TA5566 | 中国春-中间偃麦草?Ai附加系 CS- Th. intermedium?Ai addition | 0 | |
| JIC-46 | 小麦品种Hobbit 'sib' Wheat variety Hobbit 'sib' | 4 | TA5567 | 国春-中间偃麦草?Ai附加系 CS- Th. intermedium?Ai addition | 4 | |
| JIC-10 | Hobbit 'sib'-顶芒山羊草2M 附加系 Hobbit 'sib'-Ae. comosa 2M addition | 4 | TA3664 | 中国春-长穗偃麦草1E附加系 CS-A. elongatum 1E addition | 4 | |
| JIC-11 | Hobbit 'sib'-顶芒山羊草2M(2D)代换系 Hobbit 'sib'-Ae. comosa 2M(2D) substitution | 4 | TA3665 | 中国春-长穗偃麦草2E附加系 CS-A. elongatum 2E addition | 3 | |
| JIC-12 | Hobbit 'sib'-顶芒山羊草2M/2A易位系 Hobbit 'sib'-Ae. comosa 2M/2A translocation | 4 | TA3666 | 中国春-长穗偃麦草3E附加系 CS-A. elongatum 3E addition | 4 | |
| JIC-17 | Hobbit 'sib'-顶芒山羊草2M/2D易位系 Hobbit 'sib'-Ae.comosa 2M/2D translocation | 4 | TA3667 | 中国春-长穗偃麦草4E附加系 CS-A. elongatum 4E addition | 4 | |
| JIC-38 | Holdfast-二角山羊草3Sb附加系 Holdfast -Ae. bicornis 3Sb addition | 4 | TA3704 | 中国春-长穗偃麦草5E附加系 CS-A. elongatum 5E addition | 3 | |
| JIC-40 | Holdfast-二角山羊草3Sb(3A )代换系 Holdfast -Ae.bicornis 3Sb(3A) substitution | 4 | TA3668 | 中国春-长穗偃麦草6E附加系 CS-A. elongatum 6E addition | 4 | |
| JIC-41 | Holdfast-二角山羊草3Sb(3B)代换系 Holdfast -Ae.bicornis 3Sb(3B) substitution | 4 | TA3706 | 中国春-长穗偃麦草7E附加系 CS-A. elongatum 7E addition | 3 | |
| JIC-42 | Holdfast-二角山羊草3Sb(3D)代换系 Holdfast -Ae.bicornis 3Sb(3D) substitution | 4 | TA3698 | 中国春-大麦2H附加系 CS-Barley 2H addition | 4 | |
| JIC-44 | Holdfast-二角山羊草7Sb(7B)代换系 Holdfast -Ae. bicornis 7Sb(7B)substitution | 4 | TA3699 | 中国春-大麦3H附加系 CS-Barley 3H addition | 4 | |
| TA7552 | 中国春-粗穗披碱草1Ht附加系 CS-E. trachycaulus 1Ht addition | 4 | TA3700 | 中国春-大麦4H附加系 CS-Barley 4H addition | 4 | |
| TA7557 | 中国春-粗穗披碱草5Ht附加系 CS-E. trachycaulus 5Ht addition | 4 | TA3701 | 中国春-大麦5H附加系 CS-Barley 5H addition | 4 | |
| TA7558 | 中国春-粗穗披碱草6Ht附加系 CS-E. trachycaulus 6Ht addition | 4 | TA3702 | 中国春-大麦6H附加系 CS-Barley 6H addition | 3 | |
| TA7559 | 中国春-粗穗披碱草7Ht附加系 CS-E. trachycaulus 7Ht addition | 4 | TA3697 | 中国春-大麦7H单体附加系 CS-Barley 2H monosomic addition | 4 | |
| TA7556 | 中国春-粗穗披碱草1St附加系 CS-E. trachycaulus 1St addition | 4 | TA7562 | 中国春-小伞山羊草1U#1附加系 CS-Ae. umbellulata1U#1addition | 4 | |
| 续附表1 Continued table S1 | ||||||
| 编号 Accession No. | 材料 Material | 侵染型 Infection type | 编号 Accession No. | 材料 Material | 侵染型 Infection type | |
| TA5532 | 中国春-粗穗披碱草T2HtS.5HtL附加系 CS-E. trachycaulus T2HtS.5HtLaddition | 4 | TA7564 | 中国春-小伞山羊草4U#1单体附加系 CS-Ae.umbellulata 4U#1monosomic addition | 4 | |
| TA7580 | 中国春-粗穗披碱草5St单体附加系 CS-E. trachycaulus 5St monosomic addition | 4 | TA7565 | 中国春-小伞山羊草5U#1附加系 CS-Ae. umbellulata 5U#1addition | 4 | |
| TA5072 | 中国春-粗穗披碱草1HtS.1BL罗伯逊易位系 CS-E. trachycaulus 1HtS.1BLRobertsonian translocation | 4 | TA7566 | 中国春-小伞山羊草6U#1附加系 CS-Ae. umbellulata 6U#1addition | 4 | |
| TA7543 | 中国春-高大山羊草1Sl#3附加系 CS- Ae. longissima 1Sl#3 addition | 4 | TA7567 | 中国春-小伞山羊草7U#1附加系 CS-Ae. umbellulata 7U#1addition | 4 | |
| TA7544 | 中国春-高大山羊草2Sl#3附加系 CS- Ae. longissima 2Sl#3 addition | 4 | TA7643 | 中国春-大赖草2Lr#1附加系 CS- L. racemosus 2Lr#1 addition | 4 | |
| TA7545 | 中国春-高大山羊草3Sl#3附加系 CS- Ae. longissima 3Sl#3 addition | 4 | TA7648 | 中国春-大赖草7Lr#1附加系 CS- L. racemosus 7Lr#1 addition | 4 | |
| TA7546 | 中国春-高大山羊草4Sl#3附加系 CS- Ae. longissima 4Sl#3 addition | 4 | TA7649 | 中国春-大赖草7Lr#1S端体附加系 CS- L. racemosus 7Lr#1S monosomic addition | 4 | |
| TA7547 | 中国春-高大山羊草5Sl#3附加系 CS- Ae. longissima 5Sl#3 addition | 4 | CSDA1N | 中国春-单芒山羊草1N附加系 CS- Ae. uniaristata 1N addition | 4 | |
| TA7548 | 中国春-高大山羊草6Sl#3附加系 CS- Ae. longissima 6Sl#3 addition | 1 | CSDA2N | 中国春-单芒山羊草2N附加系 CS- Ae. uniaristata 2N addition | 3 | |
| TA7549 | 中国春-高大山羊草7Sl#3单体附加系 CS- Ae. longissima 7Sl#3 addition | 4 | CSDA3N | 中国春-单芒山羊草3N附加系 CS- Ae. uniaristata 3N addition | 4 | |
| TA3573 | 中国春-高大山羊草1Sl#2附加系 CS- Ae. longissima 1Sl#2 addition | 4 | CSDA4N | 中国春-单芒山羊草4N附加系 CS- Ae. uniaristata 4N addition | 4 | |
| TA3575 | 中国春-高大山羊草3Sl#2附加系 CS- Ae. longissima 3Sl#2 addition | 4 | 6N | 中国春-单芒山羊草6N附加系 CS- Ae. uniaristata 6N addition | 0 | |
| TA3577 | 中国春-高大山羊草5Sl#2附加系 CS- Ae. longissima 5Sl#2 addition | 4 | 7N | 中国春-单芒山羊草7N附加系 CS- Ae. uniaristata 7N addition | 4 | |
| TA 6506 | 中国春-高大山羊草2Sl#2(2D)代换系 CS- Ae. longissima 2Sl#2(2D) substitution | 4 | TA7594 | 中国春-易变山羊草1Sv#1附加系 CS-Ae. variabilis 1Sv#1 addition | 4 | |
| TA 6512 | 中国春-高大山羊草4Sl#2(4D)代换系 CS- Ae.longissima 4Sl#2(4D)substitution | 4 | TA7595 | 中国春-易变山羊草2Sv#1附加系 CS-Ae. variabilis 2Sv#1 addition | 4 | |
| TA6516 | 中国春-高大山羊草6Sl#2(6B)代换系 CS- Ae. longissima 6Sl#2(6B)substitution | 1+ | TA7596 | 中国春-易变山羊草3Sv#1附加系 CS-Ae. variabilis 3Sv#1 addition | 4 | |
| TA3580 | 中国春-希尔斯山羊草1Ss#1附加系 CS-Ae. searsii 1Ss#1 addition | 4 | TA7597 | 中国春-易变山羊草4Sv#1附加系 CS-Ae. variabilis 4Sv#1 addition | 4 | |
| TA3581 | 中国春-希尔斯山羊草2Ss#1附加系 CS-Ae. searsii 2Ss#1 addition | 4 | TA7598 | 中国春-易变山羊草5Sv#1附加系 CS-Ae. variabilis 5Sv#1 addition | 4 | |
| TA3582 | 中国春-希尔斯山羊草3Ss#1附加系 CS-Ae. searsii 3Ss#1 addition | 1- | TA7599 | 中国春-易变山羊草6SvS端体附加系 CS-Ae. variabilis 6SvS telosomic addition | 0 | |
| TA3583 | 中国春-希尔斯山羊草4Ss#1附加系 CS-Ae. searsii 4Ss#1 addition | 4 | TA7600 | 中国春-易变山羊草7Sv#1附加系 CS-Ae. variabilis 7Sv#1 addition | 4 | |
| TA3584 | 中国春-希尔斯山羊草5Ss#1附加系 CS-Ae. searsii 5Ss#1 addition | 3+ | TA7614 | 中国春-易变山羊草1Uv#1附加系 CS-Ae. variabilis 1Uv#1 addition | 4 | |
| TA3585 | 中国春-希尔斯山羊草6Ss#1附加系 CS-Ae. searsii 6Ss#1 addition | 4 | TA7615 | 中国春-易变山羊草2Uv#1附加系 CS-Ae. variabilis 2Uv#1 addition | 4 | |
| TA3586 | 中国春-希尔斯山羊草7Ss#1附加系 CS-Ae. searsii 7Ss#1 addition | 4 | TA7616 | 中国春-易变山羊草3Uv#1附加系 CS-Ae. variabilis 3Uv#1 addition | 4 | |
| 续附表1 Continued table S1 | ||||||
| 编号 Accession No. | 材料 Material | 侵染型 Infection type | 编号 Accession No. | 材料 Material | 侵染型 Infection type | |
| TA7718 | 中国春-沙融山羊草4Ssh#8附加系 CS-Ae. sharonensis 4Ssh#8 addition | 4 | TA7617 | 中国春-易变山羊草4Uv#1附加系 CS-Ae. variabilis4Uv#1 addition | 4 | |
| JIC-32 | 中国春-沙融山羊草2Ssh附加系 CS-Ae. sharonensis 2Ssh addition | 4 | TA7618 | 中国春-易变山羊草5Uv#1附加系 CS-Ae. variabilis 5Uv#1 addition | 4 | |
| JIC-33 | 中国春-沙融山羊草4Ssh附加系 CS-Ae. sharonensis 4Ssh addition | 4 | TA7619 | 中国春-易变山羊草6Uv1附加系 CS-Ae. variabilis6Uv#1 addition | 4 | |
| JIC-36 | 中国春-沙融山羊草6Ssh附加系 CS-Ae. sharonensis 6Ssh addition | 4 | TA7705 | 中国春-纤毛披碱草2Sc附加系 CS-El.ciliaris 2Sc addition | 4 | |
| JIC-37 | 中国春-沙融山羊草7Ssh附加系 CS-Ae. sharonensis 7Ssh addition | 4 | TA7706 | 中国春-纤毛披碱草3Sc附加系 CS-El.ciliaris 3Sc addition | 4 | |
| TA7689 | 中国春-拟斯卑尔脱山羊草1S#3附加系 CS- Ae. speltoides 1S#3 addition | 4 | TA7707 | 中国春-纤毛披碱草7Sc附加系 CS-El.ciliaris 7Sc addition | 4 | |
| TA7690 | 中国春-拟斯卑尔脱山羊草2S#3附加系 CS- Ae. speltoides 2S#3 addition | 1+ | TA7584 | 中国春-纤毛披碱草1Yc附加系 CS-El.ciliaris 1Yc addition | 4 | |
| TA7691 | 中国春-拟斯卑尔脱山羊草3S#3附加系 CS- Ae. speltoides 3S#3 addition | 4 | TA7708 | 中国春-纤毛披碱草5Yc附加系 CS-El.ciliaris 5Yc addition | 4 | |
| TA7692 | 中国春-拟斯卑尔脱山羊草4S#3附加系 CS- Ae. speltoides 4S#3 addition | 4 | TA7709 | 中国春-纤毛披碱草7Yc附加系 CS-El.ciliaris 7Yc addition | 4 | |
| TA7693 | 中国春-拟斯卑尔脱山羊草5S#3附加系 CS- Ae. speltoides 5S#3 addition | 3 | TA7684 | 中国春-筑紫披碱草1Ets#1附加系 CS-E.tsukushienis 1Ets#1 addition | 4 | |
| TA7694 | 中国春-拟斯卑尔脱山羊草6S#3附加系 CS- Ae. speltoides6S#3 addition | 4 | TA7685 | 中国春-筑紫披碱草3Ets#1附加系 CS- E. tsukushienis 3Ets#1 addition | 4 | |
| TA7695 | 中国春-拟斯卑尔脱山羊草7S#3附加系 CS- Ae. speltoides 7S#3 addition | 4 | TA7687 | 中国春-筑紫披碱草5Ets#1单体附加系 CS- E. tsukushienis 5Ets#1 monosomic addition | 4 | |
| TA7655 | 中国春-卵穗山羊草1Mg#1附加系 CS- Ae. geniculata 1Mg#1 addition | 4 | TA5660 | 中国春-筑紫披碱草T1AL.1AS-1Ets#1S易位系 CS- E. tsukushienis T1AL.1AS-1Ets#1S translocation | 4 | |
| TA7656 | 中国春-卵穗山羊草2Mg#1附加系 CS- Ae. geniculata 2Mg#1 addition | 4 | TA5661 | 中国春-筑紫披碱草TiWL-1Ets#1S-WS易位系 CS-E. tsukushienis TiWL-1Ets#1S-WS translocation | 4 | |
| TA7657 | 中国春-卵穗山羊草3Mg#1附加系 CS- Ae. geniculata 3Mg#1 addition | 4 | TA6664 | 中国春-提莫菲维2G(2B)代换系 CS-T. timopheevii 2G(2B)substitution | 3 | |
| TA7658 | 中国春-卵穗山羊草4Mg#1附加系 CS- Ae. geniculata 4Mg#1 addition | 4 | TA6666 | 中国春-提莫菲维4G(4B)代换系 CS-T. timopheevii 4G(4B)substitution | 3 | |
| TA7659 | 中国春-卵穗山羊草5Mg#1附加系 CS- Ae. geniculata 5Mg#1 addition | 3- | TA6667 | 中国春-提莫菲维5G(5B)代换系 CS-T. timopheevii 5G(5B)substitution | 4 | |
| TA7660 | 中国春-卵穗山羊草6Mg#1附加系 CS- Ae. geniculata 6Mg#1 addition | 4 | JIC-2 | 中国春-顶芒山羊草2/7M附加系 CS- Ae. comosa 2/7M addition | 4 | |
| TA7661 | 中国春-卵穗山羊草7Mg#1附加系 CS- Ae. geniculata 7Mg#1 addition | 0 | JIC-3 | 中国春-顶芒山羊草 2M附加系 CS- Ae. \comosa 2M addition | 3 | |
| TA7662 | 中国春-卵穗山羊草1Ug#1附加系 CS- Ae. geniculate 1Ug#1 addition | 3 | JIC-4 | 中国春-顶芒山羊草3M附加系 CS- Ae. comosa 3M addition | 4 | |
| TA7663 | 中国春-卵穗山羊草2Ug#1附加系 CS- Ae. geniculata 2Ug #1 addition | 3+ | JIC-5 | 中国春-顶芒山羊草4M附加系 CS- Ae. comosa 4M addition | 4 | |
| TA7688 | 中国春-卵穗山羊草3Ug#1单体附加系 CS- Ae. geniculata 3Ug #1 monosomic addition | 4 | JIC-6 | 中国春-顶芒山羊草5M附加系 CS- Ae. comosa 5M addition | 3 | |
| 续附表1 Continued table S1 | ||||||
| 编号 Accession No. | 材料 Material | 侵染型 Infection type | 编号 Accession No. | 材料 Material | 侵染型 Infection type | |
| TA7664 | 中国春-卵穗山羊草4Ug#1附加系 CS- Ae. geniculate 4Ug#1 addition | 4 | JIC-7 | 中国春-顶芒山羊草6M附加系 CS- Ae. comosa 6M addition | 4 | |
| TA7665 | 中国春-卵穗山羊草5Ug#1附加系 CS- Ae. geniculata 5Ug#1 addition | 4 | JIC-8 | 中国春-顶芒山羊草7M附加系 CS- Ae. comosa 7M addition | 3 | |
| TA7666 | 中国春-卵穗山羊草6Ug#1附加系 CS- Ae. geniculata 6Ug #1 addition | 4 | JIC-21 | 中国春-无芒山羊草2T?附加系 CS- Ae. mutica 2T? addition | 4 | |
| TA6647 | 中国春-卵穗山羊草7Mg#1(7B)代换系 CS- Ae. geniculata 7Mg#1(7B) addition | 0 | JIC-25 | 中国春-无芒山羊草7T附加系 CS- Ae. mutica 7T addition | 4 | |
| TA6648 | 中国春-卵穗山羊草7Mg#1(7D)代换系 CS- Ae. geniculata 7Mg#1(7D) addition | 1 | JIC-27 | 中国春-无芒山羊草7T?附加系 CS- Ae. mutica 7T?addition | 3 | |
| TA7725 | 中国春-两芒山羊草1Ubi#1附加系 CS-Ae. biuncialis 1Ubi#1 addition | 4 | JIC-29 | 中国春-无芒山羊草2T?附加系 CS- Ae. mutica 2T? addition | 4 | |
| TA7726 | 中国春-两芒山羊草2Ubi#1附加系 CS-Ae. biuncialis 2Ubi#1 addition | 4 | XX029 | 中国春-智利大麦1Hch+1HchS附加系 CS-Chile barley 1Hch+1HchS addition | 4 | |
| TA7729 | 中国春-两芒山羊草5Ubi#1附加系 CS-Ae. biuncialis 5Ubi#1 addition | 4 | TA7588 | 中国春-智利大麦4Hch附加系 CS-Chile barley 4Hch addition | 4 | |
| TA7733 | 中国春-两芒山羊草2Mbi#1附加系 CS-Ae. biuncialis 2Mbi#1 addition | 4 | TA7589 | 中国春-智利大麦5Hch附加系 CS-Chile barley 5Hch addition | 4 | |
| TA7734 | 中国春-两芒山羊草3Mbi#1附加系 CS-Ae. biuncialis 3Mbi#1 addition | 3+ | TA7590 | 中国春-智利大麦6Hch附加系 CS-Chile barley 6Hch addition | 0 | |
| TA7735 | 中国春-两芒山羊草4Mbi#1单体附加系 CS-Ae. biuncialis 4Mbi#1 monosomic addition | 4 | TA7591 | 中国春-智利大麦7Hch附加系 CS-Chile barley 7Hch addition | 4 | |
“0” Immune; “;” Nearly immune; “1” Highly resistance; “2” Moderate resistance; “3” Medium susceptible; “4” Highly susceptible; “+” Uredinia are larger than expected; “-” Uredinia are smaller then expected
“0”免疫;“;”近免疫;“1”高抗;“2”中抗;“3”中感;“4”高感;“+”比预想的孢子堆大;“-”比预想的孢子堆小
新窗口打开
1.2 秆锈病苗期抗性鉴定
试验于2016年10月至2017年3月在玻璃温室内进行,供试小麦材料按编号播种于直径10 cm的瓦盆中,以感病品种LC为对照,在温室中当幼苗长至一叶一心时(叶龄期在7 d左右),进行接种鉴定:将新鲜的按照1﹕1(v/v)混合的34MKGQM和21C3CTHSM用干燥滑石粉以1﹕10(v/v)稀释后喷粉接种于已喷雾(0.01%吐温20)的幼苗上,于16—18℃下保湿12—14 h后,置于18—20℃的温室中培养,待感病对照LC充分发病后进行抗病性鉴定。鉴定设3次重复,分别调查记载。侵染型按0—4级进行记载[9],其中,0为免疫,;为近免疫,1为高抗,2为中抗,3为中感,4为高感[7]。1.3 DNA提取与PLUG-PCR
取供试材料的一叶一心期幼叶,利用SDS方法[10]提取其基因组DNA。PLUG引物序列参照文献[11],由成都瑞信生物公司合成。反应体系包含2.0 μL 10×PCR buffer(含Mg2+)、2.0 μL 2.5 mmol·L-1 dNTP,上下游引物各1 μL(10 μmol·L-1)、0.2 μL5 U·μL-1Taq酶、50—100 ng基因组DNA,加ddH2O补至20 μL。反应程序为94℃ 3 min;94℃ 45 s,57℃ 45 s,72℃ 2 min,35个循环;72℃ 10 min。PCR产物分别用限制性内切酶TaqⅠ和HaeⅢ进行酶切,酶切产物用2%琼脂糖凝胶电泳进行检测。1.4 染色体特异PLUG标记的建立
利用免疫、近免疫和高抗小麦秆锈病的小麦-近缘植物染色体系和中国春为材料筛选染色体第6和7同源群的101对PLUG引物。相比中国春,选择能在小麦-近缘物种染色体系中扩增出多态性的引物。将上述引物用于扩增所涉及近缘物种的整套染色体系,确定相应扩增多态性片段为物种该染色体特异标记。2 结果
2.1 小麦-近缘植物染色体系抗秆锈鉴定
用当前中国小麦秆锈菌流行小种34MKGQM和21C3CTHSM的混合菌种对供试的168份材料进行抗秆锈鉴定,结果显示,中国春-卵穗山羊草7Mg#1附加系、中国春-卵穗山羊草7Mg#1(7A)和7Mg#1(7B)代换系、中国春-帝国黑麦1R附加系、中国春-中间偃麦草?Ai附加系(TA3681和TA5566)(?表示未鉴定同源群)、中国春-单芒山羊草6N附加系、中国春-易变山羊草6SvS端体附加系和中国春-智利大麦6Hch附加系等9份材料对秆锈病表现为免疫或近免疫(电子附表1);ALCD-尾状山羊草7C#1附加系、中国春-卵穗山羊草7Mg#1(7D)代换系、中国春-帝国黑麦6R附加系、中国春-高大山羊草6Sl#3附加系、中国春-高大山羊草6Sl#2(6B)代换系、中国春-希尔斯山羊草3S#1附加系和中国春-拟斯卑尔脱山羊草2Sg#3附加系等7份材料对秆锈病表现为高抗;其余材料均表现为中感或高感(电子附表1)。对照六倍体小麦ALCD和中国春对小麦秆锈病分别表现为中感和高感,ALCD-尾状山羊草2C—6C#1附加系、中国春-高大山羊草1Sl#3—5Sl#3、7Sl#3附加系、中国春-高大山羊草1Sl#2、3Sl#2、5Sl#2附加系、2Sl#2(2D)和4Sl#2(4D)代换系、中国春-卵穗山羊草1Mg-6Mg附加系和1Ug—7Ug附加系对小麦秆锈病均表现为中感和高感,而ALCD-尾状山羊草7C#1附加系、中国春-卵穗山羊草7Mg#1附加系、中国春-卵穗山羊草7Mg#1(7A)、7Mg#1(7B)、7Mg#1(7D)代换系、中国春-高大山羊草6Sl#3附加系和中国春-高大山羊草6Sl#2(6B)代换系对小麦秆锈病均表现为高抗(电子附表1),表明尾状山羊草7C染色体、卵穗山羊草7Mg#1染色体、高大山羊草6Sl#2和6Sl#3染色体上均含有抗秆锈基因。
中国春-希尔斯山羊草3Ss#1附加系、中国春-拟斯卑尔脱山羊草2S#3附加系和中国春-单芒山羊草6N附加系分别对小麦秆锈病表现为高抗和近免疫,而对照中国春、中国春-希尔斯山羊草1Ss#1、2Ss#1、4Ss#1—7Ss#1附加系、中国春-拟斯卑尔脱山羊草1Sg#3、3Sg#3—7Sg#3附加系、中国春-单芒山羊草1N—4N和7N附加系均中感或高感秆锈病(附表1),这表明,希尔斯山羊草3Ss#1染色体、拟斯卑尔脱山羊草2S#3染色体和单芒山羊草6N染色体上均含有抗小麦秆锈病基因。中国春-希尔斯山羊草3Ss#1附加系抗病可能是因为在希尔斯山羊草3Ss上存在抗秆锈基因Sr51的缘故;中国春-拟斯卑尔脱山羊草2S#3附加系抗病可能是因为拟斯卑尔脱山羊草2S上存在有抗病基因Sr32、Sr39和Sr47的原因。因为单芒山羊草N染色体是偏凸山羊草Nv染色体组的供体[12],而偏凸山羊草6Nv染色体含有抗小麦秆锈基因Sr38,因此,中国春-单芒山羊草6N附加系近免疫小麦秆锈病,可能是6N染色体上含有Sr38的缘故。
对照小麦中国春、中国春-帝国黑麦2R—5R和7R附加系高感秆锈病,中国春-帝国黑麦1R和6R附加系分别对小麦秆锈病表现为免疫和高抗(电子附表1),说明帝国黑麦1R和6R含有抗秆锈病基因,前者抗秆锈病可能是因为1R染色体上存在抗秆锈基因Sr31和Sr50。中国春-智利大麦1Hch+1HchS、4Hch—5Hch附加系高感小麦秆锈病,但中国春-智利大麦6Hch附加系表现为高抗(电子附表1),说明智利大麦6Hch含有抗2个小种的基因。2份中国春-中间偃麦草?Ai附加系(TA3681和TA5566)均近免疫小麦秆锈病(电子附表1),说明中间偃麦草?Ai染色体上含有抗秆锈基因。
2.2 含抗秆锈基因材料中外源染色体多态性引物筛选
为了建立秆锈病抗性较好材料中外源染色体特异的标记,利用中国春-高大山羊草6Sl#3附加系、中国春-高大山羊草6Sl#2(6B)代换系、中国春-帝国黑麦6R附加系、中国春-单芒山羊草6N附加系、中国春-易变山羊草6SvS端体附加系、中国春-智利大麦6Hch附加系、中国春和济麦系列小麦品种对染色体第6同源群的53对PLUG引物进行筛选。结果发现,相比小麦对照,共有13对引物可在相应附加/代换系中扩增出额外的多态性片段(表1),其中,TNAC1740、TNAC1751和TNAC1756能够同时在中国春-高大山羊草6Sl#3附加系和6Sl#2(6B)代换系中扩增出多态性片段。TNAC1715、TNAC1718和TNAC1676等10对引物可在中国春-帝国黑麦6R附加系中扩增出多态性片段,其中TNAC1676等5对引物已被文献[13,14]报道,因此,TNAC1715等5对引物扩增的多态性片段是黑麦6R染色体的新标记。利用含卵穗山羊草7Mg#1染色体的材料与小麦对照筛选第7同源群的48对PLUG引物,未获得多态性。TNAC1740、TNAC1751、TNAC1715、TNAC1718、TNAC1753和TNAC1748引物的扩增结果如图1所示。Table 1
表1
表113个特异PLUG标记信息
Table 1Detailed information of 13 PLUG markers
| 序号 No. | 引物 Primer | 序列 Sequence (5′-3′) | 染色体位置 Chromosomal location | 所用内切酶 Enzyme used | 标记长度 Product size (bp) | |
|---|---|---|---|---|---|---|
| 小麦 T. aestivum | 外源物种 Alien species | |||||
| 1 | TNAC1740 | F:CGGAAGTGCTCGATTGTATCT R:GCGGGTTTCTTCTCAACCTT | 6AL7-0.88-0.90 6BL5-0.40-0.66 6DL6-0.29-0.47 | 6Sl | TaqⅠ | 1100 |
| 2 | TNAC1751 | F:CTTCCTTTGCTTGTGATCCTG R:GCCTGAGGACTTGAAGTGGTA | 6AL8-0.90-1.00 6BL1-0.70-1.00 6DL12-0.68-0.74 | 6Sl | TaqⅠ | 700 |
| 3 | TNAC1756 | F:CTCCATGGACAATTCCTGCTA R:AAGGCCAGTTCCAGATTCAGT | 6B | 6Sl | TaqⅠ | 700 |
| 4 | TNAC1715 | F:CGGTAAAGATAGCCATCATCG R:TTGAGATGACCGTGAGGAATG | 6B | 6R | HaeⅢ | 750 550 |
| 5 | TNAC1718 | F:CAAGGTGATCTCAAGCTTCGT R:AGTCCAAAGAACGCACAAAGA | 6D | 6R | HaeⅢ | 750 |
| 6 | TNAC1737 | F:CCCGCTGTAGACATCCTCTCT R:GGACGATGGTCGGAATCTT | 6B 6D | 6R | HaeⅢ | 750 |
| 7 | TNAC1739 | F:ACATCGAGAAGATCGAGTTGC R:TGGAAGCCCAGTTCTCCTTAT | 6B 6D | 6R | TaqⅠ | 500 |
| 8 | TNAC1753 | F:TCTGAGCTCCTTCACAAGAGG R:CCTGCGCTTACATCTGAAACT | 6A | 6R | HaeⅢ | 500 |
| 9 | TNAC1676※ | F:ATTGACGCGATTGATAGTGAA R:AGTGGGCTACTGCTTCAGATG | 6AS5-0.65-1.006 6BS-Sat 6DS4-0.79-0.99 | 6R | HaeⅢ | 1100 |
| 10 | TNAC1678※ | F:AAATCTACTCTGCGAGGTTTGC R:TGGGAAGCGCTTACTATTGTG | 6AS5-0.65-1.00 6BS-Sat 6DS4-0.79-0.99 | 6R | HaeⅢ | 1000 |
| 11 | TNAC1685※ | F:ATGGATAGCGGAAGCGACTC R:AGCGTTTCCTCCGGTCTT | C-6AS1-0.35 6BS1-0.25-0.46 6DS1-0.54-0.79 | 6R | TaqⅠ | 1100 |
| 12 | TNAC1743※ | F:CAGTTGATCAGGGCATTCAGT R:TTCCAGTCCTTTGTTTCCACA | 6AL7-0.88-0.90 6BL8-0.66-0.70 6DL1-0.47-0.68 | 6R | TaqⅠ | 300 |
| 13 | TNAC1748※ | F:TCGTAGAATTGGTCGACGATG R:ATGGATTGGCAAAGAAAGATG | 6AL7-0.88-0.90 6BL8-0.66-0.70 6DL1-0.47-0.68 | 6R | TaqⅠ | 200 450 |
新窗口打开
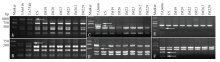 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1引物TNAC1740(A)、TNAC1751(B)、TNAC1715(C)、TNAC1718(D)TNAC1753(E)和TNAC1748(F)的扩增
-->Fig. 1PCR amplification patterns of primer pairs TNAC1740 (A), TNAC1751 (B), TNAC1715 (C), TNAC1718 (D), TNAC1753 (E) and TNAC1748 (F)
-->
2.3 抗秆锈基因所在染色体特异分子标记的建立
为了确定上述多态性片段是相应外源物种染色体特异标记,利用引物TNAC1740、TNAC1751和TNAC1756对中国春-高大山羊草1Sl#3—7Sl#3附加系、中国春-高大山羊草1Sl#2、3Sl#2和5Sl#2附加系、中国春-高大山羊草2Sl#2(2D)、4Sl#2(4D)和6Sl#2 (6B)代换系进行扩增(图2),发现6Sl#3附加系和6Sl#2(6B)代换系均能扩增出多态性片段,说明3对引物扩增出的多态性片段是高大山羊草6Sl染色体特异标记。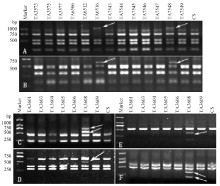 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2引物TNAC1740(A)、TNAC1751(B)、TNAC1715(C)、TNAC1718(D)TNAC1753(E)和TNAC1748(F)所获多态性条带的染色体定位
-->Fig. 2Chromosome localization of polymorphism bands that primer pairs TNAC1740 (A), TNAC1751 (B),TNAC1715 (C), TNAC1718 (D), TNAC1753 (E) and TNAC1748 (F) amplified
-->
利用TNAC1715等10对PLUG引物对中国春-帝国黑麦1R—7R附加系进行扩增(图2),发现这10对引物仅在中国春-帝国黑麦6R附加系中扩增出多态性片段,说明这10对引物扩增的多态性片段是帝国黑麦6R染色体的特异标记。
3 讨论
3.1 高大山羊草6Sl染色体抗秆锈病新基因
高大山羊草(Ae. longissima)染色体组为SlSl,高抗小麦白粉病、叶锈病、麦二叉蚜和眼斑病,抗干旱胁迫和盐胁迫[15,16],并能够显著提高小麦籽粒Fe和Zn元素含量。WANG等[17]和GARG等[18]发现高大山羊草1Sl染色体导入小麦可显著提高小麦品质;NEELAM等[19]和SHARMA等[20]先后从小麦-高大山羊草杂交后代中选育出了籽粒微量元素含量高的后代材料。CENCI等[21]发现高大山羊草3Sl染色体短臂上含有抗白粉病基因Pm13;SHENG等[22]发现高大山羊草1Sl、3Sl和5Sl上含有抗眼斑病基因;然而,未见高大山羊草染色体上存在抗秆锈基因的报道。本研究发现中国春-高大山羊草6Sl#3附加系和中国春-高大山羊草6Sl#2(6B)代换系均高抗两个秆锈生理小种34MKGQM和21C3CTHSM,说明高大山羊草6Sl上含有抗秆锈新基因。
3.2 黑麦6R染色体抗秆锈病新基因
黑麦是最早也是最成功地用于改良小麦的近缘植物之一,具有诸多优良性状,如抗病虫、抗逆、抗倒伏、分孽力强等。黑麦基因组中的抗白粉病基因Pm1~Pm8、Pm17、Pm20和抗锈基因Yr9、Lr25、Lr26、Sr27、Sr31、Sr50、Sr59已被导入小麦被广泛应用[23,24,25,26],其中的抗秆锈基因Sr27、Sr31、Sr50和Sr59分别位于黑麦3R、1RS、1RS和2RL染色体上。本研究发现中国春-帝国黑麦1R和6R附加系分别免疫和高抗小麦秆锈病,而2R—5R和7R附加系高感秆锈病。小麦-帝国黑麦1R附加系表现为免疫的原因可能是1R染色体上含有的Sr31是2个生理小种的有效抗病基因。迄今为止,未发现黑麦6R染色体上抗小麦秆锈病的报道,而本研究发现帝国黑麦6R染色体导入中国春之后导致该附加系对秆锈病表现为高抗,因此,6R染色体上含有未被报道的抗秆锈新基因,值得利用染色体工程的方法将其抗病基因通过小片段易位的形式转移到栽培小麦中。本研究中小麦-帝国黑麦2R和3R附加系均高感秆锈病,可能是Sr27和Sr59对中国当前流行的秆锈菌生理小种34MKGQM和21C3CTHSM失去了抗性。韩建东[27]对中国优势秆锈小种的致病性进行研究的结果也支持Sr27已经对中国当前流行的秆锈菌生理小种34MKGQM和21C3CTHSM失去了抗性的结论(文献[27]发表时,Sr59还未被报道)。3.3 卵穗山羊草染色体抗秆锈病新基因
卵穗山羊草(Ae. geniculata)染色体组为UgUgMgMg,是山羊草属中的一个异源四倍体种,高抗白粉病、叶锈病和条锈病[28,29,30,31]。来自该物种的抗白粉病基因Pm29被定位在卵穗山羊草第七同源群染色体上[32],抗叶锈基因Lr57和抗条锈病基因Yr40被定位在卵穗山羊草5Mg染色体上[31]。迄今,未见有关于卵穗山羊草基因组上存在秆锈基因的报道。本研究发现对照中国春高感秆锈病,而含卵穗山羊草7Mg#1染色体的附加系/代换系均对秆锈菌生理小种34MKGQM和21C3CTHSM均表现为高抗以上抗性,这表明卵穗山羊草7Mg#1染色体上含有抗秆锈基因。由于卵穗山羊草Mg染色体组来源于顶芒山羊草(Ae. comosa.),然而,在本研究的供试材料中中国春-顶芒山羊草7M附加系中感秆锈病,这可能是由于用于本研究的中国春-顶芒山羊草7M附加系中的顶芒山羊草不是中国春-卵穗山羊草7Mg附加系中卵穗山羊草的染色体组直接供体造成的。3.4 智利大麦6Hch染色体上的抗秆锈病新基因
智利大麦(Hordeum chilense)染色体组HchHch,是一个主要分布于智利和阿根廷的二倍体物种。具有高抗叶锈病、白粉病、全蚀病、颖枯病、腥黑穗病、麦二叉蚜、麦双尾蚜和禾谷孢囊线虫等优良性状[33]。研究发现,智利大麦4Hch染色体上含有小麦叶枯病抗性基因,7Hch上含有腥黑穗病抗病基因[34]和控制类胡萝卜素含量的基因[35],1Hch、4Hch和5Hch上含有抗盐基因,1Hch上还含有禾谷孢囊线虫抗性基因[36]。迄今,未见有关于智利大麦染色体上存在秆锈基因的报道。本研究发现中国春智利大麦6Hch附加系对秆锈菌生理小种34MKGQM和21C3CTHSM表现为免疫,说明智利大麦6Hch染色体上含有新的抗秆锈基因。3.5 尾状山羊草7C染色体上的抗秆锈病新基因
尾状山羊草(Ae. caudata L.)染色体组成为CC,是山羊草属的一个重要二倍体种,高抗白粉病[37]、叶锈病[38]、小麦瘿蚊病和蚜虫[37]。FRIEBE等[39]鉴定出了一套小麦-尾状山羊草附加系B#-F#1和5C(5A)、5C(5D)代换系,因此,可以用这套材料来定位抗性基因所在染色体和作为桥梁进一步向小麦转育其抗性。尾状山羊草抗小麦叶锈基因已成功转移到小麦中[38]。迄今,未见有关于尾状山羊草染色体上存在秆锈基因的报道。本研究发现ALCD感秆锈病,但ALCD-尾状山羊草7C#1附加系却高抗秆锈病,说明尾状山羊草7C染色体上存在有抗秆锈新基因。3.6 长穗偃麦草染色体上的抗秆锈基因
长穗偃麦草(Ag. elongatum)是偃麦草属多年生野生草本植物,具有二倍体(2n=2x=14,EE或EeEe或E1E1)、四倍体(2n=4x=28,EeEeEbEb或E1E1E2E2)和十倍体(2n=10x=70,EeEeEbEbExExStStStSt或EEE1E1E2E2E4E4E5E5)之分[40],具有抗寒、耐旱、耐盐、抗病和抗虫等优异性状[40,41]。分别位于长穗偃麦草3AeL、7AeL、6AeL和7el2染色体上的Sr24、Sr25、Sr26和Sr43已经被导入到小麦中。但在本研究中中国春-长穗偃麦草1E—7E附加系均高感或中感秆锈,这可能是由于(1)本研究中长穗偃麦草与其他研究的长穗偃麦草的来源不同;(2)Sr24和Sr43对于本研究所用到的2个秆锈生理小种34MKGQM和21C3CTHSM已经失去抗性[ 42]。3.5 抗秆锈基因所在染色体特异分子标记
高大山羊草染色体标记建立方面,刘晓明等[16]建立了高大山羊草1Sl染色体特异分子标记9个;覃碧等[43]建立2SlL染色体EST-STS标记5个;CENCI等[21]建立了3SlS染色体上与抗白粉病基因Pm13连锁的分子标记。本研究发现引物TNAC1740、TNAC1751和TNAC1756在中国春-高大山羊草6Sl#3附加系和中国春-高大山羊草6Sl#2(6B)代换系均能扩增出1 100、700和700 bp的多态性片段,而在中国春-高大山羊草的其他几个附加系和小麦对照中扩增不出相应多态性片段,说明3对引物所扩增的多态性片段是高大山羊草6Sl染色体所特有的片段。黑麦染色体分子标记方面,刘成等[10]建立了黑麦基因组SCAR标记用于小麦背景中黑麦染色质的鉴定。KOEBNER等[44]合成了黑麦1R染色体的特异引物,能够鉴定小麦背景下1RS染色质的存在。KOFLER等[45]通过构建BCA文库开发了74个黑麦1RS特异的分子标记。王春梅等[46]检测到5个EST-STS标记定位到黑麦1RS染色体上。唐宗祥等[47]筛选到黑麦的6R染色体的SSR标记。本研究建立了黑麦6R染色体PLUG标记10个,其中5个标记与前人报道相同[13,14],引物TNAC1715、TNAC1718、TNAC1737、TNAC1739和TNAC1753所扩增的多态性片段为被报道过的新标记,增加了黑麦6R染色体标记信息。
4 结论
高大山羊草6Sl#2和6Sl#3、帝国黑麦6R、智利大麦6Hch、卵穗山羊草7Mg#1、尾状山羊草7C、中间偃麦草?Ai染色体(同源群未鉴定)上含有抗秆锈新基因,值得利用染色体工程创制相应抗秆锈病小片段易位系;获得的8个新的多态性标记丰富了黑麦6R和高大山羊草6Sl染色体标记信息。The authors have declared that no competing interests exist.

{kind=link}
{kind=link}
{kind=link}
{kind=link}