 ,, 焦裕冰, 罗健达, 宋丽云, 李莹, 申莉莉,, 杨金广,, 王凤龙中国农业科学院烟草研究所,山东青岛 266101
,, 焦裕冰, 罗健达, 宋丽云, 李莹, 申莉莉,, 杨金广,, 王凤龙中国农业科学院烟草研究所,山东青岛 266101Cloning of Nicotiana benthamiana NAC062 and Its Inhibitory Effect on Potato Virus Y Infection
QU XiaoLing,, JIAO YuBing, LUO JianDa, SONG LiYun, LI Ying, SHEN LilLi,, YANG JinGuang,, WANG FengLongTobacco Research Institute, Chinese Academy of Agricultural Sciences, Qingdao 266101, Shandong通讯作者:
责任编辑: 岳梅
收稿日期:2021-02-22接受日期:2021-04-14
| 基金资助: |
Received:2021-02-22Accepted:2021-04-14
作者简介 About authors
曲潇玲,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (2050KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
曲潇玲, 焦裕冰, 罗健达, 宋丽云, 李莹, 申莉莉, 杨金广, 王凤龙. 本氏烟NbNAC062的克隆及对马铃薯Y病毒侵染的抑制作用. 中国农业科学, 2021, 54(19): 4110-4120 doi:10.3864/j.issn.0578-1752.2021.19.007
QU XiaoLing, JIAO YuBing, LUO JianDa, SONG LiYun, LI Ying, SHEN LilLi, YANG JinGuang, WANG FengLong.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】马铃薯Y病毒(potato virus Y,PVY)是一种单链正义RNA病毒[1],寄主范围广泛,能侵染34个属170余种植物[2]。烟草马铃薯Y病毒病会造成烟株花叶、脉坏死、茎坏死、点刻条斑等症状[3,4],目前已成为危害我国烟草生产最主要的病毒病之一[5,6,7]。NAC类转录因子与植物抗病、抗逆相关[8],研究其在PVY侵染中的调控作用,可为烟草抗病毒药剂的开发提供靶标,对防控马铃薯Y病毒病具有重要意义。【前人研究进展】NAC是一类植物特有的转录因子[9],在整个生命周期中起到重要作用。在植物生长发育中,NAC类转录因子参与调控种子萌发[10]、次生细胞壁生长[11]、叶片与果实衰老[12,13]等过程。在植物抗逆与激素信号传导中,NAC类转录因子参与调控植物抗病、冷胁迫、干旱胁迫以及水杨酸(salicylic acid,SA)[14]、脱落酸(abscisic acid,ABA)[15]、茉莉酸(jasmonic acid,JA)[16]等应答反应。内质网是真核生物蛋白形成加工的重要场所[17],蛋白的折叠合成过程复杂,易受外界环境影响。当植物处于逆境,细胞内未折叠蛋白与错误蛋白累积过量时,内质网会启动未折叠蛋白应答(unfolded protein response,UPR),调节一系列下游基因如分子伴侣BiP的表达,帮助蛋白正确折叠,维持内质网稳态[18]。植物膜结合转录因子在UPR信号传导过程中起到重要作用,NAC062则是一种参与UPR的NAC类膜结合转录因子。以往研究发现,NAC062主要在干旱、寒冷、高盐等逆境过程中起调控作用。拟南芥中NAC062蛋白在ABA介导的抗旱反应中起调节作用,过表达NAC062的植株表现出强的抗旱性[19]。另外NAC062还能将冷信号整合到植物防御反应中。当受到冷胁迫时,NAC062蛋白发生水解[20],直接与病程相关蛋白(pathogenesis-related protein,PR)PR1、PR2、PR5结合,激发病原体抗性反应[21],促使植株对丁香假单胞菌(Pseudomonas syringae)的侵染表现出较强的抗性。【本研究切入点】目前,关于NAC062的研究多数集中在模式植物拟南芥的冷害和干旱等非生物胁迫中,而在生物胁迫尤其是在病毒侵染中的功能研究较少。生物信息学分析表明NbNAC062启动子序列中包含抗逆、抗病等多种胁迫响应相关的顺式作用元件,但目前尚无NbNAC062基因克隆及其在PVY侵染中作用的研究,鉴于NAC062在内质网胁迫应答以及抗逆过程中的重要作用,本文探究烟草NAC062转录因子在PVY侵染中的功能及相关的调控信号。【拟解决的关键问题】以本氏烟(Nicotiana benthamiana)为材料,克隆NbNAC062,进行生物信息学分析与亚细胞定位,构建NbNAC062沉默与过表达载体,研究其在PVY侵染过程中的生物学功能。1 材料与方法
试验于2019—2020年在中国农业科学院烟草研究所完成。1.1 试验材料
植物材料:本氏烟,中国农业科学院烟草研究所保存,试验用烟均为5—6叶期温室土培烟苗,温室培养条件为温度(25±1)℃,光周期16 h光照/8 h黑暗,光照强度2 000 lx,相对湿度65%;病毒:马铃薯Y病毒,中国农业科学院烟草研究所病毒课题组保存于枯斑三生烟(Nicotiana tabacum var. Samsun NN)活体上;侵染性克隆:PVY-GFP[22]由中国农业科学院烟草研究所病毒课题组提供,于TE缓冲液中,-80℃保存。1.2 NbNAC062克隆与分析
根据National Center for Biotechnology Information(Table 1
表1
表1本试验所用引物
Table 1
| 引物Primer | 序列Sequence |
|---|---|
| NbNAC062 F | ATGATGGCAGTACTTCCTGG |
| NbNAC062 R | TACTCGCACTCTAAAGTATTCCC |
| TRV-NbNAC F | TAAGGTTACCGAATTCTTGGATGGATCACACCCTGGC |
| TRV-NbNAC R | AGACGCGTGAGCTCGGTACCTTCTGTATCATCAGCAATACAGC |
| Fu-NbNAC F | CTTTAGATCTTCTAGAATGATGGCAGTACTTCCTGG |
| Fu-NbNAC R | AGGAGGCCATGAATTCTACTCGCACTCTAAAGTATTCCC |
| PVY-CP-F | GATGAATGGGCTTATGGTTTGGTG |
| PVY-CP-R | GATTTGCCTAAGGGTTGGTTTCG |
| Actin-F | CAAGGAAATCACCGCTTTGG |
| Actin-R | AAGGGATGCGAGGATGGA |
| qPCR-NbNAC062 F | TGGACAAGAATTGGCATCGC |
| qPCR-NbNAC062 R | AACACCTCGGGCTCAAAGAAG |
| qPCR-BiP F | GCCACAGAAGAAGCTACCAAGTTG |
| qPCR-BiP R | GGTCCTCTCTGGGTTAACAGCG |
新窗口打开|下载CSV
1.3 PVY侵染对NbNAC062的影响
取PVY侵染的枯斑三生烟病叶1 g,加入PBS缓冲液(0.01 mol·L-1,pH 7.2—7.4)40 mL,于灭菌研钵中充分研磨后滤残渣,得病毒接种液。对本氏烟第3—4片真叶浸润病毒接种液,每片叶200 µL[23],以浸润PBS缓冲液为对照,接种后1、3、5、7 d取接种叶进行qRT-PCR。1.4 NbNAC062沉默载体的构建
根据NbNAC062序列设计含有EcoRⅠ、KpnⅠ酶切位点的引物TRV-NbNAC F/R(表1),以本氏烟cDNA为模板,PCR扩增TRV-NbNAC062沉默片段。利用ClonExpress II One Step Cloning Kit(Vazyme)将TRV-NbNAC062连接至pTRV载体,构建pTRV:: NbNAC062重组载体;将含有pTRV::NbNAC062载体的农杆菌(OD600=0.5)浸润本氏烟下表皮,每片叶500 µL,以浸润含有pTRV::PDS载体的农杆菌为阳性对照,浸润含有pTRV00空载的农杆菌为阴性对照,浸润15 d检测沉默效率并浸润PVY接种液。接种后1、3、5、7 d取接种叶,进行qRT-PCR与Western blot检测PVY CP积累量及UPR相关基因BiP表达量。另取部分沉默植株,浸润PVY-GFP侵染性克隆,5 d后在手持紫外灯下观察叶片荧光情况。1.5 NbNAC062瞬时过表达载体的构建
根据NbNAC062序列设计含有XbaⅠ、EcoRⅠ酶切位点的引物Fu-NbNAC F/R(表1),以本氏烟cDNA为模板,PCR扩增NbNAC062,扩增产物连接Fu46- RFP载体,构建入门载体Fu46::RFP::NbNAC062。利用LR ClonaseTM II enzyme mix(Invitrogen)将入门载体同源重组至pEarleyGate100载体,最终构建pEarleyGate100::RFP::NbNAC062表达载体[24]。将含有pEarleyGate100::RFP::NbNAC062载体的农杆菌(OD600=0.5)浸润本氏烟下表皮,每片叶500 µL,以浸润含有pEarleyGate100::RFP的农杆菌为对照。浸润农杆菌12 h后,统一浸润PVY接种液,接种后24、48、72 h取接种叶进行qRT-PCR与Western blot检测PVY CP积累量及UPR相关基因BiP表达量。1.6 NbNAC062蛋白的亚细胞定位
将含有pEarleyGate100::RFP::NbNAC062载体的农杆菌(OD600=0.5)浸润本氏烟下表皮,每片叶500 µL,12 h后接种PVY,温室培养48 h。利用DAPI细胞核蓝色染液(358 nm/461 nm,Solarbio)、BBcellProbe M01细胞膜绿色染液(488 nm/500 nm,Bestbio),进行细胞染色后,激光共聚焦观察NbNAC062蛋白亚细胞定位。1.7 qRT-PCR与Western blot
取各时间段处理组与对照组样品,利用RNA isolater Total RNA Extraction Reagent(Vazyme)与Plant Protein Extraction Kit(CWBIO)提取总RNA与总蛋白。根据不同样品的序列设计特异性荧光定量检测引物(表1),总RNA反转录成cDNA(Vazyme),利用ChamQTM Universal SYBR® qPCR Master Mix(Vazyme)试剂盒进行qRT-PCR。以Actin为内参基因,2-ΔΔCt法计算相对表达量,反应程序:95℃ 30 s;95℃ 10 s,60℃ 30 s,40个循环;95℃ 15 s,60℃ 60 s,95℃ 15 s。总蛋白经SDS-PAGE电泳(180 V、32 min),湿法转膜(100 V、90 min)后,分别4℃过夜孵育PVY抗体(Agdia,兔源),Actin抗体(CWBIO,鼠源),对应二抗Goat Anti-Rabbit IgG(CWBIO),Goat Anti-Mouse IgG(CWBIO)25℃孵育2 h后观察Western blot结果。2 结果
2.1 NbNAC062克隆与分析
以本氏烟cDNA为模板,NbNAC062 F/R为引物(表1),PCR扩增出1 944 bp CDS序列,编码646个氨基酸。CDS序列于NCBI全网比对,发现与渐狭叶烟草(Nicotiana attenuata)、栽培烟草(Nicotiana tabacum)、林烟草(Nicotiana sylvestris)、绒毛状烟草(Nicotiana tomentosiformis)的NAC062序列最为相似,相似度分别为96.04%、95.88%、95.88%、94.91%,对应蛋白序列相似度为95.83%、94.28%、94.28%、93.04%。其他物种核苷酸序列相似度为83.71%—85.27%,对应蛋白序列相似度为74.73%—79.35%。利用MEGA7,邻接法、自举抽样各分支置信度为1 000构建系统发育树(图1)[25]。利用DNAMAN进行氨基酸序列比对(图2-A),结果均显示,扩增序列与渐狭叶烟草NAC062序列相似度最高,表明扩增序列为本氏烟NAC062的同源序列,命名为NbNAC062。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1NbNAC062系统发育分析
Fig. 1Phylogenetic analyses of NbNAC062
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2NbNAC062蛋白序列及结构域分析
A:NbNAC062蛋白氨基酸序列对比分析。深蓝色为氨基酸完全相同,粉色为有一个氨基酸不同,浅蓝色为两个氨基酸不同。红、黄、绿色箭头线分别代表NAC结构域、DNA结合区域、TMD跨膜结构域;B:NbNAC062蛋白结构域分析
Fig. 2NbNAC062 protein sequence and domain analysis
Comparative analysis of NbNAC062 protein amino acid sequence. Dark blue indicates that the amino acids are identical, pink indicates that there is only one amino acid difference, light blue indicates that there are two amino acids differences. The red, yellow, and green arrow lines represent the NAC domain, the DNA binding region, and the TMD transmembrane domain in turn; Protein domain analysis of NbNAC062
利用UniProt[26]、SMART[27,28,29]、TMHMM Server 2.0[30]对NbNAC062蛋白进行序列分析,发现第28—179 aa为NAC结构域,第129—185 aa为DNA结合区域,第621—643 aa为C端疏水跨膜域(图2-B)。
NbNAC062序列于Sol Genomics Network网站比对获取其启动子。PlantCARE分析发现NbNAC062启动子序列中包含多种胁迫反应顺式作用元件,包括脱落酸反应顺式作用元件ABRE、茉莉酸甲酯反应顺式作用元件TGACG-motif和CGTCA-motif、参与PR转录表达的水杨酸响应元件as-1,以及参与干旱、高盐、低温等胁迫应答相关的顺式作用元件DRE、MYB、MYC。
2.2 PVY侵染对NbNAC062的影响
本氏烟接种PVY后1、3、5、7 d取接种叶进行qRT-PCR与统计分析。结果表明PVY侵染导致NbNAC062表达量上调,其中接种PVY 5 d,NbNAC062 mRNA水平显著高于接种PBS对照组,为对照组的2.52倍;接种PVY 7 d,NbNAC062 mRNA水平极显著高于对照组,为对照组的1.95倍(图3-A)。qRT-PCR检测UPR相关基因BiP发现,接种PVY后1、3、5 d,BiP mRNA表达量逐渐上调,其中接种PVY 3 d,BiP mRNA水平极显著高于对照组,为对照组的2.39倍,接种后7 d,BiP表达量极显著低于对照组,下调表达56.77%(图3-B),说明NbNAC062及BiP可能在PVY侵染中发挥作用。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3PVY侵染本氏烟后NbNAC062与BiP表达量变化
*表示差异显著P<0.05,**表示差异极显著P<0.01。下同
Fig. 3Changes of NbNAC062 and BiP expression after PVY infecting N. benthamiana
* indicates that difference is significant at the 0.05 level, P<0.05. ** indicates that difference is significant at the 0.01 level, P<0.01. The same as below
含pEarleyGate100::RFP::NbNAC062载体的农杆菌介导浸润本氏烟下表皮,12 h后接种PVY,温室培养48 h进行激光共聚焦观察。结果表明,PVY侵染前,NbNAC062蛋白定位于细胞膜,PVY侵染后部分NbNAC062蛋白转移至细胞核(图4)。
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4PVY侵染前后NbNAC062蛋白亚细胞定位
A:NbNAC062蛋白亚细胞定位,BBcellProbe M01为细胞膜绿色染料,激发波长/发射波长为488 nm/500 nm Subcellular localization of NbNAC062 protein, BBcellProbe M01 is cell membrane green dye, excitation wavelength/emission wavelength is 488 nm/500 nm;B:PVY侵染后NbNAC062蛋白亚细胞定位,DAPI为细胞核蓝色染料,激发波长/发射波长为358 nm/461 nm Subcellular localization of NbNAC062 protein after PVY infection, DAPI is nuclear blue dye, excitation wavelength/emission wavelength is 358 nm/461 nm
Fig. 4Subcellular localization of NbNAC062 protein before and after PVY infection
2.3 沉默NbNAC062对PVY侵染的影响
含pTRV::NbNAC062沉默载体的农杆菌介导浸润本氏烟第7天,沉默组与阴性对照组pTRV00表型无明显变化,阳性对照组pTRV::PDS叶片开始出现白化现象。第15天,qRT-PCR检测沉默效率为84.25%(图5)。接种PVY后,1、3、5、7 d取接种叶进行qRT-PCR与Western blot检测PVY CP积累量。qRT-PCR结果显示,3、5、7 d沉默组本氏烟PVY CP mRNA显著高于对照组,分别为对照组的2.12、2.41、1.38倍。Western blot显示5、7 d沉默组PVY CP蛋白含量显著高于对照组。通过PVY CP mRNA和蛋白在沉默植株中积累的分析表明,NbNAC062表达的下调显著促进了PVY的积累。qRT-PCR检测BiP表达,3、5、7 d沉默组BiP mRNA表达量逐渐下调,分别下调表达28.19%、58.11%、10.77%,说明沉默NAC062抑制了BiP的表达(图6)。手持紫外灯下观察,沉默组PVY-GFP荧光强度高于对照组(图5),也说明本氏烟沉默NbNAC062促进了PVY的侵染。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5沉默NbNAC062表型分析与沉默效率
A:沉默第7天处理组pTRV::NbNAC062、阴性对照组pTRV00、阳性对照组pTRV::PDS表型 On the 7th day of silence, the phenotype of pTRV::NbNAC062 treatment group, pTRV00 negative control group and pTRV::PDS positive control group;B:沉默NbNAC062第15天沉默效率 Silencing efficiency of NbNAC062 on the 15th day;C:NbNAC062沉默植株接种PVY-GFP第5天叶片荧光情况 Fluorescence of leaves on the 5th day after silencing NbNAC062 inoculated with PVY-GFP
Fig. 5Phenotype analysis and silencing efficiency of NbNAC062 silencing
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6沉默NbNAC062后PVY与BiP积累量变化
A:沉默NbNAC062后PVY侵染1、3、5、7 d,qRT-PCR检测PVY CP mRNA变化Changes of PVY CP mRNA are detected by qRT-PCR when PVY infection was 1, 3, 5, 7 days after silencing NbNAC062;B:沉默NbNAC062后PVY侵染1、3、5、7 d,qRT-PCR检测BiP变化Changes of BiP are detected by qRT-PCR when PVY infection was 1, 3, 5, 7 days after silencing NbNAC062;C:沉默NbNAC062后PVY侵染1、3、5、7 d,Western blot检测PVY蛋白量变化。每天取样组中左侧为对照右侧为处理Changes of PVY protein are detected by Western blot when PVY infection was 1, 3, 5, 7 days after silencing NbNAC062. In the daily sampling group, the left side is the control and the right side is the treatment;D:C对应的Actin Western blot蛋白杂交图The Western blot protein hybridization map of Actin corresponding to C
Fig. 6Changes in PVY and BiP accumulation after silence of NbNAC062
2.4 过表达NbNAC062对PVY侵染的影响
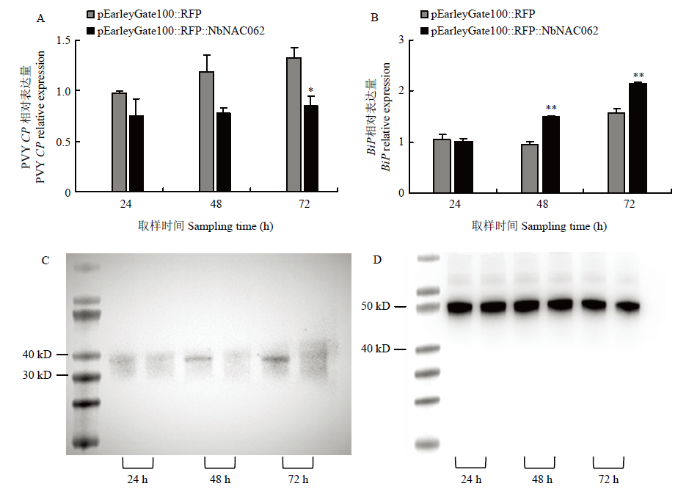
含过表达载体的农杆菌介导浸润本氏烟12 h后,浸润PVY接种液,接种后24、48、72 h取接种叶进行qRT-PCR与Western blot检测PVY CP积累量,结果表明,过表达组PVY CP mRNA含量基本持平,对照组呈上升趋势,接种72 h,过表达组PVY CP mRNA含量显著低于对照组,下调表达36.21%。Western blot结果显示,接种48、72 h,过表达组PVY蛋白含量低于对照组,说明过表达NbNAC062抑制PVY的侵染。qRT-PCR检测BiP表达,接种48、72 h,过表达组BiP mRNA表达量极显著高于对照组,分别为对照组的1.56、1.35倍,说明过表达NAC062促进了BiP的表达(图7)。图7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7过表达NbNAC062后PVY与BiP积累量变化
A:过表达NbNAC062后PVY侵染1、3、5、7 d,qRT-PCR检测PVY CP mRNA变化Changes of PVY CP mRNA are detected by qRT-PCR when PVY infection was 1, 3, 5, 7 days after overexpression NbNAC062;B:过表达NbNAC062后PVY侵染1、3、5、7 d,qRT-PCR检测BiP变化Changes of BiP are detected by qRT-PCR when PVY infection was 1, 3, 5, 7 days after overexpression NbNAC062;C:过表达NbNAC062后PVY侵染1、3、5、7 d,Western blot检测PVY蛋白量变化。每天取样组中左侧为对照右侧为处理Changes of PVY protein are detected by Western blot when PVY infection was 1, 3, 5, 7 days after overexpression NbNAC062. In the daily sampling group, the left side is the control and the right side is the treatment;D:C对应的Actin Western blot蛋白杂交图The Western blot protein hybridization map of Actin corresponding to C
Fig. 7Changes in PVY and BiP accumulation after overexpression of NbNAC062
3 讨论
NAC062是一种膜结合转录因子,属于NAC类转录因子,N端有NAC保守结构域,C末端有疏水跨膜结构[31],主要参与植物抗逆反应、激素信号传导以及生长发育等过程。拟南芥中,AtNAC062被发现可被高盐等非生物胁迫以及ABA等植物激素触发,参与植物抗旱反应;冷胁迫可使AtNAC062蛋白从细胞膜释放并进入细胞核,调节PR的表达,增强植株对病原菌的抗性反应。本研究以本氏烟cDNA为模板,扩增得到NbNAC062序列,编码646个氨基酸,N端28—179 aa为NAC结构域,129—185 aa为DNA结合区域,C末端621—643 aa为疏水跨膜结构,与AtNAC062结构相同。NbNAC062启动子序列中包含多种胁迫反应顺式作用元件,例如PR转录表达的水杨酸响应元件as-1,脱落酸反应顺式作用元件ABRE以及DRE、MYB、MYC等参与干旱、高盐、低温等胁迫诱导相关的顺式作用元件,说明NbNAC062同AtNAC062类似,可参与抗逆、抗病反应。除冷胁迫外,内质网胁迫(endoplasmic reticulum stress,ER stress)也可激活NAC062发挥作用。许多RNA病毒可以利用内质网的蛋白合成功能附着在内质网上,完成自身的复制组装[32,33,34]。作为RNA病毒的PVY,其进入宿主细胞后,诱导内质网形成囊泡状结构,以构建病毒复制复合体(viral replication complex,VRC),完成自身增殖,因而PVY的复制增殖与内质网密切相关[35]。与此同时,大量病毒蛋白的聚集会造成内质网胁迫[36],细胞启动UPR,以维持内质网稳态。NAC类转录因子在UPR信号传导中起到重要作用。拟南芥中,AtNAC062通常以休眠形式定位于细胞膜,内质网胁迫诱导剂衣霉素(tunicamycin,TM)处理后,引发ER stress,AtNAC062被激活从细胞膜转移至细胞核,调节下游UPR相关基因BiP等的表达[37]。分子伴侣是蛋白质折叠过程中十分重要的辅助因子,能与非自然状态的蛋白短暂互作,帮助它们稳定原生状态,防止错误折叠。BiP(binding protein)蛋白是一种常见的分子伴侣,在内质网腔中含量较高,是UPR途径中维持内质网稳态的重要基因。本研究发现PVY侵染本氏烟前后,激光共聚焦观察NbNAC062蛋白从细胞膜转移至细胞核,PVY侵染前期,UPR相关基因BiP表达量逐渐上调,接种PVY 3 d,BiP表达量为对照组的2.39倍,接种7 d后,BiP表达量开始下调,下调表达56.77%。说明PVY侵染会激活NbNAC062蛋白至细胞核发挥转录作用,NbNAC062及UPR相关基因BiP在PVY侵染中发挥作用。伴随NbNAC062的沉默与过量表达,BiP的表达量也相应发生下调和上调,BiP分别下调10.77%—58.11%,上调1.35—1.56倍,说明NbNAC062转录因子可能调控BiP的表达,内质网胁迫时,NbNAC062通过加强转录BIP达到维持内质网稳态的作用,本研究还发现PVY侵染引起NbNAC062表达上调,综上推测,PVY利用内质网完成自身复制增殖时造成ER stress,寄主通过NbNAC062转录因子,上调表达BiP维持内质网稳态,调节细胞稳定性来增强寄主基础抗性。
目前,关于NAC062的研究多数集中在非生物胁迫中,关于其增强病原菌抗性的研究也是基于冷胁迫诱导,其在生物胁迫中的功能研究较少,尤其是在病毒侵染方面。本研究利用病毒介导的基因沉默(virus- induced gene silencing,VIGS)与瞬时过表达技术,研究NbNAC062在PVY侵染过程中的作用,发现NbNAC062的表达量与PVY的积累量呈负相关,证明NbNAC062对PVY侵染具有抑制作用,并调节下游相关基因的表达,后续研究会通过敲除和过表达突变体,进一步确认这一结果。本试验为研究PVY侵染过程中寄主自我防卫机制提供了新思路。目前,我国传统的农业、物理和化学防治方法对于病毒病的防治效果并不理想[38],鉴于NbNAC062对PVY的抑制作用,未来可考虑利用纳米材料包裹,开发抗病毒纳米药剂。
4 结论
NbNAC062属于NAC类膜结合转录因子,PVY侵染本氏烟导致NbNAC062表达上调,激活其从细胞膜转移至细胞核,NbNAC062调控UPR相关基因BiP表达,维持细胞稳态。本氏烟沉默NbNAC062导致PVY积累量上升;过表达NbNAC062则使PVY积累量下降,说明NbNAC062对PVY侵染有一定抑制作用,未来或可作为抗病毒药剂开发的靶标。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1099/vir.0.014142-0URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s00299-016-2080-1URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
DOI:10.1016/j.jplph.2009.05.013URL [本文引用: 1]
DOI:10.1093/mp/ssq062URL [本文引用: 1]
DOI:10.1038/srep23609URL [本文引用: 1]
DOI:10.1016/j.jplph.2016.01.014URL [本文引用: 1]
DOI:10.1111/tpj.2005.43.issue-5URL [本文引用: 1]
DOI:10.1016/j.fob.2013.07.006URL [本文引用: 1]
DOI:10.1111/tpj.2010.61.issue-5URL [本文引用: 1]
DOI:10.1152/physrev.00015.2006URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1042/BJ20120244URL [本文引用: 1]
DOI:10.1042/BJ20091762URL [本文引用: 1]
DOI:10.1111/tpj.2010.61.issue-4URL [本文引用: 1]
DOI:10.1016/j.virusres.2005.03.001URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/nph.15310URL [本文引用: 1]
DOI:10.1093/molbev/msw054URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1093/nar/gkaa937URL [本文引用: 1]
DOI:10.1093/nar/gkx922URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.coviro.2011.09.009URL [本文引用: 1]
DOI:10.1146/annurev-phyto-073009-114239URL [本文引用: 1]
DOI:10.1016/j.virol.2013.09.001URL [本文引用: 1]
DOI:10.1128/JVI.01824-09URL [本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
DOI:10.1111/tpj.12604URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}