Cloning and Analysis of P-glycoprotein Gene and Its Transcriptional Response to Insecticide in Chilo suppressalis
MENG XiangKun,, WU ZhaoLu, YANG XueMei, GUAN DaoJie, WANG JianJun,College of Horticulture and Plant Protection, Yangzhou University, Yangzhou 225009, Jiangsu
Abstract 【Objective】The CsPgp was cloned from Chilo suppressalis, and the molecular characteristics and expression profiles of CsPgp were analyzed. Transcriptional responses as well as the potential transcriptional regulation mechanism of CsPgp to two common used insecticides (chlorantraniliprole and abamectin) were also studied.【Method】The full length of CsPgp was cloned from C. suppressalis using the gene cloning technology. The molecular characteristics and the transcription factor binding sites in 5′ transcriptional regulatory region of CsPgp were analyzed employing the bioinformatics technologies. Expression profiles of CsPgp in different stages and tissues of C. suppressalis, and the transcriptional responses of CsPgp to different doses of chlorantraniliprole and abamectin treatment were determined using the real-time quantitative PCR.【Result】The full length of CsPgp cDNA is 4 584 bp and consists of 23 exons. The encoding protein has 1 259 amino acids containing two transmembrane regions and two nucleotide binding domains and the typical structural features of ABC transporter family such as the Walker A, Walker B and D, H, P, Q-Loop which have important function in substrate transfer. CsPgp was mainly expressed in larval stage of C. suppressalis, especially in the 3rd and 4th instar larvae, while CsPgp showed low expression levels in the pupal and adult stages. Analysis of the tissue expressions showed that CsPgp was predominately expressed in the foregut and midgut, and had very low expression levels in other tissues including hindgut, fat body and Malpighian tubule. No significant change of CsPgp expression was found in the 3rd instar larvae of C. suppressalis after treated with LC30 and LC70 of chlorantraniliprole for 12 and 24 h, respectively, when compared with the control groups. However, the expressions of CsPgp were significantly up-regulated in larvae after treated with LC30 of chlorantraniliprole for 36 h, while the expressions of CsPgp were significantly down-regulated in larvae after treated with LC70 of chlorantraniliprole for 36 h. In the 0.05 mg·L-1 of abamectin treatment, CsPgp was remarkably down-regulated at 12 h post-treatment, while the expressions of CsPgp were not significantly changed at 24 and 36 h post-treatment, respectively. However, CsPgp was significantly induced in larvae after treated with 0.15 mg·L-1 of abamectin for 24 and 36 h, respectively. Sequence analysis of the 5′ transcriptional regulatory region of CsPgp showed that multiple transcription factor binding sites were predicted in the 5′ transcriptional regulatory region of CsPgp, including five potential CncC binding sites.【Conclusion】CsPgp was highly expressed in the midgut of C. suppressalis and could be induced by chlorantraniliprole and abamectin, which indicated that CsPgp might involve in the detoxification metabolism of chlorantraniliprole and abamectin in C. suppressalis. Multiple CncC binding sites were found in the 5′ transcriptional regulatory region of CsPgp which might have important regulatory effects on the expression of CsPgp. It was speculated that CsPgp might be regulated by transcription factor CncC and participated in the detoxification metabolism of chlorantraniliprole or abamectin when C. suppressalis was exposed to chlorantraniliprole or abamectin. Keywords:Chilo suppressalis;P-glycoprotein;molecular characteristic;insecticide induction;transcriptional regulation
PDF (4406KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 孟祥坤, 吴赵露, 杨雪梅, 官道杰, 王建军. 二化螟P糖蛋白基因的克隆分析及对杀虫剂的诱导响应. 中国农业科学, 2021, 54(19): 4121-4131 doi:10.3864/j.issn.0578-1752.2021.19.008 MENG XiangKun, WU ZhaoLu, YANG XueMei, GUAN DaoJie, WANG JianJun. Cloning and Analysis of P-glycoprotein Gene and Its Transcriptional Response to Insecticide in Chilo suppressalis. Scientia Acricultura Sinica, 2021, 54(19): 4121-4131 doi:10.3864/j.issn.0578-1752.2021.19.008
A:CsPgp核酸序列及其编码的氨基酸序列,深灰色背景标注的为CsPgp跨膜区序列;浅灰色背景标注的为CsPgp核苷酸结合区,其中重要的结构特征使用单下划线标出,斜体氨基酸为ATP结合位点Nucleotide and deduced amino acid sequences of CsPgp. The transmembrane domains are marked by dark grey background, and the nucleotide-binding domains are marked by light grey background. The important structural characteristics in nucleotide-binding domains are underlined, and the italic amino acids indicated as the binding sites of ATP。B:CsPgp基因组结构Genomics structure of CsPgp。Intron:内含子;Exon外显子;UTR:非翻译区Untranslated region Fig. 1Sequence characteristics analysis of CsPgp
柱上标有不同字母表示CsPgp在不同时期表达量差异显著(P<0.05) Histograms with different letters indicate significant difference of CsPgp expression in different stages (P<0.05)。1—6龄幼虫:1st-6th instar larva;预蛹:Prepupa;1、3、5日蛹:1st, 3rd and 5th day-old pupa;0、12、24、36、48、72小时成虫:0, 12, 24, 36, 48 and 72 hour-old adult Fig. 3Relative expression levels of CsPgp in different developmental stages of C. suppressalis
柱上标有不同字母表示CsPgp在不同组织表达量差异显著(P<0.05) Histograms with different letters indicate significant difference of CsPgp expression in different tissues (P<0.05)。脑:Brain;神经索:Nerve cord;前肠:Foregut;中肠:Midgut;后肠:Hindgut;卵巢:Ovary;马氏管:Malpighian tubule;表皮:Cuticula;血淋巴:Hemolymph;脂肪体:Fat body Fig. 4Relative expression levels of CsPgp in different tissues of C. suppressalis
基于二化螟基因组序列信息,克隆了CsPgp上游5′侧翼区约2 000 bp DNA序列,并对序列中的转录因子结合位点、启动子序列及转录起始位点等转录调控元件进行预测分析。分析结果显示,在CsPgp 5′上游调控区中预测到一个潜在的启动子序列(Score=0.91)及多个转录因子结合位点(Score>0.8)(图6)。预测的转录因子结合位点中包括5个Cnc::maf-S,3个EcR::usp及br、Deaf1、dl、tin、Dfd等。
单下划线标注的为预测的转录因子结合位点(Score>0.8),双下划线标注的为预测的启动子序列(Score=0.91),其中粗体字母为预测的转录起始位点。以CsPgp编码区起始密码子的第一个碱基位置标为“+1”,其上游碱基标为“-” Fig. 6Sequence analysis of 5′ DNA fragment of CsPgp
The predicted transcription factors are marked by single underline, and the predicted promoter sequence is marked by double underline. The transcription start site is indicated by bold letter. The first base of initiation code of CsPgp is indicated as “+1”, and its upstream sequences are indicated as “-”
LIU WC, LIU ZD, HUANGC, LU MH, LIUJ, YANG QP. Statistics and analysis of crop yield losses caused by main diseases and insect pests in recent 10 years Plant Protection, 2016, 42(5):1-9. (in Chinese) [本文引用: 1]
National Agricultural Technology Extension Service Center. Monitoring results of pesticide resistance of agricultural pests and suggestions for scientific pesticide use in China in 2017 China Plant Protection, 2018, 38(4):52-56. (in Chinese) [本文引用: 2]
National Agricultural Technology Extension Service Center. Monitoring results of pesticide resistance of agricultural pests and suggestions for scientific pesticide use in China in 2018 China Plant Protection, 2019, 39(3):63-67, 72. (in Chinese) [本文引用: 2]
National Agricultural Technology Extension Service Center. Monitoring results of pesticide resistance of agricultural pests and suggestions for scientific pesticide use in China in 2019 China Plant Protection, 2020, 40(3):64-69. (in Chinese) [本文引用: 2]
WEI YB, YANR, ZHOU QL, QIAO LY, ZHU GN, CHEN ML. Monitoring and mechanisms of chlorantraniliprole resistance in Chilo suppressalis (Lepidoptera: Crambidae) in China Journal of Economic Entomology, 2019, 112(3):1348-1353. DOI:10.1093/jee/toz001URL [本文引用: 2]
LU YH, WANG GR, ZHONG LQ, ZHANG FC, BAIQ, ZHENG XS, LU ZX. Resistance monitoring of Chilo suppressalis (Walker) (Lepidoptera: Crambidae) to chlorantraniliprole in eight field populations from east and central China Crop Protection, 2017, 100:196-202. DOI:10.1016/j.cropro.2017.07.006URL [本文引用: 4]
MAO KK, LI WH, LIAOX, LIU CY, QINY, REN ZJ, QIN XY, WANH, SHENGF, LI JH. Dynamics of insecticide resistance in different geographical populations of Chilo suppressalis (Lepidoptera: Crambidae) in China 2016-2018 Journal of Economic Entomology, 2019, 112(4):1866-1874. DOI:10.1093/jee/toz109URL [本文引用: 2]
KALSIM, PALLI SR. Cap n collar transcription factor regulates multiple genes coding for proteins involved in insecticide detoxification in the red flour beetle, Tribolium castaneum Insect Biochemistry and Molecular Biology, 2017, 90:43-52. DOI:10.1016/j.ibmb.2017.09.009URL [本文引用: 2]
MENG XK, YANG XM, WU ZL, SHEN QW, MIAO LJ, ZHENGY, QIANK, WANG JJ. Identification and transcriptional response of ATP-binding cassette transporters to chlorantraniliprole in the rice striped stem borer, Chilo suppressalis Pest Management Science, 2020, 76(11):3626-3635. DOI:10.1002/ps.v76.11URL [本文引用: 3]
DERMAUWW, VAN LEEUWENT. The ABC gene family in arthropods: Comparative genomics and role in insecticide transport and resistance Insect Biochemistry and Molecular Biology, 2014, 45:89-110. DOI:10.1016/j.ibmb.2013.11.001URL [本文引用: 5]
SUNY, XUL, CHENQ, QIN WJ, HUANG SJ, JIANGY, QIN HG. Chlorantraniliprole resistance and its biochemical and new molecular target mechanisms in laboratory and field strains of Chilo suppressalis (Walker) Pest Management Science, 2018, 74(6):1416-1423. DOI:10.1002/ps.2018.74.issue-6URL [本文引用: 2]
XUL, ZHAOJ, SUNY, XU DJ, XU GC, XU XL, ZHANG YL, HUANG SJ, HAN ZJ, GU ZY. Constitutive overexpression of cytochrome P450 monooxygenase genes contributes to chlorantraniliprole resistance in Chilo suppressalis (Walker) Pest Management Science, 2019, 75(3):718-725. DOI:10.1002/ps.2019.75.issue-3URL [本文引用: 1]
ZHAOJ, XUL, SUNY, SONG PP, HAN ZJ. UDP- glycosyltransferase genes in the striped rice stem borer, Chilo suppressalis (Walker), and their contribution to chlorantraniliprole resistance International Journal of Molecular Sciences, 2019, 20(5):1064. DOI:10.3390/ijms20051064URL [本文引用: 3]
LIB, HAN LZ, PENG YF. Development of a standardized artificial diet and rearing technique for the striped stem borer, Chilo suppressalis Walker (Lepidoptera: Crambidae) Chinese Journal of Applied Entomology, 2015, 52(2):498-503. (in Chinese) [本文引用: 1]
MENG XK, DONGF, QIANK, MIAO LJ, YANG XM, GE HC, WU ZL, WANG JJ. Transcriptome analysis reveals global gene expression changes of Chilo suppressalis in response to sublethal dose of chlorantraniliprole Chemosphere, 2019, 234:648-657. DOI:10.1016/j.chemosphere.2019.06.129URL [本文引用: 3]
XU HX, WANG GR, LU YH, YANG YJ, ZHENG XS, TIAN JC, LÜ ZX. Screening reference genes and evaluating of their expression stability for qRT-PCR normalization in Chilo suppressalis (Lepidoptera: Pyralididae) Chinese Journal of Rice Science, 2019, 33(1):75-84. (in Chinese) [本文引用: 1]
XUJ, LU MX, CUI YD, DU YZ. Selection and evaluation of reference genes for expression analysis using qRT-PCR in Chilo suppressalis (Lepidoptera: Pyralidae) Journal of Economic Entomology, 2017, 110(2):683-691. [本文引用: 1]
DERMAUWW, ILIASA, RIGAM, TSAGKARAKOUA, GRBICM, TIRRYL, VAN LEEUWENT, VONTASJ. The cys-loop ligand-gated ion channel gene family of Tetranychus urticae: Implications for acaricide toxicology and a novel mutation associated with abamectin resistance Insect Biochemistry and Molecular Biology, 2012, 42(7):455-465. DOI:10.1016/j.ibmb.2012.03.002URL [本文引用: 1]
LIAO CY, XIA WK, FENG YC, LIG, LIUH, DOUW, WANG JJ. Characterization and functional analysis of a novel glutathione S-transferase gene potentially associated with the abamectin resistance in Panonychus citri (McGregor) Pesticide Biochemistry and Physiology, 2016, 132:72-80. DOI:10.1016/j.pestbp.2015.11.002URL [本文引用: 1]
RIGAM, TSAKIRELID, ILIASA, MOROUE, MYRIDAKISA, STEPHANOU EG, NAUENR, DERMAUWW, VAN LEEUWENT, PAINEM, VONTASJ. Abamectin is metabolized by CYP392A16, a cytochrome P450 associated with high levels of acaricide resistance in Tetranychus urticae Insect Biochemistry and Molecular Biology, 2014, 46:43-53. DOI:10.1016/j.ibmb.2014.01.006URL [本文引用: 1]
WANG XL, PUINEAN AM, O’REILLYA O, WILLIAMSONM S, SMELTC L C, MILLARN S, WUY D. Mutations on M3 helix of Plutella xylostella glutamate-gated chloride channel confer unequal resistance to abamectin by two different mechanisms Insect Biochemistry and Molecular Biology, 2017, 86:50-57. DOI:10.1016/j.ibmb.2017.05.006URL [本文引用: 1]
YINQ, QIANL, SONG PP, JIAN TY, HAN ZJ. Molecular mechanisms conferring asymmetrical cross-resistance between tebufenozide and abamectin in Plutella xylostella. Journal of Asia-Pacific Entomology, 2019, 22(1):189-193. [本文引用: 2]
LUOL, SUN YJ, WU YJ. Abamectin resistance in Drosophila is related to increased expression of P-glycoprotein via the dEGFR and dAkt pathways Insect Biochemistry and Molecular Biology, 2013, 43(8):627-634. DOI:10.1016/j.ibmb.2013.04.006URL [本文引用: 1]
XIANGM, ZHANGL, LUY, TANG QL, LIANGP, SHI XY, SONG DL, GAO XW. A P-glycoprotein gene serves as a component of the protective mechanisms against 2-tridecanone and abamectin in Helicoverpa armigera Gene, 2017, 627:63-71. DOI:10.1016/j.gene.2017.06.010URL [本文引用: 5]
TIAN LX, YANG JQ, HOU WJ, XU BY, XIEW, WANG SL, ZHANG YJ, ZHOU XG, WU QJ. Molecular cloning and characterization of a P-glycoprotein from the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae) International Journal of Molecular Sciences, 2013, 14(11):22891-22905. DOI:10.3390/ijms141122891URL [本文引用: 2]
ENDERS LS, RAULT LC, HENG-MOSST M, SIEGFRIEDB D, MILLER NJ. Transcriptional responses of soybean aphids to sublethal insecticide exposure Insect Biochemistry and Molecular Biology, 2020, 118:103285. DOI:10.1016/j.ibmb.2019.103285URL [本文引用: 1]
HEC, LIANG JJ, LIU SN, WANG SL, WU QJ, XIEW, ZHANG YJ. Changes in the expression of four ABC transporter genes in response to imidacloprid in Bemisia tabaci Q (Hemiptera: Aleyrodidae) Pesticide Biochemistry and Physiology, 2019, 153:136-143. DOI:10.1016/j.pestbp.2018.11.014URL [本文引用: 1]
JIN MH, LIAO CY, CHAKRABARTYS, ZHENG WG, WU KM, XIAO YT. Transcriptional response of ATP-binding cassette (ABC) transporters to insecticides in the cotton bollworm, Helicoverpa armigera Pesticide Biochemistry and Physiology, 2019, 154:46-59. DOI:10.1016/j.pestbp.2018.12.007URL
MERZENDORFERH. ABC transporters and their role in protecting insects from pesticides and their metabolites//COHEN E. Target Receptors in the Control of Insect Pests: Part II. 2014, 46:1-72.
SUNH, PUJ, CHENF, WANG JD, HAN ZJ. Multiple ATP-binding cassette transporters are involved in insecticide resistance in the small brown planthopper, Laodelphax striatellus Insect Molecular Biology, 2017, 26(3):343-355. DOI:10.1111/imb.2017.26.issue-3URL [本文引用: 1]
XU ZF, SHIL, PENG JF, SHEN GM, WEIP, WUQ, HEL. Analysis of the relationship between P-glycoprotein and abamectin resistance in Tetranychus cinnabarinus (Boisduval) Pesticide Biochemistry and Physiology, 2016, 129:75-82. DOI:10.1016/j.pestbp.2015.10.021URL [本文引用: 1]
BAKSHIM, OELMÜLLERR. WRKY transcription factors: Jack of many trades in plants Plant Signaling and Behavior, 2014, 9(2):e27700. DOI:10.4161/psb.27700URL [本文引用: 1]
HUB, HU SZ, HUANGH, WEIQ, REN MM, HUANG SF, TIAN XR, SU JY. Insecticides induce the co-expression of glutathione S-transferases through ROS/CncC pathway in Spodoptera exigua Pesticide Biochemistry and Physiology, 2019, 155:58-71. DOI:10.1016/j.pestbp.2019.01.008URL [本文引用: 1]
WILDING CS. Regulating resistance: CncC:Maf, antioxidant response elements and the overexpression of detoxification genes in insecticide resistance Current Opinion in Insect Science, 2018, 27:89-96. DOI:10.1016/j.cois.2018.04.006URL [本文引用: 1]
CHENL, ZHANG TT, GE MY, LIU YH, XING YP, LIUL, LI FL, CHENG LG. The Nrf2-Keap1 pathway: A secret weapon against pesticide persecution in Drosophila Kc cells Pesticide Biochemistry and Physiology, 2020, 164:47-57. DOI:10.1016/j.pestbp.2019.12.008URL [本文引用: 1]
CHENG XY, HU JH, LI JX, CHENJ, WANGH, MAO TT, XUEB, LIB. The silk gland damage and the transcriptional response to detoxifying enzymes-related genes of Bombyx mori under phoxim exposure Chemosphere, 2018, 209:964-971. DOI:10.1016/j.chemosphere.2018.06.167URL [本文引用: 1]
MAO TT, LI FC, FANG YL, WANGH, CHENJ, LI MX, LU ZT, QU JW, LI JX, HU JH, CHENG XY, NIM, LIB. Effects of chlorantraniliprole exposure on detoxification enzyme activities and detoxification-related gene expression in the fat body of the silkworm, Bombyx mori Ecotoxicology and Environmental Safety, 2019, 176:58-63. DOI:10.1016/j.ecoenv.2019.03.074URL [本文引用: 1]
SHIL, SHIY, LIU MF, ZHANGY, LIAOX L. Transcription factor CncC potentially regulates the expression of multiple detoxification genes that mediate indoxacarb resistance in Spodoptera litura Insect Science, 2021, https://doi.org/10.1111/1744-7917.12860. URL [本文引用: 1]
 ,, 吴赵露, 杨雪梅, 官道杰, 王建军
,, 吴赵露, 杨雪梅, 官道杰, 王建军

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT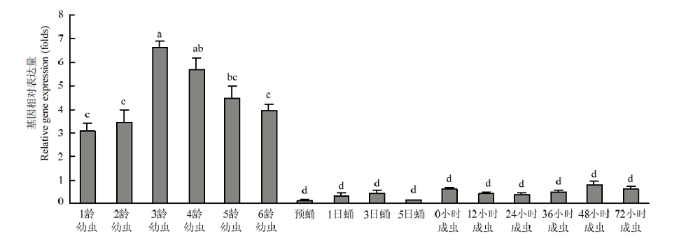 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT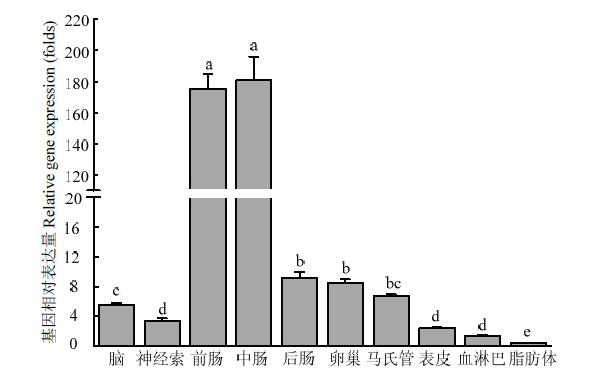 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}