Generation of ospin9 Mutants in Rice by CRISPR/Cas9 Genome Editing Technology
WU ShiYang,, YANG XiaoYi, ZHANG YanWen, HOU DianYun, XU HuaWei,College of Agriculture, Henan University of Science and Technology, Luoyang 471000, Henan
Abstract 【Objective】Auxin efflux protein family PIN-FORMED (PIN) is a key protein family in controlling polar auxin transport (PAT). OsPIN9 is one of the monocot-specific PIN genes in rice, while its biological function still needs to be further elucidated. In this study, OsPIN9 was edited and ospin9 homozygous mutants were obtained using CRISPR/Cas9 genome editing technology. The resultant ospin9 mutant lines could provide a basis for further research on the function of OsPIN9.【Method】The specific target sequence was designed according to OsPIN9 genome sequence and OsPIN9 genome editing vector was constructed. Nippobare (Oryza sativa japonica) was used as the material and the hygromycin-resistant rice was obtained by Agrobacterium-mediated transformation. The positive transgenic lines were screened by PCR. The mutation sites were confirmed by the combination of PCR and subsequent analysis of sequencing results, the homozygous mutants were obtained and the difference of amino acid sequence and tertiary structure of OsPIN9 protein was analyzed between WT and ospin9 mutants. The expression of OsPINs genes in mutant roots was performed by quantitative real-time PCR (qRT-PCR), and the phenotype of ospin9 mutants was analyzed at the seedling stage. The effects of 1-naphthaleneacetic acid (NAA) treatment on seedling development were also analyzed under 0.05 μmol·L -1 NAA for 7 d.【Result】The target site sequence was designed based on the sequence of exon1 of OsPIN9 and, subsequently, the OsPIN9 genome editing recombinant vector was constructed. A total of 18 independent transgenic lines were obtained by transformation. Sequencing analysis revealed that three different mutation types were present in 7 T0 generation lines, including 3 lines with T insertion, 3 lines with G insertion and 1 line with C insertion, and all the mutation sites happened at the 18 th base of the target sequence. Two homozygous mutation lines were further identified in the T1 generation. BLAST analysis showed that the two types of OsPIN9 mutations caused frame-shift mutation and premature termination of translation, and the mutation protein was shortened from 426 aa in WT to 172 aa, thus leading to the complete disappearance of the transmembrane helices. qPCR analysis indicated that the transcription abundance of OsPIN9 significantly decreased in ospin9 mutants compared with WT, OsPIN1a and OsPIN5b were up-regulated, while OsPIN5a was down-regulated in ospin9 mutants. Both the shoot height and the number of adventitious roots of ospin9 mutants were reduced significantly than that of WT, while its root length was comparable to that of WT. The plant growth was inhibited and the adventitious root number was still less than that of WT under NAA treatment, but no significant difference was found between ospin9 mutants and WT plants. 【Conclusion】 Auxin efflux carrier OsPIN9 was directionally edited by using CRISPR/Cas9 technology, and two transgene-free homozygous ospin9 mutants were obtained. The mutation of OsPIN9 affected the expression level of other OsPINs genes, the shoot and root development were inhibited in ospin9 mutants at the seedling stage and NAA treatment partially rescued the development of adventitious roots in ospin9 mutants. Keywords:rice;OsPIN9;polar auxin transport;CRISPR/Cas9
PDF (2333KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 吴世洋, 杨晓祎, 张艳雯, 侯典云, 胥华伟. 利用CRISPR/Cas9基因编辑技术构建水稻ospin9突变体. 中国农业科学, 2021, 54(18): 3805-3817 doi:10.3864/j.issn.0578-1752.2021.18.002 WU ShiYang, YANG XiaoYi, ZHANG YanWen, HOU DianYun, XU HuaWei. Generation of ospin9 Mutants in Rice by CRISPR/Cas9 Genome Editing Technology. Scientia Acricultura Sinica, 2021, 54(18): 3805-3817 doi:10.3864/j.issn.0578-1752.2021.18.002
以MEGA7进行蛋白序列比对[45],比对结果进一步通过Jalview进行修饰[46]。通过在线网站(TMHMM Server v. 2.0: http://www.cbs.dtu.dk/services/TMHMM/)分析蛋白的跨膜螺旋结构域[47]。通过SWISS MODEL网站(https://swissmodel.expasy.org/)预测蛋白三级结构[48]。
OsPIN9主要在根及根茎结合部表达,此外,水稻根中还检测到OsPIN1a、OsPIN1b、OsPIN1c、OsPIN2、OsPIN5a和OsPIN5b的表达[20],以14 d幼苗根为材料对目的基因及根中OsPINs进行qRT-PCR表达分析。用RNAiso Plus(TaKaRa BIO INC)提取总RNA,运用HiScript III RT SuperMix for qPCR(+gDNA wiper)(南京诺唯赞生物科技股份有限公司)反转录得到cDNA。使用AceQ Universal SYBR qPCR Master Mix(南京诺唯赞生物科技股份有限公司)在Lightcycle® 96 qPCR仪上进行qRT-PCR分析,以水稻OsACTIN1(Os03g0718100)作为内参,以2-ΔΔCT法计算基因的相对表达量。试验进行3次生物学重复,每次3个技术重复。所用引物见表1。
A:幼苗期ospin9突变体照片;B:表型数据统计分析(n≥14)。黑色圆点表示测量值;**表示P<0.01水平差异;***表示P<0.001水平差异。下同 Fig. 7Phenotype of ospin9 mutants at the seedling stage
A: Photograph of ospin9 mutants at the seedling stage; B: Statistical analysis of the phenotypic data (n≥14). Black dot indicates measured data; * indicates difference at the P<0.01 level; *** indicates difference at the P<0.001 level. The same as below
A、B:对照植株株高及不定根数统计(n=12);C:0.05 μmol·L-1 NAA处理7 d株高和不定根数统计(n=12)。*表示P<0.05水平差异 Fig. 8Effects of NAA treatment on seedling shoot height and adventitious root number
A, B: Statistical analysis of shoot height and adventitious root number in untreated plants (n=12); C: Statistical analysis of shoot height and adventitious root number under 0.05 μmol·L-1 NAA treated for 7 d (n=12). * indicates difference at the P<0.05 level
KARKIS, RIZALG, QUICK WP. Improvement of photosynthesis in rice (Oryza sativa L.) by inserting the C4 pathway Rice, 2013, 6:28. DOI:10.1186/1939-8433-6-28URL [本文引用: 1]
DUBROVSKY JG, SAUERM, NAPSUCIALY-MENDIVILS, IVANCHENKO MG, FRIMLJ, SHISHKOVAS, CELENZAJ, BENKOVAE. Auxin acts as a local morphogenetic trigger to specify lateral root founder cells Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(25): 8790-8794. [本文引用: 1]
MRAVECJ, SKUPAP, BAILLYA, HOYEROVAK, KRECEKP, BIELACHA, PETRASEKJ, ZHANGJ, GAYKOVAV, STIERHOF YD, DOBREV PI, SCHWARZEROVAK, ROLCIKJ, SEIFERTOVAD, LUSCHNIGC, BENKOVAE, ZAZIMALOVAE, GEISLERM, FRIMLJ. Subcellular homeostasis of phytohormone auxin is mediated by the ER-localized PIN5 transporter Nature, 2009, 459(7250): 1136-1140. DOI:10.1038/nature08066URL [本文引用: 2]
HAGAK, SAKAIT. Differential roles of auxin efflux carrier PIN proteins in hypocotyl phototropism of etiolated Arabidopsis seedlings depend on the direction of light stimulus Plant Signalling & Behavior, 2013, 8(1): e22556. [本文引用: 1]
ZHANG KX, XU HH, YUAN TT, ZHANGL, LU YT. Blue-light-induced PIN3 polarization for root negative phototropic response in Arabidopsis The Plant Journal, 2013, 76(2): 308-321. [本文引用: 1]
CHENR, HILSONP, SEDBROOKJ, ROSENE, CASPART, MASSON PH. The Arabidopsis thaliana AGRAVITROPIC 1 gene encodes a component of the polar-auxin-transport efflux carrier Proceedings of the National Academy of Sciences of the United States of America, 1998, 95(25): 15112-15117. [本文引用: 1]
KLEINE-VEHNJ, LEITNERJ, ZWIEWKAM, SAUERM, ABASL, LUSCHNIGC, FRIMLJ. Differential degradation of PIN2 auxin efflux carrier by retromer-dependent vacuolar targeting Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(46): 17812-17817. [本文引用: 1]
RAHMANA, TAKAHASHIM, SHIBASAKIK, WUS, INABAT, TSURUMIS, BASKIN TI. Gravitropism of Arabidopsis thaliana roots requires the polarization of PIN2 toward the root tip in meristematic cortical cells The Plant Cell, 2010, 22(6): 1762-1776. DOI:10.1105/tpc.110.075317URL [本文引用: 1]
YANGY, HAMMES UZ, TAYLOR CG, SCHACHTMAN DP, NIELSENE. High-affinity auxin transport by the AUX1 influx carrier protein Current Biology, 2006, 16(11): 1123-1127. DOI:10.1016/j.cub.2006.04.029URL [本文引用: 1]
NOHB, MURPHY AS, SPALDING EP. Multidrug resistance-like genes of Arabidopsis required for auxin transport and auxin-mediated development The Plant Cell, 2001, 13(11): 2441-2454. [本文引用: 1]
GEISLERM, MURPHY AS. The ABC of auxin transport: the role of p-glycoproteins in plant development FEBS Letters, 2006, 580(4): 1094-1102. DOI:10.1016/j.febslet.2005.11.054URL [本文引用: 1]
GRIENEISEN VA, XUJ, MAREE AF, HOGEWEGP, SCHERESB. Auxin transport is sufficient to generate a maximum and gradient guiding root growth Nature, 2007, 449(7165): 1008-1013. DOI:10.1038/nature06215URL [本文引用: 1]
WANGJ, HUH, WANGG, LIJ, CHENJ, WUP. Expression of PIN genes in rice (Oryza sativa L.): Tissue specificity and regulation by hormones Molecular Plant, 2009, 2(4): 823-831. DOI:10.1093/mp/ssp023URL [本文引用: 4]
MIYASHITAY, TAKASUGIT, ITOY. Identification and expression analysis of PIN genes in rice Plant Science, 2010, 178(5): 424-428. DOI:10.1016/j.plantsci.2010.02.018URL [本文引用: 1]
KRECEKP, SKUPAP, LIBUSJ, NARAMOTOS, TEJOSR, FRIMLJ, ZAZIMALOVA E. The PIN-FORMED (PIN) protein family of auxin transporters Genome Biology, 2009, 10(12): 249. DOI:10.1186/gb-2009-10-12-249URL [本文引用: 2]
ADAMOWSKIM, FRIMLJ. PIN-dependent auxin transport: action, regulation, and evolution The Plant Cell, 2015, 27(1): 20-32. DOI:10.1105/tpc.114.134874URL [本文引用: 1]
DAL BOSCOC, DOVZHENKOA, LIUX, WOERNERN, RENSCHT, EISMANNM, EIMERS, HEGERMANNJ, PAPONOV IA, RUPERTIB, HEBERLE-BORSE, TOURAEVA, COHEN JD, PALMEK. The endoplasmic reticulum localized PIN8 is a pollen-specific auxin carrier involved in intracellular auxin homeostasis The Plant Journal, 2012, 71(5): 860-870. DOI:10.1111/tpj.2012.71.issue-5URL [本文引用: 1]
DINGZ, WANGB, MORENOI, DUPLAKOVAN, SIMONS, CARRARON, REEMMERJ, PENCIKA, CHENX, TEJOSR, SKUPAP, POLLMANNS, MRAVECJ, PETRASEKJ, ZAZIMALOVAE, HONYSD, ROLCIKJ, MURPHYA, ORELLANAA, GEISLERM, FRIMLJ. ER-localized auxin transporter PIN8 regulates auxin homeostasis and male gametophyte development in Arabidopsis Nature Communications, 2012, 3:941. DOI:10.1038/ncomms1941URL
FERARUE, VOSOLSOBES, FERARU MI, PETRASEKJ, KLEINE-VEHNJ. Evolution and structural diversification of PILS putative auxin carriers in plants Frontiers in Plant Science, 2012, 3:227.
WANGY, CHAIC, VALLIYODANB, MAUPINC, ANNENB, NGUYEN HT. Genome-wide analysis and expression profiling of the PIN auxin transporter gene family in soybean (Glycine max) BMC Genomics, 2015, 16:951. DOI:10.1186/s12864-015-2149-1URL [本文引用: 2]
ZHANGY, HEP, YANGZ, HUANGG, WANGL, PANGC, XIAOH, ZHAOP, YUJ, XIAOG. A genome-scale analysis of the PIN gene family reveals its functions in cotton fiber development Frontiers in Plant Science, 2017, 8:461. [本文引用: 2]
LIY, ZHUJ, WUL, SHAOY, WUY, MAOC. Functional divergence of PIN1 paralogous genes in rice Plant and Cell Physiology, 2019, 60(12): 2720-2732. DOI:10.1093/pcp/pcz159URL [本文引用: 4]
CHENY, FANX, SONGW, ZHANGY, XUG. Over-expression of OsPIN2 leads to increased tiller numbers, angle and shorter plant height through suppression of OsLAZY1 Plant Biotechnology Journal, 2012, 10(2): 139-149. DOI:10.1111/pbi.2011.10.issue-2URL [本文引用: 1]
WANGL, GUOM, LIY, RUANW, MOX, WUZ, STURROCK CJ, YUH, LUC, PENGJ, MAOC. LARGE ROOT ANGLE1, encoding OsPIN2, is involved in root system architecture in rice Journal of Experimental Botany, 2018, 69(3): 385-397. DOI:10.1093/jxb/erx427URL [本文引用: 1]
INAHASHIH, SHELLEY IJ, YAMAUCHIT, NISHIUCHIS, TAKAHASHI NOSAKAM, MATSUNAMIM, OGAWAA, NODAY, INUKAIY. OsPIN2, which encodes a member of the auxin efflux carrier proteins, is involved in root elongation growth and lateral root formation patterns via the regulation of auxin distribution in rice Physiologia Plantarum, 2018, 164(2): 216-225. DOI:10.1111/ppl.2018.164.issue-2URL [本文引用: 1]
WUD, SHENH, YOKAWAK, BALUSKAF. Alleviation of aluminium-induced cell rigidity by overexpression of OsPIN2 in rice roots Journal of Experimental Botany, 2014, 65(18): 5305-5315. DOI:10.1093/jxb/eru292URL [本文引用: 1]
WUD, SHENH, YOKAWAK, BALUŠKAF. Overexpressing OsPIN2 enhances aluminium internalization by elevating vesicular trafficking in rice root apex Journal of Experimental Botany, 2015, 66(21): 6791-6801. DOI:10.1093/jxb/erv385URL [本文引用: 1]
LUG, CONEVAV, CASARETTO JA, YINGS, MAHMOODK, LIUF, NAMBARAE, BI YM, ROTHSTEIN SJ. OsPIN5b modulates rice (Oryza sativa) plant architecture and yield by changing auxin homeostasis, transport and distribution The Plant Journal, 2015, 83(5): 913-925. DOI:10.1111/tpj.2015.83.issue-5URL [本文引用: 1]
ZHANGQ, LIJ, ZHANGW, YANS, WANGR, ZHAOJ, LIY, QIZ, SUNZ, ZHUZ. The putative auxin efflux carrier OsPIN3t is involved in the drought stress response and drought tolerance The Plant Journal, 2012, 72(5): 805-816. DOI:10.1111/tpj.2012.72.issue-5URL [本文引用: 1]
HOU MM, LUO FF, WU DX, ZHANG XH, LOU MM, SHEN DF, YANM, MAO CZ, FAN XR, XU GH, ZHANG YL. OsPIN9, an auxin efflux carrier, is required for the regulation of rice tiller bud outgrowth by ammonium New Phytologist, 2021, 229:935-949. DOI:10.1111/nph.v229.2URL [本文引用: 5]
HSIEH PH, KAN CC, WU HY, YANG HC, HSIEH MH. Early molecular events associated with nitrogen deficiency in rice seedling roots Scientific Reports, 2018, 8(1): 12207. DOI:10.1038/s41598-018-30632-1URL [本文引用: 1]
QI YB, ZHANG LX, WANG LY, SONGJ, WANG JJ. CRISPR/Cas9 targeted editing for the fragrant gene Badh2 in rice Scientia Agricultura Sinica, 2020, 53(8): 1501-1509. (in Chinese) [本文引用: 2]
LIU YG, LI GS, ZHANG YL, CHEN LT. Current advances on CRISPR/Cas genome editing technologies in plants Journal of South China Agricultural University, 2019, 40(5): 38-49. (in Chinese) [本文引用: 2]
LIU WZ, XIE XR, MA XL, LIJ, CHEN JH, LIU YG. DSDecode: A web-based tool for decoding of sequencing chromatograms for genotyping of targeted mutations Molecular Plant, 2015, 8(9): 1431-1433. DOI:10.1016/j.molp.2015.05.009URL [本文引用: 1]
MA XL, CHEN LT, ZHU QL, CHEN YL, LIU YG. Rapid decoding of sequence-specific nuclease-induced heterozygous and biallelic mutations by direct sequencing of PCR products Molecular Plant, 2015, 8(8): 1285-1287. DOI:10.1016/j.molp.2015.02.012URL [本文引用: 1]
KUMARS, STECHERG, TAMURAK. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets Molecular Biology and Evolution, 2016, 33(7): 1870-1874. DOI:10.1093/molbev/msw054URL [本文引用: 1]
WATERHOUSE AM, PROCTER JB, MARTIND M A, CLAMPM, BARTONG J. Jalview Version 2-a multiple sequence alignment editor and analysis workbench Bioinformatics, 2009, 25(9): 1189-1191. DOI:10.1093/bioinformatics/btp033URL [本文引用: 1]
KROGHA, LARSSONB, VON HEIJNEG, SONNHAMMER EL. Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes Journal of Molecular Biology, 2001, 305(3): 567-580. DOI:10.1006/jmbi.2000.4315URL [本文引用: 1]
MADEIRAF, PARK YM, LEEJ, BUSON, GURT, MADHUSOODANANN, BASUTKARP, TIVEYA, POTTER SC, FINN RD, LOPEZR. The EMBL-EBI search and sequence analysis tools APIs in 2019 Nucleic Acids Research, 2019, 47(W1): W636-W641. DOI:10.1093/nar/gkz268URL [本文引用: 1]
YOSHIDAS, FORNO DA, COCK JH, GOMEZ KA. Laboratory Manual for Physiological Studies of Rice Manila: International Rice Research Institute, 1976. [本文引用: 1]
BENNETTT, BROCKINGTON SF, ROTHFELSC, GRAHAM SW, STEVENSOND, KUTCHANT, ROLFM, THOMASP, WONG GK, LEYSERO, GLOVER BJ, HARRISON CJ. Paralogous radiations of PIN proteins with multiple origins of noncanonical PIN structure Molecular Biology and Evolution, 2014, 31(8): 2042-2060. DOI:10.1093/molbev/msu147URL [本文引用: 2]
MA XL, ZHANG QY, ZHU QL, LIUW, CHENY, QIUR, WANGB, YANG ZF, LI HY, LIN YY, XIE YY, SHEN RX, CHEN SF, WANGZ, CHEN YL, GUO JX, CHEN LT, ZHAO XC, DONG ZC, LIU YG. A robust CRISPR/Cas9 system for convenient, high-efficiency multiplex genome editing in monocot and dicot plants Molecular Plant, 2015, 8(8): 1274-1284. DOI:10.1016/j.molp.2015.04.007URL [本文引用: 1]
WANG MG, MAO YF, LU YM, TAO XP, ZHU JK. Multiplex gene editing in rice using the CRISPR-Cpf1 system Molecular Plant, 2017, 10(7): 1011-1013. DOI:10.1016/j.molp.2017.03.001URL [本文引用: 1]
ZHANGY, LIANGZ, ZONGY, WANG YP, LIU JX, CHEN KL, QIU JL, GAO CX. Efficient and transgene-free genome editing in wheat through transient expression of CRISPR/Cas9 DNA or RNA Nature Communications, 2016, 7(1): 12617-12617. DOI:10.1038/ncomms12617URL [本文引用: 1]
LIANGZ, CHEN KL, LI TD, ZHANGY, WANG YP, ZHAOQ, LIU JX, ZHANG HW, LIU CM, RAN YD, GAO CX. Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes Nature Communications, 2017, 8:14261. DOI:10.1038/ncomms14261URL [本文引用: 1]
HUANG ZM, ZHOU YB, TANG XD, ZHAO XH, ZHOU ZW, FU XX, WANGK, SHI JW, LI YF, FU CJ, YANG YZ. Construction of tms5 mutants in rice based on CRISPR/Cas9 technology Acta Agronomica Sinica, 2018, 44(6): 844-851. (in Chinese) DOI:10.3724/SP.J.1006.2018.00844URL [本文引用: 1]
WANG MN, PENG JJ, WANG KJ, AN WJ, LIU YF, LI KJ, LIANG WH. Editing ROP gene OsRac5 of rice by CRISPR/Cas9 technique Chinese Journal of Biochemistry and Molecular Biology, 2018, 34(12): 1350-1357. (in Chinese) [本文引用: 1]
XUP, WANGH, TU RR, LIU QE, WU WX, FU XM, CAO LY, SHEN XH. Orientation improvement of blast resistance in rice via CRISPR/Cas9 system Chinese Journal of Rice Science, 2019, 33(4): 313-322. (in Chinese) [本文引用: 1]
LONG QZ, HUANG YL, TANG XY, WANG HM, LUM, YUAN LF, WAN JL. Creation of low-Cd-accumulating indica rice by disruption of OsNramp5 gene via CRISPR/Cas9 Chinese Journal of Rice Science, 2019, 33(5): 407-420. (in Chinese) [本文引用: 1]
XU SB, ZHENG HL, LIU LF, BU QY, LI XF, ZOU DT. Improvement of grain shape and fragrance by using CRISPR/Cas9 system Chinese Journal of Rice Science, 2020, 34(5): 406-412. (in Chinese) [本文引用: 1]
ZHOU TS, YUD, LIUL, OU YN, YUAN GL, DUAN MJ, YUAN DY. CRISPR/Cas9-mediated editing of AFP1 improves rice stress tolerance Chinese Journal of Rice Science, 2021, 35(1): 11-18. (in Chinese) [本文引用: 1]
XUM, ZHUL, SHOU HX, WUP. A PIN1 family gene, OsPIN1, involved in auxin-dependent adventitious root emergence and tillering in rice Plant and Cell Physiology, 2005, 46(10): 1674-1681. DOI:10.1093/pcp/pci183URL [本文引用: 1]
WANGT, LI CX, WU ZH, JIA YC, WANGH, SUN SY, MAO CZ, WANG XL. Abscisic acid regulates auxin homeostasis in rice root tips to promote root hair elongation Frontiers in Plant Science, 2017, 8:1121. DOI:10.3389/fpls.2017.01121URL [本文引用: 1]
ZHANG XW, LI JP, LIU AL, ZOUJ, ZHOU XY, XIANG JH, RERKSIRIW, PENGY, XIONG XY, CHEN XB. Expression profile in rice panicle: Insights into heat response mechanism at reproductive stage PLoS ONE, 2012, 7(11): e49652. DOI:10.1371/journal.pone.0049652URL [本文引用: 1]
 ,, 杨晓祎, 张艳雯, 侯典云, 胥华伟
,, 杨晓祎, 张艳雯, 侯典云, 胥华伟

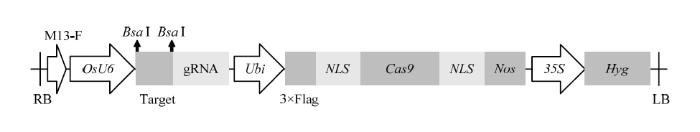 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT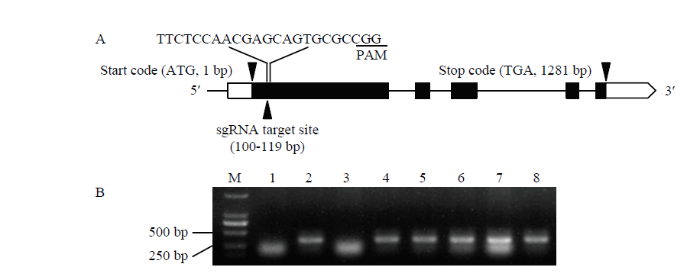 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT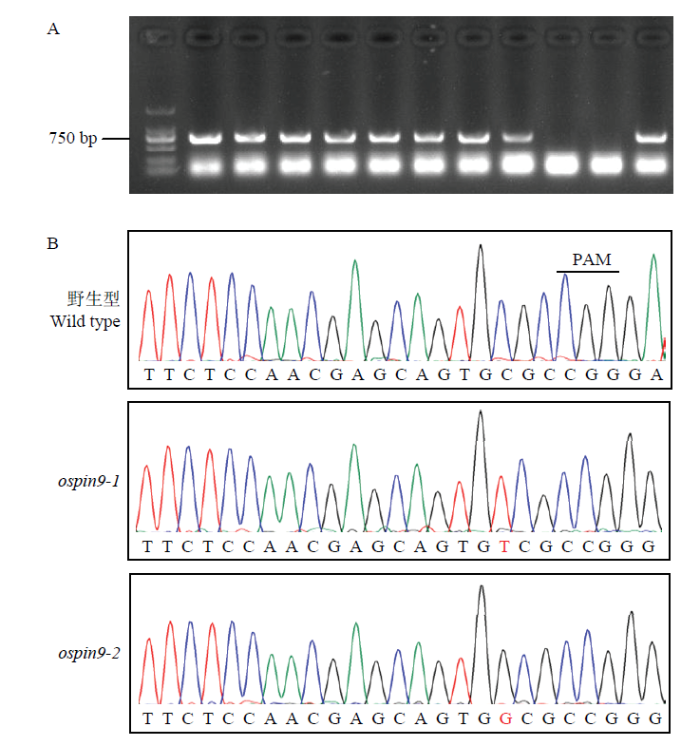 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT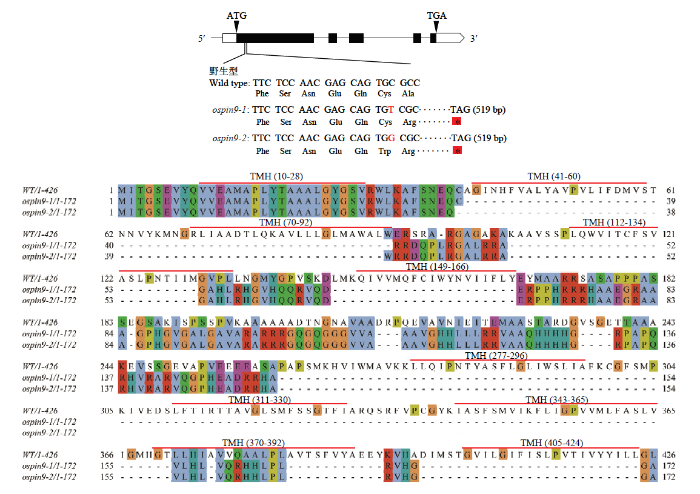 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT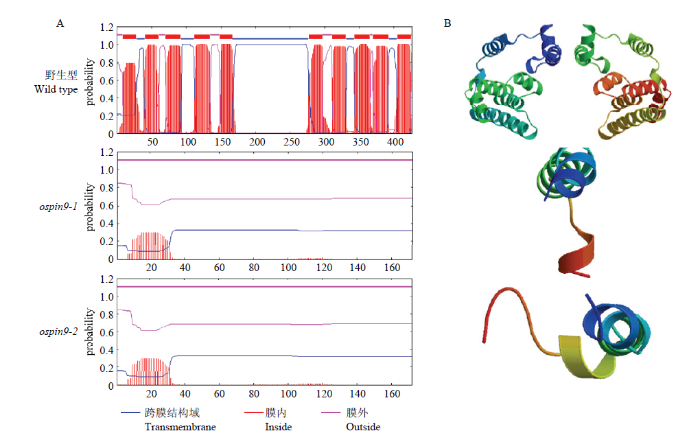 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT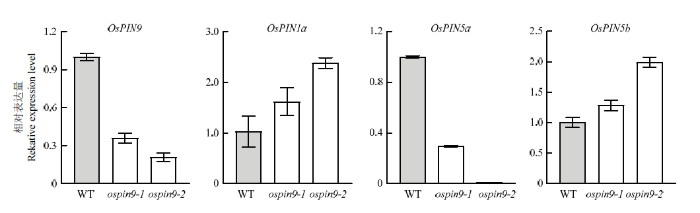 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT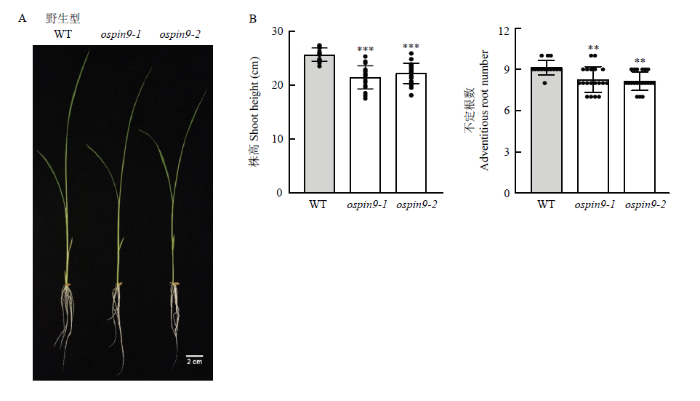 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT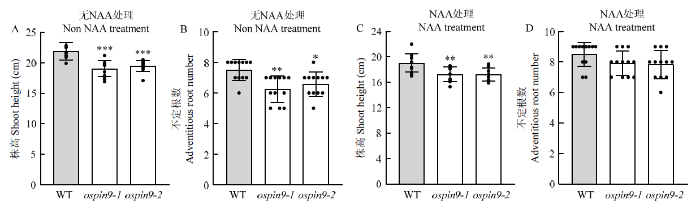 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}