Screening of ARF-Aux/IAA Interaction Combinations Involved in Apple Fruit Size
ZHOU Zhe,, BIAN ShuXun, ZHANG HengTao, ZHANG RuiPing, GAO QiMing, LIU ZhenZhen, YAN ZhenLi,Zhengzhou Fruit Research Institute, Chinese Academy of Agricultural Sciences, Zhengzhou 459000
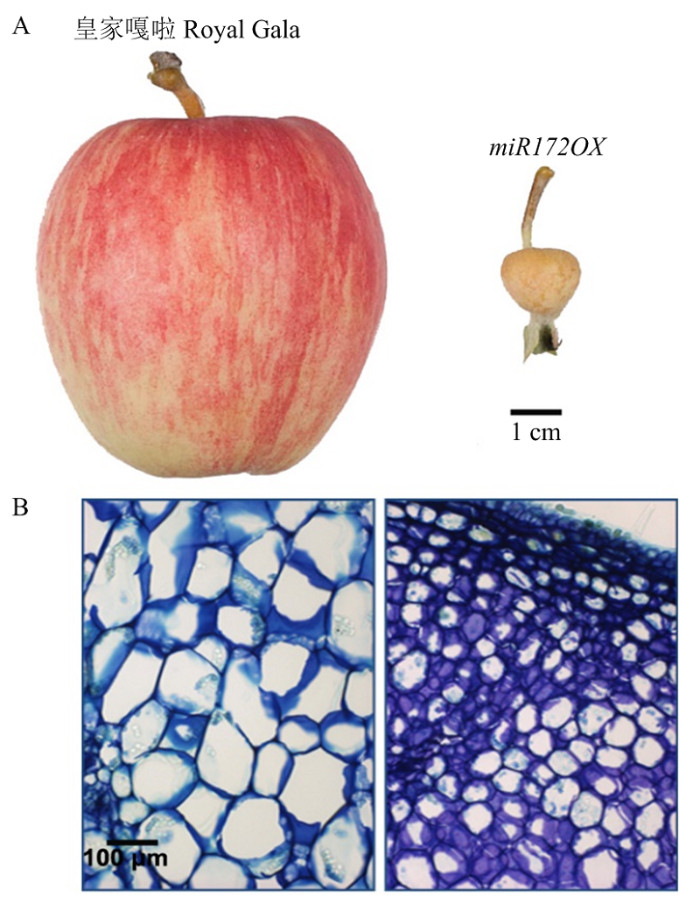
Abstract 【Objective】The transcriptomics and bioinformatics methods were used to carry out the genome-wide potential interacting MdARFs and MdIAAs pairs screening, so as to build a foundation for clarifying the function of related genes and elucidating the molecular mechanism underlying auxin-regulated apple fruit size.【Method】 Two apple genotypes, Royal Gala (WT) and 35S::miRNA172p transgenic Royal Gala (miR172OX ) were used as test materials in this study. Fruit materials from different developmental stages and tissue types were collected for both genotypes and were subjected to transcriptome sequencing. Clean reads were aligned to the reference genome and the differential expression analysis was performed. Based on the transcriptome data obtained by sequencing the transgenic small fruit and wild-type large fruit, the pairwise expression analysis was performed across MdARFs and MdIAAs families. The amino acid sequences of 23 ARFs and 34 Aux/IAAs were downloaded from Arabidopsis genome, and 21 ARFs and 25 Aux/IAAs were downloaded from tomato genome, which were further compared with candidate MdARFs and MdIAAs to construction phylogenetic trees. The MEME and TBtools were used to carry out the Motif analysis for candidate MdARFs and MdIAAs. Pairs with high interacting possibilities were further confirmed by a protein-protein interacting network constructed in apple to finalize combinations with the highest probability of involvement in fruit development. 【Result】 The whole fruit at 2 weeks post full bloom and the fruit skin, fruit flesh and fruit core at 4 WPFB were collected from WT and miR172OX, respectively. To achieve research objectives, transcriptome sequencing was carried out. A total of 178.19 Gb paired-end reads of 125 bp/150 bp were generated. All indexes indicated that the three biological replicates had highly consistent transcriptome profiles across all tissue types. FPKM values in at least one library was over 2 were used as a standard to eliminate the low expressed genes, so a total of 38MdARFs and 27 MdIAAs were expressed. In our fruit developmental transcriptome data, eight pairs of MdARF-MdIAA were obtained through Pearson correlation analysis, whose Pearson correlation coefficient was over 0.9 or below -0.9. The systematic phylogenetic analysis showed that MdARF6 and MdARF19 belonged to the same branch with AtARFs, which played a role in transcription activation, while MdARF2, MdARF4, and MdARF9 were closely related to transcriptional inhibitory AtARFs. Motif analysis results showed that both the candidate MdARFs and MdIAAs proteins contained Motif 2 and Motif 5, which were corresponded to the conserved domains Motif IV and Motif III in the IAA protein, respectively. After homolog mapping inspection with Arabidopsis, two potential MdARF-MdIAA interacting pairs were selected for future functional identification. 【Conclusion】Among apple MdARF and MdIAA family members, eight pairs of MdARF-MdIAA showed significant correlations in terms of their expression patterns during fruit development. Further homology mapping confirmed two pairs of them, including MdARF4-MdIAA17 and MdARF4-MdIAA19, were most likely to participate in the regulation of apple fruit development through mediating auxin signal transduction. Keywords:apple;ARF;Aux/IAA;interacting-pair screening
PDF (3387KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 周喆, 卞书迅, 张恒涛, 张瑞萍, 高启明, 刘珍珍, 阎振立. 苹果果实大小相关的ARF-Aux/IAA互作组合筛选[J]. 中国农业科学, 2021, 54(14): 3088-3096 doi:10.3864/j.issn.0578-1752.2021.14.014 ZHOU Zhe, BIAN ShuXun, ZHANG HengTao, ZHANG RuiPing, GAO QiMing, LIU ZhenZhen, YAN ZhenLi. Screening of ARF-Aux/IAA Interaction Combinations Involved in Apple Fruit Size[J]. Scientia Acricultura Sinica, 2021, 54(14): 3088-3096 doi:10.3864/j.issn.0578-1752.2021.14.014
A:热图显示苹果ARF和IAA基因家族表达量的聚类分析;B:苹果ARF和IAA基因家族中的差异表达基因及差异表达趋势 Fig. 2Expression analysis of MdARF and MdIAA encoding genes
A: Transcriptional heat maps showing expression levels of apple ARF and IAA encoding genes; B: Differentially expressed genes in apple ARF and IAA gene families and their differential expression trends
A:苹果候选ARF-Aux/IAA互作对中MdARFs和MdIAAs的系统发育分析;B:苹果候选ARF-Aux/IAA互作对中MdARFs和MdIAAs的保守结构域分析 Fig. 4The phylogenetic tree and conserved motif analysis of interacting candidate combination members from ARF and IAA gene families in apple
A: Phylogenetic analysis of MdARFs and MdIAAs in apple candidate ARF-Aux/IAA interaction pairs; B: Conserved domain analysis of MdARFs and MdIAAs in apple candidate ARF-Aux/IAA interaction pairs
节点大小对应节点连接数量,节点颜色对应Log2(Foldchange)值,节点间连线颜色对应PCC值 Fig. 5Putative Protein-Protein Interaction network involving in MdARFs and MdIAAs in apple
Node sizes correspond to the degree of connectivity of the nodes, nodes are color-coded according to the Log2 (Foldchange) values and edge are color-coded according to the PCC values
SUGIMOTO-SHIRASUK, ROBERTSK. "Big it up": Endoreduplication and cell-size control in plants Current Opinion in Plant Biology, 2003, 6:544-553. DOI:10.1016/j.pbi.2003.09.009URL [本文引用: 1]
HARADAT, KURAHASHIW, YANAIM, WAKASAY, SATOHT. Involvement of cell proliferation and cell enlargement in increasing the fruit size of Malus species Scientia Horticulturae, 2005, 105(4):447-456. DOI:10.1016/j.scienta.2005.02.006URL [本文引用: 1]
MARIOTTIL, PICCIARELLIP, LOMBARDIL, CECCARELLIN. Fruit-set and early fruit growth in tomato are associated with increases in indoleacetic acid, cytokinin, and bioactive gibberellin contents Journal of Plant Growth Regulation, 2011, 30:405-415. DOI:10.1007/s00344-011-9204-1URL [本文引用: 1]
PEI MS, CAO SH, WUL, WANG GM, XIE ZH, GUC, LING ZS. Comparative transcriptome analyses of fruit development among pears, peaches, and strawberries provide new insights into single sigmoid patterns BMC Plant Biology, 2020, 20(1):108. DOI:10.1186/s12870-020-2317-6URL [本文引用: 1]
PIYAS, SHRESTHA SK, BINDERB, NEAL STEWART JRC, HEWEZIT. Protein-protein interaction and gene co-expression maps of ARFs and Aux/IAAs in Arabidopsis Frontiers in Plant Science, 2014, 5:744. [本文引用: 1]
AUDRAN-DELALANDEC, BASSAC, MILAI, REGADF, ZOUINEM, BOUZAYENM. Genome-wide identification, functional analysis and expression profiling of the Aux/IAA gene family in tomato Plant and Cell Physiology, 2012, 53(4):659-672. DOI:10.1093/pcp/pcs022URL [本文引用: 2]
HUX, HOUX, YUANX, GUAND, LIU RP. Research progress on mechanism of ARF and Aux/IAA regulating fruit development and ripening Biotechnology Bulletin, 2017, 33(12):37-44. (in Chinese) [本文引用: 1]
LISCUME, REED JW. Genetics of Aux/IAA and ARF action in plant growth and development Plant Molecular Biology, 2002, 49(3/4):387-400. DOI:10.1023/A:1015255030047URL [本文引用: 2]
ULMASOVT, HAGENG, GUILFOYLE TJ. Activation and repression of transcription by auxin-response factors Proceedings of the National Academy of Sciences of the United States of America, 1999, 96(10):5844-5849. [本文引用: 2]
TIWARI SB, HAGENG, GUILFOYLE TJ. Aux/IAA proteins contain a potent transcriptional repression domain Plant Cell, 2004, 16(2):533-543. DOI:10.1105/tpc.017384URL [本文引用: 1]
SZEMENYEIH, HANNONM, LONG JA. TOPLESS mediates auxin-dependent transcriptional repression during Arabidopsis embryogenesis Science, 2008, 319(5868):1384-1386. DOI:10.1126/science.1151461URL [本文引用: 1]
ULMASOVT, HAGENG, GUILFOYLE TJ. ARF1, a transcription factor that binds to auxin response elements Science, 1997, 276(5320):1865-1868. DOI:10.1126/science.276.5320.1865URL [本文引用: 1]
SHEN CJ, YUE RQ, SUNT, ZHANGL, XU LQ, TIE SG, WANG HZ, YANG YJ. Genome-wide identification and expression analysis of auxin response factor gene family in Medicago truncatula Frontiers in Plant Science, 2015, 6:73. [本文引用: 1]
LI JN, YAN XJ, LI SH, ZHANG RS. Plants AUX/IAA gene family: Research progress Chinese Agricultural Science Bulletin, 2018, 34(15):89-92. (in Chinese) [本文引用: 1]
VERNOUXT, BRUNOUDG, FARCOTE, MORINV, VAN DEN DAELEH, LEGRANDJ, OLIVAM, DASP, LARRIEUA, WELLSD, GUEDONY, ARMITAGEL, PICARDF, GUYOMARC'HS, CELLIERC, PARRYG, KOUMPROGLOUR, DOONAN JH, ESTELLEM, GODINC, KEPINSKIS, BENNETTM, DE VEYLDERL, TRAASJ. The auxin signalling network translates dynamic input into robust patterning at the shoot apex Molecular Systems Biology, 2011, 7:508. DOI:10.1038/msb.2011.39URL [本文引用: 2]
LLERESD, SWIFTS, LAMOND AI. Detecting protein-protein interactions in vivo with FRET using multiphoton fluorescence lifetime imaging microscopy (FLIM) Current Protocols in Cytometry, 2007, 42(1). [本文引用: 1]
SHINOZAKIY, NICOLASP, FERNANDEZ-POZON, MAQ, EVANICH DJ, SHIY, XUY, ZHENGY, SNYDER SI, MARTIN L BB, RUIZ-MAYE, THANNHAUSER TW, CHENK, DOMOZYCH DS, CATALAC, FEIZ, MUELLER LA, GIOVANNONI JJ, ROSE J KC. High-resolution spatiotemporal transcriptome mapping of tomato fruit development and ripening Nature Communications, 2018, 9(1):364. DOI:10.1038/s41467-017-02782-9URL [本文引用: 2]
TATEMATSUK, KUMAGAIS, MUTOH, SATOA, WATAHIKI MK, HARPER RM, LISCUME, YAMAMOTO KT. MASSUGU2 encodes Aux/IAA19, an auxin-regulated protein that functions together with the transcriptional activator NPH4/ARF7 to regulate differential growth responses of hypocotyl and formation of lateral roots in Arabidopsis thaliana Plant Cell, 2004, 16(2):379-393. DOI:10.1105/tpc.018630URL [本文引用: 1]
ARASEF, NISHITANIH, EGUSAM, NISHIMOTON, SAKURAIS, SAKAMOTON, KAMINAKAH. IAA8 involved in lateral root formation interacts with the TIR1 auxin receptor and ARF transcription factors in Arabidopsis PLoS ONE, 2012, 7(8):e43414. DOI:10.1371/journal.pone.0043414URL [本文引用: 1]
SHENC, WANGS, BAIY, WUY, ZHANGS, CHENM, GUILFOYLE TJ, WUP, QIY. Functional analysis of the structural domain of ARF proteins in rice (Oryza sativaL.) Journal of Experimental Botany, 2010, 61(14):3971-3981. DOI:10.1093/jxb/erq208URL [本文引用: 1]
WANG YC, WANGN, XU HF, JIANG SH, FANG HC, SU MY, ZHANG ZY, ZHANG TL, CHEN XS. Auxin regulates anthocyanin biosynthesis through the Aux/IAA-ARF signaling pathway in apple Horticulture Research, 2018, 5:59. DOI:10.1038/s41438-018-0068-4URL [本文引用: 1]
YAO JL, XUJ, CORNILLEA, TOMESS, KARUNAIRETNAMS, LUOZ, BASSETTH, WHITWORTHC, REES-GEORGEJ, RANATUNGAC, SNIRCA, CROWHURSTR, DE SILVAN, WARRENB, DENGC, KUMARS, CHAGNED, BUS VG, VOLZ RK, RIKKERINK EH, GARDINER SE, GIRAUDT, MACDIARMIDR, GLEAVE AP. A microRNA allele that emerged prior to apple domestication may underlie fruit size evolution Plant Journal, 2015, 84(2):417-427. DOI:10.1111/tpj.13021URL [本文引用: 2]
ZHOUZ, CONG PH, TIANY, ZHU YM. Using RNA-seq data to select reference genes for normalizing gene expression in apple roots PLoS ONE, 2017, 12(9):e0185288. DOI:10.1371/journal.pone.0185288URL [本文引用: 1]
DACCORDN, CELTON JM, LINSMITHG, BECKERC, CHOISNEN, SCHIJLENE, VAN DE GEESTH, BIANCOL, MICHELETTID, VELASCOR, DI PIERRO EA, GOUZYJ, REES D JG, GUERIFP, MURANTYH, DUREL CE, LAURENSF, LESPINASSEY, GAILLARDS, AUBOURGS, QUESNEVILLEH, WEIGELD, VAN DE WEGE, TROGGIOM, BUCHERE. High-quality de novo assembly of the apple genome and methylome dynamics of early fruit development Nature Genetics, 2017, 49(7):1099-1106. DOI:10.1038/ng.3886URL [本文引用: 1]
KUMARR, TYAGI AK, SHARMA AK. Genome-wide analysis of auxin response factor (ARF) gene family from tomato and analysis of their role in flower and fruit development Molecular Genetics and Genomics, 2011, 285:245-260. DOI:10.1007/s00438-011-0602-7URL [本文引用: 1]
WUJ, WANG FY, CHENGL, KONG FL, PENGZ, LIU SS, YU XL, LUG. Identification, isolation and expression analysis of auxin response factor (ARF) genes in Solanum lycopersicum Plant Cell Reports, 2011, 30(11):2059-2073. DOI:10.1007/s00299-011-1113-zURL [本文引用: 1]
DE JONGM, WOLTERS-ARTSM, FERONR, MARIANIC, VRIEZEN WH. The Solanum lycopersicum auxin response factor 7 (SlARF7) regulates auxin signaling during tomato fruit set and development Plant Journal, 2009, 57(1):160-170. DOI:10.1111/tpj.2008.57.issue-1URL [本文引用: 1]
SAGARM, CHERVINC, MILAI, HAOY, ROUSTANJ-P, BENICHOUM, GIBONY, BIAISB, MAURYP, LATCHEA, PECHJ-C, BOUZAYENM, ZOUINEM. SlARF4, an auxin response factor involved in the control of sugar metabolism during tomato fruit development Plant Physiology, 2013, 161(3):1362-1374. DOI:10.1104/pp.113.213843URL [本文引用: 1]
CAUSIERB, ASHWORTHM, GUOW, DAVIESB. The TOPLESS interactome: A framework for gene repression inArabidopsis Plant Physiology, 2012, 158(1):423-438. DOI:10.1104/pp.111.186999URL [本文引用: 1]
 ,, 卞书迅, 张恒涛, 张瑞萍, 高启明, 刘珍珍, 阎振立
,, 卞书迅, 张恒涛, 张瑞萍, 高启明, 刘珍珍, 阎振立

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT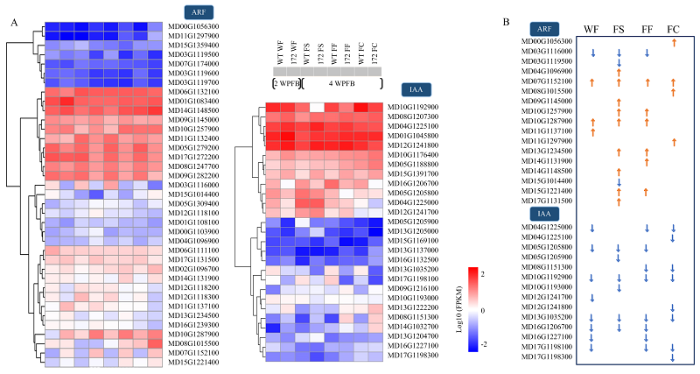 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT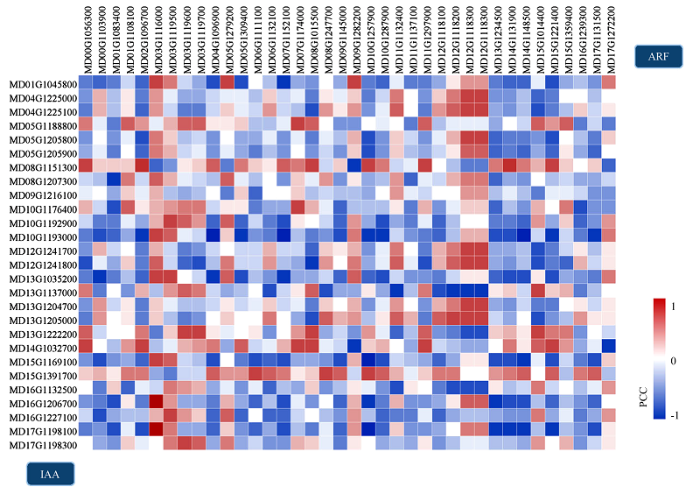 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT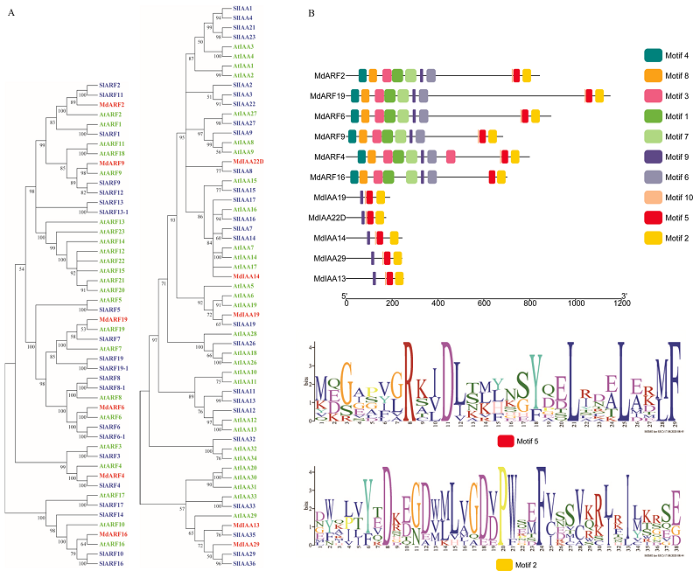 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT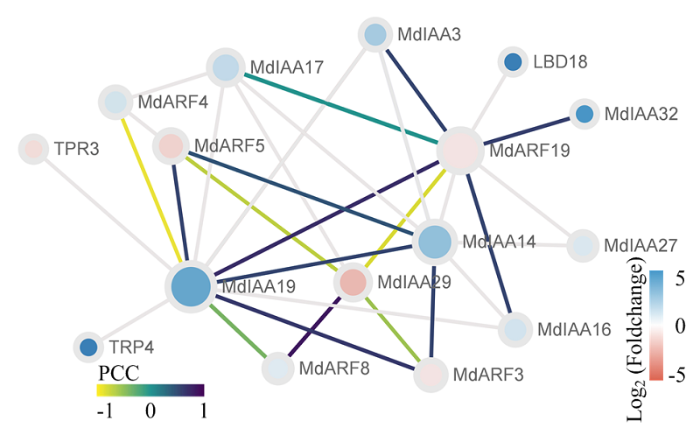 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}