Genome-Wide Identification and Expression Pattern Analysis of LRR-RLK Gene Family in Apple
HUANG JinFeng,1, LÜ TianXing1, WANG Xu2, WANG YingDa1, WANG DongMei1, YAN ZhongYe1, LIU Zhi,11Liaoning Institute of Pomology, Yingkou 115009, Liaoning 2College of Horticultural Science and Engineering, Shangdong Agricultural University, Tai’an 271018, Shangdong
Abstract 【Objective】 The study was carried out to explore the whole genome characteristics and expression patterns ofLRR-RLKs in apple, to reveal the expression specificity of family members in different tissues and their responses to biological and abiotic stress, and further understand its biological function in apple. 【Method】 The members of LRR-RLK gene family in the whole genome of apple were identified based on the local BLAST database and Pfam database. The LRR-RLK amino acid sequence prediction, subcellular localization prediction, domain analysis, phylogenetic tree and chromosome localization were completed by software of ExPASy Proteomics Server, Cell-PLoc, CD-Search Tool, MEGAX and MG2C. In addition, the expression pattern of 12 LRR-RLK genes in different tissues and stress were analyzed by real-time fluorescent quantitative PCR (qRT-PCR).【Result】 A total of 378 LRR-RLKgenes were identified from apple genome. TheseLRR-RLKgenes encoded proteins containing 318-1 827 amino acid, and the theoretical isoelectric point ranged from 6.14 to 9.01. The prediction subcellular localization of apple LRR-RLK proteins was all distributed in the cell membrane. The gene family could be divided into 15 subgroups, containing 1-111 genes. The 378 genes in this family were distributed on all 17 chromosomes of the apple, and the chromosome 7 contained 40 genes. The LRR-RLK gene family had two conserved domains, namely the leucine-rich repeat structure and the protein kinase domain. Irregular curl and α-helix was the main secondary structure in the LRR-RLK gene family, and the rotation of β-turn was very small. It was found that the 12 selected family members were expressed in all tissues (except MD00G1105400), and most genes were expressed at relatively high levels in stem. Seven genes were up-regulated under low temperature conditions, and the expression of MD09G1153800 was the most obvious. The expression of MD09G1153800 was raised to 6.8 times of that under the control. While MD06G1170200 and MD05G1061600 were both down-regulated. Eight genes were up-regulated under drought conditions, and MD00G1105400 was the most obvious one. The expression of MD00G1105400 was raised to 9.6 times of that under the control; under salt conditions, MD04G1150400, MD13G1108000 and MD02G1071800 were always up-regulated. Among them, MD02G1071800 had the highest expression after 24 hours of salt stress treatment, which was 14.9 times of that under the control. After inoculating Botryosphaeria dothidea, the expression of 12 LRR-RLKgenes increased first and then decreased. After inoculating, the expression level of Wangshanhong was higher on the first day, however the expression level of Jiguan was higher on the third day. The expression of MD09G1153800 and MD05G1065800 were up-regulated significantly in Jiguan, in relative to have no change in Wangshanhong, suggesting that these twoLRR-RLKscould serve as candidate genes for further functional characterization. 【Conclusion】 A total of 378 LRR-RLK members were identified from apple whole genome sequences, which could be divided into 15 groups and distributed on 17 chromosomes, and the most of LRR-RLK genes were responsive to stress stimulus and Botryosphaeria dothidea. Keywords:apple;LRR-RLK;gene family;identification;expression analysis
PDF (9601KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 黄金凤, 吕天星, 王寻, 王颖达, 王冬梅, 闫忠业, 刘志. 苹果LRR-RLK基因家族鉴定和表达分析[J]. 中国农业科学, 2021, 54(14): 3097-3112 doi:10.3864/j.issn.0578-1752.2021.14.015 HUANG JinFeng, LÜ TianXing, WANG Xu, WANG YingDa, WANG DongMei, YAN ZhongYe, LIU Zhi. Genome-Wide Identification and Expression Pattern Analysis of LRR-RLK Gene Family in Apple[J]. Scientia Acricultura Sinica, 2021, 54(14): 3097-3112 doi:10.3864/j.issn.0578-1752.2021.14.015
SHIU SH, BLEECKER AB. Receptor-like kinases from Arabidopsisform a monophyletic gene family related to animal receptor kinases Proceedings of the National Academy of Sciences of the United States of America, 2001, 98(19):10763-10768. [本文引用: 1]
XIANGY, CAO YL, XU CG, LI XH, WANG SP. Xa3, conferring resistance for rice bacterial blight and encoding a receptor kinase-like protein, is the same as Xa26 Theoretical and Applied Genetics, 2006, 113(7):1347-1355. DOI:10.1007/s00122-006-0388-xURL [本文引用: 1]
DE LORENZOL, MERCHANF, LAPORTEP, THOMPSONR, CLARKEJ, SOUSAC, CRESPIM. A novel plant leucine-rich repeat receptor kinase regulates the response ofMedicago truncatula roots to salt stress Plant Cell, 2009, 21(2):668-680. DOI:10.1105/tpc.108.059576URL [本文引用: 1]
LI XX, SALMANA, GUOC, YUJ, CAO SX, GAO XM, LIW, LIH, GUO YF. Identification and characterization of LRR-RLKfamily genes in potato reveal their involvement in peptide signaling of cell fate decisions and biotic/abiotic stress responses Cells, 2018, 7:120. DOI:10.3390/cells7090120URL [本文引用: 3]
TICHTINSKYG, VANNOOSTHUYSEV, COCK JM, GAUDET. Making inroads into plant receptor kinase signalling pathways Trends in Plant Science, 2003, 8(5):231-237. DOI:10.1016/S1360-1385(03)00062-1URL [本文引用: 1]
LEHTI-SHIU MD, SHIU SH. Diversity, classification and function of the plant protein kinase superfamily Philosophical Transactions of The Royal Society Biological Sciences, 2012, 367:2619-2639. DOI:10.1098/rstb.2012.0003URL [本文引用: 1]
SUN JM, LI LT, WANGP, ZHANG SL, WU JY. Genome-wide characterization, evolution, and expression analysis of the leucine-rich repeat receptor-like protein kinase (LRR-RLK) gene family in Rosaceae genomes BMC Genomics, 2017, 18:763. DOI:10.1186/s12864-017-4155-yURL [本文引用: 6]
GOMEZ-GOMEZL, BAUERZ, BOLLERT. Both the extracellular leucine-richrepeat domain and the kinase activity of FLS2are required for flagellin binding and signaling in Arabidopsis Plant Cell, 2001, 13(5):1155-1163. DOI:10.1105/tpc.13.5.1155URL [本文引用: 1]
KUNZEG, ZIPFELC, ROBATZEKS, NIEHAUSK, BOLLERT, FELIXG. The N terminus of bacterial elongation factor Tu elicits innate immunity in Arabidopsis plants Plant Cell, 2004, 16:3496-3507. DOI:10.1105/tpc.104.026765URL [本文引用: 1]
SONG WY, WANG GL, CHEN LL, KIM HS, PILY, HOLSTENT, GARDNERJ, WANGB, ZHAI WX, ZHU LH, FAUQUETC, RONALDP. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21 Science, 1995, 270(5243):1804-1806. DOI:10.1126/science.270.5243.1804URL [本文引用: 1]
HUH, WANGJ, SHIC, YUANC, PENG CF, YIN JJ, LI WT, HEM, WANG JC, MA BT, WANG YP, LI SG, CHEN XW. A receptor like kinase gene with expressional responsiveness on Xanthomonas oryzaepv.oryzae is essential for Xa21-mediated disease resistance Rice, 2015, 8: 1. https://doi.org/10.1186/s12284-014-0034-1. DOI:10.1186/s12284-014-0034-1URL [本文引用: 1]
CHEN TT, XIAOJ, XUJ, WANWT, QINB, CAO AZ, CHENW, XING LP, DUC, GAO XQ, ZHANG SZ, ZHANG RQ, SHEN WB, WANG HY, WANG XE. Two members of TaRLK family confer powdery mildew resistance in common wheat BMC Plant Biology, 2016, 16:27. DOI:10.1186/s12870-016-0713-8URL [本文引用: 1]
BLAUM BS, MAZZOTTAS, NOLDEKE ER, HALTERT, MADLUNGJ, KEMMERLINGB, STEHLET. Structure of the pseudokinase domain of BIR2, a regulator of BAK1-mediated immune signaling inArabidopsis Journal of Structural Biology, 2014, 186(1):112-121. DOI:10.1016/j.jsb.2014.02.005URL [本文引用: 1]
IMKAMPEJ, HALTERT, HUANG SH, SCHULZES, MAZZOTTAS, SCHMIDTN, MANSTRETTAR, POSTELS, WIERZBAM, YANGY, VAN DONGEN W M AM, STAHLM, ZIPFELC, GOSHE MB, CLOUSES, DE VRIES SC, TAXF, WANG XF, KEMMERLINGB. The Arabidopsis leucine-rich repeat receptor kinase BIR3negatively regulates BAK1 Receptor complex formation and stabilizes BAK1. Plant Cell, 2017, 29(9):2285-2303. DOI:10.1105/tpc.17.00376URL [本文引用: 1]
SUN RB, WANG SH, MAD, LIU CL. Genome-wide analysis of LRR-RLK gene family in four Gossypium species and expression analysis during cotton development and stress responses Genes, 2018, 9:592. DOI:10.3390/genes9120592URL [本文引用: 2]
LIH, HAN XD, LIU XX, ZHOU MY, RENW, ZHAO BB, JU CL, LIUY, ZHAO JR. A leucine-rich repeat-receptor-like kinase gene SbER2-1 from sorghum (Sorghum bicolor L.) confers drought tolerance in maize BMC Genomics, 2019, 20:737. DOI:10.1186/s12864-019-6143-xURL [本文引用: 1]
LIN FM, LIS, WANGK, TIANH, GAO JF, ZHAO QZ, DU CQ. A leucine-rich repeat receptor-like kinase, OsSTLK, modulates salt tolerance in rice Plant Science, 2020, 296:110465. DOI:10.1016/j.plantsci.2020.110465URL [本文引用: 1]
LIR, AN JP, YOU CX, WANG XF, HAO YJ. Molecular cloning and functional characterization of the CEP RECEPTOR1 gene MdCEPR1 of apple (Malus × domestica) Plant Cell, Tissue and Organ Culture, 2020, 140(3):539-550. DOI:10.1007/s11240-019-01745-wURL [本文引用: 1]
LIAO YL, HU CQ, ZHANG XW, CAO XF, XU ZJ, GAO XL, LI LH, ZHU JQ, CHEN RJ. Isolation of a novel leucine-rich repeat receptorlike kinase (OsLRR2) gene from rice and analysis of its relation to abiotic stress responses Biotechnology & Biotechnological Equipment, 2017, 31(1):51-57. [本文引用: 1]
WANG JH, KUCUKOGLUM, ZHANG LB, CHENP, DANIELD, NILSSONO, JONESB, SANDBERGG, ZHENGB. The ArabidopsisLRR-RLK, PXC1, is a regulator of secondary wall formation correlated with the TDIF-PXY/TDR-WOX4 signaling pathway BMC Plant Biology, 2013, 13:94. DOI:10.1186/1471-2229-13-94URL [本文引用: 1]
YANGM, HAN YZ, A YJ, HA BK, WANG CL. Study of RLK6, one member of leucine-rich repeat receptor-like kinases(LRR-RLKs) subfamily gene, on process of flowering in Arabidopsis Journal of Nuclear Agricultural Sciences, 2017, 31(4):654-662. (in Chinese) [本文引用: 1]
YU YX, SONG JL, TIAN XH, ZHANG HW, LI LG, ZHU HF. Arabidopsis PRK6 interacts specifically with AtRopGEF8/12 and induces depolarized growth of pollen tubes when over expressed Science China-Life Sciences, 2018, 61:100-112. DOI:10.1007/s11427-016-9107-3URL [本文引用: 1]
DIMITROVAI, TAX FE. Lateral root growth in Arabidopsisis controlled by short and long distance signaling through the LRR RLKs XIP1/CEPR1 and CEPR2 Plant Signaling and Behavior, 2018, 13(6):e1489667. DOI:10.1080/15592324.2018.1489667URL [本文引用: 1]
ZOUY, LIU XY, WANGQ, CHENY, LIUC, QIUY, ZHANGW. OsRPK1, a novel leucine-rich repeat receptor-like kinase, negatively regulates polar auxin transport and root development in rice Biochimica et Biophysica Acta (BBA)-General Subjects, 2014, 1840(6):1676-1685. DOI:10.1016/j.bbagen.2014.01.003URL [本文引用: 1]
VELASCOR, ZHARKIKHA, AFFOURTITJ, DHINGRAA, CESTAROA, KALYANARAMANA, FONTANAP, BHATNAGAR SK, TROGGIOM, PRUSSD, SALVIS, PINDOM, BALDIP, CASTELLETTIS, CAVAIUOLOM, COPPOLAG, COSTAF, COVAV, RI AD, GOREMYKINV, et al. The genome of the domesticated apple (Malus×domestica Borkh.) Nature Genetics, 2010, 42(10):833-839. DOI:10.1038/ng.654URL [本文引用: 1]
DACCORDN, CELTON JM, LINSMITHG, BECKERC, CHOISNEN, SCHIJLENE, VAN DE GEESTH, BIANCOL, MICHELETTID, WELASCOR, DI PIERRO EA, GOUZYJ, REES D JG, GUERIFP, MURANTYH, DUREL CE, LAURENSF, LESPINASSEY, GAILLARDS, AUBOURGS, QUESNEVILLEH, WEIGELD, VAN DE WEGE, TROGGIOM, BUCHERE. High-quality de novo assembly of the apple genome and methylome dynamics of early fruit development Nature Genetics, 2017, 49:1099-1106. DOI:10.1038/ng.3886URL [本文引用: 1]
YUAN GP, HAN XL, BIAN SX, ZHANG LY, TIANY, ZHANG CX, CONG PH. Bioinformatics and expression analysis of theLIMgene family in apple Scientia Agricultura Sinica, 2019, 52(23):4322-4332. (in Chinese) [本文引用: 1]
WAI CM, POWELLB, MINGR, MIN XJ. Genome-wide identification and analysis of genes encoding proteolytic enzymes in pineapple Tropical Plant Biology, 2016, 9:161-175. DOI:10.1007/s12042-016-9172-5URL [本文引用: 1]
ZHANGY, HUANG TH, ZHANG XL, LIU TK, HOU XL, LIY. Cloning and expression analysis of BcSERK1 from non-heading Chinese cabbage Journal of Nanjing Agricultural University, 2019, 42(6):1014-1021. (in Chinese) [本文引用: 1]
ZHOU Fl, GUOY, QIU LJ. Genome-wide identification and evolutionary analysis of leucine-rich repeat receptor-like protein kinase genes in soybean BMC Plant Biology, 2016, 16:58. DOI:10.1186/s12870-016-0744-1URL [本文引用: 2]
YUANN, RAI KM, BALASUBRAMANIAN VK, UPADHYAY SK, LUOH, MENDUV. Genome-wide identification and characterization of LRR-RLKs reveal functional conservation of the SIFsubfamily in cotton (Gossypium hirsutum) BMC Plant Biology, 2018, 18:185. DOI:10.1186/s12870-018-1395-1URL [本文引用: 1]
MAGALHAES DM, SCHOLTE L LS, SILVA NV, OLIVEIRA GC, ZIPFELC, TAKITA MA, DE SOUZA AA. LRR-RLK family from two Citrus species: Genome-wide identification and evolutionary aspects BMC Genomics, 2016, 17:623. DOI:10.1186/s12864-016-2930-9URL [本文引用: 1]
WEI ZR, WANG JH, YANG SH, SONG YJ. Identification and expression analysis of the LRR-RLK gene family in tomato (Solanum lycopersicum) Heinz 1706 Genome, 2015, 58(4):121-134. DOI:10.1139/gen-2015-0035URL [本文引用: 1]
LIU PL, DUL, HUANGY, GAO SM, YUM. Origin and diversification of leucine-rich repeat receptor-like protein kinase (LRR-RLK) genes in plants BMC Evolutionary Biology, 2017, 17:47. DOI:10.1186/s12862-017-0891-5URL [本文引用: 1]
BETTEMBOURGM, DAL-SOGLIOM, BUREAUC, VERNETA, DARDOUXA, PORTEFAIXM, BESM, MEYNARDD, MIEULETD, CAYROLB, PERINC, COURTOISB, MA JF, DIEVARTA. Root cone angle is enlarged in docs1 LRR-RLK mutants in rice Rice, 2017, 10:50. DOI:10.1186/s12284-017-0190-1URL [本文引用: 1]
HOU BZ, CHENGX, SHEN YY. A leu-rich repeat receptor-like protein kinase, FaRIPK1, interacts with the ABA receptor, FaABAR, to regulate fruit ripening in strawberry Journal of Experimental Botany, 2018, 69(7):1569-1582. DOI:10.1093/jxb/erx488URL [本文引用: 1]
PARK SJ, MOON JC, PARK YC, KIMJ, KIM DS, JANG CS. Molecular dissection of the response of a rice leucine-rich repeat receptor-like kinase (LRR-RLK) gene to abiotic stresses Journal of Plant Physiology, 2014, 171(17):1645-1653. DOI:10.1016/j.jplph.2014.08.002URL [本文引用: 1]
XIE RJ, LI YJ, HE SL, ZHENG YQ, YI SL, LVQ, DENGL. Genome-wide analysis of citrusR2R3MYBgenes and their spatiotemporal expression under stresses and hormone treatments PLoS ONE, 2014, 9(12):e113971. DOI:10.1371/journal.pone.0113971URL [本文引用: 1]
LIUH, DUOH, ZHAOD, SUNE, MA FP, MA CL, ZUO CW. Identification ofCRKgene family in pear and its members in response to signals of Valsa pyri Acta Horticulturae Sinica, 2020, 47(4):963-973. (in Chinese) [本文引用: 1]
LINH, LENGH, GUO YS, KONDOS, ZHAO YH, SHI GL, GUO XW. QTLs and candidate genes for downy mildew resistance conferred by interspecific grape (V. vinifera L. × V. amurensis Rupr.) crossing Scientia Horticulturae, 2019, 244:200-207. DOI:10.1016/j.scienta.2018.09.045URL [本文引用: 1]
FAIZEM, FAIZEM, FAIZEL, ISHIIH. Characterization of a leucine-rich repeat receptor-like protein kinase (LRPK) gene from Japanese pear and its possible involvement in scab resistance Journal of General Plant Pahtology, 2007, 73(2):104-112. [本文引用: 1]
KOMJANCM, FESTIS, RIZZOTTIL, CATTIVELLIL, CERVONEF, DE LORENZOG. A leucine-rich repeat receptor-like protein kinase(LRPKm1) gene is induced in Malus × domestica by Venturia inaequalis infection and salicylic acid treatment Plant Molecular Biology, 1999, 40:945-957. DOI:10.1023/A:1006275924882URL [本文引用: 1]
PADMARASUS, SARGENT DJ, PATOCCHIA, TROGGOIM, BALDIP, LINSMITHG, POLESL, JANSCHM, KELLERHALSM, TATTARINIS, VELASCOR. Identification of a leucine-rich repeat receptor-like serine/threonine-protein kinase as a candidate gene for Rvi12(Vb)-based apple scab resistance Molecular Breeding, 2018, 38:73. DOI:10.1007/s11032-018-0825-yURL [本文引用: 1]
COVAV, PARISR, PASSEROTTIS, ZINIE, GESSLERC, PERTOTI, LOIN, MUSETTIR, KOMJANCM. Mapping and functional analysis of four apple receptor-like protein kinases related to LRPKm1 in HcrVf2-transgenic and wild-type apple plants Tree Genetics & Genomes, 2010, 6:389-403. [本文引用: 1]
 ,1, 吕天星1, 王寻2, 王颖达1, 王冬梅1, 闫忠业1, 刘志
,1, 吕天星1, 王寻2, 王颖达1, 王冬梅1, 闫忠业1, 刘志

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT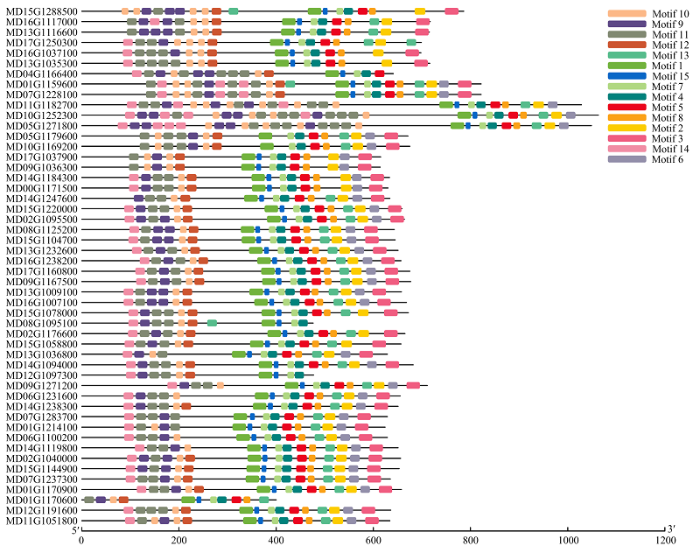 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT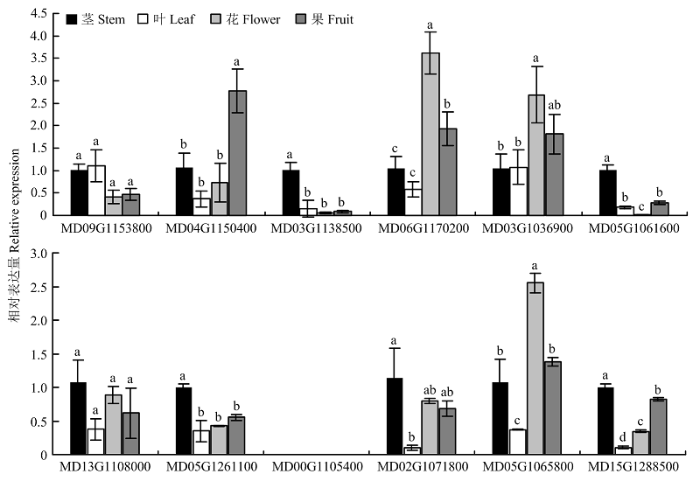 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT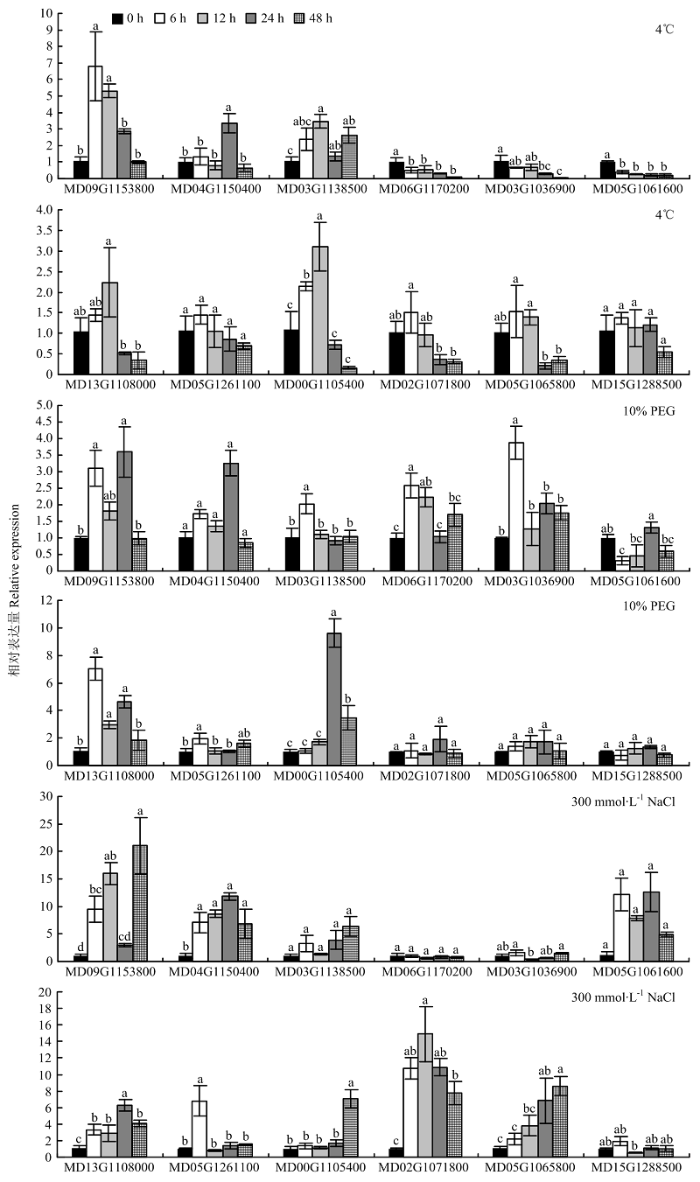 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT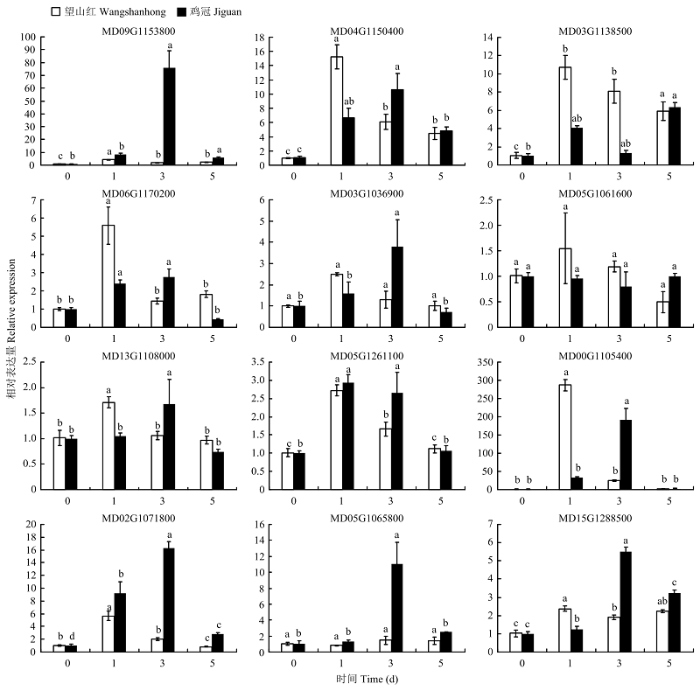 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}