Advances of Biosynthesis and Toxicity of Cereulide Produced by Emetic Bacillus cereus
CUI YiFang,1, ZHENG Min2, DING ShuangYang2, ZHU Kui,21Beijing Key Laboratory for Prevention and Control of Infectious Diseases in Livestock and Poultry/Institute of Animal Husbandry and Veterinary Medicine, Beijing Academy of Agricultural and Forestry Sciences, Beijing 100097 2National Center for Veterinary Drug Evaluation/College of Veterinary Medicine, China Agricultural University, Beijing 100193
Abstract Bacillus cereus (B. cereus) is a gram-positive facultative anaerobe that can produce spores to survive adverse environments. And it is widely present in soil, water, air and a variety of foods. Pathogenic B. cereus is one of the most common food-borne pathogens, and the toxins produced by B. cereus are the main cause of food poisoning. Cereulide is a major toxin produced by pathogenic B. cereus, which is a small molecule lipophilic cyclic dodecadepsipeptide with stable structural properties. Cereulide can cause mild food poisoning with emetic symptoms, such as nausea and vomiting, and it may induce severe fatal diseases such as hepatic encephalopathy or acute liver failure. Current researches believed that cereulide caused vomiting by stimulating the vagus nerve, and induced the loss of mitochondrial membrane potential by acting as a potassium ionophore, which ultimately led to cell death. However, the toxic mechanism of hepatic encephalopathy or acute liver failure caused by cereulide remains unclear. Cereulide is encoded by the cereulide synthetase gene cluster (ces) and is synthesized by the non-ribosomal peptide synthetase (NRPS) system. Cereulide is composed by two hydroxy acids and two amino acid residues [-D-HIC-D-Ala-L-HIV-L-Val-], which forms a trimer phenolphthalein after three iterations and shows a structural specificity and representativeness. However, isocereulides may be produced due to the flexibility of the NRPS system. Therefore, the toxicity of cereulide is closely related to its biosynthesis process. Based on previous studies, this review summarized and proposed the biosynthesis mechanism of cereulide. Firstly, the CesA and CesB domained in ces recognize D-α-ketocarboxylic acid, L-alanine, L-α-ketoisovalerate and L-valine, respectively, which formed the main synthetic unit dipeptide of cereulide by covalent bonding. Secondly, a tetrapeptide was synthesized by repeating the above process. Thirdly, the second tetrapeptide was synthesized through repeated reactions, and the two tetrapeptides formed an octapeptide through esterification. Fourthly, the above reaction was repeated to form a ternary complex product peptide. Lastly, because the surface structure of the active center of the thioesterase domain in ces-NRPS prevented external water molecules from entering, it induced an internal nucleophilic attack reaction and finally released a circular cereulide. The risk of food poisoning caused by cereulide producing B. cereus was underestimated. In addition, our previous studies have found that some probiotic Bacillus products were contaminated with cereulide-producing B. cereus strains. This posed a potential risk to food safety and public health. This review briefly summarized the characteristics and toxic mechanisms of cereulide, which would provide a scientific basis for the prevention of cereulide. This review also summarized and proposed the biosynthesis process of cereulide. Functions of two domains in the synthesis process need to be focused. The main function of ketoreductase (KR) domain was that it could catalyze the formation of esters of keto acid at the beginning of the biosynthesis processes. The important role of thioesterase (TE) domain was to form repeating units and the cyclic peptide in the last link of synthesis. These could serve as a model for other cyclic peptides synthesized by the non-ribosomal peptide synthetase system. Keywords:Bacillus cereus;cereulide;biosynthesis;non-ribosomal peptide synthetase;toxicity
PDF (2807KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 崔一芳, 郑敏, 丁双阳, 朱奎. 蜡样芽孢杆菌致吐毒素的毒性作用与生物合成研究进展[J]. 中国农业科学, 2021, 54(12): 2666-2674 doi:10.3864/j.issn.0578-1752.2021.12.016 CUI YiFang, ZHENG Min, DING ShuangYang, ZHU Kui. Advances of Biosynthesis and Toxicity of Cereulide Produced by Emetic Bacillus cereus[J]. Scientia Acricultura Sinica, 2021, 54(12): 2666-2674 doi:10.3864/j.issn.0578-1752.2021.12.016
值得注意的是,部分蜡样芽孢杆菌作为益生菌被广泛应用于人类医疗保健、畜牧业、农业和水产环境等多个领域[13]。例如,美国食品药品监督管理局(Food and Drug Administration, FDA)和饲料工业协会(American Feed Industry Association, AFIA)在1989年就批准了蜡样芽孢杆菌可作为直接应用于畜牧生产的菌种之一[14]。我国农业农村部1996年也正式批准蜡样芽孢杆菌可作为饲料级微生物添加剂的菌种之一[15]。尽管目前益生菌的总体使用记录显示出较令人满意的安全性结果,但相对于其应用的广泛度来说,益生菌的安全性并未受到充分鉴定[16]。例如,本团队前期对益生芽孢杆菌的毒性和耐药性进行了系统评估,发现菌株可产生多种毒素,其中便包括cereulide[17,18]。此外,详细综述了蜡样芽孢杆菌可能产生的多种毒素,包括毒素特征、毒性作用、合成机制及检测方法,评估其对蜡样芽孢杆菌作为益生菌使用的潜在危害[19]。
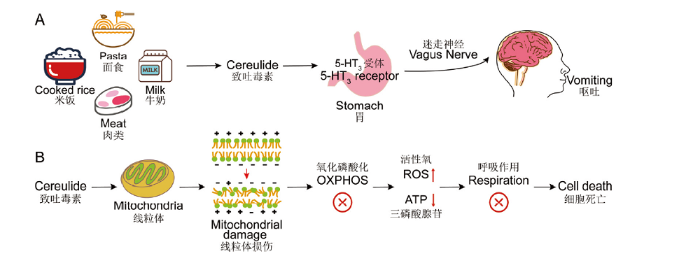
A:Cereulide存在于多种食物中,通过与胃肠道内分布的5-HT3受体结合,刺激迷走神经进而引发呕吐症状;B:Cereulide作为钾离子载体,会破坏细胞脂质膜上的电化学电位梯度,通过影响呼吸作用,最终导致细胞死亡 Fig. 1Toxicity and mechanism of cereulide
A: Cereulide is present in a variety of foods. It induces vomiting by binding to 5-HT3 receptors in the gastrointestinal tract and stimulating the vagus nerve; B: Cereulide, as a potassium ionophore, disrupts the electrochemical potential gradient on lipid membranes which affects respiration, and ultimately leads to cell death
NRPS系统的A型生物合成途径可合成线性肽,例如枯草芽孢杆菌(B. subtilis)中的表面活性素(surfactin)。B型途径合成迭代肽,典型代表为蜡样芽孢杆菌(B. cereus)的致吐毒素(cereulide)。C型途径控制合成非线性肽,例如霍乱弧菌(Vibrio cholerae)的弧菌杆菌素(vibriobactin) Fig. 2Three biosynthesis pathways of NRPSs
Type A biosynthesis pathway of NRPS system synthesizes linear peptides, such as surfactin in B. subtilis. Type B pathway synthesizes iterative peptides like cereulide in B. cereus. Type C pathway encodes nonlinear peptides, such as vibriobactin in Vibrio cholerae
CEUPPENSS, BOONN, UYTTENDAELEM. Diversity of Bacillus cereus group strains is reflected in their broad range of pathogenicity and diverse ecological lifestyles FEMS Microbiology Ecology, 2013,84(3):433-450. DOI:10.1111/femsec.2013.84.issue-3URL [本文引用: 1]
MAHLERH, PASIA, KRAMER JM, SCHULTEP, SCOGING AC, B?RW, KRAHENBUHLS. Fulminant liver failure in association with the emetic toxin of Bacillus cereus. The New England Journal of Medicine, 1997,336(16):1142-1148. [本文引用: 1]
RAJKOVICA. Microbial toxins and low level of foodborne exposure Trends in Food Science & Technology, 2014,38(2):149-157. [本文引用: 1]
BENNETT SD, WALSH KA, GOULD LH. Foodborne disease outbreaks caused by Bacillus cereus, Clostridium perfringens, and Staphylococcus aureus-United States, 1998-2008 Clinical Infectious Diseases, 2013,57(3):425-433. DOI:10.1093/cid/cit244URL [本文引用: 1]
GLASSETB, HERBINS, GUILLIERL, CADEL-SIXS, VIGNAUD ML, GROUTJ, PAIRAUDS, MICHELV, HENNEKINNE JA, RAMARAON, BRISABOISA. Bacillus cereus-induced food-borne outbreaks in France, 2007 to 2014: epidemiology and genetic characterisation EURO Surveillance, 2016,21(48):30413. [本文引用: 1]
MESSELH?USSERU, FRENZELE, BL?CHINGERC, ZUCKERR, K?MPFP, EHLING-SCHULZM. Emetic Bacillus cereus are more volatile than thought: recent foodborne outbreaks and prevalence studies in Bavaria (2007-2013) BioMed Research International, 2014,2014:465603. [本文引用: 2]
TER BEEKA, BRULS. To kill or not to kill Bacilli: opportunities for food biotechnology Current Opinion in Biotechnology, 2010,21(2):168-174. DOI:10.1016/j.copbio.2010.03.014URL [本文引用: 1]
GáLVEZA, ABRIOUELH, LóPEZ RL, BEN OMARN. Bacteriocin-based strategies for food biopreservation International Journal of Food Microbiology, 2007,120(1-2):51-70. DOI:10.1016/j.ijfoodmicro.2007.06.001URL [本文引用: 1]
IN 'T VELDP H, VAN DER LAAKL F J, VAN ZONM, BIESTA-PETERS EG. Elaboration and validation of the method for the quantification of the emetic toxin of Bacillus cereus as described in EN-ISO 18465-Microbiology of the food chain - Quantitative determination of emetic toxin (cereulide) using LC-MS/MS International Journal of Food Microbiology, 2019,288:91-96. DOI:10.1016/j.ijfoodmicro.2018.03.021URL [本文引用: 1]
ELSHAGHABEE F MF, ROKANAN, GULHANE RD, SHARMAC, PANWARH. Bacillus as potential probiotics: status, concerns, and future perspectives Frontiers in Microbiology, 2017,8:1490. DOI:10.3389/fmicb.2017.01490URL [本文引用: 1]
FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS (FAO). Probiotics in animal nutrition production, impact and regulation FAO Animal Production and Health Paper 2016, No. 179, http://www.fao.org/3/a-i5933e.pdf. URL [本文引用: 1]
MINISTRY OF AGRICULTURE AND RURAL AFFIRS OF THE PEOPLE’S REPUBLIC OF CHINA. Ministry of Agticulture Announcement No. 2045 (Category of Feed Additives 2013), 2013. (in Chinese) [本文引用: 1]
DORONS, SNYDMAN DR. Risk and safety of probiotics Clinical Infectious Diseases, 2015,60(Suppl. 2):S129-S134. DOI:10.1093/cid/civ085URL [本文引用: 1]
ZHUK, HOLZEL CS, CUI YF, MAYERR, WANGY, DIETRICHR, DIDIERA, BASSITTAR, MARTLBAUERE, DING SY. Probiotic Bacillus cereus strains, a potential risk for public health in China Frontiers in Microbiology, 2016,7:718. [本文引用: 1]
CUI YF, WANG SL, DING SY, SHEN JZ, ZHUK. Toxins and mobile antimicrobial resistance genes in Bacillus probiotics constitute a potential risk for One Health Journal of Hazardous Materials, 2020,382:121266. DOI:10.1016/j.jhazmat.2019.121266URL [本文引用: 1]
CUI YF, M?RTLBAUERE, DIETRICHR, LUO HL, DINGS Y, ZHUK. Multifaceted toxin profile, an approach toward a better understanding of probiotic Bacillus cereus. Critical Reviews in Toxicology, 2019,49(4):342-356. [本文引用: 2]
MAGARVEY NA, EHLING-SCHULZM, WALSH CT. Characterization of the cereulide NRPS alpha-hydroxy acid specifying modules: activation of alpha-keto acids and chiral reduction on the assembly line Journal of American Chemical Society, 2006,128(33):10698-10699. DOI:10.1021/ja0640187URL [本文引用: 3]
MARXENS, STARK TD, RUTSCHLEA, LüCKINGG, FRENZELE, SCHERERS, EHLING-SCHULZM, HOFMANNT. Depsipeptide intermediates interrogate proposed biosynthesis of cereulide, the emetic toxin of Bacillus cereus Scientific Reports, 2015,5:10637. DOI:10.1038/srep10637URL [本文引用: 2]
TEPLOVA VV, MIKKOLAR, TONSHIN AA, SARIS NE, SALKINOJA-SALONENM S. The higher toxicity of cereulide relative to valinomycin is due to its higher affinity for potassium at physiological plasma concentration Toxicology and Applied Pharmacology, 2006,210(1-2):39-46. DOI:10.1016/j.taap.2005.06.012URL [本文引用: 1]
GRANUMP E. Spotlight on Bacillus cereus and its food poisoning toxins FEMS Microbiology Letters, 2017,364(10):fnx071. [本文引用: 1]
STENFORS ARNESEN LP, FAGERLUNDA, GRANUM PE. From soil to gut: Bacillus cereus and its food poisoning toxins FEMS Microbiology Reviews, 2008,32(4):579-606. DOI:10.1111/j.1574-6976.2008.00112.xURL [本文引用: 1]
KRANZLERM, STOLLEWERKK, ROUZEAU-SZYNALSKIK, BLAYOL, SULYOKM, EHLING-SCHULZM. Temperature exerts control of Bacillus cereus emetic toxin production on post-transcriptional levels Frontiers in Microbiology, 2016,7:1640. [本文引用: 1]
RAJKOVICA, UYTTENDAELEM, VERMEULENA, ANDJELKOVICM, FITZ-JAMESI, IN 'T VELDP, DENONQ, VéRHER, DEBEVEREJ. Heat resistance of Bacillus cereus emetic toxin, cereulide Letters in Applied Microbiology, 2008,46(5):536-541. DOI:10.1111/j.1472-765X.2008.02350.xURL [本文引用: 1]
CARROLL LM, WIEDMANNM. Cereulide synthetase acquisition and loss events within the evolutionary history of Group III Bacillus cereus sensu lato facilitate the transition between emetic and diarrheal foodborne pathogen BioRxiv, 2020, doi: https://doi.org/10.1101/2020.05.12.090951. URL [本文引用: 1]
CARROLL LM, WIEDMANNM, MUKHERJEEM, NICHOLAS DC, MINGLE LA, DUMAS NB, COLE JA, KOVACJ. Characterization of emetic and diarrheal Bacillus cereus strains from a 2016 foodborne outbreak using whole-genome sequencing: addressing the microbiological, epidemiological, and bioinformatic challenges Frontiers in Microbiology, 2019,10:144. DOI:10.3389/fmicb.2019.00144URL [本文引用: 2]
RAMARAON, TRAN SL, MARINM, VIDICJ. Advanced methods for detection of Bacillus cereus and its pathogenic factors Sensors, 2020,20(9):2667. DOI:10.3390/s20092667URL [本文引用: 1]
AGATAN, OHTA M. MORIM, ISOBEM. A novel dodecadepsipeptide, cereulide, is an emetic toxin of Bacillus cereus FEMS Microbiology Letters, 1995,129(1):17-19. [本文引用: 1]
SHINAGAWAK, KONUMAH, SEKITAH, SUGIIS. Emesis of rhesus monkeys induced by intragastric administration with the HEp-2 vacuolation factor (cereulide) produced by Bacillus cereus FEMS Microbiology Letters, 1995,130(1):87-90. [本文引用: 1]
DELBRASSINNEL, ANDJELKOVICM, DIERICKK, DENAYERS, MAHILLONJ, VAN LOCOJ. Prevalence and levels of Bacillus cereus emetic toxin in rice dishes randomly collected from restaurants and comparison with the levels measured in a recent foodborne outbreak Foodborne Pathogens and Disease, 2012,9(9):809-814. DOI:10.1089/fpd.2012.1168URL [本文引用: 1]
R?NNING HT, ASP TN, GRANUM PE. Determination and quantification of the emetic toxin cereulide from Bacillus cereus in pasta, rice and cream with liquid chromatography-tandem mass spectrometry Food Additives and Contaminants Part a-Chemistry Analysis Control Exposure & Risk Assessment, 2015,32(6):911-921. [本文引用: 1]
VANGOITSENHOVENR, RONDASD, CRèVECOEURI, D'HERTOGW, BAATSENP, MASINIM, ANDJELKOVIM, VANL OCO J, MATTHYSC, MATHIEUC, OVERBERGHL, VAN DERSCHUEREN B. Foodborne cereulide causes beta-cell dysfunction and apoptosis PLoS ONE, 2014,9(8):e104866. DOI:10.1371/journal.pone.0104866URL [本文引用: 2]
PAANANENA, MIKKOLAR, SARENEVAT, MATIKAINENS, HESSM, ANDERSSONM, JULKUNENI, SALKINOJA-SALONENM S, TIMONENT. Inhibition of human natural killer cell activity by cereulide, an emetic toxin from Bacillus cereus Clinical and Experimental Immunology, 2002,129(3):420-428. DOI:10.1046/j.1365-2249.2002.01898.xURL [本文引用: 2]
DECLEERM, JOVANOVICJ, VAKULAA, UDOVICKIB, AGOUAR E K, MADDERA, DE SAEGERS, RAJKOVICA. Oxygen consumption rate analysis of mitochondrial dysfunction caused by Bacillus cereus cereulide in Caco-2 and HepG2 cells Toxins, 2018,10(7):266. DOI:10.3390/toxins10070266URL [本文引用: 2]
J??SKEL?INEN EL, TEPLOVAV, ANDERSSON MA, ANDERSSON LC, TAMMELAP, ANDERSSON MC, PIRHONEN TI, SARIS NE, VUORELAP, SALKINOJA-SALONEN MS. In vitro assay for human toxicity of cereulide, the emetic mitochondrial toxin produced by food poisoning Bacillus cereus Toxicology in Vitro, 2003,17(5-6):737-744. DOI:10.1016/S0887-2333(03)00096-1URL [本文引用: 1]
VANGOITSENHOVENR, MASINIM, OVERBERGHL, VAN LOCOJ, MATHIEUC, VAN DERSCHUEREN B. Cereulide food toxin, beta cell function and diabetes: facts and hypotheses Diabetes Research and Clinical Practice, 2015,109(1):1-5. DOI:10.1016/j.diabres.2015.04.029URL [本文引用: 1]
CHOIW, NAMKUNGJ, HWANGI, KIMH, LIMA, PARK HJ, LEE HW, HAN KH, PARKS, JEONG JS, BANGG, KIM YH, YADAV VK, KARSENTYG, JU YS, CHOIC, SUH JM, PARK JY, PARKS, KIMH. Serotonin signals through a gut-liver axis to regulate hepatic steatosis Nature Communications, 2018,9(1):4824. DOI:10.1038/s41467-018-07287-7URL [本文引用: 1]
CUI YF, LIUY, LIU XY, XIAX, DING SY, ZHUK. Evaluation of the toxicity and toxicokinetics of cereulide from an emetic Bacillus cereus strain of milk origin Toxins, 2016,8(6):156. DOI:10.3390/toxins8060156URL [本文引用: 1]
BOZHüYüK KA, MICKLEFIELDJ, WILKINSONB. Engineering enzymatic assembly lines to produce new antibiotics Current Opinion in Microbiology, 2019,51:88-96. DOI:10.1016/j.mib.2019.10.007URL [本文引用: 1]
BROWN AS, CALCOTT MJ, OWEN JG, ACKERLEY DF. Structural, functional and evolutionary perspectives on effective re-engineering of non-ribosomal peptide synthetase assembly lines Natural Product Reports, 2018,35(11):1210-1228. DOI:10.1039/C8NP00036KURL [本文引用: 1]
EHLING-SCHULZM, VUKOVN, SCHULZA, SHAHEENR, ANDERSSONM, M?RTLBAUERE, SCHERERS. Identification and partial characterization of the nonribosomal peptide synthetase gene responsible for cereulide production in emetic Bacillus cereus Applied and Environmental Microbiology, 2005,71(1):105-113. DOI:10.1128/AEM.71.1.105-113.2005URL [本文引用: 1]
EHLING-SCHULZM, FRICKERM, GRALLERTH, RIECKP, WAGNERM, SCHERERS. Cereulide synthetase gene cluster from emetic Bacillus cereus: structure and location on a mega virulence plasmid related to Bacillus anthracis toxin plasmid pXO1 BMC Microbiology, 2006,6:20. DOI:10.1186/1471-2180-6-20URL [本文引用: 1]
DOMMEL MK, LüCKINGG, SCHERERS, EHLING-SCHULZM. Transcriptional kinetic analyses of cereulide synthetase genes with respect to growth, sporulation and emetic toxin production in Bacillus cereus Food Microbiology, 2011,28(2):284-290. DOI:10.1016/j.fm.2010.07.001URL [本文引用: 1]
LüCKINGG, FRENZELE, RüTSCHLEA, MARXENS, STARK TD, HOFMANNT, SCHERERS, EHLING-SCHULZM. Ces locus embedded proteins control the non-ribosomal synthesis of the cereulide toxin in emetic Bacillus cereus on multiple levels Frontiers in Microbiology, 2015,6:1101. [本文引用: 1]
EHLING-SCHULZM, FRENZELE, GOHARM. Food-bacteria interplay: pathometabolism of emetic Bacillus cereus Frontiers in Microbiology, 2015,6:704. [本文引用: 1]
LüCKINGG, DOMMEL MK, SCHERERS, FOUETA, EHLING-SCHULZM. Cereulide synthesis in emetic Bacillus cereus is controlled by the transition state regulator AbrB, but not by the virulence regulator PlcR Microbiology, 2009,155(Pt 3):922-931. DOI:10.1099/mic.0.024125-0URL [本文引用: 1]
GUéRINA, R?NNING HT, DARGAIGNARATZC, CLAVELT, BROUSSOLLEV, MAHILLONJ, GRANUM PE, NGUYEN-THEC. Cereulide production by Bacillus weihenstrphanensis strains during growth at different pH values and temperatures Food Microbiology, 2017,65:130-135. DOI:10.1016/j.fm.2017.02.006URL [本文引用: 1]
GENG PL, HU YM, ZHOU GP, YUAN ZM, HU XM. Characterization of three autolysins with activity against cereulide- producing Bacillus isolates in food matrices International Journal of Food Microbiology, 2017,241:291-297. DOI:10.1016/j.ijfoodmicro.2016.10.030URL [本文引用: 1]
AONZO DA, MAGARVEY NA, SCHMEING TM. Characterization of cereulide synthetase, a toxin-producing macromolecular machine PLoS ONE, 2015,10(6):e0128569. DOI:10.1371/journal.pone.0128569URL [本文引用: 2]
KUO MM, HAYNES WJ, LOUKIN SH, KUNGC, SAIMIY. Prokaryotic K+ channels: from crystal structures to diversity FEMS Microbiology Reviews, 2005,29(5):961-985. DOI:10.1016/j.femsre.2005.03.003URL [本文引用: 1]
SANDYM, RUIZ, GALLAGHERJ, ZHANGW. Enzymatic synthesis of dilactone scaffold of antimycins ACS Chemical Biology, 2012,7(12):1956-1961. DOI:10.1021/cb300416wURL [本文引用: 1]
SüSSMUTHR, MüLLERJ, VON D?HRENH, MOLNRáRI. Fungal cyclooligomer depsipeptides: from classical biochemistry to combinatorial biosynthesis Natural Product Reports, 2011,28(1):99-124. DOI:10.1039/C001463JURL [本文引用: 1]
GAOL, LIUH, MAZ, HANJ, LUZ, DAIC, LVF, BIEX. Translocation of the thioesterase domain for the redesign of plipastatin synthetase Scientific Reports, 2016. 6:38467. DOI:10.1038/srep38467URL [本文引用: 1]
SIEBER SA, MARAHIEL MA. Molecular mechanisms underlying nonribosomal peptide synthesis: approaches to new antibiotics Chemical Reviews, 2005,105(2):715-738. DOI:10.1021/cr0301191URL [本文引用: 2]
BRUNER SD, WEBERT, KOHLI RM, SCHWARZERD, MARAHIEL MA, WALSH CT, STUBBS MT. Structural basis for the cyclization of the lipopeptide antibiotic surfactin by the thioesterase domain SrfTE Structure, 2002,10(3):301-310. DOI:10.1016/S0969-2126(02)00716-5URL [本文引用: 1]
HUR GH, VICKERY CR, BURKART MD. Explorations of catalytic domains in non-ribosomal peptide synthetase enzymology Natural Product Reports, 2012,29(10):1074-1098. DOI:10.1039/c2np20025bURL [本文引用: 1]
 ,1, 郑敏2, 丁双阳2, 朱奎
,1, 郑敏2, 丁双阳2, 朱奎

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT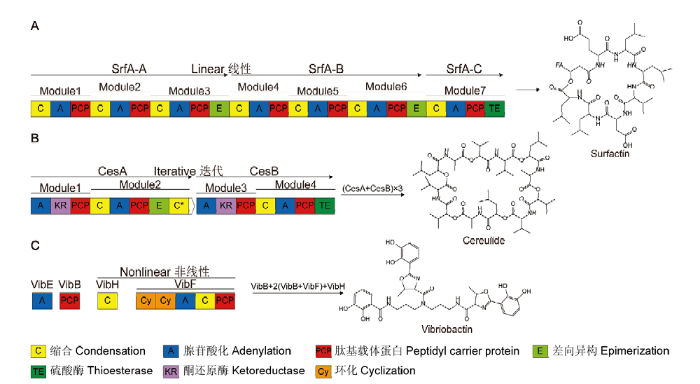 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT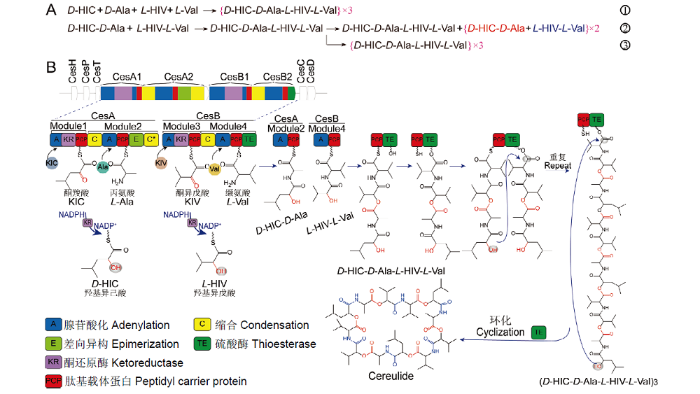 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}