 ,1,2, 何旭江1, 江武军3, 王子龙1, 曾志将,1
,1,2, 何旭江1, 江武军3, 王子龙1, 曾志将,1The Capping Pheromone Contents and Putative Biosynthetic Pathways in Larvae of Honeybees Apis cernana
QIN QiuHong,1,2, HE XuJiang1, JIANG WuJun3, WANG ZiLong1, ZENG ZhiJiang,1通讯作者:
责任编辑: 岳梅
收稿日期:2020-08-24接受日期:2020-11-9网络出版日期:2021-06-01
| 基金资助: |
Received:2020-08-24Accepted:2020-11-9Online:2021-06-01
作者简介 About authors
秦秋红,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (1321KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
秦秋红, 何旭江, 江武军, 王子龙, 曾志将. 东方蜜蜂幼虫封盖信息素含量及生物合成通路[J]. 中国农业科学, 2021, 54(11): 2464-2475 doi:10.3864/j.issn.0578-1752.2021.11.017
QIN QiuHong, HE XuJiang, JIANG WuJun, WANG ZiLong, ZENG ZhiJiang.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】蜜蜂是一种高度社会化的昆虫,蜂群成员间需要进行信息交流来协调个体活动,以确保群体能够健康生存和持续繁衍。作为完全依赖群体的成员,蜜蜂幼虫通过各种化学信号向成年工蜂表明自己的生物状态和需求,以便得到必需的喂养和照料[1,2,3,4]。从分子水平研究东方蜜蜂(Apis cernana)幼虫封盖信息素的生物合成通路,进一步探索蜜蜂幼虫诱导成年工蜂封盖幼虫巢房行为的分子机理,可为深入了解东方蜜蜂信息素交流的内在机制提供新的启示。【前人研究进展】LE CONTE等首次从西方蜜蜂(Apis mellifera)雄蜂幼虫中鉴定出10种幼虫酯类信息素(the brood ester pheromone,BEP):甲基棕榈酸酯(methyl palmitate,MP)、甲基油酸酯(methyl oleate,MO)、甲基亚油酸酯(methyl linoleate,ML)、甲基硬脂酸酯(methyl stearate,MS)、甲基亚麻酸酯(methyl linolenate,MLN)、乙基棕榈酸酯(ethyl palmitate,EP)、乙基油酸酯(ethyl oleate,EO)、乙基亚油酸酯(ethyl linoleate,EL)、乙基硬脂酸酯(ethyl stearate,ES)和乙基亚麻酸酯(ethyl linolenate,ELN)[5]。近30年来科研工作者发现,蜜蜂幼虫信息素中单一或组合的脂肪酸酯能够引起工蜂不同的行为与发育变化,这些信息素成分随着幼虫日龄和性别的不同而发生变化(工蜂或雄蜂),成年工蜂也相应地调整它们对幼虫的行为反应[1,6-7]。然而目前大多数的研究仅涉及信息素的化学成分鉴定和功能描述,对蜜蜂信息素的合成途径研究较少。HE等研究发现E-β-罗勒烯可能是工蜂幼虫的饥饿信息素,并利用RNA-seq技术分析推测出E-β-罗勒烯在工蜂幼虫体内通过乙酰辅酶A和乙酰乙酰辅酶A从头合成的生物合成通路及其调控候选基因[8]。进一步研究表明,蜜蜂蜂王与雄蜂幼虫通过同样的通路在体内从头合成E-β-罗勒烯作为其饥饿信息素来乞求食物[9]。蜜蜂是一种变态发育的昆虫,其生活史要经过卵、幼虫、蛹和成虫4个阶段。在正常的蜂巢环境中(巢房温度约35℃),卵孵化成幼虫并发育到一定时期(蜂王幼虫5 d、工蜂幼虫6 d、雄蜂幼虫7 d),成年工蜂就会从蜡腺分泌蜂蜡对其巢房进行封盖,以利于幼虫在一个干净、稳定的封闭环境中化蛹[10]。LE CONTE等研究发现,蜜蜂幼虫信息素中MP、MO、ML和MLN 4种成分中的单一或混合成分都可以诱导成年工蜂对幼虫巢房的封盖行为[11]。QIN等利用RNA-seq技术分析推测出西方蜜蜂工蜂与雄蜂幼虫利用乙酰辅酶A合成MP、MO、ML和MLN的生物合成通路以及12个调控候选基因,并利用稳定同位素示踪剂证实了这些封盖信息素成分是由幼虫合成的,而不是从它们的食物中获得[12]。蜂螨是危害蜜蜂最严重的寄生性病害之一,导致全球蜜蜂很高的死亡率[13]。研究表明,大蜂螨能利用蜜蜂幼虫信息素找到即将封盖的工蜂或雄蜂幼虫[5,11],在幼虫被封盖前潜入其巢房内,吸食幼虫血淋巴,繁殖后代,而蜂螨更偏好于寄生在雄蜂的幼虫巢房内[10]。【本研究切入点】东方蜜蜂和西方蜜蜂是目前世界上广泛饲养的两个蜂种,它们在外部形态、个体发育、生活习性和学习记忆等生物学特性方面存在较大差异[14,15,16]。目前对蜜蜂信息素的研究大多集中在以意大利蜜蜂(Apis mellifera ligustica)为代表的西方蜜蜂上。虽然近年来我国****逐渐对东方蜜蜂信息素展开研究[17,18],获得了一些东方蜜蜂化学通讯的知识,但相对于西方蜜蜂而言,人们对东方蜜蜂信息素仍知之甚少,尤其是信息素分子水平层面的研究较为缺乏。【拟解决的关键问题】为了丰富对东方蜜蜂信息素的了解,分别利用GC/MS技术和RNA-seq技术,分析封盖信息素化学成分在不同封盖时期的东方蜜蜂工蜂与雄蜂幼虫体内的含量变化及转录组差异,验证MP、MO、ML和MLN是与东方蜜蜂封盖行为相关的信息素并探究其生物合成分子机理,进一步揭示东方蜜蜂幼虫与成年工蜂之间的信息素交流内在机制。1 材料与方法
信息素鉴定试验于2017年4—5月在江西中烟工业有限责任公司完成,转录组测序分析于2017年4—8月在北京百迈客生物科技有限公司完成,荧光定量PCR验证于2018年4—5月在江西农业大学蜜蜂研究所完成。1.1 供试昆虫
供试样品均采自中华蜜蜂(Apis cernana cernana)蜂群,由江西农业大学蜜蜂研究所(28.46°N,115.49°E)根据标准的养蜂技术饲养。幼虫样品均分为以下3个不同封盖时期:4日龄未封盖的工蜂与雄蜂幼虫(4-day-old WL/DL);正在封盖的工蜂与雄蜂幼虫(capping WL/DL),即蜂房上的蜡盖正在被筑建的幼虫;已封盖的工蜂与雄蜂幼虫(capped WL/DL),样品取自完全封盖的蜂房,蜡盖下有一层薄薄的茧衣,工蜂约8日龄,雄蜂约9日龄。各组幼虫均为从蜂箱中直接取出巢脾新鲜采集,每个生物学重复分别来自于群势相同的不同健康蜂群。
1.2 气相色谱-质谱(GC/MS)分析
3个不同封盖时期的幼虫样品数量分别为4日龄未封盖的工蜂与雄蜂幼虫各40只,正在封盖和已封盖的工蜂幼虫各20只,正在封盖和已封盖的雄蜂幼虫各10只,每组样品进行4个生物学重复。将采集的幼虫样品(未经清洗)分别放入装有3 mL二氯甲烷(分析纯,南京化学试剂股份有限公司)的玻璃瓶中,并添加20 μL甲基十九烷酸酯(≥98%,标准品,10 μg·mL-1,Sigma)作为内标。随后,将装有幼虫的玻璃小瓶放置在水平脱色摇床(ZD-9560,江苏盛蓝仪器制造有限公司)上轻轻摇晃30 min(120 r/min),最后将上清液转移到干净的玻璃瓶中并用氮气浓缩至约20 μL。分别取1 μL浓缩后的样品注入气相色谱-质谱(GC/MS)系统(7890A/5975C,Agilent)进行检测,具体参数参照QIN等[12]的报道。在MP、MO、ML和MLN的基峰重建了单个离子监测色谱图,内部标准分别为质荷比(m/z)270、264、294、292和312。对于感兴趣的化合物,通过参考每种化合物的校准曲线,将峰面积转换为数量。外部标准品为MP、MO、ML和MLN(≥99.5%,Sigma),7个浓度的外标(0.25、0.5、1、2、5、10和25 μg·mL-1)被用来构建标准曲线。所构建的4条标准曲线的相关系数均大于99.96%,说明GC/MS系统足够稳定[19],可用于后期对蜜蜂幼虫MP、MO、ML和MLN的定量分析。
1.3 RNA-seq分析
采集未封盖、正在封盖和已封盖的工蜂与雄蜂幼虫各6只,每组样品进行3个生物学重复。取样时将幼虫放入5 mL EP管中,随后迅速放入液氮速冻,并于-80℃保存。按标准TRlzol Reagent法提取RNA(Life technologies,California,USA),并使用琼脂糖凝胶电泳和核酸蛋白分析仪(IMPLEN)对RNA进行质量检测,将检测合格的RNA样品用于cDNA文库构建。cDNA文库的构建和测序由北京百迈客生物科技有限公司完成,具体操作方法参见余爱丽等[20]的报道。对测序获得的18个样品的Raw Data进行去接头、去引物序列、过滤低质量Reads等质量控制,最终获得高质量Clean Reads。采用中华蜜蜂的全基因组序列(ftp://ftp.ncbi.nlm.nih.gov/genomes/all/GCA/002/290/385/GCA_002290385.1_ApisCC1.0/)作为参考基因进行比对。以FPKM(Fragments per Kilobase of transcript per Million fragments mapped)作为衡量指标,对测序样品的基因表达量进行统计。用斯皮尔曼相关系数r2评估各组样品3个生物学重复的相关性[21]。以fold change≥2且FDR<0.05作为筛选差异表达基因(differential expressed gene,DEG)的标准检测各幼虫样品组间的基因表达差异。
为了进一步解读差异表达基因的功能,探究东方蜜蜂幼虫封盖信息素的生物合成途径,利用KEGG(Kyoto Encyclopedia of Genes and Genomes)数据库对差异表达基因进行通路(Pathway)注释和富集分析。首先将差异表达基因最长的转录本转化为蛋白序列,然后利用BLAST软件与KEGG蛋白数据库进行比较,界定标准为e<10-5。为了识别RNA-seq数据中假定的代谢通路富集,使用基于KEGG Orthology的注释系统2.0(KOBAS 2.0)分析KEGG映射的结果。KOBAS 2.0将提供的基因数据库与代谢和信号通路的多个现有数据库进行对照,以识别RNA-seq样本中富集的通路[22]。
1.4 差异表达基因qRT-PCR验证
从本研究的转录组测序结果中推测出东方蜜蜂工蜂与雄蜂幼虫封盖信息素的生物合成通路及参与该通路的11个基因。为了验证转录组测序结果的可靠性,以中华蜜蜂工蜂幼虫为实验材料,随机选取5个参与该通路的候选基因进行qRT-PCR定量分析:Kat(gene- APICC_00421:3-ketoacyl-CoA thiolase)、Hadha(gene-APICC_06057:Trifunctional enzyme subunit alpha)、Mecr(gene-APICC_00389:PREDICTED: probable trans-2-enoyl-CoA reductase, mitochondrial)、Hacd(gene-APICC_02088:PREDICTED: very-long- chain(3R)-3-hydroxyacyl-CoA dehydratase hpo-8)和Δ11 desaturase(gene-APICC_01118:Acyl-CoA Delta(11)desaturase)。采集未封盖、正在封盖和已封盖的工蜂幼虫各3只,每组样品进行3个生物学重复。将采集的样品迅速放入液氮中速冻,并于-80℃保存。按标准TRlzol Reagent法提取RNA并检测RNA样品的质量,随后用反转录试剂盒(TaKaRa)将检测合格的总RNA反转录合成第一链cDNA。用Primer 5.0软件进行基因引物设计,并由上海生物工程有限公司进行引物合成(引物序列详见表1)。选用β-actin为“管家基因”。qRT-PCR反应体系(10 μL): cDNA模板1 μL,上、下游引物各0.4 μL(10 μmol·L-1),SYBR? Premix Ex TaqTM Ⅱ 5 μL,无RNA酶水3.2 μL。qRT-PCR反应条件:95℃预变性30 s;95℃ 10 s,60℃ 1 min,40个循环。扩增反应结束后继续从50℃缓慢加热到90℃,建立PCR产物的熔解曲线。对于每个基因,每个样品进行3个技术重复。用公式2-ΔΔCt计算各个基因在各样品中的相对表达水平[23,24],然后做平方根转换后进行方差分析,检验各基因在工蜂幼虫不同封盖时期的表达量差异情况。
Table 1
表1
表1实时荧光定量PCR引物
Table 1
| 基因编号 Gene ID | 基因名称 Gene name | 正向引物 Forward primer (5′-3′) | 反向引物 Reverse primer (5′-3′) |
|---|---|---|---|
| gene-APICC_00421 | Kat | CCACGGCCTCAAACGACTC | CGGCACCATCAGAAATACCAG |
| gene-APICC_06057 | Hadha | GGAGGAGGCTTGGAGATGG | GCACCAGGCAAGATACCTAACA |
| gene-APICC_00389 | Mecr | TGCCTTCACCAAAATTAGCCC | CCATGCTGTCATCCAAAATCC |
| gene-APICC_01118 | Δ11 desaturase | GCTTGTCAATTCCGCTGCTC | AACCTTCGCCAAGTGCTCCTA |
| gene-APICC_02088 | Hacd | GCAGCTTCATCTTTTGCTCTTC | CTATAACTCCATGCTTCTGGGTG |
| β-actin | TCCTGGAATCGCAGATAGAATG | GGAAGGTGGACAAAGAAGCAAG |
新窗口打开|下载CSV
1.5 数据统计方法
使用StatView 5.01(SAS Institute,Cary,NC,USA)分析GC/MS和qRT-PCR的数据。数据采用“ANOVA and t-test”中的“ANOVA or ANCOVA”进行统计分析,各组间用Fisher’s PLSD进行差异显著性比较分析。组间差异被认为是显著的概率水平为0.05。2 结果
2.1 不同封盖时期幼虫封盖信息素含量
在工蜂中,4个封盖信息素成分在正在封盖和已封盖幼虫体内的含量均显著高于4日龄未封盖的幼虫。其中,MP和MO的含量均随幼虫年龄的增长而增加,且在3个封盖时期间差异显著(P<0.05),而ML和MLN的含量在正在封盖和已封盖的幼虫中差异不显著(P>0.05)(图1)。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1工蜂不同封盖时期幼虫封盖信息素含量
图中数据为平均值±标准误,柱上标不同字母表示差异显著(P<0.05)。
Fig. 1The contents of capping pheromone extracted from worker larvae at different capping stages
The data in the figure were mean±S.E. Different letters on the bars indicated significant differences (P<0.05). The same as
在雄蜂中,从4日龄未封盖幼虫到已封盖幼虫,4种信息素成分含量均随年龄增长而增加,且已封盖幼虫的信息素含量显著高于4日龄和正在封盖的幼虫(P<0.05)。此外,除了ML在正在封盖幼虫体内的含量显著高于4日龄未封盖幼虫外(P<0.05),其余3种信息素成分在这两个封盖时期的幼虫中均差异不显著(P>0.05)(图2)。
图2
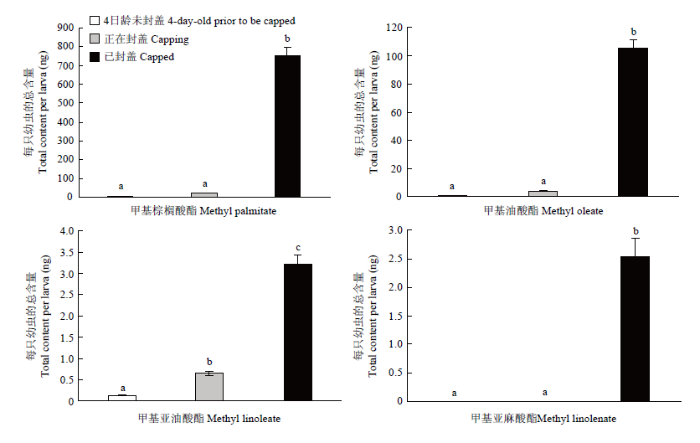 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2雄蜂不同封盖时期幼虫封盖信息素含量
Fig. 2The contents of capping pheromone extracted from drone larvae at different capping stages
2.2 RNA-seq分析
完成了18个样品RNA-seq测序,经过质量控制,共获得57.26 Gb Clean Data,各样品Clean Data均达到2.65 Gb,且Q30碱基百分比均≥94.74%。分别将各样品的Clean Reads与中华蜜蜂参考基因组进行序列比对,比对效率均在88.89%—95.37%,且Unique Reads的比对率≥87.99%,数据利用率正常。各试验组3个生物学重复的斯皮尔曼相关系数r2均>0.75,说明本研究中工蜂与雄蜂各封盖时期幼虫3个重复样品的相关性较强。对工蜂与雄蜂3个不同封盖时期幼虫的基因表达量进行组间比较分析,分别获得4 299和3 926个差异表达基因。在未封盖与正在封盖、正在封盖与已封盖以及未封盖与已封盖3个比较组中,工蜂幼虫分别有62、3 288和3 701个基因存在表达差异,其中表达上调的基因数分别为20、1 812和2 022个,表达下调的基因数分别为42、1 476和1 679个(图3-A);雄蜂幼虫分别有355、2 343和3 489个基因存在表达差异,其中表达上调的基因数分别为204、1 517和1 936个,表达下调的基因数分别为151、826和1 553个(图3-C)。分别对工蜂与雄蜂幼虫的差异表达基因集进行集合分析(
图3
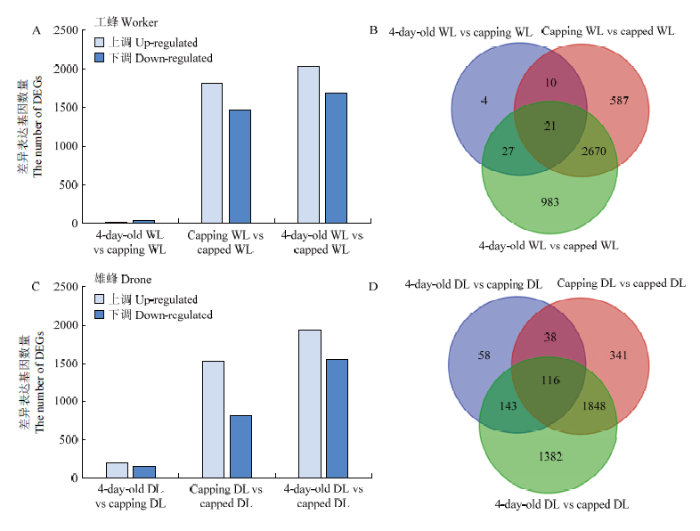 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3工蜂与雄蜂不同封盖时期幼虫差异表达基因
Fig. 3The number of DEGs in worker and drone larvae of different capping stages
在KEGG注释分析中,工蜂不同封盖时期幼虫间的所有差异表达基因共注释到152个KEGG通路,其中未封盖与正在封盖、正在封盖与已封盖以及未封盖与已封盖3个比较组中的差异表达基因分别注释到31、99和22个通路。雄蜂不同封盖时期幼虫间的所有差异表达基因共注释到130个KEGG通路,其中上述3个比较组中的差异表达基因分别注释到65、128和125个通路。KEGG分类注释结果表明,上述KEGG通路涉及到新陈代谢、细胞过程、遗传信息处理、环境信息处理和有机系统5个方面,其中与代谢有关的通路数量最多。
进一步对注释到的所有KEGG通路进行分析,发现两个脂肪酸延伸通路(ko00062:Fatty acid elongation)和不饱和脂肪酸生物合成通路(ko01040:Biosynthesis of unsaturated fatty acids),它们可能涉及长碳链的生物合成途径:首先由乙酰辅酶A通过脂肪酸延伸通路从头合成十六盐酸(C16),然后以C16为前体通过不饱和脂肪酸生物合成通路形成一个或多个双键的不饱和脂肪酸,如油酸和亚油酸。据报道,一些昆虫通过这一途径进行信息素的生物合成[25,26,27]。因此推测,这两个KEGG通路可能与东方蜜蜂工蜂与雄蜂幼虫封盖信息素的生物合成有关。
在工蜂与雄蜂幼虫中,共有11个差异表达基因富集在脂肪酸延伸通路和不饱和脂肪酸生物合成通路。其中5个基因gene-APICC_00389、gene-APICC_00421、gene-APICC_05477、gene-APICC_06057和gene- APICC_10032富集在脂肪酸延伸通路,4个基因gene- APICC_01118、gene-APICC_01590、gene-APICC_03413和gene-APICC_06057富集在不饱和脂肪酸生物合成通路,此外还有2个基因gene-APICC_05451和gene-APICC_02088同时富集到上述两个通路中。推测这11个基因可能参与调控东方蜜蜂工蜂与雄蜂幼虫封盖信息素MP、MO、ML和MLN的生物合成过程(表2)。
Table 2
表2
表2工蜂与雄蜂幼虫封盖信息素生物合成相关候选基因注释
Table 2
| 基因编号Gene ID | 基因注释Gene annotation |
|---|---|
| gene-APICC_00421 | 3-ketoacyl-CoA thiolase (Apis cerana cerana) |
| gene-APICC_06057 | Trifunctional enzyme subunit alpha (Apis cerana cerana) |
| gene-APICC_10032 | Enoyl-CoA hydratase (Apis cerana cerana) |
| gene-APICC_00389 | PREDICTED: probable trans-2-enoyl-CoA reductase, mitochondrial (Apis cerana) |
| gene-APICC_05477 | PREDICTED: elongation of very long chain fatty acids protein 4-like isoform X1 (Apis dorsata) |
| gene-APICC_02088 | PREDICTED: very-long-chain (3R)-3-hydroxyacyl-CoA dehydratase hpo-8 (Apis cerana) |
| gene-APICC_05451 | Trans-2,3-enoyl-CoA reductase (Apis cerana cerana) |
| gene-APICC_01118 | Acyl-CoA Delta (11) desaturase (Apis cerana cerana) |
| gene-APICC_08222 | Acyl-CoA desaturase (Apis cerana cerana) |
| gene-APICC_01590 | PREDICTED: acyl-CoA Delta (11) desaturase-like (Apis cerana) |
| gene-APICC_03413 | Acyl-CoA desaturase (Apis cerana cerana) |
新窗口打开|下载CSV
根据上述结果,提出东方蜜蜂工蜂与雄蜂幼虫封盖信息素的生物合成通路,以及调控这些通路的相关基因(图4)。
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4东方蜜蜂工蜂与雄蜂幼虫封盖信息素生物合成通路及相关调控基因
网格表示工蜂与雄蜂不同封盖时期幼虫的基因表达量。每个格子的灰度等级代表各个基因在幼虫中的绝对表达量,各灰度等级对应的FPKM值分别为0—10、10—20、20—40、40—80、80—160、160—320、320—640、640—1280、1280—2560和2560—5500
Fig. 4Proposed biosynthetic pathways involved in the synthesis of capping pheromone of A. cerana, and expression of related genes in worker and drone larvae of different capping stages
The grids illustrate the gene expression levels for different capping stages larvae of workers and drones. Grey scale density of each cell shows the absolute expression magnitude of honeybee larvae, with the FPKM values 0-10, 10-20, 20-40, 40-80, 80-160, 160-320, 320-640, 640-1280, 1280-2560 and 2560-5500 represented by gray scale levels, respectively
2.3 qRT-PCR
调控东方蜜蜂工蜂与雄蜂幼虫封盖信息素生物合成的5个候选基因Kat、Hadha、Mecr、Hacd和Δ11 desaturase在封盖前后3个时期的表达量上、下调趋势和差异情况均与转录组测序结果相一致(图5)。qRT-PCR结果验证了转录组测序结果的可靠性。图5
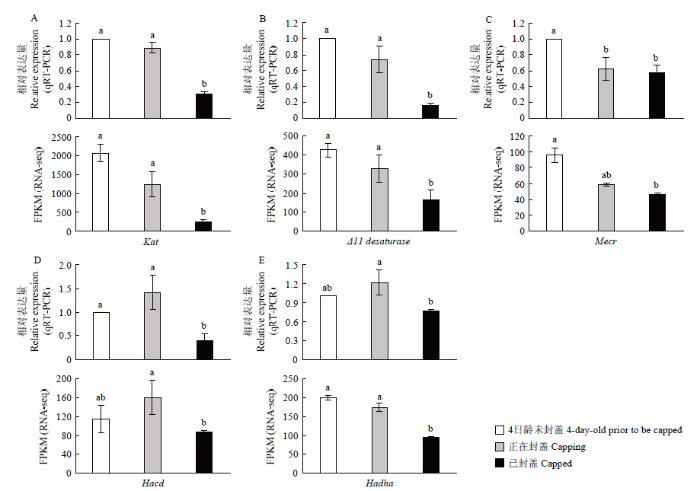 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图55个封盖信息素生物合成候选基因在不同封盖时期幼虫中的表达量
图中数据为平均值±标准误,柱上标不同字母表示表达量差异显著(qRT-PCR:P<0.05;RNA-seq:FDR≤0.05)
Fig. 5Expression levels of five genes possibly implicated in the biosynthesis of capping pheromone components in larvae of different capping stages
The data in the figure were mean±S.E. Different letters on the bars indicated significant differences (qRT-PCR: P<0.05; RNA-seq: FDR≤0.05)
3 讨论
LE CONTE等提出MP、MO、ML和MLN是诱导蜜蜂封盖行为的重要信号[11]。本研究表明,工蜂与雄蜂幼虫在被封蜡盖的关键阶段均增加了这4种信息素成分的释放量,这与前人报道的西方蜜蜂结果相一致[12,28],进一步验证了MP、MO、ML和MLN是与东方蜜蜂封盖行为相关的信息素。在蜂螨的生活周期里,可分为体外寄生期和蜂房繁殖期两个不同的时期。体外寄生期的雌性成螨寄生在巢房外的成年蜂体上,依靠吸取成年蜂的血淋巴生活,当巢房内的蜜蜂幼虫将要被工蜂封盖时,便潜入到幼虫巢房中,并于巢房封盖后转移到幼虫或预蛹体上,靠吮吸其血淋巴补充营养,随后产卵繁殖。蜂螨更偏好于寄生在雄蜂的幼虫巢房内[10]。据报道,在蜂群中有3种BEP成分对蜂螨有吸引作用,其中MP对蜂螨的吸引力最强,而EP和ML的作用相对较弱[5]。在本研究中,MP和ML在正在封盖的雄蜂幼虫中的含量均显著高于同发育时期的工蜂幼虫,试验数据支持早期的研究结果,即这些BEP成分可以被蜂螨用来从正在封盖的工蜂幼虫中区分和挑选出雄蜂幼虫。
颜伟玉等[17]测定了10种幼虫信息素成分在中华蜜蜂不同日龄幼虫和成年蜂个体中的含量分布情况,张含等[18]和曾云峰等[29]先后研究了幼虫信息素3种酯类成分MP、EP和EO对中华蜜蜂工蜂和蜂王个体发育以及工蜂采集、哺育和封盖等行为的影响,并与意大利蜜蜂作比较,均发现中华蜜蜂幼虫与意大利蜜蜂幼虫所分泌的幼虫信息素的成分和含量不同,且这两个蜂种的工蜂对幼虫信息素的反应灵敏度也不同。本研究中,东方蜜蜂工蜂与雄蜂幼虫的MO、ML和MLN含量整体来说均低于已报道的相同实验条件下的西方蜜蜂[12],笔者推测这可能是由于东方蜜蜂嗅觉更灵敏[16],较低的信息素水平即可引起工蜂对幼虫的封盖行为。然而,MP在东方蜜蜂工蜂与雄蜂幼虫中的含量却高于西方蜜蜂。除了能引发工蜂的封盖行为外,MP还被报道与工蜂的发育和采集等行为有关[18,29],但东方蜜蜂幼虫MP含量高的具体原因还有待进一步研究。
信息素的前体主要有3个来源:从头合成、宿主植物或基质提供的前体的转化以及宿主分子的直接结合,但大部分的信息素还是通过昆虫自身的从头合成而来[30]。脂类信息素广泛存在于鳞翅目昆虫中且种类繁多,其生物合成通常是在一系列酶的作用下,利用软脂酸和硬脂酸等脂肪酸中间产物作为前体,经过碳链缩短、去饱和化和官能团修饰等步骤逐步形成信息素。本研究利用转录组测序技术分析了3个不同封盖时期的东方蜜蜂工蜂与雄蜂幼虫各组间的差异表达基因,并对差异表达基因进行KEGG注释和富集分析,发现脂肪酸延伸和不饱和脂肪酸生物合成通路有差异表达基因显著富集,由此笔者推测出东方蜜蜂工蜂与雄蜂幼虫封盖信息素可能的生物合成通路及相关的调控基因。这些生物合成通路是由乙酰辅酶A从头合成C16,然后以C16为前体通过去饱和作用形成一个或多个双键的不饱和脂肪酸。在许多昆虫中,乙酰辅酶A是主导脂肪酸和酯类生物合成的几个不同代谢途径的前体[31,32,33],昆虫能利用上述途径进行信息素的生物合成[26,27,28],如在家蚕、钩端螺旋体和大豆中均发现了类似的甲酯合成途径[26-27,34-35]。这些研究支持关于东方蜜蜂幼虫4种封盖信息素成分生物合成通路的推测。本研究中提出的东方蜜蜂工蜂与雄蜂幼虫封盖信息素生物合成通路与QIN等[12]报道的西方蜜蜂工蜂与雄蜂幼虫封盖信息素生物合成通路相同。东方蜜蜂与西方蜜蜂生存的自然环境、气候条件和植物区系等因素不同,其生物学特性存在较大的差异,如外部形态、个体发育、生活习性和学习记忆等[14,15,16]。东方蜜蜂和西方蜜蜂是同属于蜜蜂属的两个蜂种,其遗传背景相似,都具有蜜蜂属的特性,如高度的社会性生活、采蜜和贮蜜以及筑造双面六边形巢房等。已有研究表明,东方蜜蜂幼虫含有与西方蜜蜂幼虫相同的10种脂肪酸酯类信息素[17],其中一些信息素成分已被证实在影响工蜂发育和行为方面具有与西方蜜蜂相似的效应。因此,笔者推测东方蜜蜂与西方蜜蜂有可能利用相同的生物合成途径进行信息素的生物合成。
作用于信息素合成过程的脱氢酶基因已在许多昆虫中被发现。在本研究中,2个Δ11-desaturase基因(gene-APICC_01118和gene-APICC_01590)以及2个Acyl-CoA desaturase基因(gene-APICC_08222和gene-APICC_03413)富集在笔者提出的东方蜜蜂幼虫封盖信息素的生物合成通路,且GO注释表明gene-APICC_08222和gene-APICC_03413具有Δ11-desaturase的生物功能。Δ11脱氢酶是蛾类和蝴蝶类在C11上形成双键最主要的酶[36,37,38,39,40,41]。Δ11-desaturase是一种生物功能酶,具有类似于Δ11和Δ12的去饱和活性,在昆虫信息素生物合成过程中发挥着重要作用[42,43,44,45,46]。ROELOFS等研究发现,红带卷叶蛾、大白菜尺蛾和家蚕中的油酸酯、亚油酸酯和亚麻酸酯是从头合成的,并由Δ11-desaturase去饱和[47]。因此,推测这4个基因可能参与了东方蜜蜂幼虫封盖信息素成分的生物合成途径。将参与调控东方蜜蜂封盖信息素生物合成通路的11个候选基因与已报道的西方蜜蜂的12个候选基因进行比对,发现东方蜜蜂中的10个候选基因(gene-APICC_05477除外)与西方蜜蜂中的10个候选基因有相同的注释或基因功能,这些基因分别在东方蜜蜂和西方蜜蜂幼虫封盖信息素生物合成通路中发挥着相同的调控作用。进一步比较这些候选基因在未封盖、正在封盖和已封盖工蜂幼虫与雄蜂幼虫中的表达量可知,这些基因在同一封盖时期的工蜂与雄蜂幼虫中的表达量相近。上述比较结果进一步为这些基因作为参与蜜蜂幼虫封盖信息素生物合成通路的调控基因提供了证据,但确切的调控机制还有待进一步研究。
4 结论
东方蜜蜂工蜂幼虫与雄蜂幼虫在被封蜡盖的关键阶段增加了甲基棕榈酸酯(MP)、甲基油酸酯(MO)、甲基亚油酸酯(ML)和甲基亚麻酸酯(MLN)的释放量,进一步验证了它们是与蜜蜂封盖行为相关的信息素,推测这些信息素可能是在相关基因的调控下由乙酰辅酶A从头合成,而东方蜜蜂幼虫与西方蜜蜂幼虫可能利用相同的生物合成通路进行信息素的生物合成。致谢:
本研究得到了江西中烟工业有限责任公司郭磊和江西农业大学麻俊武老师的帮助,在此表示感谢!参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1016/S0003-3472(83)80077-3URL [本文引用: 2]
DOI:10.1146/annurev.en.15.010170.001043URL [本文引用: 1]
DOI:10.1007/BF01048280URL [本文引用: 1]
DOI:10.1007/BF01054610URL [本文引用: 1]
DOI:10.1126/science.245.4918.638URL [本文引用: 3]
DOI:10.1007/s10886-005-7623-9URL [本文引用: 1]
DOI:10.1007/BF01136651URL [本文引用: 1]
DOI:10.1038/srep22359URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
DOI:10.1007/BF01138390URL [本文引用: 3]
DOI:10.1007/s13592-019-00686-9URL [本文引用: 5]
DOI:10.1007/BF00981926URL [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
DOI:10.1007/s00359-012-0747-9URL [本文引用: 3]
[本文引用: 3]
[本文引用: 3]
[本文引用: 3]
[本文引用: 3]
DOI:10.1016/j.chroma.2011.10.077URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1186/s13059-014-0550-8URL [本文引用: 1]
DOI:10.1093/nar/gkr483URL [本文引用: 1]
DOI:10.1006/abio.2001.5530URL [本文引用: 1]
DOI:10.1093/bioinformatics/btp578URL [本文引用: 1]
DOI:10.1016/0045-2068(84)90011-7URL [本文引用: 1]
[本文引用: 3]
[本文引用: 3]
DOI:10.1051/apido/2010017URL [本文引用: 2]
[本文引用: 2]
[本文引用: 2]
DOI:10.1016/S0965-1748(99)00016-8URL [本文引用: 1]
DOI:10.1016/S0021-9258(19)41198-8URL [本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.ibmb.2003.06.008URL [本文引用: 1]
DOI:10.1111/ejb.1969.8.issue-1URL [本文引用: 1]
DOI:10.1007/s11032-008-9246-7URL [本文引用: 1]
[本文引用: 1]
DOI:10.1186/1471-2148-8-270URL [本文引用: 1]
DOI:10.1016/j.ibmb.2011.05.003URL [本文引用: 1]
DOI:10.1371/journal.pone.0037230URL [本文引用: 1]
DOI:10.1038/ncomms4957URL [本文引用: 1]
DOI:10.1007/s10886-014-0452-yURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.ibmb.2006.05.005URL [本文引用: 1]
DOI:10.1007/BF01940732URL [本文引用: 1]
DOI:10.1016/0965-1748(92)90152-5URL [本文引用: 1]
DOI:10.1002/(ISSN)1520-6327URL [本文引用: 1]
DOI:10.1016/0045-2068(84)90011-7URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}