Fusarium verticillioides Resistant Maize Inbred Line Development Using Host-Induced Gene Silencing Technology
HE KeWei,1, CHEN JiaFa2, ZHOU ZiJian,2, WU JianYu,1,21College of Agronomy, Henan Agricultural University, Zhengzhou 450002 2College of Life Sciences of Henan Agricultural University, Zhengzhou 450002
Abstract 【Objective】Fusarium verticillioides (F. verticillioides) is a common pathogen, which can cause ear rot, stalk rot, seedling blight, and seed rot in maize. These diseases caused by Fusarium verticillioides not only affected the yield and quality of maize, but also seriously threatened to the safety of human and livestock by a variety of fungal toxins such as fumonisin which produced during the metabolic process of the pathogen. So far, there is no report about the major resistance gene cloned and utilized for Fusarium verticillioides in maize. Using host-induced gene silencing technology provides a new strategy for resistance breeding in maize. 【Method】Key genes associated with the Fusarium verticillioides development were cloned using homologous gene sequence method, and the dsRNA were produced by in-vitro transcription (IVT) assay. The dsRNA for different genes was premixed with suspension spores of Fusarium verticillioides used for RNA silencing experiment in vitro. For investigate the degree of disease, the seeds of the susceptible inbred line Xi502 were sterilized and inoculated, and then were cultured in a petri dish at 28℃ in the dark for 48 h. For investigate the incidence of the seeds after inoculation, glucose was added to the spore suspension mixed with dsRNA, then spore germination and mycelia growth were observed under the microscope after 25℃ culture for 24 h. The Xi502 seedlings of the trifoliate stage were transferred to the spore suspension with premixed dsRNA for culture, and the incidence of seedlings blight was observed after 7 days. In order to select the target gene for HIGS, combine the seed morphological observation result after inoculation and seedling inoculation result. Then, the silent vector about these key target genes were constructed and transferred into the susceptible inbred line Xi502. The transgenic seeds were evaluated by artificial inoculation. The total RNA of the transgenic seeds after inoculation was extracted, and the relative expression of target genes in F. verticillioide was analyzed by qRT-PCR to determine the silencing effect of HIGS line. 【Result】Eighteen candidate genes related to growth were cloned by homologous cloning method in Fusarium verticillioides. It was found that the disease level of seeds was significantly reduced after 11 candidate genes silencing by seed inoculation experiments. Furthermore, six of the 11 candidate target genes, deo, Ras2, Dpdc, Hsp90, Frp1, and Atg15, were found that response to the spore germination and mycelium growth after gene silencing. Finally, based on the results of seedling inoculation, 3 silencing target genes deo, Atg15 and Frp1 with significant inhibitory effect in vitro were selected. Then the silencing vector was constructed by combine three specific segments from the three target genes, transgenic plants were obtained. It was found that the resistance level was highly increased in T2-generation seeds compared to the none-transgenic plants. As well as the expression levels of all the three target genes were significantly decreased in Fusarium verticillioides. 【Conclusion】Three genes, deo, Atg15 and Frp1, are important for development of Fusarium verticillioides. By constructing transgenic HIGS plants for target gene deo, Atg15 and Frp1, the increase the resistance to Fusarium verticillioides in maize. Keywords:maize (Zea mays L.);Fusarium verticillioides;host-induced gene silencing;transgenic;ear rot
PDF (2384KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 赫可伟, 陈甲法, 周子键, 吴建宇. 基于宿主诱导的基因沉默技术创制抗拟轮枝镰孢玉米自交系[J]. 中国农业科学, 2021, 54(9): 1835-1845 doi:10.3864/j.issn.0578-1752.2021.09.002 HE KeWei, CHEN JiaFa, ZHOU ZiJian, WU JianYu. Fusarium verticillioides Resistant Maize Inbred Line Development Using Host-Induced Gene Silencing Technology[J]. Scientia Acricultura Sinica, 2021, 54(9): 1835-1845 doi:10.3864/j.issn.0578-1752.2021.09.002
A: Seeds appearance of WT and HIGS transgenic lines at 7 days post inoculation. B: Statistics of the disease grade of seed rot at 7 days post inoculation in WT and HIGS transgenic lines. C: The relative expression of 3 target genes at 3 days post inoculation in WT and HIGS transgenic lines
WANGX M, JINQ M, SHIJ, WANGZ Y, LIX. The status of maize diseases and the possible effect of variety resistance on disease occurrence in the future Acta Phytopathologica Sinica, 2006,36(1):1-11. (in Chinese) [本文引用: 1]
KAMLEM, MAHATOD K, DEVIS, LEEK E, KANGS G, KUMARP. Fumonisins: Impact on agriculture, food, and human health and their management strategies Toxins, 2019,11(6):328. [本文引用: 1]
KNUTSENH K, ALEXANDERJ, BARREG?RDL, BIGNAMIM, BRüSCHWEILERB, CECCATELLIS, COTTRILLB, DINOVIM, EDLERL, GRASL-KRAUPPB, HOGSTRANDC, HOOGENBOOML, NEBBIAC S, PETERSENA, ROSEM, ROUDOTA C, SCHWERDTLET, VLEMINCKXC, VOLLMERG, WALLACEC, DALL'ASTAG S, TARANUI, ALTIERIA, ROLDáN-TORRES R, OSWALDI P. Risks for animal health related to the presence of fumonisins, their modified forms and hidden forms in feed EFSA Journal, 2018,16(5):5242. [本文引用: 1]
ROSSP F, RICEL G, PLATTNERR D, OSWEILERG D, WILSONT M, OWENSD L, NELSONH A, RICHARDJ L. Concentrations of fumonisin B1 in feeds associated with animal health problems Mycopathologia, 1991,114(3):129-135. [本文引用: 1]
MARASASW F O. Fumonisins: Their implications for human and animal health Natural Toxins, 1995,3(4):193-198. [本文引用: 1]
GELDERBLOMW C A, JASKIEWICZK, MARASASW F O, THIELP G, HORAKR M, VLEGGAARR, KRIEKN P. Fumonisins: Novel mycotoxins with cancer-promoting activity produced by Fusarium moniliforme Applied & Environmental Microbiology, 1988,54(7):1806-1811. [本文引用: 1]
YOSHIZAWAT, YAMASHITAA, LUOY. Fumonisin occurrence in corn form high-risk and low-risk areas for human esophageal cancer in China Applied and Environmental Microbiology, 1994,60(5):1626-1629. [本文引用: 1]
UENOY, IIJIMAK, WANGS D, SUGIURAY, SEKIJIMAM, TANAKAT, CHENC, YUS Z. Fumonisins as a possible contributory risk factor for primary liver cancer: A 3-year study of corn harvested in Haimen, China, by HPLC and ELISA Food and Chemical Toxicology, 1997,35(12):1143-1150. [本文引用: 1]
ZUOW, CHAOQ, ZHANGN, YEJ, TANG, LIB, XINGY, ZHANGB, LIUH, FENGLERK A, ZHAOJ, ZHAOX, CHENY, LAIJ, YANJ, XUM. A maize wall-associated kinase confers quantitative resistance to head smut Nature Genetics, 2015,47(2):151-157. [本文引用: 1]
WANGC, YANGQ, WANGW, LIY, GUOY, ZHANGD, MAX, SONGW, ZHAOJ, XUM. A transposon-directed epigenetic change in ZmCCT underlies quantitative resistance to Gibberella stalk rot in maize The New Phytologist, 2017,215(4):1503-1515. [本文引用: 1]
LUNSFORDJ N, FUTRELLM C, SCOTTG E. Maternal influence on response of corn to Fusarium moniliforme Phytopathology, 1974,65:223-225. [本文引用: 1]
ROBERTSON-HOYTL A, JINESM P, BALINT-KURTIP J, KLEINSCHMIDTC E, WHITED G, PAYNEG A, MARAGOSC M, MOLNáRT L, HOLLANDJ B. QTL mapping for Fusarium ear rot and fumonisin contamination resistance in two maize populations Crop Science, 2006,46(4):1734-1745. [本文引用: 1]
MASCHIETTOV, COLOMBIC, PIRONAR, PEAG, STROZZIF, MAROCCOA, ROSSINIL, LANUBILEA. QTL mapping and candidate genes for resistance to Fusarium ear rot and fumonisin contamination in maize BMC Plant Biology, 2017,17(1):20. [本文引用: 1]
SEPTIANIP, LANUBILEA, STAGNATIL, BUSCONIM, NELISSENH, MARIOENRICO P, DELL’ACQUAM, MAROCCOA. Unravelling the genetic basis of Fusarium seedling rot resistance in the MAGIC maize population: Novel targets for breeding Scientific Reports, 2019,9(1):5665. [本文引用: 1]
ZHANGF, WANX Q, PANG T. Molecular mapping of QTL for resistance to maize ear rot caused by Fusarium moniliforme Acta Agronomica Sinica, 2007,33(3):491-496. (in Chinese) [本文引用: 1]
DINGJ Q, WANGX M, CHANDERS, YANJ B, LIJ S. QTL mapping of resistance to Fusarium ear rot using a RIL population in maize Molecular Breeding, 2008,22(3):395-403. [本文引用: 1]
STAGNATIL, LANUBILEA, SAMAYOAL F, BRAGALANTIM, GIORNIP, BUSCONIM, HOLLANDJ B, MAROCCOA. A genome wide association study reveals markers and genes associated with resistance to Fusarium verticillioides infection of seedlings in a maize diversity panel Genes, Genomes, Genetics, 2019,9(2):571-579. [本文引用: 1]
ZILAC T, OGUTF, ROMAYM C, GARDNERC A, BUCKLERE S, HOLLANDJ B. Genome-wide association study of Fusarium ear rot disease in the U.S.A. maize inbred line collection BMC Plant Biology, 2014,14:372. [本文引用: 1]
LANUBILEA, FERRARINIA, MASCHIETTOV, DELLEDONNEM, MAROCCOA, BELLIND. Functional genomic analysis of constitutive and inducible defense responses to Fusarium verticillioides infection in maize genotypes with contrasting ear rot resistance BMC Genomics, 2014,15(1):710. [本文引用: 1]
YAOL, LIY, MAC, TONGL, DUF, XUM. Combined genome-wide association study and transcriptome analysis reveal candidate genes for resistance to Fusarium ear rot in maize Journal of Integrative Plant Biology, 2020,62(10):1535-1551. [本文引用: 1]
SUDARSHANAM R, ROYG, FALKB W. Methods for engineering resistance to plant viruses Methods in Molecular Biology, 2007,354:183-195. [本文引用: 1]
WATERHOUSEP M, FUSAROA F. Viruses face a double defense by plant small RNAs Science, 2006,313(5783):54-55. [本文引用: 1]
BAUMJ A, BOGAERTT, CLINTONW, HECKG R, FELDMANNP, ILAGANO, JOHNSONS, PLAETINCKG, MUNYIKWAT, PLEAUM, VAUGHNT, ROBERTSJ. Control of coleopteran insect pests through RNA interference Nature Biotechnology, 2007,25(11):1322-1326. [本文引用: 1]
KHATRIM, RAJAMM V. Targeting polyamines of Aspergillus nidulansby siRNA specific to fungal ornithine decarboxylase gene Medical Mycology, 2007,45(3):211-220. [本文引用: 2]
TINOCOM, BáRBARAD, DALL'ASTTAR, JO?OP, ARAG?OF. In vivo trans-specific gene silencing in fungal cells by in planta expression of a double-stranded RNA BMC Biology, 2010,8:27. [本文引用: 2]
NOWARAD, GAYA, LACOMMEC, SHAWJ, RIDOUTC, DOUCHKOVD, HENSELG, KUMLEHNJ, SCHWEIZERP. HIGS: Host-induced gene silencing in the obligate biotrophic fungal pathogen Blumeria graminis The Plant Cell, 2010,22(9):3130-3141. [本文引用: 2]
KOCHA, KUMARN, WEBERL, KELLERH, IMANIJ, KOGELK H. Host-induced gene silencing of cytochrome P450 lanosterol C14 -demethylase-encoding genes confers strong resistance to Fusarium species Proceedings of the National Academy of Sciences of the USA, 2013,110(48):19324-19329. [本文引用: 6]
THAKARED, ZHANGJ, WINGR A, COTTYP J, SCHMIDTM A. Aflatoxin-free transgenic maize using host-induced gene silencing Science Advances, 2017,3(3):e1602382. [本文引用: 5]
BLUHMB H, ZHAOX, FLAHERTYJ E, XUJ R, DUNKLEL D. RAS2 regulates growth and pathogenesis in Fusarium graminearum Molecular Plant-Microbe Interactions, 2007,20(6):627-636. [本文引用: 2]
JIANGJ, LIUX, YINY, MAZ. Involvement of a velvet protein FgVea in the regulation of asexual development, lipid and secondary metabolisms and virulence in Fusarium graminearum PLoS ONE, 2011,6(11):e28291. [本文引用: 1]
ADáMA L, KOHUTG, HORNOKL. Fphog1, a hog-type map kinase gene, is involved in multistress response in Fusarium proliferatum Journal of Basic Microbiology, 2010,48(3):151-159. [本文引用: 1]
COLABARDINIA C, BROWNN A, SAVOLDIM, GOLDMANM H S, GOLDMANG H. Functional characterization of Aspergillus nidulans ypka, a homologue of the mammalian kinase SGK PLoS ONE, 2013,8(3):e57630. [本文引用: 2]
CHOIY E, SHIMW B. Functional characterization of Fusarium verticillioides CPP1, a gene encoding a putative protein phosphatase 2A catalytic subunit Microbiology, 2008,154(1):326-336. [本文引用: 1]
EATONC J, CABRERAI E, SERVINJ A, WRIGHTS J, COXM P, BORKOVICHK A. The guanine nucleotide exchange factor RIC8 regulates conidial germination through Gα proteins in Neurospora crassa PLoS ONE, 2012,7(10):e48026. [本文引用: 3]
VIJAIB, SABINEB, ALBERTV, GOPALANS, WEIY D. Alanine: glyoxylate aminotransferase 1 is required for mobilization and utilization of triglycerides during infection process of the rice blast pathogen, Magnaporthe oryzae Plant Signaling & Behavior, 2012,7(9):1206-1208. [本文引用: 1]
NGUYENL N, J?RGB, LEG T, ST?RKELC, SCH?FERW. Autophagy-related lipase FgATG15 of Fusarium graminearum is important for lipid turnover and plant infection Fungal Genetics and Biology, 2011,48(3):217-224. [本文引用: 2]
ZHANGH, GUOJ, VOEGELER T, ZHANGJ, DUANY, LUOH, KANGZ. Functional characterization of calcineurin homologs pscna1/pscnb1 in Puccinia striiformis f sp tritici using a host-induced RNAi system PLoS ONE, 2012,7(11):e49262. [本文引用: 2]
DUYVESTEIJNR G E, WIJKR V, BOERY, REPM, HARINGM A. Frp1 is a Fusarium oxysporum f-box protein required for pathogenicity on tomato Molecular Microbiology, 2005,57(4):1051-1063. [本文引用: 2]
LAMOTHF, JUVVADIP R, FORTWENDELJ R, STEINBACHW J. Heat shock protein 90 is required for conidiation and cell wall integrity in Aspergillus fumigatus Eukaryotic Cell, 2012,11(11):1324-1332. [本文引用: 1]
ZHAOP B, RENA Z, XUH J, LID C. The gene fpk1, encoding a camp-dependent protein kinase catalytic subunit homolog, is required for hyphal growth, spore germination, and plant infection in Fusarium verticillioides Journal of Microbiology & Biotechnology, 2010,20(1):208. [本文引用: 1]
WORIEDHM, HAUBERI, MARTINEZ-ROCHAA L, VOIGTC, MAIERF J, SCHR?DERM, MEIERC, HAUBERJ, SCH?FERW. Preventing fusarium head blight of wheat and cob rot of maize by inhibition of fungal deoxyhypusine synthase Molecular Plant-Microbe Interactions, 2011,24(5):619-627. [本文引用: 2]
CHENJ, DINGJ, LIH, LIZ, SUNX, LIJ, WANGR, DAIX, DONGH, SONGW, CHENW, XIAZ, WUJ. Detection and verification of quantitative trait loci for resistance to Fusarium ear rot in maize Molecular Breeding, 2012,30(4):1649-1656. [本文引用: 1]
MUC, GAOJ, ZHOUZ, WANGZ, SUNX, ZHANGX, DONGH, HANY, LIX, WUY, SONGY, MAP, DONGC, CHENJ, WUJ. Genetic analysis of cob resistance to F. verticillioides: Another step towards the protection of maize from ear rot Theoretical and Applied Genetics, 2019,132(4):1049-1059. [本文引用: 1]
DOLORESB R, FERNANDOC G, CARLOSL G A, VICTORH A R, JOSEL C S. Responses of maize landrace seedlings to inoculations of Fusarium spp Open Access Library Journal, 2017,4(6):1-14. [本文引用: 1]
JUM, ZHOUZ, MUC, ZHANGX, GAOJ, LIANGY, CHENJ, WUY, LIX, WANGS, WENJ, YANGL, WUJ. Dissecting the genetic architecture of Fusarium verticillioides seed rot resistance in maize by combining QTL mapping and genome-wide association analysis Scientific Reports, 2017,7:46446. [本文引用: 1]
 ,1, 陈甲法2, 周子键
,1, 陈甲法2, 周子键

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT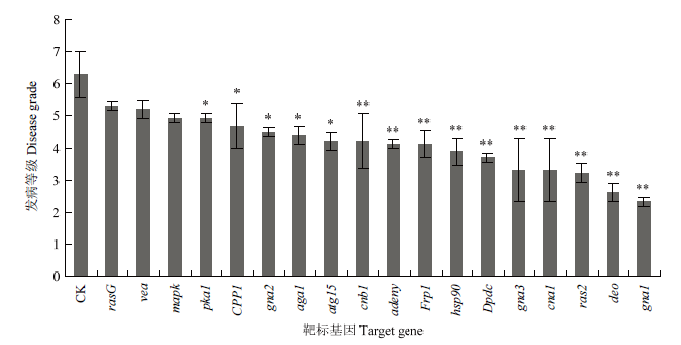 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT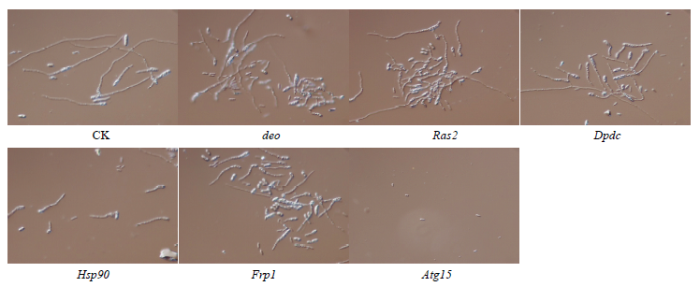 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT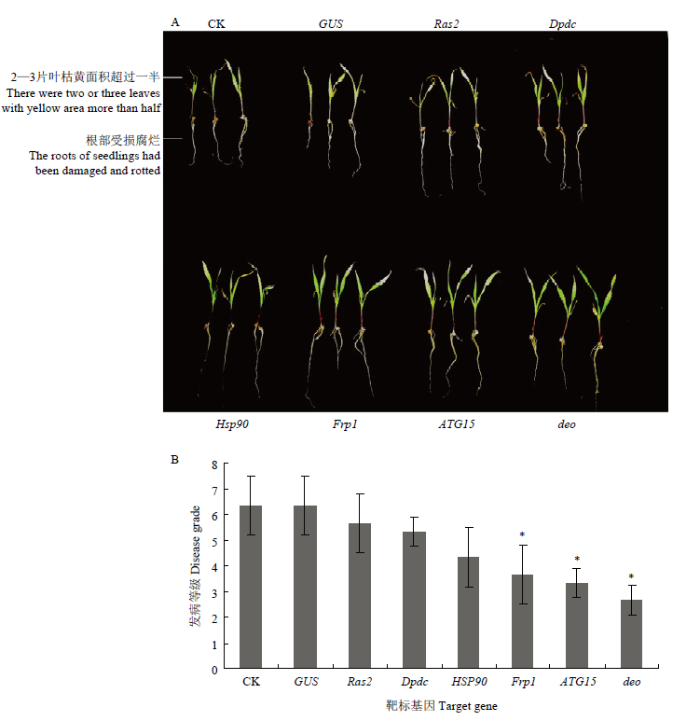 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT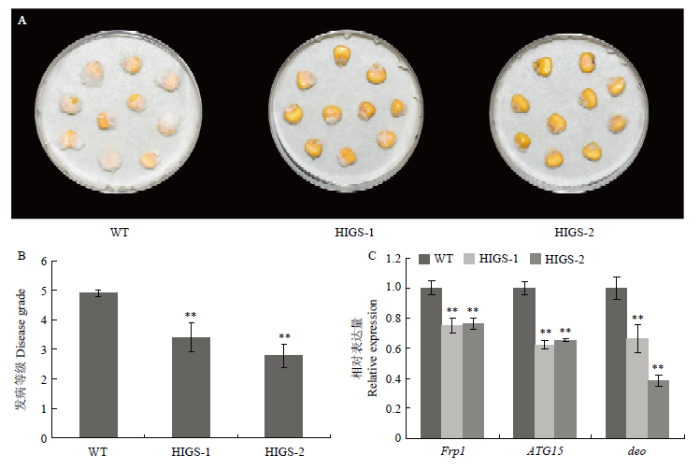 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}